
Submitted:
18 April 2024
Posted:
18 April 2024
You are already at the latest version
Abstract
Amidst worsening climate change, drought stress imperils global agriculture, jeopardizing crop yields and food security, necessitating urgent exploration of sustainable methods like biopriming, harnessing beneficial microorganisms to bolster plant resilience. Recent research has revealed diverse biological compounds with versatile applications produced by Schizophyllum commune, rendering this fungus as a promising contender for biopriming applications. For the first time this study aimed to investigate,the potential of S. commune exo- (EPSH) and intra-PSH (IPSH), isolated from two strains: Italian (ITA) and Serbian (SRB) under submerged cultivation, in enhancing the resilience of Pisum sativum L. seeds through biopriming technique. Testing of seed quality of bioprimed, hydroprimed, and unprimed seeds was done using a germination test, under optimal and drought conditions. In optimal conditions, the IPSH SRB increased germination energy by 5.50% compared to the control, while the highest percentage of germination (94.70%) was shown after biopriming with PSH isolated from the ITA strain. Additionally, all assessed treatments resulted in a boost in seedling growth and biomass accumulation, where the ITA strain demonstrated greater effectiveness in optimal conditions, whereas the SRB strain showed superiority in drought conditions. The drought tolerance indices increased significantly in response to all examined treatments during the drought, with EPSH ITA (23.00%) and EPSH SRB (24.00%) demonstrating the greatest effects. Results demonstrate a significant positive influence of fungal PSH, indicating their potential as biopriming agents and offers insights into novel strategies for agricultural resilience in the face of changing environmental conditions.
Keywords:
agricultural application
; fungi
; biopriming
; stress condition
; submerged cultivation
; S. commune
; P. sativum
1. Introduction
In the face of escalating climate change, drought stress poses a significant threat to global agriculture, compromising crop productivity and food security [1]. Finding sustainable and efficient methods to bolster plant resilience against drought stress has become a paramount objective in agricultural research. Among the various strategies to mitigate drought stress, biopriming—an eco-friendly technique involving the use of beneficial microorganisms—has emerged as a promising approach to enhance crop resilience [2].
Biopriming is a cutting-edge seed treatment technique that involves the application of beneficial microorganisms, such as fungi, bacteria, or actinomycetes or their products onto the seed surface to enhance seed germination, seedling vigor, and overall plant health [3]. This innovative method aims to improve seed quality, uniformity, and establishment while mitigating the adverse effects of various biotic and abiotic stresses on plants. Biopriming with these beneficial microorganisms has shown promising results in enhancing plant growth, promoting antioxidative defense systems, and increasing biomass and yield in various crops [4]. Moreover, it helps plants cope with abiotic stresses like drought, salinity, or low temperatures by improving water uptake and enhancing root and shoot growth [5]. Also, this technique is considered an eco-friendly alternative to chemical treatments, contributing to sustainable agriculture practices [6].
Pea (Pisum sativum L.) stands as an emblematic plant species that has long served as a model organism for understanding various aspects of plant biology, and is cultivated globally for its nutritional value and versatility in culinary applications [7]. Consumption of P. sativum L. has been associated with various health benefits beyond basic nutrition, since the plant's components, including starch, protein, fiber, vitamins, minerals, and phytochemicals, offer antioxidant properties, gastrointestinal health benefits, reduced glycemic index due to its intermediate amylose content, and potential bioactivities like angiotensin I-converting enzyme inhibition and antioxidant activity [8]. Hence, vegetable peas in the last 20 years have gained significant attention all over the world [9]. Moreover, pea plays a vital role in agriculture as a legume crop that provides protein-rich feed and contributes to sustainable farming practices through nitrogen fixation [10]. Its cultivation supports soil health by enhancing nitrogen levels through symbiotic relationships with nitrogen-fixing bacteria [10]. However, pea susceptibility to drought stress, underscores the urgent need to fortify its defenses against such environmental challenges [11].
Among many fungal species, Schizophyllum commune Fries 1815 stands out for its remarkable adaptability, while recent studies have unveiled the different biological compounds synthesized by S. commune as potent bioactive compounds with multifaceted applications [12,13,14]. Research has shown that this fungus produces compounds like alkaloids, flavonoids, phenols, saponins, and tannins that can affect other microorganisms [15]. Additionally, this species exhibits increased metabolism, leading to the production of various metabolites involved in glycolysis, fatty acid synthesis, and amino acid synthesis. Moreover, during years several studies have highlighted polysaccharides (PSH) of S. commune with immunomodulatory effects on humans, antioxidant, and antimicrobial properties against Escherichia coli, Bacillus cereus, Staphylococcus aureus and Salmonella sp. [16,17]. The most investigated PSH from S. commune is beta-glucan, schizophyllan, that has been found to exhibit diverse biological effects, including antitumor, immunomodulatory, and anti-inflammatory activities [18]. In our previous research we have identified PSHs isolated from Italian (ITA) and Serbian (SRB) strains of S. commune after submerged cultivation as schizophyllan [17] and have proven its anti-acetylcholinesterase and antioxidant properties. The potential of its bioactive compounds and metabolic activities extends beyond medicine to agriculture application, making S. commune a promising candidate as a biopriming agent that could positively influence seed germination and plant growth and development. However, the potential of these PSH in conferring drought tolerance to agricultural crops remains largely unexplored, even though understanding how these fungal PSH fortify pea seeds could revolutionize crop resilience in the face of changing climate patterns.
This research aims to unveil the hidden armor that fungi and their biological compounds like PSH provide to plants, shedding light on a novel approach to enhance crop survival under adverse environmental conditions. Therefore, in this research, we have examined and contrasted the impact of PSH derived from submerged cultivation of two S. commune strains originating from Italy and Serbia, on the seed quality performance of P. sativum L. subsequent to biopriming under both, optimal and drought conditions.
2. Materials and Methods
2.1. Fungal Material
Two dikaryon strains of the wild-growing S. commune Fries 1815, belonging to the Phylum Basidiomycota, Class Agaricomycetes, Order Auriculariales, and Family Schizopyllaceae, were collected in 2016 near Bologna (Italy, IT), and in 2012 in Zmajevac (Fruška Gora low Mountain chain) in Serbia (SRB). The identification of fungal species was conducted through a study of their fungal morphology both macroscopically (considering color, shape, size, and hyphae) and microscopically. Mycelia were extracted from the fruiting bodies of both isolates and then cultured at 26 °C for 10 days on malt agar (Torlak, Serbia). These mycelia from both isolates were preserved in the fungal culture collection FUNGICULT at the ProFungi laboratory (Department of Biology and Ecology, Faculty of Sciences, University of Novi Sad; https://www.pmf.uns.ac.rs/en/research/groups/profungi/ ). Each isolate was assigned a specific reference number: 0043 for S. commune SRB and 0047 for S. commune IT as presented in Mišković et al. [17].
2.2. Fungal Cultivation and Polysaccharide Extraction Process
The polysaccharide (PSH) extracts were prepared following a modified method outlined by Chen et al. [19]. PSH extracts from both S. commune strains (SRB and ITA) were simultaneously extracted to obtain exo-polysaccharide (EPSH) from the filtrate (F) and intra-polysaccharide (IPSH) from the mycelia biomass (BM). The extraction process began with inoculating the respective fungal strains from the culture collection (FUNGICULT, ProFungi laboratory, Faculty of Sciences, University of Novi Sad) onto Malt agar (Torlak, Serbia) and cultivating them in a thermostat (IKA-Werke GmbH & Co. KG, Staufen in Breisgau, Germany) at 26 °C for 12 days. Subsequently, plaques (1 cm2) were transferred into 100 mL of liquid medium [17] for submerged cultivation on a thermostatic shaker (120 rpm, 26 °C; IKA, KS 4000, Germany) for 14 days. After cultivation, the samples were filtered to separate BM from the F components. The EPSH was then precipitated overnight at +4 °C using absolute ethanol (Sigma-Aldrich (Steinheim, Germany), followed by filtration and centrifugation twice at +4 °C (10,000 g, 20 min; Centrifuge 5810 R, Eppendorf, Germany) to remove the supernatant. The resulting precipitate was dried at +60 °C (Memmert UF55, Germany) for 20 min to eliminate residual water and ethanol, and subsequently rehydrated in distilled water (dH2O) on a magnetic stirrer (Velp Scientifica, Italy) at +85 °C, 100 rpm for one hour.
BM underwent freezing, lyophilization (Christ Alpha 2-4 LD plus, Germany) and grinding (IKA A11, Germany). Subsequently, the BM was hydrated (0.1 g in 10 mL dH2O) and dried at +121 °C for 20 min (Memmert UF55 oven, Germany). This hydration and drying process was repeated three times prior to centrifugation at 8,000 rpm for 10 min (Centrifuge 5810 R, Eppendorf, Germany). Each drying cycle was followed by the addition of dH2O. The resulting supernatant contained the extracted intraPSH (IPSH).
2.3. Plant Material and Seed Priming
The plant material utilized in this study was the garden pea (Pisum sativum L.) cv. Dunav, developed at the Institute of Field and Vegetable Crops, National Institute of the Republic of Serbia, Novi Sad, Serbia, within the Department of Vegetable and Alternative Crops. The seeds of the chosen pea cultivar were produced at the Rimski Šančevi (N 45°19′, E 19°50′), Serbia, in 2022.
For seed priming, the pea seeds underwent sterilization with 5% sodium hypochlorite solution (NaClO) (Sigma Aldrich, St. Louis, MO, USA) followed by triple rinsing with distilled water. Seed priming was carried out by immersing the pea seeds in dH2O (hydropriming - HP) and aqueous solutions containing PSH extracts at a concentration of 1% (10 mg/mL) (both EPSH and IPSH) in a ratio of 1:5 (w/v) as per Farooq et al. [20] for 6 hours following Arafa et al. [21]. Biopriming involved immersion in EPSH and IPSH solutions from both ITA and SRB fungal strains, while the control group remained unprimed. Subsequently, the seeds were rinsed thoroughly with dH2O and air-dried on filter paper until they regained their initial weight.
2.4. Examination of seed quality and initial growth of pea plants under optimal and drought conditions
2.4.1. Seed Germination Assessment
The study involved a working sample comprising 3 sets of 100 randomly selected seeds each. These seeds, post-priming along with the control group, were placed in plastic boxes sized 240×150 mm, with sterilized sand serving as the growth medium. The experiment consisted of a total of 36 boxes, grouped into two sets, where one set contained 18 boxes that were optimally supplied with water, and the other contained 18 boxes in which drought was simulated using a solution of polyethylene glycol-0.5 MPa solution of polyethylene glycol (PEG-6000) (Sigma Aldrich, St. Louis, MO, USA), as outlined by Tamindžić et al. [22]. Both sets were placed in a germination chamber (Conviron CMP4030, Canada) at +20 °C for eight days following ISTA Rules [23]. Germination energy (GE) (first count) was assessed on the fifth day post-sowing by counting only normal seedlings with well-developed essential structures, while seed germination (SG) and the percentage of abnormal seedlings (AS) were evaluated on the eighth day post-sowing.
2.4.2. Determination of the Shoot and Root Length and the Root/Shoot Ratio
The determination of the shoot length (SL) and the root length (RL), as well as the root/shoot ratio (R/S ratio), was conducted by placing 25 seeds per replicate on filter paper moistened with water (optimal conditions) and a -0.5 MPa PEG solution (simulating water deficit - drought). This setup was then incubated in a germination chamber (Conviron CMP4030, Canada) at 20 °C for eight days. The shoot and root length of 10 normal seedlings was measured on the fifth and eighth day using a ruler [24]. The Root/Shoot ratio (R/S ratio) was calculated on the eighth day based on a formula provided by Bayat et al. [25]:
Root/Shoot ratio= Average root length (mm)/Average shoot length(mm)
2.4.3. Determination of the Fresh and Dry Shoot and Root Biomass Accumulation
On the eighth day following seed placement on filter paper, the shoot fresh weight (SFW) and root fresh weight (RFW) was measured using an analytical balance (Kern 770-13, KERN & Sohn GmbH, Ballingen, Germany). Subsequently, the samples underwent drying in an oven for 24 hours at 80 °C (Heraeus UT 12 Oven, Heraeus Instruments, Germany), after which the shoot dry weight (SDW) and root dry weight (RDW) of the pea seedlings was determined.
2.4.4. Determination of Shoot Elongation Rate (SER) and Root Elongation Rate (RER)
The shoot elongation rate (SER) and root elongation rate (RER) were calculated following the method of Channaoui et al. [26] in three replicates. The formulas used to determine these rates were based on the methodology outlined in the provided research:
where SLE represents the shoot length of, and RLE the root length determined on the fifth day, SLS, RLS show the shoot and root length of seedlings determined on the eighth day, while TE and TS are the periods (days) between the two measurements.
𝑆𝐸𝑅=(𝑆𝐿𝐸−𝑆𝐿𝑆)/(𝑇𝐸−𝑇𝑆)
𝑅𝐸𝑅=(𝑅𝐿𝐸−𝑅𝐿𝑆)/(𝑇𝐸−𝑇𝑆)
2.4.5. Determination of Seed Vigor Index
Seedling Vigor Index (SVI) [27] was determined based on formula:
where SL is seedling length (cm), and FG represents final germination (%).
SVI = SL × FG
2.4.6. Electrolyte Permeability Assay
The impact on membrane permeability was evaluated by assessing electrolyte leakage (EL) following the protocol outlined by Blum and Ebercon [28] as detailed by Farooq et al. [20]. Six leaf discs from pea plants were washed with distilled water, soaked in 6 mL of distilled water for 12 hours, and then the electrical conductivity of the solution (S1) was measured using a laboratory conductometer (HI5321, Hanna Instruments, Woonsocket, RI, USA). Subsequently, the samples were subjected to boiling water for 20 min, cooled to room temperature, and the electrical conductivity of the solution (S2) was recorded. Electrolyte permeability (EL) is calculated as the ratio of S1 to S2.
2.4.7. Determination of Membrane Stability Index
The membrane stability index (MSI), as defined by Sairam [29] and elaborated by Tamindžić et al. [24], was determined through a procedure involving two sets of test tubes. Each set contained 0.10 g of fresh leaf mass and 10 mL of distilled water. One set was subjected to heating at 30 °C for 30 min using a water bath (VIMS elektrik, WKP-14, Republic of Serbia), followed by measurement of electrical conductivity (S1) using a laboratory conductometer (Laboratory Research Grade Benchtop EC/TDS/Salinity/Resistivity Meter — HI5321, Hanna Instruments, Woonsocket, RI, USA). The second set underwent heating at 100 °C for 15 min, after which the electrical conductivity (S2) was determined. The MSI was then calculated using the provided formula:
MSI = (1-C1/C2) × 100
2.4.8. Determination of Relative Water Content
The relative water content (RWC) in pea leaves was determined using the procedure outlined in Farooq et al. [20]. For this test, approximately 0.50 g of fresh leaves mass (Wf) was measured, then the leaves were rinsed and immersed in tubes filled with water until fully saturated, followed by another measurement (Ws). The saturated leaves were then dried in an oven for 24 hours (at 80 °C), and their mass was measured again (Wd). The relative water content was subsequently calculated using the following formula:
RWC = (Wf-Wd)/(Ws-Wd) x 100
2.4.9. Determination of Pea Stress Tolerance Indices
The drought tolerance index (DTI), i.e., plant tolerance to water deficit or stressful conditions (drought), recalculated according to Maiti et al. [30]:
The drought tolerance index (DTI) = Dry plant biomass in control group (g)/Dry plant biomass in treated group (g)
The shoot length stress tolerance index (SLSI) and root length stress tolerance index (RLSI) were determined using the following formulas [25]:
SLSI = Average shoot length in treatment (mm) / Average shoot length in control (mm)
RLSI = Average root length in treatment (mm) / Average root length in control (mm)
2.5. Statistical Analysis
The obtained data were processed statistically using analysis of variance (One-way and Two-way ANOVA), while the significance of the differences between means was tested using the Tukey HSD test at the significance level of p < 0.05. Correlation analysis was performed using Pearson's product-moment correlation. The aforementioned statistical analyses were performed in IBM SPSS statistical software (version 22.0 for Windows) and Statistica software version 12.01 (StatSoft Inc., Tulsa, OK, USA), while the heat map was performed in Microsoft Excel (version 2016).
3. Results
3.1. Effect of Seed Bio-Priming Treatments on Seed Germination and Initial Seedling Growth of Pea
The two-factor analysis of variance (Table 1) showed that the stress factor (S) had a significant effect on all examined parameters at a significance level of p < 0.001, except for the AS parameter (p < 0.01). Also, the treatment factor (T) had a statistically significant effect on all examined parameters at the significance level of p < 0.001, except for the AS and SFW parameters where the significance was manifested at the p < 0.01 level. In addition, the interaction of S × T had a statistically significant effect on all examined parameters (p < 0.001), except for the AS parameter, which did not show significance, while the RWC parameter showed significance at the level of p < 0.01.
Examining the impact of biopriming with PSH derived from S. commune under distinct growth conditions (optimal conditions, drought stress) revealed a notable effect on enhancing pea GE and SG under optimal conditions. Specifically, the IPSH SRB extract demonstrated a 5.50% increase in GE compared to the control, with the highest SG percentage observed in seeds treated with PSH (EPSH and IPSH) isolated from the ITA strain (94.70%) (Figure 1). However, no significant differences were observed in the occurrence of AS among the treatments investigated. Conversely, drought stress induced by PEG solution significantly reduced the GE of pea seeds by 34.50% in the control group, with a 12.60% reduction in SG under drought conditions. Nonetheless, analysis of variance highlighted a positive impact of PSH on GE and SG compared to control and HP (p < 0.05). Particularly, biopriming with IPSH ITA extract led to a 46.00% increase in GE and a 16.00% increase in SG under drought conditions (Figure 1 and Figure 2). Seeds treated with PSH from the SRB strain exhibited 31.60% (EPSH SRB) and 35.20% (IPSH SRB) higher GE and 9.70% and 13.90% higher SG compared to the control group (Figure 1). Notably, the occurrence of AS decreased under both optimal and drought conditions (Figure 1 and Figure 2).
The quality parameters of initial pea growth following biopriming with fungal PSH were examined under optimal conditions and drought stress as well. Biopriming treatments significantly affected pea growth parameters in both conditions, as indicated in Table 2 and 3.
In optimal conditions, the pea SL averaged 55 mm in the control group. However, all tested treatments exhibited a statistically significant increase in SL compared to the control (p < 0.05). The most considerable increase was observed with biopriming using ITA strain extracts (17.30% and 10.60%, respectively). Drought stress resulted in a 52.70% reduction in SL, with all investigated treatments showing a positive effect on this parameter. Nevertheless, under drought conditions, SRB strain extracts demonstrated a superior effect compared to other treatments, leading to a 45% and 43.50% increase in SL, respectively, compared to the control. Additionally, biopriming significantly impacted shoot RL under both optimal and drought conditions compared to control and HP. Notably, the highest values of RL were recorded with biopriming using SRB strain extracts (Table 2). Biopriming of seeds under optimal conditions did not have a statistically significant effect on the SFW compared to the control. However, in conditions of water deficit, treatments with PSH of the SRB strain had a significant effect on this parameter, since it was increased by 37.30% after biopriming with EPSH SRB and by 20.30% after treatment with IPSH SRB compared to the control. When it comes to the RFW, it can be clearly observed that the drought stress had a negative effect on this parameter, where the analysis of the results determined that the reduction was 53.00% compared to the control (Table 2). Also, analysis of variance showed a significant effect of treatment on this parameter both in optimal conditions and in drought stress conditions. All treatments led to an increase in the fresh mass of roots under optimal conditions, only in the treatment with IPSH SRB extracts, the increase was not statistically significant. The highest values of this parameter were recorded after biopriming with EPSH ITA and IPSH SRB extracts (17.80% and 13.90%, respectively) compared to the control. In conditions of water deficit, analysis of variance showed that all treatments had a positive and statistically significant effect on this parameter. Also, a similar pattern of biopriming influence was observed under optimal conditions, where EPSH ITA and IPSH SRB extracts exhibited the best effect on RFW compared to the control (15.50% and 27.50%, respectively) (Table 2).
Similar to previous parameters, drought led to a reduction in SDW by 44.80% and a reduction in RDW by 40.10% compared to the control. Under optimal conditions, the only significant increase in SDW was observed after treatment with EPSH ITA extracts (24.50%) compared to the control. In contrast, under stressful drought conditions, all examined treatments significantly increased the SDW compared to the control. The highest value of SDW was recorded following biopriming with EPSH SRB extracts (0.1434 g), showing a 25.90% increase over the control (Table 2). Additionally, analysis highlighted a significant impact of stress on RDW, with a 40.10% reduction observed in the control. Among the tested treatments, biopriming with EPSH ITA emerged as the most effective, with a 30.20% increase in RDW compared to the control. Moreover, the research findings unequivocally demonstrated that drought stress conditions significantly impacted SVI, resulting in a 53.40% reduction of SVI in the control group (Table 2). Notably, all treatments investigated exhibited a significant effect on this parameter compared to the control, both under optimal and drought-stressed conditions. Under optimal conditions, the treatment with IPSH SRB showed the highest SVI values compared to the control (1694.60), representing a notable 24.90% increase over the control. Moreover, it is evident that PSH derived from the SRB strain, consistent with previous findings, exerted the most beneficial effects under drought conditions. A significant increase of 100.00% (EPSH SRB) and 109.00% (IPSH SRB) relative to the control was observed, emphasizing their efficacy in enhancing seed vigor even under challenging conditions.
Examining the effects of pea biopriming with PSH isolated from S. commune included the monitoring of the SER, RER, R/S ratio, MSI and EL as well (Table 3). The results showed, as with the previous parameters, that the drought significantly reduced the rate of SER and RER in the control. Also, the analysis of variance showed a significant increase in SER in the treatments with EPSH ITA and EPSH SRB (11.40% and 17.60%, respectively) compared to the control under optimal conditions. In drought conditions, all tested treatments significantly increased this parameter, and among them the treatments with extracts of the SRB strain stand out the most (54.70% and 50.80%). Under drought conditions, the PSH of the ITA strain also showed a positive effect on SER, but to a lesser extent than the PSH of the SRB strain. Likewise, the results also revealed a significant effect of treatment on the R/S ratio compared to the control. Under optimal conditions, a notable increase in this parameter was observed following biopriming with EPSH SRB (2.03) compared to the control (1.72), while a significant decrease was noted after biopriming with IPSH SRB (1.00). In water deficit conditions, a significant increase was recorded following HP and biopriming (2.65 ± 0.21, EPSH ITA = 2.63 ± 0.48 and EPSH SRB = 2.83 ± 0.09, respectively)with PSH from the ITA strain.
Conversely, treatments involving PSH from the SRB strain exhibited a significant decrease in the R/S ratio compared to the control. Furthermore, the obtained results showed a significant variation of the MSI and EL parameters between the examined treatments. The results of the research showed that the drought affected the reduction of the MSI (10.30%) and the increase of EL (43.90%) in the control. Also, a similar but reversed pattern was observed in the response to the investigated treatments, so in optimal conditions it was recorded that biopriming with IPSH ITA had the most significant positive effect on these parameters compared to the control. In conditions of water deficit, the best effect was shown by the PSH of the ITA strain compared to the control (Table 3).
The study also monitored RWC, DTI, SLSI, and RLSI (Table 4). Analysis of variance revealed that stress significantly impacted RWC, with a reduction of 11.10% observed in the control group due to drought. The PSH from the SRB strain exhibited the most notable effect on increasing this parameter, with an increase of 3.20% (IPSH SRB) under optimal conditions and 8.10% (IPSH SRB) under drought stress. Moreover, the tested treatments did not significantly affect the DTI under optimal conditions, except for biopriming with EPSH ITA, which showed a significant increase of 13.00% compared to the control. However, under water deficit conditions, all treatments led to a significant increase in the DTI compared to the control. EPSH ITA (23.00%) and EPSH SRB (24.00%) exhibited the most favorable effects among the tested treatments. Regarding the SLSI and RLSI, the treatments had varying effects on these parameters. Under optimal conditions, HP and ITA strain PSH led to a significant increase in the SLSI compared to the control. In drought conditions, a statistically significant increase in the SLSI was observed after biopriming with ITA and SRB strain PSH. Additionally, all examined treatments except HP had a positive effect on the RLSI under both optimal and drought stress conditions (Table 4).
3.2. Correlation analysis
Correlation analysis confirmed the significant effect of biopriming with PSHs isolated from S. commune on seed quality and initial growth of pea under optimal and drought stress conditions (Figure S1 and S2). Results indicated that under optimal conditions, there were highly significant positive correlations between various parameters: the SL and RFW, as well as the RLSI; RL and RER, alongside the SVI; SFW and SDW, coupled with the MSI; RFW and RLSI; RDW and RER, alongside EL. Conversely, significant negative correlations were observed between the SFW and EL, RDW and MSI, RER and MSI, and between the MSI and EL (Figure S1).
A strong and significant correlation (r2> 0.5) was identified among various plant parameters and drought tolerance indices. These included relationships between GE and SG, RFW and RDW, SL and RL, SER and RER, SVI, RWC, SLSI, and RLSI, as well as other key factors. Conversely, negative correlations (r2< 0) were observed between the SER and the R/S ratio, as well as MSI and EL under-drought stress (Figure S2).
4. Discussion
The stages of seed germination and emergence are crucial for achieving an optimal crop composition. Various factors, such as conditions during seed development and subsequent storage, play a significant role in seed germination and vigor, thus greatly impacting crop yields [4]. Moreover, drought stress negatively impacts plant growth and productivity, but seed priming has been shown to mitigate these effects by inducing a primed state in seeds, resulting in early and uniform germination, enhanced growth features, and improved stress response in plants [31]. The effects of drought on plants range from morphological to molecular levels, affecting crop growth, yield, and various physiological processes like cell division and enlargement [32,33]. Therefore, assessing the quality of pea seeds and their capacity to generate healthy shoots under both optimal and drought-stressed conditions is very important.
The observed decrease in GE and SG under drought stress in this study is likely attributed to prolonged imbibition phase III duration and the hindrance of oxygen supply to seeds during germination due to the high viscosity of PEG and limited O2 diffusion [22,34,35]. However, the findings reveal that PSH derived from S. commune, under submerged cultivation, promotes both the GE and SG of pea seeds. Furthermore, they demonstrate a beneficial impact on the initial plant growth and development, under both examined conditions. These results mark a pioneering contribution, as there is no literature data to the best of our knowledge on the utilization of PSH from this fungus in seed biopriming.
Traditionally, seed biopriming involves the application of various bioinoculants, such as plant extracts, beneficial microorganisms (bacteria, algae, fungi, etc.), or biological products (e.g., chitosan, humic acid), known to enhance specialized characteristics to mitigate the adverse effects of abiotic stress and boost yields [2]. For instance, previous studies have shown that biopriming with cattail extract (Typha angustifolia) increased the germination energy of pea seeds by 15.00% under stressful conditions like salinity [36].
In contrast, our research demonstrates an even more significant improvement, with a 31.50% increase in germination energy after biopriming with PSH (IPSH ITA) in drought conditions. Similarly, biopriming with PSH isolated from the microalgae Chlorella vulgaris has been shown to enhance seed germination and initial plant growth parameters in wheat and beans [37], while treatment with EPSH derived from rhizobacteria significantly stimulated seed germination, growth, and yield in wheat [38]. On the other hand, priming of garden peas with calcium chloride (osmopriming) and salicylic acid (hormopriming) significantly influenced all analyzed parameters of seed germination and initial plant growth, except for the proportion of abnormal seedlings, both in optimal and stressful conditions (heat stress) [24]. Compared to EPSH and IPSH isolated from the submerged culture of S. commune, GE under optimal conditions after hormo- and osmopriming was increased by 0.10 to 7.00%, respectively, on average, i.e. under heat stress conditions by 2.30 to 14.30% [24].
Furthermore, Miljaković et al. [4] showed that biopriming of soybean seeds with bacteria B. japonicum and B. megaterium significantly affects the increase of seed quality as well as improved plant growth. The greater SL and RL compared to the control may be due to increased divisions in the apical meristem, which caused an increase in initial growth, as is the case after biopriming of pea seeds with cattail extract [36]. Research by Shaffique et al. [39] showed a positive impact of biopriming with a bacterial strain similar toKlebsiella spp. which produces an EPSH matrix on seed germination, vigor and wheat biomass production under drought conditions, which is in agreement with the results of this study. Also, biopriming of pea seeds with Trichoderma asperellum affected the increase in the SL up to 22%, RL up to 23% and the RDW (60%) and SDW (21%) under optimal conditions after 40 days [40]. Furthermore, Chandra Nayaka et al. [41] in their research indicated a positive effect of bioprimed corn seeds with Trichoderma harzianum on SG and vigor index, which agrees with the obtained results. Ghezal et al. [36] attributed the higher seed germination rate and germination uniformity to metabolic repair during imbibition, as well as the accumulation of secondary metabolites that increase germination. Namely, seed germination, as an essential phase of the plant growth, development, and successful establishment of crops, is threatened in drought conditions due to weaker activation of the necessary hydrolytic enzymes for starch breakdown, solubilization and transport of carbohydrates, which leads to a decrease in vigor and germination index [42]. These comparisons underscore the remarkable potential of PSH from S. commune in seed biopriming, offering promising prospects for enhancing crop productivity, particularly under challenging environmental conditions.
Given that seed quality affects the speed and uniformity of pea emergence, as well as the initial plant growth [24], quality parameters of initial pea growth after biopriming with fungal PSH in optimal conditions and conditions of water deficit - drought were also examined. The qualitative parameter of SVI was closely monitored, serving as a reliable indicator of seed vitality and robustness. Seed vigor index encompasses a multitude of traits, including tolerance to aging, dormancy, viability, rapid germination, and shoot formation, particularly under adverse conditions. It is influenced by various genetic factors and external environmental influences [43]. Moreover, the R/S ratio serves as a crucial parameter indicating the balance between plant tissue dedicated to support functions (root) and that promoting growth (shoots). During the initial stages of the growing season, root growth profoundly impacts the development of shoots, with a higher R/S ratio correlating with increased crop yield. This parameter is heavily influenced by nutrient availability and seed mass [44], making its monitoring essential in these studies. The monitoring of MSI and EL was of great importance, considering that these parameters indicate the impact of oxidative stress in drought conditions and damage to lipid membranes due to the creation of reactive oxygen species, especially in photosynthetic organelles [45]. According to Almeselmani et al. [46], this can lead to direct oxidation of lipid membranes so that their permeability increases and causes ion leakage. Moreover, RWC serves as a crucial indicator of plant water status, reflecting the balance between leaf tissue water supply and transpiration rate [47]. The findings of this study revealed that PSH extracted from two strains of S. commune positively impacted all these parameters, particularly in drought conditions, further affirming the potential of these metabolites as biopriming agents.
Priming is recognized for its capacity to rectify damage induced by seed aging and exposure to abiotic stresses [36]. The observed biostimulatory effects on seed germination and initial growth parameters attributed to isolated EPSH and IPSH can likely be ascribed to the PSH' ability to mitigate water loss. Previous studies on cyanobacterial PSH, like those from C. vulgaris, have demonstrated their capability to alleviate osmotic disturbances in seeds, ensuring adequate moisture for germination [37,48].Moreover, considering the positive correlation between GE and initial growth parameters under drought conditions, it is evident that biopriming with PSH from S. commune holds critical importance for early-stage development (germination) and subsequent biomass increase and yield enhancement during pea development. This aligns with findings by Saha et al. [42] highlighting the increase in drought tolerance and enhanced seed germination under drought conditions attributed to the growth-promoting effects of biopriming. Furthermore, the presented results underscore the significance of biopriming in maintaining and enhancing physiological parameters like MSI and RWC under water deficit conditions. Drought's impact on morphological, physiological, biochemical, and molecular characteristics during germination and emergence phases can significantly impede shoot growth [49]. In this context, priming plays a crucial role in improving germination and plant growth by activating numerous stress-responsive genes, regulating proteins and genes involved in various cellular processes (such as drought-responsive RD1 and RD2 genes of the AP2/ERF TF family as well as P5CSA encoding pyrroline-5- carboxylate synthase A, a key enzyme in proline synthesis), and facilitating the mobilization of reserve substances, among other functions [42,50]. For instance, cyanobacterial filtrates have been found to stimulate the synthesis of bioactive compounds, including cytokinins, auxins, and gibberellins, influencing root and shoot growth in germinated wheat seeds [37,51]. Additionally, studies on biopriming of Trichoderma harzianum wheat seeds under drought conditions have demonstrated its effectiveness in improving MSI and RWC [52], consistent with the findings of this study. Bouremani et al. [53] also noted a significant reduction in RWC due to drought, leading to protein and enzyme denaturation, membrane instability, and metabolic imbalance in cells. Their research on plant growth-promoting rhizobacteria (PGPR) treatments to mitigate drought effects highlighted favorable impacts on RWC and membrane stability, indicating the need for further investigation at cellular and molecular levels to elucidate underlying mechanisms.
5. Conclusions
In conclusion, this investigation into fortifying P. sativum L. seeds in optimal conditions and under drought stress using PSH from S. commune sheds light on a promising avenue for sustainable agriculture in the face of escalating environmental challenges. Our findings underscore the significant potential of IPSH and EPSH, with emphasis on SRB strain as biostimulants for enhancing plant resilience to drought stress.
It is important to emphasize that these results are considered pioneering, because there is no previous literature data on the use of PSH from this fungal species in the context of seed biopriming. The results showed that PSH from S. commune, cultivated in submerged culture, positively contributes to GE and SG of pea seeds, with an emphasis on PSH isolated from the SRB strain. Likewise, PSH have a beneficial effect on the initial growth and development of plants when exposed to drought stress, since all tested treatments led to a significant increase in the DTI compared to the control, and the best effect was shown by EPSH ITA (23.00%) and EPSH SRB (24.00%).
Significant improvement of the tested parameters, as well as a positive correlation between GE and initial growth parameters under drought conditions indicate that the application of these PSH may have long-term benefits for the plant. Furthermore, the utilization of PSH extracted from S. commune through biopriming might be crucial for triggering the initial phase of growth, specifically seed germination, as well as for increasing biomass and yield during subsequent stages of pea maturation.
Further research is warranted to explore the broader applicability of S. commune PSH across different plant species and environmental contexts. Additionally, investigations into the molecular mechanisms underlying the interaction between these PSH and plant physiology will deepen our understanding and facilitate the development of tailored strategies for enhancing crop resilience.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Figure S1: Heat map based on correlation analysis of investigated parameters of vegetable peas in optimal conditions; Figure S2: Heat map based on correlation analysis of investigated parameters of vegetable peas under drought.
Author Contributions
Conceptualization, M.K., G.T., J.M., and M.R..; methodology, J.M., G.T., and M.R.; software, J.M. and N.K.; validation, M.K. and G.T.; formal analysis, J.M., G.T., and M.R..; investigation, J.M., G.T., and M.R.; resources, J.M. and M.R.; data curation, J.M.; writing—original draft preparation, J.M., G.T. and M.R.; writing—review and editing, M.K., G.T., J.M., M.R., M.I., and N.K.; visualization, J.M.; supervision, M.K. and G.T.; funding acquisition, M.K. and G.T.; All authors have read and agreed to the published version of the manuscript.
Funding
The Ministry of Science, Technological Development, and Innovation of the Republic of Serbia supported this study through Grants No. 451-03-66/2024-03/200125, 451-03-65/2024-03/200125, and 451-03-66/2024-03/200032.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy.
Acknowledgments
Prof. Maja Karaman and other authors express their gratitude to prof. Alessandra Zambonelli and prof. Federica Piattoni from the Department of Agricultural and Food Sciences, University of Bologna, Italy for the help of obtaining the ITA strain of S. commune near the city of Bologna during the Erasmus Mundus Exchange Program in 2016. We warmly thank the Center of Excellence for Legumes, Institute of Field and Vegetable Crops, Novi Sad, Serbia, for supporting the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lesk, C.; Anderson, W.B.; Rigden, A.J.; Coast, O.; Jägermeyr, J.; McDermid, S.P.; Davis, K.F.; Konar, M. Compound heat and moisture extreme impacts on global crop yields under climate change. Nat. Rev. Earth Environ. 2022, 3, 872–889. [Google Scholar] [CrossRef]
- Chakraborti, S.; Bera, K.; Sadhukhan, S.; Dutta, P. Bio-priming of seeds: Plant stress management and its underlying cellular, biochemical and molecular mechanisms. Plant Stress 2022, 3, 100052. [Google Scholar] [CrossRef]
- Verma, P.; Hiremani, N.S.; Gawande, S.P.; Sain, S.K.; Nagrale, D.T. , Narkhedkar, N.G., Prasad, Y.G. Modulation of plant growth and antioxidative defense system through endophyte biopriming in cotton (Gossypium spp.) and non-host crops. Heliyon 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Miljaković, D.; Marinković, J.; Tamindžić, G.; Đorđević, V.; Tintor, B.; Milošević, D.; Ignjatov, M.; Nikolić, Z. Bio-priming of soybean with Bradyrhizobium japonicum and Bacillus megaterium: Strategy to improve seed germination and the initial seedling growth. Plants 2022, 11, 1927. [Google Scholar] [CrossRef]
- Metwally, R.A.; Abdelhameed, R.E.; Soliman, S.A.; Al-Badwy, A.H. Potential use of beneficial fungal microorganisms and C-phycocyanin extract for enhancing seed germination, seedling growth and biochemical traits of Solanum lycopersicum L. BMC Microbiol. 2022, 22, 108. [Google Scholar] [CrossRef]
- Deshmukh, A.J.; Jaiman, R.S.; Bambharolia, R.P.; Patil, V.A. Seed biopriming-a review. Int. J. Econ. Plants 2020, 7, 038–043. [Google Scholar] [CrossRef]
- Rasskazova, I.; Kirse-Ozolina, A. Field pea Pisum sativum L. as a perspective ingredient for vegan foods: A review. Food Sci. 2020, 35, 125–131. [Google Scholar]
- Dahl, W.J.; Foster, L.M.; Tyler, R.T. Review of the health benefits of peas (Pisum sativum L.). Br. J. Nutr. 2012, 108, S3–S10. [Google Scholar] [CrossRef] [PubMed]
- Tulbek, M.C.; Wang, Y.L.; Hounjet, M. Pea—A sustainable vegetable protein crop. In: Sustainable Protein Sources, Academic Press, Chapter 9, pp. 143-–162. 2024. [CrossRef]
- Wu, D.T.; Li, W.X.; Wan, J.J.; Hu, Y.C.; Gan, R.Y.; Zou, L. A comprehensive review of pea (Pisum sativum L.): chemical composition, processing, health benefits, and food applications. Food 2023, 12, 2527. [Google Scholar] [CrossRef]
- Al-Turki, A.; Murali, M. Recent advances in PGPR-mediated resilience toward interactive effects of drought and salt stress in plants. Front. Microbiol. 2023, 14, 1214845. [Google Scholar] [CrossRef]
- Ţura, D.; Wasser, S.P.; Zmitrovich, I.V. Wood-inhabiting fungi: Applied aspects. In: Fungi: and Management Strategies, CRC Press, 2018, Chapter 12, pp. 245–292.
- Mišković, J.; Rašeta, M.; Krsmanović, N.; Karaman, M. Update on mycochemical profile and selected biological activities of genus Schizophyllum Fr. 1815. Microbiol. Res. 2023, 14, 409–429. [Google Scholar] [CrossRef]
- Abd Razak, D.L.; Abd Ghani, A.; Lazim, M.I.M.; Khulidin, K.A.; Shahidi, F.; Ismail, A. Schizophyllum commune (Fries) mushroom: A review on its nutritional components, antioxidative and anti-inflammatory properties. Curr. Opin. Food Sci. 2024, 56, 101129. [Google Scholar] [CrossRef]
- Krause, K.; Jung, E.M.; Lindner, J.; Hardiman, I.; Poetschner, J.; Madhavan, S.; Matthäus, C.; Kai, M.; Menezes, R.C.; Popp, J.; Svatoš, A.; Koth, E. Response of the wood-decay fungus Schizophyllum commune to co-occurring microorganisms. PLoS One 2020, 15, e0232145. [Google Scholar] [CrossRef]
- Rangkhawong, P.; Issaranuwat, P.; Gaensakoo, R. Mycelial fermentation in submerged culture of Schizophyllum commune and its properties. SCJMSU 2014, 33, 420–428. [Google Scholar]
- Mišković, J.; Karaman, M.; Rašeta, M.; Krsmanović, N.; Berežni, S.; Jakovljević, D.; Piattoni, F.; Zambonelli, A.; Gargano, M.L.; Venturella, G. Comparison of two Schizophyllum commune strains in production of acetylcholinesterase inhibitors and antioxidants from submerged cultivation. J. Fungi 2021, 7, 115. [Google Scholar] [CrossRef]
- Kumar, A.; Bharti, A.K.; Bezie, Y. Schizophyllum commune: A fungal cell-factory for production of valuable metabolites and enzymes. BioResources 2022, 17, 5420–5436. [Google Scholar] [CrossRef]
- Chen, C.-C.; Nargotra, P.; Kuo, C.-H.; Liu, Y.-C. High-molecular-weight exopolysaccharides production from Tuber borchii cultivated by submerged fermentation. Int. J. Mol.Sci. 2023, 24, 4875. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Aziz, T.; Basra, S.M.A.; Cheema, M.A.; Rehman, H. Chilling tolerance in maize hybrid induced by seed priming with salicylic acid. J. Agron. Crop Sci. 2008, 194, 161–168. [Google Scholar] [CrossRef]
- Arafa, S.A.; Attia, K.A.; Niedbała, G.; Piekutowska, M.; Alamery, S.; Abdelaal, K.; Attallah, S.Y. Seed priming boost adaptation in pea plants under drought stress. Plants 2021, 10, 2201. [Google Scholar] [CrossRef]
- Tamindžić, G.; Červenski, J.; Milošević, D.; Vlajić, S.; Nikolić, Z.; Ignjatov, M. The response of garden pea cultivars to simulated drought. Acta Agric. Serb. 2021, 26, 167–173. [Google Scholar] [CrossRef]
- ISTA. International Rules for Seed Testing; Seed Science and Technology: Zurich, Switzerland, 2022. [Google Scholar]
- Tamindžić, G.; Ignjatov, M.; Miljaković, D.; Červenski, J.; Milošević, D.; Nikolić, Z.; Vasiljević, S. Seed priming treatments to improve heat stress tolerance of garden pea (Pisum sativum L.). Agriculture 2023, 13, 439. [Google Scholar] [CrossRef]
- Bayat, M.; Zargar, M.; Murtazova, K.M.S.; Nakhaev, M.R.; Shkurkin, S.I. Ameliorating seed germination and seedling growth of nano-primed wheat and flax seeds using seven biogenic metal-based nanoparticles. Agronomy 2022, 12, 811. [Google Scholar] [CrossRef]
- Channaoui, S.; El Idrissi, I.S.; Mazouz, H.; Nabloussi, A. Reaction of some rapeseed (Brassica napus L.) genotypes to different drought stress levels during germination and seedling growth stages. OCL 2019, 26, 23. [Google Scholar] [CrossRef]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor determination in soybean seed by multiple criteria. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Sairam, R.K. Effect of moisture-stress on physiological activities of two contrasting wheat genotypes. Indian J. Exp. Biol. 1994, 32, 594–594. [Google Scholar]
- Maiti, R.K.; de la Rosa-Ibarra, M.; Sandoval, N.D. Genotypic variability in glossy sorghum lines for resistance to drought, salinity and temperature stress at the seedling stage. J. Plant Physiol. 1994, 143, 241–244. [Google Scholar] [CrossRef]
- Aswathi, K.R.; Kalaji, H.M.; Puthur, J.T. Seed priming of plants aiding in drought stress tolerance and faster recovery: A review. Plant Growth Regul. 2022, 97, 235–253. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants' physio-biochemical and phyto-hormonal responses to alleviate the adverse effects of drought stress: A comprehensive review. Plants 2022, 11, 1620. [Google Scholar] [CrossRef]
- Braccini, A.L.; Ruiz, H.A.; Braccini, M.C.L.; Reis, M.S. Germinaçáo e vigor de sementes de soja sob estresse hidrico induzido por soluções de cloreto de sódio, mannitol e polietilenglicol. Rev. Bras. Sementes 1996, 18, 10–16. [Google Scholar] [CrossRef]
- Pereira, I.C.; Catão, H.; Caixteta, F. Seed physiological quality and seedling growth of pea under water and salt stress. Rev. Bras. Eng. Agric. Ambient. 2020, 24, 95–100. [Google Scholar] [CrossRef]
- Ghezal, N.; Rinez, I.; Sbai, H.; Saad, I.; Farooq, M.; Rinez, A.; Haouala, R. Improvement of Pisum sativum salt stress tolerance by bio-priming their seeds using Typha angustifolia leaves aqueous extract. S. Afr. J. Bot. 2016, 105, 240–250. [Google Scholar] [CrossRef]
- Dalal, S.R.; Hussein, M.H.; El-Naggar, N.E.A.; Mostafa, S.I.; Shaaban-Dessuuki, S.A. Characterization of alginate extracted from Sargassum latifolium and its use in Chlorella vulgaris growth promotion and riboflavin drug delivery. Sci. Rep. 2021, 11, 16741. [Google Scholar] [CrossRef] [PubMed]
- Prasad, J.; Dey, R.; Raghuwanshi, R. Exopolysaccharide-producing rhizospheric bacteria enhance yield via promoting wheat (Triticum aestivum L.) growth at early stages. Microbiology 2022, 91, 757–769. [Google Scholar] [CrossRef]
- Shaffique, S.; Khan, M.A.; Wani, S.H.; Imran, M.; Kang, S.M.; Pande, A.; Lee, I.J. Biopriming of maize seeds with a novel bacterial strain SH-6 to enhance drought tolerance in South Korea. Plants 2022, 11, 1674. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Dwivedi, P. Trichoderma-induced promotion of nitrogen use efficiency is mediated by nitric oxide generation leading to improved growth and yield in pea (Pisum sativum L.) plants. J. Plant Growth Regul. 2022, 42, 6397–6412. [Google Scholar] [CrossRef]
- Chandra Nayaka, S.; Niranjana, S.R.; Uday Shankar, A.C.; Niranjan Raj, S.; Reddy, M.S.; Prakash, H.S.; Mortensen, C.N. Seed biopriming with novel strain of Trichoderma harzianum for the control of toxigenic Fusarium verticillioides and fumonisins in maize. Arch. Phytopathol. Pflanzenschutz. 2010, 43, 264–282. [Google Scholar] [CrossRef]
- Saha, D.; Choyal, P.; Mishra, U.N.; Dey, P.; Bose, B.; Prathibha, M.D.; Singhal, R.K. Drought stress responses and inducing tolerance by seed priming approach in plants. Plant Stress 2022, 4, 100066. [Google Scholar] [CrossRef]
- Reed, R.C.; Bradford, K.J.; Khanday, I. Seed germination and vigor: Ensuring crop sustainability in a changing climate. Heredity 2022, 128, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Mašková, T.; Herben, T. Root: shoot ratio in developing seedlings: How seedlings change their allocation in response to seed mass and ambient nutrient supply. Ecol. Evol. 2018, 8, 7143–7150. [Google Scholar] [CrossRef]
- Sakya, A.T.; Sulistyaningsih, E.; Indradewa, D.; Purwanto, B.H. Physiological characters and tomato yield under drought stress. IOP Conf. Ser. Earth Environ. Sci. 2018, 200, 012043. [Google Scholar] [CrossRef]
- Almeselmani, M.; Abdullah, F.; Hareri, F.; Naaesan, M.; Ammar, M.A.; ZuherKanbar, O. Effect of drought on different physiological characters and yield component in different varieties of Syrian durum wheat. J. Agric. Sci. 2011, 3, 127. [Google Scholar] [CrossRef]
- Soltys-Kalina, D.; Plich, J.; Strzelczyk-Żyta, D.; Śliwka, J.; Marczewski, W. The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of ‘Katahdin’-derived potato cultivars. Breed. Sci. 2016, 66, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, G.; Moreno-Casasola, P.; Barrera, O. Interaction between algae and seed germination in tropical dune slack species: a facilitation process. Aquat. Bot. 1998, 60, 409–416. [Google Scholar] [CrossRef]
- Yigit, N.; Sevik, H.; Cetin, M.; Kaya, N. Determination of the effect of drought stress on the seed germination in some plant species.In: Water Stress in Plants, Rahman, I.M.M.; Begum, M.Z.A.; Hasegawa, H. (Eds.), InTechOpen, 2016, 43, 62. [CrossRef]
- Kubala, S.; Wojtyla, Ł.; Quinet, M.; Lechowska, K.; Lutts, S.; Garnczarska, M. Enhanced expression of the proline synthesis gene P5CSA in relation to seed osmopriming improvement of Brassica napus germination under salinity stress. J. Plant Physiol. 2015, 183, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Stirk, W.A.; Ördög, V.; Van Staden, J. Identification of the cytokinin isopentenyladenine in a strain of Arthronema africanum (Cyanobacteria). J. Phycol. 1999, 35, 89–92. [Google Scholar] [CrossRef]
- Shukla, N.; Awasthi, R.P.; Rawat, L.; Kumar, J. Seed biopriming with drought tolerant isolates of Trichoderma harzianum promote growth and drought tolerance in Triticum aestivum. Ann. Appl. Biol. 2015, 166, 171–182. [Google Scholar] [CrossRef]
- Bouremani, N.; Cherif-Silini, H.; Silini, A.; Bouket, A.C.; Luptakova, L.; Alenezi, F.N.; Belbahri, L. Plant growth-promoting rhizobacteria (PGPR): A rampart against the adverse effects of drought stress. Water 2023, 15, 418. [Google Scholar] [CrossRef]
Figure 1.
Graphic representation of the effect of biopriming with PSHs isolated from both strains S. commune (SRB, ITA) under optimal conditions on the parameters of pea seed quality testing.
Figure 1.
Graphic representation of the effect of biopriming with PSHs isolated from both strains S. commune (SRB, ITA) under optimal conditions on the parameters of pea seed quality testing.
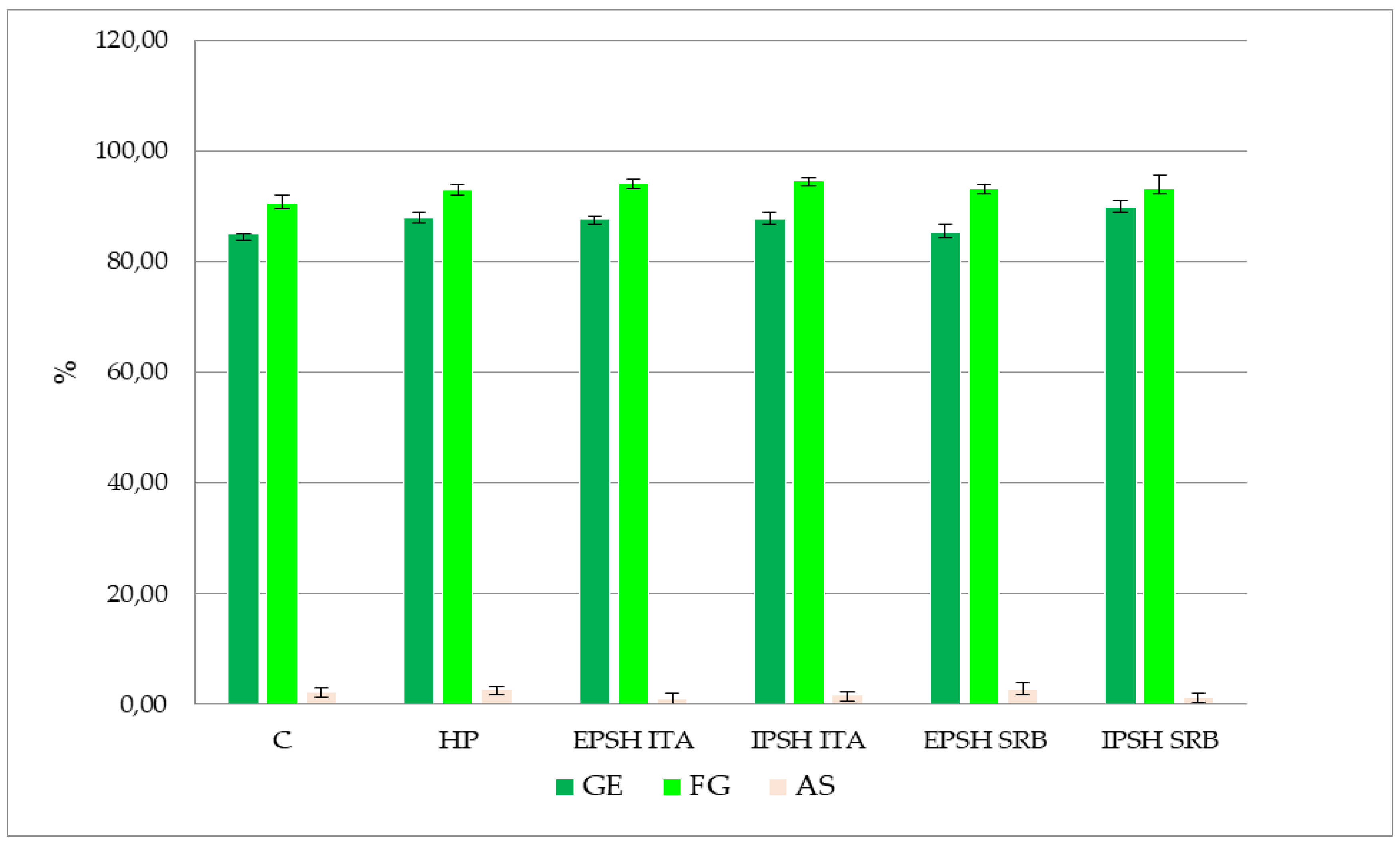
Figure 2.
Graphic representation of the effect of biopriming with PSHs isolated from S. commune under drought conditions on the parameters of pea seed quality testing.
Figure 2.
Graphic representation of the effect of biopriming with PSHs isolated from S. commune under drought conditions on the parameters of pea seed quality testing.
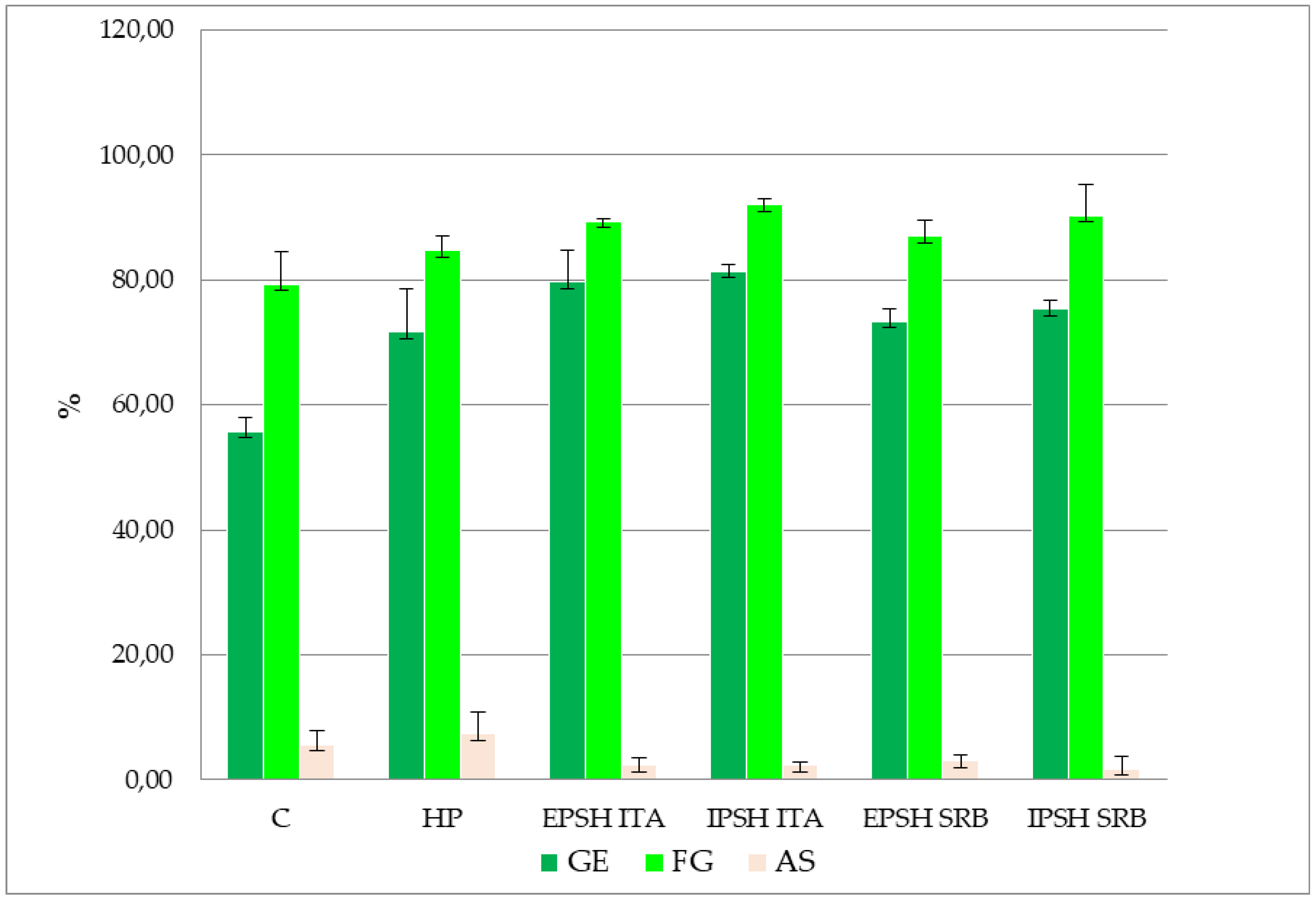

Table 1.
Two-factor analysis of variance of quality parameters of pea after biopriming with PSH isolated from S. commune under optimal and drought stress conditions.
Table 1.
Two-factor analysis of variance of quality parameters of pea after biopriming with PSH isolated from S. commune under optimal and drought stress conditions.
| Trait | S | Т | S × Т |
| Germination energy (GE) | *** | *** | *** |
| Seed Germination (SG) | *** | *** | *** |
| Abnormal seedlings (AS) | ** | ** | NS |
| Shoot length (SL) | *** | *** | *** |
| Root length (RL) | *** | *** | *** |
| Shoot fresh weight (SFW) | *** | ** | *** |
| Root fresh weight (RFW) | *** | *** | *** |
| Shoot dry weight (SDW) | *** | *** | *** |
| Root dry weight (RDW) | *** | *** | *** |
| Shoot elongation rate (SER) | *** | *** | *** |
| Root elongation rate (RER) | *** | *** | *** |
| Seedling vigor index (SVI) | *** | *** | *** |
| Membrane stability index (MSI) | *** | *** | *** |
| Electrolyte leakage (EL) | *** | *** | *** |
| Relative water content (RWC) | *** | *** | ** |
| Drought tolerance index (DTI) | *** | *** | *** |
| Shoot length tolerance index (SLSI) | *** | *** | *** |
| Root length tolerance index (RLSI) | *** | *** | *** |
*p < 0,05; **p < 0,01; ***p < 0,001; NS – not significant.
Table 2.
Effect of biopriming with PSH isolated from S. commune on seedling quality parameters (SH, RH, SFW, RFW, SDW, RDW and SVI) in optimal and stressful conditions (drought).
Table 2.
Effect of biopriming with PSH isolated from S. commune on seedling quality parameters (SH, RH, SFW, RFW, SDW, RDW and SVI) in optimal and stressful conditions (drought).
| Тreatment* | SL** (mm) | RL (mm) | SFW (g) | RFW (g) | SDW (g) | RDW (g) | SVI |
|---|---|---|---|---|---|---|---|
| Optimal conditions | |||||||
| C | 55.00 ± 1.50d | 94.70 ± 2.90d | 2.37 ± 0.17 ab | 2.08 ± 0.04 c | 0.2064 ± 0.009 bc | 0.1820 ± 0.001 c | 1356.80 ± 7.30e |
| HP | 59.0 ± 0.90bc | 97.0 ± 2.0 d | 2.48 ± 0.11 b | 2.16 ± 0.02 b | 0.2421 ± 0.015 ab | 0.1785 ± 0.002 c | 1450.80 ± 21.50 d |
| EPSH ITA | 64.50 ± 0.09 a | 114.5 ± 3.30b | 2.76 ± 0.11 b | 2.45 ± 0.02 a | 0.2569 ± 0.023 a | 0.1831 ± 0.001 c | 168.70 ± 39.70ab |
| IPSH ITA | 60.80 ± 0.70b | 105.7 ± 3.3 c | 2.48 ± 0.06 b | 2.37 ± 0.05 a | 0.2092 ± 0.005bc | 0.1847 ± 0.001 c | 1576.1 ± 30.6 c |
| EPSH SRB | 57.20 ± 0.70cd | 115.90 ± 1.10b | 1.98 ± 0.16 a | 2.20 ± 0.01 b | 0.1844 ± 0.008 c | 0.2020 ± 0.005 a | 1615.50 ± 24.00bc |
| IPSH SRB | 57.60 ± 2.20bcd | 123.90 ± 1.50a | 2.03 ± 0.25 a | 2.13 ± 0.02 bc | 0.1957 ± 0.021 c | 0.1950 ± 0.002 b | 1694.60 ± 29.80a |
| Drought conditions | |||||||
| C | 26.00 ± 0.50d | 53.50 ± 4.30e | 1.18 ± 0.12 c | 1.42 ± 0.03 d | 0.1139 ± 0.003 c | 0.1091 ± 0.005 c | 632.00 ± 73.40d |
| HP | 27.30 ± 1.30d | 72.50 ± 3.50d | 1.37 ± 0.05 bc | 1.61 ± 0.06 bc | 0.1288 ± 0.006 b | 0.1291 ± 0.002 b | 844.70 ± 9.50c |
| EPSH ITA | 33.90 ± 0.90b | 88.80 ± 4.20c | 1.35 ± 0.07 bc | 1.64 ± 0.01 b | 0.1315 ± 0.002 b | 0.1420 ± 0.006 a | 1096.5 ± 9.50c |
| IPSH ITA | 30.80 ± 1.10c | 87.30 ± 2.90c | 1.31 ± 0.04 bc | 1.81 ± 0.04 a | 0.1324 ± 0.003 b | 0.1360 ± 0.002 ab | 1087.40 ± 44.10b |
| EPSH SRB | 37.70 ± 0.90a | 119.80 ± 1.90a | 1.62 ± 0.10 a | 1.56 ± 0.02 bc | 0.1434 ± 0.002 bc | 0.1325 ± 0.002 ab | 1369.80 ± 42.60a |
| IPSH SRB | 37.30 ± 0.50a | 108.9 ± 2.20b | 1.42 ± 0.09 ab | 1.54 ± 0.03 c | 0.1329 ± 0.003 b | 0.1332 ± 0.006 ab | 1320.90 ± 88.90a |
* Data are presented as the mean of three replicates ± standard deviation. Differences between treatments were analyzed using ANOVA, as well as the post-hoc Tukey HSD test (p < 0.05). Each column (optimal conditions and drought) has a different superscript indicating statistical significance. ** SL – shoot length, RL – root length, SFW – shoot fresh weight, RFW – root fresh weight, SDW – shoot dry weight, RDW – root dry weight, SVI – seedling vigor index.
Table 3.
Effect of biopriming with PSH isolated from S. commune on seedling quality parameters (SER, RER, R/S ratio, MSI and EL) in optimal and stressful conditions (drought).
Table 3.
Effect of biopriming with PSH isolated from S. commune on seedling quality parameters (SER, RER, R/S ratio, MSI and EL) in optimal and stressful conditions (drought).
| Treatment* | SER** | RER | R/S ratio | MSI (%) | EL (%) |
|---|---|---|---|---|---|
| Optimal conditions | |||||
| C | 20.21 ± 0.41c | 9.19 ± 1.10b | 1.72 ± 0.10b | 81.00 ± 0.18b | 19.00 ± 0.18b |
| HP | 10.21 ± 0.22c | 9.14 ± 0.91b | 1.64 ± 0.06b | 81.88 ± 0.52b | 18.12 ± 0.52b |
| EPSH ITA | 11.37 ± 0.21ab | 10.94 ± 1.22b | 1.78 ± 0.06b | 81.78 ± 0.38b | 18.22 ± 0.38b |
| IPSH ITA | 10.47 ± 0.37bc | 10.80 ± 1.36b | 1.74 ± 0.05b | 83.99 ± 0.22a | 16.01 ± 0.22c |
| EPSH SRB | 12.01 ± 0.39a | 14.78 ± 0.47a | 2.03 ± 0.04a | 71.07 ± 0.51c | 28.93 ± 0.51a |
| IPSH SRB | 10.07 ± 0.62c | 13.92 ± 0.65a | 1.00 ± 0.07c | 71.67 ± 1.06c | 28.33 ± 1.06a |
| Drought conditions | |||||
| C | 5.91 ± 0.30c | 5.73 ± 1.50 d | 2.05 ± 0.38 b | 72.66 ± 1.04 b | 27.34 ± 1.04a |
| HP | 4.40 ± 0.41d | 6.83 ± 1.25 d | 2.65 ± 0.21 a | 72.48 ± 0.79 b | 27.52 ± 0.79a |
| EPSH ITA | 7.03 ± 0.36b | 13.31 ± 1.73 c | 2.63 ± 0.48 a | 81.99 ± 0.93 a | 18.01 ± 0.93b |
| IPSH ITA | 6.09 ± 0.35c | 11.50 ± 1.17c | 2.83 ± 0.09a | 82.37 ± 0.81a | 17.63 ± 0.81b |
| EPSH SRB | 9.14 ± 0.28a | 22.23 ± 1.69a | 0.92 ± 0.02c | 71.93 ± 0.67b | 28.07 ± 0.67a |
| IPSH SRB | 8.91 ± 0.25a | 18.23 ± 0.58b | 1.00 ± 0.11c | 72.15 ± 1.04b | 27.85 ± 1.04a |
* Data are presented as the mean of three replicates ± standard deviation. Differences between treatments were analyzed using ANOVA, as well as the post-hoc Tukey HSD test (p < 0.05). Each column (optimal conditions and drought) has a different superscript indicating statistical significance. ** SER – shoot elongation rate, RER – root elongation rate, R/S ratio – root/shoot ratio, MSI – membrane stability index, EL – electrolyte leakage.
Table 4.
Effect of biopriming with PSH isolated from S. commune on seedling quality parameters (RWC, DTI, SLSI and RLSI) in optimal and stressful conditions (drought).
Table 4.
Effect of biopriming with PSH isolated from S. commune on seedling quality parameters (RWC, DTI, SLSI and RLSI) in optimal and stressful conditions (drought).
| Treatment* | RWC** | DTI | SLSI | RLSI |
|---|---|---|---|---|
| Optimal condition | ||||
| C | 81.80 ± 0.70bc | 1.00 ± 0.00 b | 100.00 ± 0.00c | 100.00 ± 0.00d |
| HP | 81.90 ± 0.30bc | 1.08 ± 0.03 ab | 107.30 ± 1.80b | 102.50 ± 1.70cd |
| EPSH ITA | 81.30 ± 1.20c | 1.13 ± 0.06 a | 117.30 ± 2.00a | 121.20 ± 6.70ab |
| IPSH ITA | 82.30 ± 0.50bc | 1.01 ± 0.03 b | 110.70 ± 3.80ab | 111.70 ± 4.40bc |
| EPSH SRB | 83.10 ± 1.10ab | 0.99 ± 0.02 b | 104.00 ± 3.40bc | 122.60 ± 4.60ab |
| IPSH SRB | 84.40 ± 0.50a | 1.01 ± 0.04 b | 104.70 ± 2.10bc | 131.00 ± 3.20a |
| Drought conditions | ||||
| C | 72.70 ± 1.10bc | 1.00 ± 0.00 c | 100.00 ± 0.00d | 100.00 ± 0.00d |
| HP | 71.80 ± 0.60c | 1.16 ± 0.03 b | 105.20 ± 6.00d | 135.90 ± 8.80cd |
| EPSH ITA | 75.30 ± 0.20ab | 1.23 ± 0.01 a | 130.40 ± 0.80b | 167.20 ± 22.20bc |
| IPSH ITA | 74.20 ± 0.8 bc | 1.20 ± 0.01 ab | 118.60 ± 5.00c | 164.20 ± 18.10bc |
| EPSH SRB | 75.80 ± 1.90ab | 1.24 ± 0.01 a | 144.90 ± 2.60a | 224.80 ± 15.40a |
| IPSH SRB | 78.60 ± 1.80a | 1.19 ± 0.03 ab | 143.40 ± 4.30a | 204.50 ± 18.06 ab |
* Data are presented as the mean of three replicates ± standard deviation. Differences between treatments were analyzed using ANOVA, as well as the post-hoc Tukey HSD test (p < 0.05). Each column (optimal conditions and drought) has a different superscript indicating statistical significance. ** RWC – relative water content, DTI – drought tolerance index, SLSI –shoot length tolerance index, RLSI – root length tolerance index.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



