
Submitted:
18 April 2024
Posted:
22 April 2024
You are already at the latest version
Abstract
The main focus of the in vitro toxicity assessment methods is on the viability of the cells, which is usually based on metabolism changes. Yet, when exposed to toxic substances, the cell triggers multiple signals in response. With this in mind, we have developed a promising cell-based toxicity method that observes various cell responses when exposed to toxic substances (either death, division or remaining viable). Based on the collective cell response, we observed and predicted the dynamics of the cell population to determine the toxicity of the toxicant. The method was tested with two different conformations: In the first conformation, we exposed a monoculture model of blood macrophages to UV light, hydrogen peroxide, nutrient deprivation, tetrabromobisphenol A, fatty acids and 5-fluorouracil. In the second, we exposed a coculture liver model consisting of hepatocytes, hepatic stellate cells, Kupffer cells and liver sinusoidal endothelial cells to rifampicin, ibuprofen and 5-fluorouracil. The method showed good accuracy compared to established toxicity assessment methods. In addition, this approach provided more representative information on the toxic effects of the compounds as it considers the different cellular responses induced by toxic agents.
Keywords:
cell viability
; in vitro toxicology
; dynamic direct cell death assay
1. Introduction
Cell death is a dynamic process with different pathways (apoptosis, necroptosis, ferroptosis…) that lead to different responses. Each pathway has many different subroutines with many different molecular mechanisms that can overlap [1]. Most subroutines last between 3 and 48 hours and usually end in cell death. However, during this period, the cell can reverse the cell death pathway (a process known as anastasis). Anastasis can be triggered at any point along the death pathway until the cell is no longer able to reverse the death pathway [2]. In addition, certain cell death pathways release cytokines that trigger the proliferation of neighbouring cells (of the same or a different cell type). This is a mechanism that allows the tissue to compensate for the death of neighbouring cells and is called compensatory proliferation (a pathway involved in normal homeostatic mechanisms) [3]. Thus, when a toxic agent is injected into a cell population, the cells either die, remain intact or divide [4,5].
In this sense, evaluating the toxicity of a compound using a cell model can be misleading if not all the possible outcomes are considered. For example, a toxic compound may induce a decrease in the cell number of a cell population which may afterwards renew it. Moreover, this cell population may be a functional cell population to the same extent as before the exposure [4,6]. To solve this problem, we propose a new method to determine substance’s toxicity that encompasses the three outcomes mentioned above. Based on these outcomes we observed and predicted cell population dynamics to determine the toxicity of the toxic agent. We tested the suitability and accuracy of the proposed method with two different conformations to have a more comprehensive data in regards of its usability. In the first conformation, we exposed a monoculture model of blood macrophages to different toxic agents. In the second, we exposed a coculture liver model to various hepatotoxic compounds. We observed that the proposed method is consistent with the toxicity assessment of other well-established toxicity assessment methods. Moreover, the toxicity assessment of the hepatotoxic drugs with this new method is in line with the available clinical data [7,8,9,10,11].
2. Results
2.1. Toxicity Assessment of Various Compounds with Different Methods.
MTT toxicity assessment was concordant with the cell count method in all the samples with the exception of the FA and H2O2 treatments (Figure 1a). The toxicity assessment with the MTT assay overestimate the toxic effect of the FA and H2O2 exposure in TLT cells (Figure 1a). Similar trend was observed when depraving the TLT cells of nutrients, but it was not statistically significant.
PI toxicity assessment underestimate the toxicity of FA, H2O2, TBBPA and UV in comparison to MTT and cell count methods (Figure 1a). The toxicity assessment with the PI method and cell count method were similar when starving the cells or treating them with 5-FU (Figure 1a).
When analysing the cell number with the DD Cell-Tox method in the untreated samples (control), we count more cells on Day 1 than on Day 0 (Figure 1b) despite they already formed monolayer at Day 0. On Day 1, the dead cells ratio was around 12%, while the ratio of dividing cells was 20% (Table 1). Thus, a pro-proliferative effect was expected in the following day (Day 1+).
Samples treated with FA and H2O2 had less cells on Day 1 than on Day 0 (Figure 1b). On Day 1, both groups didn`t have any dividing cells, and the dead cell ratio was high (35% and 33% for FA and H2O2 treatments, respectively) (Table 1). Thus, an anti-proliferative effect was expected (Day 1+), which was confirmed by the analysis with the cell count method at Day 2 (Figure 1c). Indeed, we observed a significantly lower cell number in both treatments when comparing them to the untreated samples.
The samples treated with TBBPA and UV had less cells on Day 1 than on Day 0 (Figure 1b). On Day 1, we observed cell division in 3% and 7% of the samples in TBBPA and UV treatments, respectively (Table 1). While most of the cells were dead (dying), 45% and 26% in samples treated with TBBPA and UV treatments, respectively (Table 1). Thus, an overall anti-proliferative effect is expected in both treatments (Day 1+), as it was observed by the cell count method on Day 2 (Figure 1c). This differences on cell number between the treated and untreated samples were statistically significant.
We counted less cells on Day 1 than on Day 0 in samples treated with nutrient starvation and 5-FU (Figure 1b). On Day 1, 26% and 39% of cell division ratio was determined in 5-FU and nutrient starvation, respectively (Table 1). While the cell death ratio on both treatments was lower than the cell division (7% and 12% in 5-FU and nutrient starvation, respectively) (Table 1). Thus, a pro-proliferative effect is expected (Day 1+), which was corroborated by the cell count method on Day 2 (Figure 1c).
The toxicity assessment (given as cell number predictions) with the DD Cell-Tox method at Day 1+ (after the treatment) (Figure 1b) for the coming days were similar to the cell number counted at Day 2 with the Cell Count method (Figure 1c). It is noteworthy that the DD Cell-Tox method predict accurately the treatments that had an overall pro-proliferative effect and anti-proliferative effect. This is visible as cell number prediction on Day 1+ (Figure 1b) and was then corroborated with the cell count method at Day 2 (Figure 1c).
2.2. Toxicity Evaluation of DD Cell-Tox Method in Complex Cell-Based Liver Model in Comparison to Clinical Data.
The cell number increased in all samples of the liver model (Table 2). We didn’t observe any toxic response in the samples treated with RIF, IBU or 5-FU (Figure 2 a). However, the prediction of the cell number infers that the cell number may be reduced in the future. This is consistent with the observations made in patients treated with rifampicin, ibuprofen or 5-FU (Figure 2b) [7,8,9,10,11]. In the first week of treatment, no toxic effect was observed in the liver of the patients. In the following weeks, however, the patients’ liver markers rose above normal values, indicating a possible toxic effect on the liver (Figure 2b).
3. Discussion
Cells are constantly dying and reproducing to maintain the homeostasis of the organ. Thus, assessing the viability of a cell population at a given time may not describe the dynamics of the cell population (in terms of its viability) [4,5]. This is especially true when inoculating a toxic substance. A group of cells that is categorised as dead may reverse course and even induce neighbouring cells to proliferate. The frequency of these events (known as false positives) could be lower if viability is determined using a method that considers the dynamics of the cell population. To estimate the imminent dynamics of a cell population (t1+h), we can calculate the difference in cell number between two different time points (t0 and t1), count the dividing cells and subtract the dead cells at time point t1.
Most cell death pathways last 48 hours [1,12,13,14], and most cells divide for 24 hours [15]. In addition, tissue-repairing signals that trigger proliferation are initiated approximately 6 hours after the onset of programmed cell death [3]. Therefore, 24-48 hours after inoculation with the toxic compound, the majority of cells would either die, maintain their integrity or divide.
Previous studies observed that exposure of a toxic compound to a cell population can be accompanied not only by cell death but also by cell proliferation (known as compensatory proliferation) [16,17]. This situation can be even more pronounced in heterogeneous cell populations; certain cell types can be induced to die while other cell types can be induced to proliferate [16,17]. In complex models such as the liver can be particularly striking, where dying hepatocytes (which make up 80% of the liver) can induce fibroblast proliferation through compensatory proliferation [18,19]. If these assumptions are contemplated, the toxicity of a given compound can be determined with greater accuracy than existing methods because all possible outcomes are considered (in contrast to existing methods). In this sense, the proposed method predicted cell population dynamics in two models (monoculture of TLT and complex liver model with four different cell types) with high accuracy.
When exposing the TLT cells to 12.5 mM H2O2 we expected to observe a higher frequency of necroptotic cells [20]. On the other hand, 266 µM TBBPA or 500 µM 5-FU exposure should induce apoptosis on TLT cells [21,22]. Cell death (mainly by autophagy) was also induced by nutrient starvation [23]. For this purpose, 50 microliters of the cell medium were added, and the medium was not replenished for 72 hours. To induce ferroptosis (via lipid peroxidation) [24], 15 µL of FA was added to the cell medium. Finally, cell death (mainly parthanatos and apoptosis) was also induced by UV irradiation of the cells for 5 minutes [25]. After the exposure to the above mentioned toxic agents, we observed a reduction in cell viability in all cases, which was expected [20,21,22,23,24,25]. At this point, we replenish the media in all samples with cell culture medium without the toxic substances. 24 hours after changing the media, the cells exposed to nutrient deprivation and 500 µM 5-FU proliferated, while the remaining cells showed reduced viability. The nutrient deprivation triggers an autophagy response [23], which we observed in the first 24 hours (Figure 1b, Day 1). However, as previously observed by other studies [26,27,28], after the cells were supplied with new nutrients, they changed this pathway and favoured cell division (Figure 1b, Day 1+). 5-FU induces cell death arrest in cells [29], and subsequently to cell death, as we observed in the samples treated with it (Figure 1b, Day 1). But after changing the medium, the cells reversed the cell death pathway and induced proliferation (Figure 1b, Day 1+). As a result, the cells showed a higher cell number 24 hours after cell renewal (Figure 1c, Day 2). As observed in a previous study, cells can re-enter the cell cycle after cell cycle arrest [30]. In another study, the mechanisms between cell cycle arrest switching and proliferation were also discussed (growth factors and focal adhesion kinase activation of cell cycle and the arrest induced by yes-associated protein phosphorylation) [31]. Once the cell cycle arrest molecule is washed away, cells can resume or continue the cell cycle and proliferate [31].
It is noteworthy that the untreated sample in the TLT cell experiment showed a relatively high ratio of cell death to cell division. However, considering that the blood macrophages formed a monolayer at the beginning of the experiment and had a doubling time of 20 hours, further TLT cultivation is expected to induce them to die and proliferate [32].
We also tested the DD Cell-Tox method in a liver model with four different types of liver cells (LSEC, Kupffer, HSC and hepatocytes). We exposed the model to three different hepatotoxic compounds and evaluated the cell population dynamics using the DD Cell-Tox method. The cell population predictions were consistent with the clinical data (Figure 2b) [7,8,9,10,11].
RIF is an antibiotic that is commonly used to treat tuberculosis. Treatment with RIF can lead to toxicity after the first week of exposure. The mechanism of hepatotoxicity is related to the induction of various cytochrome P450 (CYP) enzymes, including CYP3A4 and ABC C2. The toxicity of the resulting metabolites may be the main cause of toxicity [7]. In this sense, no toxic response was observed during the first week, as documented in clinical data (Figure 2b) [10].
IBU is a non-steroidal anti-inflammatory drug (NSAID) that is used to treat inflammation and mild pain in patients. IBU can have potentially hepatotoxic effects with prolonged constant use (over a week of constant use at high doses), possibly due to the accumulation of toxic metabolic by-products [33]. During the first week of exposure, we didn’t observe any toxic response, as it happened with the clinical reference we found (Figure 2b) [11].
5-FU is used as a cytotoxic anticancer agent for the treatment of various types of cancer (especially in the colon). 5-FU has been associated with clinically apparent acute liver injury in certain cases. Clinical hepatotoxic markers increase slightly when patients are treated with 5-FU. This is mainly due to the accumulation of toxic metabolic by-products and the inhibition of thymidylate synthase [8,34,35,36]. The latter may induce a cell population reduction in comparison to the untreated sample during the first week of exposure (Table 2). Longer exposure to 5-FU would have a significant toxic effect on the liver model, as observed in clinical references (Figure 2b) [9].
Thus, the method is compatible and accurate with a complex model with different cell types. In addition, this method allows a more informative observation of potential cell replication and cell death events. With the DD Cell-Tox method, we can observe cell death or proliferation activity in the first 24 hours and predict possible cell deaths and cell doublings in the next 24 hours. In this way, we cover a 48-hour window, which is normally the longest period for programmed cell death or cell doubling.
Highlights
- The DD Cell-Tox method directly determines the viability of cells. Therefore, it has the potential to be more accurate than methods based on an indirect determination of cell viability.
- The DD Cell-Tox method provides a more comprehensive assessment of toxic compound-induced cell outcomes.
- The application of the DD Cell-Tox method can be of particular importance in toxicological studies where compensatory proliferation is expected.
Limitations
At a certain point, the cell can initiate the cell death process, but the test does not classify it as dead because the test only determines the cells after the “point of no return”. However, counting a cell that is dead before the point of no return can lead to a false positive result, as the cell can reverse the process.
Innovative methods for testing carcinogenic cells continue to be a focus of research and development in oncology and cancer biology [37,38]. Cancer treatments aim to preserve the function of healthy cells by maintaining homeostasis, while inducing cell death of cancer cells to control the disease [39]. This method is of particular interest for research with carcinogenic cells for use in cancer models as it assesses cell death and proliferation.
4. Materials and Methods
4.1. Cells
i. Human blood macrophages, called TLT, were isolated from the blood of a healthy donor at the University of Maribor (Maribor, Slovenia) (10). TLT cells were cultured in colourless Williams E medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 5 wt.% fetal bovine serum (Gibco, Thermo Fisher Scientific, Waltham, MA USA), respectively. L-glutamine (2 mM, Sigma), penicillin (100 U mL−1, Sigma), and streptomycin (1 mg*/mL−1, Fluka, Buchs, Switzerland) were also added. Cells were cultured in 25 cm2 culture flasks (Corning, New York, USA) at 37 °C and 5% CO2.
ii. A liver model was built by coculturing liver sinusoidal endothelial cells (LSEC), human stellate cells (HSC), Kupffer cells (nonparenchymal liver cells, NPC), from ZEN-BIO (USA), and hepatocytes from Lonza (Switzerland). NPC and hepatocytes were grown separately in a 25 cm2 flask (NUNC, Denmark) in a controlled environment at 37º C and 5% CO2 and later seeded together in a 96-well microplate (NUNC, Denmark) to build the liver model.
4.2. Chemicals
Chemical 3-(4,5-dimethylthiazole-2-yl)-2,5-phenyl tetrazolium bromide (MTT) was acquired from Sigma-Aldrich (Saint Louis, MO, USA). Phosphate-buffered saline solution (PBS), FluoVolt™ (FV) Membrane Potential Kit, Vybrant™ DyeCycle™ Ruby Stain (Vybrant), and propidium iodide (PI) were purchased from Thermo Fisher Scientific (Waltham, MA USA).
Hydrogen peroxide (30%) (H2O2), tetrabromobisphenol A (TBBPA), Rifampicin (RIF), ibuprofen (IBU), and 5-fluorouracil (5- FU) were purchased from Merck (Merck KGaA, Darmstadt, Germany). Fatty acids (FA) were obtained from oil digestion with lipase (Merck KGaA, Germany).
4.2. Protocol
The protocol of the DD-Cell-Tox method is the same as the protocol described in Appendix A.3. in the manuscript “A Promising Method for the Determination of Cell Viability: The Membrane Potential Cell Viability Assay” [40] (Figure 3a-b). As described in this manuscript, the cells with FV intensity outside the “viable FV intensity range” (term described in the manuscript) were categorised as dead. In contrast, the cells with FV intensity within the “viable FV intensity range” were categorised as viable cells. However, in the DD-Cell-Tox method, we count the number of dividing cells: The cells that were temporally depolarized and had a higher DNA content (2n) were categorised as duplicating cells [41]. By doing so, we were able to observe and predict the cell number in the following days by calculating the cell number at the time of the experiment, counting the dividing cells and subtracting the dead cells at the time of measurement. This method provides the cell dynamics, and the toxicity of the drug is determined by it (Figure 3c).
4.3. Testing the Method in the First Configuration (Monoculture)
We seeded TLT cells in two 96-well microplate and waited until they reached 100% confluency. After cell monolayer formation, the cells on a 96-well microplate were treated with different toxic substances 12.5 mM H2O2, 266 µM TBBPA, 500 µM 5-FU, nutrient deprivation, 15 µL FA for 24 hours. We exposed the cells seeded in the other 96-well microplate to UV light for 5 minutes. At this point, we tested the viability of the samples with three different methods: DD Cell-Tox method, PI method [42] and MTT assay [43] (Figure 4c). After the treatments, we renewed the cell medium in all samples with new cell culture medium without the toxic agents. And we tested the viability of the samples with the Cell count method [44] (Figure 4d) and compared its toxicity assessment with the DD Cell-Tox method, PI method and MTT assay (Figure 4e).
4.4. Testing the Method in the Second Arrangement (Coculture Model)
We cultured the liver cells separately until confluency and then reseeded them to build the liver model. We seeded 20,000 hepatocytes and 5,000 NPC (Figure 5a). We cultured them with Hep medium (as described in the manuscript “In Vitro Human Liver Model for Toxicity Assessment with Clinical and Preclinical Instrumentation” [45]). We individually added three different hepatotoxic drugs to the cell culture media at the following final concentrations: 50 µmol/l RIF, 1 mmol/l IBU or 500 µmol/l 5-FU. Two days after the addition we renew the same culture medium (Figure 5c). 5 days after the initial exposure, we measured the viability of the cells with the DD Cell-Tox method and compared to the clinical data obtained from the literature [7,8,9,10,11] (Figure 5d).
4.5. Statistical Analysis
We used the ANOVA-tukey HSD test to compare the viability assessment of the cell count, MTT and PI methods. We used Mann-Whitney to test the difference between the cell number in DD Cell-Tox and Cell count methods.
5. Conclusions
Cell viability tests should determine cell viability as a changing parameter and not as a single moment. Based on this basic observation regarding the measurement of viability, we recommend a dynamic approach to measure an observed cell population rather than cell viability at a specific point in time. This is particularly important when evaluating the effect of potential anticarcinogenic compounds, where different cell responses are expected in cancerogenic and healthy cells after their exposure to them. The DD Cell-Tox method incorporates these premises and has been successfully tested in different situations. However, many alternative tests should be performed to evaluate the potential broad application of the method.
Author Contributions
Conceptualization, E.M.; methodology, E.M.; software, E.M.; validation, E.M.; formal analysis, E.M.; investigation, E.M.; resources, E.M.; data curation, E.M.; writing—original draft preparation, E.M.; writing—review and editing, E.M, L.K.S., M.R. and M.M. All authors have read and agreed to the published version of the manuscript.”
Funding
The research leading to these results has received funding from the European Union’s Seventh Framework Programme (FP7/2007-2013) under grant agreement no 311820 (ECsafeSEAFOOD).
Data Availability Statement
Authors agree to make data and materials supporting the results or analyses presented in their paper available upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Gupta, K.H.; Goldufsky, J.W.; Wood, S.J.; Tardi, N.J.; Moorthy, G.S.; Gilbert, D.Z.; Zayas, J.P.; Hahm, E.; Altintas, M.M.; Reiser, J.; Shafikhani, S.H. , Apoptosis and Compensatory Proliferation Signaling Are Coupled by CrkI-Containing Microvesicles. Dev Cell 2017, 41, 674–684e5. [Google Scholar] [CrossRef] [PubMed]
- Gudipaty, S.A.; Conner, C.M.; Rosenblatt, J.; Montell, D.J. Unconventional Ways to Live and Die: Cell Death and Survival in Development, Homeostasis, and Disease. Annu. Rev. Cell Dev. Biol. 2018, 34, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Alenzi, F.Q. , Links between apoptosis, proliferation and the cell cycle. Br J Biomed Sci 2004, 61, 99–102. [Google Scholar] [CrossRef]
- Krafts, K. , Tissue Repair. In Encyclopedia of Toxicology (Third Edition), Wexler, P., Ed. Academic Press: Oxford, 2014; pp 577-583.
- Cusack, R.; Chawke, L.; O’brien, D.; O’connor, B.; O’connor, T. Predictors of hepatotoxicity among patients treated with antituberculous medication. Qjm: Int. J. Med. [CrossRef]
- Vincenzi, B.; Imperatori, M.; Picardi, A.; Gentilucci, U.V.; Gallo, P.; Fausti, V.; Ceruso, M.S.; Santini, D.; Tonini, G. Liver toxicity in colorectal cancer patients treated with first-line FOLFIRI-containing regimen: a single institution experience. Expert Rev. Anticancer. Ther. 2015, 15, 971–976. [Google Scholar] [CrossRef]
- Diseases, B.M.N.I. o. D. a. D. a. K. , LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]-fluorouracil. In livertox.nih.gov: 2018.
- Diseases, B.M.N.I. o. D. a. D. a. K. , LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]- rifampicin. In livertox.nih.gov: 2018.
- Diseases, B.M.N.I. o. D. a. D. a. K. , LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]- ibuprofen. In livertox.nih.gov: 2018.
- Messam, C.A.; Pittman, R.N. Asynchrony and Commitment to Die during Apoptosis. Exp. Cell Res. 1998, 238, 389–398. [Google Scholar] [CrossRef]
- Aragane, Y.; Kulms, D.; Metze, D.; Wilkes, G.; Pöppelmann, B.; Luger, T.A.; Schwarz, T. Ultraviolet Light Induces Apoptosis via Direct Activation of CD95 (Fas/APO-1) Independently of Its Ligand CD95L. J. Cell Biol. 1998, 140, 171–182. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Bernard, S.; Herzel, H. , Why do cells cycle with a 24 hour period? Genome Inform 2006, 17, 72–79. [Google Scholar]
- Nishikawa, S.; Takamatsu, A. Effects of cell death-induced proliferation on a cell competition system. Math. Biosci. 2019, 316, 108241. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, H.D.; Bergmann, A. The Role of Apoptosis-Induced Proliferation for Regeneration and Cancer. Cold Spring Harb. Perspect. Biol. 2012, 4, a008797. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Li, J.; Wei, Y.; Lu, J.; Dong, R. Hepatic Stellate Cell: A Double-Edged Sword in the Liver. Physiol. Res. 2021, 70, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, M.; Yang, C.; Martino, M.; Duncan, M.B.; Rieder, F.; Tanjore, H.; Kalluri, R. Fibroblasts Derive from Hepatocytes in Liver Fibrosis via Epithelial to Mesenchymal Transition. J. Biol. Chem. 2007, 282, 23337–23347. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Wan, C.; Guo, R.; Guo, D. , Is Hydrogen Peroxide a Suitable Apoptosis Inducer for All Cell Types? Biomed Res Int 2016, 2016, 7343965. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Chen, C.; An, J.; Shang, Y.; Li, H.; Xia, H.; Yu, J.; Wang, C.; Liu, Y.; et al. Regulation of TBBPA-induced oxidative stress on mitochondrial apoptosis in L02 cells through the Nrf2 signaling pathway. Chemosphere 2019, 226, 463–471. [Google Scholar] [CrossRef]
- Cho, K.-H.; Choi, S.-M.; Kim, B.-C.; Lee, S.-H.; Park, M.-S.; Kim, M.-K.; Kim, J.-K. 5-fluorouracil-induced oligodendrocyte death and inhibitory effect of cycloheximide, trolox, and Z-VAD-FMK in murine cortical culture. Cancer 2004, 100, 1484–1490. [Google Scholar] [CrossRef]
- Noguchi, M.; Hirata, N.; Tanaka, T.; Suizu, F.; Nakajima, H.; Chiorini, J.A. Autophagy as a modulator of cell death machinery. Cell Death Dis. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. , Ferroptosis: past, present and future. Cell Death Dis 2020, 11, 88. [Google Scholar] [CrossRef]
- Salucci, S.; Burattini, S.; Battistelli, M.; Baldassarri, V.; Maltarello, M.C.; Falcieri, E. Ultraviolet B (UVB) Irradiation-Induced Apoptosis in Various Cell Lineages in Vitro. Int. J. Mol. Sci. 2012, 14, 532–546. [Google Scholar] [CrossRef]
- Aldewachi, H.S.; A Wright, N.; Appleton, D.R.; Watson, A.J. The effect of starvation and refeeding on cell population kinetics in the rat small bowel mucosa. J. Anat. 1975, 119, 105–121. [Google Scholar] [PubMed]
- Goodlad, R.A.; Wright, N.A. The effects of starvation and refeeding on intestinal cell proliferation in the mouse. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1984, 45, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Tessitore, L. Apoptosis and Cell Proliferation Are Involved in the Initiation of Liver Carcinogenesis by a Subnecrogenic Dose of Diethylnitrosamine in Refed Rats. J. Nutr. 2000, 130, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, K.; Jacob, S.T. An alternative molecular mechanism of action of 5-fluorouracil, a potent anticancer drug. Biochem. Pharmacol. 1997, 53, 1569–1575. [Google Scholar] [CrossRef] [PubMed]
- Granada, A.E.; Jiménez, A.; Stewart-Ornstein, J.; Blüthgen, N.; Reber, S.; Jambhekar, A.; Lahav, G. The effects of proliferation status and cell cycle phase on the responses of single cells to chemotherapy. Mol. Biol. Cell 2020, 31, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Gérard, C.; Goldbeter, A. , The balance between cell cycle arrest and cell proliferation: control by the extracellular matrix and by contact inhibition. Interface Focus 2014, 4, 20130075. [Google Scholar] [CrossRef] [PubMed]
- Chitu, V.; Yeung, Y.; Yu, W.; Nandi, S.; Stanley, E.R. Measurement of Macrophage Growth and Differentiation. Curr. Protoc. Immunol. 2011, 92, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Mazaleuskaya, L.L.; Theken, K.N.; Gong, L.; Thorn, C.F.; FitzGerald, G.A.; Altman, R.B.; Klein, T.E. , PharmGKB summary: ibuprofen pathways. Pharmacogenet Genomics 2015, 25, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Peters, G.; Backus, H.; Freemantle, S.; van Triest, B.; Codacci-Pisanelli, G.; van der Wilt, C.; Smid, K.; Lunec, J.; Calvert, A.; Marsh, S.; et al. Induction of thymidylate synthase as a 5-fluorouracil resistance mechanism. Biochim. et Biophys. Acta (BBA) - Mol. Basis Dis. 2002, 1587, 194–205. [Google Scholar] [CrossRef]
- Advani, P.P.; Fakih, M.G. 5-FU-induced hyperammonemic encephalopathy in a case of metastatic rectal adenocarcinoid successfully rechallenged with the fluoropyrimidine analog, capecitabine. Anticancer Res. 2011, 31, 335–338. [Google Scholar]
- Zhang, C.; Wang, H.; Ning, Z.; Xu, L.; Zhuang, L.; Wang, P.; Meng, Z. Serum liver enzymes serve as prognostic factors in patients with intrahepatic cholangiocarcinoma. OncoTargets Ther. 2017, ume 10, 1441–1449. [Google Scholar] [CrossRef]
- Barguilla, I.; Maguer-Satta, V.; Guyot, B.; Pastor, S.; Marcos, R.; Hernández, A. In Vitro Approaches to Determine the Potential Carcinogenic Risk of Environmental Pollutants. Int. J. Mol. Sci. 2023, 24, 7851. [Google Scholar] [CrossRef] [PubMed]
- Audebert, M.; Assmann, A.-S.; Azqueta, A.; Babica, P.; Benfenati, E.; Bortoli, S.; Bouwman, P.; Braeuning, A.; Burgdorf, T.; Coumoul, X.; et al. New approach methodologies to facilitate and improve the hazard assessment of non-genotoxic carcinogens—a PARC project. Front. Toxicol. 2023, 5, 1220998. [Google Scholar] [CrossRef] [PubMed]
- Loftus, L.V.; Amend, S.R.; Pienta, K.J. Interplay between Cell Death and Cell Proliferation Reveals New Strategies for Cancer Therapy. Int. J. Mol. Sci. 2022, 23, 4723. [Google Scholar] [CrossRef] [PubMed]
- Madorran, E.; Stožer, A.; Arsov, Z.; Maver, U.; Rožanc, J. A Promising Method for the Determination of Cell Viability: The Membrane Potential Cell Viability Assay. Cells 2022, 11, 2314. [Google Scholar] [CrossRef] [PubMed]
- Madorran, E.; Stožer, A.; Arsov, Z.; Maver, U.; Rožanc, J. A Promising Method for the Determination of Cell Viability: The Membrane Potential Cell Viability Assay. Cells 2022, 11, 2314. [Google Scholar] [CrossRef] [PubMed]
- Rieger, A.M.; Nelson, K.L.; Konowalchuk, J.D.; Barreda, D.R. , Modified annexin V/propidium iodide apoptosis assay for accurate assessment of cell death. Journal of visualized experiments : JoVE 2011.
- Van Merloo, J.; Kaspers, G.J.; Cloos, J. Cell sensitivity assays: The MTT assay. Methods Mol. Biol. 2011, 731, 237–245. [Google Scholar]
- Luk, H.-Y.; McFarlin, B.K.; Vingren, J.L. Using image-based flow cytometry to monitor satellite cells proliferation and differentiation in vitro. Methods 2017, 112, 175–181. [Google Scholar] [CrossRef]
- Madorran, E.; Kocbek Šaherl, L.; Rakuša, M.; Munda, M. , In Vitro Human Liver Model for Toxicity Assessment with Clinical and Preclinical Instrumentation. In Preprints, Preprints: 2024.
Figure 1.
Viability assessment of the various methods in comparison to cell count method. a) Viability assessment comparison between Cell count (CC), MTT (MTT) and PI methods (PI). b) Viability assessment with the DD Cell-Tox method. c) Cell count of the samples at different time points. (> 0.05, * 0.05, ** 0.01, *** 0.001). CC: Cell count method, MTT: MTT assay, PI: PI method; 5-FU: 5-fluorouracil, FA: Fatty acids, H2O2, NS: Nutrient starvation, TBBPA: tetrabromobisphenol A.
Figure 1.
Viability assessment of the various methods in comparison to cell count method. a) Viability assessment comparison between Cell count (CC), MTT (MTT) and PI methods (PI). b) Viability assessment with the DD Cell-Tox method. c) Cell count of the samples at different time points. (> 0.05, * 0.05, ** 0.01, *** 0.001). CC: Cell count method, MTT: MTT assay, PI: PI method; 5-FU: 5-fluorouracil, FA: Fatty acids, H2O2, NS: Nutrient starvation, TBBPA: tetrabromobisphenol A.
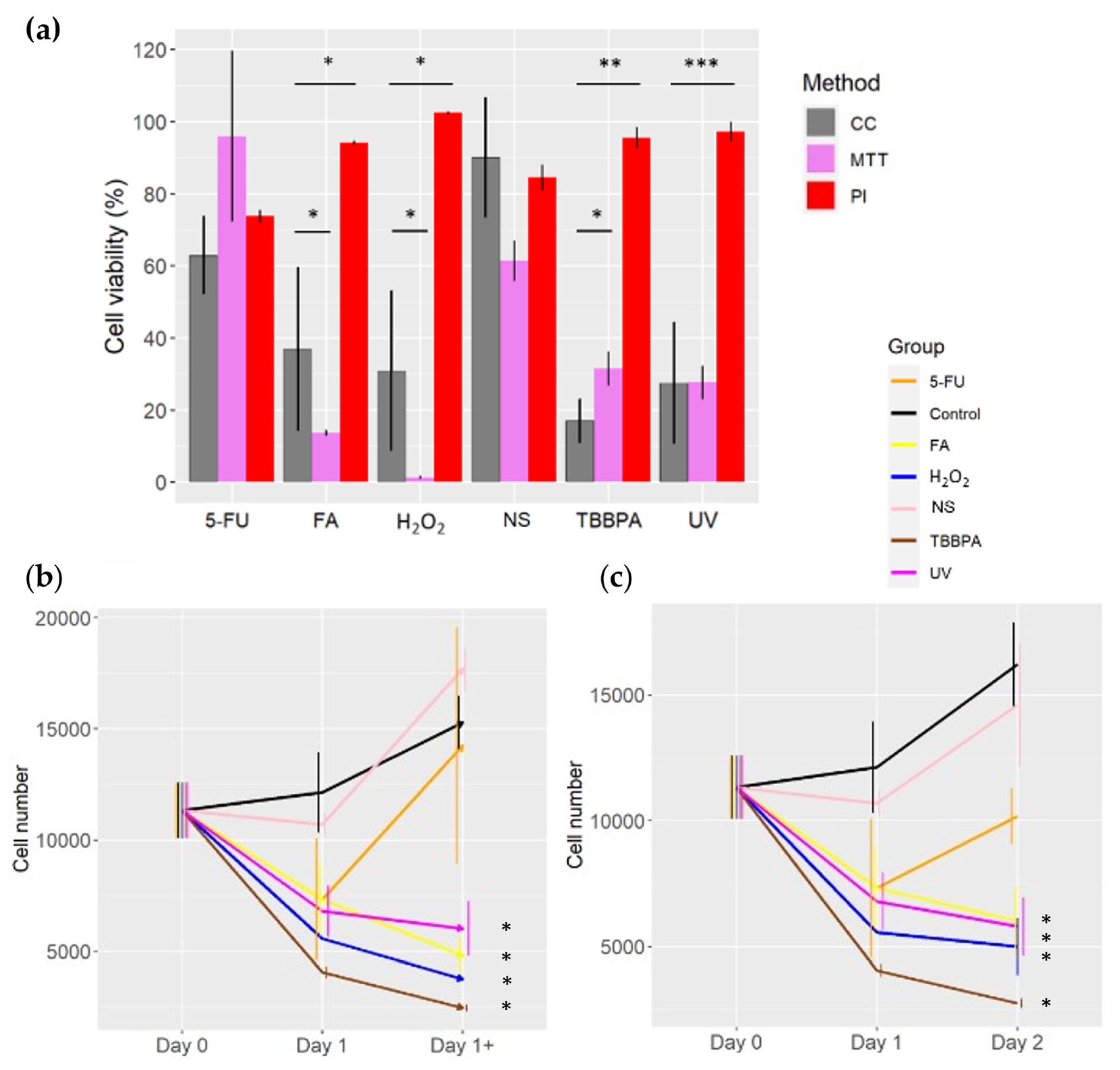
Figure 2.
Comparison between the DD Cell-Tox method toxicity assessment in a liver model and the existing clinical data for RIF, IBU and 5-FU. a) DD Cell-Tox method toxicity evaluation of RIF, IBU and 5-FU. b) ALT, AST and Bilirubin values of patients treated with RIF, IBU and 5-FU. (> 0.05, * 0.05, ** 0.01, *** 0.001). 5-FU: 5-fluorouracil, IBU: ibuprofen, RIF: rifampicin.
Figure 2.
Comparison between the DD Cell-Tox method toxicity assessment in a liver model and the existing clinical data for RIF, IBU and 5-FU. a) DD Cell-Tox method toxicity evaluation of RIF, IBU and 5-FU. b) ALT, AST and Bilirubin values of patients treated with RIF, IBU and 5-FU. (> 0.05, * 0.05, ** 0.01, *** 0.001). 5-FU: 5-fluorouracil, IBU: ibuprofen, RIF: rifampicin.
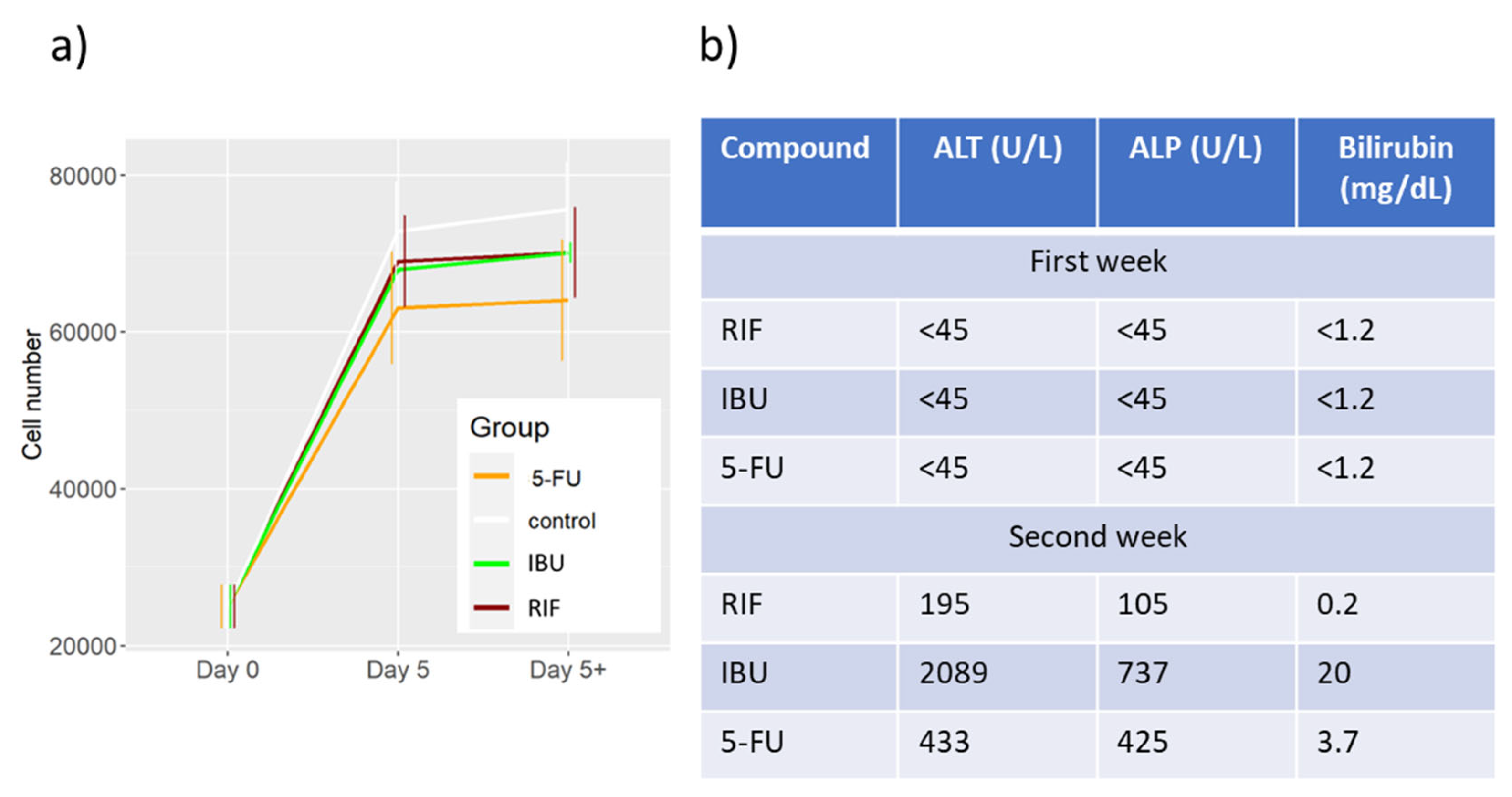
Figure 3.
DD Cell-Tox method toxicity assessment scheme. a) The method’s protocol. b) Cell response measurement with the Imaging flow cytometer MK2 (ISX). c) Diagram (analysis) of the cell dynamics for the toxicity assessment of substances.
Figure 3.
DD Cell-Tox method toxicity assessment scheme. a) The method’s protocol. b) Cell response measurement with the Imaging flow cytometer MK2 (ISX). c) Diagram (analysis) of the cell dynamics for the toxicity assessment of substances.
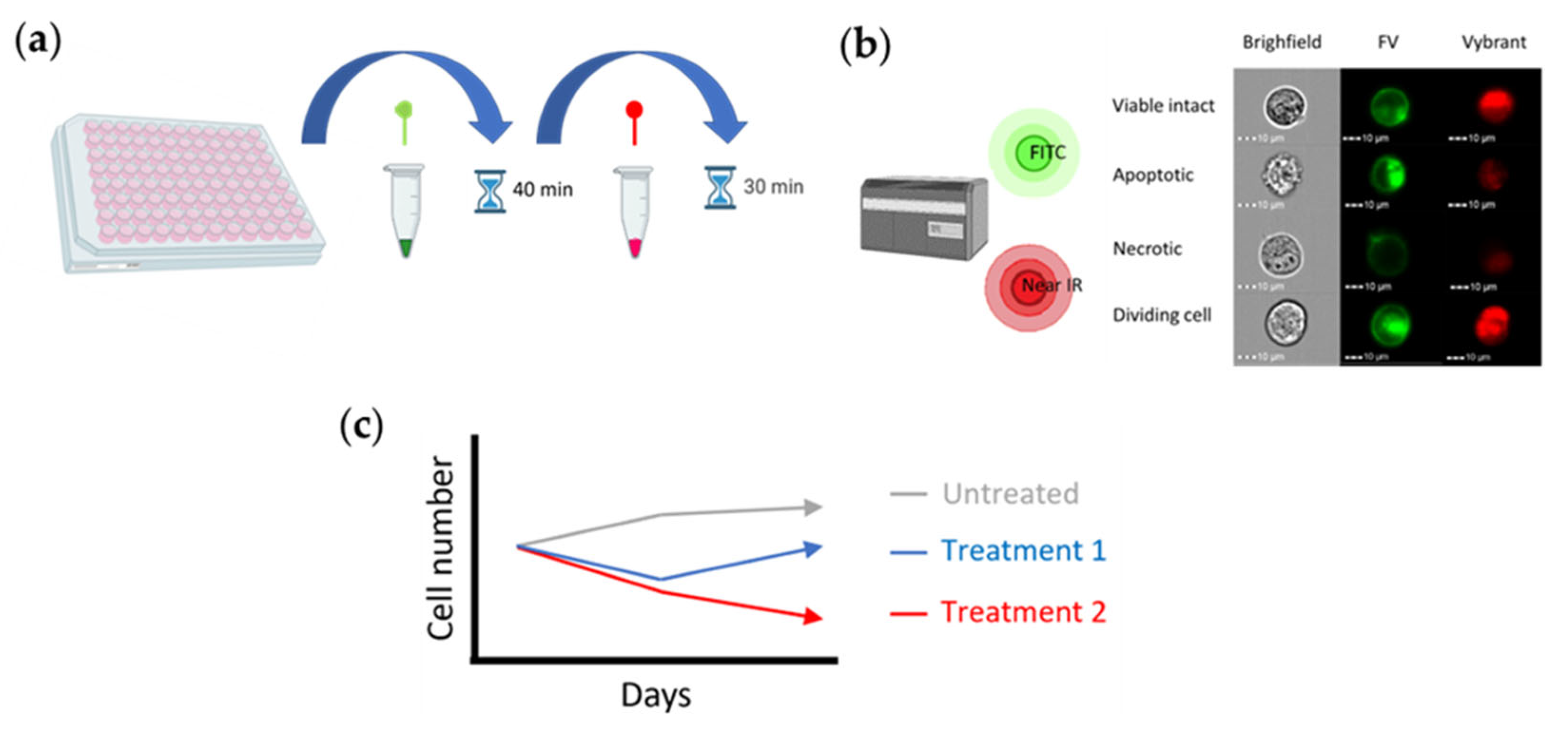
Figure 4.
The experimental setup for DD Cell-Tox accuracy assessment in blood macrophages. a) Cells were seeded and cultured till 100% confluency was achieved. b) After cell attachment, the samples were treated with hydrogen peroxide, TBBPA, 5-FU, FA, nutrient starvation and UV c) 24 hours after the treatment, cell viability was assessed with the MTT assay and the PI method. At the same time, DD Cell-Tox method was performed. The formula to calculate the cell number prediction of the population is depicted in the graph below the formula used for the calculation. d) 48 hours after the treatment, cell viability was assessed with the cell count method. e) The accuracy of the cell count method 48 hours after treatment was compared to the DD Cell-Tox method, MTT assay, and the PI method. 5-FU: 5-fluorouracil, FA: Fatty acids, H2O2, NS: Nutrient starvation, TBBPA: tetrabromobisphenol A.
Figure 4.
The experimental setup for DD Cell-Tox accuracy assessment in blood macrophages. a) Cells were seeded and cultured till 100% confluency was achieved. b) After cell attachment, the samples were treated with hydrogen peroxide, TBBPA, 5-FU, FA, nutrient starvation and UV c) 24 hours after the treatment, cell viability was assessed with the MTT assay and the PI method. At the same time, DD Cell-Tox method was performed. The formula to calculate the cell number prediction of the population is depicted in the graph below the formula used for the calculation. d) 48 hours after the treatment, cell viability was assessed with the cell count method. e) The accuracy of the cell count method 48 hours after treatment was compared to the DD Cell-Tox method, MTT assay, and the PI method. 5-FU: 5-fluorouracil, FA: Fatty acids, H2O2, NS: Nutrient starvation, TBBPA: tetrabromobisphenol A.
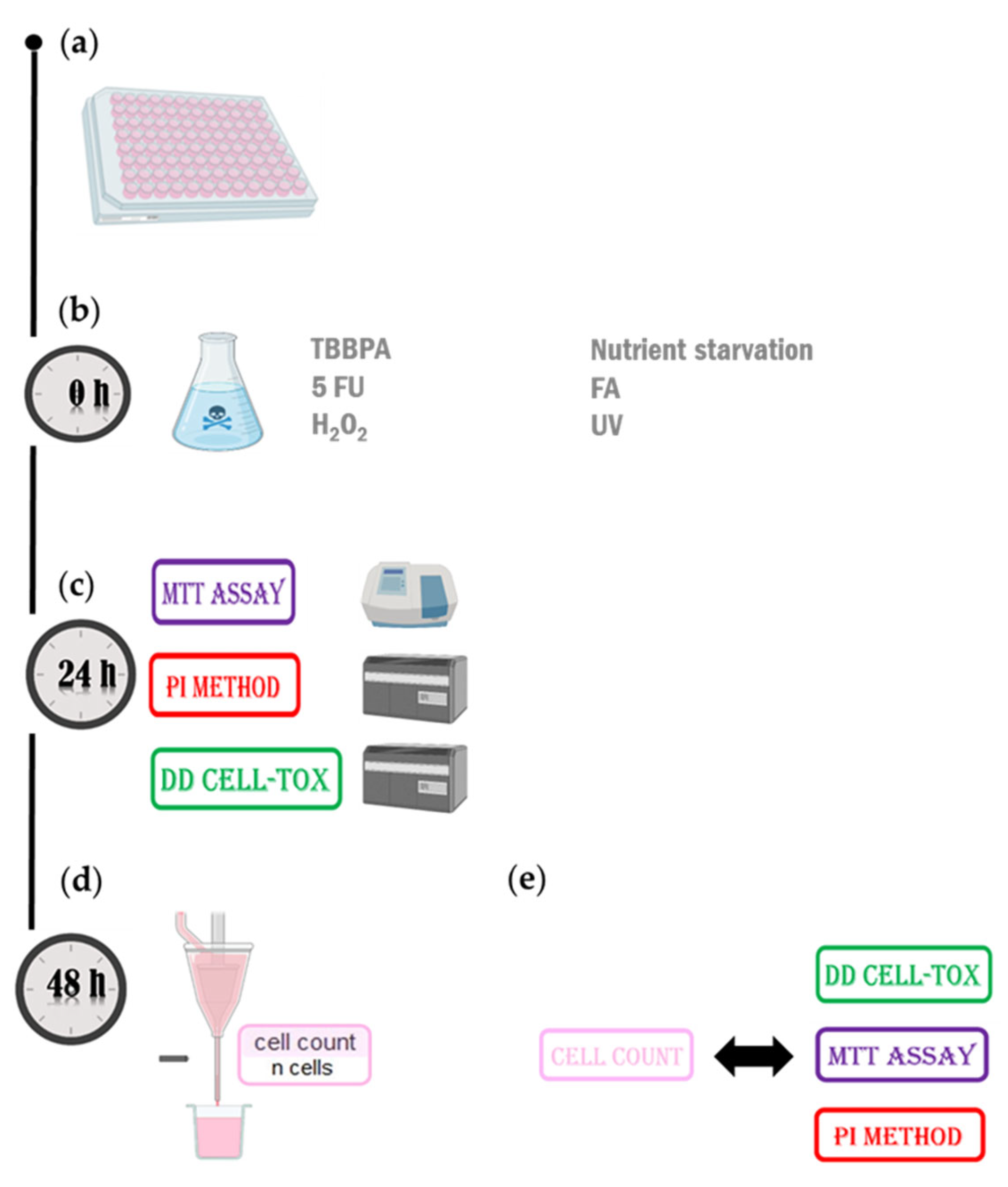
Figure 5.
The experimental setup for DD Cell-Tox accuracy assessment in a cell-based liver model. a) We designed a liver model with four cell types: LSEC, HSC, Kupffer cells and hepatocytes. b) Brightfield image at 20x magnification obtained with an inverted microscope (Axiovert 40 CFL, ZEISS, Germany). c) We exposed the liver model to RIF, IBU, and 5-FU for 5 days (we renew the medium after 2 days). d) We measured the cell viability with the DD Cell-Tox method. 5-FU: 5-fluorouracil, IBU: ibuprofen, RIF: rifampicin.
Figure 5.
The experimental setup for DD Cell-Tox accuracy assessment in a cell-based liver model. a) We designed a liver model with four cell types: LSEC, HSC, Kupffer cells and hepatocytes. b) Brightfield image at 20x magnification obtained with an inverted microscope (Axiovert 40 CFL, ZEISS, Germany). c) We exposed the liver model to RIF, IBU, and 5-FU for 5 days (we renew the medium after 2 days). d) We measured the cell viability with the DD Cell-Tox method. 5-FU: 5-fluorouracil, IBU: ibuprofen, RIF: rifampicin.
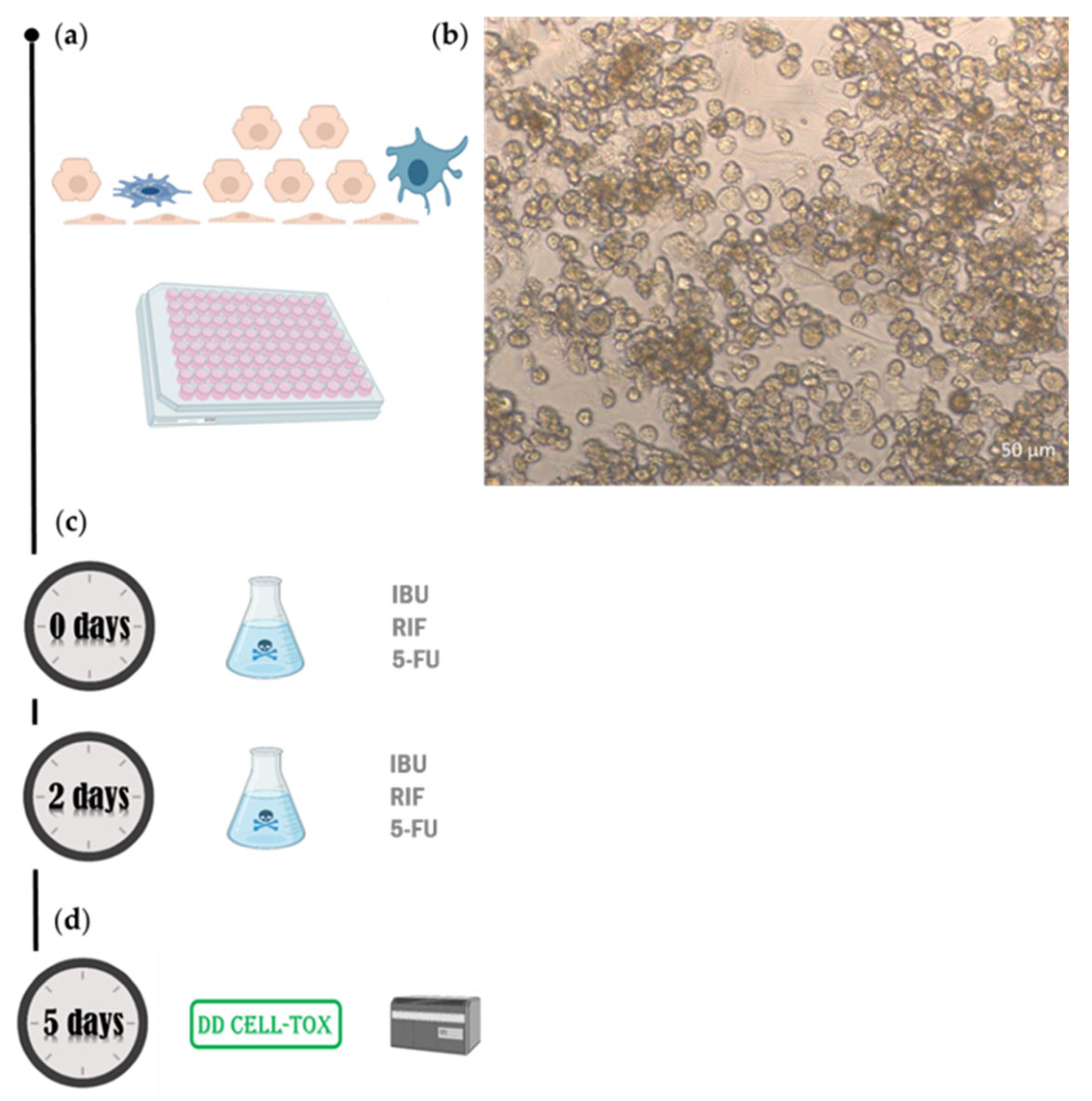

Table 1.
Summarizing the effect of contaminants in TLT.
| Group | Number of initial cells | Number of final cells | Dead cells (%) | Division cells (%) |
|---|---|---|---|---|
| Control | 11329 | 12130 | 12 | 20 |
| 5-FU | 11329 | 7327 | 7 | 26 |
| FA | 11329 | 7318 | 35 | 0 |
| H2O2 | 11329 | 5564 | 33 | 0 |
| NS | 11329 | 10694 | 12 | 39 |
| TBBPA | 11329 | 4064 | 45 | 3 |
| UV | 11329 | 6797 | 26 | 7 |
5-FU: 5-fluorouracil, FA: Fatty acids, H2O2, NS: Nutrient starvation, TBBPA: tetrabromobisphenol A.
Table 2.
Summary of the DD cell-Tox method toxicity evaluation of three well-known hepatotoxic drugs.
Table 2.
Summary of the DD cell-Tox method toxicity evaluation of three well-known hepatotoxic drugs.
| Group | Number of initial cells | Number of final cells | Cell number increase (%) | Dead cells (%) | Division cells (%) |
|---|---|---|---|---|---|
| Control | 25000 | 72797 | 291 | 3 | 2 |
| 5-FU | 25000 | 63056 | 252 | 4 | 3 |
| IBU | 25000 | 67919 | 272 | 3 | 4 |
| RIF | 25000 | 68995 | 276 | 2 | 2 |
5-FU: 5-fluorouracil, IBU: ibuprofen, RIF: rifampicin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



