
Submitted:
19 April 2024
Posted:
19 April 2024
You are already at the latest version
Abstract
Epithelial barrier damage plays a central role in the development and maintenance of allergic inflammation.
The rise in airways epithelial barrier permeability alters the tissue homeostasis and allows the allergen and other external agents penetration.
Different factors contribute to barrier impairment, such as eosinophilic infiltrate and allergens proteases action: the eosinophilic cationic proteins and the allergens' proteolytic activity have a central role in contributing to epithelial damage.
In the airways, the allergens proteases degrade the epithelial junctional proteins allowing the allergen penetration and the uptake by dendritic cells.
The increase in allergen-immune system interactions induce the release of the alarmins and the activation of type 2 inflammatory pathways causing, or worsening, the main symptoms at skin, bowel and respiratory levels.
We aim to highlight the molecular mechanisms underlying allergenic proteases-induced epithelial barrier damage and the role of immune response in allergic asthma onset, maintenance and progression.
We further will explore clinical and radiological potential biomarkers of airways remodeling in allergic asthma patients.
Keywords:
allergic asthma
; epithelial barrier damage
; protease allergen
; type 2 inflammation
; airway remodeling
1. Introduction
Epithelial barrier damage is a crucial feature of inflammatory allergic diseases. The epithelium plays a double protective action: mechanical, through maintaining skin and mucosal barrier integrity, and immunological, through the action of a rich set of molecules that ensure the immune tolerance.
The epithelial barrier is a structured entity in which cell-cell adhesion complexes ensure integrity and effectiveness [1].
Tight Junctions (TJs) include transmembrane proteins of the claudin family, occludin, tricellulin, junctional adhesion molecules, and cytoplasmic proteins (such as the Zonula Occludens (ZO)-1 ZO-2, ZO-3).
Adherens junctions are composed of cadherin-catenin complexes and they act as key regulators of paracellular permeability [2].
Desmosomes provide mechanical stability and hemidesmosomes contribute to the epithelial layer-basal membrane attachment.
In normal conditions, a functional physical barrier contributes to the regulation of epithelial permeability, cell proliferation and differentiation. The loss of barrier integrity increases the exposure to environmental, allergic and toxic substances, breaking the immune tolerance and inducing the activation of different pro-inflammatory pathways.
Epithelial barrier damage characterizes different inflammatory diseases such as asthma, Chronic Rhinosinusitis with Nasal Polyps (CRSwNP), Eosinophilic Esophagitis (EoE) and Atopic Dermatitis (AD); different mechanisms could contribute to the barrier dysfunction.
In allergic diseases, the disruption of the epithelial barrier is associated with TJ defects and with a reduction of the adherence junctions and desmosomes number [3,4,5,6].
Zonulin is a regulator of epithelial and endothelial barrier function. It regulates intestinal permeability through disrupting tight junctions. Defective epithelial barrier function is a hallmark of airway inflammation in asthma [3].
Several susceptibility genes have been associated with epithelial barrier differentiation and homeostasis [9,10]. Structural airways remodeling signs have been found in genetically predisposed asthma children [11,12,13].
Moreover, the environmental context with the action of viruses [14], pollutants [15], cigarette smoke [16] and allergens play a central role in epithelial injury. Moreover, industrialization and consumption of highly processed food, can contribute to altering the gut microbiota and the intestinal barrier, increasing the susceptibility to allergic sensitization [17].
Several allergens, or allergens components, such as the House-Dust Mite (HDM) Der p1, have shown the ability to disrupt the TJs through both direct and indirect proteolytic activity [18].
The increase in epithelium permeability leads to Type 2 (T2) cytokines production and eosinophils activation and proliferation in the airways. Both T2 cytokines and eosinophil mediators interfere with TJs, contributing to the lack of barrier response. In asthma, the inflammatory processes start from the release of alarmins; the entity of cytokines’ release correlates with the clinical symptoms, the disease severity and the airway remodeling process [19] (Figure 1).
We aim to explore the molecular mechanisms underlying allergenic proteases-induced damage in allergic asthma onset, maintenance and progression. Further, we highlight the role of both epithelial barrier dysfunction and immune response in airways remodeling, exploring potential clinical and radiological biomarkers and the role of different therapeutic options.
2. Molecular Mechanisms in Allergic Asthma: The Allergens Proteases
Allergens proteases are proteolytic enzymes with a primary role in the pathogenesis of respiratory allergies, facilitating the allergen-host interactions and promoting the development of allergic sensitization.
Allergens and pathogens with proteolytic activity can intrinsically overcome the host tolerance, activating various immunological pathways. Proteases enhance antigen-presenting cells in airways, inducing specific Immunoglobulin-E (IgE) production, eosinophils recruitment and inflammatory mediators release in airways, skin or other barrier tissues.
Proteases could be released as enzymatically inactive zymogens, requiring additional adjuvants for the activation, or they could show independent proteolytic activity, such as HDM [20,21,22,23].
The HDM major protease Der p 1 is a cysteine protease which has shown self-maturation capacity in acidic ambient [24]; moreover, it has demonstrated to promote the maturation of other HDM proteases, such as requiring enzymatic activation-HDM serine protease Der p 3, Der p 6 and Der p 9 [25].
Beyond activating factors, specific and non-specific protease inhibitors also participate in the regulation of proteolytic activity, ensuring the maintenance of tissue homeostasis [23].
Although HDM Der p 1 has been the first allergen protease to be characterized [26], more recently, different HDM, cockroaches’ [27], fungal [28] and plant proteases have been described [29].
Based on the catalysis mechanism, according to the location of the cleavage site and the nature of their active site residues, proteases have been classified into five classes: aspartic, metallo, cysteine, serine and threonine proteases [30].
Although all five classes of proteases are found in the human genome [31], only aspartic, cysteine and serine proteases have been identified as allergens [32].
Most of cysteine proteases share structural homologies with Der p 1, having cysteine-histidine-asparagine as active site residues; while serine protease allergens are structurally similar to trypsin with serine-histidine-aspartic acid as active site residues.
House Dust Mite
Mite allergens proteases include the papain-like cysteine proteases from group 1 (Der p 1, Der f 1); the group 2 (Der p 2, Der f 2), which are lipid-binding proteins causing sensitization in more than 90% of mite-allergic patients [33]; the proteases from groups 3 (Der p 3, Der f 3), group 6 (Der p 6, Der f 6) and group 9 (Der p 9) are serine proteases with trypsin, chymotrypsin and collagenase activity, respectively.
Fungi
Fungal proteases are strong activators of T2 inflammation and play a major role in epithelial damage mechanisms. Fungal proteases, as well as HDM ones, can act as adjuvants of endogenous proteases and degraders of protease inhibitors.
The major fungal proteases are serine proteases, such as those of Cladosporium cladosporioides [38], Penicillium and Aspergillus species [39]; however aspartate proteases have been described in Alternaria alternata [40].
Aspergillus fumigatus, Alternaria alternata, and Cladosporium herbarum proteases induce morphologic changes and cell desquamation in the cultured airway epithelial cells, favoring the release of proinflammatory cytokines [41].
In particular, A. alternata proteases induce intense eosinophilic activation: the addition of aspartate protease inhibitors to A. alternata extract has shown to attenuate the eosinophils response [40].
Fungal proteases, as well as Der p1, interact with the kinin system, the coagulation cascade and the fibrinolytic mechanism. The release of fibrinogen cleavage products induced by prothrombinase activity stimulates the innate immune response through the activation of Toll-Like Receptor-4 (TLR4) [42,43]. Moreover, thrombin is involved in different signaling pathways inducing the IgE-independent cytokine production [44].
Cockroaches
The only cockroaches allergen showing proteolytic activity is the Periplaneta americana serine protease Per a 10 [45], which provides both self activation and adjuvant effects on inactive proteases. Although Blattella germanica extract is rich in proteases that show direct proinflammatory effects on the airway epithelial, none of them have shown a direct proteolytic activity [46]. The aerosolized cockroach extract induced airway eosinophilic inflammation in animal models [47,48].
Foods
Food components, such as melon, kiwi, papaya fruit and others, could induce allergic sensitization and have shown serine and cysteine proteolytic action. The papain, a papaya fruit-derived allergen, belongs to the same family of cysteine proteases of HDM major group 1 allergens. Papain has shown to activate both innate and Th2 immune responses [49], inducing the alarmins release [50,51], the activation of mouse basophils in vitro [52] and lung eosinophilia in mice [53,54].
Pollen
Pollen proteolytic activity has been attributed to both allergenic and non-allergenic cysteine, serine, and metalloprotease [55]. IgE-reactive cysteine proteases are present on the coat of Cynodon dactylon, Sorghum halepense and Phleum pratense pollen [56]. In Ambrosia artemisiifolia pollen, the allergenic cysteine protease Amb a 11 has been isolated [29]. Betula verrucosa contains proteases potentially homologue to Der p 1 [57].
Taken together, allergens proteases alter epithelial cells and cell junctions, promoting the external agents’ penetration and the activation of different inflammatory pathways.
2.1. The Allergens Proteases in Epithelial Barrier Damage and Inflammatory Signals
Airways, skin and gastrointestinal tract are the main tissues involved in allergen proteases-driven epithelial barrier damage [58,59].
In airway epithelium, HDM exerts proteolytic activity through both direct and indirect mechanisms involving the direct occludin and claudin degradation [60,61] and the Protease-Activated Receptors (PARs) activation, respectively [23,60,62]. A primary role has been attributed to Der p 1, although HDM serine peptidases have also shown the ability to damage epithelium [23].
It has been observed that Der p 1 could cause the detachment of bronchial epithelial cells [18,63,64].
The main role of Der p 1 is confirmed by the substantial inhibition of HDM activity if Der p 1-selective inhibition is provided [23].
Similar mechanisms are exploited by the fungal serine proteases Pen c 13 and Asp f 13 and the cockroach Per a 10 [65,66]. Moreover, in human bronchial epithelial cells, the fungal Pen c 13 has shown to downregulate the expression of CD44, which is involved in epithelial repair mechanisms [67].
In the gastrointestinal tract, it has been demonstrated that allergen proteases, such as the kiwi fruit actinidin (Act d 1), affect occludin and ZO-1, increasing the intestinal permeability [68].
Damaged epithelia are easily crossed by allergens and other external agents that directly reach Dendritic Cells (DCs), inducing modifications in cell surface receptors and unbalancing the immune response toward a T2 phenotype.
Active proteases induce the proteolytic cleavage of CD40, which results in reducing the Type 1 (T1) inflammatory mediators release, with a weak IL-12 production and, in contrast, increased levels of IL-4 and IL-13 [69].
Der p 1 can not only induce the soluble CD40 directly from DCs surface [69], but also cleave the DC-SIGN (CD209) [70], a receptor involved in T1 cells differentiation [71]. Moreover, Der p 1 upregulates the expression of CD86 favoring the expression, in DCs, of chemokines involved in T2 response [72].
Der p 1, as well as Per a 10, has been shown to modulate both T and B cells through the direct cleavage of CD25 (the alpha chain of the IL-2 Receptor), and CD23 (the low-affinity Receptor for IgE) [73], resulting in less IL-12 and Interferon (INF) release, increased T2 cytokines levels and IgE synthesis.
In addition, the CD23 cleavage could furtherly increase the IgE synthesis disrupting the negative feedback between the membrane bound form of IgE Receptor and the IgE production [74].
Allergens proteases could activate the mast cells through both IgE-mediated and non- IgE-mediated mechanisms. Non-IgE-mediated mast cells-activation involves the cell surface PARs [75,76].
PAR-1, -3 and -4 are activated by thrombin, while PAR-2 is activated by trypsin, which shares molecular features with allergic proteases [75].
Epithelial cells, mast cells, basophils, eosinophil and other cellular types are all involved in PAR-2 mediated- allergen proteases response, as observed after Der p 1, Der p 3 and Der p 9 exposure [77,78].
In airway epithelial cells, PAR-2 activation induces cellular morphologic changes, cell desquamation and the release of cytokines, growth factors and prostanoids [50].
The inflammatory environment enhances PAR expression [79], this has been demonstrated in asthma patient’s bronchial epithelium biopsies, compared with control [80].
Moreover, allergens can upregulate PAR-2 levels on pulmonary and bone marrow derived myeloid Dendritic Cells (mDCs) [81]. An overexpression of PAR-2 and PAR-3 mRNAs has been described in nasal polyp epithelial cells stimulated with Aspergillus, Alternaria, and Cladosporium [82].
PAR-2 favors the alveolar macrophages [83], neutrophil and eosinophil recruitment. In particular, the role of PAR-2 in eosinophils activation has been confirmed by the inhibition of eosinophilic response - stimulated with the cell-free extract of A. alternata exposure- in presence of protease ligands and PAR-2 antagonist peptide [40].
Active Der p 1, Der f 1 or papain lead to the superoxide anion production through the direct eosinophil activation [84].
Basophils exposed to proteolytically active Der p 1 or papain secrete Thymic Stromal Lymphopoietin (TSLP) and IL-4 in an-IgE independent way [20,85]; the specific mechanism is unknown, although nociceptive primary sensory neurons, Mas-related G-protein- coupled receptors (Mrgprs), seem to be involved [86].
The early role of type 2 Innate Lymphoid Cells (ILC2) in T2 immune response is well known. Damaged epithelium produces alarmines, such as TSLP, IL-25, IL-33 which activate ILC2 to produce large amounts of IL-4, IL-5 and IL-13, promoting Th2 differentiation. Th2 cells contribute to T2 cytokines release and mediate the allergen-specific IgE production [87].
In allergic patients, Der p 1 and Aspergillus have shown to induce the ILC2 recruitment and activation [54,88].
A contribution to tissue injury is provided by the alterations of the protease/anti-protease balance.
As mentioned above, the physiological cellular protective function is ensured by the action of anti-proteases, such as 1-antitrypsin, elafin and Secretory Leukocyte Proteinase Inhibitor (SLPI). The proteases/anti-proteases balance is critical to protect lung tissue being, the loss of their homeostasis, a feature of emphysema and asthma [34].
Beyond the allergen proteases (such as papain, Der p 1, cat skin) degrading effect [34], genetic factors could cause the loss of protease inhibitors expression, contributing to exogenous damage [89,90].
The mechanical and immunological epithelial barrier dysfunction induces molecular, cellular and tissue modifications that are features of allergic asthma.
In this context, the chronic inflammation predisposes to- (and enhances) the airway remodeling and the asthma exacerbations occurrence.
3. Airway Remodeling and Its Role in Asthma Exacerbations
3.1. Biomarkers of Airway Remodeling
Asthma-related airway remodeling includes structural changes like sub-epithelial fibrosis, thicker Airway Smooth Muscle (ASM), mucous gland hyperplasia, angiogenesis, and damaged epithelial layers, resulting in stiffer airway walls [91]. Such remodeling significantly contributes to persistent symptoms and severity in severe asthma cases [92]. Notably, airway remodeling can begin early, even before asthma diagnosis, as observed in preschool children with confirmed wheezing [93]. The identification of potential biomarkers could aid in detecting early signs of remodeling.
3.1.1. Epithelial Remodeling
Asthma-induced airway epithelium remodeling involves epithelial cell deterioration, loss, decreased ciliated cells, and increased goblet cells [94]. The Epithelial-Mesenchymal Transition (EMT) is crucial in this process, driven by Transforming Growth Factor beta (TGF-β), leading to epithelial cells transforming into mesenchymal cells [95]. Markers include reduced E-cadherin and increased N-cadherin [96]. The IL-33/CD146 axis influences EMT in asthma, with HDM extract boosting IL-33 and CD146. Epithelial cell interactions with the immune system may involve Extracellular Vesicles (EVs), with altered microRNA (miRNAs) content in response to stress or activation, playing a role in asthma development [97]. The communication between lung epithelial cells and the immune system may involve EVs carrying miRNAs . These miRNAs, which change due to cellular stress or activation [98], are crucial in asthma [99,100,101], showing different levels in asthma patients’ bronchoalveolar lavage fluid compared to healthy individuals [102]. Research indicates specific miRNAs in EVs from airway epithelium, like miR-34a, miR-92b, and miR-210, could be key in initiating Th2 responses and asthma development [103].
3.1.2. Reticular Basement Membrane Thickening
Research links Reticular Basement membrane (RBM) thickening in asthma to gene expressions influencing airway growth and fibrosis, affecting various physiological processes [104]. Identifying specific fibrocytes in Bronchoalveolar Lavage Fluid (BALF) marked by CD34/CD45RO/α-SMA/procollagen I, indicative of basement membrane thickening, suggests a role in mild asthma’s airway remodeling, with future non-invasive detection possibilities [105]. A study on severe asthma identifies galectin-3 as a biomarker in omalizumab-treated patients, distinguishing responders by their protein profiles related to smooth muscle and extracellular matrix [106].
3.1.3. Subepithelial Fibrosis
TGFβ plays a crucial role in asthma by transforming airway fibroblasts into myofibroblasts, leading to subepithelial fibrosis [107]. The severity of fibrosis correlates with TGF-β1 mRNA levels in bronchial biopsies [108], and elevated αvβ8 integrins in asthma indicate their potential as biomarkers [109]. Periostin, associated with IL-4 and IL-13, impacts fibrosis and inflammation, marking the efficacy of Th2 antagonists [110]. Follistatin-like 1 (FSTL1)-induced autophagy may promote epithelial-mesenchymal transition, suggesting new asthma treatments [111].
3.1.4. Airway Smooth Muscle
Many ASM cell mitogens are involved in asthma, such as Platelet-Derived Growth Factor (PDGF), TGF-β, Epidermal Growth Factor (EGF), Heparin-Binding EGF, and Vascular Endothelial Growth Factor (VEGF) [112].
ASM, histologically assessed by endobronchial biopsies, has been recognized as a valuable biomarker in phenotyping airways disease, especially in the context of personalized medicine [113].
TGF-β stands out as a potential biomarker for this particular mechanism, as it becomes activated when ASM cells and the airways contract. TGF-β is known as a cytokine that promotes remodeling processes [114]. Additionally, pharmacological means to inhibit Transient Receptor Potential Vanilloid-1 (TRPV1), a factor that can influence the tone of ASM and effectively mitigate airway remodeling in living organisms, are promising [115].
Recent research highlights the absence of a complete molecular marker system for ASM cells (ASMCs), yet remains hopeful for future developments. It has been discovered that Myosin Heavy chain 11 (MYH11) serves as a marker for mature SMCs, and Transgelin (TAGLN) indicates early SMC differentiation. This suggests the possibility of using various molecular markers or their combinations to identify the properties and origins of increased ASMCs in asthma-related airway remodeling, depending on the stage of differentiation and research requirements [116].
DNA methylation changes in severe asthma, particularly in ASMCs, illuminate disease mechanisms. Asthma shows reduced methylation in the Phosphodiesterase 4D (PDE4D) promoter area, impacting ASMCs proliferation [117]. These patterns relate to asthma’s severity and correlate with gene and miRNA changes, affecting ASMCs function. This suggests the potential use of demethylating agents in severe asthma treatment [118]. Integrins, crucial in ASM contraction and remodeling, mediate ASM and extracellular matrix interactions. Fibronectin-binding α5β1, α2β1, and α9β1 integrins could be therapeutic targets [112].
3.1.5. Mucus
3.1.6. Vasculature
Many studies have observed changes in the bronchial vascular network in asthma, including increased blood vessel number, size, density, vascular leakage, and plasma engorgement. This neovascularization, a key element of airway remodeling, has uncertain effects on bronchial walls and lung function. Contributing factors include extracellular matrix alterations and growth factor dysregulation [121]. VEGF, a key stimulator of endothelial cell growth and vascular permeability, is elevated in asthma, and specific integrins like αvβ3 and αvβ5 play vital roles in blood vessel development [112].
3.2. Airway Remodeling: Radiological Pathways and Key Points
High-resolution Computed Tomography (HRCT) is crucial in identifying radiological markers in asthma, revealing both static and dynamic airway changes as small as 1 mm in diameter [122,123]. In patients with stable asthma who undergo computer CT scans, three primary measurements are acknowledged as efficient in assessing airway remodeling: the percentage of bronchial Wall Thickness (WT%), the Bronchial-to-Arterial (BA) diameter ratio (BA ratio), and the level of Airway Collapsibility (AC) during both inhalation and exhalation. This evaluation of airway remodeling relies on the post-bronchodilator [124].
In a significant study, about 80% of severe asthma patients showed chest CT abnormalities, highlighting CT’s value in assessing this condition [125].
Hartley et al. discovered a negative correlation between Wall Area percentage (WA%) and Forced Expiratory Volume in 1 second (FEV1) in non-smoking asthma patients. This indicates that WA% is a crucial marker for assessing airway remodeling in severe asthma, highlighting the relationship between airway WT and lung function impairment [126].
Quantitative CT (QCT) scans are effective biomarkers for airway remodeling, significantly enhancing the precise analysis and understanding of severe asthma [92,127,128,129]. QCT biomarkers like WT%, WA%, and air trapping (measured as low attenuation area) are higher in asthma patients compared to controls [130] and are particularly elevated in severe cases [131]. These QCT measures correlate closely with asthma severity, and histological findings, making them effective for both studying and monitoring asthma [132].
3.2.1. Radiological Indicators for Assessing Severity, Early Identification, and Involvement of Small Airways
Bronchial WT (BWT) and emphysema are more common in patients with severe asthma compared to mild asthma [125,133,134,135]. However, other studies have not found a link with the severity of asthma [133,136,137]. WT% is a meaningful radiological marker in assessing lung function changes in asthma. In the Severe Asthma Research Program (SARP) study, which focused on never smokers or ex-smokers with less than a 10-pack year history, it was found that the WT% was notably higher in asthmatics who experienced a significant decline in lung function over a three-year period compared to those whose lung function remained normal or improved [138]. Similarly, in HRCT imaging, patients with lower bronchodilator-responsive FEV1 had twice the WT compared to those with normal FEV1 (about 90% predicted), underscoring a significant relationship between increased BWT and diminished lung function [139].
Emphysema-linked changes notably impact lung function in asthma patients, regardless of smoking habits [140,141] indicates the permanent nature of persistent airway obstruction in severe asthma patients, particularly in those with a significant reduction in baseline bronchodilator-responsive FEV1. In the study by Kim YH et al., emphysema scores were four times higher in the Tr5 group compared to the Tr4 group, a trend also observed among non-smokers [139]. Research has shown that 15% to 39% of people with asthma, including non-smokers, experience these changes [142]. CT-measured air trapping in asthma patients is linked to the severity of their asthma and an increased likelihood of experiencing severe exacerbations [143]. Patients undergoing three months of inhaled corticosteroid therapy showed reduced air trapping in CT scans [144]. Additionally, a study by Haldar et al on 26 patients with severe, eosinophilic, refractory asthma revealed that one-year treatment with mepolizumab, an anti-IL5 monoclonal antibody, significantly lowered average wall area compared to a placebo [145].
Delta Lumen, defined as the percent change in airway lumen area between Functional Reserve Capacity (FRC) and Total Lung Capacity (TLC), is a new metric in a study of 152 asthma patients. It negatively correlates with wall thickness WT% and low attenuation area, especially in severe cases like refractory asthma requiring systemic corticosteroids or hospitalization due to exacerbation. This suggests that a reduced Delta Lumen, as measured by QCT, could be a useful biomarker for identifying severe, unstable asthma [146].
The more pronounced thickening of airway walls observed in HRCT images can act as an early indicator of airway remodeling in asthma cases, even when lung function tests like FEV1 are normal [147].
In persistent asthma, the tBTW is linked to increased resistance and reactance in peripheral airways, a higher frequency of severe exacerbations, and the presence of nasal polyposis [148]. QCT showed a strong correlation between bronchial lumen area and inner diameter with lung function tests in a study of 83 long-term asthma patients. Notably, these measures were reduced from the seventh to the ninth bronchial generations, indicating airway remodeling predominantly in medium and small airways [149].
3.2.2. Radiological Pathway in Patient Phenotyping
Various attempts have been made to phenotype asthmatic patients through radiological pathways.
The WA% is significantly higher in asthma patients than those with Eosinophilic Bronchitis (EB), with a difference of 72 (3.1)% versus 54 (2.1)%. This suggests that in asthma, increased WA% might play a more critical role in airway hyperresponsiveness than factors like air trapping or centrilobular prominence, which are typically considered. In contrast, the WA% in EB patients is not as prominently different [150].
In a cluster-based study of asthma patients, clusters with higher bronchial wall area in the right upper lobe’s bronchus, as assessed by CT, were associated with elevated sputum neutrophil levels [151]. In 2014, Gupta et al., through cluster analysis, identified three novel asthma phenotypes with unique clinical and radiological characteristics. Cluster 1 showed an increase in the lumen volume and a decrease in the percentage wall volume of the right upper lobe apical segmental bronchus. In contrast, Cluster 3 had the smallest lumen volume but the highest percentage wall volume in the same bronchus. Cluster 2, however, displayed an absence of proximal airway remodeling. These findings suggest distinct structural changes in the airways of different asthma phenotypes [152].
In a HRCT study of 109 untreated asthma patients, key findings included airway remodeling, bronchiectasis, and mucus plugs, more pronounced than in healthy individuals. A notable inverse relationship existed between airway WT and mid-expiratory flow [153].
In a study of 61 asthmatic patients, four QCT-based clusters were identified, differing in asthma severity and lung function decline over five years. Cluster C1 consisted of non-severe asthmatics with increased wall thickness; C2 had a mix of severe and non-severe cases with limited bronchodilator response; C3 and C4 included severe asthmatics, with C3 focusing on severe allergic asthma without small airway disease, and C4 on ex-smokers with significant small airway disease and emphysema [154]. Kim S. et al. categorized asthma airway remodeling into three types: Large Airway Involvement (LA), Small Airway Involvement (SA), and Normal/Near-Normal (NN). In their study of 91 severe asthma patients, 81.3% showed bronchial thickening and bronchiectasis, 6.6% had small airway remodeling associated with airflow obstruction and smoking, while 26% displayed no significant remodeling and required fewer oral corticosteroids [155].
4. Relevant Therapeutic Options in Airway Remodeling
4.1. The Role of Standard Therapy in Airway Remodeling (LAMA)
Prior randomized trials have shown that Inhaled Corticosteroids (ICS) can lead to a reduction in subepithelial fibrosis [156,157]. The use of Inhaled Corticosteroids/ Long-Acting Beta2 Agonists (ICS/LABAs) is known to reduce airway inflammatory and remodeling pathways. For instance, in post-ICS-LABA treatment, there’s a noted downregulation in the expression of various elements like nuclear receptor transcription coactivator, N-acetyltransferase, protein tyrosine kinase, nuclear receptor, and RNA polymerase II-activating transcription factor [158].
Muscarinic M1-3 receptors, present in the lungs, are crucial for the bronchodilatory effect of Long-Acting Muscarinic Antagonists (LAMAs), primarily through M3 inhibition. M3 receptors also influence mucus secretion, making LAMAs effective in reducing it. Muscarinic receptors are found in various lung cells, including epithelial cells, fibroblasts, smooth muscle cells, and inflammatory cells. This indicates that non-neuronal cells can also produce and release Acetylcholine (ACh), contributing to different biological responses in an autocrine or paracrine manner [159].
In animal and in vitro studies, LAMAs have shown significant anti-inflammatory and anti-proliferative effects. They are capable of inhibiting airway remodeling triggered by allergens [160].
ACh plays a role in airway inflammation and remodeling, also influencing the growth of ASM. Studies have shown the benefits of using muscarinic ACh Receptor (mAChR) antagonists, especially long-acting types [161] (LAMAs), in targeting these effects by blocking ACh’s activation of mAChRs.
In earlier stages of the condition, the challenge in prescribing LAMAs lies in the high variability of patient responses and the lack of detailed patient phenotyping. Enhancing the characterization of parasympathetic tone activity could lead to more effective LAMA prescriptions [162].
Adding LAMAs to ICS/LABA therapy enhances lung function, decreases exacerbations, and slightly improves asthma control in moderate to severe asthma patients not fully controlled by ICS/LABA alone. LAMAs are effective across various asthma phenotypes and endotypes. Three LAMA molecules- Tiotropium (TIO), Glycopyrronium (GLY), and umeclidinium- have been studied as add-ons, each with slightly different action onsets and half-lives. GLY, in particular, acts slightly faster than TIO and umeclidinium may have similar properties [163].
The impact of anticholinergic drugs on airway remodeling remains unclear. Further research is needed to understand the anti-inflammatory effects of anti-muscarinic drugs on human airway inflammation and remodeling processes.
4.2. The Role of Biological Drugs in Airway Remodeling
The specific mechanisms of how environmental factors trigger inflammatory responses leading to airway remodeling in asthma are not completely clear. Alarmins, cytokines from epithelial cells, start these immune processes, contributing to remodeling. Biological therapies can improve airflow by addressing inflammation and may reverse fixed remodeling caused by structural changes. Differentiating the immediate and long-term effects of biologics is vital for evaluating their impact on severe asthma’s airway remodeling [164].
Omalizumab, a humanized IgG1-κ monoclonal antibody, targets the Fc fragment of IgE [165]. It has been shown to reduce the thickness of the basement membrane and decrease fibronectin deposits in the airways of asthma patients [166].
Mepolizumab treatment in asthma patients not only reduced the number of eosinophils in the bronchial passages but also decreased TGF-β1 positive eosinophils, the thickness and tenascin immunoreactivity of the airways, and the levels of TGF-β1 in bronchoalveolar lavage fluid [167].
In biopsies from severe eosinophilic asthma patients, benralizumab significantly reduced eosinophils in the bronchial lamina propria and airway smooth muscle mass, without affecting myofibroblast numbers. This reduction was linked to the depletion of TGF-β1 positive eosinophils [168]. Additionally, a single dose of benralizumab notably improved ventilation in patients with uncontrolled asthma and significant mucus plugging [169].
In a mouse model of asthma, the use of dupilumab, which blocks both IL-4 and IL-13, was effective in preventing eosinophils from infiltrating lung tissues, while it did not impact the levels of circulating eosinophils [170]. In a different mouse model, blocking the IL-4Rα receptor improved lung function. This effect was achieved by influencing various factors involved in inflammation and the remodeling process in the lungs [171].
TSLP, overexpressed in asthmatics’ airway epithelium, activates lung fibroblasts, promoting airway remodeling [172]. Tezepelumab, a human IgG2-λ monoclonal antibody, targets TSLP. Studies show TSLP’ role in fibrotic lung disease and its blockade reduces inflammation, TGF-β1 levels, and airway remodeling in animal models [173,174]. The CASCADE study revealed Tezepelumab significantly reduces airway submucosal eosinophils in moderate-to-severe asthma patients compared to placebo [175]. Lebrikizumab, a humanized monoclonal antibody, targets and inhibits soluble IL-13, blocking its downstream signaling. In exploratory analyses, treatment with lebrikizumab has been linked to a decrease in subepithelial fibrosis, a characteristic of airway remodeling [176].
The EMT in airway remodeling is influenced by the IL-33/CD146 axis. IL-33, derived from HDM extract-treated alveolar epithelial cells, stimulates CD146 expression. This process promotes EMT in the context of chronic allergic inflammation caused by HDM exposure. These findings highlight the potential of targeting the IL-33/CD146 pathway as a therapeutic approach in airway remodeling [97].
5. Discussion
The loss of epithelial barrier function and the airways remodeling are both features of allergic asthma.
The first one, generally occurs in the early stages of the disease, not rarely preceding the allergic sensitization. The second one has been longer considered as resulting from a long lasting disease, however, evidence has shown that airway remodeling may occur in asthma patients even prior to diagnosis [93].
The barrier damage and the airways remodeling appear to be linked, defining a more complex clinical phenotype: epithelial permeability is higher in severe asthma compared with mild asthma [177], as well as in CRSwNP compared with CRS without nasal polyps [178].
Allergen proteases, such as Der p 1 or Asp f 13, directly induced Airway Hyperresponsiveness (AHR) in animal models [179,180] and provoked morphological and molecular modifications in human ASM cells. These effects have been described not only in allergic patients, but also in subjects without prior allergic sensitization [87].
The complete inactivation of Aspergillus protease activity totally prevented the T2 airway inflammation in a murine model of asthma [21]. Moreover, selective inhibition of Der p 1 not only reduced the levels of blood allergen-specific IgE, but also suppressed AHR in rats, avoiding the chronic inflammation and the predisposition to airway remodeling [181].
The redox ambient in bronchial lumen regulates the response to allergens proteases. Der p 1 activity is enhanced by the bronchial epithelium-secreted glutathione-S-transferase-pi and by the presence of the antioxidant glutathione, both of which are highly present in the human epithelial lung fluid [182]. In damaged epithelia of asthmatic patients this effect is favored by anti-protease and mucociliary clearance impairment [183]. Moreover, in allergens proteases-induced inflammation, the production of mitochondrial Reactive Oxygen Species (ROS) is increased, feeding the inflammatory vicious circle. Downregulation of Indoleamine 2,3-dioxygenase (IDO) is observed in bronchial epithelial cells after Aspergillus, Der p 1 and HDM extracts exposure [184,185].
Cystatin SN (CST1) inhibits the cysteine protease activity and its expression is enhanced in asthma epithelium. In a recent study, sputum and serum CST1 protein levels were negatively correlated with lung function in asthma patients; CST1 protein levels were significantly lower in the serum of HDM-specific IgE-positive asthmatics than in sIgE-negative asthmatics. Moreover, the HDM-induced epithelial barrier function disruption was suppressed by recombinant human CST1 protein in vitro and in vivo, reducing asthma symptoms. CST1 has been considered as a potential biomarker for monitoring asthma control [186].
The complex of different chemical and biological agents, which humans are daily exposed to, is known as the “exposome” [187] and includes microorganisms, pollution, hygiene-derived products, HDM, natural toxins, and food additives. The exposure to these factors alters the cell functions and favors the allergic response activation [188,189].
Proteases deriving from microorganisms, chemicals, environmental pollution, cigarette smoke and other noxious agents may damage the epithelial barrier, similarly to allergen proteases, contributing to the inflammatory process, the AHR and the airway remodeling [190].
Considering infective agents, the exposure to viruses during infancy and childhood predisposes to asthma development [191].
Staphylococcus aureus produces a wide range of proteins including toxins, serine-protease-like proteins, and protein A and its role in severe asthma and CRSwNP is well known. Staphylococcal enterotoxin B-IgE sensitization has been considered as a possible independent risk factor for asthma development and, in severe asthma patients, it has been associated with the presence of CRSwNP as comorbidity [192]. Moreover, Staphylococcus aureus, regardless of enterotoxins production, may damage the airway epithelial cells, inducing the release of IL-25, IL-33 and TSLP, which can activate the ILC2 and the T2 response [193].
Recent evidence has demonstrated that, in the skin of a preclinical mouse model, eosinophil-recruiting chemokines (and eosinophil infiltration) are induced after Staphylococcus aureus epicutaneous exposure, the IL-36α-IL-36R pathway is involved [194].
The exposure to tobacco smoke has been strongly associated with asthma prevalence in children [195] and exacerbates asthma and rhinitis symptoms in adults, decreasing muco-ciliary clearance [196]. The cigarette smoke can directly damage the TJ of the pulmonary epithelium [197], promoting the T2 response through epigenetic modifications, such as decreasing gene methylation of IL-4, IL-13 or increasing FOXP3 methylation after HDM challenge [198,199].
The airborne microplastics inhalation caused pulmonary inflammatory cell infiltration, bronchoalveolar macrophage aggregation and increased the TNF-α levels in both healthy and asthmatic mice [200].
Air pollutants exalt the action of aeroallergens, damaging the pollen cell wall and facilitating the release of allergenic proteins into the environment [201,202]. The direct allergenic proteins - air pollutants contact promotes chemical protein modification before inhalation and deposition in the respiratory tract. Especially high ozone (O3) and nitrogen dioxide (NO2) levels have been shown to efficiently nitrate and cross-link the proteins [203,204]. The concentrations of smog and industrial contamination-associated O3, NO2 and particles in suspension are geographically associated with a higher rate of infant asthma [205]; also maternal exposure to NO2 leads to enhanced sensitivity to allergens and increased AHR [206].
Increased T2 response and accumulation of ILC2 cells was observed in a diesel exhaust-enhanced allergic mice model [207]; O3 and NO2 promote the release of cytokines and chemokines, such as IL-33, IL-25 and TSLP, in both normal and asthmatic patients’ bronchial epithelial cells [208,209].
Recently it has been observed that TLR4 is enhanced in Phl p 5, but not in Bet v 1, after the ROS and nitrogen species exposure; subsequently, chemical modification and increased protein-receptor interactions occur. These events might contribute to the growing prevalence of respiratory allergies in industrialized countries [210].
The allergens proteases cause the airways remodeling both directly and indirectly, through chronic inflammation-induced modifications.
HDM proteses directly induce CX3CL1 chemokine activating the T2 response [211] and the proliferation of ASMCs [212].
HDM proteases, signaling through EGF receptor and TGF-β1, have shown to promote epithelial-to-mesenchymal transition in human bronchial epithelium cells, contributing to airway remodeling in asthma [213].
The chronic inflammation induces a chronic repair reaction leading to the continuous release of growth factors, the uncontrolled proliferation of fibroblasts, ASMCs, goblet cells, and the deposition of extracellular matrix molecules [214,215] which are all features of the airway remodeling process.
Due to the different emerging asthma phenotypes and to the increasing number of factors included in the exposoma concept, which contribute to inflammatory damage and enhance asthma incidence and severity, the identification of clinical and radiological biomarkers is a concrete need in asthma. In a previous study, we have explored the role of serum free light chains -κ and -λ in asthma patients, showing their value as qualitative and quantitative (severity indicator) potential biomarkers, respectively [216].
Focusing on airway remodeling, the examination of the physio-pathological and radiological features in allergic asthma should be considered for patients’ clusterization.
6. Conclusions and Future Directions
The role of allergen proteases in the pathogenesis of allergic asthma has been early identified and it is currently well known. Molecular mechanisms underlying the allergen proteases-induced damage, starting from the epithelial barrier loss, have been better defined over the years and new allergen proteases have been identified.
In our review, the clinical features and the radiological patterns of airways remodeling have been explored, in order to emphasize the importance of biomarker identification in a disease with multiple endotypes and phenotypes.
The aim of this review was to draw a continuous thread between molecular mechanisms of allergen protease exposure, epithelium damage, chronic inflammation and airway remodeling in allergic asthma patients.
Although, as previously remarked, these “steps” are not necessary subsequently, in many cases they may take part in an evolutive process. The lack of available biomarkers, in particular for monitoring airway inflammation and remodeling, doesn’t allow an optimization for the therapeutic management and follow-up of allergic asthma patients.
Considering T2 inflammation, beyond the blockage of IL-5, IL-4, IL-13 and IgE, the epithelial damage-derived cytokines are a new available therapeutic target.
In a future perspective, a better endo- pheno-typization of asthmatic patients will ensure the selection of the appropriate therapeutic option, between the increasing number of available drugs. Currently, different available molecules have shown positive effects on airway remodeling. However, the identification of new potential therapeutic targets in the molecular pathways involved in the airway remodeling process should be achieved, considering the increase of allergic, environmental, and chemical stimuli in the industrialized exposome.
Author Contributions
IB: SC, VNQ Data curation, Methodology, Supervision, Writing, Review and editing. IB: Figure. SD: Tables. CC, DS, FM, GEC, IB, SC, SD, VNQ: Data curation, Methodology, Project administration, Supervision, Review and editing.
Availability statement
The study materials will be available upon reasonable request to the corresponding author.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| Tight Junctions | TJs |
| Zonula Occludens | ZO |
| Chronic Rhinosinusitis with Nasal Polyps | CRSwNP |
| Eosinophilic Esophagitis | EoE |
| Atopic Dermatitis | AD |
| House-Dust Mite | HDM |
| Type 2 | T2 |
| Immunoglobulin-E | IgE |
| Toll-Like Receptor-4 | TLR4 |
| Protease-Activated Receptors | PARs |
| Dendritic Cells | DCs |
| Type 1 | T1 |
| Interferon | INF |
| myeloid Dendritic Cells | mDCs |
| Thymic Stromal Lymphopoietin | TSLP |
| type 2 Innate Lymphoid Cells | ILC2 |
| Secretory Leukocyte Proteinase Inhibitor | SLPI |
| Airway Smooth Muscle | ASM |
| Epithelial-Mesenchymal Transition | EMT |
| Transforming Growth Factor-β | TGF-β |
| Extracellular Vesicles | EVs |
| microRNA | miRNAs |
| Reticular Basement membrane | RBM |
| Bronchoalveolar Lavage Fluid | BALF |
| Follistatin-like 1 | FSTL1 |
| Platelet-Derived Growth Factor | PDGF |
| Epidermal Growth Factor | EGF |
| Heparin-Binding | EGF |
| Vascular Endothelial Growth Factor | VEGF |
| Transient Receptor Potential Vanilloid-1 | TRPV1 |
| ASM cells (ASMCs); Myosin Heavy chain 11 | MYH11 |
| Transgelin | TAGLN |
| Phosphodiesterase 4D | PDE4D |
| High-resolution Computer Tomography | HRCT |
| Wall Thickness percentage | WT% |
| Bronchial-to-Arterial | BA |
| Airway Collapsibility | AC |
| Wall Area percentage | WA% |
| Forced Expiratory Volume in 1 second | FEV1 |
| Quantitative CT | QCT |
| Bronchial WT | BWT |
| Severe Asthma Research Program | SARP |
| Functional Reserve Capacity | FRC |
| Total Lung Capacity | TLC |
| Eosinophilic Bronchitis | EB |
| Large Airway Involvement | LA |
| Small Airway Involvement | SA |
| Normal/Near-Normal | NN |
| Inhaled Corticosteroids | ICS |
| Inhaled Corticosteroids/ Long-Acting Beta2 Agonists | ICS/LABAs |
| Long-Acting Muscarinic Antagonists | LAMAs |
| Acetylcholine | Ach |
| muscarinic ACh Receptor | mAChR |
| Tiotropium | TIO |
| Glycopyrronium | GLY |
| Airway Hyperresponsiveness | AHR |
| Reactive Oxygen Species | ROS |
| Indoleamine 2,3-dioxygenase | IDO |
| Cystatin SN | CST1 |
| Toll-like Receptor | 4TLR4 |
| Ozone | O3 |
| Nitrogen dioxide | NO2 |
References
- Steelant B, Seys SF, Boeckxstaens G, Akdis CA, Ceuppens JL, Hellings PW. Restoring airway epithelial barrier dysfunction: a new therapeutic challenge in allergic airway disease. Rhinology 2016;54:195-205.
- Hartsock, A.; Nelson, W.J. Adherens and tight junctions: Structure, function and connections to the actin cytoskeleton. Biochim. et Biophys. Acta (BBA) - Biomembr. 2008, 1778, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.Y.; Shin, E.; Byeon, S.-J.; Hong, S.J.; Kang, S.H.; Lee, T.; Kim, T.-B.; Choi, J.-H. Serum Zonulin Is a Biomarker for Severe Asthma. Allergy, Asthma Immunol. Res. 2023, 15, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Shahana, S.; Björnsson, E.; Lúdvíksdóttir, D.; Janson, C.; Nettelbladt, O.; Venge, P.; Roomans, G. Ultrastructure of bronchial biopsies from patients with allergic and non-allergic asthma. Respir. Med. 2005, 99, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Xiao C, Puddicombe SM, Field S, Haywood J, Broughton-Head V, Puxeddu I, et al. Defective epithelial barrier function in asthma. J Allergy Clin Immunol 2011;128:549-56.e1-12.
- Caruso, C.; Giancaspro, R.; Guida, G.; Macchi, A.; Landi, M.; Heffler, E.; Gelardi, M. Nasal Cytology: A Easy Diagnostic Tool in Precision Medicine for Inflammation in Epithelial Barrier Damage in the Nose. A Perspective Mini Review. Front. Allergy 2022, 3, 768408. [Google Scholar] [CrossRef] [PubMed]
- Steelant, B. Epithelial dysfunction in chronic respiratory diseases, a shared endotype? Curr. Opin. Pulm. Med. 2020, 26, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.E. Epithelial Barrier Function and Immunity in Asthma. Ann. Am. Thorac. Soc. 2014, 11, S244–S251. [Google Scholar] [CrossRef] [PubMed]
- Koppelman, G.H.; Meyers, D.A.; Howard, T.D.; Zheng, S.L.; Hawkins, G.A.; Ampleford, E.J.; Xu, J.; Koning, H.; Bruinenberg, M.; Nolte, I.M.; et al. Identification of PCDH1 as a Novel Susceptibility Gene for Bronchial Hyperresponsiveness. Am. J. Respir. Crit. Care Med. 2009, 180, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Loxham, M.; Davies, D.E. Phenotypic and genetic aspects of epithelial barrier function in asthmatic patients. J. Allergy Clin. Immunol. 2017, 139, 1736–1751. [Google Scholar] [CrossRef] [PubMed]
- A Fedorov, I.; Wilson, S.J.; E Davies, D.; Holgate, S.T. Epithelial stress and structural remodelling in childhood asthma. Thorax 2005, 60, 389–394. [Google Scholar] [CrossRef]
- Payne DN, Rogers AV, Adelroth E, Bandi V, Guntupalli KK, Bush A, et al. Early thickening of the reticular basement membrane in children with difficult asthma Am J Respir Crit Care Med 2003;167:78-82.
- Cokugras H, Akcakaya N, Seckin, Camcioglu Y, Sarimurat N, Aksoy F. Ultrastructural examination of bronchial biopsy specimens from children with moderate asthma. Thorax 2001;56:25-9.
- Teoh, K.-T.; Siu, Y.-L.; Chan, W.-L.; Schlüter, M.A.; Liu, C.-J.; Peiris, J.S.M.; Bruzzone, R.; Margolis, B.; Nal, B.; Stucke, V.M.; et al. The SARS Coronavirus E Protein Interacts with PALS1 and Alters Tight Junction Formation and Epithelial Morphogenesis. Mol. Biol. Cell 2010, 21, 3838–3852. [Google Scholar] [CrossRef]
- Lehmann, A.D.; Blank, F.; Baum, O.; Gehr, P.; Rothen-Rutishauser, B.M. Diesel exhaust particles modulate the tight junction protein occludin in lung cells in vitro. Part. Fibre Toxicol. 2009, 6, 26. [Google Scholar] [CrossRef]
- Petecchia L, Sabatini F, Varesio L, Camoirano A, Usai C, Pezzolo A, et al. Bronchial airway epithelial cell damage following exposure to cigarette smoke includes disassembly of tight junction components mediated by the extracellular signal-regulated kinase 1/2 pathway. Chest 2009;135:1502-12.
- Poto, R.; Fusco, W.; Rinninella, E.; Cintoni, M.; Kaitsas, F.; Raoul, P.; Caruso, C.; Mele, M.C.; Varricchi, G.; Gasbarrini, A.; et al. The Role of Gut Microbiota and Leaky Gut in the Pathogenesis of Food Allergy. Nutrients 2023, 16, 92. [Google Scholar] [CrossRef]
- Wan, H.; Winton, H.L.; Soeller, C.; Tovey, E.R.; Gruenert, D.C.; Thompson, P.J.; Stewart, G.A.; Taylor, G.W.; Garrod, D.R.; Cannell, M.B.; et al. Der p 1 facilitates transepithelial allergen delivery by disruption of tight junctions. J. Clin. Investig. 1999, 104, 123–133. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. et Biophys. Acta (BBA) - Biomembr. 2009, 1788, 864–871. [Google Scholar] [CrossRef]
- Sokol, C.L.; Barton, G.M.; Farr, A.G.; Medzhitov, R. A mechanism for the initiation of allergen-induced T helper type 2 responses. Nat. Immunol. 2008, 9, 310–318. [Google Scholar] [CrossRef]
- Kheradmand, F.; Kiss, A.; Xu, J.; Lee, S.-H.; Kolattukudy, P.E.; Corry, D.B. A Protease-Activated Pathway Underlying Th Cell Type 2 Activation and Allergic Lung Disease. J. Immunol. 2002, 169, 5904–5911. [Google Scholar] [CrossRef]
- Phipps, S.; Lam, C.E.; Kaiko, G.E.; Foo, S.Y.; Collison, A.; Mattes, J.; Barry, J.; Davidson, S.; Oreo, K.; Smith, L.; et al. Toll/IL-1 Signaling Is Critical for House Dust Mite–specific Th1 and Th2 Responses. Am. J. Respir. Crit. Care Med. 2009, 179, 883–893. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Newton, G.K.; Perrior, T.R.; Robinson, C. Allergen Delivery Inhibitors: A Rationale for Targeting Sentinel Innate Immune Signaling of Group 1 House Dust Mite Allergens through Structure-Based Protease Inhibitor Design. Mol. Pharmacol. 2018, 94, 1007–1030. [Google Scholar] [CrossRef]
- Chevigné, A.; Barumandzadeh, R.; Groslambert, S.; Cloes, B.; Dehareng, D.; Filée, P.; Marx, J.-C.; Frère, J.-M.; Matagne, A.; Jacquet, A.; et al. Relationship between Propeptide pH Unfolding and Inhibitory Ability during ProDer p 1 Activation Mechanism. J. Mol. Biol. 2007, 374, 170–185. [Google Scholar] [CrossRef]
- Herman J, Thelen N, Smargiasso N, et al. Der p 1 is the primary activator of Der p 3, Der p 6 and Der p 9 the proteolytic allergens produced by the house dust mite Dermatophagoides pteronyssinus. Biochim Biophys Acta. 2014;1840(3):1117-1124. [CrossRef]
- Tovey, E.R.; Chapman, M.D.; Platts-Mills, T.A.E. Mite faeces are a major source of house dust allergens. Nature 1981, 289, 592–593. [Google Scholar] [CrossRef]
- Sudha, V.T.; Arora, N.; Gaur, S.N.; Pasha, S.; Singh, B.P. Identification of a serine protease as a major allergen (Per a 10) of Periplaneta americana. Allergy 2008, 63, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.-D.; Tam, M.F.; Chou, H.; Han, S.-H. The Importance of Serine Proteinases as Aeroallergens Associated with Asthma. Int. Arch. Allergy Immunol. 1999, 119, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Bouley, J.; Groeme, R.; Le Mignon, M.; Jain, K.; Chabre, H.; Bordas-Le Floch, V.; Couret, M.-N.; Bussières, L.; Lautrette, A.; Naveau, M.; et al. Identification of the cysteine protease Amb a 11 as a novel major allergen from short ragweed. J. Allergy Clin. Immunol. 2015, 136, 1055–1064. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Finn, R. Twenty years of the MEROPS database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2016, 44, D343–D350. [Google Scholar] [CrossRef] [PubMed]
- Puente, X.S.; Sánchez, L.M.; Overall, C.M.; López-Otín, C. Human and mouse proteases: a comparative genomic approach. Nat. Rev. Genet. 2003, 4, 544–558. [Google Scholar] [CrossRef]
- Radauer, C.; Breiteneder, H. Pollen allergens are restricted to few protein families and show distinct patterns of species distribution. J. Allergy Clin. Immunol. 2006, 117, 141–147. [Google Scholar] [CrossRef]
- Gruber, A.; Manček, M.; Wagner, H.; Kirschning, C.J.; Jerala, R.; Kim, B.-C.; van Gelder, H.; Kim, T.A.; Lee, H.-J.; Baik, K.G.; et al. Structural Model of MD-2 and Functional Role of Its Basic Amino Acid Clusters Involved in Cellular Lipopolysaccharide Recognition. J. Biol. Chem. 2004, 279, 28475–28482. [Google Scholar] [CrossRef]
- Brown, A.; Farmer, K.; MacDonald, L.; Kalsheker, N.; Pritchard, D.; Haslett, C.; Lamb, J.; Sallenave, J.-M. House Dust Mite Der p 1 Downregulates Defenses of the Lung by Inactivating Elastase Inhibitors. Am. J. Respir. Cell Mol. Biol. 2003, 29, 381–389. [Google Scholar] [CrossRef]
- Deb, R.; Shakib, F.; Reid, K.; Clark, H. Major House Dust Mite Allergens Dermatophagoides pteronyssinus 1 and Dermatophagoides farinae 1 Degrade and Inactivate Lung Surfactant Proteins A and D. J. Biol. Chem. 2007, 282, 36808–36819. [Google Scholar] [CrossRef]
- Gough, L.; Schulz, O.; Sewell, H.F.; Shakib, F. The Cysteine Protease Activity of the Major Dust Mite Allergen Der P 1 Selectively Enhances the Immunoglobulin E Antibody Response. J. Exp. Med. 1999, 190, 1897–1902. [Google Scholar] [CrossRef] [PubMed]
- Ghaemmaghami, A.M.; Shakib, F. Human T cells that have been conditioned by the proteolytic activity of the major dust mite allergen Der p 1 trigger enhanced immunoglobulin E synthesis by B cells. Clin. Exp. Allergy 2002, 32, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.; Tam, M.F.; Lee, L.-H.; Chiang, C.-H.; Tai, H.-Y.; Panzani, R.C.; Shen, H.-D. Vacuolar Serine Protease Is a Major Allergen of Cladosporium cladosporioides. Int. Arch. Allergy Immunol. 2008, 146, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.-D.; Tam, M.F.; Tang, R.-B.; Chou, H. Aspergillus and Penicillium allergens: Focus on proteases. Curr. Allergy Asthma Rep. 2007, 7, 351–356. [Google Scholar] [CrossRef]
- Matsuwaki, Y.; Wada, K.; White, T.A.; Benson, L.M.; Charlesworth, M.C.; Checkel, J.L.; Inoue, Y.; Hotta, K.; Ponikau, J.U.; Lawrence, C.B.; et al. Recognition of Fungal Protease Activities Induces Cellular Activation and Eosinophil-Derived Neurotoxin Release in Human Eosinophils. J. Immunol. 2009, 183, 6708–6716. [Google Scholar] [CrossRef]
- Kauffman HK, Tomee JFC, Marjolein A, van de Riet A, Timmerman JB, Borger P. Protease-dependent activation of epithelial cells by fungal allergens leads to morphologic changes and cytokine production. J Allergy Clin Immunol. 2000;105:1185–93.
- Millien, V.O.; Lu, W.; Shaw, J.; Yuan, X.; Mak, G.; Roberts, L.; Song, L.-Z.; Knight, J.M.; Creighton, C.J.; Luong, A.; et al. Cleavage of Fibrinogen by Proteinases Elicits Allergic Responses Through Toll-Like Receptor 4. Science 2013, 341, 792–796. [Google Scholar] [CrossRef]
- Landers, C.T.; Tung, H.-Y.; Knight, J.M.; Madison, M.C.; Wu, Y.; Zeng, Z.; Porter, P.C.; Rodriguez, A.; Flick, M.J.; Kheradmand, F.; et al. Selective cleavage of fibrinogen by diverse proteinases initiates innate allergic and antifungal immunity through CD11b. J. Biol. Chem. 2019, 294, 8834–8847. [Google Scholar] [CrossRef]
- Burzynski, L.C.; Humphry, M.; Pyrillou, K.; Wiggins, K.A.; Chan, J.N.; Figg, N.; Kitt, L.L.; Summers, C.; Tatham, K.C.; Martin, P.B.; et al. The Coagulation and Immune Systems Are Directly Linked through the Activation of Interleukin-1α by Thrombin. Immunity 2019, 50, 1033–1042. [Google Scholar] [CrossRef]
- Sudha, V.T.; Arora, N.; Singh, B.P. Serine protease activity of Per a 10 augments allergen-induced airway inflammation in a mouse model. Eur. J. Clin. Investig. 2009, 39, 507–516. [Google Scholar] [CrossRef]
- Pomes A, Chapman MD, Vailes LD, et al. Cockroach allergen Bla g 2; structure, function, and implications for allergic sensitization. Am J Respir Crit Care Med. 2002;165:391–7.
- Campbell, E.M.; Kunkel, S.L.; Strieter, R.M.; Lukacs, N.W. Temporal Role of Chemokines in a Murine Model of Cockroach Allergen-Induced Airway Hyperreactivity and Eosinophilia. J. Immunol. 1998, 161, 7047–7053. [Google Scholar] [CrossRef]
- Papouchado, B.G.; Chapoval, S.P.; Marietta, E.V.; Weiler, C.R.; David, C.S. Cockroach Allergen-Induced Eosinophilic Airway Inflammation in HLA-DQ/Human CD4+ Transgenic Mice. J. Immunol. 2001, 167, 4627–4634. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Cao, W.; Kasturi, S.P.; Ravindran, R.; I Nakaya, H.; Kundu, K.; Murthy, N.; Kepler, T.B.; Malissen, B.; Pulendran, B. The T helper type 2 response to cysteine proteases requires dendritic cell–basophil cooperation via ROS-mediated signaling. Nat. Immunol. 2010, 11, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Kouzaki, H.; O’Grady, S.M.; Lawrence, C.B.; Kita, H. Proteases Induce Production of Thymic Stromal Lymphopoietin by Airway Epithelial Cells through Protease-Activated Receptor-2. J. Immunol. 2009, 183, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.S.; Angkasekwinai, P.; Chang, S.H.; Chung, Y.; Dong, C. Protease Allergens Induce the Expression of IL-25 via Erk and p38 MAPK Pathway. J. Korean Med Sci. 2010, 25, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Sokol C. L., Chu N. Q., Yu S., Nish S. A., Laufer T. M., Medzhitov R.. 2009. Basophils function as antigen-presenting cells for an allergen-induced T helper type 2 response. Nat. Immunol. 10: 713–720.
- Oboki, K.; Ohno, T.; Kajiwara, N.; Arae, K.; Morita, H.; Ishii, A.; Nambu, A.; Abe, T.; Kiyonari, H.; Matsumoto, K.; et al. IL-33 is a crucial amplifier of innate rather than acquired immunity. Proc. Natl. Acad. Sci. USA 2010, 107, 18581–18586. [Google Scholar] [CrossRef] [PubMed]
- Kamijo, S.; Takeda, H.; Tokura, T.; Suzuki, M.; Inui, K.; Hara, M.; Matsuda, H.; Matsuda, A.; Oboki, K.; Ohno, T.; et al. IL-33–Mediated Innate Response and Adaptive Immune Cells Contribute to Maximum Responses of Protease Allergen–Induced Allergic Airway Inflammation. J. Immunol. 2013, 190, 4489–4499. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, R.; de Matos, M.R.; Cortes, L.; Nunes-Correia, I.; Todo-Bom, A.; Pires, E.; Veríssimo, P. Pollen Proteases Play Multiple Roles in Allergic Disorders. Int. J. Mol. Sci. 2020, 21, 3578. [Google Scholar] [CrossRef] [PubMed]
- Bashir, M.E.H.; Ward, J.M.; Cummings, M.; Karrar, E.E.; Root, M.; Mohamed, A.B.A.; Naclerio, R.M.; Preuss, D. Dual Function of Novel Pollen Coat (Surface) Proteins: IgE-binding Capacity and Proteolytic Activity Disrupting the Airway Epithelial Barrier. PLOS ONE 2013, 8, e53337. [Google Scholar] [CrossRef] [PubMed]
- Höllbacher, B.; Schmitt, A.O.; Hofer, H.; Ferreira, F.; Lackner, P. Identification of Proteases and Protease Inhibitors in Allergenic and Non-Allergenic Pollen. Int. J. Mol. Sci. 2017, 18, 1199. [Google Scholar] [CrossRef]
- Tulic, M.K.; Vivinus-Nébot, M.; Rekima, A.; Medeiros, S.R.; Bonnart, C.; Shi, H.; Walker, A.; Dainese, R.; Boyer, J.; Vergnolle, N.; et al. Presence of commensal house dust mite allergen in human gastrointestinal tract: a potential contributor to intestinal barrier dysfunction. Gut 2016, 65, 757–766. [Google Scholar] [CrossRef]
- Baglivo, I.; Colantuono, S.; Lumaca, A.; Papa, A.; Gasbarrini, A.; Caruso, C. The last step to achieve barrier damage control. Front. Immunol. 2024, 15, 1354556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, J.; Zuo, J.; Newton, G.K.; Stewart, M.R.; Perrior, T.R.; Garrod, D.R.; Robinson, C. Allergen Delivery Inhibitors: Characterisation of Potent and Selective Inhibitors of Der p 1 and Their Attenuation of Airway Responses to House Dust Mite Allergens. Int. J. Mol. Sci. 2018, 19, 3166. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Robinson, C. Cellular and Molecular Events in the Airway Epithelium Defining the Interaction Between House Dust Mite Group 1 Allergens and Innate Defences. Int. J. Mol. Sci. 2018, 19, 3549. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Yu, S.-J.; Tsai, J.-J.; Yu, C.-H.; Liao, E.-C. Antagonism of Protease Activated Receptor-2 by GB88 Reduces Inflammation Triggered by Protease Allergen Tyr-p3. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Wan, H.; Winton, H.L.; Soeller, C.; Taylor, G.W.; Gruenert, D.C.; Thompson, P.J.; Cannell, M.B.; Stewart, G.A.; Garrod, D.R.; Robinson, C. The transmembrane protein occludin of epithelial tight junctions is a functional target for serine peptidases from faecal pellets of Dermatophagoides pteronyssinus. Clin. Exp. Allergy 2001, 31, 279–294. [Google Scholar] [CrossRef] [PubMed]
- Tomee JF, van Weissenbruch R, de Monchy JG, Kauffman HF: Interactions between inhalant allergen extracts and airway epithelial cells: effect on cytokine production and cell detachment. J Allergy Clin Immunol 1998, 102:75–85.
- Schwab, C.J.; Cooley, J.D.; Brasel, T.; Jumper, C.A.; Graham, S.C.; Straus, D.C. Characterization of Exposure to Low Levels of Viable Penicillium chrysogenum Conidia and Allergic Sensitization Induced by a Protease Allergen Extract from Viable P. chrysogenum Conidia in Mice. Int. Arch. Allergy Immunol. 2003, 130, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Kurup, V.; Xia, J.-Q.; Crameri, R.; Rickaby, D.; Choi, H.; Flückiger, S.; Blaser, K.; Dawson, C.; Kelly, K. Purified Recombinant A. fumigatus Allergens Induce Different Responses in Mice. Clin. Immunol. 2001, 98, 327–336. [Google Scholar] [CrossRef]
- Tai, H.-Y.; Tam, M.F.; Chou, H.; Perng, D.-W.; Shen, H.-D. Pen ch 13 Major Fungal Allergen Decreases CD44 Expression in Human Bronchial Epithelial Cells. Int. Arch. Allergy Immunol. 2010, 153, 367–371. [Google Scholar] [CrossRef]
- Grozdanovic, M.M.; Čavić, M.; Nešić, A.; Andjelković, U.; Akbari, P.; Smit, J.J.; Gavrović-Jankulović, M. Kiwifruit cysteine protease actinidin compromises the intestinal barrier by disrupting tight junctions. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 2016, 1860, 516–526. [Google Scholar] [CrossRef]
- Ghaemmaghami, A.M.; Gough, L.; Sewell, H.F.; Shakib, F. The proteolytic activity of the major dust mite allergen Der p 1 conditions dendritic cells to produce less interleukin-12: allergen-induced Th2 bias determined at the dendritic cell level. Clin. Exp. Allergy 2002, 32, 1468–1475. [Google Scholar] [CrossRef]
- Furmonaviciene, R.; Ghaemmaghami, A.M.; Boyd, S.E.; Jones, N.S.; Bailey, K.; Willis, A.C.; Sewell, H.F.; Mitchell, D.A.; Shakib, F. The protease allergen Der p 1 cleaves cell surface DC-SIGN and DC-SIGNR: experimental analysis of in silico substrate identification and implications in allergic responses. Clin. Exp. Allergy 2007, 37, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Cheong, C.; Matos, I.; Choi, J.-H.; Dandamudi, D.B.; Shrestha, E.; Longhi, M.P.; Jeffrey, K.L.; Anthony, R.M.; Kluger, C.; Nchinda, G.; et al. Microbial Stimulation Fully Differentiates Monocytes to DC-SIGN/CD209+ Dendritic Cells for Immune T Cell Areas. Cell 2010, 143, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Smits, H.H.; Ratajczak, C.; Nithiananthan, A.; A Wierenga, E.; A Stewart, G.; Jacquet, A.; Tonnel, A.-B.; Pestel, J. Monocyte-derived dendritic cells exposed to Der p 1 allergen enhance the recruitment of Th2 cells: major involvement of the chemokines TARC/CCL17 and MDC/CCL22. . 2003, 14, 219–28. [Google Scholar]
- Takai, T.; Kato, T.; Ota, M.; Yasueda, H.; Kuhara, T.; Okumura, K.; Ogawa, H. Recombinant Der p 1 and Der f 1 with in vitro Enzymatic Activity to Cleave Human CD23, CD25 and α1-Antitrypsin, and in vivo IgE-Eliciting Activity in Mice. Int. Arch. Allergy Immunol. 2005, 137, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Engeroff, P.; Vogel, M. The role of CD23 in the regulation of allergic responses. Allergy 2021, 76, 1981–1989. [Google Scholar] [CrossRef]
- Reed CE, Kita H: The role of protease activation of inflammation in allergic respiratory diseases. J Allergy Clin Immunol 2004;114:997–1008.
- Yu CK, Chen CL: Activation of mast cells is essential for development of house dust mite Dermatophagoides farinae-induced allergic airway inflammation in mice. J Immunol 2003;171: 3808–3815.
- Asokananthan N, Graham PT, Stewart DJ, et al.: House dust mite allergens induce proinflammatory cytokines from respiratory epithelial cells: the cysteine protease allergen, Der p 1, activates protease-activated receptor (PAR)-2 and inactivates PAR-1. J Immunol 2002, 169:4572–4578.
- Sun, G.; Stacey, M.A.; Schmidt, M.; Mori, L.; Mattoli, S. Interaction of Mite Allergens Der P3 and Der P9 with Protease-Activated Receptor-2 Expressed by Lung Epithelial Cells. J. Immunol. 2001, 167, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, E.; Reiser, G. A novel therapeutic target in various lung diseases: Airway proteases and protease-activated receptors. Pharmacol. Ther. 2007, 115, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Knight DA, Lim S, Scaffidi AK, Roche N, Chung KF, Stewart GA, Thompson PJ. Protease activated receptors in human airways: upregulation of PAR-2 in respiratory epithelial cells from patients with asthma. J Allergy Clin Immunol. 2001;108:797–803.
- Lewkowich, I.P.; Day, S.B.; Ledford, J.R.; Zhou, P.; Dienger, K.; Wills-Karp, M.; Page, K. Protease-activated receptor 2 activation of myeloid dendritic cells regulates allergic airway inflammation. Respir. Res. 2011, 12, 122–122. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.-H.; Lee, Y.-H.; Jeon, C.-H. Protease-dependent activation of nasal polyp epithelial cells by airborne fungi leads to migration of eosinophils and neutrophils. Acta Oto-Laryngologica 2006, 126, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Day, S.B.; Zhou, P.; Ledford, J.R.; Page, K. German Cockroach Frass Proteases Modulate the Innate Immune Response via Activation of Protease-Activated Receptor-2. J. Innate Immun. 2010, 2, 495–504. [Google Scholar] [CrossRef]
- Miike, S.; Kita, H. Human eosinophils are activated by cysteine proteases and release inflammatory mediators. J. Allergy Clin. Immunol. 2003, 111, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.; Coward, W.R.; I Pritchard, D.; A Hewitt, C.R.; Pritchard, D.I.; Hewitt, C.R.A. Basophils express a type 2 cytokine profile on exposure to proteases from helminths and house dust mites. J. Leukoc. Biol. 2003, 73, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Serhan N, Cenac N, Basso L, Gaudenzio N. Mas-related G proteincoupled receptors (Mrgprs) – key regulators of neuroimmune interactions. Neurosci Lett. 2021;749:135724. [CrossRef]
- Soh, W.T.; Zhang, J.; Hollenberg, M.D.; Vliagoftis, H.; Rothenberg, M.E.; Sokol, C.L.; Robinson, C.; Jacquet, A. Protease allergens as initiators–regulators of allergic inflammation. Allergy 2023, 78, 1148–1168. [Google Scholar] [CrossRef] [PubMed]
- Halim, T.Y.; Steer, C.A.; Mathä, L.; Gold, M.J.; Martinez-Gonzalez, I.; McNagny, K.M.; McKenzie, A.N.J.; Takei, F. Group 2 Innate Lymphoid Cells Are Critical for the Initiation of Adaptive T Helper 2 Cell-Mediated Allergic Lung Inflammation. Immunity 2014, 40, 425–435. [Google Scholar] [CrossRef]
- Azouz, N.P.; Ynga-Durand, M.A.; Caldwell, J.M.; Jain, A.; Rochman, M.; Fischesser, D.M.; Ray, L.M.; Bedard, M.C.; Mingler, M.K.; Forney, C.; et al. The antiprotease SPINK7 serves as an inhibitory checkpoint for esophageal epithelial inflammatory responses. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Azouz NP, Klingler AM, Pathre P, et al. Functional role of kallikrein 5 and proteinase-activated receptor 2 in eosinophilic esophagitis. Sci Transl Med. 2020;12(545):eaaz7773. [CrossRef]
- Kozlik, P.; Zuk, J.; Bartyzel, S.; Zarychta, J.; Okon, K.; Zareba, L.; Bazan, J.G.; Kosalka, J.; Soja, J.; Musial, J.; et al. The relationship of airway structural changes to blood and bronchoalveolar lavage biomarkers, and lung function abnormalities in asthma. Clin. Exp. Allergy 2019, 50, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Aysola, R.S.; Hoffman, E.A.; Gierada, D.; Wenzel, S.; Cook-Granroth, J.; Tarsi, J.; Zheng, J.; Schechtman, K.B.; Ramkumar, T.P.; Cochran, R.; et al. Airway Remodeling Measured by Multidetector CT Is Increased in Severe Asthma and Correlates With Pathology. Chest 2008, 134, 1183–1191. [Google Scholar] [CrossRef]
- Lezmi, G.; Gosset, P.; Deschildre, A.; Abou-Taam, R.; Mahut, B.; Beydon, N.; de Blic, J. Airway Remodeling in Preschool Children with Severe Recurrent Wheeze. Am. J. Respir. Crit. Care Med. 2015, 192, 164–171. [Google Scholar] [CrossRef]
- Bahmer, T.; Sand, J.M.B.; Weckmann, M. Lost in transition: biomarkers of remodeling in patients with asthma. Curr. Opin. Pulm. Med. 2020, 26, 40–46. [Google Scholar] [CrossRef]
- Yang, Z.; Qu, Z.; Yi, M.; Shan, Y.; Ran, N.; Xu, L.; Liu, X. MiR-448-5p inhibits TGF-β1-induced epithelial-mesenchymal transition and pulmonary fibrosis by targeting Six1 in asthma. J. Cell. Physiol. 2018, 234, 8804–8814. [Google Scholar] [CrossRef] [PubMed]
- Rout-Pitt, N.; Farrow, N.; Parsons, D.; Donnelley, M. Epithelial mesenchymal transition (EMT): a universal process in lung diseases with implications for cystic fibrosis pathophysiology. Respir. Res. 2018, 19, 1–10. [Google Scholar] [CrossRef]
- Sun, Z.; Ji, N.; Ma, Q.; Zhu, R.; Chen, Z.; Wang, Z.; Qian, Y.; Wu, C.; Hu, F.; Huang, M.; et al. Epithelial-Mesenchymal Transition in Asthma Airway Remodeling Is Regulated by the IL-33/CD146 Axis. Front. Immunol. 2020, 11, 1598. [Google Scholar] [CrossRef] [PubMed]
- Mateescu, B.; Kowal, E.J.K.; Van Balkom, B.W.M.; Bartel, S.; Bhattacharyya, S.N.; Buzás, E.I.; Buck, A.H.; de Candia, P.; Chow, F.W.N.; Das, S.; et al. Obstacles and opportunities in the functional analysis of extracellular vesicle RNA—An ISEV position paper. J. Extracell. Vesicles 2017, 6, 1286095. [Google Scholar] [CrossRef]
- Milger, K.; Götschke, J.; Krause, L.; Nathan, P.; Alessandrini, F.; Tufman, A.; Fischer, R.; Bartel, S.; Theis, F.J.; Behr, J.; et al. Identification of a plasma miRNA biomarker signature for allergic asthma: A translational approach. Allergy 2017, 72, 1962–1971. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kumar, M.; Ahmad, T.; Mabalirajan, U.; Aich, J.; Agrawal, A.; Ghosh, B. Antagonism of mmu-mir-106a attenuates asthma features in allergic murine model. J. Appl. Physiol. 2012, 113, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.Y.; Horvat, J.C.; Pinkerton, J.W.; Starkey, M.R.; Essilfie, A.T.; Mayall, J.R.; Nair, P.M.; Hansbro, N.G.; Jones, B.; Haw, T.J.; et al. MicroRNA-21 drives severe, steroid-insensitive experimental asthma by amplifying phosphoinositide 3-kinase–mediated suppression of histone deacetylase 2. J. Allergy Clin. Immunol. 2016, 139, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Levänen, B.; Bhakta, N.R.; Paredes, P.T.; Barbeau, R.; Hiltbrunner, S.; Pollack, J.L.; Sköld, C.M.; Svartengren, M.; Grunewald, J.; Gabrielsson, S.; et al. Altered microRNA profiles in bronchoalveolar lavage fluid exosomes in asthmatic patients. J. Allergy Clin. Immunol. 2013, 131, 894–903. [Google Scholar] [CrossRef]
- Bartel, S.; La Grutta, S.; Cilluffo, G.; Perconti, G.; Bongiovanni, A.; Giallongo, A.; Behrends, J.; Kruppa, J.; Hermann, S.; Chiang, D.; et al. Human airway epithelial extracellular vesicle miRNA signature is altered upon asthma development. Allergy 2019, 75, 346–356. [Google Scholar] [CrossRef]
- Bazan-Socha, S.; Buregwa-Czuma, S.; Jakiela, B.; Zareba, L.; Zawlik, I.; Myszka, A.; Soja, J.; Okon, K.; Zarychta, J.; Kozlik, P.; et al. Reticular Basement Membrane Thickness Is Associated with Growth- and Fibrosis-Promoting Airway Transcriptome Profile-Study in Asthma Patients. Int. J. Mol. Sci. 2021, 22, 998. [Google Scholar] [CrossRef]
- Nihlberg, K.; Larsen, K.; Hultgårdh-Nilsson, A.; Malmström, A.; Bjermer, L.; Westergren-Thorsson, G. Tissue fibrocytes in patients with mild asthma: A possible link to thickness of reticular basement membrane? Respir. Res. 2006, 7, 50–50. [Google Scholar] [CrossRef] [PubMed]
- Mauri, P.; Riccio, A.M.; Rossi, R.; Di Silvestre, D.; Benazzi, L.; De Ferrari, L.; Negro, R.W.D.; Holgate, S.T.; Canonica, G.W. Proteomics of bronchial biopsies: Galectin-3 as a predictive biomarker of airway remodelling modulation in omalizumab-treated severe asthma patients. Immunol. Lett. 2014, 162, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, S.S.; Yuan, S.; Innes, A.L.; Kerr, S.; Woodruff, P.G.; Hou, L.; Muller, S.J.; Fahy, J.V. Roles of epithelial cell-derived periostin in TGF-β activation, collagen production, and collagen gel elasticity in asthma. Proc. Natl. Acad. Sci. 2010, 107, 14170–14175. [Google Scholar] [CrossRef]
- Vignola, A.M.; Chanez, P.; Chiappara, G.; Merendino, A.; Pace, E.; Rizzo, A.; la Rocca, A.M.; Bellia, V.; Bonsignore, G.; Bousquet, J. Transforming Growth Factor- β Expression in Mucosal Biopsies in Asthma and Chronic Bronchitis. Am. J. Respir. Crit. Care Med. 1997, 156, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.-M.; Sutanto, E.N.; Iosifidis, T.; Kicic-Starcevich, E.; Looi, K.; Garratt, L.W.; Martinovich, K.M.; Lannigan, F.J.; Knight, D.A.; Stick, S.M.; et al. Reduced transforming growth factor β1 (TGF-β1) in the repair of airway epithelial cells of children with asthma. Respirology 2016, 21, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Izuhara, K.; Arima, K.; Ohta, S.; Suzuki, S.; Inamitsu, M.; Yamamoto, K.-I. Periostin in Allergic Inflammation. Allergol. Int. 2014, 63, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liu, Y.; Miller, M.; Cao, L.; Zhao, J.; Wu, J.; Wang, J.; Liu, L.; Li, S.; Zou, M.; et al. Autophagy plays a role in FSTL1-induced epithelial mesenchymal transition and airway remodeling in asthma. Am. J. Physiol. Cell. Mol. Physiol. 2017, 313, L27–L40. [Google Scholar] [CrossRef] [PubMed]
- Joseph, C.; Tatler, A. Pathobiology of Airway Remodeling in Asthma: The Emerging Role of Integrins. J. Asthma Allergy 2022, ume 15, 595–610. [Google Scholar] [CrossRef]
- Sha, J.; Rorke, S.; Langton, D. Airway smooth muscle as an underutilised biomarker: a case report. BMC Pulm. Med. 2019, 19, 1–4. [Google Scholar] [CrossRef]
- Oenema, T.A.; Maarsingh, H.; Smit, M.; Groothuis, G.M.M.; Meurs, H.; Gosens, R. Bronchoconstriction Induces TGF-β Release and Airway Remodelling in Guinea Pig Lung Slices. PLOS ONE 2013, 8, e65580. [Google Scholar] [CrossRef]
- Choi, J.Y.; Lee, H.Y.; Hur, J.; Kim, K.H.; Kang, J.Y.; Rhee, C.K.; Lee, S.Y. TRPV1 Blocking Alleviates Airway Inflammation and Remodeling in a Chronic Asthma Murine Model. Allergy, Asthma Immunol. Res. 2018, 10, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Qiu, C.; Chen, R. A narrative review of research advances in the study of molecular markers of airway smooth muscle cells. Ann. Transl. Med. 2021, 10, 375. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.H.Y.; Shang, Y.; Mitzner, W.; Sham, J.S.K.; Tang, W.-Y. Aberrant DNA Methylation of Phosphodiestarase 4D Alters Airway Smooth Muscle Cell Phenotypes. Am. J. Respir. Cell Mol. Biol. 2016, 54, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.M.; Lavender, P.; Kuo, C.-H.S.; Galea, F.; Michaeloudes, C.; Flanagan, J.M.; Chung, K.F.; Adcock, I.M. DNA methylation modules in airway smooth muscle are associated with asthma severity. Eur. Respir. J. 2018, 51, 1701068. [Google Scholar] [CrossRef] [PubMed]
- Bahmer, T.; Sand, J.M.B.; Weckmann, M. Lost in transition: biomarkers of remodeling in patients with asthma. Curr. Opin. Pulm. Med. 2020, 26, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.Y.; Rogers, D.F. Mucus hypersecretion in asthma: intracellular signalling pathways as targets for pharmacotherapy. Curr. Opin. Allergy Clin. Immunol. 2010, 10, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Harkness, L.M.; Ashton, A.W.; Burgess, J.K. Asthma is not only an airway disease, but also a vascular disease. Pharmacol. Ther. 2015, 148, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.H.; Mitzner, W. Invited Review: Understanding airway pathophysiology with computed tomograpy. J. Appl. Physiol. 2003, 95, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.I.S.; Colby, T.V.; Müller, N.L. Asthma and Associated Conditions: High-Resolution CT and Pathologic Findings. Am. J. Roentgenol. 2004, 183, 817–824. [Google Scholar] [CrossRef]
- Chae, E.J.; Kim, T.-B.; Cho, Y.S.; Park, C.-S.; Seo, J.B.; Kim, N.; Moon, H.-B. Airway Measurement for Airway Remodeling Defined by Post-Bronchodilator FEV1/FVC in Asthma: Investigation Using Inspiration-Expiration Computed Tomography. Allergy, Asthma Immunol. Res. 2011, 3, 111–117. [Google Scholar] [CrossRef]
- Gupta, S.; Siddiqui, S.; Haldar, P.; Raj, J.V.; Entwisle, J.J.; Wardlaw, A.J.; Bradding, P.; Pavord, I.D.; Green, R.H.; Brightling, C.E. Qualitative Analysis of High-Resolution CT Scans in Severe Asthma. Chest 2009, 136, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Siddiqui, S.; Haldar, P.; Raj, J.V.; Entwisle, J.J.; Wardlaw, A.J.; Bradding, P.; Pavord, I.D.; Green, R.H.; Brightling, C.E. Qualitative Analysis of High-Resolution CT Scans in Severe Asthma. Chest 2009, 136, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.; Gupta, S.; Hartley, R.; Brightling, C.E. Computed tomography scans in severe asthma. Curr. Opin. Pulm. Med. 2012, 18, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, A.; Hall, C.; Hoffman, E.A.; Woods, J.C.; Gierada, D.S.; Castro, M. Using imaging as a biomarker for asthma. J. Allergy Clin. Immunol. 2017, 139, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.E. Quantitative computed tomography. Eur. J. Radiol. 2009, 71, 415–424. [Google Scholar] [CrossRef] [PubMed]
- A Little, S.; Sproule, M.W.; Cowan, M.D.; Macleod, K.J.; Robertson, M.; Love, J.G.; Chalmers, G.W.; McSharry, C.P.; Thomson, N.C. High resolution computed tomographic assessment of airway wall thickness in chronic asthma: reproducibility and relationship with lung function and severity. Thorax 2002, 57, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Bumbacea, D.; Campbell, D.; Nguyen, L.; Carr, D.; Barnes, P.; Robinson, D.; Chung, K. Parameters associated with persistent airflow obstruction in chronic severe asthma. Eur. Respir. J. 2004, 24, 122–128. [Google Scholar] [CrossRef] [PubMed]
- DeBoer, E.M.; Spielberg, D.R.; Brody, A.S. Clinical potential for imaging in patients with asthma and other lung disorders. J. Allergy Clin. Immunol. 2017, 139, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Harmanci, E.; Kebapci, M.; Metintas, M.; Ozkan, R. High-Resolution Computed Tomography Findings Are Correlated with Disease Severity in Asthma. Respiration 2002, 69, 420–426. [Google Scholar] [CrossRef]
- Gupta, S.; Hartley, R.; Khan, U.T.; Singapuri, A.; Hargadon, B.; Monteiro, W.; Pavord, I.D.; Sousa, A.R.; Marshall, R.P.; Subramanian, D.; et al. Quantitative computed tomography–derived clusters: Redefining airway remodeling in asthmatic patients. J. Allergy Clin. Immunol. 2014, 133, 729–738. [Google Scholar] [CrossRef]
- Wang, D.; Luo, J.; Du, W.; Zhang, L.-L.; He, L.-X.; Liu, C.-T. A morphologic study of the airway structure abnormalities in patients with asthma by high-resolution computed tomography. J. Thorac. Dis. 2016, 8, 2697–2708. [Google Scholar] [CrossRef]
- Paganin, F.; Trussard, V.; Seneterre, E.; Chanez, P.; Giron, J.; Godard, P.; Sénac, J.P.; Michel, F.B.; Bousquet, J. Chest Radiography and High Resolution Computed Tomography of the Lungs in Asthma. Am. Rev. Respir. Dis. 1992, 146, 1084–1087. [Google Scholar] [CrossRef]
- Obojski, A.; Patyk, M.; Zaleska-Dorobisz, U. Airway remodeling assessed by quantitative computed tomography at various stages of asthma severity defined according to Global Initiative for Asthma report: a single-center study. Pol. Arch. Intern. Med. 2022, 132. [Google Scholar] [CrossRef]
- Krings, J.G.; Goss, C.W.; Lew, D.; Samant, M.; McGregor, M.C.; Boomer, J.; Bacharier, L.B.; Sheshadri, A.; Hall, C.; Brownell, J.; et al. Quantitative CT metrics are associated with longitudinal lung function decline and future asthma exacerbations: Results from SARP-3. J. Allergy Clin. Immunol. 2021, 148, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Shin, K.E.; Chang, H.S.; Lee, J.-U.; Park, S.-L.; Park, J.S.; Park, J.S.; Park, C.-S. Relationships Between High-Resolution Computed Tomographic Features and Lung Function Trajectory in Patients With Asthma. Allergy, Asthma Immunol. Res. 2023, 15, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Tonga, K.O.; Chapman, D.G.; Farah, C.S.; Oliver, B.G.; Zimmermann, S.C.; Milne, S.; Sanai, F.; Jetmalani, K.; Berend, N.; Thamrin, C.; et al. Reduced lung elastic recoil and fixed airflow obstruction in asthma. Respirology 2019, 25, 613–619. [Google Scholar] [CrossRef]
- Shimizu, K.; Tanabe, N.; Oguma, A.; Kimura, H.; Suzuki, M.; Yokota, I.; Makita, H.; Sato, S.; Hirai, T.; Nishimura, M.; et al. Parenchymal destruction in asthma: Fixed airflow obstruction and lung function trajectory. J. Allergy Clin. Immunol. 2022, 149, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Gelb, A.F.; Yamamoto, A.; Verbeken, E.K.; Nadel, J.A. Unraveling the Pathophysiology of the Asthma-COPD Overlap Syndrome. Chest 2015, 148, 313–320. [Google Scholar] [CrossRef]
- Busacker, A.; Newell, J.D.; Keefe, T.; Hoffman, E.A.; Granroth, J.C.; Castro, M.; Fain, S.; Wenzel, S. A Multivariate Analysis of Risk Factors for the Air-Trapping Asthmatic Phenotype as Measured by Quantitative CT Analysis. Chest 2009, 135, 48–56. [Google Scholar] [CrossRef]
- Tunon-De-Lara, J.-M.; Laurent, F.; Giraud, V.; Perez, T.; Aguilaniu, B.; Meziane, H.; Basset-Merle, A.; Chanez, P. Air trapping in mild and moderate asthma: Effect of inhaled corticosteroids. J. Allergy Clin. Immunol. 2007, 119, 583–590. [Google Scholar] [CrossRef]
- Haldar, P.; Brightling, C.E.; Hargadon, B.; Gupta, S.; Monteiro, W.; Sousa, A.; Marshall, R.P.; Bradding, P.; Green, R.H.; Wardlaw, A.J.; et al. Mepolizumab and Exacerbations of Refractory Eosinophilic Asthma. N. Engl. J. Med. 2009, 360, 973–984. [Google Scholar] [CrossRef]
- Shim, S.S.; Schiebler, M.L.; Evans, M.D.; Jarjour, N.; Sorkness, R.L.; Denlinger, L.C.; Rodriguez, A.; Wenzel, S.; Hoffman, E.A.; Lin, C.-L.; et al. Lumen area change (Delta Lumen) between inspiratory and expiratory multidetector computed tomography as a measure of severe outcomes in asthmatic patients. J. Allergy Clin. Immunol. 2018, 142, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Wang, Z.; Shen, C.; Jin, C.; Yu, N.; Wang, J.; Yin, N.; Guo, Y. Small airway dysfunction may be an indicator of early asthma: findings from high-resolution CT. Ann. Allergy, Asthma Immunol. 2019, 122, 498–501. [Google Scholar] [CrossRef]
- Chan, R.; Duraikannu, C.; Thouseef, M.J.; Lipworth, B. Impaired Respiratory System Resistance and Reactance Are Associated With Bronchial Wall Thickening in Persistent Asthma. J. Allergy Clin. Immunol. Pr. 2023, 11, 1459–1462. [Google Scholar] [CrossRef]
- Patyk, M.; Obojski, A.; Sokołowska-Dąbek, D.; Parkitna-Patyk, M.; Zaleska-Dorobisz, U. Airway wall thickness and airflow limitations in asthma assessed in quantitative computed tomography. Ther. Adv. Respir. Dis. 2020, 14. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-W.; Park, J.-S.; Lee, Y.-M.; Lee, J.-H.; Jang, A.-S.; Kim, D.-J.; Hwangbo, Y.; Uh, S.-T.; Kim, Y.-H.; Park, C.-S. Differences in radiological/HRCT findings in eosinophilic bronchitis and asthma: implication for bronchial responsiveness. Thorax 2006, 61, 41–47. [Google Scholar] [CrossRef]
- Gupta, S.; Siddiqui, S.; Haldar, P.; Entwisle, J.J.; Mawby, D.; Wardlaw, A.J.; Bradding, P.; Pavord, I.D.; Green, R.H.; Brightling, C.E. Quantitative analysis of high-resolution computed tomography scans in severe asthma subphenotypes. Thorax 2010, 65, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Hartley, R.; Khan, U.T.; Singapuri, A.; Hargadon, B.; Monteiro, W.; Pavord, I.D.; Sousa, A.R.; Marshall, R.P.; Subramanian, D.; et al. Quantitative computed tomography–derived clusters: Redefining airway remodeling in asthmatic patients. J. Allergy Clin. Immunol. 2014, 133, 729–738. [Google Scholar] [CrossRef]
- Yang, Z.; Qin, L.; Qiao, J.; Cheng, C.; Liu, Y.; Zhang, S.; Fang, X.; Li, Z.; Renz, H.; Liu, X.; et al. Novel imaging phenotypes of naïve asthma patients with distinctive clinical characteristics and T2 inflammation traits. Ther. Adv. Chronic Dis. 2022, 13. [Google Scholar] [CrossRef]
- Kim, S.; Choi, S.; Kim, T.; Jin, K.N.; Cho, S.-H.; Lee, C.H.; Kang, H.-R. Phenotypic clusters on computed tomography reflects asthma heterogeneity and severity. World Allergy Organ. J. 2022, 15, 100628. [Google Scholar] [CrossRef]
- Kim, S.; Lee, C.; Jin, K.; Cho, S.; Kang, H. Severe Asthma Phenotypes Classified by Site of Airway Involvement and Remodeling via Chest CT Scan. J. Investig. Allergol. Clin. Immunol. 2018, 28, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, M.; Nakamura, Y.; Sim, J.J.; Yamashiro, Y.; Uchida, K.; Hosaka, K.; Isogai, S. Inhaled corticosteroid reduced lamina reticularis of the basement membrane by modulation of insulin-like growth factor (IGF)-I expression in bronchial asthma. Clin. Exp. Allergy 1998, 28, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, D.; Chetta, A.; Del Donno, M.; Bertorelli, G.; Casalini, A.; Pesci, A.; Testi, R.; Foresi, A. Effect of short-term treatment with low-dose inhaled fluticasone propionate on airway inflammation and remodeling in mild asthma: a placebo-controlled study. Am. J. Respir. Crit. Care Med. 1997, 155, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.-R.; Liang, Y.-R.; Tzeng, I.-S.; Tzeng, I.-S.; Hsieh, P.-C.; Hsieh, P.-C.; Kuo, C.-Y.; Kuo, C.-Y.; Huang, S.-Y.; Huang, S.-Y.; et al. Transcriptome analysis in patients with asthma after inhaled combination therapy with long-acting β2-agonists and corticosteroids. Int. J. Med Sci. 2022, 19, 1770–1778. [Google Scholar] [CrossRef]
- Koarai, A.; Ichinose, M. Possible involvement of acetylcholine-mediated inflammation in airway diseases. Allergol. Int. 2018, 67, 460–466. [Google Scholar] [CrossRef]
- Cazzola, M.; Ora, J.; Rogliani, P.; Matera, M.G. Role of muscarinic antagonists in asthma therapy. Expert Rev. Respir. Med. 2017, 11, 239–253. [Google Scholar] [CrossRef]
- Cazzola, M.; Calzetta, L.; Matera, M.G. Long-acting muscarinic antagonists and small airways in asthma: Which link? Allergy 2021, 76, 1990–2001. [Google Scholar] [CrossRef] [PubMed]
- Ora, J.; Calzetta, L.; Ritondo, B.L.; Matera, M.G.; Rogliani, P. Current long-acting muscarinic antagonists for the treatment of asthma. Expert Opin. Pharmacother. 2021, 22, 2343–2357. [Google Scholar] [CrossRef]
- Muiser, S.; Gosens, R.; Berge, M.v.D.; Kerstjens, H.A. Understanding the role of long-acting muscarinic antagonists in asthma treatment. Ann. Allergy, Asthma Immunol. 2022, 128, 352–360. [Google Scholar] [CrossRef]
- Varricchi, G.; Ferri, S.; Pepys, J.; Poto, R.; Spadaro, G.; Nappi, E.; Paoletti, G.; Virchow, J.C.; Heffler, E.; Canonica, W.G. Biologics and airway remodeling in severe asthma. Allergy 2022, 77, 3538–3552. [Google Scholar] [CrossRef]
- Schulman, E.S. Development of a Monoclonal Anti-Immunoglobulin E Antibody (Omalizumab) for the Treatment of Allergic Respiratory Disorders. Am. J. Respir. Crit. Care Med. 2001, 164, S6–S11. [Google Scholar] [CrossRef] [PubMed]
- Zastrzeżyńska, W.; Przybyszowski, M.; Bazan-Socha, S.; Gawlewicz-Mroczka, A.; Sadowski, P.; Okoń, K.; Jakieła, B.; Plutecka, H.; Ćmiel, A.; Sładek, K.; et al. Omalizumab may decrease the thickness of the reticular basement membrane and fibronectin deposit in the bronchial mucosa of severe allergic asthmatics. J. Asthma 2020, 57, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Flood-Page, P.; Menzies-Gow, A.; Phipps, S.; Ying, S.; Wangoo, A.; Ludwig, M.S.; Barnes, N.; Robinson, D.; Kay, A.B. Anti-IL-5 treatment reduces deposition of ECM proteins in the bronchial subepithelial basement membrane of mild atopic asthmatics. J. Clin. Investig. 2003, 112, 1029–1036. [Google Scholar] [CrossRef]
- Chachi, L.; Diver, S.; Kaul, H.; Rebelatto, M.C.; Boutrin, A.; Nisa, P.; Newbold, P.; Brightling, C. Computational modelling prediction and clinical validation of impact of benralizumab on airway smooth muscle mass in asthma. Eur. Respir. J. 2019, 54, 1900930. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, M.J.; Kooner, H.K.; Eddy, R.L.; Jeimy, S.; Licskai, C.; Mackenzie, C.A.; Svenningsen, S.; Nair, P.; Yamashita, C.; Parraga, G. Asthma Control, Airway Mucus, and 129Xe MRI Ventilation After a Single Benralizumab Dose. Chest 2022, 162, 520–533. [Google Scholar] [CrossRef] [PubMed]
- Le Floc’h, A.; Allinne, J.; Nagashima, K.; Scott, G.; Birchard, D.; Asrat, S.; Bai, Y.; Lim, W.K.; Martin, J.; Huang, T.; et al. Dual blockade of IL-4 and IL-13 with dupilumab, an IL-4Rα antibody, is required to broadly inhibit type 2 inflammation. Allergy 2020, 75, 1188–1204. [Google Scholar] [CrossRef]
- Scott, G.; Asrat, S.; Allinne, J.; Lim, W.K.; Nagashima, K.; Birchard, D.; Srivatsan, S.; Ajithdoss, D.K.; Oyejide, A.; Ben, L.-H.; et al. IL-4 and IL-13, not eosinophils, drive type 2 airway inflammation, remodeling and lung function decline. Cytokine 2023, 162, 156091. [Google Scholar] [CrossRef]
- Cao, L.; Liu, F.; Liu, Y.; Liu, T.; Wu, J.; Zhao, J.; Wang, J.; Li, S.; Xu, J.; Dong, L. TSLP promotes asthmatic airway remodeling via p38-STAT3 signaling pathway in human lung fibroblast. Exp. Lung Res. 2018, 44, 288–301. [Google Scholar] [CrossRef]
- Chen, Z.-G.; Zhang, T.-T.; Li, H.-T.; Chen, F.-H.; Zou, X.-L.; Ji, J.-Z.; Chen, H. Neutralization of TSLP Inhibits Airway Remodeling in a Murine Model of Allergic Asthma Induced by Chronic Exposure to House Dust Mite. PLOS ONE 2013, 8, e51268. [Google Scholar] [CrossRef]
- Lin, S.-C.; Chou, H.-C.; Chen, C.-M.; Chiang, B.-L. Anti-thymic stromal lymphopoietin antibody suppresses airway remodeling in asthma through reduction of MMP and CTGF. Pediatr. Res. 2019, 86, 181–187. [Google Scholar] [CrossRef]
- Diver, S.; Khalfaoui, L.; Emson, C.; Wenzel, S.E.; Menzies-Gow, A.; Wechsler, M.E.; Johnston, J.; Molfino, N.; Parnes, J.R.; Megally, A.; et al. Effect of tezepelumab on airway inflammatory cells, remodelling, and hyperresponsiveness in patients with moderate-to-severe uncontrolled asthma (CASCADE): a double-blind, randomised, placebo-controlled, phase 2 trial. Lancet Respir. Med. 2021, 9, 1299–1312. [Google Scholar] [CrossRef]
- Austin, C.D.; Edick, M.G.; Ferrando, R.E.; Solon, M.; Baca, M.; Mesh, K.; Bradding, P.; Gauvreau, G.M.; Sumino, K.; FitzGerald, J.M.; et al. A randomized, placebo-controlled trial evaluating effects of lebrikizumab on airway eosinophilic inflammation and remodelling in uncontrolled asthma (CLAVIER). Clin. Exp. Allergy 2020, 50, 1342–1351. [Google Scholar] [CrossRef]
- Hackett, T.-L.; Singhera, G.K.; Shaheen, F.; Hayden, P.; Jackson, G.R.; Hegele, R.G.; Van Eeden, S.; Bai, T.R.; Dorscheid, D.R.; Knight, D.A. Intrinsic Phenotypic Differences of Asthmatic Epithelium and Its Inflammatory Responses to Respiratory Syncytial Virus and Air Pollution. Am. J. Respir. Cell Mol. Biol. 2011, 45, 1090–1100. [Google Scholar] [CrossRef]
- Soyka MB, Wawrzyniak P, Eiwegger T, Holzmann D, Treis A, Wanke K, et al. Defective epithelial barrier in chronic rhinosinusitis: the regulation of tight junctions by IFNgamma and IL-4. J Allergy Clin Immunol 2012;130:1087-96.e1010.
- Balenga, N.A.; Klichinsky, M.; Xie, Z.; Chan, E.C.; Zhao, M.; Jude, J.; Laviolette, M.; Panettieri, R.A., Jr.; Druey, K.M. A fungal protease allergen provokes airway hyper-responsiveness in asthma. Nat. Commun. 2015, 6, 6763. [Google Scholar] [CrossRef] [PubMed]
- Grunstein, M.M.; Veler, H.; Shan, X.; Larson, J.; Grunstein, J.S.; Chuang, S. Proasthmatic effects and mechanisms of action of the dust mite allergen, Der p 1, in airway smooth muscle. J. Allergy Clin. Immunol. 2005, 116, 94–101. [Google Scholar] [CrossRef] [PubMed]
- John RJ, Rusznak C, Ramjee M, Lamont AG, Abrahamson M, Hewitt EL: Functional effects of the inhibition of the cysteine protease activity of the major house dust mite allergen Der p 1 by a novel peptide-based inhibitor. Clin Exp Allergy 2000;30:784–793.
- López-Rodríguez, J.C.; Manosalva, J.; Cabrera-García, J.D.; Escribese, M.M.; Villalba, M.; Barber, D.; Martínez-Ruiz, A.; Batanero, E. Human glutathione-S-transferase pi potentiates the cysteine-protease activity of the Der p 1 allergen from house dust mite through a cysteine redox mechanism. Redox Biol. 2019, 26, 101256. [Google Scholar] [CrossRef] [PubMed]
- McKelvey, M.C.; Brown, R.; Ryan, S.; Mall, M.A.; Weldon, S.; Taggart, C.C. Proteases, Mucus, and Mucosal Immunity in Chronic Lung Disease. Int. J. Mol. Sci. 2021, 22, 5018. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Lee, S.-H. TGF-β/SMAD4 mediated UCP2 downregulation contributes to Aspergillus protease-induced inflammation in primary bronchial epithelial cells. Redox Biol. 2018, 18, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Aldajani, W.A.; Salazar, F.; Sewell, H.F.; Knox, A.; Ghaemmaghami, A.M. Expression and regulation of immune-modulatory enzyme indoleamine 2,3-dioxygenase (IDO) by human airway epithelial cells and its effect on T cell activation. Oncotarget 2016, 7, 57606–57617. [Google Scholar] [CrossRef]
- Yao, L.; Yuan, X.; Fu, H.; Guo, Q.; Wu, Y.; Xuan, S.; Kermani, N.Z.; Adcock, I.M.; Zeng, X.; Liu, Y.; et al. Epithelium-derived cystatin SN inhibits house dust mite protease activity in allergic asthma. Allergy 2023, 78, 1507–1523. [Google Scholar] [CrossRef]
- Rappaport, S.M.; Smith, M.T. Environment and Disease Risks. Science 2010, 330, 460–461. [Google Scholar] [CrossRef] [PubMed]
- Sozener, Z.C.; Ozturk, B.O.; Cerci, P.; Turk, M.; Akin, B.G.; Akdis, M.; Altiner, S.; Ozbey, U.; Ogulur, I.; Mitamura, Y.; et al. Epithelial barrier hypothesis: Effect of the external exposome on the microbiome and epithelial barriers in allergic disease. Allergy 2022, 77, 1418–1449. [Google Scholar] [CrossRef] [PubMed]
- López-Rodríguez, J.C.; Rodríguez-Coira, J.; Benedé, S.; Barbas, C.; Barber, D.; Villalba, M.T.; Escribese, M.M.; Villaseñor, A.; Batanero, E. Comparative metabolomics analysis of bronchial epithelium during barrier establishment after allergen exposure. Clin. Transl. Allergy 2021, 11, e12051. [Google Scholar] [CrossRef] [PubMed]
- Parrón-Ballesteros, J.; Gordo, R.G.; López-Rodríguez, J.C.; Olmo, N.; Villalba, M.; Batanero, E.; Turnay, J. Beyond allergic progression: From molecules to microbes as barrier modulators in the gut-lung axis functionality. Front. Allergy 2023, 4. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie KJ, Anderton SM, Schwarze J. Viral respiratory tract infections and asthma in early life: cause and effect? Clin Exp Allergy. (2014) 44(1):9–19. Clin. Exp. Allergy. [CrossRef]
- Caruso, C.; Colantuono, S.; Ciasca, G.; Basile, U.; Di Santo, R.; Bagnasco, D.; Passalacqua, G.; Caminati, M.; Michele, S.; Senna, G.; et al. Different aspects of severe asthma in real life: Role of Staphylococcus aureus enterotoxins and correlation to comorbidities and disease severity. Allergy 2023, 78, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Martens, K.; Seys, S.F.; Alpizar, Y.A.; Schrijvers, R.; Bullens, D.M.A.; Breynaert, C.; Lebeer, S.; Steelant, B. Staphylococcus aureus enterotoxin B disrupts nasal epithelial barrier integrity. Clin. Exp. Allergy 2021, 51, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Kline, S.N.; Orlando, N.A.; Lee, A.J.; Wu, M.-J.; Zhang, J.; Youn, C.; Feller, L.E.; Pontaza, C.; Dikeman, D.; Limjunyawong, N.; et al. Staphylococcus aureus proteases trigger eosinophil-mediated skin inflammation. Proc. Natl. Acad. Sci. 2024, 121. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Seiber, E.E.; Ferketich, A.K. Secondhand smoke and asthma: What are the effects on healthcare utilization among children? Prev. Med. 2013, 57, 125–128. [Google Scholar] [CrossRef]
- E Baena-Cagnani, C.; Gómez, R.M.; Baena-Cagnani, R.; Canonica, G.W. Impact of environmental tobacco smoke and active tobacco smoking on the development and outcomes of asthma and rhinitis. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 136–140. [Google Scholar] [CrossRef]
- Sun, Y.-B.; Liu, M.; Fan, X.-S.; Zhou, L.-P.; Li, M.-W.; Hu, F.-Y.; Yue, Q.-F.; Zhang, Y.-M. Effects of cigarette smoke on the aggravation of ovalbumin-induced asthma and the expressions of TRPA1 and tight junctions in mice. Mol. Immunol. 2021, 135, 62–72. [Google Scholar] [CrossRef]
- Wu, C.-C.; Hsu, T.-Y.; Chang, J.-C.; Ou, C.-Y.; Kuo, H.-C.; Liu, C.-A.; Wang, C.-L.; Chuang, H.; Chen, C.-P.; Yang, K.D. Paternal Tobacco Smoke Correlated to Offspring Asthma and Prenatal Epigenetic Programming. Front. Genet. 2019, 10, 471. [Google Scholar] [CrossRef]
- Christensen, S.; Jaffar, Z.; Cole, E.; Porter, V.; Ferrini, M.; Postma, B.; Pinkerton, K.E.; Yang, M.; Kim, Y.J.; Montrose, L.; et al. Prenatal environmental tobacco smoke exposure increases allergic asthma risk with methylation changes in mice. Environ. Mol. Mutagen. 2017, 58, 423–433. [Google Scholar] [CrossRef]
- Lu, K.; Lai, K.P.; Stoeger, T.; Ji, S.; Lin, Z.; Lin, X.; Chan, T.F.; Fang, J.K.-H.; Lo, M.; Gao, L.; et al. Detrimental effects of microplastic exposure on normal and asthmatic pulmonary physiology. J. Hazard. Mater. 2021, 416, 126069. [Google Scholar] [CrossRef]
- Ouyang, Y.; Xu, Z.; Fan, E.; Li, Y.; Zhang, L. Effect of nitrogen dioxide and sulfur dioxide on viability and morphology of oak pollen. Int. Forum Allergy Rhinol. 2016, 6, 95–100. [Google Scholar] [CrossRef]
- Motta, A.; Marliere, M.; Peltre, G.; Sterenberg, P.; Lacroix, G. Traffic-Related Air Pollutants Induce the Release of Allergen-Containing Cytoplasmic Granules from Grass Pollen. Int. Arch. Allergy Immunol. 2006, 139, 294–298. [Google Scholar] [CrossRef]
- Reinmuth-Selzle, K.; Ackaert, C.; Kampf, C.J.; Samonig, M.; Shiraiwa, M.; Kofler, S.; Yang, H.; Gadermaier, G.; Brandstetter, H.; Huber, C.G.; et al. Nitration of the Birch Pollen Allergen Bet v 1.0101: Efficiency and Site-Selectivity of Liquid and Gaseous Nitrating Agents. J. Proteome Res. 2014, 13, 1570–1577. [Google Scholar] [CrossRef]
- Backes, A.T.; Reinmuth-Selzle, K.; Leifke, A.L.; Ziegler, K.; Krevert, C.S.; Tscheuschner, G.; Lucas, K.; Weller, M.G.; Berkemeier, T.; Pöschl, U.; et al. Oligomerization and Nitration of the Grass Pollen Allergen Phl p 5 by Ozone, Nitrogen Dioxide, and Peroxynitrite: Reaction Products, Kinetics, and Health Effects. Int. J. Mol. Sci. 2021, 22, 7616. [Google Scholar] [CrossRef]
- Khreis, H.; Kelly, C.; Tate, J.; Parslow, R.; Lucas, K.; Nieuwenhuijsen, M. Exposure to traffic-related air pollution and risk of development of childhood asthma: A systematic review and meta-analysis. Environ. Int. 2017, 100, 1–31. [Google Scholar] [CrossRef]
- Yue, H.; Yan, W.; Ji, X.; Zhang, Y.; Li, G.; Sang, N. Maternal exposure to NO2 enhances airway sensitivity to allergens in BALB/c mice through the JAK-STAT6 pathway. Chemosphere 2018, 200, 455–463. [Google Scholar] [CrossRef]
- De Grove, K.C.; Provoost, S.; Hendriks, R.W.; McKenzie, A.N.; Seys, L.J.; Kumar, S.; Maes, T.; Brusselle, G.G.; Joos, G.F. Dysregulation of type 2 innate lymphoid cells and T H 2 cells impairs pollutant-induced allergic airway responses. J. Allergy Clin. Immunol. 2017, 139, 246–257. [Google Scholar] [CrossRef]
- Mirowsky, J.E.; Dailey, L.A.; Devlin, R.B. Differential expression of pro-inflammatory and oxidative stress mediators induced by nitrogen dioxide and ozone in primary human bronchial epithelial cells. Inhal. Toxicol. 2016, 28, 374–382. [Google Scholar] [CrossRef]
- Bayram, H.; Sapsford, R.J.; Abdelaziz, M.M.; Khair, O.A. Effect of ozone and nitrogen dioxide on the release of proinflammatory mediators from bronchial epithelial cells of nonatopic nonasthmatic subjects and atopic asthmatic patients in vitro. J. Allergy Clin. Immunol. 2001, 107, 287–294. [Google Scholar] [CrossRef]
- Reinmuth-Selzle, K.; Bellinghausen, I.; Leifke, A.L.; Backes, A.T.; Bothen, N.; Ziegler, K.; Weller, M.G.; Saloga, J.; Schuppan, D.; Lucas, K.; et al. Chemical modification by peroxynitrite enhances TLR4 activation of the grass pollen allergen Phl p 5. Front. Allergy 2023, 4, 1066392. [Google Scholar] [CrossRef]
- Loxham, M.; E Smart, D.; Bedke, N.J.; Smithers, N.P.; Filippi, I.; Blume, C.; Swindle, E.J.; Tariq, K.; Howarth, P.H.; Holgate, S.T.; et al. Allergenic proteases cleave the chemokine CX3CL1 directly from the surface of airway epithelium and augment the effect of rhinovirus. Mucosal Immunol. 2018, 11, 404–414. [Google Scholar] [CrossRef]
- Perros, F.; Dorfmuller, P.; Souza, R.; Durand-Gasselin, I.; Godot, V.; Capel, F.; Adnot, S.; Eddahibi, S.; Mazmanian, M.; Fadel, E.; et al. Fractalkine-induced smooth muscle cell proliferation in pulmonary hypertension. Eur. Respir. J. 2007, 29, 937–943. [Google Scholar] [CrossRef]
- Heijink, I.H.; Postma, D.S.; Noordhoek, J.A.; Broekema, M.; Kapus, A. House Dust Mite–Promoted Epithelial-to-Mesenchymal Transition in Human Bronchial Epithelium. Am. J. Respir. Cell Mol. Biol. 2010, 42, 69–79. [Google Scholar] [CrossRef]
- Gao, F.; Qiao, J.; Zhang, Y.; Jin, X. Chronic intranasal administration of Aspergillus fumigatus spores leads to aggravation of airway inflammation and remodelling in asthmatic rats. Respirology 2009, 14, 360–370. [Google Scholar] [CrossRef]
- Kurup, V.P.; Grunig, G. Animal models of allergic bronchopulmonary aspergillosis. Mycopathologia 2002, 153, 165–177. [Google Scholar] [CrossRef]
- Caruso, C.; Ciasca, G.; Baglivo, I.; Di Santo, R.; Gasbarrini, A.; Firinu, D.; Bagnasco, D.; Passalacqua, G.; Schiappoli, M.; Caminati, M.; et al. Immunoglobulin free light chains in severe asthma patient: Could they be a new biomarker? Allergy 2024. [Google Scholar] [CrossRef]
Figure 1.
The role of allergens proteases in epithelial barrier damage, chronic inflammation and airway remodeling. AHR: Airway Hyperresponsiveness.
Figure 1.
The role of allergens proteases in epithelial barrier damage, chronic inflammation and airway remodeling. AHR: Airway Hyperresponsiveness.
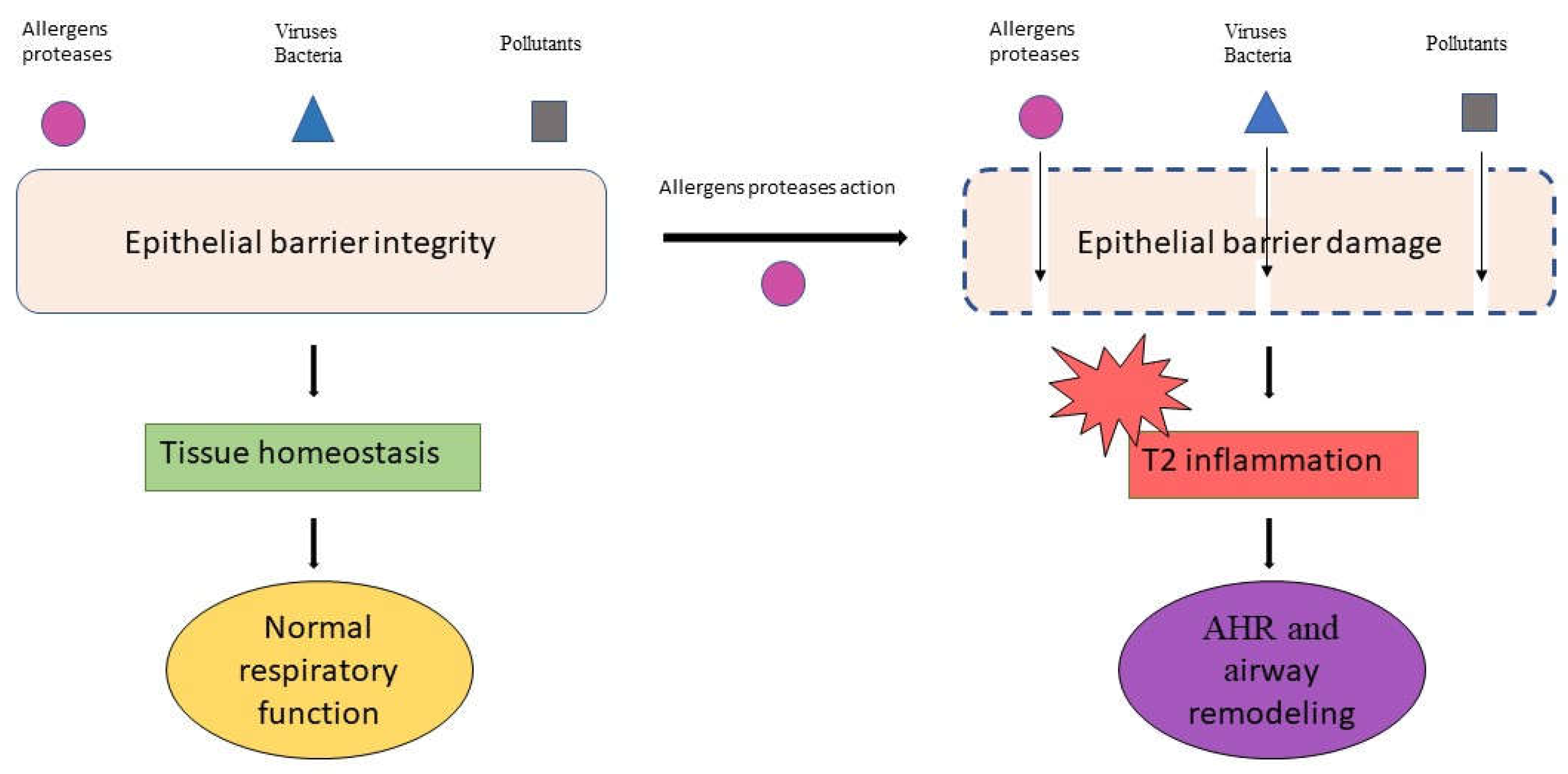

Table 1.
Markers of airway remodeling.
| Marker | Description | Reference |
|---|---|---|
| High-resolution CT (HRCT) | HRCT is crucial for identifying static and dynamic airway changes in asthma, revealing details as small as 1 mm in diameter. | Brown RH, Mitzner W, 1985; Silva CI, Colby TV, 2004 |
| Bronchial Wall Thickness (% WT) | % WT, the bronchial-to-arterial diameter ratio (BA ratio), and the level of airway collapsibility (AC) are acknowledged as efficient measurements for assessing airway remodeling in CT scans. | Chae EJ, Kim TB, Cho YS, et al., 2010 |
| Wall Area Percentage (WA%) | WA% is a crucial marker for assessing airway remodeling in severe asthma, with a negative correlation between WA% and FEV1 observed, indicating the relationship between airway wall thickness and lung function impairment. | Hartley RA, Barker BL, Newby, 2016 |
| Quantitative CT (qCT) scans | QCT scans serve as effective biomarkers for airway remodeling, enhancing the precise analysis of severe asthma. Biomarkers such as wall thickness percentage (WT%), wall area percentage (WA%), and air trapping are higher in asthma patients and particularly elevated in severe cases. | Walker C, Gupta S, Hartley et al., 2012; Aysola RS, Hoffman EA, et al., 2008; Trivedi A, Hall C, Hoffman EA, 2017; Adams JE, 2009 |
| Bronchial Wall Thickness (BWT) and Emphysema | BWT and emphysema are more prevalent in patients with severe asthma, indicating their role as radiological markers for lung function changes in asthma. | Gupta S, Siddiqui, 2009; Harmanci E, Kebapci M, 2002; Gupta S, Hartley R, Khan UT, et al., 2014; Wang D, Luo J, Du W, Zhang LL, 2016 |
Table 2.
Biomarkers of airway remodeling.
| Biomarker | Description | Reference |
|---|---|---|
| Sub-epithelial fibrosis | Characterized by thicker airway smooth muscle, mucous gland hyperplasia, angiogenesis, and damaged epithelial layers, contributing to stiffer airway walls. | Kozlik P, 2020; Aysola RS, chest 2008 |
| Epithelial remodeling | Involves deterioration of epithelial cells, loss of ciliated cells, and an increase in goblet cells. The epithelial-mesenchymal transition (EMT) driven by TGF-β is a key process, with markers like reduced E-cadherin and increased N-cadherin. | Bahmer T, Sand JMB, 2020; Yang ZC, Qu ZH, Yi MJ, 2019; Rout-Pitt N, Farrow N, 2018 |
| Reticular Basement Membrane (RBM) thickening | Linked to gene expressions affecting airway growth and fibrosis. Identifying specific fibrocytes in BALF as markers suggests a role in airway remodeling. | Bazan-Socha S, Buregwa-Czuma, 2021; Nihlberg K, Larsen K, e coll., 2006 |
| Subepithelial Fibrosis | TGFβ’s role in transforming airway fibroblasts into myofibroblasts leading to subepithelial fibrosis. The severity of fibrosis correlates with TGFB1 mRNA levels, and periostin’s association with IL-4 and IL-13 impacts fibrosis and inflammation. | Sidhu SS, Yuan S, Innes AL, 2010; Vignola AM, Chanez P, 1997; Izuhara K, Arima K, Ohta, 2014 |
| Airway Smooth Muscle (ASM) | ASM cell mitogens, such as PDGF, TGFβ, EGF, are involved in asthma. Histology assessed through endobronchial biopsies serves as a valuable biomarker. | Joseph C, Tatler AL, 2022; Sha J, Rorke S, Langton D, et al., 2019 |
| Mucus | Hypersecretion of mucins MUC5AC and MUC5B by goblet cells contributes to airway remodeling, with targeting MUC5AC secretion as a potential therapeutic strategy. | Bahmer T, Sand JMB, Weckmann, 2016; Lai HY, Rogers DF, 2010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



