
Submitted:
18 April 2024
Posted:
19 April 2024
You are already at the latest version
Abstract
Parkinson's disease (PD) is recognized as the second most prevalent primary chronic neurodegenerative disorder of the central nervous system. Clinically, PD is characterized as a movement disorder, exhibiting an incidence and mortality rate that is increasing faster than any other neurological condition. In recent years, there has been a growing interest concerning the role of the gut microbiome in the etiology and pathophysiology of PD. The establishment of a brain-gut microbiome axis is now real, with evidence denoting a bidirectional communication between the brain and the gut microbiota through metabolic, immune, neuronal, and endocrine mechanisms and pathways. Among these, the vagus nerve represents the most direct form of communication between the brain and the gut. Given the potential interactions between bacteria and drugs, it has been observed that the therapies for PD can have an impact on the composition of the microbiome. Therefore, in the scope of the present review, we will discuss the current understanding of gut microbiome on PD and whether this may be a new paradigm for treating this devastating disease.
Keywords:
Parkinson’s Disease
; Microbiome
; Gut-Brain Axis
1. Introduction
Parkinson’s disease (PD) is the second most common neurodegenerative disease of the central nervous system (CNS)[1,2]. Currently, this movement disorder affects 10 million people worldwide, with 75,000 new cases per year [3,4]. According to the World Health Organization (WHO), PD's incidence and death rate is increasing faster than any other neurological condition, and its prevalence has doubled over the past 25 years [5].
PD is a complex and multifactorial disease, presenting distinctive pathological hallmarks, including the depletion of dopaminergic neurons in the substantia nigra pars compacta (SNpc). This depletion arises from the accumulation of a pathological misfolding alpha-synuclein (αSyn) protein, leading to the formation of Lewy neurites in the (remaining) neurons. αSyn is a monomeric protein and thus is expected to undergo genetic and post-translational modifications, including phosphorylation, ubiquitination, nitration, and oxidation, which can predispose it to misfold [2,6]. This misfolding makes this protein insoluble since it tends to form β-sheet-rich amyloid aggregates that accumulate and form intracellular eosinophilic inclusions [1,3]. Consequently, these changes can significantly impact the correct functioning of the central, peripheral, and enteric nervous systems (ENS) [1,3,7,8]. In fact, the presence of αSyn aggregates, combined with other factors, such as mitochondrial dysfunction, ubiquitin-proteasome system (UPS), synaptic dysfunction, e.g., contributes to neuronal degeneration and death, leading to the appearance of motor (slow movements, resting tremors, rigidity, postural instability, and bradykinesia) and non-motor (dementia, gastrointestinal dysfunction, behavioral and cognitive impairments, among others) symptoms [1,6,7,9,10,11,12].
Despite all this knowledge, the etiology of PD is still unclear, although genetic and environmental factors provide the most plausible explanation for the onset of this pathology [7]. From the clinical point of view, PD displays heterogeneity, with different onset and progression patterns [13,14]. Advanced age at diagnosis is the most critical risk factor, although young Parkinsonism (even if rare) cannot be excluded. Nevertheless, in addition to aging, other risk factors should also be considered, namely family history of the disease, gender (in which men are more affected than women), and environmental factors such as exposure to pesticides and rural living [1,6]. Notwithstanding, it is known that cellular and molecular alterations can also contribute to PD through high levels of oxidative stress, mitochondrial dysfunction, neuroinflammation, xenobiotic toxins, and altered dopamine metabolism [6].
At the moment, both pharmacological and non-pharmacological therapies are available for PD. The current gold-standard PD treatment is pharmacological, based on dopamine replacement. However, non-pharmacological alternative approaches such as deep brain stimulation (DBS), an established safe neurosurgical symptomatic therapy, can be used in certain patients in more advanced stages of the disease, in which the treatment had no effectiveness or in which the medication itself produced severe side effects [13,14]. Nevertheless, these treatments promote temporary relief, being ineffective all over time, leading to the necessity of increasing drug dosages (in case of pharmacological therapies) or DBS intensities (in case of non-pharmacological approaches), which in most cases results in the appearance of adverse effects such as sleep disturbance, impulsivity, and addiction [1,4]. Hence, addressing this remains a noteworthy constraint in managing PD and a significant obstacle in developing or repurposing innovative therapies to slow down or halt its progression [17]. Moreover, it poses the question of how we can effectively manage PD and determine the optimal target for its treatment?
Despite numerous other avenues of research, in recent years, significant emphasis has been placed on exploring the vast potential of the gut microbiome and its intricate interconnection. As a result, efforts have been made to comprehend how the gut microbiome may respond under PD conditions, specifically whether it may contribute to the development or progression of the disease or offer a new therapeutic target to address PD. Therefore, in the present review, we will explore the bidirectional association between the gut microbiome and PD, discussing the current understanding of the gut microbiome in the pathophysiology of PD and as a potential target for novel therapeutic approaches.
Gut Microbiome
The human body is over 99% microbial in terms of genes. Our gut holds almost 100 trillion microbes, meaning there are 10-100 times more bacteria than eukaryotic cells in the human body [10,18]. Nevertheless, bacterial composition and diversity differ according to different parts of the body [6]. The colonization of the gut is thought to begin immediately after birth, and it can be influenced by various factors such as the method of removing the fetus, namely vaginal or caesarean, or the type of breastfeeding (breastfeeding or formula), environmental stimuli, and antibiotic use [19,20,21]. Remarkably, this colonization is considered in the first 3-5 years of life. Although it is generally stable within healthy adult individuals throughout life, it is a dynamic entity whose composition is susceptible to change due to external disturbances [6,22]. Factors such as genetics, stress, environmental exposures, age, metabolism, geography, surgeries, physical activity, antibiotic intake, and diet can influence the stability of human gut microbiota [21,22].
Microbiota is the complex ecological community composed of all the microorganisms that reside in our gut, and it has been called microbiome to the entire genetic material (genome) of the microbiota [21]. Specifically, microbiota comprises bacteria, eukaryotic cells, viruses, archaeobacteria, fungi, and protozoa [23]. These microbiota constituents interact with each other and the host, impacting health through their metabolic activities and host interactions and in normal human physiology and disease contexts [6,18]. Bacteria predominately control the gut microbiome's metabolic activities, such as modulation of energy metabolism, nutrition absorption, and regulation of gut microbiota composition [24]. Thus, gut microbes can play a harmless or beneficial effect on the host, being essential to keep the gut homeostasis [4,25]. In addition, the microbiota interacts with the immune system, facilitating the normal development of immune functions during homeostasis and the emission of signals to promote the maturation of immune cells [25].
The adult microbiota harbors more than 1,000 phylotypes of bacteria at the species level, with Firmicutes and Bacteroides emerging as the two most abundant phyla, representing at least 70-75% of the total number of microorganisms [26]. Actinobacteria, Proteobacteria, and Verrucomicrobia can also be found, albeit in a smaller amount [22,27]. Scientific reports have been highlighting that it is essential to understand the stability of the microbiota within an individual over time to predict health status and to be able to develop therapies to correct dysbiosis, the term used to define any imbalance between the bacterial community and the host. Indeed, gut dysbiosis is associated with several pathologies, including systemic or neurological disorders [18,27]. Accordingly, recent studies have claimed that gut microbiome can be an important piece of evidence to understand human pathology and physiology better [3]. Along with that, even though there was variability in adult gut communities, certain bacterial populations were common across different groups of individuals. Thus, a concept called “enterotype” emerged, which has been used to categorize individuals according to the composition of their intestinal microbiota [7]. Based on this concept, it was then proposed that three distinct symbiotic host-microbial states could be formed, driven by groups of dominant genera, namely Bacteroides (this has recently been subdivided into two groups: B1 and B2), Prevotella, or Ruminococcus. However, it should be noted that this concept of enterotype remains to be fully addressed [7,18,20,22]. Nevertheless, several factors may also play a role in this distinction and characterization. For instance, there is considerable intra- and inter-individual diversity in the microbiota composition of healthy control subjects and in the context of multiple disorders, where intestinal dysbiosis has been described, indicating that the gut microbiota is not uniform across populations. Environmental factors (e.g., smoking, alcohol, and diet), which significantly influence the structure and composition of microbial communities, can be used to explain this heterogeneity [7,28]. Besides age, genetics, environment, and lifestyle be factors affecting microbiota (see Figure 1), these are also critical factors contributing to PD onset and modulation throughout life [27].
Age
Microbiota colonization begins immediately after birth and plays an essential role in developing the ENS and postnatal immune system [4,7]. This colonization process is primarily influenced by various perinatal conditions such as the type of birth, nutrition, and the use or non-use of medication in the initial stages [29]. During the first years of life, the gut microbiota experiences various compositional and functional changes, which can significantly impact individual susceptibility to immune-related diseases in adulthood [7]. In the case of neurodegenerative diseases, aging is a well-defined hallmark associated with the accumulation of mitochondrial DNA defects, oxidative damage, and neuromelanin [27,30]. Furthermore, it is also recognized that problems caused by the gut or gut bacteria that eventually lead to dysbiosis have a higher frequency in older people [3,27].
Genetics
PD is mainly considered an idiopathic condition, with only approximately 10-15% of cases having a hereditary component associated with several genetic mutations [31]. Extensive research has been focusing on SNCA (αSyn), LRRK2 (leucine-rich repeat kinase-2), PINK-1 (phosphatase and tensin [PTEN] homologue-induced kinase-1), and PARK genes [32,33]. SNCA gene encodes for αSyn, and its mutations impact the protein’s expression levels, thus becoming a risk factor for PD. This gene has been widely associated with increased opportunistic pathogens in the intestine of PD patients. Interestingly, mutations in SNCA are not the prevalent ones in PD patients. Indeed, LRRK-2 is the most frequent genetic cause of the autosomal dominant form of PD, accounting for 10–40% of familial cases in various ethnicities [34]. Mutations in this enzyme are also closely associated with inflammatory bowel diseases (IBDs) [34,35].Regarding the PINK-1 gene, it plays a role in the removal of damaged mitochondria, being mutations in this gene associated with mitochondrial dysfunction seen in the autosomal recessive early-onset PD [36]. The PARK gene encodes the Parkin protein, which can undergo several mutations resulting in a cellular accumulation of damaged mitochondria [32]. In addition, mutations in this protein are also known to disrupt the ubiquitin-proteasome pathway, which ultimately triggers the accumulation of misfolded protein aggregates (e.g., αSyn) [32,37]. Curiously, exposure to pathogenic bacteria or bacterial metabolites was shown to cause epigenetic changes in the genes mentioned above. Hence, this interaction could be involved in most sporadic cases of PD. However, the relationship between genetic predisposition and the bacterial community in a PD context remains poorly understood [6].
Environment
Increased xenobiotic degradation in the gut, particularly herbicides and pesticides, has been found in the gut of PD patients [38,39]. Preclinical studies have demonstrated that exposure to these compounds can induce the death of dopaminergic neurons, leading to movement impairments in mice [6,40]. Still, considering the wide variety of pesticides in use, their long half-life, and the potential for accumulation in food chains, the associated risk may be even greater [41]. Therefore, more extensive studies should be conducted to approach this potential interconnection and how this may interplay with microbiome and neurologic diseases.
Lifestyle
Although the three factors mentioned above (age, genetics, and environment) have major effects on the pathophysiology of PD, lifestyle can also play a role in PD, in which diet, physical exercise, caffeine, and nicotine consumption could be contributors (Figure 2) [41]. Despite all the information regarding possible microbiome modulators and their impact on such disorders, a fundamental question remains: how can microbiome data analysis be correlated with a diseased state?
The main challenge in addressing this question lies in several perspectives, namely in the heterogeneity profile between patients and diseases and in the technological availability to deeply discriminate the inherent variances. The conjugation between advanced techniques (proteomics, metabolomics, transcriptomics) and bioinformatic tools will be crucial to analyzing and integrating large amounts of data, based on the idea that only then will it be possible to capture the multilayered structure of the data.
Microbiome Characterization Techniques
The study of the human microbiome diversity began in the 1680s, when Anton Van Leeuwenhoek compared the oral and fecal microbiomes and found that microorganisms vary not only from region to region but also between states of health or disease regardless of location [42,43,44]. The initial investigations into the microbiome, namely the identification and characterization of microorganisms, were conducted through methodologies focused on cultivating and isolating bacteria [45]. Nevertheless, it quickly became evident that all existing species' cultivation and phenotypic characterization were unfeasible, either due to a lack of favorable conditions or to slow bacterial growth [46]. Nevertheless, developing novel analytical and sequencing techniques for studying the human microbiome has become a priority [44]. Indeed, the study of the microbiome has been a significant focus of various diseases in recent years, with an increasing number of areas recognizing the microbiome as a "Key Player" in several pathological conditions [47,48,49]. Presently, a multitude of molecular biological analysis tools can be employed in the study of the gut microbiome, from conventional to more advanced techniques, as outlined in Table 1 [50,51,52]. Although the development of traditional methods has been essential to the initial investigations of the gut microbiome, the development of novel methodologies, such as microbiome shotgun sequencing, has made it possible to overcome the biases associated with traditional culture, thus enabling a better understanding of the composition, diversity, and interrelationships among the microorganisms constituting the microbiome, contributing significantly to our understanding of their role in states of health or disease [52].
The microbiome is studied by analyzing the structure and dynamics of bacterial communities and the interactions that they can establish with each other [72]. Regardless of the chosen analytical technique, the study comprises four essential phases (Figure 1): 1) fecal sample collection, 2) DNA extraction, 3) DNA analysis (utilizing the most suitable technique for the intended purpose (techniques outlined in Table 1)), and 4) bioinformatic analysis of the results obtained. These analyses can be done to obtain a taxonomy profiling or else analysis of gene functions of the gut microbiome [50,73]. Subsequently, correlations between the healthy and altered gut microbiome in a pathological context can be made [50,74,75].
Bearing this in mind, to perform microbiome research accurately, it is essential that, in combination with standard methodologies, appropriate pipelines (such as Quantitative Insights Into Microbial Ecology (QIIME) [76], and the Metagenomic Rapid An- notation using Subsystem Technology (MG-RAST) [77] should be established for research regarding the human gut microbiome [45,50,78]. This is crucial since methodological differences could lead to inconsistent data, thus limiting knowledge in this area [50,79,80]. Projections indicate that, in the forthcoming years, analyses of the gut microbiome could yield pivotal insights, facilitating the provision of personalized medicine for different diseases and thereby exerting a profound impact on individual human health [81,82,83].
Despite the substantial growth in this area in recent years, it is still impossible to have direct correlations between the microorganisms in our gut microbiome and their role in the onset and progression of specific pathologies, such as PD [52,84]. Consequently, there is a paramount need to allocate resources towards comprehending intestinal microorganisms' diversity and their bioactive metabolites' functionality in the context of diverse diseases, including PD [85,86]. This strategic investment is essential for the eventual utilization of the intestinal microbiome in diagnosing and treating PD.
Microbiome and PD: From Diagnosis to Treatment
Exposure to pathogenic bacteria, or bacterial metabolites, is one of the factors shown to cause epigenetic changes when they interact with (PD) genes [6]. It has been proposed that this interaction may be involved in most sporadic cases of PD. However, the relationship between genetic predisposition and the gut bacterial community in PD needs to be better understood [6]. These features lead to a particular interest in exploring the gut microbiome when studying PD. From a scientific and clinical perspective, this holds a vast potential for diagnosis, prognosis, and even for understanding disease pathogenesis, as dysbiosis can induce peripheral inflammation, which, in turn, may contribute to the pathophysiology of PD through humoral and neural pathways [34,87]. Actually, numerous studies have revealed a correlation between the prevalence of specific bacterial taxa and the longevity of the disease, severity, motor and non-motor symptom scores, and the use of antiparkinsonian therapies [6]. Additionally, thirteen studies conducted across three continents have demonstrated differences in the gut bacterial community between PD patients and healthy individuals [88].
This bidirectional communication between the brain and the gut microbiota, known as the Brain-Gut-Microbiome Axis (GBrA), refers to the interaction between the gut microbes and the CNS via metabolic, immune, neuronal, and endocrine signaling pathways and mechanisms (Figure 2) [89,90]. Still, one pathway is offering a more direct form of communication: the vagus nerve [3,4,6,89,91]. The vagus nerve is a complex bidirectional system that allows the communication between the hypothalamic-pituitary-adrenal axis hormones, the CNS, the autonomic nervous system, and the ENS [4,27]. Modifications in this axis have been postulated to impact the development of PD directly [3,4]. The discovery of αSyn aggregates in peripheral locations, such as the ENS, supports the concept of the "GBrA". The ENS, which innervates the gastrointestinal (GI) system and is located near the intestinal lumen, provides a significant region for interaction with bacteria [6,7]. From the gut to the brain, there is an ascending aggregation of αSyn and Lewy Bodies (LB) formation, leading to a progressive neurodegeneration [4]. Having mentioned this, a study conducted by Sampson et al. underscores the significance of GBrA in the pathogenesis of PD [92]. The research involving transgenic mice with PD reveals that alterations in the intestinal composition contribute to the disease's pathogenesis [92]. Notably, αSyn aggregates demonstrate a propensity to readily spread from the enteric nervous system to the brain, shedding light on the pivotal role of the GBrA in the progression of PD [4,92].
In this way, it can be assumed that the intestinal microbiome profoundly influences this entire neuronal network, facilitating the absorption of nutrients, vitamins, and medications while modulating the immune system [4]. In 2017, Stolzenberg et al. demonstrated a positive correlation between inflammation of the intestinal wall in pediatric patients and the expression of αSyn in the ENS [93]. This finding suggests an inflammatory response that precedes GI infections commonly associated with the pathogenesis of PD [94]. Additionally, it further indicates that the expression of this protein aggregates within the ENS is part of its normal immune defense mechanism, which supports the hypothesis of Braak et al. [6,93].
Microbiome vs αSyn – Potential Contributions of the Microbiome to PD: Braak’s Hypothesis
Interestingly, PD not only affects the brain, but recent studies have also demonstrated that the disease extends to other organs, namely belonging to the GI system [34]. In 2003, Braak and colleagues postulated a “dual-hit hypothesis” suggesting that αSyn aggregation, triggered by a neurotrophic agent or an unknown pathogen, propagates in a prion-like manner from the ENS and the olfactory bulb to the CNS via the dorsal motor nucleus of the vagus nerve [4,27,91,95,96]. This invasion creates a pro-inflammatory environment, increasing the permeability of the intestinal barrier and leading to reactive oxygen species (ROS) accumulation. Consequently, this will result in a dysregulation in homeostasis, activating various immunological mechanisms that might promote αSyn aggregation [34,97]. Based on such postulation, Braak et al. proposed a six-stage progression system for PD in the brain and in surrounding olfactory regions based on the propagation patterns of αSyn, which can be further correlated with several characteristic hallmarks of the disease [34,98]. With this in mind, Holmqvist et al. conducted studies that provided direct evidence supporting Braak’s hypothesis [8]. This study observed that all forms of αSyn (monomers, oligomers, and fibrils) were actively transported from the gut to the brain through the dorsal motor of the vagus nerve. To support this theory, the same authors employed human brain lysate from PD patients containing various forms of αSyn and recombinant αSyn in an in vivo model (Adult wild-type Sprague Dawley rats) [8]. In both situations, it was deduced that αSyn was transported from the gut to the brain via the vagus nerve [8]. Following such pieces of evidence, S. Kim et al., upon injection of misfolded αSyn into the gut of healthy mice, discovered an accumulation of this protein in both the vagus nerve and the brain. This finding points to the fact that PD may originate in the gut in specific subsets of patients [91,99]. Yet, this assumption remains under debate.
Given the divergent nature of the symptoms associated with PD, these findings support the hypothesis that this disease may be divided into two variants. One variant postulates that PD originates in the gut, explaining why some patients initially present intestinal discomfort problems. The other variant suggests that the disease initiates in the brain, being justified by the early onset of neurological symptoms [91,100].
Gut Microbiome and Neuroinflammation
The cell structure of microbes varies, presenting a tendency to initiate signaling pathways for pattern recognition receptors that can cause inflammation [4]. Inflammation is widely recognized as a prominent pathophysiological characteristic of PD [9]. Patients with PD have been observed to display heightened levels of several inflammatory molecules (IL-1β (Interleukin-1 beta), IL-6 (Interleukin-6), INF-γ (interferon-γ), MCP-1 (Monocyte chemoattractant protein-1) and TNF-α (Tumor necrosis factor- α)) in their bloodstream, cerebrospinal fluid (CSF), and brain [101]. (Neuro)Inflammation is widely recognized as a prominent pathophysiological characteristic of PD, being a major contributor to disease progression and severity, so it can be assumed that it typically develops due to protein aggregation and dopaminergic cell death [9,27,88]. Indeed, several studies have demonstrated the ability of αSyn to stimulate the production of pro-inflammatory cytokines in both microglia and monocytes [6]. Nevertheless, it should be noted that the same inflammatory markers are not always implicated in PD. Even so, some specific cytokines and chemokines are frequently dysregulated (in the brain, CSF and blood) such as IL-6, TNF (Tumor necrosis factor), IL-1 (Interleukin-1), CRP (C-reactive protein), IL-10 (Interleukin 10), CCL5 (Chemokine (C-C motif) ligand 5), INF-γ, and IL-2 (Interleukin-2) [4,27]. This inflammatory process may be triggered by a breakdown in intestinal barrier function (caused, for example, by bacterial infections, oxidative stress, and dysbiosis), leading to increased systemic exposure to inflammatory microbial products, causing αSyn deposition, a characteristic of intestinal hyper-permeability in PD (Figure 2) [4,6,88,102]. This may be related to the etiology and symptomatology of PD [95]. Furthermore, systemic inflammation can disrupt the blood-brain barrier (BBB) permeability, resulting in inflammatory cytokines or systemic immune cells' entrance into the CNS. As a result, the association between low-grade inflammation in the gut and PD has gained considerable attention [27,88].
The GI tract is coated on the inside by the intestinal mucosa, which is a physical and immunological barrier that separates the external environment from the host’s bloodstream. Numerous factors can compromise this barrier's proper functioning and permeability, including bacteria and their metabolic by-products. For instance, disruption of the bacterial balance causes an increase in permeability, subsequently triggering an inflammatory intestinal response, which leads to a state of (neuro)inflammation [6]. Indeed, comparative studies have shown that individuals with PD exhibit an overabundance of a pro-inflammatory profile (such as Streptococcus, Bifidobacterium, Lactobacillus, Akkermansia, and Desulfovibrio) in their gut microbiota, along with increased expression of genes related to pro-inflammatory cytokines and chemokines in their intestinal tissue, compared to control subjects [27,88,103]. In addition, a significant population of neurons near the intestinal epithelium directly connects to the brainstem via the vagus nerve [27]. In a study conducted by Sampson et al. in 2016, oral administration of microbial metabolites into germ-free mice promotes neuroinflammation (αSyn-Dependent Microglia Activation) and subsequently led to the manifestation of characteristic motor symptoms of PD [34,92]. Nevertheless, the role of inflammation is still a matter of discussion in PD since studies defend that it can exert a dual role, being either neurotoxic or neuroprotective. [9].
Molecular Mediators: Toll-Like Receptors (TLR)
The vagus nerve is frequently involved in how the microbiota influences the CNS, specifically through neuroimmune and neuroendocrine mechanisms [89,104]. This communication is facilitated by microbe-derived molecules known as modulators that interact with enteroendocrine cells (EECs), enterochromaffin cells (ECCs), and the mucosal immune system to propagate signals [89,105]. TLRs are innate immune receptors expressed in microglia, immune and non-immune cells [106]. They are capable of recognizing various exogenous and endogenous stimuli to mediate inflammatory responses [106,107]. Most studies indicated an increased expression of TLRs in the brain and peripheral blood cells of PD samples (TLR2, TLR4, and TLR9 having the most prominent roles) [106,107]. We can hypothesize that by increasing both TLRs and inflammatory cytokines (whose function is to serve as endogenous damage-associated molecular patterns (DAMPs)), αSyn may directly impact microglial cells [89,108]. The signaling of TLRs triggers NF-κB activation, which is essential for an increase in nitric oxide production and dopaminergic neuron apoptosis [6,109]. In fact, according to some researchers, when gut dysbiosis occurs, the barrier becomes dysfunctional, activating TLRs, which subsequently recognize bacterial lipopolysaccharide (LPS) [110]. The presence of LPS impairs the function of the intestinal barrier and promotes the production of various pro-inflammatory cytokines that will cross the BBB, thereby inducing a state of neuroinflammation and consequently leading to neuronal cell death in PD [6,110].
Molecular Mediators: LPS and Lipopolysaccharide-Binding Protein (LBP)
Maintaining the integrity of the intestinal barrier is essential for regulating how luminal substances travel across the gut wall, both through paracellular and transcellular pathways. As previously mentioned, a compromised intestinal barrier increases the susceptibility of systemic circulation of inflammatory microbial products such as LPS [27,88]. This leads to intestinal inflammation and oxidative stress, further disrupting mucosal permeability and triggering the aggregation of αSyn in the ENS [4]. Actually, LPS is the major component of the outer surface membrane specific to gram-negative bacteria [111]. Studies regarding the blood and plasma of PD patients have shown increased systemic exposure to LPS, supporting the hypothesis that PD is associated with intestinal barrier dysfunction [88]. Given that oscillations between LPS and LBP levels occur, there have been indications assuming this is a potential marker for dysbiosis [27]. For instance, a study by Paul M. Carvey et al. showed that prenatal exposure to LPS leads to long-term loss of dopaminergic neurons in SNpc of a PD rat model. [112]. Such finding is in line with the study of Adam Jangula and colleagues, which revealed that the expression of αSyn enhances LPS-induced BBB permeability in preclinical models of PD (Snca−/− (KO) mice) [113].
Metabolic Mediators: Short-Chain Fatty Acids (SCFAs)
With the help of EECs and ECCs, SCFAs play a crucial role as neuroactive molecules in the communication between host and microbes [6,89]. The concentration and distribution of SCFA types significantly influence gut health and homeostatic functions, such as mucin production, tight-junction formation, glucose homeostasis, and immunomodulation [4,114]. From a physiological perspective, these compounds are produced in the gut through bacterial metabolism of carbohydrates and protein, serving as an energy source for microbial growth and proliferation [6,88]. The three main types of SCFAs are acetic, propionic, and butyric acid [88]. Nevertheless, in environments with high pH levels, SCFAs may exist as salts, which limits their penetration through the bacterial cell wall [89].
One of the primary mediators contributing to the impact of the gut on PD symptoms is the reduced concentrations of a specific SCFA, butyrate [95]. Such decreased levels of these SCFAs in the intestine severely affect the integrity and function of the intestinal barrier, promoting inflammation and increasing the risk of αSyn deposition [6]. Thus, when a host has a diet low in fermentable fiber, the microbes nourish themselves on mucus glycans using alternative energy sources, which are, in turn, less favorable. As a result, fermentative activity decreases, reducing SCFA production [89]. Butyrate exerts its effects through multiple mechanisms, including activating specific receptors and inhibiting the histone deacetylase (HDAC) enzyme [115]. By inhibiting HDAC, butyrate can regulate gene expression and epigenetic activity, influencing various cellular processes (such as proliferation and apoptosis) [115] . Moreover, butyrate enhance mitochondrial performance and prevent cell death by blocking a key mediator (mitochondrial complex Ι) [95,116]. Additionally, it is thought that the microbiota may influence the synthesis of molecules such as neurotransmitters (e.g., gamma amino butyric acid) and the products of fermentation (e.g., the short chain fatty acids butyrate, propionate, and acetate) promoting the functioning of the nervous system [6,117]. While contrasting fecal SCFA levels in PD patients and control subjects, decreased concentrations of SCFAs, particularly butyric acid, were found in PD patients, which were linked to microbiota alterations [88]. In fact, several bacterial taxa reportedly reduced in PD are SCFA producers [2,3,27,88]. These molecules are important as an energy source in the body, as well as by stimulating colonic blood flow, capturing fluids and electrolytes, influencing inflammatory responses, and proliferating the mucosa [88,89].
Thus, through the actions of SCFAs on tight junction protein expression, they can modulate the permeability of both intestinal and blood-brain barriers, exhibiting anti-inflammatory and antioxidant properties [6,27].
Age, environment, and genetics are the three major factors modulating the microbiota. In addition, there are other lifestyle factors that appear to play a role in PD pathophysiology. Diet can modulate our microbiome through the availability of macro- and micronutrients. For example, there are certain compounds that may trigger the PD onset/pathology and others that have neuroprotective properties [118,119]. Exercise has been associated with beneficial alterations in microbiome composition, impacting energy homeostasis and regulation [4,120]. In addition to medication, it is clinically recommended to prescribe activation therapies, such as physiotherapy, which is a widely employed therapeutic approach [41]. Caffeine consumption is one of the most studied nutritional habits and is highly correlated with a decreased PD risk, suggesting its potential to modulate the pathophysiology of the disease [91,95,121,122]. As regards nicotine, there are a few studies that have consistently demonstrated an inverse association between smoking and the risk of developing PD [91,95]. Former smokers and smokers had a lower risk of developing PD compared to non-smokers [122]. These factors will cause an alteration in the microbiota, increasing harmful bacteria and decreasing SCFAs, which will lead to a state of dysbiosis, this causes the integrity and function of the intestinal barrier to be affected, increasing metabolites such as LPS and TLRs and decreasing LBP, promoting an increase in various pro-inflammatory cytokines that in turn will cross the BBB reaching the CNS. This state of dysbiosis will also cause an aggregation of αSyn, which via the vagus nerve (Braak's Theory), will reach the CNS. In the brain, this aggregation of αSyn increased pro-inflammatory cytokines, and increased ROS will lead to a state of neuroinflammation and consequently lead to death of dopaminergic neurons. Parkinson Disease (PD); Short-chain fatty acids (SCFAs); bacterial lipopolysaccharide (LPS); lipopolysaccharide-binding protein (LBP); Toll-like receptors (TLRs); Blood-Brain Barrier (BBB); Enteric Nervous System (ENS); Central Nervous System (CNS); alpha-synuclein (α-Syn); Reactive Oxygen Species (ROS). Figure generated using BioRender.com (accessed on March 2024)).
Microbiome: A New Source for PD Biomarkers?
As previously indicated, in addition to the fact that there is no effective treatment to halt the progression of the disease, when patients are diagnosed, they already have a high level of degeneration of the dopaminergic neurons [75,123]. Consequently, there is an urgent need to develop sensitive and practical biomarkers that might allow to detect PD at earlier stages, particularly within the symptomatic prodromic phases of the disease [124,125,126].
Numerous molecules in the CSF, including αSyn, DJ-1, amyloid-beta, tau, and lysosomal enzymes, are currently under investigation as potential biomarkers for PD [7,127,128,129]. Recently, Siderowf et al., show that α-Syn seed amplification assays (SAAs) might be able to distinguish PD patients and healthy subjects profiles, which can be tested, in a near future, as a potential biomarker [130]. Up to date, no definitive and specific biomarkers have been identified. Nevertheless, an in-depth analysis of gut dysbiosis, might be an important step to successfully originate specific PD biomarkers.
Current research indicates that the intestinal microbiota and the GI environment exert influence and may be used as non-motor biomarkers for PD [92]. As previously noted, alongside the characteristic motor symptoms, individuals afflicted with PD commonly experience GI dysfunctions, primarily involving the intestinal tract [131,132]. Constipation, a common GI disorder in PD, may be partly caused by changes in gut microbiota composition and its byproducts [133,134]. Consequently, constipation, dysbiosis, modified intestinal permeability, enteric accumulation of αSyn, and GI inflammation are evident in individuals, indicative of PD at various points in their lives [135,136,137]. It is hypothesized that by profiling the microbiota of PD patients and those at a heightened risk of developing PD, alterations in the GI system could be employed as potential biomarkers for the early detection of PD [138]. However, more studies are needed to fully support this possibility.
The genomic characterization of fecal samples offers a potential avenue for identifying biomarkers associated with PD [139,140]. Qian and colleagues successfully identified twenty-five genetic markers within the intestinal microbiota, which is beneficial for PD diagnosis and to distinguish this pathology from others, such as Alzheimer's disease (AD) and multisystem atrophy (MSA) [140]. Furthermore, investigations into the fecal metabolome of individuals with PD, in comparison to healthy controls, revealed a diminished concentration of SCFAs and noteworthy alterations in the metabolism of amino acids, lipids, and vitamins [141,142]. Therefore, bacterial metabolites and perturbed metabolic pathways may also serve as biomarkers for PD [142]. Despite these promising findings, to date no studies have specifically evaluated the fecal metabolome as a diagnostic biomarker for PD. While the obtained developments are encouraging, replication in larger population samples is imperative to ascertain their reproducibility. Additionally, an emphasis on characterizing the intestinal microbiota, not only through gene sequencing but also at the level of the fecal metabolome, is essential to uncover novel possibilities for biomarkers associated with PD. Additionally, exploring biomarkers, underlying causes of the disease, and symptom management strategies offer potential clinical applications in the future. Pursuing these avenues holds the potential not only to enhance our understanding of PD but also to contribute to the development of effective and early interventions aimed at symptom management, slowing down the disease progression in the future and could make it possible to identify the patients who would most benefit from experimental therapy and to quantify the effectiveness of future drugs [143].
Microbiome: Are We Looking for a New Therapeutic Target?
The existing therapies for PD, including the gold standard, levodopa, predominantly alleviate symptoms with limited efficacy and lack substantial prophylactic effects [135,144,145]. Given the prevalence of GI dysfunction in over 80% of PD individuals, it is plausible to hypothesize that alterations in our microbiota may influence PD. Therefore, this suggests the potential role of a compromised GBrA in the pathogenesis of the disease [146,147].
In contrast to predictive biomarkers, which might indicate or demonstrate a causal relationship, therapeutic interventions need a direct connection between microbes and disease states [148]. If imbalances in the microbiome were considered, as synonymous of disease, restoring a healthy microbiome emerges as a compelling therapeutic alternative [149]. In this context, diverse approaches have been investigated to model the microbiome and reinstate equilibrium. Indeed, microbiome-based therapies encompass various strategies, including live biotherapeutics, fecal microbiota transplant, microbiome mimetics, dietary interventions, prebiotics, probiotics, symbiotics, antibiotics, and phage therapy [148,150,151].
Since the intestinal microbiome is significantly influenced by diet, dietary interventions also emerge as a potential therapeutic target for preventing, modifying, or delaying PD [152,153,154,155]. Notably, nutritional interventions (such as probiotics) can restore the composition of the intestinal microbiota, thereby improving the prognosis of the disease [135,156].
As mentioned above, patients with this disease often exhibit a dysregulated gut microbiome characterized by reduced SCFA bacteria and increased LPS bacteria [152,157,158]. Addressing this imbalance, a treatment method could involve using prebiotics, such as fibers, which can stimulate the growth of SCFA-producing bacteria, increasing SCFAs in the colon [159,160]. In addition, it was noted that administering prebiotics to individuals diagnosed with PD may potentially modify the course of the disease. Therefore, assumptions have been made postulating that an increase in SCFA levels might mitigate issues such as intestinal permeability to endotoxin, colonic inflammation, and neuroinflammation, ultimately contributing to a reduction in the loss of dopaminergic neurons [7,157,161].
Furthermore, Perez-Pardo et al. discovered that a dietary intervention combining two membrane synthesis precursors—uridine and docosahexaenoic acid (DHA)—which enhance phospholipid synthesis, along with prebiotic fibers, exhibited a neuroprotective effect in a PD model-induced by rotenone [162]. Their study demonstrated partial alleviation of both motor and non-motor symptoms induced by rotenone and the restoration of DAT levels in the striatum [162,163,164]. This suggests that the combination of these nutritional elements may benefit the progression and symptomatology of PD. Subsequent investigations have further corroborated this view, demonstrating that diet, probiotics, and prebiotics can potentially normalize dysbiotic microbiota [7,165,166,167]. Therefore, these interventions could hold promise as potentially beneficial strategies in treating PD.
Another potential use of microbiota-targeted intervention is in treating PD patients to optimize the efficacy of current PD drugs since a better understanding of the influence of GM as a predictive biomarker for drug metabolism will give us information on the bioavailability of the medication, its responsiveness, and the direct results of the treatment [4,168,169,170]. Notably, research demonstrated that eradicating Helicobacter pylori (H. pylori) improved levodopa absorption, a key drug used in PD treatment, resulting in reduced motor symptoms [171,172,173]. In PD patients, an H. pylori infection correlates with ongoing motor deterioration [172,173]. Therefore, H. pylori infection interferes with the pharmacokinetics of levodopa, leading to increased delays and periods of ineffectiveness [174]. These findings provide a compelling rationale for considering microbiota-targeted interventions to optimize levodopa therapy, particularly for individuals requiring high and frequent doses of levodopa who may experience significant side effects [175]. Such interventions could potentially enhance the therapeutic outcomes and alleviate the life-altering impact of levodopa-related side effects in PD patients.
While the prospect of interventions targeting the microbiota in PD is promising, it is crucial to acknowledge that additional clinical trials are necessary to ascertain the actual benefits in this context. Presently, there are 39 clinical trials underway (including those exploring the relationship between PD and gut dysbiosis and potential therapeutics), as outlined in Table 2 (Search conducted on the National Library of Medicine's Clinical Trials Registry Platform with the terms "Parkinson's" and "Gut Microbiome"). However, it's essential to note that pre-clinical and clinical studies investigating gut therapeutic strategies for diverse PD symptoms come with certain limitations. Importantly, small sample sizes, insufficient duration for therapeutic intervention, variations in disease severity among patients, inadequacies in outcome measures, and a lack of standardization in key reading variables are some of those limitations [135,176]. Furthermore, the substantial heterogeneity in the microbiome across individuals raises concerns regarding gut microbiota variability in PD patients [177,178]. This emphasizes the significance of individualized approaches during clinical assessment, diagnosis, and treatment selection in pursuing more effective outcomes [179].
Gut Microbiome-Drug Interaction for the Treatment of PD
Notably, the efficacy of pharmaceutical drugs exhibits considerable variability among individuals, owing to the intricate nature of the human body [180,181]. The complexity of this system is further underscored by the pivotal role played by the gut microbiome [182]. Recent research has illuminated the significant influence exerted by gut bacteria on the modulation of drug mechanisms and associated side effects [183,184]. Importantly, these interactions between gut microbes and pharmaceutical agents are bidirectional [185]. In other words, drugs can induce alterations in the composition and functionality of the gut microbiota, subsequently impacting the metabolism and utilization of these drugs within the body [186,187].
The spectrum of pharmaceuticals demonstrating discernible connections to the microbiota is extensive. However, the specific alterations incurred by various drugs on the microbiota exhibit distinctive patterns. Consequences may encompass a diminution in microbial diversity and an augmentation in the proliferation of particular potentially pathogenic bacteria, such as Proteobacteria, Bifidobacteriaceae, Ruminococcaceae, Verrucomicrobiaceae, and Christensenellaceae [188,189,190,191,192]. Nevertheless, the mechanisms through which medications impact the microbiota predominantly operate through indirect mechanisms, wherein alterations are introduced to various facets of the gut environment, including nutrient composition, thereby exerting an influence on bacterial growth [193]. Notably, certain medications undergo direct metabolic transformations by bacteria, thereby dictating their subsequent distribution and functionality within the body [184]. An illustrative instance of this phenomenon is evident in the case of levodopa. In this context, bacteria directly metabolize levodopa, ultimately shaping its trajectory and effects within the organism.
Following oral administration, the absorption of levodopa is imperative through the small intestine for it to traverse the BBB and gain access to the brain [194,195]. Within the brain, the human enzyme aromatic amino acid decarboxylase (AAAD), also called DOPA decarboxylase, catalyzes the conversion of levodopa into therapeutically active dopamine [196,197,198]. The pivotal determinant of the drug's efficacy lies in the bioavailability of levodopa to the brain [199]. Consequently, levodopa is frequently co-administered with catechol metabolism inhibitors, exemplified by carbidopa, to impede its metabolism at sites other than its intended action, thereby optimizing its availability for therapeutic impact [196,200]. This is crucial, as certain gut bacteria and peripheral human enzymes can metabolize the drug before it crosses the BBB [105,201]. This preemptive metabolism could substantially diminish the availability and consequently undermine the efficacy of levodopa [201]. In light of this, the interplay between levodopa and H. pylori, as previously mentioned, underscores the intricate relationship between pharmaceutical agents and gut microbiota [174].
Recent research has brought another dimension of microbial involvement in levodopa metabolism, specifically through microbial decarboxylases in the gut [202]. Notably, a newly identified bacterial metabolism of levodopa, facilitated by tyrosine decarboxylases (tyrDCs), has been determined, with Enterococcus faecalis (E. faecalis) predominantly carrying out this process [169]. Intriguingly, the potential to enhance the drug's efficacy is evident through manipulating this microbial metabolism. Mutation of tyrDCs in E. faecalis has been shown to impede the bacterial conversion of Levodopa into dopamine, presenting a promising avenue for improving the drug's therapeutic effectiveness [169,203].
Nevertheless, considering the role of the intestinal microbiome in drug metabolism, coupled with the substantial impact of medications on the microbiome's composition, the possible interaction between PD medication and the microbiome's composition is not surprising. Actually, insights derived from these studies support the plausible existence of a clinically significant relationship between the microbiota and drug metabolism in individuals with PD. Consequently, mapping the human microbiome becomes imperative to unravel the underlying mechanisms governing these intricate microbiota-drug interactions [204]. Such exploration is pivotal not only for elucidating the dynamics of this interaction but also for unveiling how it influences the efficacy of medication in the context of PD.
Future Perspectives
Although high-throughput sequencing technology has allowed outstanding advances in gut microbiota research, much remains to be unraveled. There is plenty of indirect evidence to support the active role of the microbiome in PD, but there is limited conclusive support. And the real question remains unanswered: Is GM dysregulation a cause or a consequence of PD? It is tough to demonstrate the exact molecular and cellular pathways through which the microbiome may promote the pathogenesis of PD. This is because the microbiome is highly heterogeneous between individuals. Most of the existing studies on gut microbes in PD are cross-sectional studies, which cannot sufficiently indicate the causal relationships between gut microbes and PD pathogenesis, resulting in a significant gap in the literature and much divergence in the results. Therefore, considering the high heterogeneity of gut microbes between different individuals, it is necessary to emphasize more longitudinal research to advance our knowledge about the mechanisms underlying the correlation between gut microbes and PD so that in the future, we can use GM as a biomarker and so that more targeted treatment strategies based on the gut microbiome can be developed (either alone or as an adjuvant for existing therapies) (Figure 3).
Conclusions
PD is a highly heterogeneous disease with an unclear etiology. Whether or not the microbiome is the initial cause of PD remains uncertain, but it has been hypothesized to play a key role in the distinct stages of the disease. This is supported by various pro-inflammatory mediators associated with bacteria, which might contribute to or facilitate a neuroinflammatory state in PD. The GBrA assumes particular importance in PD, namely in the formation of αSyn aggregates and in the bidirectional communication and transport of αSyn via the vagus nerve. Therefore, even though it is an exciting perspective, there are still questions that should be addressed: is microbiome a cause or a consequence of PD? Can microbiome be a potential source of biomarkers or a therapeutic target for PD? Can the microbiome be a critical modulator of the current/available treatment pipeline?
Considering this, future randomized controlled trials are needed to evaluate the potential impact of microbiomes in the different stages of PD and distinct treatment regimens. By doing so, new concepts in the pathophysiology and therapeutic setting of the disease might be established.
Author Contributions
Conceptualization, CV and FGT; Methodology, CV; Investigation, CV; Writing – Original Draft Preparation, CV; Writing – Review & Editing, BA, CSG, RCS, JMM, CT, EG, CP, MAV, FGT; Visualization, MAV and FGT; Supervision, MAV and FGT.; Project Administration, FGT; Funding Acquisition, FGT. All authors have read and agreed to the published version of the manuscript.
Funding
The present work was supported by Prémios Santa Casa Neurociências—Prize Mantero Belard for Neurodegenerative Diseases Research (MB-28-2019), the Portuguese Foundation for Science and Technology (FCT) to F.G.T. (2021.00643.CEECIND), B.A. (2022.13292.BD), CSG (2023.03466.BD), R.C.S. (2022.13323.BD), JMM (2023.00893.BD), and CT (2022.12641.BD).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors acknowledge BioRender for providing the available illustrations that have been used to create figures of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Balestrino, R.; Schapira, A. H. V. Parkinson Disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Parnetti, L.; Gaetani, L.; Eusebi, P.; Paciotti, S.; Hansson, O.; El-Agnaf, O.; Mollenhauer, B.; Blennow, K.; Calabresi, P. CSF and Blood Biomarkers for Parkinson’s Disease. Lancet. Neurol. 2019, 18, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, B.; Lou, P.; Dai, T.; Chen, Y.; Zhuge, A.; Yuan, Y.; Li, L. The Relationship Between the Gut Microbiome and Neurodegenerative Diseases. Neurosci. Bull. 2021, 37, 1510–1522. [Google Scholar] [CrossRef] [PubMed]
- Lubomski, M.; Tan, A. H.; Lim, S.-Y.; Holmes, A. J.; Davis, R. L.; Sue, C. M. Parkinson’s Disease and the Gastrointestinal Microbiome. J. Neurol. 2020, 267, 2507–2523. [Google Scholar] [CrossRef] [PubMed]
- 5. World Health Organization. Parkinson disease, 20 April.
- Pavan, S.; Prabhu, A. N.; Prasad Gorthi, S.; Das, B.; Mutreja, A.; Shetty, V.; Ramamurthy, T.; Ballal, M. Exploring the Multifactorial Aspects of Gut Microbiome in Parkinson’s Disease. Folia Microbiol. (Praha). 0123. [Google Scholar] [CrossRef]
- Keshavarzian, A.; Engen, P.; Bonvegna, S.; Cilia, R. The Gut Microbiome in Parkinson’s Disease: A Culprit or a Bystander? Prog. Brain Res. 2020, 252, 357–450. [Google Scholar] [CrossRef] [PubMed]
- Holmqvist, S.; Chutna, O.; Bousset, L.; Aldrin-Kirk, P.; Li, W.; Björklund, T.; Wang, Z.-Y.; Roybon, L.; Melki, R.; Li, J.-Y. Direct Evidence of Parkinson Pathology Spread from the Gastrointestinal Tract to the Brain in Rats. Acta Neuropathol. 2014, 128, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Hassanzadeh, K.; Rahimmi, A. Oxidative Stress and Neuroinflammation in the Story of Parkinson’s Disease: Could Targeting These Pathways Write a Good Ending? J. Cell. Physiol. 2018, 234, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Sun, M. F.; Shen, Y. Q. Dysbiosis of Gut Microbiota and Microbial Metabolites in Parkinson’s Disease. Ageing Res. Rev. 2018, 45, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Lim, K. L.; Tan, J. M. M. Role of the Ubiquitin Proteasome System in Parkinson’s Disease. BMC Biochem. 2007, 8 (SUPPL. 1), 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Benito, M.; Granado, N.; García-Sanz, P.; Michel, A.; Dumoulin, M.; Moratalla, R. Modeling Parkinson’s Disease With the Alpha-Synuclein Protein. Front. Pharmacol. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wüllner, U.; Borghammer, P.; Choe, C. un; Csoti, I.; Falkenburger, B.; Gasser, T.; Lingor, P.; Riederer, P. The Heterogeneity of Parkinson’s Disease. J. Neural Transm. 2023, 130, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Greenland, J. C.; Williams-Gray, C. H.; Barker, R. A. The Clinical Heterogeneity of Parkinson’s Disease and Its Therapeutic Implications. Eur. J. Neurosci. 2019, 49, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Mahlknecht, P.; Foltynie, T.; Limousin, P.; Poewe, W. How Does Deep Brain Stimulation Change the Course of Parkinson’s Disease? Mov. Disord. 2022, 37, 1581–1592. [Google Scholar] [CrossRef] [PubMed]
- Hariz, M.; Blomstedt, P. Deep Brain Stimulation for Parkinson’s Disease. J. Intern. Med. 2022, 292, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Virel, A.; Dudka, I.; Laterveer, R.; Af Bjerkén, S. (1)H NMR Profiling of the 6-OHDA Parkinsonian Rat Brain Reveals Metabolic Alterations and Signs of Recovery after N-Acetylcysteine Treatment. Mol. Cell. Neurosci. 2019, 98, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J. C.; Ursell, L. K.; Parfrey, L. W.; Knight, R. The Impact of the Gut Microbiota on Human Health: An Integrative View. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Diaz Heijtz, R.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M. L.; Forssberg, H.; Pettersson, S. Normal Gut Microbiota Modulates Brain Development and Behavior. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kasper, L. H. The Role of Microbiome in Central Nervous System Disorders. Brain. Behav. Immun. 2014, 38, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J. F.; O’Riordan, K. J.; Sandhu, K.; Peterson, V.; Dinan, T. G. The Gut Microbiome in Neurological Disorders. Lancet. Neurol. 2020, 19, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Foster, J. A.; McVey Neufeld, K.-A. Gut-Brain Axis: How the Microbiome Influences Anxiety and Depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Cox, M. J.; Cookson, W. O. C. M.; Moffatt, M. F. Sequencing the Human Microbiome in Health and Disease. Hum. Mol. Genet. 2013, 22, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tan, Y.; Cheng, H.; Zhang, D.; Feng, W.; Peng, C. Functions of Gut Microbiota Metabolites, Current Status and Future Perspectives. Aging Dis. 2022, 13, 1106–1126. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Liu, X.; Ye, Y.; Yan, X.; Cheng, Y.; Zhao, L.; Chen, F.; Ling, Z. Gut Microbiota: A Novel Therapeutic Target for Parkinson’s Disease. Front. Immunol. 2022, 13, 937555. [Google Scholar] [CrossRef] [PubMed]
- Bull, M. J.; Plummer, N. T. Part 1: The Human Gut Microbiome in Health and Disease. Integr. Med. (Encinitas). 2014, 13, 17–22. [Google Scholar] [PubMed]
- Shannon, K. M. Gut-Derived Sterile Inflammation and Parkinson’s Disease. Front. Neurol. 2022, 13, 831090. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Hayes, R. B. Environmental Influences on the Human Microbiome and Implications for Noncommunicable Disease. Annu. Rev. Public Health 2021, 42, 277–292. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; Lugli, G. A.; Rodriguez, J. M.; Bode, L.; de Vos, W.; Gueimonde, M.; Margolles, A.; van Sinderen, D.; Ventura, M. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81. [Google Scholar] [CrossRef] [PubMed]
- Reeve, A.; Simcox, E.; Turnbull, D. Ageing and Parkinson ’ s Disease : Why Is Advancing Age the Biggest Risk Factor ? Ageing Res. Rev. 2014, 14, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Wang, P.; Jankovic, J. The Genetics of Parkinson Disease. Ageing Res. Rev. 2018, 42, 72–85. [Google Scholar] [CrossRef]
- Biswas, S.; Bagchi, A. Study of the Effects of Nicotine and Caffeine for the Treatment of Parkinson’s Disease. Appl. Biochem. Biotechnol. 2022, 639–654. [Google Scholar] [CrossRef]
- Nuytemans, K.; Theuns, J.; Cruts, M.; Van Broeckhoven, C. Genetic Etiology of Parkinson Disease Associated with Mutations in the SNCA, PARK2, PINK1, PARK7, and LRRK2 Genes: A Mutation Update. Hum. Mutat. 2010, 31, 763–780. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.-X.; Gulam, M. Y.; Chia, N. S. J.; Feng, L.; Rotzschke, O.; Tan, E.-K. Gut-Brain Axis: Potential Factors Involved in the Pathogenesis of Parkinson’s Disease. Front. Neurol. 2020, 11, 849. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Lenardo, M. J. The Role of LRRK2 in Inflammatory Bowel Disease. Cell Res. 2012, 22, 1092–1094. [Google Scholar] [CrossRef] [PubMed]
- Vizziello, M.; Borellini, L.; Franco, G.; Ardolino, G. Disruption of Mitochondrial Homeostasis: The Role of Pink1 in Parkinson’s Disease. Cells 2021, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hernán, M. A.; Takkouche, B.; Caamaño-Isorna, F.; Gestal-Otero, J. J. A Meta-Analysis of Coffee Drinking, Cigarette Smoking, and the Risk of Parkinson’s Disease. Ann. Neurol. 2002, 52, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Hill-Burns, E. M.; Debelius, J. W.; Morton, J. T.; Wissemann, W. T.; Lewis, M. R.; Wallen, Z. D.; Peddada, S. D.; Factor, S. A.; Molho, E.; Zabetian, C. P.; Knight, R.; Payami, H. Parkinson’s Disease and Parkinson’s Disease Medications Have Distinct Signatures of the Gut Microbiome. Mov. Disord. 2017, 32, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Pavan, S.; Prabhu, A. N.; Prasad Gorthi, S.; Das, B.; Mutreja, A.; Shetty, V.; Ramamurthy, T.; Ballal, M. Exploring the Multifactorial Aspects of Gut Microbiome in Parkinson’s Disease. Folia Microbiol. (Praha). 2022, 67, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Blesa, J.; Phani, S.; Jackson-Lewis, V.; Przedborski, S. Classic and New Animal Models of Parkinson’s Disease. J. Biomed. Biotechnol. 2012, 2012. [Google Scholar] [CrossRef]
- Reichmann, H.; Csoti, I.; Koschel, J.; Lorenzl, S.; Schrader, C.; Winkler, J.; Wüllner, U. Life Style and Parkinson’s Disease. J. Neural Transm. 2022, 129, 1235–1245. [Google Scholar] [CrossRef]
- Leevven-, A.; Obfervatioiis, M. An Ab/Lratt of A.
- Power, A.; Dobell, C. $ection of Tbe Lbtitor. of !Hebicine. The Discovery of the Intestinal Protozoa of Man.’. Proc R Soc Med.
- Ursell, L. K.; Metcalf, J. L.; Parfrey, L. W.; Knight, R. Defining the Human Microbiome. Nutr. Rev. 2012, 70 (SUPPL. 1). [Google Scholar] [CrossRef]
- Grice, E. A.; Segre, J. A. Published in Final Edited Form as: The Human Microbiome: Our Second Genome. Annu. Rev. Genomics Hum. Genet. 2012, 13, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Hollister, E. B.; Gao, C.; Versalovic, J. Compositional and Functional Features of the Gastrointestinal Microbiome and Their Effects on Human Health. Gastroenterology 2014, 146, 1449–1458. [Google Scholar] [CrossRef]
- Blum, H. E. The Microbiome: A Key Player in Human Health and Disease. J. Healthc. Commun. 2017, 02, 1–5. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z. X.; Chen, X. Y.; Wang, J. Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J. B.; Wei, L.; Li, J.; Chen, Z. S. Microbiota in Health and Diseases. Signal Transduct. Target. Ther. 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- Afzaal, M.; Saeed, F.; Shah, Y. A.; Hussain, M.; Rabail, R.; Socol, C. T.; Hassoun, A.; Pateiro, M.; Lorenzo, J. M.; Rusu, A. V.; Aadil, R. M. Human Gut Microbiota in Health and Disease: Unveiling the Relationship. Front. Microbiol. 2022, 13, 1–14. [Google Scholar] [CrossRef]
- Song, E. J.; Lee, E. S.; Nam, Y. Do. Progress of Analytical Tools and Techniques for Human Gut Microbiome Research. J. Microbiol. 2018, 56, 693–705. [Google Scholar] [CrossRef]
- Fraher, M. H.; O’Toole, P. W.; Quigley, E. M. M. Techniques Used to Characterize the Gut Microbiota: A Guide for the Clinician. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 312–322. [Google Scholar] [CrossRef]
- Rezasoltani, S.; Ahmadi Bashirzadeh, D.; Nazemalhosseini Mojarad, E.; Asadzadeh Aghdaei, H.; Norouzinia, M.; Shahrokh, S. Signature of Gut Microbiome by Conventional and Advanced Analysis Techniques: Advantages and Disadvantages. Middle East J. Dig. Dis. 2020, 12, 5–11. [Google Scholar] [CrossRef]
- Cani, P. D. Gutmicrobiota and Obesity: Lessons from the Microbiome. Brief. Funct. Genomics 2013, 12, 381–387. [Google Scholar] [CrossRef]
- Scales, B. S.; Huffnagle, G. B. The Microbiome in Wound Repair and Tissue Fibrosis. J. Pathol. 2013, 229, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Suau, A.; Bonnet, R.; Sutren, M.; Godon, J. J.; Gibson, G. R.; Collins, M. D.; Doré, J. Direct Analysis of Genes Encoding 16S RRNA from Complex Communities Reveals Many Novel Molecular Species within the Human Gut. Appl. Environ. Microbiol. 1999, 65, 4799–4807. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Sakamoto, M.; Benno, Y. Phylogenetic Analysis of the Human Gut Microbiota Using 16S RDNA Clone Libraries and Strictly Anaerobic Culture-Based Methods. Microbiol. Immunol. 2002, 46, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Komiya, S.; Yasumizu, Y.; Yasuoka, Y.; Mizushima, K.; Takagi, T.; Kryukov, K.; Fukuda, A.; Morimoto, Y.; Naito, Y.; Okada, H.; Bono, H.; Nakagawa, S.; Hirota, K. Full-Length 16S RRNA Gene Amplicon Analysis of Human Gut Microbiota Using MinIONTM Nanopore Sequencing Confers Species-Level Resolution. BMC Microbiol. 2021, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Ghoshal, U.; Dhole, T. N.; Ghoshal, U. C. Fecal Microbiota in Patients with Irritable Bowel Syndrome Compared with Healthy Controls Using Real-Time Polymerase Chain Reaction: An Evidence of Dysbiosis. Dig. Dis. Sci. 2015, 60, 2953–2962. [Google Scholar] [CrossRef]
- Furet, J. P.; Firmesse, O.; Gourmelon, M.; Bridonneau, C.; Tap, J.; Mondot, S.; Doré, J.; Corthier, G. Comparative Assessment of Human and Farm Animal Faecal Microbiota Using Real-Time Quantitative PCR. FEMS Microbiol. Ecol. 2009, 68, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, J. F.; Rôças, I. N.; Rosado, A. S. Application of Denaturing Gradient Gel Electrophoresis (DGGE) to the Analysis of Endodontic Infections. J. Endod. 2005, 31, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Imaeda, H.; Fujimoto, T.; Takahashi, K.; Kasumi, E.; Fujiyama, Y.; Andoh, A. Terminal-Restriction Fragment Length Polymorphism (T-RFLP) Analysis for Changes in the Gut Microbiota Profiles of Indomethacin-and Rebamipide-Treated Mice. Digestion 2012, 86, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Namsolleck, P.; Thiel, R.; Lawson, P.; Holmstrøm, K.; Rajilic, M.; Vaughan, E. E.; Rigottier-Gois, L.; Collins, M. D.; De Vos, W. M.; Blaut, M. Molecular Methods for the Analysis of Gut Microbiota. Microb. Ecol. Health Dis. 2004, 16, (2–3). [Google Scholar] [CrossRef]
- Wang, W. L.; Xu, S. Y.; Ren, Z. G.; Tao, L.; Jiang, J. W.; Zheng, S. Sen. Application of Metagenomics in the Human Gut Microbiome. World J. Gastroenterol. 2015, 21, 803–814. [Google Scholar] [CrossRef]
- Bashiardes, S.; Zilberman-Schapira, G.; Elinav, E. Use of Metatranscriptomics in Microbiome Research. Bioinform. Biol. Insights 2016, 10, 19–25. [Google Scholar] [CrossRef]
- Ojala, T.; Kankuri, E.; Kankainen, M. Understanding Human Health through Metatranscriptomics. Trends Mol. Med. 2023, 29, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Issa Isaac, N.; Philippe, D.; Nicholas, A.; Raoult, D.; Eric, C. Metaproteomics of the Human Gut Microbiota: Challenges and Contributions to Other OMICS. Clin. Mass Spectrom. 2019, 14, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Peters, D. L.; Wang, W.; Zhang, X.; Ning, Z.; Mayne, J.; Figeys, D. Metaproteomic and Metabolomic Approaches for Characterizing the Gut Microbiome. Proteomics 2019, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, Y.; Xiao, X.; Zheng, J.; Zhou, H. Metaproteomics: A Strategy to Study the Taxonomy and Functionality of the Gut Microbiota. J. Proteomics 2020, 219. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, K. S.; Maier, T. V.; Walker, A.; Heinzmann, S. S.; Forcisi, S.; Martinez, I.; Walter, J.; Schmitt-Kopplin, P. Challenges of Metabolomics in Human Gut Microbiota Research. Int. J. Med. Microbiol. 2016, 306, 266–279. [Google Scholar] [CrossRef] [PubMed]
- Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut Microbiota Profiling: Metabolomics Based Approach to Unravel Compounds Affecting Human Health. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. F.; Beggs, M. L.; Erickson, B. D.; Cerniglia, C. E. DNA Microarray Analysis of Predominant Human Intestinal Bacteria in Fecal Samples. Mol. Cell. Probes 2004, 18, 223–234. [Google Scholar] [CrossRef]
- Bikel, S.; Valdez-Lara, A.; Cornejo-Granados, F.; Rico, K.; Canizales-Quinteros, S.; Soberón, X.; Del Pozo-Yauner, L.; Ochoa-Leyva, A. Combining Metagenomics, Metatranscriptomics and Viromics to Explore Novel Microbial Interactions: Towards a Systems-Level Understanding of Human Microbiome. Comput. Struct. Biotechnol. J. 2015, 13, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Nearing, J. T.; Comeau, A. M.; Langille, M. G. I. Identifying Biases and Their Potential Solutions in Human Microbiome Studies. Microbiome 2021, 9, 1–22. [Google Scholar] [CrossRef]
- Madhogaria, B.; Bhowmik, P.; Kundu, A. Correlation between Human Gut Microbiome and Diseases. Infect. Med. 2022, 1, 180–191. [Google Scholar] [CrossRef]
- Vijay, A.; Valdes, A. M. Role of the Gut Microbiome in Chronic Diseases: A Narrative Review. Eur. J. Clin. Nutr. 2022, 76, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J. G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F. D.; Costello, E. K.; Fierer, N.; Peña, A. G.; Goodrich, J. K.; Gordon, J. I.; Huttley, G. A.; Kelley, S. T.; Knights, D.; Koenig, J. E.; Ley, R. E.; Lozupone, C. A.; McDonald, D.; Muegge, B. D.; Pirrung, M.; Reeder, J.; Sevinsky, J. R.; Turnbaugh, P. J.; Walters, W. A.; Widmann, J.; Yatsunenko, T.; Zaneveld, J.; Knight, R. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nature methods, 20 May; 10. [CrossRef]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E. M.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; Wilkening, J.; Edwards, R. A. The Metagenomics RAST Server - a Public Resource for the Automatic Phylogenetic and Functional Analysis of Metagenomes. BMC Bioinformatics 2008, 9, 386. [Google Scholar] [CrossRef]
- Morgan, X. C.; Huttenhower, C. Meta’omic Analytic Techniques for Studying the Intestinal Microbiome. Gastroenterology 2014, 146, 1437–1448. [Google Scholar] [CrossRef] [PubMed]
- Lyu, R.; Qu, Y.; Divaris, K.; Wu, D. Methodological Considerations in Longitudinal Analyses of Microbiome Data: A Comprehensive Review. Genes (Basel). 2023, 15, 51. [Google Scholar] [CrossRef]
- Custer, G. F.; Gans, M.; van Diepen, L. T. A.; Dini-Andreote, F.; Buerkle, C. A. Comparative Analysis of Core Microbiome Assignments: Implications for Ecological Synthesis. mSystems 2023, 8. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Joardar, N.; Sengupta, S.; Sinha Babu, S. P. Gut Microbes as Future Therapeutics in Treating Inflammatory and Infectious Diseases: Lessons from Recent Findings. J. Nutr. Biochem. 2018, 61, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Behrouzi, A.; Nafari, A. H.; Siadat, S. D. The Significance of Microbiome in Personalized Medicine. Clin. Transl. Med. 2019, 8. [Google Scholar] [CrossRef]
- Kashyap, P. C.; Chia, N.; Nelson, H.; Segal, E.; Elinav, E. Microbiome at the Frontier of Personalized Medicine. Mayo Clin. Proc. 2017, 92, 1855–1864. [Google Scholar] [CrossRef]
- Ji, B.; Nielsen, J. From Next-Generation Sequencing to Systematic Modeling of the Gut Microbiome. Front. Genet. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Benameur, T.; Porro, C.; Twfieg, M. E.; Benameur, N.; Panaro, M. A.; Filannino, F. M.; Hasan, A. Emerging Paradigms in Inflammatory Disease Management: Exploring Bioactive Compounds and the Gut Microbiota. Brain Sci. 2023, 13. [Google Scholar] [CrossRef]
- Krishnan, S.; Alden, N.; Lee, K. Pathways and Functions of Gut Microbiota Metabolism Impacting Host Physiology. Curr. Opin. Biotechnol. 2015, 36, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Chen, Q.; Zhang, Y. Multidirectional Associations between the Gut Microbiota and Parkinson ’ s Disease, Updated Information from the Perspectives of Humoral Pathway, Cellular Immune Pathway and Neuronal Pathway. 2023, No. December, 1–20. [CrossRef]
- Aho, V. T. E.; Houser, M. C.; Pereira, P. A. B.; Chang, J.; Rudi, K.; Paulin, L.; Hertzberg, V.; Auvinen, P.; Tansey, M. G.; Scheperjans, F. Relationships of Gut Microbiota, Short-Chain Fatty Acids, Inflammation, and the Gut Barrier in Parkinson’s Disease. Mol. Neurodegener. 2021, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Martin, C. R.; Osadchiy, V.; Kalani, A.; Mayer, E. A. The Brain-Gut-Microbiome Axis. Cell. Mol. Gastroenterol. Hepatol. 2018, 6, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Kubista, M.; Andrade, J. M.; Bengtsson, M.; Forootan, A.; Jonák, J.; Lind, K.; Sindelka, R.; Sjöback, R.; Sjögreen, B.; Strömbom, L.; Ståhlberg, A.; Zoric, N. The Real-Time Polymerase Chain Reaction. Mol. Aspects Med. 2006, 27, (2–3). [Google Scholar] [CrossRef] [PubMed]
- Considine, E.; Yin, L.; Hartmann, M. A Review of the Primary Nutritional and Environmental Factors Associated with Parkinson’s Disease. J. Student Res. 2021, 10, 1–14. [Google Scholar] [CrossRef]
- Sampson, T. R.; Debelius, J. W.; Thron, T.; Janssen, S.; Shastri, G. G.; Ilhan, Z. E.; Challis, C.; Schretter, C. E.; Rocha, S.; Gradinaru, V.; Chesselet, M. F.; Keshavarzian, A.; Shannon, K. M.; Krajmalnik-Brown, R.; Wittung-Stafshede, P.; Knight, R.; Mazmanian, S. K. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Stolzenberg, E.; Berry, D.; Yang, D.; Lee, E. Y.; Kroemer, A.; Kaufman, S.; Wong, G. C. L.; Oppenheim, J. J.; Sen, S.; Fishbein, T.; Bax, A.; Harris, B.; Barbut, D.; Zasloff, M. A. A Role for Neuronal Alpha-Synuclein in Gastrointestinal Immunity. J. Innate Immun. 2017, 9, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q. Q.; Haikal, C.; Li, W.; Li, J. Y. Gut Inflammation in Association With Pathogenesis of Parkinson’s Disease. Front. Mol. Neurosci. 2019, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Dadar, M.; Anderson, G.; Chirumbolo, S.; Maes, M. Preventive Treatments to Slow Substantia Nigra Damage and Parkinson’s Disease Progression: A Critical Perspective Review. Pharmacol. Res. 2020, 161, 105065. [Google Scholar] [CrossRef]
- Visanji, N. P.; Brooks, P. L.; Hazrati, L. N.; Lang, A. E. The Prion Hypothesis in Parkinson’s Disease: Braak to the Future. Acta Neuropathol. Commun. 2014, 2, 1–12. [Google Scholar] [CrossRef]
- Rietdijk, C. D.; Perez-Pardo, P.; Garssen, J.; van Wezel, R. J. A.; Kraneveld, A. D. Exploring Braak’s Hypothesis of Parkinson’s Disease. Front. Neurol. 2017, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C. H.; Del Tredici, K.; Braak, H. Parkinson’s Disease: A Dual-Hit Hypothesis. Neuropathol. Appl. Neurobiol. 2007, 33, 599–614. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kwon, S. H.; Kam, T. I.; Panicker, N.; Karuppagounder, S. S.; Lee, S.; Lee, J. H.; Kim, W. R.; Kook, M.; Foss, C. A.; Shen, C.; Lee, H.; Kulkarni, S.; Pasricha, P. J.; Lee, G.; Pomper, M. G.; Dawson, V. L.; Dawson, T. M.; Ko, H. S. Transneuronal Propagation of Pathologic α-Synuclein from the Gut to the Brain Models Parkinson’s Disease. Neuron 2019, 103, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Borghammer, P.; Van Den Berge, N. Brain-First versus Gut-First Parkinson’s Disease: A Hypothesis. J. Parkinsons. Dis. 2019, 9, S281–S295. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Brockmann, K. Blood and Cerebrospinal Fluid Biomarkers of Inflammation in Parkinson’s Disease. J. Parkinsons. Dis. 2022, 12, S183–S200. [Google Scholar] [CrossRef] [PubMed]
- Di Vincenzo, F.; Del Gaudio, A.; Petito, V.; Lopetuso, L. R.; Scaldaferri, F. Gut Microbiota, Intestinal Permeability, and Systemic Inflammation: A Narrative Review. Intern. Emerg. Med. 2023, 19, 275–293. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Wang, J.; Deng, Y.; Ye, Z.; Ge, Y. Inflammatory Microbes and Genes as Potential Biomarkers of Parkinson’s Disease. npj Biofilms Microbiomes 2022, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, S. Z.; Yu, Y. J.; Adeli, K. Role of Gut Microbiota in Neuroendocrine Regulation of Carbohydrate and Lipid Metabolism via the Microbiota-Gut-Brain-Liver Axis. Microorganisms 2020, 8, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Dicks, L. M. T. Gut Bacteria and Neurotransmitters. Microorganisms 2022, 10, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Heidari, A.; Yazdanpanah, N.; Rezaei, N. The Role of Toll-like Receptors and Neuroinflammation in Parkinson’s Disease. J. Neuroinflammation 2022, 19, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, Z.; Lv, A.; Fan, C. How Toll-like Receptors Influence Parkinson’s Disease in the Microbiome–Gut–Brain Axis. Front. Immunol. 2023, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Isik, S.; Yeman Kiyak, B.; Akbayir, R.; Seyhali, R.; Arpaci, T. Microglia Mediated Neuroinflammation in Parkinson’s Disease. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Tansey, M. G.; Wallings, R. L.; Houser, M. C.; Herrick, M. K.; Keating, C. E.; Joers, V. Inflammation and Immune Dysfunction in Parkinson Disease. Nat. Rev. Immunol. 2022, 22, 657–673. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-F.; Shen, Y.-Q. Dysbiosis of Gut Microbiota and Microbial Metabolites in Parkinson’s Disease. Ageing Res. Rev. 2018, 45, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Alexander, C.; Rietschel, E. T. Bacterial Lipopolysaccharides and Innate Immunity. J. Endotoxin Res. 2001, 7, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Carvey, P. M.; Chang, Q.; Lipton, J. W.; Ling, Z. Prenatal Exposure to the Bacteriotoxin Lipopolysaccharide Leads to Long-Term Losses of Dopamine Neurons in Offspring: A Potential, New Model of Parkinson’s Disease. Front. Biosci. 2003, No. 10, 826–837. [Google Scholar] [CrossRef]
- Jangula, A.; Murphy, E. J. Lipopolysaccharide-Induced Blood Brain Barrier Permeability Is Enhanced by Alpha-Synuclein Expression. Neurosci. Lett. 2013, 551, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D. Q. H.; Sperandio, M.; Di Ciaula, A. Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Ji, X.; Song, Z.; Song, W.; Huang, Q.; Yu, T.; Shi, D.; Wang, F.; Xue, X.; Guo, J. Butyrate Inhibits the Mitochondrial Complex Ι to Mediate Mitochondria-Dependent Apoptosis of Cervical Cancer Cells. BMC Complement. Med. Ther. 2023, 23, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bienenstock, J.; Kunze, W.; Forsythe, P. Microbiota and the Gut-Brain Axis. Nutr. Rev. 2015, 73, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Gubert, C.; Kong, G.; Renoir, T.; Hannan, A. J. Exercise, Diet and Stress as Modulators of Gut Microbiota: Implications for Neurodegenerative Diseases. Neurobiol. Dis. 2020, 134. [Google Scholar] [CrossRef] [PubMed]
- Yemula, N.; Dietrich, C.; Dostal, V.; Hornberger, M. Parkinson’s Disease and the Gut: Symptoms, Nutrition, and Microbiota. J. Parkinsons. Dis. 2021, 11, 1491–1505. [Google Scholar] [CrossRef] [PubMed]
- Gaßner, H.; Steib, S.; Klamroth, S.; Pasluosta, C. F.; Adler, W.; Eskofier, B. M.; Pfeifer, K.; Winkler, J.; Klucken, J. Perturbation Treadmill Training Improves Clinical Characteristics of Gait and Balance in Parkinson’s Disease. J. Parkinsons. Dis. 2019, 9, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Postuma, R. B.; Lang, A. E.; Munhoz, R. P.; Charland, K.; Pelletier, A.; Moscovich, M.; Filla, L.; Zanatta, D.; Romenets, S. R.; Altman, R.; Chuang, R.; Shah, B. Caffeine for Treatment of Parkinson Disease: A Randomized Controlled Trial. Neurology 2012, 79, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Gallo, V.; Vineis, P.; Cancellieri, M.; Chiodini, P.; Barker, R. A.; Brayne, C.; Pearce, N.; Vermeulen, R.; Panico, S.; Bueno-De-Mesquita, B.; Vanacore, N.; Forsgren, L.; Ramat, S.; Ardanaz, E.; Arriola, L.; Peterson, J.; Hansson, O.; Gavrila, D.; Sacerdote, C.; Sieri, S.; Kühn, T.; Katzke, V. A.; Van Der Schouw, Y. T.; Kyrozis, A.; Masala, G.; Mattiello, A.; Perneczky, R.; Middleton, L.; Saracci, R.; Riboli, E. Exploring Causality of the Association between Smoking and Parkinson’s Disease. Int. J. Epidemiol. 2019, 48, 912–925. [Google Scholar] [CrossRef] [PubMed]
- Lotankar, S.; Prabhavalkar, K. S.; Bhatt, L. K. Biomarkers for Parkinson’s Disease: Recent Advancement. Neurosci. Bull. 2017, 33, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Le, W.; Dong, J.; Li, S.; Korczyn, A. D. Can Biomarkers Help the Early Diagnosis of Parkinson’s Disease? Neurosci. Bull. 2017, 33, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, E.; Garrido, A.; Scholz, S. W.; Poewe, W. Challenges in the Diagnosis of Parkinson’s Disease. Lancet Neurol. 2021, 20, 385–397. [Google Scholar] [CrossRef]
- Delenclos, M.; Jones, D. R.; McLean, P. J.; Uitti, R. J. Biomarkers in Parkinson’s Disease: Advances and Strategies. Park. Relat. Disord. 2016, 22, S106–S110. [Google Scholar] [CrossRef]
- Dos Santos, M. C. T.; Scheller, D.; Schulte, C.; Mesa, I. R.; Colman, P.; Bujac, S. R.; Bell, R.; Berteau, C.; Perez, L. T.; Lachmann, I.; Berg, D.; Maetzler, W.; Da Costa, A. N. Evaluation of Cerebrospinal Fluid Proteins as Potential Biomarkers for Early Stage Parkinson’s Disease Diagnosis. PLoS One 2018, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, P.; Lashuel, H. A. Opportunities and Challenges of Alpha-Synuclein as a Potential Biomarker for Parkinson’s Disease and Other Synucleinopathies. npj Park. Dis. 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E. H.; Tennagels, S.; Gold, R.; Gerwert, K.; Beyer, L.; Tönges, L. Update on CSF Biomarkers in Parkinson’s Disease. Biomolecules 2022, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Siderowf, A.; Concha-Marambio, L.; Lafontant, D. E.; Farris, C. M.; Ma, Y.; Urenia, P. A.; Nguyen, H.; Alcalay, R. N.; Chahine, L. M.; Foroud, T.; Galasko, D.; Kieburtz, K.; Merchant, K.; Mollenhauer, B.; Poston, K. L.; Seibyl, J.; Simuni, T.; Tanner, C. M.; Weintraub, D.; Videnovic, A.; Choi, S. H.; Kurth, R.; Caspell-Garcia, C.; Coffey, C. S.; Frasier, M.; Oliveira, L. M. A.; Hutten, S. J.; Sherer, T.; Marek, K.; Soto, C. Assessment of Heterogeneity among Participants in the Parkinson’s Progression Markers Initiative Cohort Using α-Synuclein Seed Amplification: A Cross-Sectional Study. Lancet Neurol. 2023, 22, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Meng, L. B.; Chen, L. J.; Shi, X.; Tu, L.; Zhou, Q.; Yu, J. L.; Liao, X.; Zeng, Y.; Yuan, Q. Y. The Role of the Microbiota-Gut-Brain Axis and Intestinal Microbiome Dysregulation in Parkinson’s Disease. Front. Neurol. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Claudino Dos Santos, J. C.; Oliveira, L. F.; Noleto, F. M.; Gusmão, C. T. P.; Brito, G. A. de C.; Viana, G. S. de B. Gut-Microbiome-Brain Axis: The Crosstalk between the Vagus Nerve, Alpha-Synuclein and the Brain in Parkinson’s Disease. Neural Regen. Res. 2023, 18, 2611–2614. [Google Scholar] [CrossRef] [PubMed]
- Weis, S.; Schwiertz, A.; Unger, M. M.; Becker, A.; Faßbender, K.; Ratering, S.; Kohl, M.; Schnell, S.; Schäfer, K.-H.; Egert, M. Effect of Parkinson’s Disease and Related Medications on the Composition of the Fecal Bacterial Microbiota. NPJ Park. Dis. 2019, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, Y.-B. Intestinal Microbiota and Chronic Constipation. Springerplus 2016, 5, 1130. [Google Scholar] [CrossRef] [PubMed]
- Salim, S.; Ahmad, F.; Banu, A.; Mohammad, F. Gut Microbiome and Parkinson’s Disease: Perspective on Pathogenesis and Treatment. J. Adv. Res. 2023, 50, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Gorecki, A. M.; Preskey, L.; Bakeberg, M. C.; Kenna, J. E.; Gildenhuys, C.; MacDougall, G.; Dunlop, S. A.; Mastaglia, F. L.; Anthony Akkari, P.; Koengten, F.; Anderton, R. S. Altered Gut Microbiome in Parkinson’s Disease and the Influence of Lipopolysaccharide in a Human α-Synuclein over-Expressing Mouse Model. Front. Neurosci. 2019, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, S.; Le, W. Intestinal Permeability, Dysbiosis, Inflammation and Enteric Glia Cells: The Intestinal Etiology of Parkinson’s Disease. Aging Dis. 2022, 13, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Haikal, C.; Chen, Q.-Q.; Li, J.-Y. Microbiome Changes: An Indicator of Parkinson’s Disease? Transl. Neurodegener. 2019, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Feng, X.; Zhou, X.; Zhao, M.; Xiao, H.; Li, R.; Shen, H. Identification of Gut Metabolites Associated with Parkinson’s Disease Using Bioinformatic Analyses. Front. Aging Neurosci. 2022, 14, 1–14. [Google Scholar] [CrossRef]
- Qian, Y.; Yang, X.; Xu, S.; Huang, P.; Li, B.; Du, J.; He, Y.; Su, B.; Xu, L. M.; Wang, L.; Huang, R.; Chen, S.; Xiao, Q. Gut Metagenomics-Derived Genes as Potential Biomarkers of Parkinson’s Disease. Brain 2020, 143, 2474–2489. [Google Scholar] [CrossRef] [PubMed]
- Hashish, S.; Salama, M. The Role of an Altered Gut Microbiome in Parkinson’s Disease: A Narrative Review. Appl. Microbiol. 2023, 3, 429–447. [Google Scholar] [CrossRef]
- Zacharias, H. U.; Kaleta, C.; Cossais, F.; Schaeffer, E.; Berndt, H.; Best, L.; Dost, T.; Glüsing, S.; Groussin, M.; Poyet, M.; Heinzel, S.; Bang, C.; Siebert, L.; Demetrowitsch, T.; Leypoldt, F.; Adelung, R.; Bartsch, T.; Bosy-Westphal, A.; Schwarz, K.; Berg, D. Microbiome and Metabolome Insights into the Role of the Gastrointestinal–Brain Axis in Parkinson’s and Alzheimer’s Disease: Unveiling Potential Therapeutic Targets. Metabolites 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K. Y.; Bhoopatiraju, S.; Silverglate, B. D.; Grossberg, G. T. Biomarkers in Parkinson’s Disease: A State of the Art Review. Biomarkers in Neuropsychiatry 2023, 9. [Google Scholar] [CrossRef]
- Jankovic, J.; Aguilar, L. G. Current Approaches to the Treatment of Parkinson’s Disease. Neuropsychiatr. Dis. Treat. 2008, 4, 743–757. [Google Scholar] [CrossRef] [PubMed]
- Poewe, W.; Antonini, A.; Zijlmans, J. C.; Burkhard, P. R.; Vingerhoets, F. Levodopa in the Treatment of Parkinson’s Disease: An Old Drug Still Going Strong. Clin. Interv. Aging 2010, 5, 229–238. [Google Scholar] [CrossRef]
- Mulak, A.; Bonaz, B. Brain-Gut-Microbiota Axis in Parkinson’s Disease. World J. Gastroenterol. 2015, 21, 10609–10620. [Google Scholar] [CrossRef]
- Toledo, A. R. L.; Monroy, G. R.; Salazar, F. E.; Lee, J. Y.; Jain, S.; Yadav, H.; Borlongan, C. V. Gut–Brain Axis as a Pathological and Therapeutic Target for Neurodegenerative Disorders. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Gulliver, E. L.; Young, R. B.; Chonwerawong, M.; D’Adamo, G. L.; Thomason, T.; Widdop, J. T.; Rutten, E. L.; Rossetto Marcelino, V.; Bryant, R. V.; Costello, S. P.; O’Brien, C. L.; Hold, G. L.; Giles, E. M.; Forster, S. C. Review Article: The Future of Microbiome-Based Therapeutics. Aliment. Pharmacol. Ther. 2022, 56, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, N.; Kitano, S.; Puah, G. R. Y.; Kittelmann, S.; Hwang, I. Y.; Chang, M. W. Microbiome and Human Health: Current Understanding, Engineering, and Enabling Technologies. Chem. Rev. 2023, 123, 31–72. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J. S.; Ng, S. C.; Schnabl, B. Promises of Microbiome-Based Therapies. J. Hepatol. 2022, 76, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Wong, A. C. New Approaches to Microbiome-Based Therapies ABSTRACT.
- Jackson, A.; Forsyth, C. B.; Shaikh, M.; Voigt, R. M.; Engen, P. A.; Ramirez, V.; Keshavarzian, A. Diet in Parkinson’s Disease: Critical Role for the Microbiome. Front. Neurol. 2019, 10, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hegelmaier, T.; Lebbing, M.; Duscha, A.; Tomaske, L.; Tönges, L.; Holm, J. B.; Bjørn Nielsen, H.; Gatermann, S. G.; Przuntek, H.; Haghikia, A. Interventional Influence of the Intestinal Microbiome Through Dietary Intervention and Bowel Cleansing Might Improve Motor Symptoms in Parkinson’s Disease. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Singh, R. K.; Chang, H. W.; Yan, D.; Lee, K. M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T. H.; Bhutani, T.; Liao, W. Influence of Diet on the Gut Microbiome and Implications for Human Health. J. Transl. Med. 2017, 15, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Purdel, C.; Margină, D.; Adam-Dima, I.; Ungurianu, A. The Beneficial Effects of Dietary Interventions on Gut Microbiota—An Up-to-Date Critical Review and Future Perspectives. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Leeming, E. R.; Johnson, A. J.; Spector, T. D.; Roy, C. I. L. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 1–28. [Google Scholar] [CrossRef]
- Duan, W. X.; Wang, F.; Liu, J. Y.; Liu, C. F. Relationship Between Short-Chain Fatty Acids and Parkinson’s Disease: A Review from Pathology to Clinic. Neurosci. Bull. 0123. [Google Scholar] [CrossRef]
- Suganya, K.; Koo, B. S. Gut–Brain Axis: Role of Gut Microbiota on Neurological Disorders and How Probiotics/Prebiotics Beneficially Modulate Microbial and Immune Pathways to Improve Brain Functions. Int. J. Mol. Sci. 2020, 21, 1–29. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Deehan, E. C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Liu, X.; Ye, Y.; Yan, X.; Cheng, Y.; Zhao, L.; Chen, F.; Ling, Z. Gut Microbiota: A Novel Therapeutic Target for Parkinson’s Disease. Front. Immunol. 2022, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Perez-Pardo, P.; de Jong, E. M.; Broersen, L. M.; Wijk, N. van; Attali, A.; Garssen, J.; Kraneveld, A. D. Promising Effects of Neurorestorative Diets on Motor, Cognitive, and Gastrointestinal Dysfunction after Symptom Development in a Mouse Model of Parkinson’s Disease. Front. Aging Neurosci. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Luo, Y.; Ray Chaudhuri, K.; Reynolds, R.; Tan, E. K.; Pettersson, S. The Role of Gut Dysbiosis in Parkinson’s Disease: Mechanistic Insights and Therapeutic Options. Brain 2021, 144, 2571–2593. [Google Scholar] [CrossRef] [PubMed]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaź, P. The Role of Microbiota-Gut-Brain Axis in Neuropsychiatric and Neurological Disorders. Pharmacol. Res. 2021, 172. [Google Scholar] [CrossRef] [PubMed]
- Soundharrajan, I.; Kuppusamy, P.; Srisesharam, S.; Lee, J. C.; Sivanesan, R.; Kim, D.; Choi, K. C. Positive Metabolic Effects of Selected Probiotic Bacteria on Diet-Induced Obesity in Mice Are Associated with Improvement of Dysbiotic Gut Microbiota. FASEB J. 2020, 34, 12289–12307. [Google Scholar] [CrossRef] [PubMed]
- Alard, J.; Lehrter, V.; Rhimi, M.; Mangin, I.; Peucelle, V.; Abraham, A. L.; Mariadassou, M.; Maguin, E.; Waligora-Dupriet, A. J.; Pot, B.; Wolowczuk, I.; Grangette, C. Beneficial Metabolic Effects of Selected Probiotics on Diet-Induced Obesity and Insulin Resistance in Mice Are Associated with Improvement of Dysbiotic Gut Microbiota. Environ. Microbiol. 2016, 18, 1484–1497. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, F.; Fiasca, F.; Minelli, M.; Maio, D.; Mattei, A.; Vergallo, I.; Cifone, M. G.; Cinque, B.; Minelli, M. The Effects of Low-Nickel Diet Combined with Oral Administration of Selected Probiotics on Patients with Systemic Nickel Allergy Syndrome (SNAS) and Gut Dysbiosis. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Lubomski, M.; Davis, R. L.; Sue, C. M. The Gut Microbiota: A Novel Therapeutic Target in Parkinson’s Disease? Park. Relat. Disord. 2019, 66, 265–266. [Google Scholar] [CrossRef] [PubMed]
- Rekdal, V. M.; Bess, E. N.; Bisanz, J. E.; Turnbaugh, P. J.; Balskus, E. P. Discovery and Inhibition of an Interspecies Gut Bacterial Pathway for Levodopa Metabolism. Science (80-. ). 2019, 364. [Google Scholar] [CrossRef] [PubMed]
- van Kessel, S. P.; Frye, A. K.; El-Gendy, A. O.; Castejon, M.; Keshavarzian, A.; van Dijk, G.; El Aidy, S. Gut Bacterial Tyrosine Decarboxylases Restrict Levels of Levodopa in the Treatment of Parkinson’s Disease. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Su, W.; Li, S.; Du, W.; Ma, X.; Jin, Y.; Li, K.; Chen, H. Eradication of Helicobacter Pylori Infection Might Improve Clinical Status of Patients with Parkinson’s Disease, Especially on Bradykinesia. Clin. Neurol. Neurosurg. 2017, 160, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Lolekha, P.; Sriphanom, T.; Vilaichone, R. K. Helicobacter Pylori Eradication Improves Motor Fluctuations in Advanced Parkinson’s Disease Patients: A Prospective Cohort Study (HP-PD Trial). PLoS One 2021, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hashim, H.; Azmin, S.; Razlan, H.; Yahya, N. W.; Tan, H. J.; Manaf, M. R. A.; Ibrahim, N. M. Eradication of Helicobacter Pylori Infection Improves Levodopa Action, Clinical Symptoms and Quality of Life in Patients with Parkinson’s Disease. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Visanji, N. P.; Liu, L. W. C.; Lang, A. E.; Pfeiffer, R. F. Gastrointestinal Dysfunction in Parkinson’s Disease. Lancet Neurol. 2015, 14, 625–639. [Google Scholar] [CrossRef] [PubMed]
- Boelens Keun, J. T.; Arnoldussen, I. A.; Vriend, C.; Van De Rest, O. Dietary Approaches to Improve Efficacy and Control Side Effects of Levodopa Therapy in Parkinson’s Disease: A Systematic Review. Adv. Nutr. 2021, 12, 2265–2287. [Google Scholar] [CrossRef] [PubMed]
- Vaswani, P. A.; Tropea, T. F.; Dahodwala, N. Overcoming Barriers to Parkinson Disease Trial Participation: Increasing Diversity and Novel Designs for Recruitment and Retention. Neurotherapeutics 2020, 17, 1724–1735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yue, L.; Fang, X.; Wang, G.; Li, C.; Sun, X.; Jia, X.; Yang, J.; Song, J.; Zhang, Y.; Guo, C.; Ma, G.; Sang, M.; Chen, F.; Wang, P. Altered Gut Microbiota in Parkinson’s Disease Patients/Healthy Spouses and Its Association with Clinical Features. Park. Relat. Disord. 2020, 81, 84–88. [Google Scholar] [CrossRef]
- Fonseca, D. C.; Marques Gomes da Rocha, I.; Depieri Balmant, B.; Callado, L.; Aguiar Prudêncio, A. P.; Tepedino Martins Alves, J.; Torrinhas, R. S.; da Rocha Fernandes, G.; Linetzky Waitzberg, D. Evaluation of Gut Microbiota Predictive Potential Associated with Phenotypic Characteristics to Identify Multifactorial Diseases. Gut Microbes 2024, 16, 2297815. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Sutton, J. Personalized Medicine Could Transform Healthcare (Review). Biomed. Reports 2017, 7, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Naylor, S.; Chen, J. Y. Unraveling Human Complexity and Disease with Systems Biology and Personalized Medicine. Per. Med. 2010, 7, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Goetz, L. H.; Schork, N. J. Personalized Medicine: Motivation, Challenges, and Progress. Fertil. Steril. 2018, 109, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Chen, Y.; Huang, W.; Zhou, H.; Zhang, W. Drug-Microbiota Interactions: An Emerging Priority for Precision Medicine. Signal Transduct. Target. Ther. 2023, 8. [Google Scholar] [CrossRef] [PubMed]
- Wilson, I. D.; Nicholson, J. K. Gut Microbiome Interactions with Drug Metabolism, Efficacy and Toxicity Europe PMC Funders Author Manuscripts The Gut Microbiota Have the Capability of Preforming a Wide Range of Metabolic Reactions On. Transl. Res. 2017, 179, 204–222. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.; Griffin, B. T.; Clarke, G.; Hyland, N. P. Drug–Gut Microbiota Interactions: Implications for Neuropharmacology. Br. J. Pharmacol. 2018, 175, 4415–4429. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Zuo, T. Interplays between Drugs and the Gut Microbiome. Gastroenterol. Rep. 2022, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Lim, S. H. Bidirectional Interaction between Intestinal Microbiome and Cancer: Opportunities for Therapeutic Interventions. Biomark. Res. 2020, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Weersma, R. K.; Zhernakova, A.; Fu, J. Interaction between Drugs and the Gut Microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef] [PubMed]
- Misera, A.; Łoniewski, I.; Palma, J.; Kulaszyńska, M.; Czarnecka, W.; Kaczmarczyk, M.; Liśkiewicz, P.; Samochowiec, J.; Skonieczna-Żydecka, K. Clinical Significance of Microbiota Changes under the Influence of Psychotropic Drugs. An Updated Narrative Review. Front. Microbiol. 2023, 14, 1–16. [Google Scholar] [CrossRef]
- Enright, E. F.; Gahan, C. G. M.; Joyce, S. A.; Griffin, B. T. The Impact of the Gut Microbiota on Drug Metabolism and Clinical Outcome. Yale J. Biol. Med. 2016, 89, 375–382. [Google Scholar] [PubMed]
- Fan, H. X.; Sheng, S.; Zhang, F. New Hope for Parkinson’s Disease Treatment: Targeting Gut Microbiota. CNS Neurosci. Ther. 2022, 28, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Yue, Y.; He, T.; Huang, C.; Qu, B.; Lv, W.; Lai, H. Y. The Association Between the Gut Microbiota and Parkinson’s Disease, a Meta-Analysis. Front. Aging Neurosci. 2021, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Keshavarzian, A.; Green, S. J.; Engen, P. A.; Voigt, R. M.; Naqib, A.; Forsyth, C. B.; Mutlu, E.; Shannon, K. M. Colonic Bacterial Composition in Parkinson’s Disease. Mov. Disord. 2015, 30, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Jiang, J.; Wang, X.; Jiang, K.; Cao, H. Exposure to Prescribed Medication in Early Life and Impacts on Gut Microbiota and Disease Development. eClinicalMedicine 2024, 68, 102428. [Google Scholar] [CrossRef] [PubMed]
- WADE, D. N.; MEARRICK, P. T.; MORRIS, J. L. Active Transport of L-Dopa in the Intestine. Nature 1973, 242, 463–465. [Google Scholar] [CrossRef] [PubMed]
- Ngwuluka, N.; Pillay, V.; Du Toit, L. C.; Ndesendo, V.; Choonara, Y.; Modi, G.; Naidoo, D. Levodopa Delivery Systems: Advancements in Delivery of the Gold Standard. Expert Opin. Drug Deliv. 2010, 7, 203–224. [Google Scholar] [CrossRef] [PubMed]
- Smith-Hicks, C.; Raymond, G. V. CHAPTER 11 - Developmental Disorders; Irani, D. N. B. T.-C. F. in C. P., Ed.; W.B. Saunders: Philadelphia, 2009. [Google Scholar] [CrossRef]
- Gonçalves, S.; Nunes-Costa, D.; Cardoso, S. M.; Empadinhas, N.; Marugg, J. D. Enzyme Promiscuity in Serotonin Biosynthesis, From Bacteria to Plants and Humans. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Gershanik, O. S. Improving L-Dopa Therapy: The Development of Enzyme Inhibitors. Mov. Disord. 2015, 30, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Jenner, P. Stalevo(®) : A Pioneering Treatment for OFF Periods in Parkinsons Disease. Eur. J. Neurol. 2023, 30 Suppl 2, 3–8. [Google Scholar] [CrossRef]
- Tambasco, N.; Romoli, M.; Calabresi, P. Levodopa in Parkinson’s Disease: Current Status and Future Developments. Curr. Neuropharmacol. 2017, 16, 1239–1252. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, S. M.; Gonzales, C.; Jarmusch, A. K.; Momper, J. D.; Ma, J. D. Contribution of the Gut Microbiome to Drug Disposition, Pharmacokinetic and Pharmacodynamic Variability. Clin. Pharmacokinet. 2021, 60, 971–984. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Sheng, S.; Zhang, F. Relationship Between Gut Bacteria and Levodopa Metabolism. Curr. Neuropharmacol. 2022, 21, 1536–1547. [Google Scholar] [CrossRef] [PubMed]
- Jameson, K. G.; Hsiao, E. Y. A Novel Pathway for Microbial Metabolism of Levodopa. Nat. Med. 2019, 25, 1195–1197. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A. L. Mapping Human Microbiome Drug Metabolism by Gut Bacteria and Their Genes. Nature 2019, 570, 462–467. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phases of Gut Microbiome Study. The study of microbiome comprises four essential phases, namely (1) fecal sample collection, (2) DNA extraction, (3) DNA analysis, utilizing the most suitable technique for the intended purpose (techniques outlined in Table 1), and (4) bioinformatic analysis of the obtained results. These analyses can be done to obtain a taxonomy profiling or else analysis of gene functions of the gut microbiome [50,73].
Figure 1.
Phases of Gut Microbiome Study. The study of microbiome comprises four essential phases, namely (1) fecal sample collection, (2) DNA extraction, (3) DNA analysis, utilizing the most suitable technique for the intended purpose (techniques outlined in Table 1), and (4) bioinformatic analysis of the obtained results. These analyses can be done to obtain a taxonomy profiling or else analysis of gene functions of the gut microbiome [50,73].
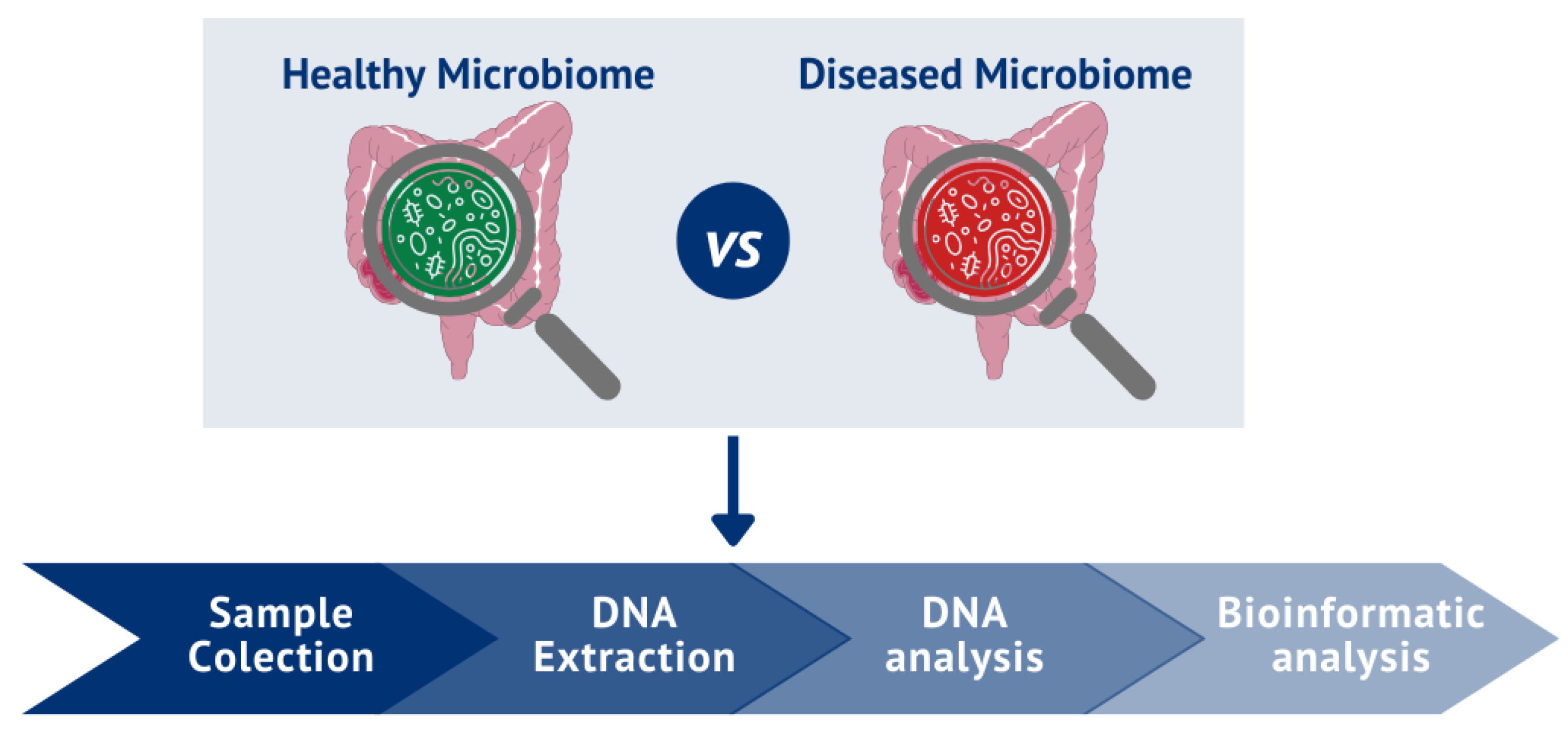
Figure 2.
Gut-Brain Axis Model.

Figure 3.
Gut microbiome and Parkinson's disease. The gut microbiome is an incredibly complex ecosystem, having a denoted and multifaceted impact on people’s health. Indeed, it is nowadays well accepted that it might play a role in the pathogenesis of numerous diseases, including neurodegenerative diseases such as PD. This appreciation of the importance and complexity of the gut microbiome leads to several questions that are currently important to address, with the correct application of technological assays to the same set of samples in hopes of capturing multiple layers of information about the microbiome's involvement in disease. Parkinson’s Disease (PD); Gut Microbiome (GM); Short-chain fatty acids (SCFAs); bacterial lipopolysaccharide (LPS).
Figure 3.
Gut microbiome and Parkinson's disease. The gut microbiome is an incredibly complex ecosystem, having a denoted and multifaceted impact on people’s health. Indeed, it is nowadays well accepted that it might play a role in the pathogenesis of numerous diseases, including neurodegenerative diseases such as PD. This appreciation of the importance and complexity of the gut microbiome leads to several questions that are currently important to address, with the correct application of technological assays to the same set of samples in hopes of capturing multiple layers of information about the microbiome's involvement in disease. Parkinson’s Disease (PD); Gut Microbiome (GM); Short-chain fatty acids (SCFAs); bacterial lipopolysaccharide (LPS).
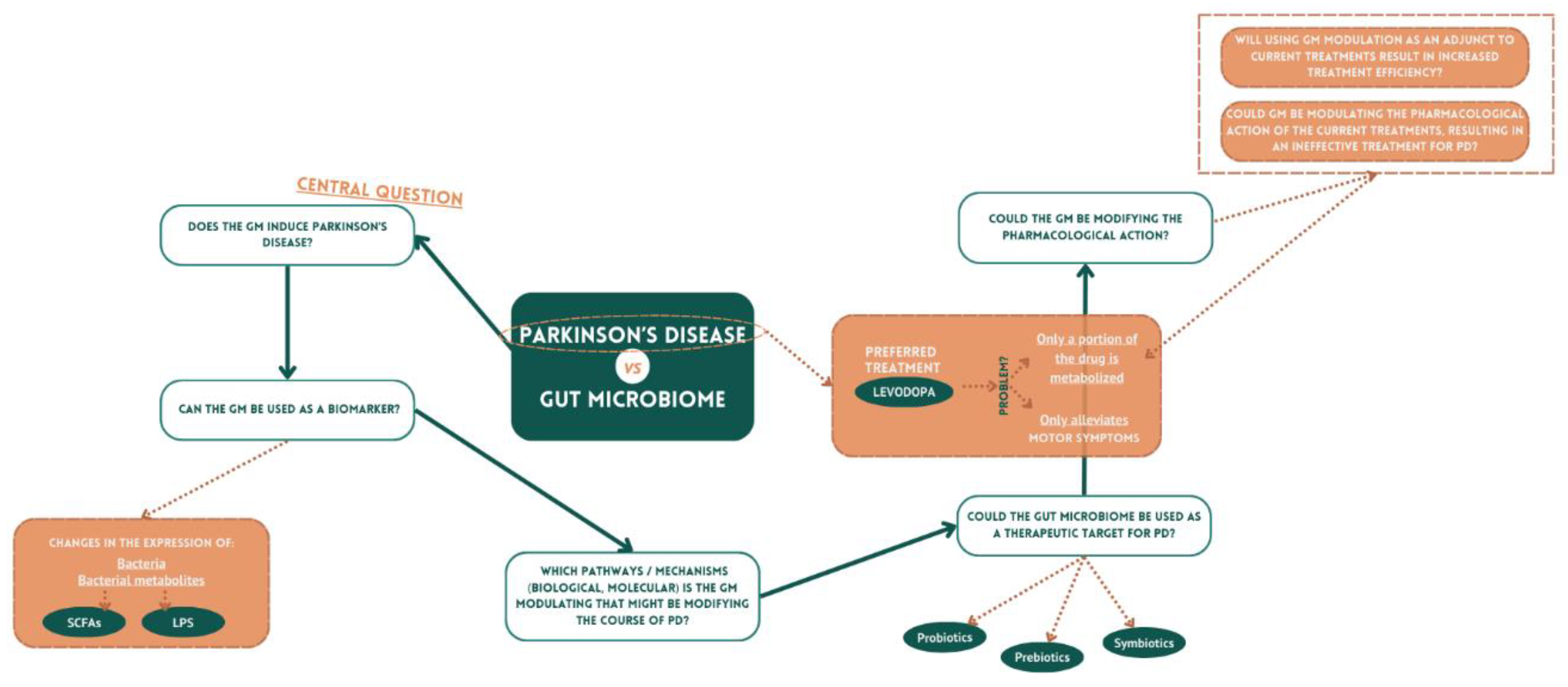

Table 1.
Techniques used to characterize the gut microbiota.
| Technique | Description | References |
|---|---|---|
| Culture-based | Isolation/ Growing of bacteria on selective media | [51,52,53,54] |
| 16S rRNA (16S ribossomal RNA) gene sequencing based on cloning |
The process involves cloning the entire 16S rRNA amplicon, followed by Sanger sequencing and capillary electrophoresis for analysis. | [51,52,55,56] |
| Direct sequencing of 16S rRNA amplicons |
High-throughput parallel sequencing of partial 16S rRNA amplicons. | [51,52,57] |
| Quantitative real-time polymerase chain reaction (qPCR) | 16 S rRNA amplification and quantification. A substance in the reaction mixture exhibits fluorescence when is attached to a double-stranded DNA. | [51,52,58,59] |
| Denaturing gradient gel electrophoresis (DGGE) / Temperature Gradient Gel Electrophoresis (TGGE) | 16S rRNA amplicons separated on a gel using a denaturant/ temperature gradient | [51,52,60] |
| Terminal restriction fragment length polymorphism (T-RFLP) |
After the quantification of fluorescently labeled primers, the 16S rRNA amplicon undergoes digestion utilizing restriction enzymes. Subsequently, the resulting digested fragments are separated through gel electrophoresis. | [51,52,61] |
| Fluorescence in situ hybridization (FISH) |
Oligonucleotide probes, labeled with fluorescent markers, bind to sequences that are complementary to the target 16S rRNA. The fluorescence generated during this hybridization process can be quantified using flow cytometry. | [51,52,62] |
| Microbiome Shotgun sequencing | Extensive parallel sequencing of the entire genome. | [51,52] |
| Metagenomics | Exploring high-resolution profiling of gut microbiota genomes and characterizing gene structures of uncultivated microbiota. | [51,63] |
| Metatranscriptomics | High-resolution gene expression profiling is achieved through sequencing messenger RNA (mRNA) or complementary DNA (cDNA). This approach is used to explore differential microbial gene expression under various physiological or environmental conditions, providing insights into microbial adaptation and responses. | [64,65] |
| Metaproteomics | High-resolution protein monitoring and profiling involve the identification of proteins and peptides, enabling the examination of differential microbial protein production in diverse physiological or environmental conditions. | [66,67,68] |
| Metabolomics | Metabolites undergo analysis to profile the metabolic activity of microbial hosts. | [67,69,70] |
| DNA microarrays | Oligonucleotide probes labeled with fluorescent markers undergo hybridization with complementary nucleotide sequences. The resulting fluorescence is detected using a fluorescence laser detector. | [51,71] |
Table 2.
Summary table of clinical cases registered (Clinical Trials.gov: access on February 04, 2024)
Table 2.
Summary table of clinical cases registered (Clinical Trials.gov: access on February 04, 2024)
| Clinical Trial Title | Study Type | Sample Size | Study Phase | ClinicalTrials.gov Identifier |
|---|---|---|---|---|
| Ketogenic Diet Interventions in Parkinson's Disease: Safeguarding the Gut Microbiome (KIM) | Interventional | 50 | Not yet recruiting | NCT05469997 |
| Characterization of Fecal Microbiome Changes After Administration of PRIM-DJ2727 in Parkinson's Disease Patients | Interventional | 0 | Withdrawn | NCT03026231 |
| The Microbiome in Parkinson's Disease | Observational | 210 | Unknown status | NCT03129451 |
| Microbiome and Diet in Parkinson's Disease - a Randomized, Controlled Phase 2 Trial (PD-Diet) | Interventional | 40 | Not yet recruiting | NCT06207136 |
| Levodopa Response and Gut Microbiome in Patients With Parkinson's Disease | Observational | 38 | Completed | NCT04956939 |
| Prebiotics in the Parkinson's Disease Microbiome | Interventional | 20 | Completed | NCT04512599 |
| Observational Small Intestine and Blood Fingerprint (SmIle) Study in Parkinson's Disease | Observational | 100 | Not yet recruiting | NCT06003608 |
| Deep Brain Stimulation Therapy and Intestinal Microbiota | Observational | 30 | Unknown status | NCT04855344 |
| Single-center Pathophysiological Study of the Role of Inflammation, Changes in the Intestinal Epithelial Barrier and the Intestinal Microbiota in Parkinson's Disease | Interventional | 77 | Terminated | NCT04652843 |
| A Pilot Study to Explore the Role of Gut Flora in Parkinson's Disease | Observational | 100 | Unknown status | NCT04148326 |
| Study of the Genome, Gut Metagenome and Diet of Patients With Incident Parkinson's Disease | Observational | 138 | Terminated | NCT04119596 |
| Resistant Maltodextrin for Gut Microbiome in Parkinson's Disease: Safety and Tolerability Study | Interventional | 30 | Active, not recruiting | NCT03667404 |
| Microbiota Modification for the Treatment of Motor Complication of Parkinson´s Disease | Interventional | 14 | Completed | NCT04730245 |
| Constipation and Changes in the Gut Flora in Parkinson's Disease | Observational | 80 | Recruiting | NCT05787756 |
| AADC/TDC in Advanced Parkinson's Disease | Observational | 50 | Recruiting | NCT05558787 |
| A Trial of Fecal Microbiome Transplantation in Parkinson's Disease Patients | Interventional | 51 | Completed | NCT04854291 |
| Gut Microbiota and Parkinson's Disease | Observational | 50 | Unknown status | NCT03710668 |
| Determining the Microbiota Composition of the Middle Meatus in Parkinson's | Observational | 48 | Completed | NCT03336697 |
| Study of the Fecal Microbiome in Patients With Parkinson's Disease | Interventional | 15 | Completed | NCT03671785 |
| Parkinson's Disease Biomarkers in Nerve Cells in the Gut | Observational | 60 | Recruiting | NCT05347407 |
| Efficacy and Safety of Fecal Microbiota Transplantation in the Treatment of Parkinson's Disease With Constipation | Interventional | 30 | Recruiting | NCT04837313 |
| Increased Gut Permeability to Lipopolysaccharides (LPS) in Parkinson's Disease | Observational | 43 | Completed | NCT01155492 |
| Fecal Microbiota Transplantation for Parkinson's Disease | Interventional | 49 | Completed | NCT03808389 |
| Microbiota Intervention to Change the Response of Parkinson's Disease | Interventional | 86 | Recruiting | NCT03575195 |
| Microbiome Composition and Function Contributes to Cognitive Impairment and Neuroinflammation in Parkinson's Disease | Observational | 100 | Recruiting | NCT05419453 |
| Gut Microbiota in the Progression of Alpha-synucleinopathies | Observational | 490 | Recruiting | NCT05353868 |
| Gut Health and Probiotics in Parkinson's (SymPD) | Interventional | 60 | Active, not recruiting | NCT05146921 |
| Effects of Probiotics on Peripheral Immunity in Parkinson's Disease | Interventional | 88 | Enrolling by invitation | NCT05173701 |
| Gut Microbiota Across Early Stages of Synucleinopathy: From High-risk Relatives, REM Sleep Behavior Disorder to Early Parkinson's Disease | Observational | 441 | Completed | NCT03645226 |
| Meridian Activation Remedy System for Parkinson's Disease (MARS-PD) | Interventional | 88 | Recruiting | NCT05621772 |
| Mediterranean Diet Intervention to Improve Gastrointestinal Function in Parkinson's Disease a Randomized, Controlled, Clinical Trial (MEDI-PD) | Interventional | 46 | Completed | NCT04683900 |
| MOVIN' CARE for PD (Risk Management) (jcpdmcP) | Interventional | 308 | Active, not recruiting | NCT06147284 |
| Fecal Microbiota Transplantation As a Potential Treatment for Parkinson's Disease | Interventional | 10 | Completed | NCT03876327 |
| Efficacy of Probiotics for Parkinson Disease (PD) | Interventional | 300 | Not yet recruiting | NCT06118294 |
| MOVIN' CARE for PD (Mind-body Interventions) (jcpdmcI) | Interventional | 154 | Active, not recruiting | NCT06078046 |
| Establishment of a Human Tissue Bank for Studying the Microbial Etiology of Neurodegenerative Diseases | Observational | 0 | Withdrawn | NCT01954875 |
| The Sunnybrook Dementia Study (SDS) | Observational | 1600 | Recruiting | NCT01800214 |
| Metabolic Cofactor Supplementation in Alzheimer's Disease (AD) and Parkinson's Disease (PD) Patients | Interventional | 120 | Completed | NCT04044131 |
| N-DOSE: A Dose Optimization Trial of Nicotinamide Riboside in Parkinson's Disease | Interventional | 80 | Recruiting | NCT05589766 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



