
Submitted:
19 April 2024
Posted:
22 April 2024
You are already at the latest version
Abstract
Alpinia oxyphylla is a traditional Chinese medicinal plant, with a medicinal history of more than 1,700 years. Ring leaf blight (RLB) disease, caused by pestalotioid species, is an important disease of A. oxyphylla, seriously affecting the yield and quality of fruits. The causal agent of RLB disease has not been systematically identified and characterized so far. In this study, thirty-six pestalotioid strains were isolated from the leaves and stems of A. oxyphylla collected from 6 cities of Hainan province, China. Based on multi-locus phylogeny (ITS, tef-1α and tub2) and morphological characteristics analyses, seventeen species belonging to three genera (Neopestalotiopsis, Pestalotiopsis and Pseudopestalotiopsis) were identified and six new species (N. baotingensis, N. oblatespora, N. olivaceous, N. oxyphylla, N. wuzhishanensis and N. yongxunensis) were described. Pathogenicity tests revealed that strains of Neopestalotiopsis species caused more severe ring leaf blight on A. oxyphylla than strains of Pestalotiopsis and Pseudopestalotiopsis under wounded inoculation conditions.
Keywords:
Pestalotioid species
; New taxa
; Alpinia oxyphylla
; Ring leaf blight
; Pathogenicity
1. Introduction:
Alpinia oxyphylla is an important Chinese herbal plant, with a medicinal history dating back to 1,700 years ago [1]. As an edible herb, the traditional medicinal effects of A. oxyphylla mainly include warming the kidney, solidifying spermatorrhea, arresting polyuria, warming the spleen as well as stopping diarrhea and saliva [2,3]. Moreover, the essential oil of A. oxyphylla has various effects including antibacteria, anticancer, antioxidant, vasodilation and improve immunity [4]. A. oxyphylla likes to grow in warm and humid environmental conditions and is commonly planted under rubber tree, areca tree and other economic forests as a semi-shade plant [5,6,7]. A. oxyphylla is mainly distributed in southern China, such as Hainan, Guangdong and Guangxi province. Among them, Hainan is the most important planting area for A. oxyphylla, accounting for 90% of the total output in China [8,9,10].
The occurrence of diseases causes serious losses to the production and quality of A. oxyphylla. Ring leaf blight (RLB) is an important disease of A. oxyphylla, occurring from seedling to fruiting stage, mainly infecting the old leaves. The disease often extends from the leaf edge or tip, forming irregular, reddish-brown spots with alternating dark and light brown, wavy concentric rings and obvious yellow halos around the periphery of the disease spots, on which scattered numerous small black conidiomata of the pathogen. The high temperature and rainy season contribute to the occurrence of RLB disease, and the high incidence of this disease is from August to September. Under suitable conditions, the proportion of diseased plants can reach more than 50%, and the area of the diseased spots can reach 1/3~1/2 of the leaf surface, even the entire leaf, which has an impressive impact to the growth of A. oxyphylla [11,12].
The pathogen of RLB disease was first reported as Pestalotia palmarum in 1986[11]. Subsequently, the classification status of P. palmarum was adjusted to the genus of Pestalotiopsis, while the genus of Pestalotia and Pestalotiopsis was used confusingly in description of A. oxyphylla diseases [13]. The ring brown spot (RBS) disease of A. oxyphylla was caused by Pestalosphaeria alpinia, a species of sexual morphs of pestalotioid fungi [25]. As asexual fungi, most pestalotioid species lack the sexual morphs, Pestalosphaeria[26]. Most of pestalotioid species are important plant pathogens, and are also found as commonly endophytes or saprophytes, mainly distributed throughout tropical and temperate regions [14,15,16]. Pestalotioid species can infect leaves, shoots, flowers, fruits, or other parts of plants, and cause a variety of diseases of multiple economic crops, including leaf spot, gray blight, shoot dieback, trunk diseases, dry flower and fruit rot [15,17,18,19,20,21,22,23,24]. Hence, pestalotioid species causing the disease of A. oxyphylla need to be reidentified and characterized based on fungi diversity, molecular systematics and pathogenicity.
The development of molecular phylogenetic analysis overcomes the limitation of overlapping conidial measurements in traditional taxonomy of pestalotioid species [14,15,27,28]. In 2014, two novel genera, Neopestalotiopsis and Pseudopestalotiopsis, were segregated from Pestalotiopsis based on conidial characters and multi-locus phylogenetic analyses. The combined sequences of ITS, tub2 and tef-1α genes were used to construct phylogenetic trees, which become an important basis for distinguishing different species within the genus of Pestalotiopsis, Neopestalotiopsis and Pseudopestalotiopsis. Morphologically, Neopestalotiopsis can be easily differentiated from Pestalotiopsis and Pseudopestalotiopsis by their versicolorous median cells of conidia, and Pseudopestalotiopsis is different from Pestalotiopsis with three darker concolorous median cells [15]. Through these methods, many novel pestalotioid species isolated from different plants have been introduced in recent years [17,20,29,30,31,32,33,34].
Therefore, the object of this study is to clarify the types, characteristics and pathogenicity of pestalotioid species related to the disease of A. oxyphylla in Hainan, China.
2. Materials and Methods
2.1. Sample Collection, Fungi Isolation and Morphological Examination
Fresh leaves and stems of A. oxyphylla with typical ring spot were collected from the main planted area at ten towns in six cities of Hainan province, including Baoting, Ledong, Qiongzhong, Sanya, Wanning, and Wuzhishan in 2022. Small pieces (5×5 mm) of leaves or stems were cut from the junction of disease and health areas, disinfected with 3% sodium hypochlorite for 3 min, then 75% ethanol for 30s, subsequently washed with sterilized water for three times. The treated tissue pieces were dried on sterilized blotting paper and then placed on PDA plates (containing 100 μg/mL streptomycin, 50 μg/mL kanamycin and 100 μg/mL ampicillin). The plates were cultured at room temperature and examined daily for 7 days, then the marginal mycelia were transferred to fresh PDA and purified by single-spore culturing.
The pestalotioid strains usually sporulated at room temperature on PDA after 10-20 days. Conidiomata were observed using dissecting microscope (CNOPTEC, SZ680, China) and the characteristics of spores and conidiophores using optical microscope (CNOPTEC, DV320, China). All the morphological characteristics of the spores were photographed and measured at least 30 individuals using OPTPro. The images were progressed by Adobe Photoshop CS6. The pure cultures of isolated fungal strains were stored in the seed health center of China Agricultural University.
2.2. DNA Extraction, Gene Sequencing, and Phylogenetic Analyses
DNA was extracted from fresh fungal mycelia using the Biomed genomic DNA extraction kit (Biomed, Beijing). The partial sequences of three genes (ITS, tef-1α and tub2) were amplified. The PCR was performed according to Table 1 and the PCR products were purified and sequenced at Beijing Tsingke Biotech.
The nucleotide sequences were checked by Chromas2.4.1, then blasted in the NCBI to assess the closest phylogenetic matches. All related sequences by blasted or referenced previous studies were downloaded from GenBank (Table 2). MAFFT v.7(https://mafft.cbrc.jp/alignment/software/) was used to align each locus sequences and MEGA v.11 was used to manually improve the sequences. The three final aligned gene sequences were concatenated by SequenceMatrix [40].
The phylogenetic analyses of the combined sequences were carried out with maximum-likelihood (ML) and Bayesian inference (BI) methods. ML analysis was performed on the CIPRES web portal (https://www.phylo.org) using RAxML-HPC BlackBox 8.2.10 with GTRGAMMA substitution model and 1,000 bootstrap replicates [91]. BI analysis was implemented using MrBayes v.3.2.7 [92], and MrModeltest 2.2 [92] was used to seek the best-fit nucleotide substitution models for each gene. Two Markov chain Monte Carlo (MCMC) were run for 1,000,000 generations, and trees were sampled every 1000th generation. The first 25% of trees, standing for the burn-in phase of the analyses, were discarded, and the remaining trees were estimated the posterior probabilities. ML tree and BI tree were viewed using Figtree v.1.4.4. and modified by WPS Office.
The new species can be further confirmed through PHI (Pairwise Homoplasy Index) analysis, which can also be used to analyze the species boundaries and related taxa [93]. The PHI test was completed in SplitsTree v.4 [94,95],and the value over 0.05 reveals no significant recombination in the dataset. The relationship among closely related species were shown by splits graphs through the LogDet transformation and split decomposition.
2.3. Pathogenicity Test
The pathogenicity of fungi was tested by wound inoculation method. Fresh and healthy leaves of A. oxyphylla with 30-40 cm long were collected from the field. The surface of the leaves was disinfected by spraying of 75% ethanol and then washed three times with sterile water. Each fungal isolate was inoculated on 6 sites of a leaf with 3 leave replicates. A piece of mycelial (6 mm diameter), which was taken from the margin of a fresh colony cultured to 2/3 of the PDA plate’s diameter, was placed on the wound of leaf injured by sterilized needle. A piece of PDA without mycelium was used as control. The inoculated leaves were placed in a box and cultured in the incubator at 26℃, 600 LUX with 16 h/8 h LED light/dark cycle. After 5 days, disease symptoms were recorded and the lesion area was measured using ImageJ and the data were analyzed by SPSS Statistics 24. The re-isolated fungi from disease lesion were identified based on Koch's postulate.
3. Results
3.1. Phylogenetic Analyses
A total of 36 pestalotioid isolates were obtained from the leaves (32 isolates) and stems (4 isolates) samples of A. oxyphylla in six cities of Hainan province. Based on ITS sequence and the color of intermediate cells of conidia, 36 strains were classified into three genera, of which 32 strains belong to Neopestalotiopsis, two strains belong to Pestalotiopsis and two strains belong to Pseudopestalotiopsis.
The phylogenetic tree of Neopestalotiopsis contained 145 taxa, with 2 outgroup taxa (P. colombiensis and P. diversiseta). A total of 1,404 characters including gaps (503 for ITS, 469 for tef-1a, and 432 for tub2) were included in the phylogenetic analysis. For the Bayesian inference, the HKY+G model with gamma-distributed rate was selected for ITS, HKY+G model with gamma-distributed rate was selected for tef1-a and the HKY+I+G model with invgamma-distributed rate was selected for tub2. Similar tree topologies were acquired by ML and BI methods, and the best scoring ML tree is shown in Figure 1. The phylogenetic tree analyzed 32 Neopestalotiopsis taxa isolated from A. oxyphylla, revealed 6 novel species.
The phylogenetic tree of Pestalotiopsis comprised 78 taxa, with the outgroup taxon (N. cubana CBS 600.96). A total of 1,475 characters including gaps (505 for ITS, 495 for tef-1a, and 475 for tub2) were included in the phylogenetic analysis. For the Bayesian inference, the GTR + I + G model with invgamma-distributed rate was selected for ITS, GTR + G model with gamma-distributed rate was selected for tef1-a and the GTR + I + G model with invgamma-distributed rate was selected for tub2. Similar tree topologies were obtained by ML and BI methods, and the best scoring ML tree is shown in Figure 2. The phylogenetic tree analyzed two Pestalotiopsis strains isolated from A. oxyphylla, clustered with the type species of P. hydei.
The alignment of Pseudopestalotiopsis contained 35 taxa, with P. trachicarpicola OP068 as outgroup taxon. A total of 1,392 characters including gaps (521 for ITS, 442 for tef-1a, and 429 for tub2) were included in the phylogenetic analysis. For the Bayesian inference, the HKY+G model with gamma-distributed rate was selected for ITS, HKY + G model with gamma-distributed rate was selected for tef1-a and the HKY + I model with propinv-distributed rate was selected for tub2. Similar tree topologies were obtained by ML and BI methods, and the best scoring ML tree is shown in Figure 3. The phylogenetic tree analyzed two Pseudopestalotiopsis taxa isolated from A. oxyphylla, clustered with the type species of Ps. avicenniae and Ps. myanmarina respectively.
3.2. PHI Analyses
The result of PHI test indicates no obvious recombination (Фw =0.1064) among N. baotingensis SX41-0706, N. oblatespora YJ11-0708 and their closely species N. saprophytica MFLUCC 12-0282, N. paeoniea CBS 318.74, N. hydeana MFLUCC 20-0132, N. egyptiaca CBS 140162, N. guajavicola FMBCC 11.4, N. mesopotamica CBS 299.74 (Figure 4a). And there is no significant recombination (Фw =0.0786) between N. olivaceous LF25-0709 and its closely species N. amomi HKAS 124563, N. zingiber GUCC 21001, N. magna MFLUCC 12-0652 (Figure 4b). N. yongxunensis YX101-0708, N. wuzhishanensis YX116-0708 and their closely taxa have no significant recombination according to the PHI test result (Фw =0.1103) (Figure 4c).
3.3. Taxonomy
Based on multi-locus phylogeny (ITS, tef-1α and tub2) and morphological characteristics analyses, 17 species were identified. Three Neopestalotiopsis strains failed to acquire spores and were not been identified to specific species. Six new species were described as below. The conidial dimension of identified isolations in this study and their closely strains are shown in Table 3.
Neopestalotiopsis baotingensis X.F. Cui and Z.G. Hao, sp. nov. (Figure 5)
Etymology: named referring to the first collection city of Baoting in Hainan Province.
Holotype: SX41-0706.
Description:
Conidiomata on PDA solitary or aggregated, globose, dark. Conidiophores often degenerated to conidiogenous cells. Conidiogenous cells spherical, hyaline. Conidia, fusiform, straight to slightly curved, 18-26×5-7.2 μm (=23.2×6.3 μm), 4 septate; basal cell conical to obtuse, hyaline, thin and smooth-walled, 3.2-6.2 μm long (=4.5 μm); three median cells, 12-17.3 μm (=14.8 μm), verruculose, versicolor, pale brown to dark brown, septa and periclinal walls darker than the rest of the cell, second cell from base pale brown to brown, paler than the two other cells, 3.2-5.5 μm long (=4.4 μm), third cell brown to dark brown, darker than the two other cells, 4-6 μm long (=4.9 μm), fourth cell brown to darker brown, 4-6 μm long (=5 μm); apical cell 2.5-5 μm long (=3.8 μm), cylindric to subcylindric; with 2-4 tubular appendages on the apical cell, often 2-3, arising from the apex of the apical cell, unbranched, 3-30.5 μm long (=19.7 μm); single basal appendage, unbranched, tubular, centric, 2.5-10 μm long (=6.3 μm). Sexual morph not observed.
Culture characteristics: The colony reached 70 mm diameter on PDA after 4 days growth at room temperature. The colony was off white, dense aerial hyphae on the surface with crenate edge, and its reverse was lemon-yellow.
Material examined: China, Hainan Province, Baoting city, Shiling Town, Shuixian village, from leaf spots of A. oxyphylla, 6 July 2022, X.F. Cui and Z.G. Hao (SX41-0706, holotype); ex-type, Hainan Province, Wuzhishan city, Shuiman Town, Yongxun village, from spots on base stem of A. oxyphylla, 8 July 2022, X.F. Cui and Z.G. Hao (YJ34-0708);
Notes: Two strains of Neopestalotiopsis baotingensis were isolated from two cities of Hainan, SX41-0706 and YJ34-0708, clustered with well-supported (ML=81%, BI=1). N. baotingensis is closed related to N. saprophytica (MFLUCC 12-0282) in the phylogenetic analysis. The conidiophores of N. baotingensis often degenerated to conidiogenous cells, while that of N. saprophytica unbranched or irregularly branched; N. baotingensis is shorter than N. saprophytica (N. baotingensis 18-26 μm, =23.2 μm vs. N. saprophytica 22–30 μm, =24.9 μm); and N. baotingensis has shorter apical appendages (N. baotingensis 3-30.5 μm, =19.7 μm vs. N. saprophytica 23-35 μm, =27.3 μm). Additionally, there are 18 bp difference of ITS~tef-1α~tub2 between N. baotingensis and N. saprophytica (4/452 in ITS; 16/784 in tef-1α; and 1/448 in tub2). The PHI test about N. baotingensis reveals that there is no obvious recombination between N. baotingensis and its closely taxa. Therefore, N. baotingensis is classified as a new species in this study.
Neopestalotiopsis oblatespora X.F. Cui and Z.G. Hao, sp. nov. (Figure 6)
Etymology: named referring to spore morphology.
Holotype: YJ11-0708.
Description:
Conidiomata not observed on PDA. Conidiophores often monopodial branched, colorless. Conidia, oblate, straight, scarcely curved, 18-23.2×5.5-6.7 μm (=20.2×6.2 μm), 4 septate; basal cell conical to subcylindrical, pale brown or hyaline, thin and smooth-walled, 2.5-4.5 μm long (=3.2 μm); three median cells, 12-15 μm (=13.6 μm), nearly concolor or versicolor, brown to dark brown, septa and periclinal walls darker than the rest of the cell, second cell from base brown to dark brown, 3.7-6 μm long (=4.7 μm), third cell dark brown, 3-5 μm long (=4.2 μm); fourth dark brown, 3.5-5.3 μm long (=4.4 μm); apical cell 2.5-4 μm long (=3.2 μm), conical, hyaline, thin and smooth-walled; 2-4 tubular appendages on the apical cell (often 3), arising from the apex of the apical cell, unbranched, 10-26.5 μm long (=18 μm); single basal appendage, unbranched, tubular, centric or lateral, 2-9 μm long (=5.6 μm). Sexual morph not observed.
Culture characteristics: The colonies reached 70 mm diameter after 4 days on PDA at room temperature, serrated edge, off white, sparse aerial hyphae on the surface appearing radiant, turning grey after sporulation.
Material examined: China, Hainan Province, Wuzhishan city, Shuiman Town, Yongxun village, from the spots on base stem of A. oxyphylla, 8 July 2022, X.F. Cui and Z.G. Hao (YJ11-0708);
Notes:
Based on multigene analyses, Neopestalotiopsis oblatespora is closely related to Neopestalotiopsis guajavicola (FMBCC 11.4), only 2 bp difference between them (1/476 in ITS, 1/378 in tef-1α). However, N. oblatespora is distinct from N. guajavicola with sporulation structure (branched conidiophores of N. oblatespora vs. conidiomata of N. guajavicola); smaller spore (N. oblatespora: 18-23×5.5-6.7 μm, =20.2×6.2 μm vs. N. guajavicola 21.7-24.9×6-7 μm, =23.3×6.5 μm); shorter apical appendages (N. oblatespora: 10-26.5 μm, =18 μm vs. N. guajavicola : 19.1-24.5 μm, =21.8 μm); additionally, having 2-4 apical appendages, while N. guajavicola carrying 2-3 appendages. Moreover, N. oblatespora has no significant recombination with its closely taxa according to the PHI test. Therefore, N. oblatespora is classified as a new species at present study.
Neopestalotiopsis olivaceous X.F. Cui and Z.G. Hao, sp. nov. (Figure 7)
Etymology: named referring to the color of colony.
Holotype: LF25-0709.
Description:
Conidiomata not observed on PDA. Conidia sometimes aggregate into globose, dark green. Conidiophore branches, with spore scars. Conidia, fusiform, straight to obviously irregular curved, 21.5-33.8×5.5-7.7 μm (=26.5×6.3 μm), 4 septate; basal cell conical, hyaline or pale olive, smooth, thin-walled, 2.7-6.2 μm long (=4.5 μm); three median cells 14 to 21.7 μm long (=17 μm), pale olivaceous to olivaceous, concolorous, wall rugose, septa darker than the rest of the cell; second cell from base, pale olivaceous to olivaceous, 3.3 to 8.5 μm long (=5.9 μm); third cell, pale olivaceous to olivaceous, 4 to 6.5 μm long (=5.1 μm); fourth cell, pale olivaceous to olivaceous, 4 to 6.5 μm long (=5.4 μm); apical cell 3.5 to 5.5 μm long (=4.5 μm), hyaline, conic to acute; with 2 to 5 (often 3-4) tubular appendages on the apical cell, inserted at different loci in a crest at the apex of the apical cell, unbranched, 9.5 to 22.5 μm (=14 μm) long; single basal appendage, occasionally no, unbranched, tubular, centric or lateral, 1.2 to 4.8 μm (=2.4 μm) long. Sexual morph not observed.
Culture characteristics: The colonies reached 70 mm diameter on PDA after 7 days growth at room temperature. Colonies appeared circular, white above, medium dense, aerial hyphae on the surface flat; and its reverse was olivaceous, gradually deepened over time.
Material examined: China, Hainan Province, Qiongzhong city, Changzheng Town, Luofan village, from leaf spots of A. oxyphylla, 9 July 2022, X.F. Cui and Z.G. Hao (LF25-0709, holotype); ex-type, Hainan Province, Baoting city, Shiling Town, Shuixian village, from leaf spots of A. oxyphylla, 6 July 2022, X.F. Cui and Z.G. Hao (SX33-0706); Hainan Province, Wuzhishan city, Shuiman Town, Yongxun village, from leaf spots of A. oxyphylla, 8 July 2022, X.F. Cui and Z.G. Hao (YX45-0708).
Notes: Three strains of Neopestalotiopsis olivaceous were isolated from three cities of Hainan, LF25-0709, SX33-0706 and YX45-0708, clustered with well-supported (ML=99%, BI=1). N. olivaceous clusters a sister group with N. amoni (HKAS 124563) and N. zingiberis (GUCC 21001). Molecularly, N. olivaceous can be differed from N. amoni (HKAS 124563) and N. zingiberis (GUCC 21001) according to ITS~ tef-1α~tub2 (1/471 of ITS, 6/347 of TEF with N. amoni; 3/447 of ITS, 14/722 of TUB and 13/358 of TEF with N. zingiberis). Morphologically, N. olivaceous is distinguished with longer conidia (21.5-33.8 μm of N. olivaceous vs. 18-30 μm of N. amoni and 21-31 μm of N. zingiberis), different numbers of apical appendages (2-5 tubular appendages of N. olivaceous vs. 2-3 of N. amoni and 1-3 of N. zingiberis) and longer apical appendages (N. olivaceous 9.5-22.5 μm vs. N. amoni 7-17 μm and N. zingiberis 12-15 μm). The result of PHI test showed no significant recombination among N. olivaceous and its closely taxa. Thus, N. olivaceous is classified as a new species at present study.
Etymology: named referring to the host species, Alpinia oxyphylla.
Holotype: LF55-0709.
Description:
Conidiomata solitary or aggregated, globose, dark, often immersed in PDA. Conidiophores distinct, often degenerated to conidiogenous cells. Conidiogenous cells spherical, hyaline. Conidia, fusiform, straight to slightly curved, 18.8-23.5×5.3-7.0 μm (=21×6.2 μm), 4 septate; basal cell conical to subcylindrical, hyaline, thin and smooth-walled, 2.3-5 μm long (=3.9 μm); three median cells, 11.3-15 μm (=13 μm), versicolor, brown to dark brown, septa and periclinal walls darker than the rest of the cell, wall with verrucae; second cell from base pale brown, paler than the other two cells, 3.3-5.2 μm long (=4.1 μm), third cell dark brown, darker than the other two, 3.5-5.0 μm long (=4.1 μm), fourth pale brown to brown, 3.7-5.4 μm long (=4.4 μm); apical cell 2.8-5 μm long (=3.8 μm), conic to acute, hyaline, thin and smooth-walled; with 2-4 tubular appendages on the apical cell (often 2-3), arising from the apex of the apical cell, occasionally branched, flexuous, 10-25.3 μm long (=18.6 μm); single basal appendage, unbranched, tubular, centric, 2.5-8 μm long (=5 μm). Sexual morph not observed.
Culture characteristics: The colonies reached 70 mm diameter after 9 days on PDA at room temperature, edge circular, off white, dense, central aerial hyphae on the surface raised, with filiform margin; fruit bodies black; reverse similar in color.
Material examined: China, Hainan Province, Qiongzhong city, Changzheng Town, Luofan village, from leaf spots of A. oxyphylla, 9 July 2022, X.F. Cui and Z.G. Hao (LF55-0709, holotype); ex-type, Hainan Province, Wuzhishan city, Maoyang Town, Maohui village, from base stem spots of A. oxyphylla, 8 July 2022, X.F. Cui and Z.G. Hao (MJ31-0708); ex-type, Hainan Province, Baoting city, Nanmao Shengli Farm, from leaf spots of A. oxyphylla, 6 July 2022, X.F. Cui and Z.G. Hao (NM44-0706).
Notes:
Based on multigene analyses, Neopestalotiopsis oxyphylla is closely related to N. brachiata (MFLUCC 17-1555), N. elaeidis (MFLUCC 15-0735), N. petila (MFLUCC 17-1738), N. aotearoa (CBS 367.54) and N. piceana (CBS 394.48), only 0-2 bp difference among them. However, N. oxyphylla is distinct from N. elaeidis with larger spore (N. oxyphylla: 18.8-23.5×5.3-7.0 μm, =21×6.2 μm vs. N. elaeidis 10-20×3-7 μm, =16×5.5 μm) and thinner spore (N. oxyphylla: 5.3-7.0 μm vs. N. aotearoa: 6.5–8.5 μm and N. piceana 7.5–9 μm); N. oxyphylla has different numbers of apical appendages (N. oxyphylla: 2-4; N. brachiata: 1-3; N. aotearoa, N. elaeidis and N. petila: 2-3; N. piceana: 3), and shorter apical appendages(N. oxyphylla: 10-25.3 μm vs. N. brachiata: 9.5-33; N. petila: 22-29 μm, N. piceana: 21-31 μm), but longer than N. aotearoa (5-12 μm). In addition, N. oxyphylla has shorter basal appendage (N. oxyphylla: 2.5-8 μm vs. N. piceana: 6-23 μm). Therefore, N. oxyphylla is classified as a new species at present study.
Neopestalotiopsis wuzhishanensis X.F. Cui and Z.G. Hao, sp. nov. (Figure 9)
Etymology: named referring to the first collection city of Wuzhishan in Hainan province.
Holotype: YX116-0708.
Description:
Conidiomata on PDA solitary, globose, dark. Conidiophores often degenerated to conidiogenous cells. Conidiogenous cells unclear. Conidia, fusiform, straight, scarcely curved, 19.5-26.5×4.5-6.3 μm (=22.4×5.2 μm), 4 septate; basal cell conical to subcylindrical, hyaline, thin and smooth-walled, 2.8-5.5 μm long (=4.2 μm); three median cells, 12.8-16 μm (=14.4 μm), nearly concolor, pale brown, hyaline, septa and periclinal walls darker than the rest of the cell; second cell from base pale brown , 4-6.2 μm long (=5.1 μm), third cell pale brown, 3.5-5.2 μm long (=4.4 μm), fourth pale brown, 3.8-6.3 μm long (=4.7 μm); apical cell 2.7-5.5 μm long (=3.6 μm), conic to acute, hyaline, thin and smooth-walled; with 1-3 tubular appendages on the apical cell (often 1-2), arising from the apex of the apical cell, unbranched, strairht to flexuous, 9-20.8 μm long (=15.4 μm); single or no basal appendage, unbranched, tubular, centric, 0.8-3.8 μm long (=1.9 μm). Sexual morph not observed.
Culture characteristics: The colonies reached 70 mm diameter after 12 days on PDA at room temperature, edge circular, white, medium dense, aerial hyphae on the surface flat, with filiform margin; fruit bodies black. And its reverse was lemon-yellow.
Material examined: China, Hainan Province, Wuzhishan city, Shuiman Town, Yongxun village, from leaf spot of A. oxyphylla, 8 July 2022, X.F. Cui and Z.G. Hao (YX116-0708).
Notes:
Neopestalotiopsis wuzhishanensis clusters a sister group to Neopestalotiopsis cubana (CBS 600.96). While, N. wuzhishanensis is different from N. cubana depending on ITS, tef-1α and tub2 sequences (3/481 in ITS, 4/446 in tef-1α, 3/720 in tub2). Additionally, there are remarkably discrepancies in morphological characteristics, N. wuzhishanensis is thinner (N. wuzhishanensis: 4.5-6.3 μm, =5.2 μm vs. N. cubana 8-9.5 μm, =8.8 μm), shorter in apical appendages (N. wuzhishanensis: 9-20.8 μm, =15.4 μm vs. N. cubana: 21-27 μm, =24 μm) and base appendage (N. wuzhishanensis: 0.8-3.8 μm, =1.9 μm vs. N. cubana: 4-7 μm); additionally, three media cells of N. wuzhishanensis are paler than N. cubana; furthermore, N. cubana having 1-3 apical appendages, while N. cubana carrying 2-4 appendages. The result of PHI test showed that N. wuzhishanensis has no significant recombination with their closely taxa. Therefore, N. wuzhishanensis is classified as a new species at present study.
Neopestalotiopsis yongxunensis X.F. Cui and Z.G. Hao, sp. nov. (Figure 10)
Etymology: named referring to the first collection village of Yongxun in Hainan province.
Holotype: YX101-0708.
Description:
Conidiomata on PDA solitary or aggregated, globose, dark, embedded or semi-immersed. Conidiophores often degenerated to conidiogenous cells. Conidiogenous cells unclear. Conidia, fusiform, straight to curved, 18.2-25.5×5.8-7.5 μm (=21.6×6.6 μm), 4 septate; basal cell conical, hyaline, thin and smooth-walled, 3.0-5.2 μm long (=4.1 μm); three median cells, 12-15.2 μm (=13.7 μm), versicolor, pale brown to brown, septa and periclinal walls darker than the rest of the cell; second cell from base pale brown, paler than the other two cells, 3.5-5.3 μm long (=4.3 μm), third cell brown, darker than the other two, 3.8-5.3 μm long (=4.6 μm), fourth cell brown, 4.0-5.2 μm long (=4.6 μm); apical cell 2.5-5.0 μm long (=3.7 μm), conic to subcylindrical, hyaline, thin and smooth-walled; with 2-4 tubular appendages on the apical cell, arising from the apex of the apical cell, filiform, unbranched, strairht to flexuous, 10.5-24.7 μm long (=18.2 μm); single basal appendage, unbranched, tubular, centric, 1.7-7 μm long (=4.2 μm). Sexual morph not observed.
Culture characteristics: The colonies reached 70 mm diameter after 4 days on PDA at room temperature, edge circular, white, dense aerial mycelium on the surface; reverse similar in color. Fruit bodies black, mostly under the hyphae, visible on the back.
Material examined: China, Hainan Province, Wuzhishan city, Shuiman Town, Yongxun village, from leaf spot of A. oxyphylla, 8 July 2022, X.F. Cui and Z.G. Hao (YX101-0708);
Notes:
Neopestalotiopsis yongxunensis is related to N. dendrobii (MFLUCC 14-0106) and N. paeonia-suffruticosa (HKAS 123212) in the phylogenetic analysis. While, N. yongxunensis can be differed from N. dendrobii and N. paeonia-suffruticosa depending on ITS, tef1-α and tub2 sequences, 7 bp difference (2/284 in tef1-α, 5/441 in tub2) with N. dendrobii and 10 bp difference (9/440 in tef1-α, 1/740 in tub2) with N. paeonia-suffruticosa. In addition, there are remarkably discrepancies in morphological characteristics, N. yongxunensis is thinner in conidia (N. yongxunensis: 5.8-7.5 μm, =6.6 μm vs. N. paeonia-suffruticosa: 9-11 μm, =9.5 μm), has different numbers of apical appendages (N. yongxunensis 2-4 vs. N. paeonia-suffruticosa 3-4) and shorter apical appendages (N. yongxunensis: 10.5-24.7 μm vs. N. paeonia-suffruticosa 22.5-34 μm). While N. yongxunensis differs from N. dendrobii in having longer apical appendages (N. yongxunensis: 10.5-24.7 μm vs. N. dendrobii 5-6.5 μm) with different numbers (N. yongxunensis 2-4 vs. N. dendrobii 2-3). Furthermore, the PHI test indicates that there is no significant recombination between N. yongxunensis and its closely species. Therefore, N. yongxunensis is classified as a new species at present study.
3.4. Pathogenicity Assay
Sixteen of the 20 tested Pestalotioid isolates were able to cause typical brown lesions after inoculation, while the other 4 isolates did not, including Neopestalotiopsis sp.4 MH133-0708, N. coffeae-arabicae NM42-0706, N. oblatespora YJ11-0708 and Neopestalotiopsis sp.3 SX11-0706. The lesion areas measured 5 days after inoculation were 54.02, 13.86, 15.57, 4.65, 11.08, 117.40, 100.63, 82.31, 8.55, 80.25, 32.03, 7.02, 104.86, 84.48, 102.04 and 16.17 mm2 for isolates of P. hydei BA11-0708, Ps. myanmarina JR34-0709, Ps. avicenniae LF48-0709, N. coffeae-arabicae BL32-0708, N. coffeae-arabicae LF51-0709, N. cubana MH51-0708, N. cubana YX112-0708, N. wuzhishanensis YX116-0708, N. yongxunensis YX101-0708, N. baotingensis SX41-0706, N. vaccinii JR31-0709, N. rosicola NM47-0708, N. oxyphylla LF55-0709, N. brachiata SX31-0706, Neopestalotiopsis sp.5 XC11-0709 and N. olivaceous LF25-0709, respectively (Figure 11). The morphology of purified fungi re-isolated from the lesion of inoculation was identical with those of the isolateds used for inoculation, which were also confirmed by PCR and gene sequences. The results of pathogenicity and phylogenetic analysis showed that the strains close to N. cubana and N. brachiata had stronger pathogenicity (Figure 1 and Figure 11B).
4. Discussion
In this study, 36 pestalotioid strains were isolated. According to multi-locus phylogeny (ITS, tef-1α and tub2) and morphological characteristics analyses, one Pestalotiopsis sp, two Pseudopestalotiopsis spp., and 14 Neopestalotiopsis spp. were identified. Six new species (N. baotingensis, N. oblatespora, N. olivaceous, N. oxyphylla, N. wuzhishanensis and N. yongxunensis) were described. Among 36 strains, the isolation frequency of N. coffeae-arabicae and N. cubana was both 16.67%, higher than the others;additionally, N. coffeae-arabicae, N. olivaceous and N. oxyphylla were isolated from 5, 3 and 3 cities separately, with more widely distribution in Hainan than others. This is the first systematic report of Neopestalotiopsis, Pestalotiopsis and Pseudopestalotiopsis fungi related with A. oxyphylla in its main planted area.
The development of molecular biology has greatly facilitated the identification of microorganisms, and the phylogeny analyses of combined ITS, tef-1α and tub2 can better distinguish Neopestalotiopsis, Pestalotiopsis and Pseudopestalotiopsis. For example, N. olivaceous and N. wuzhishanensis in this study do not conform to the morphological characteristics of versicolorous median cells depicted in Neopestalotiopsis. This phenomenon was also mentioned by Sun et al [16], so the phylogeny analyses can overcome the discrimination of the three genera only by intermediate color cells. While the three gene sequences of N. oxyphylla, N. aotearoa and N. brachiata are closely similar only with 0-2 bp difference of the combined sequence, also between N. oblatespora and N. guajavicola with 2 bp difference, while they have obvious discrepancy in morphological characteristics. Similar phenomenon was observed between N. alpapicalis MFLUCC 17-2544T and N. rhizophorae MFLUCC 17-1551T [41]. Therefore, more gene fragments need to be introduced in order to further differentiate closly related species of pestalotioid fungi.
RLB disease is an important disease in the cultivation process of A. oxyphylla according to previous reports. Its pathogen was reported as Pestalotia palmarum in 1986[11], now classified as Pestalotiopsis palmarum. While the ring brown spot (RBS) disease with similar symptom to RLB disease was caused by Pestalosphaeria alpinia, the sexual morph of Pestalotioid, reported in 1994[25]. Perhaps due to the differences in classification method and limitation on sample size, P. palmarum and P. alpinia were not isolated in this study,which explained the potential diversity of pestalotioid fungi in this host needed to be further explored. In addition, the symptoms of RLB and RBS disease are similar both caused by pestalotioid fungi with different morph, so it is recommended to merge the two diseases into one for future research and disease management.
Through the pathogenicity tests of 20 pestalotioid strains, most species can cause obvious symptoms on the leaves indicating the diversity pathogen of RLB disease, and the Neopestalotiopsis species (the lesion area over 75 mm2 of 7 species) were more tended to infect A. oxyphylla and caused more serious disease than Pestalotiopsis (the lesion area about 50 mm2) and Pseudopestalotiopsis (less than 50 mm2 both of the two). The reports that the disease caused by Neopestalotiopsis fungi were more in recent years [16]. In addition, all pathogenicity tests were carried out with a single cultivar of A. oxyphylla and constant environmental conditions. As we all know, differences in varieties and changes in environmental conditions can both affect the occurrence of diseases. Therefore, more attempts need to be performed on different varieties under different environmental conditions.
What is worth noting is that most of pestalotioid species have a broad range of hosts, and one species of pestalotioid species can infect several economic plants, while a plant can be harmed by several pestalotioid fungi. For example, N. cubana can infect rubber trees [96], Camellia oleifera [17] and Ixora chinensis [97], and a new leaf fall disease of rubber trees was caused by N. aotearoa, N. cubana and N. formicarum [96]. A. oxyphylla is a semi-shade plant mainly planted under rubber forests. In this study, 6 strains of N. cubana, one strain of Neopestalotiopsis sp.3 SX11-0706 clustered with N. formicarum and 5 strains (N. oxyphylla, N. brachiata and Neopestalotiopsis sp.5 XC11-0709) closely related to N. aotearoa were isolated. It refers that some pestalotioid species may infect both rubber tree and A. oxyphylla. The promotion of cultivation medicinal plant under the forest should also pay attention to the occurrence of cross-infection diseases, in order to prevent them in advance.
A comprehensive understanding of the species and genetic diversity of pathogens is the foundation for sustainable disease management. Since there is no research about the resistance varieties of A. oxyphylla to RLB disease. The strains with different characteristics and pathogenicity isolated in this study may provide a material basis for subsequent screening of resistant varieties, including highly active biological and chemical agents friendly to the environment.
Author Contributions
Formal analysis, Xiufen Cui; Methodology, Zhigang Hao and Yingbin Li; Resources, Xiufen Cui, Zhigang Hao and Jinan Zhang; Software, Shuang Song; Supervision, Jianqiang Li, Yixiang Liu and Laixin Luo; Visualization, Menghuai Chen; Writing – original draft, Xiufen Cui. All authors read and approved the manuscript.
Data Availability Statement
All data supporting the findings discussed in this article can be obtained from the corresponding author upon reasonable request.
Acknowledgements
This work was supported by the Major Science and Technology Project in Yunnan Province (202102AE090042-02) and the leading fund project of Sanya Research Institute of China Agricultural University (SYND-2022-11).
Conflict of Interest
The authors declare no conflict of interest.
References
- Qiu, L.Y. Investigation of germplasm resources of Alpinia oxyphylla Miq. in Hainan Island. Master's thesis,Hainan University, Hainan, China, 2015.
- Pharmacopoeia of the People's Republic of China. Alpiniae oxyphyllae fructus; The Medicine Science and Technology Press of China, China, 2020, 1, 303.
- Zhang, Q.; Zheng, Y.L.; Hu, X.J.; Hu, X.L.; Lv, W.W.; Lv, D.; Chen, J.J.; Wu, M.L.; Song, Q.C.; Shentu, J.Z. Ethnopharmacological uses, phytochemistry, biological activities, and therapeutic applications of Alpinia oxyphylla Miguel: A review. Journal of Ethnopharmacology 2018, 224, 149-168. [CrossRef]
- Zhang, J.Q.; Wang, S.; Li, Y.H.; Xu, P.; Chen, F.; Tan, Y.F.; Duan, J.A. Anti-diarrheal constituents of Alpinia oxyphylla. Fitoterapia 2013, 89, 149-156. [CrossRef]
- Zhou, J.Z.; Zhang, K.; Zhang, L.Y.; Xian, Q.Q.; Sun, P.Z.; Du, Z.Y. Geographical distribution and prediction of potential region of Alpinia oxyphylla Miq. Anhui Agric. Sci.2018, 46, 1-3+8. [CrossRef]
- Wang, Y.F. Exploration of metabolites and related genes of volatile oil from wild Alpinia oxyphylla Miq. in Hainan Province. Central University for Nationalities, Beijing, China, 2022.
- Chen, L.Z.; Liufu, Y.Q.; Lin, S.C.; Xian, S.Q. Analysis of the basic biological characteristics and high yield cultivation techniques of Alpinia oxyphylla Miq. South China Agriculture 2018, 12, 27+29. [CrossRef]
- Zhang, C.; Wu, Y.G.; Yang, H.G.; Yao, G.L.; Zhang, J.F.; Yu, J.; Yang, D.M.; Chen, P. The current planting situation and industrial development countermeasures of Alpinia oxyphylla Miq. in Hainan Province. Modern Horticulture 2021, 44, 63-67. [CrossRef]
- He, H.M. Analysis of biological characteristics and environment for high production of Alpinia Oxyphylla. Chinese Journal of Mordern Applied Pharmacy 1992, 205-207.
- Yan, X.X.; Ren, B.L.; Wang, M.Y.; Wang, Q.L.; Yang, Q.; Tang, H.; Wang, Z.N. Present situation and development strategy of Alpinia oxyphylla. China Journal of Chinese Materia Medica 2019, 44, 1960-1964. [CrossRef]
- Hong, X.Q.; Chen, J.J. Three important new diseases of Alpinia oxyphylla Miq. Chinese Journal of Tropical Agriculture 1986, 50-53.
- Xu, X.R.; Zhou, J.; Wu, X.P.; Lei, X.T. The main diseases and pests of Alpinia oxyphylla and their prevention and control. China Tropical Agriculture 2012, 48, 56-58.
- Fu, M.Y.; Zeng, X.P.; Rui, K.; Wang, H.F.; Chen, M.C. The occurrence and control of the leaf blight disease of Alpinia oxyphylla in Hainan Island. Modern Agricultural Science and Technology 2015, 655, 170-171.
- Maharachchikumbura, S.S.N.; Guo, L.D.; Cai, L.; Chukeatirote, E.; Wu, W.P.; Sun, X.; Crous, P.W.; Bhat, D.J.; McKenzie, E.H.C.; Bahkali, A.H.; et al. A multi-locus backbone tree for Pestalotiopsis, with a polyphasic characterization of 14 new species. Fungal Diversity 2012, 56, 95-129. [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Studies in Mycology 2014, 121-186. [CrossRef]
- Sun, Y.R.; Jayawardena, R.S.; Sun, J.E.; Wang, Y. Pestalotioid species associated with medicinal plants in southwest China and Thailand. Microbiology Spectrum 2023, 11, e0398722. [CrossRef]
- Li, L.L.; Yang, Q.; Li, H. Morphology, phylogeny, and pathogenicity of pestalotioid species on Camellia oleifera in China. Journal of Fungi 2021, 7, 1080. [CrossRef]
- Diogo, E.; Goncalves, C.I.; Silva, A.C.; Valente, C.; Braganca, H.; Phillips, A.J.L. Five new species of Neopestalotiopsis associated with diseased Eucalyptus spp. in Portugal. Mycological Progress 2021, 20, 1441-1456. [CrossRef]
- Lin, H.F.; Chen, T.H.; Liu, S.D. Bioactivity of antifungal substance iturin a produced by Bacillus subtilis strain BS-99-H against Pestalotiopsis eugeniae, a causal pathogen of wax apple fruit rot. Plant Pathology Bulletin 2010, 19, 225-233.
- Prasannath, K.; Shivas, R.G.; Galea, V.J.; Akinsanmi, O.A. Neopestalotiopsis species associated with flower diseases of Macadamia integrifolia in Australia. Journal of Fungi 2021, 7, 771. [CrossRef]
- Tsai, I.; Chung, C.L.; Lin, S.R.; Hung, T.H.; Shen, T.L.; Hu, C.Y.; Hozzein, W.N.; Ariyawansa, H.A. Cryptic diversity, molecular systematics, and pathogenicity of genus Pestalotiopsis and allied genera causing gray blight disease of tea in Taiwan, With a description of a new Pseudopestalotiopsis species. Plant Disease 2021, 105, 425-443. [CrossRef]
- Maharachchikumbura, S.S.N.; Larignon, P.; Hyde, K.D.; Al-Sadi, A.M.; Liu, Z.Y. Characterization of Neopestalotiopsis, Pestalotiopsis and Truncatella species associated with grapevine trunk diseases in France. Phytopathologia Mediterranea 2016, 55, 380-390. [CrossRef]
- Ayoubi, N.; Soleimani, M.J. Strawberry fruit rot caused by Neopestalotiopsis iranensis sp nov., and N. mesopotamica. Current Microbiology 2016, 72, 329-336. [CrossRef]
- Chamorro, M.; Aguado, A.; De los Santos, B. First report of root and crown rot caused by Pestalotiopsis clavispora (Neopestalotiopsis clavispora) on strawberry in Spain. Plant Disease 2016, 100, 1495-1495. [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. Pestalotiopsis-morphology, phylogeny, biochemistry and diversity. Fungal Diversity 2011, 50, 167-187. [CrossRef]
- Qi, P.K. Fungal diseases of cultivated medicinal plants in Guangdong Province; Guangdong Science and Technology Press: Guangdong, China, 1994.
- Jeewon, R.; Liew, E.C.Y.; Hyde, K.D. Phylogenetic relationships of Pestalotiopsis and allied genera inferred from ribosomal DNA sequences and morphological characters. Molecular Phylogenetics and Evolution 2002, 25, 378-392. [CrossRef]
- Jeewon, R.; Liew, E.C.Y.; Simpson, J.A.; Hodgkiss, I.J.; Hyde, K.D. Phylogenetic significance of morphological characters in the taxonomy of Pestalotiopsis species. Molecular Phylogenetics and Evolution 2003, 27, 372-383. [CrossRef]
- Gualberto, G.F.; Catarino, A.D.; Sousa, T.F.; Cruz, J.C.; Hanada, R.E.; Caniato, F.F.; Silva, G.F. Pseudopestalotiopsis gilvanii sp. nov. and Neopestalotiopsis formicarum leaves spot pathogens from guarana plant: a new threat to global tropical hosts. Phytotaxa 2021, 489, 121-139. [CrossRef]
- He, Y.K.; Yang, Q.; Sun, Y.R.; Zeng, X.Y.; Jayawardena, R.S.; Hyde, K.D.; Wang, Y. Additions to Neopestalotiopsis (Amphisphaeriales, Sporocadaceae) fungi: two new species and one new host record from China. Biodiversity Data Journal 2022, 10, e90709. [CrossRef]
- Li, W.L.; Dissanayake, A.J.; Zhang, T.; Maharachchikumbura, S.S.N.; Liu, J.K. Identification and pathogenicity of pestalotioid fungi associated with woody oil plants in Sichuan Province, China. Journal of Fungi 2022, 8. [CrossRef]
- Prematunga, C.J.Y., L.Q.; Gomdola, D.; Balasuriya, A.; Yang, Y.H.; Jayawardena, R.S.; Luo, M. An addition to pestalotioid fungi in China: Neopestalotiopsis fragariae sp. nov. causing leaf spots on Fragaria × ananassa. Asian Journal of Mycology 2022, 5, 220-238, https://asianjournalofmycology.org/pdf/AJOM_5_2_10-1.
- Yang, Q.; He, Y.K.; Yuan, J.; Wang, Y. Two new Pseudopestalotiopsis species isolated from Celtis sinensis and Indocalamus tessellatus plants in southern China. Phytotaxa 2022, 543, 274-282. [CrossRef]
- Yang, Q.; Zeng, X.Y.; Yuan, J.; Zhang, Q.; He, Y.K.; Wang, Y. Two new species of Neopestalotiopsis from southern China. Biodiversity Data Journal 2021, 9. [CrossRef]
- White, T.J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Pcr Protocols, Academic Press, Inc, 1990, 315-322.
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553-556. [CrossRef]
- O'Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proceedings of the National Academy of Sciences of the United States of America 1998, 95, 2044-2049. [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Applied and Environmental Microbiology 1995, 61, 1323-1330. [CrossRef]
- Odonnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Molecular Phylogenetics and Evolution 1997, 7, 103-116. [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171-180. [CrossRef]
- Norphanphoun, C.; Jayawardena, R.S.; Chen, Y.; Wen, T.C.; Meepol, W.; Hyde, K.D. Morphological and phylogenetic characterization of novel pestalotioid species associated with mangroves in Thailand. Mycosphere 2019, 10, 531-578. [CrossRef]
- Kumar, V.; Cheewangkoon, R.; Gentekaki, E.; Maharachchikumbura, S.S.N.; Brahmanage, R.S.; Hyde, K.D. Neopestalotiopsis alpapicalis sp. nov. a new endophyte from tropical mangrove trees in Krabi Province (Thailand). Phytotaxa 2019, 393, 251-262. [CrossRef]
- Bezerra, J.D.P.; Machado, A.R.; Firmino, A.L.; Rosado, A.W.C.; de Souza, C.A.F.; de Souza-Motta, C.M.; Freire, K.; Paiva, L.M.; Magalhaes, O.M.C.; Pereira, O.L.; et al. Mycological diversity description I. Acta Botanica Brasilica 2018, 32, 656-666. [CrossRef]
- Jiang, N.; Fan, X.; Tian, C. Identification and characterization of leaf-inhabiting fungi from Castanea Plantations in China. Journal of Fungi (Basel, Switzerland) 2021, 7. [CrossRef]
- Liu, X.F.; Tibpromma, S.; Zhang, F.; Xu, J.C.; Chethana, K.W.T.; Karunarathna, S.C.; Mortimer, P.E. Neopestalotiopsis cavernicola sp. nov. from Gem Cave in Yunnan Province, China. Phytotaxa 2021, 512, 1-27. [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: micro-fungi associated with Pandanaceae. Fungal Diversity 2018, 93, 1-160. [CrossRef]
- Song, Y.; Geng, K.; Zhang, B.; Hyde, K.D.; Zhao, W.S.; Wei, J.G.; Kang, J.C.; Wang, Y. Two new species of Pestalotiopsis from Southern China. Phytotaxa 2013, 126, 22-30. [CrossRef]
- Peng, C.; Crous, P.W.; Jiang, N.; Fan, X.; Liang, Y.; Tian, C. Diversity of Sporocadaceae (pestalotioid fungi) from rosa in China. Persoonia 2022, 49, 201-260. [CrossRef]
- Ma, X.Y.; Maharachchikumbura, S.S.N.; Chen, B.W.; Hyde, K.D.; McKenzie, E.H.C.; Chomnunti, P.; Kang, J.C. Endophytic pestalotioid taxa in Dendrobium orchids. Phytotaxa 2019, 419, 268-286. [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Le Roux, J.J.; Richardson, D.M.; Strasberg, D.; Shivas, R.G.; Alvarado, P.; Edwards, J.; Moreno, G.; Sharma, R.; et al. Fungal Planet description sheets: 371-399. Persoonia 2015, 35, 264-327. [CrossRef]
- Konta, S.; Tibpromma, S.; Karunarathna, S.C.; Samarakoon, M.C.; Steven, L.S.; Mapook, A.; Boonmee, S.; Senwanna, C.; Balasuriya, A.; Eungwanichayapant, P.D.; et al. Morphology and multigene phylogeny reveal ten novel taxa in Ascomycota from terrestrial palm substrates (Arecaceae) in Thailand. Mycosphere 2023, 14, 107-152. [CrossRef]
- Ul Haq, I.; Ijaz, S.; Khan, N.A. Genealogical concordance of phylogenetic species recognition-based delimitation of Neopestalotiopsis species associated with leaf spots and fruit canker disease affected guava plants. Pakistan Journal of Agricultural Sciences 2021, 58, 1301-1313. [CrossRef]
- Zhang, Z.; Liu, R.; Liu, S.; Mu, T.; Zhang, X.; Xia, J. Morphological and phylogenetic analyses reveal two new species of Sporocadaceae from Hainan, China. MycoKeys 2022, 88, 171-192. [CrossRef]
- Freitas, E.F.S.; Da Silva, M.; Barros, M.V.P.; Kasuya, M.C.M. Neopestalotiopsis hadrolaeliae sp. nov., a new endophytic species from the roots of the endangered orchid Hadrolaelia jongheana in Brazil. Phytotaxa 2019, 416, 211-220. [CrossRef]
- Huanaluek, N.; Jayawardena, R.S.; Maharachchikum-Bura, S.S.N.; Harishchandra, D.L. Additions to pestalotioid fungi in Thailand: Neopestalotiopsis hydeana sp. nov. and Pestalotiopsis hydei sp. nov. Phytotaxa 2021, 479, 23-43. [CrossRef]
- Akinsanmi, O.A.; Nisa, S.; Jeff-Ego, O.S.; Shivas, R.G.; Drenth, A. Dry flower disease of macadamia in Australia caused by Neopestalotiopsis macadamiae sp. nov. and Pestalotiopsis macadamiae sp. nov. Plant Disease 2017, 101, 45-53. [CrossRef]
- Maharachchikumbura, S.S.; Guo, L.D.; Chukeatirote, E.; Hyde, K.D. Improving the backbone tree for the genus Pestalotiopsis; addition of P. steyaertii and P. magna sp. nov. Mycological Progress 2014, 13, 617-624. [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.H.; Gilchrist, C.L.M.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N.; et al. Fungal Planet description sheets: 1042-1111. Persoonia 2020, 44, 301-459. [CrossRef]
- Liu, F.; Bonthond, G.; Groenewald, J.Z.; Cai, L.; Crous, P.W. Sporocadaceae, a family of coelomycetous fungi with appendage-bearing conidia. Studies in Mycology 2019, 92, 287-415. [CrossRef]
- Silverio, M.L.; Calvacanti, M.A.d.Q.; Silva, G.A.d.; Oliveira, R.J.V.d.; Bezerra, J.L. A new epifoliar species of Neopestalotiopsis from Brazil. Agrotropica 2016, 28, 151-158.
- Crous, P.W.; Summerell, B.A.; Swart, L.; Denman, S.; Taylor, J.E.; Bezuidenhout, C.M.; Palm, M.E.; Marincowitz, S.; Groenewald, J.Z. Fungal pathogens of Proteaceae. Persoonia 2011, 27, 20-45. [CrossRef]
- Chaiwan, N.; Jeewon, R.; Pem, D.; Jayawardena, R.S.; Nazurally, N.; Mapook, A.; Promputtha, I.; Hyde, K.D. Fungal species from Rhododendron sp.: Discosia rhododendricola sp. nov, Neopestalotiopsis rhododendricola sp. nov. and Diaporthe nobilis as a new host record. Journal of Fungi (Basel, Switzerland) 2022, 8. [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D. Additions to Pestalotiopsis in Taiwan. Mycosphere 2018, 9, 999-1013. [CrossRef]
- Santos, J.; Hilario, S.; Pinto, G.; Alves, A. Diversity and pathogenicity of pestalotioid fungi associated with blueberry plants in Portugal, with description of three novel species of Neopestalotiopsis. European Journal of Plant Pathology 2022, 162, 539-555. [CrossRef]
- Fiorenza, A.; Gusella, G.; Aiello, D.; Polizzi, G.; Voglmayr, H. Neopestalotiopsis siciliana sp. nov. and N. rosae causing stem lesion and dieback on avocado plants in Italy. Journal of Fungi 2022, 8. [CrossRef]
- Jayawardena, R.S.; Liu, M.; Maharachchikumbura, S.S.N.; Zhang, W.; Xing, Q.; Hyde, K.D.; Nilthong, S.; Li, X.; Yan, J. Neopestalotiopsis vitis sp. nov. causing grapevine leaf spot in China. Phytotaxa 2016, 258, 63-74. [CrossRef]
- Liu, X.F.; Tibpromma, S.; Hughes, A.C.; Chethana, K.W.T.; Wijayawardene, N.N.; Dai, D.Q.; Du, T.Y.; Elgorban, A.M.; Stephenson, S.L.; Suwannarach, N.; et al. Culturable mycota on bats in central and southern Yunnan Province, China. Mycosphere 2023, 14, 497-662. [CrossRef]
- Liu, F.; Hou, L.w.; Raza, M.; Cai, L. Pestalotiopsis and allied genera from Camellia, with description of 11 new species from China. Scientific Reports 2017, 7. [CrossRef]
- Gu, R.; Bao, D.F.; Shen, H.W.; Su, X.J.; Li, Y.X.; Luo, Z.L. Endophytic Pestalotiopsis species associated with Rhododendron in Cangshan Mountain, Yunnan Province, China. Frontiers in Microbiology 2022, 13. [CrossRef]
- Crous, P.W.; Boers, J.; Holdom, D.; Osieck, E.R.; Steinrucken, T.V.; Tan, Y.P.; Vitelli, J.S.; Shivas, R.G.; Barrett, M.; Boxshall, A.G.; et al. Fungal Planet description sheets: 1383–1435. Persoonia 2022, 48, 261-371. [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.W.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Lucking, R.; et al. Fungal diversity notes 111-252-taxonomic and phylogenetic contributions to fungal taxa. Fungal Diversity 2015, 75, 27-274. [CrossRef]
- Chaiwan, N.; Wanasinghe, D.N.; Mapook, A.; Jayawardena, R.S.; Norphanphoun, C.; Hyde, K.D. Novel species of Pestalotiopsis fungi on Dracaena from Thailand. Mycology 2020, 11, 306-315. [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1-110: taxonomic and phylogenetic contributions to fungal species. Fungal Diversity 2015, 72, 1-197. [CrossRef]
- Silva, N.; Maharachchikumbura, S.; Thambugala, K.; Bhat, D.J.; Hyde, K.D. Morpho-molecular taxonomic studies reveal a high number of endophytic fungi from Magnolia candolli and M. garrettii in China and Thailand. Mycosphere 2021, 12, 163–237.
- Yan, M.Z.; Maharachchikumbura, S.; Tian, Q.; Hyde, K.D. Pestalotiopsis species on ornamental plants in Yunnan Province, China. Sydowia -Horn 2013, 65, 113–128.
- Maharachchikumbura, S.S.N.; Chukeatirote, E.; Guo, L.D.; Crous, P.W.; Mckenzie, E.H.C.; Hyde, K.D. Pestalotiopsis species associated with Camellia sinensis (tea). Mycotaxon 2013, 123, 47-61(15). [CrossRef]
- Monteiro, P.; Gonçalves, M.F.M.; Pinto, G.; Silva, B.; Martín-García, J.; Diez, J.J.; Alves, A. Three novel species of fungi associated with pine species showing needle blight-like disease symptoms. European Journal of Plant Pathology 2022, 162, 183-202. [CrossRef]
- Hyde, K.D.; Jeewon, R.; Chen, Y.-J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.-B.; Lin, C.-G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: is the descriptive curve flattening? Fungal Diversity 2020, 103, 219-271. [CrossRef]
- Das, K.; Lee, S.Y.; Jung, H.Y. Pestalotiopsis kaki sp. nov., a Novel species isolated from persimmon tree (Diospyros kaki) Bark in Korea. Mycobiology 2020, 49, 54-60. [CrossRef]
- Chen, Y.Y.; Maharachchikumbura, S.S.N.; Liu, J.K.; Hyde, K.D.; Nanayakkara, R.R.; Zhu, G.S.; Liu, Z.Y. Fungi from Asian Karst formations I. Pestalotiopsis photinicola sp nov., causing leaf spots of Photinia serrulata. Mycosphere 2017, 8, 103-110. [CrossRef]
- Harishchandra, D.L.; Aluthmuhandiram, J.V.S.; Yan, J.; Hyde, K.D. Molecular and morpho-cultural characterisation of Neopestalotiopsis and Pestalotiopsis species associated with ornamental and forest plants in China. Lett Appl Microbiol 2020, 73(3), 352-362.
- Zhang, Y.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Hyde, K.D. A novel species of Pestalotiopsis causing leaf spots of Trachycarpus fortunei. Cryptogamie Mycologie 2012, 33, 311-318. [CrossRef]
- Wei, J.G.; Phan, C.K.; Wang, L.; Xu, T.; Luo, J.T.; Sun, X.; Guo, L.D. Pestalotiopsis yunnanensis sp. nov., an endophyte from Podocarpus macrophyllus (Podocarpaceae) based on morphology and ITS sequence data. Mycological Progress 2013, 12, 563-568. [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Liu, Z.Y.; Hyde, K.D. Pseudopestalotiopsis ignota and Ps. camelliae spp. nov associated with grey blight disease of tea in China. Mycological Progress 2016, 15. [CrossRef]
- Catarino, A.d.M.; Hanada, R.E.; de Queiroz, C.A.; Sousa, T.F.; Lima, Í.N.; Gasparotto, L.; da Silva, G.F. First report of Pseudopestalotiopsis dawaina causing spots in Caryota mitis in Brazil. Plant Disease 2020, 104, 989. [CrossRef]
- Nozawa, S.; Ando, K.; Phay, N.; Watanabe, K. Pseudopestalotiopsis dawaina sp. nov. and Ps. kawthaungina sp. nov.: two new species from Myanmar. Mycological Progress 2018, 17, 865-870. [CrossRef]
- Tsai, I.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Ariyawansa, H.A. Molecular phylogeny, morphology and pathogenicity of Pseudopestalotiopsis species on Ixora in Taiwan. Mycological Progress 2018, 17, 941-952. [CrossRef]
- Lateef, A.A.; Sepiah, M.; Bolhassan, M.h. Description of Pseudopestalotiopsis kubahensis sp. nov., a new species of microfungi from Kubah National Park, Sarawak, Malaysia. Current Research in Environmental & Applied Mycology 2015. 5, 376–381.
- Nozawa, S.; Yamaguchi, K.; Yen, L.T.H.; Hop, D.V.; Phay, N.; Ando, K.; Watanabe, K. Identification of two new species and a sexual morph from the genus Pseudopestalotiopsis. Mycoscience 2017, 58, 328-337. [CrossRef]
- Song, Y.; Tangthirasunun, N.; Maharachchikumbura, S.S.N.; Jiang, Y.L.; Xu, J.J.; Hyde, K.D.; Wang, Y. Novel Pestalotiopsis species from Thailand point to the rich undiscovered diversity of this chemically creative genus. Cryptogamie Mycologie 2014, 35, 139-149. [CrossRef]
- Miller, M.A.; Pfeiffer, W.T.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 2010, 1-8. [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572-1574. [CrossRef]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1-40. [CrossRef]
- Bruen, T.C.; Philippe, H.; Bryant, D. A simple and robust statistical test for detecting the presence of recombination. Genetics 2006, 172, 2665-2681. [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Molecular Biology and Evolution 2006, 23, 254-267. [CrossRef]
- Pornsuriya, C.; Chairin, T.; Thaochan, N.; Sunpapao, A. Identification and characterization of Neopestalotiopsis fungi associated with a novel leaf fall disease of rubber trees (Hevea brasiliensis) in Thailand. Journal of Phytopathology 2020, 168, 416-427. [CrossRef]
- Khoo, Y.W.; Tan, H.T.; Khaw, Y.S.; Li, S.F.; Chong, K.P. First report of Neopestalotiopsis cubana causing leaf blight on Ixora chinensis in Malaysia. Plant Disease 2022, 106, 2749-2749. [CrossRef]
Figure 1.
RAxML tree of Neopestalotiopsis isolates based on ITS, tef-1α and tub2 sequences. The roots of this tree are Pestalotiopsis diversiseta MFLUCC 12-0287 and P. colombiensis CBS 118553. The strains isolated in this study are marked in red. Ex-type strains are marked with T. ML bootstrap values ≥50% and BI probabilities (in red) ≥0.90 are displayed at the nodes.
Figure 1.
RAxML tree of Neopestalotiopsis isolates based on ITS, tef-1α and tub2 sequences. The roots of this tree are Pestalotiopsis diversiseta MFLUCC 12-0287 and P. colombiensis CBS 118553. The strains isolated in this study are marked in red. Ex-type strains are marked with T. ML bootstrap values ≥50% and BI probabilities (in red) ≥0.90 are displayed at the nodes.

Figure 2.
RAxML tree of Pestalotiopsis isolates based on ITS, tef-1α and tub2 sequences. The root of this tree is N. cubana CBS 600.9. The strains isolated in this study are marked in red. Ex-type strains are marked with T. ML bootstrap values ≥50% and BI probabilities (in red) ≥0.90 are displayed at the nodes.
Figure 2.
RAxML tree of Pestalotiopsis isolates based on ITS, tef-1α and tub2 sequences. The root of this tree is N. cubana CBS 600.9. The strains isolated in this study are marked in red. Ex-type strains are marked with T. ML bootstrap values ≥50% and BI probabilities (in red) ≥0.90 are displayed at the nodes.
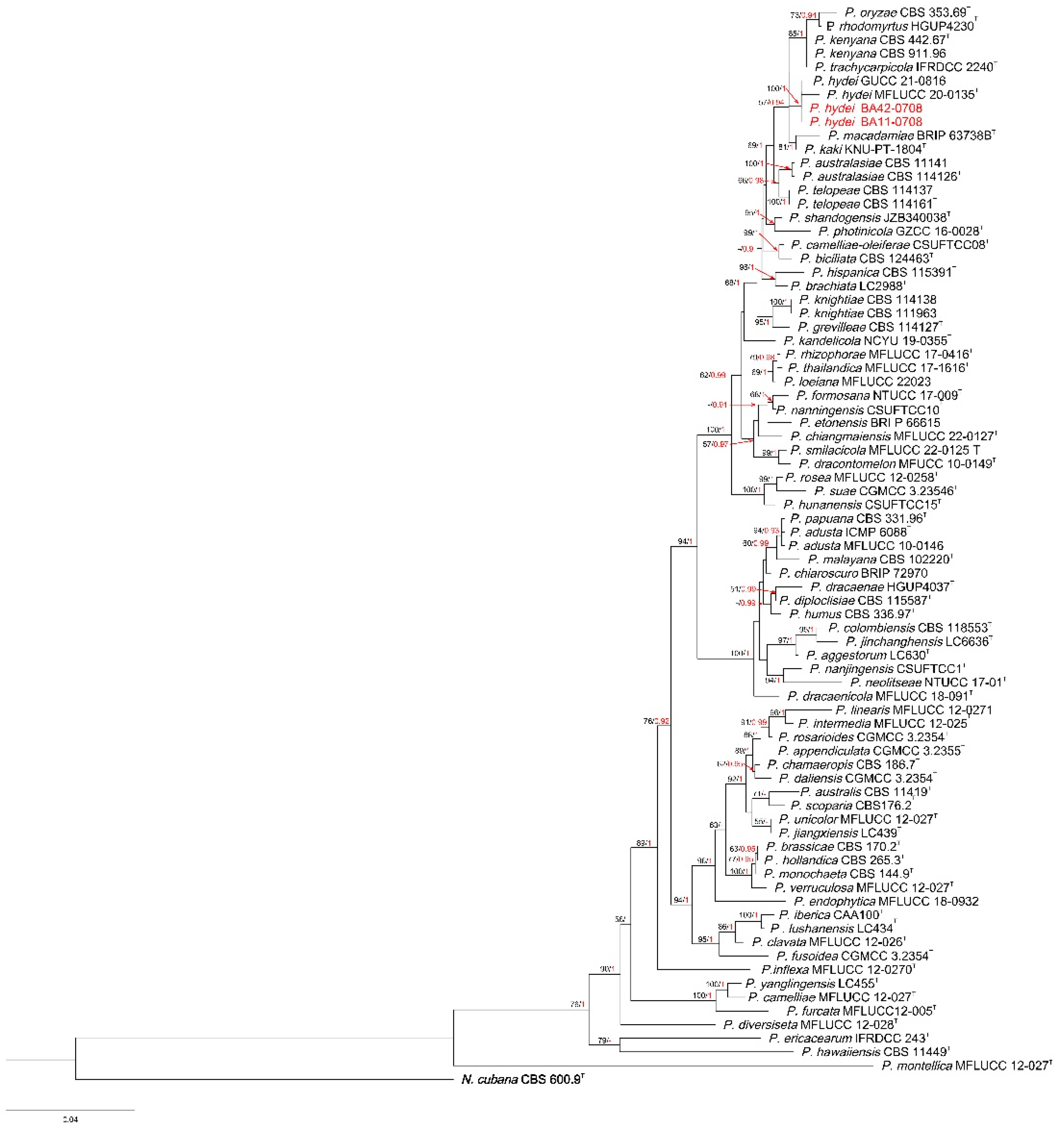
Figure 3.
RAxML tree of Pseudopestalotiopsis isolates based on ITS, tef-1α and tub2 sequences. The root of this tree is P. trachycaroicola OP068. The strains isolated in this study are marked in red. Ex-type strains are marked with T. ML bootstrap values ≥50% and BI probabilities (in red) ≥0.90 are displayed at the nodes.
Figure 3.
RAxML tree of Pseudopestalotiopsis isolates based on ITS, tef-1α and tub2 sequences. The root of this tree is P. trachycaroicola OP068. The strains isolated in this study are marked in red. Ex-type strains are marked with T. ML bootstrap values ≥50% and BI probabilities (in red) ≥0.90 are displayed at the nodes.

Figure 4.
Split graphs showing the result of PHI test of new Neopestalotiopsis species with their most closely related species. The new species in each graph is shown in red font.
Figure 4.
Split graphs showing the result of PHI test of new Neopestalotiopsis species with their most closely related species. The new species in each graph is shown in red font.
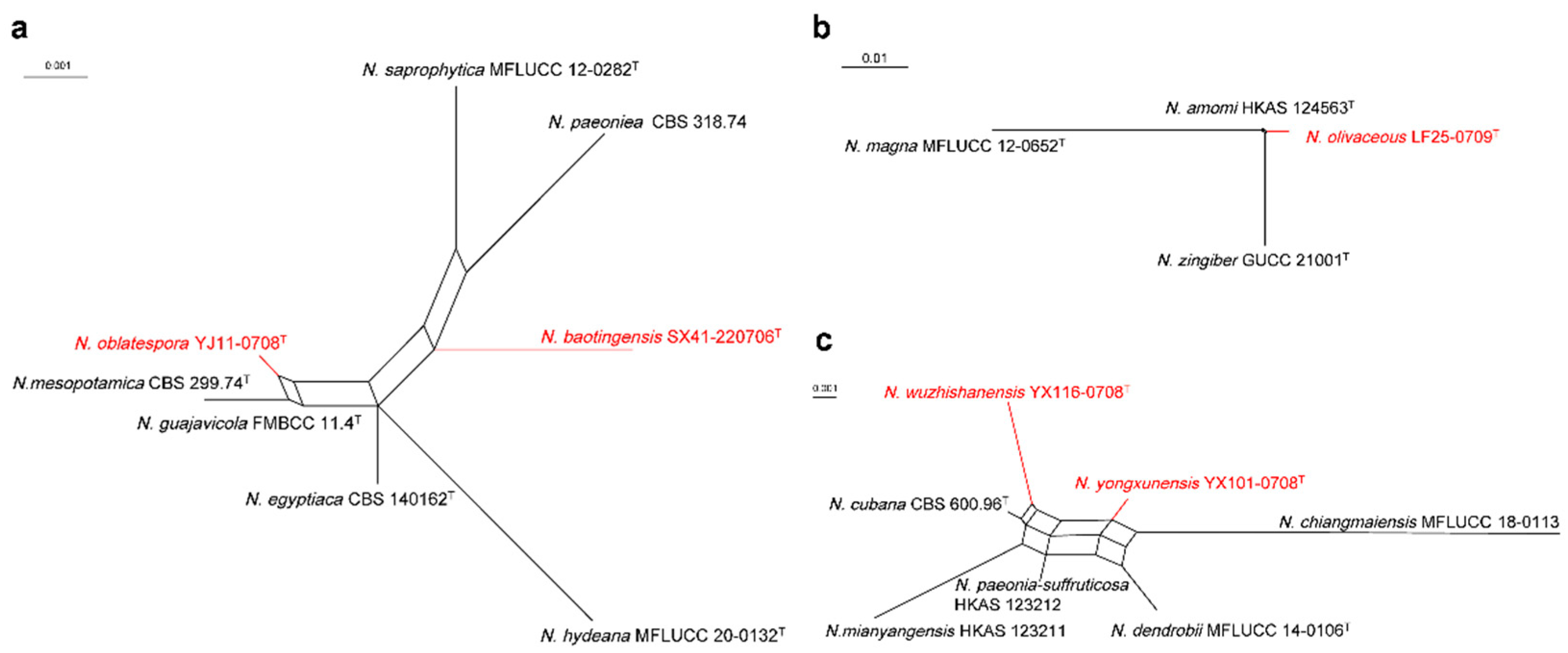
Figure 5.
Neopestalotiopsis baotingensis (SX41-0706, holotype). (a-b) Colony on PDA (above and reverse), (c-d) Conidiomata on PDA, (e-f) Conidiogenous cells, (g-l) Conidia. Scale bars = 10 μm.
Figure 5.
Neopestalotiopsis baotingensis (SX41-0706, holotype). (a-b) Colony on PDA (above and reverse), (c-d) Conidiomata on PDA, (e-f) Conidiogenous cells, (g-l) Conidia. Scale bars = 10 μm.
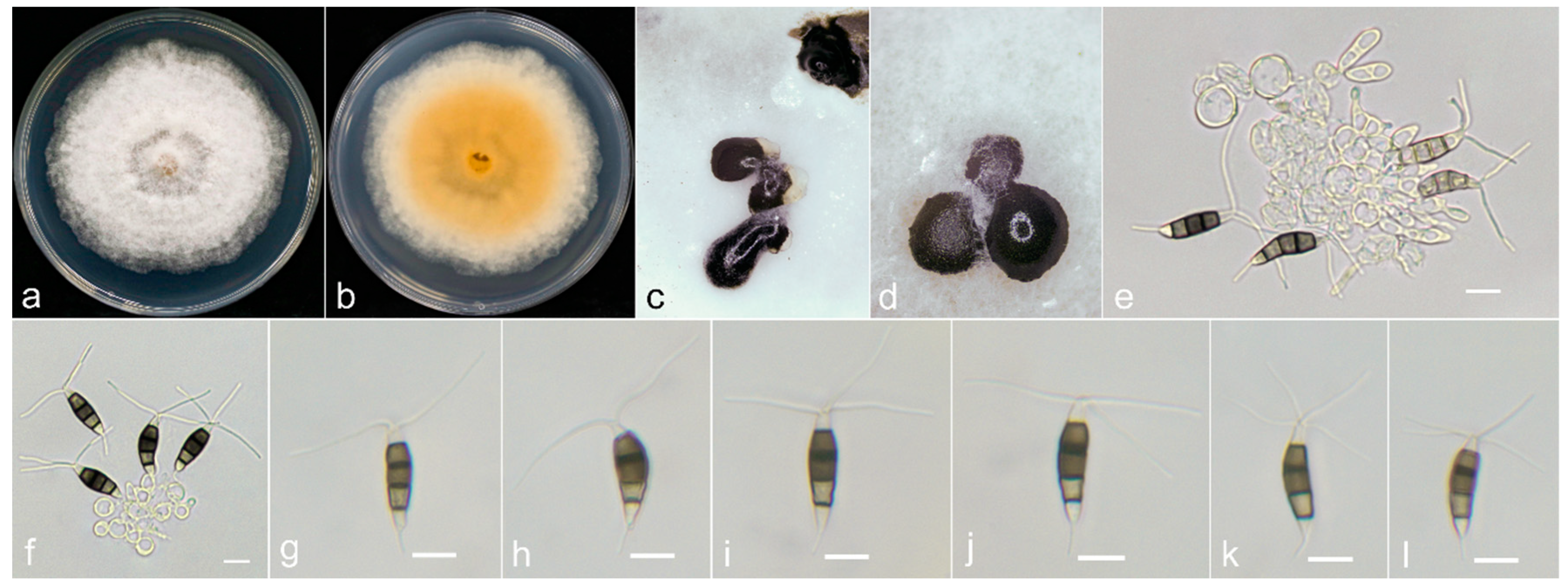
Figure 6.
Neopestalotiopsis oblatespora (YJ11-0708, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidia pile on PDA, (d-e) Conidiophores, (f-k) Conidia. Scale bars = 10 μm.
Figure 6.
Neopestalotiopsis oblatespora (YJ11-0708, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidia pile on PDA, (d-e) Conidiophores, (f-k) Conidia. Scale bars = 10 μm.

Figure 7.
Neopestalotiopsis olivaceous (LF25-0709, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidia pile on PDA, (d) Conidiophores, (e-l) Conidia. Scale bars = 10 μm.
Figure 7.
Neopestalotiopsis olivaceous (LF25-0709, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidia pile on PDA, (d) Conidiophores, (e-l) Conidia. Scale bars = 10 μm.
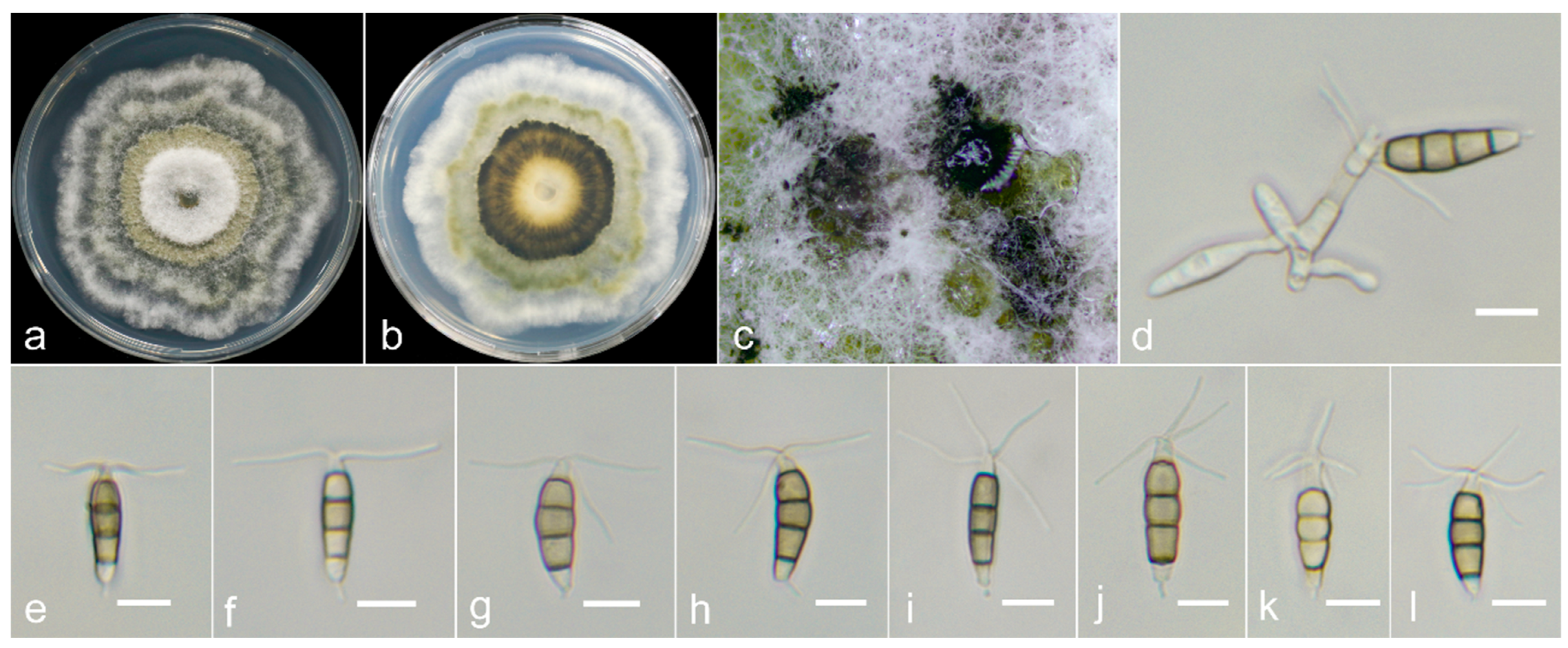
Figure 8.
Neopestalotiopsis oxyphylla (LF55-0709, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidiomata on PDA, (d-e) Conidiogenous cells, (f-k) Conidia. Scale bars = 10 μm.
Figure 8.
Neopestalotiopsis oxyphylla (LF55-0709, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidiomata on PDA, (d-e) Conidiogenous cells, (f-k) Conidia. Scale bars = 10 μm.
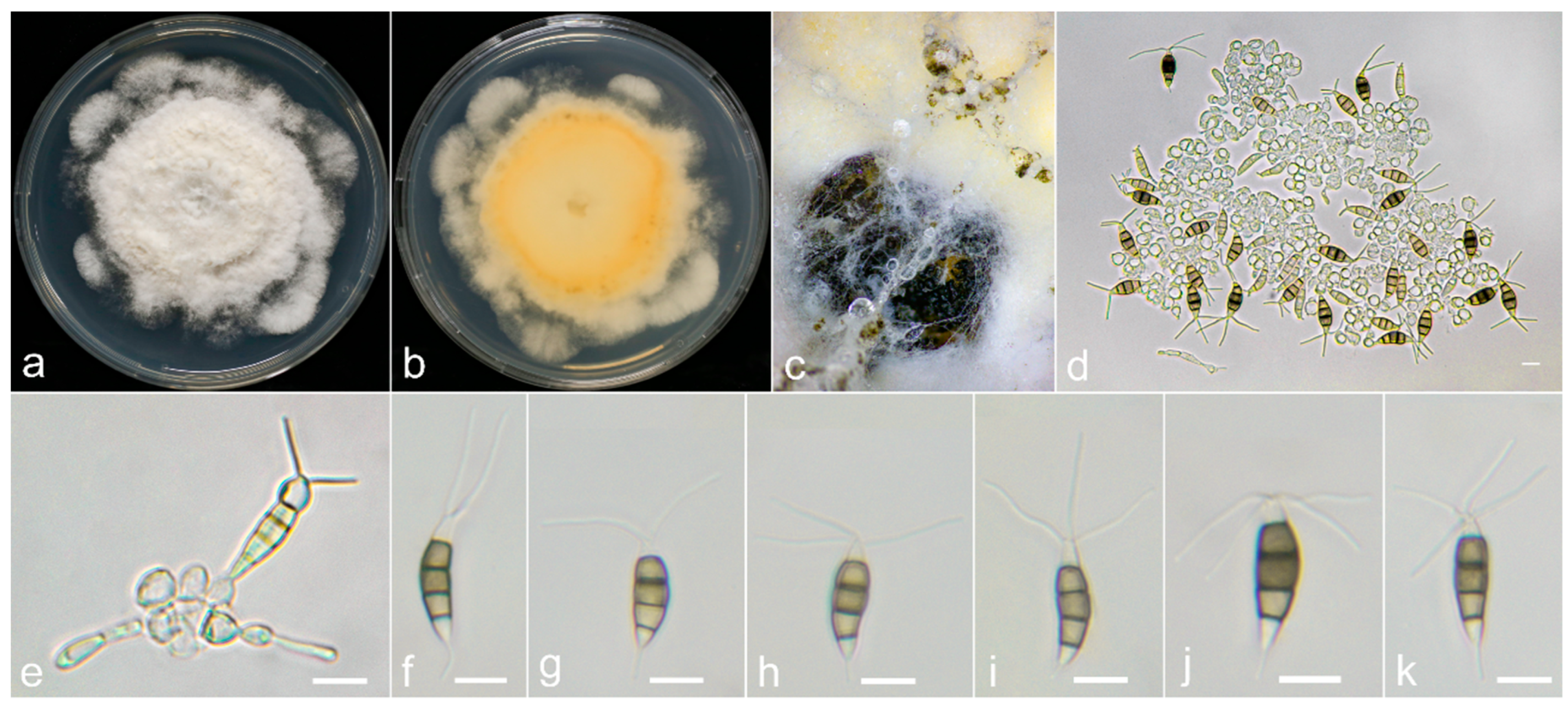
Figure 9.
Neopestalotiopsis wuzhishanensis (YX116-0708, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidiomata on PDA, (d-e) Conidiogenous cells, (f-k) Conidia. Scale bars = 10 μm.
Figure 9.
Neopestalotiopsis wuzhishanensis (YX116-0708, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidiomata on PDA, (d-e) Conidiogenous cells, (f-k) Conidia. Scale bars = 10 μm.
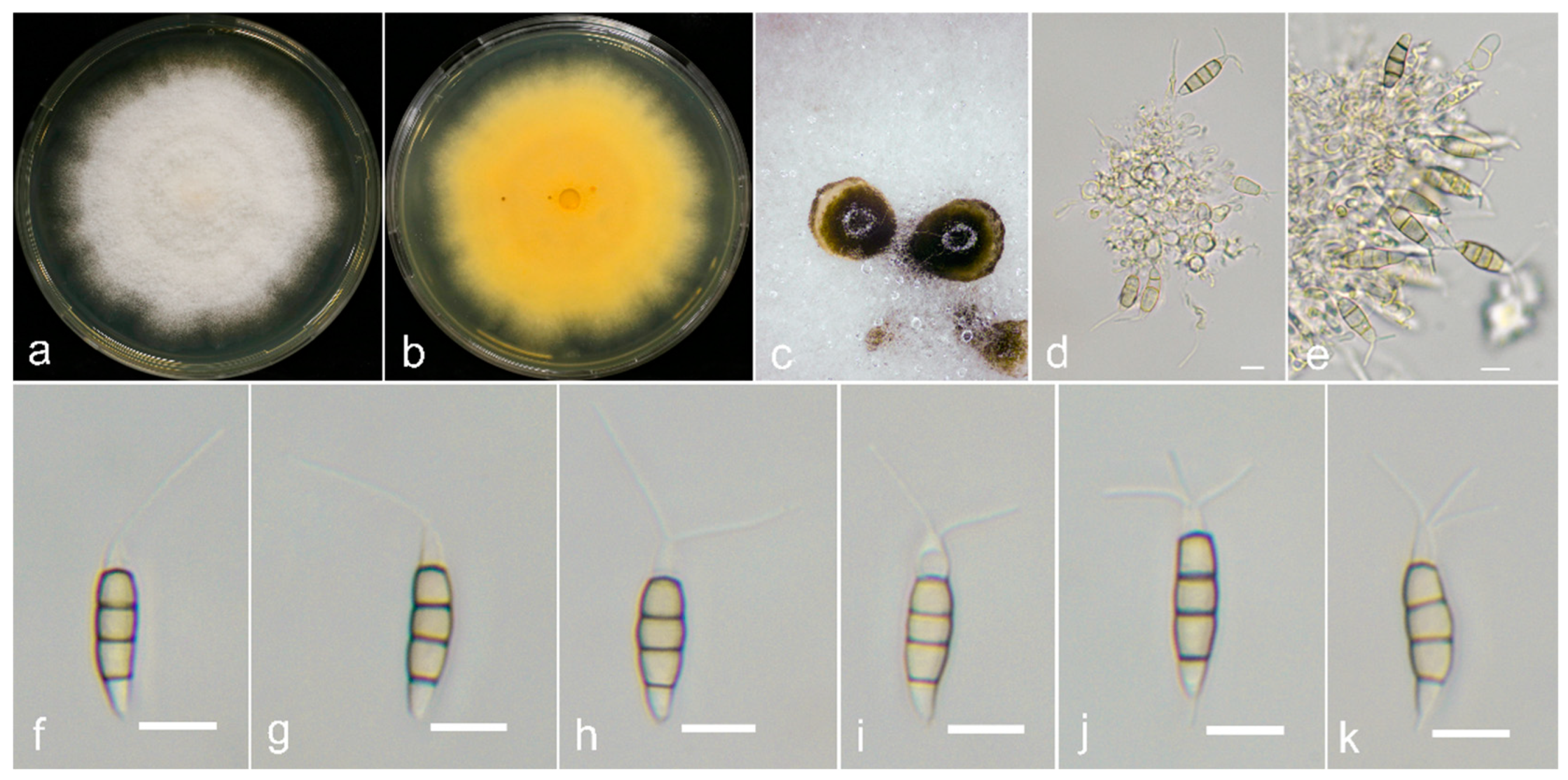
Figure 10.
Neopestalotiopsis yongxunensis (YX101-0708, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidiomata on PDA, (d-e) Conidiogenous cells, (f-k) Conidia. Scale bars = 10 μm.
Figure 10.
Neopestalotiopsis yongxunensis (YX101-0708, holotype). (a-b) Colony on PDA (above and reverse), (c) Conidiomata on PDA, (d-e) Conidiogenous cells, (f-k) Conidia. Scale bars = 10 μm.
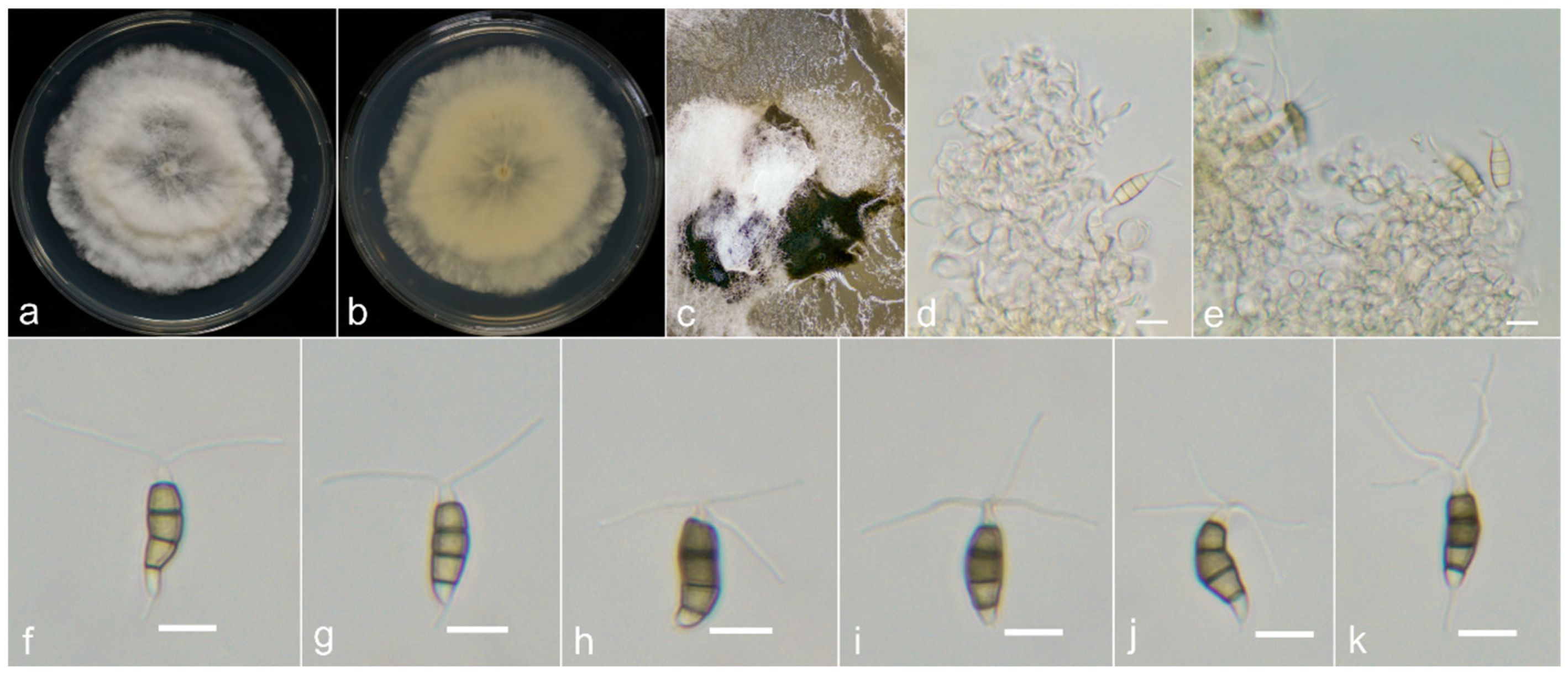
Figure 11.
Pathogenicity test results of 20 pestalotioid species on Alpinia oxyphylla leaves. (A) symptoms on leaves after 5 days. Icons in figures in sequence are CK, P. hydei BA11-0708, Ps. myanmarina JR34-0709, Ps. avicenniae LF48-0709, N. coffeae-arabicae BL32-0708, N. coffeae-arabicae LF51-0709, Neopestalotiopsis sp.4 MH133-0708, N. coffeae-arabicae NM42-0706, N. cubana MH51-0708, N. cubana YX112-0708, N. wuzhishanensis YX116-0708, N. yongxunensis YX101-0708, N. baotingensis SX41-0706, N. oblatespora YJ11-0708, N. vaccinii JR31-0709, N. rosicola NM47-0708, N. oxyphylla LF55-0709, N. brachiata, SX31-0706, Neopestalotiopsis sp.5 XC11-0709, N. olivaceous LF25-0709, Neopestalotiopsis sp.3 SX11-0706. (B) Pathogenicity of the isolates was evaluated by measuring area of the necrotic lesions after 5 days. Error bars indicate the standard deviation of the mean. Significant differences (p < 0.05) are indicated with different letters according to Duncan’s multiple range test. The abscissa designation corresponds sequentially to (A), excluding “CK”.
Figure 11.
Pathogenicity test results of 20 pestalotioid species on Alpinia oxyphylla leaves. (A) symptoms on leaves after 5 days. Icons in figures in sequence are CK, P. hydei BA11-0708, Ps. myanmarina JR34-0709, Ps. avicenniae LF48-0709, N. coffeae-arabicae BL32-0708, N. coffeae-arabicae LF51-0709, Neopestalotiopsis sp.4 MH133-0708, N. coffeae-arabicae NM42-0706, N. cubana MH51-0708, N. cubana YX112-0708, N. wuzhishanensis YX116-0708, N. yongxunensis YX101-0708, N. baotingensis SX41-0706, N. oblatespora YJ11-0708, N. vaccinii JR31-0709, N. rosicola NM47-0708, N. oxyphylla LF55-0709, N. brachiata, SX31-0706, Neopestalotiopsis sp.5 XC11-0709, N. olivaceous LF25-0709, Neopestalotiopsis sp.3 SX11-0706. (B) Pathogenicity of the isolates was evaluated by measuring area of the necrotic lesions after 5 days. Error bars indicate the standard deviation of the mean. Significant differences (p < 0.05) are indicated with different letters according to Duncan’s multiple range test. The abscissa designation corresponds sequentially to (A), excluding “CK”.
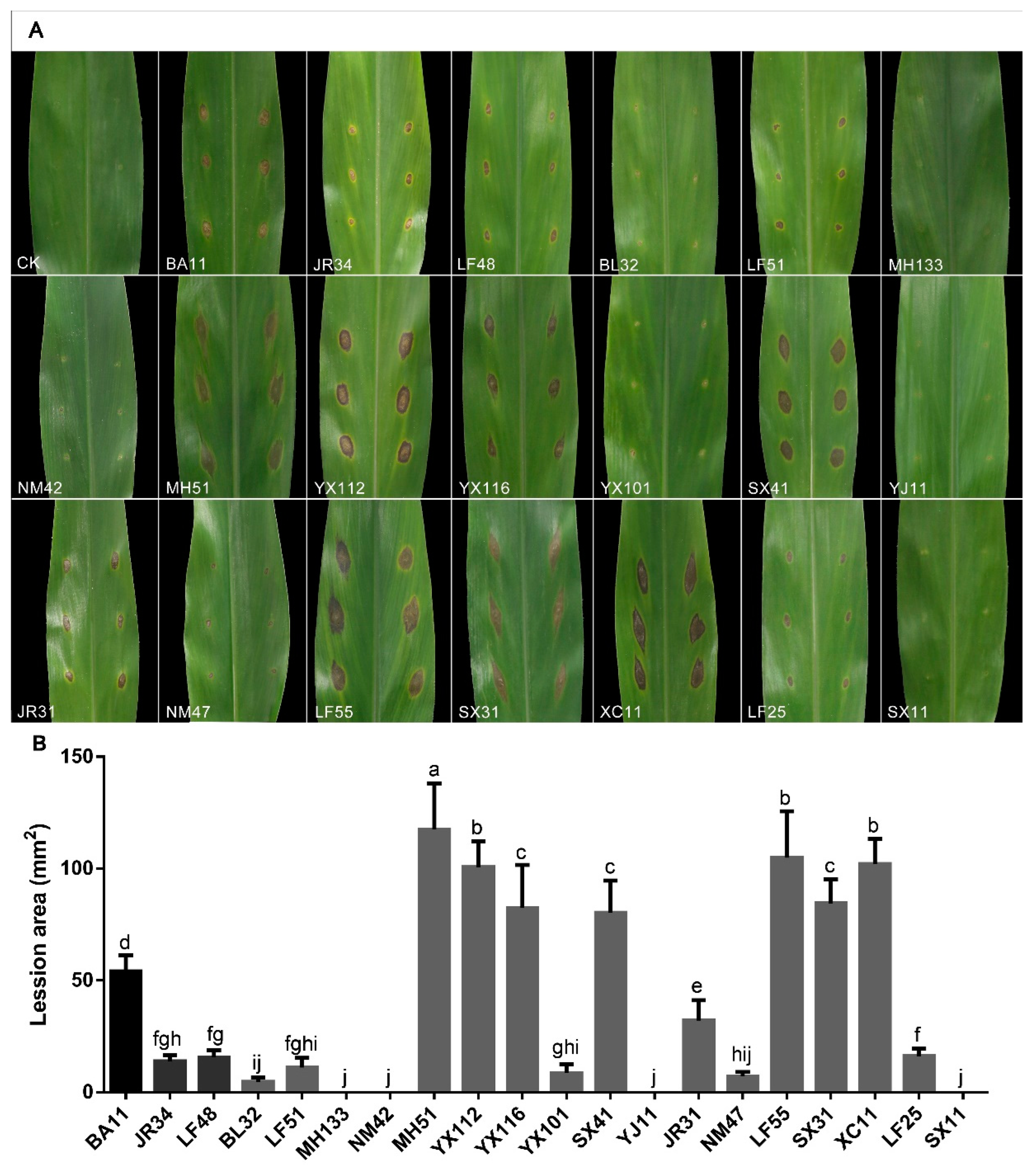

Table 1.
PCR primers and procedures used in this study.
| Locus | Primes Name | Sequence (5’to 3’) | PCR procedures | Reference |
|---|---|---|---|---|
| ITS | ITS5 | GGAAGTAAAAGTCGTAACAAGG | 95°C 5 min; 94°C 25 s; 52°C 25 s; 72°C 10 s; repeat 2 to 4 for 35 cycles; 72°C 5 min; 4°C on hold | [35] |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| tef-1α | EF1-728F | CATCGAGAAGTTCGAGAAGG | 95°C 5 min; 94°C 25 s; 52°C 25 s; 72 °C 10 s (15s); repeat 2 to 4 for 35 cycles; 72°C 5 min; 4°C on hold | [36,37] |
| EF1-526F | GTCGTYGTYATYGGHCAYGT | |||
| EF2 | GGARGTACCAGTSATCATGTT | |||
| tub2 | T1 | AACATGCGTGAGATTGTAAGT | 95°C 5 min; 94°C 25 s; 55°C 25 s; 72°C 15 s; repeat 2 to 4 for 35 cycles; 72°C 5 min; 4°C on hold | [38,39] |
| Bt2b | ACCCTCAGTGTAGTGACCCTTGGC |
Table 2.
The strains information and their genes’ accession numbers of pestalotioid species used in this study.
Table 2.
The strains information and their genes’ accession numbers of pestalotioid species used in this study.
| Taxonomic status | Strain No. | Host/Substrate | Origin | Gen Bank Accessions Numbers | References | ||
|---|---|---|---|---|---|---|---|
| ITS | tub 2 | tef-lα | |||||
| Neopestalotiopsis acrostichi | MFLUCC 17-1754T | Acrostichum aureum | Thailand | MK764272 | MK764338 | MK764316 | [41] |
| N. acrostichi | MFLUCC 17-1755 | Acrostichum aureum | Thailand | MK764273 | MK764339 | MK764317 | [41] |
| N. alpapicalis | MFLUCC 17-2544T | Rhizophora mucronata | Thailand | MK357772 | MK463545 | MK463547 | [42] |
| N. alpapicalis | MFLUCC 17-2545 | Rhizophora mucronata | Thailand | MK357773 | MK463546 | MK463548 | [42] |
| N. amomi | HKAS 124563T | Amomum villosum | China | OP498012 | OP752133 | OP653489 | [16] |
| N. amomi | HKAS 124564 | Amomum villosum | China | OP498013 | OP765913 | OP753382 | [16] |
| N. aotearoa | CBS 367.54T | Canvas | New Zealand | KM199369 | KM199454 | KM199526 | [14] |
| N. asiatica | MFLUCC 12-0286T | Prunus dulcis | China | JX398983 | JX399018 | JX399049 | [15] |
| N. australis | CBS 114159T | Telopea sp. | Australia | KM199348 | KM199432 | KM199537 | [15] |
| N. brachiata | MFLUCC 17-1555 | Rhizophora apiculata | Thailand | MK764274 | MK764340 | MK764318 | [41] |
| N. brasiliensis | COAD 2166T | Psidium guajava | Brazil | MG686469 | MG692400 | MG692402 | [43] |
| N. brasiliensis | CFCC 54341 | Castanea mollissima | China | MW166229 | MW218522 | MW199748 | [44] |
| N. camelliae-oleiferae | CSUFTCC81T | Camellia oleifera | China | OK493585 | OK562360 | OK507955 | [17] |
| N. camelliae-oleiferae | CSUFTCC82 | Camellia oleifera | China | OK493586 | OK562361 | OK507956 | [17] |
| N. cavernicola | KUMCC 20-0269T | Cave | China | MW545802 | MW557596 | MW550735 | [45] |
| N. chiangmaiensis | MFLUCC 18–0113 | Pandanus sp. | Thailand | NA | MH412725 | MH388404 | [46] |
| N. chrysea | MFLUCC 12-0261T | Dead leaves | China | JX398985 | JX399020 | JX399051 | [14] |
| N. chrysea | MFLUCC 12-0262 | Dead leaves | China | JX398986 | JX399021 | JX399052 | [14] |
| N. clavispora | MFLUCC 12-0281T | Magnolia sp. | China | JX398979 | JX399014 | JX399045 | [14] |
| N. clavispora | MFLUCC 12-0280 | Magnolia sp. | China | JX398978 | JX399013 | JX399044 | [14] |
| N. cocoes | MFLUCC 15-0152T | Cocos nucifera | Thailand | KX789687 | NA | KX789689 | [41] |
| N. coffeae-arabicae | HGUP4015 | Coffea arabica | China | KF412647 | KF412641 | KF412644 | [47] |
| N. coffeae-arabicae | HGUP4019T | Coffea arabica | China | KF412649 | KF412643 | KF412646 | [47] |
| N. concentrica | CFCC 55162T | Rosa chinensis | China | OK560707 | OM117698 | OM622433 | [48] |
| N. cubana | CBS 600.96T | Leaf litter | Cuba | KM199347 | KM199438 | KM199521 | [15] |
| N. dendrobii | MFLUCC 14-0106T | Dendrobium cariniferum | Thailand | MK993571 | MK975835 | MK975829 | [49] |
| N. dendrobii | MFLUCC 14-0099 | Dendrobium cariniferum | Thailand | MK993570 | MK975834 | MK975828 | [49] |
| N. drenthii | BRIP 72263a | Macadamia integrifolia | Australia | MZ303786 | MZ312679 | MZ344171 | [20] |
| N. drenthii | BRIP 72264aT | Macadamia integrifolia | Australia | MZ303787 | MZ312680 | MZ344172 | [20] |
| N. egyptiaca | CBS 140162T | Mangifera indica | Egypt | KP943747 | KP943746 | KP943748 | [50] |
| N. elaeagni | HGUP10002T | Elaeagnus pungens | China | MW930716 | MZ683391 | MZ203452 | [30] |
| N. elaeidis | MFLUCC 15-0735 | Elaeis guineensis | Thailand | ON650689 | NA | ON734012 | [51] |
| N. ellipsospora | MFLUCC 12-0283T | Dead plant | China | JX398980 | JX399016 | JX399047 | [14] |
| N. eucalyptorum | CBS 147684T | Eucalyptus globulus | Portugal | MW794108 | MW802841 | MW805397 | [18] |
| N. eucalypticola | CBS 264.37T | Eucalyptus globulus | NA | KM199376 | KM199431 | KM199551 | [15] |
| N. fragariae | ZHKUCC 22-0115 | Fragaria x ananassa | China | ON651146 | ON685199 | ON685197 | [32] |
| N. foedans | CGMCC 3.9123T | Mangrove plant | China | JX398987 | JX399022 | JX399053 | [14] |
| N. foedans | CGMCC 3.9178 | Neodypsis decaryi | China | JX398989 | JX399024 | JX399055 | [14] |
| N. formicarum | CBS 362.72T | Dead ant | Cuba | KM199358 | KM199455 | KM199517 | [15] |
| N. formicarum | CBS 115.83 | Plant debris | Cuba | KM199344 | KM199444 | KM199519 | [15] |
| N. guajavae | FMBCC 11.1T | Psidium guajava | Pakistan | MF783085 | MH460871 | MH460868 | [52] |
| N. guajavicola | FMBCC 11.4T | Psidium guajava | Pakistan | MH209245 | MH460873 | MH460870 | [52] |
| N. haikouensis | SAUCC212271T | Ilexchinensis sp. | China | OK087294 | OK104870 | OK104877 | [53] |
| N. hadrolaeliae | COAD 2637T | Hadrolaelia jongheana | Brazil | MK454709 | MK465120 | MK465122 | [54] |
| N. hispanica | CBS 147686T | Eucalyptus globulus | Portugal | MW794107 | MW802840 | MW805399 | [18] |
| N. honoluluana | CBS 114495T | Telopea sp. | USA | KM199364 | KM199457 | KM199548 | [15] |
| N. hydeana | MFLUCC 20-0132T | Artocarpus heterophyllus | Thailand | MW266069 | MW251119 | MW251129 | [55] |
| N. hyperici | HKAS 124561 | Hypericum monogynum | China | OP498010 | OP765908 | OP713768 | [16] |
| N. iberica | CBS 147688T | Eucalyptus globulus | Portugal | MW794111 | MW802844 | MW805402 | [18] |
| N. javaensis | CBS 257.31T | Cocos nucifera | Indonesia | KM199357 | KM199437 | KM199543 | [15] |
| N. lusitanica | CBS 147690T | Eucalyptus globulus | Portugal | MW794110 | MW802843 | MW805406 | [18] |
| N. longiappendiculata | CBS 147692T | Eucalyptus globulus | Portugal | MW794112 | MW802845 | MW805404 | [18] |
| N. macadamiae | BRIP 63737cT | Macadamia integrifolia | Australia | KX186604 | KX186654 | KX186627 | [56] |
| N. macadamiae | BRIP 63742a | Macadamia integrifolia | Australia | KX186599 | KX186657 | KX186629 | [56] |
| N. maddoxii | BRIP 72266aT | Macadamia integrifolia | Australia | MZ303782 | MZ312675 | MZ344167 | [56] |
| N. magna | MFLUCC 12-0652T | Pteridium sp. | France | KF582795 | KF582793 | KF582791 | [57] |
| N. mesopotamica | CBS 336.86T | Pinus brutia | Iraq | KM199362 | KM199441 | KM199555 | [15] |
| N. mesopotamica | CBS 299.74 | Eucalyptus sp. | Turkey | KM199361 | KM199435 | KM199541 | [15] |
| N. mianyangensis | HKAS 123211 | Paeonia suffruticosa | China | OP546681 | OP672161 | OP723490 | [31] |
| N. musae | MFLUCC 15-0776T | Musa sp. | Thailand | KX789683 | KX789686 | KX789685 | [41] |
| N. natalensis | CBS 138.41T | Acacia mollissima | South Africa | KM199377 | KM199466 | KM199552 | [15] |
| N. nebuloides | BRIP 66617T | Sporobolus elongatus | Australia | MK966338 | MK977632 | MK977633 | [58] |
| N. olumideae | BRIP 72273aT | Macadamia integrifolia | Australia | MZ303790 | MZ312683 | MZ344175 | [20] |
| N. paeoniea | CBS 318.74 | Anacardium occidentale | Nigeria | MH554031 | MH554707 | NA | [59] |
| N. paeonia-suffruticosa | HKAS 123212T | Paeonia suffruticosa | China | OP082292 | OP235980 | OP204794 | [31] |
| N. pernambucana | URM 7148-01T | Vismia guianensis | Brazil | KJ792466 | NA | KU306739 | [60] |
| N. perukae | FMBCC11.3T | Guava | Pakistan | MH209077 | MH460876 | MH523647 | [52] |
| N. petila | MFLUCC 17-1737 | Rhizophora mucronata | Thailand | MK764275 | MK764341 | MK764319 | [41] |
| N. petila | MFLUCC 17-1738T | Rhizophora mucronata | Thailand | MK764276 | MK764342 | MK764320 | [41] |
| N. phangngaensis | MFLUCC 18-0119T | Pandanus sp. | Thailand | MH388354 | MH412721 | MH388390 | [46] |
| N. piceana | CBS 254.32 | Cocos nucifera | Indonesia | KM199372 | KM199452 | KM199529 | [15] |
| N. piceana | CBS 394.48T | Picea sp. | UK | KM199368 | KM199453 | KM199527 | [15] |
| N. photiniae | MFLUCC 22-0129T | Photinia serratifolia | China | OP498008 | OP752131 | OP753368 | [16] |
| N. protearum | CBS 114178T | Leucospermum cuneiforme | Zimbabwe | JN712498 | KM199463 | LT853201 | [61] |
| N. psidii | FMBCC 11.2T | Psidium guajava | Pakistan | MF783082 | MH477870 | MH460874 | [52] |
| N. rhapidis | GUCC 21501T | Rhododendron simsii | China | MW931620 | MW980441 | MW980442 | [34] |
| N. rhizophorae | MFLUCC 17-1551T | Rhizophora mucronata | Thailand | MK764277 | MK764343 | MK764321 | [41] |
| N. rhizophorae | MFLUCC 17 1550 | Rhizophora mucronata | Thailand | MK764278 | MK764344 | MK764322 | [41] |
| N. rhododendri | GUCC 21504T | Rhododendron simsii | China | MW979577 | MW980443 | MW980444 | [34] |
| N. rhododendricola | KUN-HKAS-123204T | Rhododendron sp. | China | OK283069 | OK274147 | OK274148 | [62] |
| N. rosae | CBS 101057T | Rosa sp. | New Zealand | KM199359 | KM199429 | KM199523 | [15] |
| N. rosicola | CFCC 51992T | Rosa chinensis | China | KY885239 | KY885245 | KY885243 | [63] |
| N. rosicola | CFCC 51993 | Rosa chinensis | China | KY885240 | KY885246 | KY885244 | [63] |
| N. samarangensis | CBS 115451 | Unidentified Tree | China | KM199365 | KM199447 | KM199556 | [15] |
| N. saprophytica | MFLUCC 12-0282T | Magnolia sp. | China | JX398982 | JX399017 | JX399048 | [15] |
| N. scalabiensis | CAA 1029T | Vaccinium corymbosum | Portugal | MW969748 | MW934611 | MW959100 | [64] |
| N. sichuanensis | CFCC 54338T | Castanea mollissima | China | MW166231 | MW218524 | MW199750 | [44] |
| N. siciliana | AC46 | Persea americana | Italy | ON117813 | ON209162 | ON107273 | [65] |
| N. sonneratae | MFLUCC 17-1744 | Sonneronata alba | Thailand | MK764279 | MK764345 | MK764323 | [41] |
| N. sonneratae | MFLUCC 17-1745T | Sonneronata alba | Thailand | MK764280 | MK764346 | MK764324 | [41] |
| N. steyaertii | IMI 192475T | Eucalyptus viminalis | Australia | KF582796 | KF582794 | KF582792 | [15] |
| N. surinamensis | CBS 450.74T | Soil under Elaeis guineensis | Suriname | KM199351 | KM199465 | KM199518 | [15] |
| N. subepidermalis | CFCC 55160 | Rosa chinensis | China | OK560699 | OM117690 | OM622425 | [48] |
| N. suphanburiensis | MFLUCC 22-0126T | Unknown | Thailand | OP497994 | OP752135 | OP753372 | [16] |
| N. terricola | HKAS 123213 | Paeonia suffruticosa | China | OP082294 | OP235982 | OP204796 | [31] |
| N. thailandica | MFLUCC 17-1730T | Rhizophora mucronata | Thailand | MK764281 | MK764347 | MK764325 | [41] |
| N. thailandica | MFLUCC 17-1731 | Rhizophora mucronata | Thailand | MK764282 | MK764348 | MK764326 | [41] |
| N. umbrinospora | MFLUCC 12-0285T | Unidentified plant | China | JX398984 | JX399019 | JX399050 | [14] |
| N. vaccinii | CAA 1059T | Vaccinium corymbosum | Portugal | MW969747 | MW934610 | MW959099 | [64] |
| N. vacciniicola | CAA 1055T | Vaccinium corymbosum | Portugal | MW969751 | MW934614 | MW959103 | [64] |
| N. vheenae | BRIP 72293aT | Macadamia integrifolia | Australia | MZ303792 | MZ312685 | MZ344177 | [20] |
| N. vitis | MFLUCC 15-1265T | Vitis vinifera cv. “Summer black” | China | KU140694 | KU140685 | KU140676 | [66] |
| N. vitis | MFLUCC 15-1270 | Vitis vinifera cv. “Kyoho” | China | KU140699 | KU140690 | KU140681 | [66] |
| N. xishuangbannaensis | KUMCC 21-0424T | Kerivoula hardwickii (Bat) | China | ON426865 | OR025934 | OR025973 | [67] |
| N. xishuangbannaensis | KUMCC 21-0425 | Kerivoula hardwickii (Bat) | China | ON426866 | OR025935 | OR025974 | [67] |
| N. zakeelii | BRIP 72282aT | Macadamia integrifolia | Australia | MZ303789 | MZ312682 | MZ344174 | [20] |
| N. zimbabwana | CBS 111495T | Leucospermum cunciforme | Zimbabwe | NA | KM199456 | KM199545 | [15] |
| N. zingiber | GUCC 21001T | Zingiber officinale | China | MW930715 | MZ683390 | MZ683389 | [34] |
| Neopestalotiopsis sp.2 | CFCC 54340 | Castanea mollissima | China | MW166235 | MW218528 | MW199754 | [44] |
| Neopestalotiopsis sp.2 | ZX22B | Castanea mollissima | China | MW166236 | MW218529 | MW199755 | [44] |
| Neopestalotiopsis sp. nov. | GUCC 210003 | Unknown | China | MW930717 | MZ683392 | MZ540914 | [34] |
| Neopestalotiopsis sp.1 | CFCC 54337 | Castanea mollissima | China | MW166233 | MW218526 | MW199752 | [44] |
| Neopestalotiopsis sp.1 | ZX121 | Castanea mollissima | China | MW166234 | MW218527 | MW199753 | [44] |
| Pestalotiopsis adusta | ICMP 6088T | Refrigerator door | Fiji | JX399006 | JX399037 | JX399070 | [14] |
| P. adusta | MFLUCC 10-0146 | Syzygium sp. | Thailand | JX399007 | JX399038 | JX399071 | [14] |
| P. aggestorum | LC6301T | Camellia sinensis | China | KX895015 | KX895348 | KX895234 | [68] |
| P. appendiculata | CGMCC 3.23550T | Rhododendron sp. | China | OP082431 | OP185516 | OP185509 | [69] |
| P. australasiae | CBS 114126T | Knightia sp. | New Zealand | KM199297 | KM199409 | KM199499 | [15] |
| P. australasiae | CBS 11141 | Protea sp. | New Sout Wales | KM199298 | KM199410 | KM199501 | [15] |
| P. australis | CBS 114193T | Grevillea sp. | New South Wales | KM199332 | KM199383 | KM199475 | [15] |
| P. biciliata | CBS 124463T | Platanus×hispanica | Slovakia | KM199308 | KM199399 | KM199505 | [15] |
| P. brachiata | LC2988T | Camellia sp. | China | KX894933 | KX895265 | KX895150 | [14] |
| P. brassicae | CBS 170.26T | Brassica napus | New Zealand | KM199379 | NA | KM199558 | [15] |
| P. camelliae | MFLUCC 12-0277T | Camellia japonica | China | JX399010 | JX399041 | JX399074 | [14] |
| P. camelliae-oleiferae | CSUFTCC08T | Camellia oleifera | China | OK493593 | OK562368 | OK507963 | [17] |
| P. chamaeropis | CBS 186.71T | Chamaerops humilis | Italy | KM199326 | KM199391 | KM199473 | [14] |
| P. chiangmaiensis | MFLUCC 22-0127T | Phyllostachys edulis | Thailand | OP497990 | OP752137 | OP753374 | [16] |
| P. chiaroscuro | BRIP 72970 | Sporobolus natalensis | Australia | OK422510 | OK423752 | OK423753 | [70] |
| P. clavata | MFLUCC 12-0268T | Buxus sp. | China | JX398990 | JX399025 | JX399056 | [14] |
| P. colombiensis | CBS 118553T | Eucalyptus eurograndis | Colombia | KM199307 | KM199421 | KM199488 | [15] |
| P. daliensis | CGMCC 3.23548T | Rhododendron sp. | China | OP082429 | OP185518 | OP185511 | [69] |
| P. diploclisiae | CBS 115587T | Diploclisia glaucescens | China | KM199320 | KM199419 | KM199486 | [15] |
| P. diversiseta | MFLUCC 12-0287T | Rhododendron sp. | China | NR 120187 | JX399040 | JX399073 | [14] |
| P. dracaenae | HGUP4037T | Dracaena fragrans | China | NA | MT598645 | MT598644 | [71] |
| P. dracaenicola | MFLUCC 18-0913T | Dracaena sp. | Thailand | MN962731 | MN962733 | MN962732 | [72] |
| P. dracontomelon | MFUCC 10-0149T | Dracontomelon dao | Thailand | KP781877 | NA | KP781880 | [73] |
| P. endophytica | MFLUCC 18-0932 | Endophytic on healthy leaves of Magnolia candoll | Thailand | NR 172439 | NA | MW417119 | [74] |
| P. ericacearum | IFRDCC 2439T | Rhododendron delavayi | China | KC537807 | KC537821 | KC537814 | [75] |
| P. etonensis | BRI P 66615 | Sporobolus jacquemontii | Australia | MK966339 | MK977634 | MK977635 | [58] |
| P. formosana | NTUCC 17-009T | dead grass | China | MH809381 | MH809385 | MH809389 | [63] |
| P. furcata | MFLUCC 12-0054T | Camellia sinensis | Thailand | JQ683724 | JQ683708 | JQ683740 | [76] |
| P. fusoidea | CGMCC 3.23545T | Endophytic in fresh Rhododendron delavayi leaves. | China | OP082427 | OP185519 | OP185512 | [69] |
| P. grevilleae | CBS 114127T | Grevillea sp. | Australia | KM199300 | KM199407 | KM199504 | [15] |
| P. hawaiiensis | CBS 114491T | Leucospermum sp. | Hawaii | KM199339 | KM199428 | KM199514 | [15] |
| P. hispanica | CBS 115391T | Protea ‘Susara’ | Spain | MH553981 | MH554640 | MH554399 | [59] |
| P. hydei | MFLUCC 20-0135T | Litsea Petiolata | Thailand | MW266063 | MW251112 | MW251113 | [55] |
| P. hydei | GUCC 21-0816 | dead twigs | China | OP753660 | OP765909 | OP753383 | [16] |
| P. hollandica | CBS 265.33T | Sciadopitys verticillata | Netherlands | KM199328 | KM199388 | KM199481 | [15] |
| P. humus | CBS 336.97T | Soil | Papua New Guinea | KM199317 | KM199420 | KM199484 | [15] |
| P. hunanensis | CSUFTCC15T | Camellia oleifera | China | OK493599 | OK562374 | OK507969 | [17] |
| P. iberica | CAA1006T | Pinus radiata | Spain | MW732249 | MW759036 | MW759039 | [77] |
| P. inflexa | MFLUCC 12-0270T | Unidentified tree | China | JX399008 | JX399039 | JX399072 | [14] |
| P. intermedia | MFLUCC 12-0259T | Unidentified tree | China | JX398993 | JX399028 | JX399059 | [14] |
| P. jiangxiensis | LC4399T | Eurya sp. | China | KX895009 | KX895341 | KX895227 | [68] |
| P. jinchanghensis | LC6636T | Camellia sinensis | China | KX895028 | KX895361 | KX895247 | [68] |
| P. kandelicola | NCYU 19-0355T | Kandelia candel | China | MT560722 | MT563099 | MT563101 | [78] |
| P. kaki | KNU-PT-1804T | Diospyros kaki | Korea | LC552953 | LC552954 | LC553555 | [79] |
| P. kenyana | CBS 442.67T | Coffea sp. | Kenya | KM199302 | KM199395 | KM199502 | [15] |
| P. kenyana | CBS 911.96 | raw material from agar-agar | NA | KM199303 | KM199396 | KM199503 | [15] |
| P. knightiae | CBS 114138T | Knightia sp. | New Zealand | KM199310 | KM199408 | KM199497 | [15] |
| P. knightiae | CBS 111963 | Knightia sp. | New Zealand | KM199311 | KM199406 | KM199495 | [15] |
| P. linearis | MFLUCC 12-0271 | Trachelospermum sp. | China | JX398992 | JX399027 | JX399058 | [14] |
| P. loeiana | MFLUCC 22-0123 | dead leaves | Thailand | OP497988 | OP713769 | OP737881 | [16] |
| P. lushanensis | LC4344T | Camellia sp. | China | KX895005 | KX895337 | KX895223 | [68] |
| P. macadamiae | BRIP 63738BT | Macadamia integrifolia | Australia | KX186588 | KX186680 | KX186621 | [56] |
| P. malayana | CBS 102220T | Macaranga triloba | Malaysia | KM199306 | KM199411 | KM199482 | [15] |
| P. monochaeta | CBS 144.97T | Quercus robur | Netherlands | KM199327 | KM199386 | KM199479 | [15] |
| P. jesteri | MFLUCC 12-0279T | Fagraea bodenii | China | JX399012 | JX399043 | JX399076 | [14] |
| P. nanjingensis | CSUFTCC16T | Camellia oleifera | China | OK493602 | OK562377 | OK507972 | [17] |
| P. nanningensis | CSUFTCC10T | Camellia oleifera | China | OK493596 | OK562371 | OK507966 | [17] |
| P. neolitseae | NTUCC 17-011T | Neolitsea villosa | Taiwan | MH809383 | MH809387 | MH809391 | [63] |
| P. oryzae | CBS 353.69T | Oryza sativa | Denmark | KM199299 | KM199398 | KM199496 | [15] |
| P. papuana | CBS 331.96T | Coastal soil | Papua New Guinea | KM199321 | KM199413 | KM199491 | [15] |
| P. photinicola | GZCC 16-0028T | Photinia serrulata | China | KY092404 | KY047663 | KY047662 | [80] |
| P. rhizophorae | MFLUCC 17-0416T | Rhizophora apiculata | Thailand | MK764283 | MK764349 | MK764327 | [41] |
| P. rhodomyrtus | HGUP4230T | Rhodomyrtus tomentosa | China | KF412648 | KF412642 | KF412645 | [47] |
| P. rosarioides | CGMCC 3.23549T | Rhododendron decorum | China | OP082430 | OP185520 | OP185513 | [69] |
| P. rosea | MFLUCC 12-0258T | Pinus sp. | China | JX399005 | JX399036 | JX399069 | [14] |
| P. scoparia | CBS176.25T | Chamaecyparis sp. | NA | KM199330 | KM199393 | KM199478 | [15] |
| P. shandogensis | JZB340038T | unknow | China | MN625275 | MN626729 | MN626740 | [81] |
| P. smilacicola | MFLUCC 22-0125T | Smilax sp. | Thailand | OP497991 | OP762673 | OP753376 | [16] |
| P. suae | CGMCC 3.23546T | Rhododendron delavayi | China | OP082428 | OP185521 | OP185514 | [69] |
| P. telopeae | CBS 114161T | Telopea sp. | Australia | KM199296 | KM199403 | KM199500 | [15] |
| P. telopeae | CBS 114137 | Protea sp. | Australia | KM199301 | KM199469 | KM199559 | [15] |
| P. thailandica | MFLUCC 17-1616T | Rhizophora apiculata | Thailand | MK764285 | MK764351 | MK764329 | [41] |
| P. trachycarpicola | OP068T | Trachycarpus fortunei | China | JQ845947 | JQ845945 | JQ845946 | [82] |
| P. unicolor | MFLUCC 12-0276T | Rhododendron sp. | China | JX398999 | JX399030 | NA | [14] |
| P. verruculosa | MFLUCC 12-0274T | Rhododendron sp. | China | JX398996 | NA | JX399061 | [14] |
| P. yanglingensis | LC4553T | Camellia sinensis | China | KX895012 | KX895345 | KX895231 | [83] |
| Pseudopestalotiopsis ampullacea | LC6618T | Camellia sinensis | China | KX895025 | KX895358 | KX895244 | [68] |
| Ps. annellata | NTUCC 17-030T | Camellia sinensis | China, Taiwan | MT322087 | MT321889 | MT321988 | [21] |
| Ps. avicenniae | MFLUCC 17-0434T | Avicennia marina | Thailand | MK764287 | MK764353 | MK764331 | [41] |
| Ps. camelliae | CGMCC 3.9192 | Camellia sinensis | China | NA | KU562851 | KU562850 | [84] |
| Ps. camelliae-sinensis | NTUCC 18-031 | Camellia sinensis | China, Taiwan | MT322047 | MT321849 | MT321948 | [21] |
| Ps. camelliae-sinensis | LC3490T | Camellia sinensis | China | KX894985 | KX895316 | KX895202 | [68] |
| Ps. chinensis | NTUCC 18-066 | Camellia sinensis | China, Taiwan | MT322083 | MT321885 | MT321984 | [21] |
| Ps. chinensis | LC3011T | Camellia sinensis | China | KX894937 | KX895269 | KX895154 | [68] |
| Ps. chinensis | NTUCC 18-038 | Camellia sinensis | China, Taiwan | MT322055 | MT321857 | MT321956 | [21] |
| Ps. cocos | CBS 272.29T | Cocos nucifera | Java | MH855069 | KM199467 | KM199553 | [15] |
| Ps. celtidis | GUCC 21599T | Celtis sinensis | China | OL423535 | OL439010 | OL439012 | [33] |
| Ps. curvatispora | MFLUCC 17-1723 | Rhizophora mucronata | Thailand | MK764290 | MK764356 | MK764334 | [41] |
| Ps. curvatispora | MFLUCC 17-1722T | Rhizophora mucronata | Thailand | MK764289 | MK764355 | MK764333 | [41] |
| Ps. dawaina | INPA 2912 | Caryota mitis | Brazil | MN096659 | MN151310 | MN151308 | [85] |
| Ps. dawaina | MM14-F0015T | unknown | Dawei, Myanmar | LC324750 | LC324751 | LC324752 | [86] |
| Ps. gilvanii | INPA 2913T | Paullinia cupana | Brazil | MN385951 | MN385954 | MN385957 | [29] |
| Ps. hydeae | NTUCC 17-003.1 | Diospyros sp. | China, Taiwan | MG816313 | MG816323 | MG816333 | [87] |
| Ps. ignota | NN 42909T | Camellia sinensis | China | KU500020 | NA | KU500016 | [84] |
| Ps. indica | CBS 459.78T | Hibiscus rosa-sinensis | India | KM199381 | KM199470 | KM199560 | [15] |
| Ps. indocalami | GUCC 21600T | Indocalamus tessellatus | China | OL423536 | OL439011 | OL439013 | [33] |
| Ps. ixorae | NTUCC 17-001.1T | Lxora sp. | NA | MG816316 | MG816326 | MG816336 | [87] |
| Ps. kawthaungina | MM14F0083T | unknown | Kawthaung, Myanmar | LC324753 | LC324754 | LC324755 | [86] |
| Ps. kubahensis | UMAS-KUB-P20T | Macaranga sp. | Sarawak, Malaysia | MG818971 | NA | NA | [88] |
| Ps. myanmarina | NBRC 112264T | Averrhoa carambola | Dawei, Myanmar | LC114025 | LC114045 | LC114065 | [89] |
| Ps. rhizophorae | MFLUCC 17-1560T | Rhizophora apiculata | Thailand | MK764291 | MK764357 | MK764335 | [41] |
| Ps. simitheae | KUMCC 17-0255 | Magnolia candolli | China | MW244023 | MW602387 | MW273930 | [74] |
| Ps. simitheae | MFLUCC12-0121T | Pandanus odoratissimus | Thailand | KJ503812 | KJ503815 | KJ503818 | [90] |
| Ps. solicola | CBS 386.97T | soil in tropical forest | Papua New Guinea | MH554039 | MH554715 | MH554474 | [59] |
| Ps. Taiwanensis | NTUCC 17-002.1T | Ixora sp. | China, Taiwan | MG816319 | MG816329 | MG816339 | [87] |
| Ps. Thailandica | MFLUCC 17-1724T | Rhizophora mucronata | Thailand | MK764292 | MK764358 | MK764336 | [41] |
| Ps. Thailandica | MFLUCC 17-1725 | Rhizophora mucronata | Thailand | MK764293 | MK764359 | MK764337 | [41] |
| Ps. Theae | MFLUCC 12-0055T | Camellia sinensis | Thailand | JQ683727 | JQ683711 | JQ683743 | [14] |
| Ps. vietnamensis | NBRC 112252 | Fragaria sp. | Hue, Vietnam | LC114034 | LC114054 | LC114074 | [89] |
Holotype strains are marked with T.
Table 3.
The conidial dimension of Pestalotioid species related to this study.
| Species | Isolate Number | Conidial Size (μm) | Apical Appendages (μm) | Basal Appendage | |
|---|---|---|---|---|---|
| Number | Length | ||||
| N. baotingensis | SX41-0706 | 18-26×5-7.2 | 2-4 | 3-30.5 | 2.5-10 |
| N. Saprophytica | MFLUCC 12-0282 | 22–30×5–6 | 2-4 | 23–35 | 4-7 |
| N. brachiata | SX31-0706 | 18.5-25.3×5.5-7.5 | 1-3 | 3.7-38.7 | 2.5-8 |
| N. brachiata | MFLUCC 17-1555 | 18.5–25×5.5–6 | 1-3 | 9.5–33 | 4–9 |
| N. coffeae-arabicae | BL32-0708 | 19.2-25.3×5.3-7 | 2-4 | 10.9-22.6 | 1.4-5.4 |
| N. coffeae-arabicae | LF51-0709 | 17.8-24.2×5-7 | 2-4 | 6.6-21.6 | 2.5-6.8 |
| N. coffeae-arabicae | NM42-0706 | 17.5-23.8×5.8-7.8 | 2-4 | 12.7-31 | 2.7-9.2 |
| N. coffeae-arabicae | HGUP4019 | 16-20×5-7 | 2-4 | 11-16 | 3-5 |
| N. cubana | MH51-0708 | 19.7-30×5-6.8 | 2-4 | 15.5-32.2 | 4-7.5 |
| N. cubana | YX112-0708 | 21-29×5.6-7.3 | 2-4 | 18.7-36.5 | 3.3-10.3 |
| N. cubana | CBS 600.96 | 20-25×8-9.5 | 2-4 | 21-27 | 4-7 |
| N. oblatespora | YJ11-0708 | 18-23.2×5.5-7 | 2-4 | 10-26.5 | 2-9 |
| N. guajavicola | FMBCC 11.4 | 23.3×6.5 | 2-3 | 21.8 | 4.4 |
| N. olivaceous | LF25-0709 | 21.5-33.8×5.5-7.7 | 2-5 | 9.5-22.5 | (0)1.2-4.8 |
| N. amomi | HKAS 124563 | 18-30×4-7 | 2-3 | 7-17 | 2-5 |
| N. zingiberis | GUCC 21001 | 21‒31×6‒9.5 | 1-3 | 12-15 | 0-6 |
| N. oxyphylla | LF55-0709 | 18.8-23.5×5.3-7.0 | 2-4 | 10-25.3 | 2.5-8 |
| N. aotearoa | CBS 367.54 | 21–28×6.5–8.5 | 2-3 | 5-12 | 1.5-4 |
| N. elaeidis | MFLUCC 15-0735 | 10-20×3-7 | 2-3 | 10-20 | (0)2-6 |
| N. petila | MFLUCC 17-1738 | 21–26.5×6–7 | 2-3 | 22-29 | 3-8 |
| N. piceana | CBS 394.48 | 19.5–25×7.5–9 | 3 | 21-31 | 6-23 |
| N. samarangensis | MFLUCC 12-0233 | 18–21×6.5–7.5 | 3 | 12–18 | 3.5–5.2 |
| N. rosicola | NM47-0708 | 16.9-24.6×5.5-7.2 | 2-4 | 10-25 | 1.7-7 |
| N. rosicola | CFCC 51992 | 20.2-25.5×5.5-8 | 2-4 | 17-22.8 | 2-9.5 |
| N. vaccinii | JR31-0709 | 14.5-20.6×5.5-7.4 | 2-3 | 10-22.5 | 1.3-5.1 |
| N. vaccinii | CAA1059 | 20.9×6.4 | 2-3 | 8.9–25.3 | 1.7–6.6 |
| N. hispanica | CBS 147686 | 24.4–25.3×7.2–7.8 | 3-4 | 19.5–22.6 | 5.1–15.5 |
| N. wuzhishanensis | YX116-0708 | 19.5-26.5×4.5-6.3 | 1-3 | 9-20.8 | (0)0.8-3.8 |
| N. mianyangensis | UESTCC 22.0006 | 19–23×5.5–7 | 3 | 5.5–11 | 3–4 |
| N. yongxunensis | YX101-0708 | 18.2-25.5×5.8-7.5 | 2-4 | 10.5-24.7 | 1.7-7 |
| N. dendrobii | MFLUCC 14-0106 | 20.5–23×6.5–7.5 | 2–3 | 5–6.5 | NA |
| N. paeonia-suffruticosa | CGMCC3.23554 | 20–23×9–11 | 3–4 | 22.5–34 | 3.5–7.5 |
| P. hydei | BA11-0708 | 20.3-27.8×4.5-6.6 | 1-3 | 3.4-17.2 | 1-9.5 |
| P. hydei | MFLUCC 20-0135 | 18-35×3-6 | 1-3 | 3-12 | 2-8 |
| Ps. avicenniae | LF48-0709 | 20.8-30.7×5.8-7.9 | 1-3 | 17.2-33.3 | 2-7.8 |
| Ps. avicenniae | MFLUCC 17-0434 | 22.5-26.5×5.5-6 | 1-3 | 15.5–28.5 | 3-4 |
| Ps. myanmarina | JR34-0709 | 25.4-34.8×5.8-7.4 | 2-3 | 18.1-36.9 | 2.7-7 |
| Ps. myanmarina | NBRC 11226 | 31-38.5×6.5-9 | 2-3 | 22.5-38.5 | NA |
Isolations in this study are in bold, references see Table 2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



