
Submitted:
22 April 2024
Posted:
23 April 2024
You are already at the latest version
Abstract
This article studies the morphological parameters of vegetative and generative organs of different age groups of Crataegus ambigua from four populations in Western Karatau (Mangistau region, Kazakhstan). The study examined four populations Pop 1 Sultan Epe, Pop 2 Karakozaiym, Pop 3 Emdikorgan, and Pop 4 Samal located in various gorges of western Karatau. Some phylogenetic inference methods were applied using six genetic markers atpF-atpH, ITS, matK, psbK-psbI, rbcL, and trnH-psbA. We also used a statistical analysis of the vegetative and generative organs of plants for three age groups of plants (virgin, young generative, and adult generative). According to the age structure, Samal populations have a high concentration of young generative plants - 42.3% and adult generative plants - 30.9%. Morphological analysis showed the significance of the parameters of the generative organs and separated the Samal population into a separate group according to the main PCoA coordinates. The results of the floristic analysis showed that the populations in the Samal have a high concentration of species diversity. The comparative dendrogram of UPGMA shows the informativeness of the use of genetic markers and the psbK-psbI region can be used to judge the difference between the fourth Samal population and the other three.
Keywords:
Crataegus ambigua
; populations
; morphology
; genetic diversity
; atpF-atpH
; ITS
; matK
; psbK-psbI
; rbcL and trnH-psbA
1. Introduction
The genus Crataegus L. is one of the ancient and rich genera of species from the family Rosaceae Juss. Information about hawthorn was first found in the works of the Greek philosopher Theophrastus in 372-287. BC. [1,2,3,4]. Species of the genus Crataegus L. were found on Earth in the Cretaceous period of the Mesozoic era along with magnolias (Magnolia L.), bay trees (Laurus L.), tulip trees (Liriodendron L.), plane trees (Platanus L.) and other broad-leaved genera [5]. Representatives of the hawthorn genus spread widely in the Tertiary period [6,7].
During the Quaternary period, many hawthorns died due to glaciation, and some began migrating south [8]. After the glaciation, many species began migrating north again [9,10]. In the mountainous regions of North America, whole-leaved hawthorns appeared in the post-glacial period, and their ancestors were dissected hawthorns. A group of ancient hawthorns had deeply dissected leaves. They were the ancestors of modern Eurasian (Crataegus laevigata (Poir.) DC.) and several American dissect-leaved and small-thorned hawthorns.
The distribution range of Crataegus species is in temperate and subtropical regions of the northern hemisphere, mainly along river valleys, plains, and mountainous areas. They are found in Europe, North Africa, the mountains of Central Asia, the Caucasus, Asia Minor, Siberia, the Far East (Primorye, Sakhalin, and Kamchatka), Korea, China, Japan, Mongolia, Afghanistan, and the northwestern part of the Himalayas. The locations of most species are concentrated in the eastern part of North America - from New Foundland to the mountains of Northern Mexico. Several species are found in the western continent - from the Rocky Mountains to the Pacific coast. In a natural environment, hawthorns are usually found singly or in groups in thickets of bushes, along forest edges, in clearings and clearings, and on screes; less often they grow in sparse forests. They are distributed from sea level to the upper limit of forest vegetation in the mountains, in a wide variety of relief conditions, and on different substrates [11].
Crataegus is very polymorphic and has about 300 species [12]. Some researchers increase the diversity of hawthorn to 1000 species, since hawthorn species easily cross with each other and form many forms and hybrids [13]. Also, according to various taxonomic systems, this genus includes 1250 species, of which 1125 are American, also according to Krussmaim [14], 90 species grow in Europe and Asia and up to 800 in America, 39 species grow in the territory of the former USSR [15], in Kazakhstan, according to the flora of Kazakhstan – 7 species [16], according to M.S. Baitenov – 9 species [17]. One of the few representatives of the tree flora of Mangistau is the dubious hawthorn (Crataegus ambigua C.A.Mey. ex A.K.Becker) [18].
C. ambigua is a shrub or small tree, 3-4 m in height. The shoots are non-thorny, red-brown, covered with skin. The leaves are broadly ovate, and lobed. Petioles are short. Inflorescence 12-20 flowers. The fruits are purple-black, spherical, and broadly elliptical. This plant blooms in May bears fruit in July-August, and reproduces by seeds. Good results have been obtained from autumn sowings with freshly harvested seeds. Mesoxerophyte. narrow endemic of Western Kazakhstan. It is listed in the Red Book of Kazakhstan [19] and in the Catalog of Rare and Endangered Plant Species of the Mangistau Region [20] as a rare species in the republic. Due to the polymorphic nature of the hawthorn, there are two interpretations of its species. In several works, the name C. ambigua is retained for the Mangyshlak hawthorn [21], while other authors [22,23,24] recognize it as an independent species of C. trancaspica (Trans-Caspian hawthorn). This plant species is a narrow endemic of Western Kazakhstan [25]. It grows mainly along the bottom of ravines, chalk and gypsum gorges, and in damp places. It is found in the mountains of Western and Eastern Karatau, the Western part of Northern Aktau (Emdy Ridge), on the Tyub-Karagan Peninsula, and in Northern Ustyurt [26]. Natural populations of dubious hawthorn are found in the mountains of Western and Eastern Karatau, the Western part of Northern Aktau, the Tyubkaragan Peninsula, northern Ustyurt, outside Kazakhstan - in the southeast of the European part of Russia. It grows singly or in small groups on rocky slopes and the bottoms of wet gorges [27]. The number of dubious hawthorn individuals tends to decrease due to the harvesting of its wood for fuel and overgrazing by livestock. The plant is characterized by high salt, heat, and drought resistance. The species is listed in the Red Book of Kazakhstan and the Catalog of Rare and Endangered Plant Species of the Mangistau Region (Red Book) as a rare species in Kazakhstan [28]. The authors of the catalog of rare and endangered plant species of the Mangistau region (Red Book) (2006) recommended the organization of a reserve in the mountains of the Mangyshlak Karatau for the protection of hawthorn and several other rare plants [29].
Carrying out a comprehensive assessment of the population genetic material of C. ambigua from natural habitats is necessary to identify donors of valuable economic and biological traits. The study of the same forms under natural and introduced conditions allows us to identify the phenotypic and genotypic variability of plants. At the moment, the amount of research on the study of genetic characteristics is insignificant, but there are works on the study of the center of genetic diversity of the section Crataegus, which extends from Turkey to Iran [30]. Modern advances in the field of molecular biology make it possible to carry out genomic typing of plants from C. ambigua populations based on data on the primary DNA structure [31,32,33]. DNA barcoding technology introduced in 2003 [34] makes it possible to standardize the procedure for identifying Crataegus species using reference DNA loci. Nuclear DNA sequences, such as the ITS1 and ITS2 loci, can be reference loci. In plastid DNA, the role of reference sequences is the non-coding and spacer regions of the genes rpoB, rpoC1, rbcL, matK, psbK-psbI, trnH-psbA, atpF, atpH [35,36,37,38,39].
As is known, ITS DNA loci are located between the structural genes of ribosomal RNA 18S, 5.8S, and 26S, forming a single cluster of nuclear genes, organized in the form of tandem DNA repeats. Each ribosomal gene cluster consists of a transcribed region (genes 18S, 5.8S, and 26S), internal transcribed spacers located on either side of 5.8S, called ITS1 and ITS2, respectively, and flanking external transcribed spacers ETS1 and ETS2 [40]. ITS markers are widely used as phylogenetic markers to classify plants at different taxonomic levels, including genus, species, and subspecies [41,42].
The primary sequence of the genes rpoB, rpoC1, rbcL, matK, psbK-psbI, trnH-psbA, atpF, and atpH are used in determining the origin and relationships of Crataegus plants [43]. The non-coding regions of the rpoB, rpoC1, rbcL, matK, psbK-psbI, trnH-psbA, atpF, atpH genes are subject to large mutational events and occupy most of the plastome. In contrast, the coding regions of these genes evolve slowly and are highly conserved between species. Determination of the primary structure of intergenic spacers of these genes and introns has found wide application in the phylogenetic study of plants, thereby making a significant contribution to the processes of introduction and selection of economically valuable plants. The main goal of the study is to study new populations in the Mangistau region, based on morphological and genetic parameters and to identify the main differences between C. ambigua populations, as well as their conservation [44]. Also, C. ambigua plants are of economic value as ornamental plants, as well as producers of biologically active substances. These studies may also be associated with the use of selected natural populations and varieties of C. ambigua with their selection and introduction, which require the use of advances in molecular biology in the field of DNA marking of the original lines for selection. It is advisable to carry out the selection and assessment of promising genotypes of collection funds using DNA barcoding of constitutive loci of both genomic and cytoplasmic DNA. The DNA bar code allows the identification of C. ambigua plants used for selection and introduction at each stage of the breeding process.
2. Study Area
The study area is located in the western part of the Karatau mountain system, Mangystau region of Kazakhstan, an area with a relatively high concentration of biological diversity of various arid plant species, including valuable fruit and berry trees. The Mangistau region is located in the west of Kazakhstan, in a desert zone with various mountain systems. Karatau is a large and significant mountain system with a length of 110 km from west to east and a width of 12 km. It is divided into Western and Eastern Mangystau. The highest peak is Mount Beshoky (552 m) in Western Mangystau. The foothills of the mountain ranges are cut by ravines and potholes. The western part of Karatau has several mountain gorges forming isolated ecosystems. This mountainous terrain formed during the Permian period from metamorphic limestones, sandstones, and conglomerates. The gorge is suitable for grazing [45]. The climate is sharply continental, and extremely dry, with hot summers and moderately cold winters. This territory can be classified as an arid zone with unfavorable conditions for most species of vascular plants, which, in turn, increases interest in this territory and its natural populations [46].
3. Methods
3.1. Plant Materials
As a basis for studying morphological and genetic parameters, plant samples from four populations of C. ambigua from western Karatau, Mangystau region, were studied. Homogeneous populations were selected for the study. Populations are located in isolated gorges. Population 1 Sultan Epe in the Sultan Epe Gorge (Tyubkaragan Peninsula), Population 2 Karakozaiym in the Karakozayim Gorge (Tyubkaragan Peninsula), Population 3 Emdikorgan in the Emdikorgan Gorge (Northern Aktau Ridge), Population 4 Samal in the Samal gorge (Western Karatau Ridge). Table 1 shows the main coordinates of the population locations (Table 1).
Each population has its unique conditions. The highest-located population is Pop 4 Samal 247 meters above sea level, and the lowest-located population Pop 3 Emdikorgan is 35 meters above sea level. Figure 1 shows the location of the four study populations and photographs in natural conditions (Figure 1).
As plant material for the study of morphological indicators, individual reference plants in natural populations in the amount of ten pieces from different age groups (virginal, young generative, and adult generative) and populations were selected, as well as low trees and shrubs with brownish-gray bark, old trees with shoots are reddish-brown, with a gray coating, branching - up to the 6th order. Young shoots, often light brown, are covered with axillary spines, sometimes bare. Leaves are alternate pinnate or serrated. An analysis of the morphological parameters of the vegetative and generative organs of hawthorns of different ages was carried out (Figure 2).
Also, when studying morphological parameters, the age composition of populations was taken into account, and three groups of different-aged C. ambigua plants were identified from four populations: virgin (V) young generative (YG) adult generative (AG). The parameters of the vegetative organs of plants were studied: plant height, cm (PH); plant crown diameter, cm (CD); trunk height, cm (TH); trunk diameter, cm (TD); spike size, cm (SS); leaf length, cm (LL); leaf width, cm (SW); leaf petiole length, cm (LPL); leaf area, cm (LA). The parameters of generative organs were also studied: inflorescence diameter, cm (ID); number of flowers on 1 inflorescence, pcs (NFI); number of inflorescences on 1 branch, pcs (NIB); pedicel length, mm (PL); flower diameter, mm (FD); fruit weight (FM); fruit length (FL); fruit width(FW); number of seeds (NS); seed mass (SM).
3.2. Floristic Analysis
Population analysis of the floristic properties of plants in ontogenesis was studied using the method of O. V. Smirnova and co-author. [47]. At the same time, the names of plant species were verified against the “Flora of Kazakhstan” [48], “Illustrated Key” [49], “Identifier of Plants of Central Asia” [50], and the International Index of Names of Electronic Databases [51]. The classification of life forms of species included in the population was assessed using the method of I.G. Serebryakov [52]. To describe the populations, traditional geobotanical survey methods using ecological and morphological indicators were used. Individuals of C. ambigua were counted by age group and the floristic composition was described [53]. Plant nomenclature was determined according to POWO [54]. When describing ontogeny, we used the method of Komarov et al. [55]. The population type was determined using the method of S.V. Fedorov [56]. The following age groups were considered: young individuals (of root or seed origin); virginal (large individuals, but not reaching the generative period); young generative; and adults generative. Seedlings and senile individuals were not identified in the natural population at the time of the study.
3.3. Genetically Analysis
Fresh leaves of C. ambigua plants from the four populations studied were used for DNA extraction. Total DNA was isolated from ground leaf powder using the cetyltrimethylammonium bromide (CTAB) protocol with double chloroform purification [57]. DNA quality and concentration were assessed using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and 1% agarose gel electrophoresis. The DNA concentration was normalized to the working concentration for further analysis. The preparation of buffers was carried out by the guidelines [58]. To isolate DNA, a method using cetyltriethylammonium bromide was used [59]. DNA sequencing will be performed using the Sanger method using the BigDye™ Terminator v3.1 Cycle Sequencing Kit [60]. The obtained data on the primary structure of the DNA fragments under study will be analyzed through the NCBI genetic information database using the BLAST (Basic Local Alignment Search Tool) program [61]. To barcode the plants selected in the study, we selected DNA sequences of the chloroplast genome, such as matK, rbcL, trnH-psbA, psbK-psbI, and atpF-atpH, as well as nuclear DNA fragments ITS1 and ITS2. Table 2 shows the sequences of the primers used in the study (Table 2).
For the above primers, polymerase chain reaction modes were developed, taking into account their incomplete specificity for target sequences. Even though the primers used in the work are universal for most plants, they are not strictly specific for the plant species under study. The specificity of primers ranges from 90% to full specificity of the oligonucleotide for the target. In this connection, optimization of primer annealing was carried out in a temperature gradient from 55°C to 62°C. To perform PCR, the following concentrations of substances were used: template DNA - 120 ng; direct primer – 0.5 µM; reverse primer – 0.5 µM; dNTP 0.5 mM; 1X Pfu buffer; Pfu DNA polymerase 5 activity units; ultrapure water up to a volume of 40 µl. The amplification program for the Eppendorf Nexus Gradient thermal cycler had the following temperature conditions: +98°C (1 min) – 1 cycle; +98°C (30 sec), +55°C-62°C (1 min), +68°C (2 min) – 25 cycles; +68°C (10 min) – 1 cycle; +4°C (15 min) – 1 cycle. Separation of amplification products was carried out using DNA electrophoresis in a 1.5% agarose gel in TBE buffer with ethidium bromide (10 μg/ml). Results of PCR amplification using primers for the matK, rbcL, trnH-psbA, and ITS loci in a temperature gradient from 55°C to 62°C. For the psbI and atpF-atpH loci, primer annealing was also optimized for temperature.
The resulting DNA sequences were edited and contiguous sequences assembled (from 193 to 570 bp) using the MEGA program, version 5 (Tamura et al., 2011), and the resulting DNA sequence information was analyzed for the following basic evolutionary analyses (Kumar and Gadagkar, 2001), evolutionary distances and phylogenetic reconstructions (Nei and Kumar, 2000; Tamura et al., 2011).
3.4. Statistical Processing
Statistical processing of the results was carried out using the Statistics 10 program (StatSoft STATISTICA 10.2011) and the capabilities of the Microsoft Excel 10.1 program. Statistical processing was carried out by calculating the nonparametric Mann–Whitney test to determine the significance of differences between indicators of floristic composition between populations, as well as between morphological indicators of populations of the studied species. Mathematical processing was carried out according to the method of Kuziev R.K. and Yuldashev G. et al. [62,63]. Statistical analysis of population dynamics varies depending on the limits, which vary according to the standard deviation of the coefficient of variation, the mean error, the degree of confidence, and the degree of precision. The dendrogram was constructed using PAST 4.03 and the unweighted pair group method with arithmetic mean algorithm (UPGMA) and Boot N:1000 [64]. To compare the results, a one-way analysis of variance (ANOVA) was used, which found a significant correlation between soil type and the main parameters of plants in the population. When analyzing primary data, correlation coefficients were calculated using the statistics program R Studio (IDE) for Windows (R version 3.6.0, 2019). The mean values of the main quantitative traits of the four populations were grouped using the PCoA (principal component analysis) method. PCoA was performed using the Numerical Taxonomy and Multivariate Analysis System version 2.1. (NTSYS-pc) [65].
3.5. Results
Floristic Analysis of C. ambigua Populations
Population 1 Sultan Epe in the Sultan Epe Gorge (Tyubkaragan Peninsula). The Sultan Epe Gorge is a coastal canyon, it is a plateau with the highest absolute elevations of about 200 m. The plateau drops steeply to the sea, from which deep ravines cut into it, branching at their peaks, more numerous and further penetrating inland from the southern side. The soils on the plateau are loamy and sandy loam, gravelly, solonetzic, and solonchak. On the slopes of ridges and ridges, poorly developed eroded soils are formed, and there are outcrops of bedrock. The floristic composition of the site is heterogeneous, the hawthorn-mulberry-forb (Crataegus ambigua - Morus alba - Herba varia) community has a pronounced layering, the dominant is Crataegus ambigua, the co-dominant is Morus alba. The tree layer (3-5 m in height) occupies a significant part of the territory of the gorge and is confined to the steep upper and lower gentle parts of the slopes and is characterized by a well-developed undergrowth dominated by thickets of Crataegus ambigua, Morus alba, Morus nigra, in small quantities, Salix alba is found scattered, in a single specimen of Elaeagnus angustifolia. The shrub layer is well formed, its share in the cover is 40-50%. The dominant ones are Caragana grandiflora and Rhamnus sintenesii, they are found everywhere, both in rock outcrops, and along ravines, and ravines, and sai, evenly distributed both on the plains and on the slopes. In the shrub layer, in addition to the dominant species, there are secondary subshrubs - Nanophyton erinaceum, Artemisia lercheana, Atraphaxis replicata, Limonium suffriticosum, Salsola arbuscula. The herbaceous cover is ambiguous in composition, well-developed, and relatively rich. The most common dominants are Cardaria draba, Chorispora tenella, Psathyrostachys juncens, abundant ephemerides Poa bulbosa, Eremopyrum triticeum, E. orientale, Veronica amoema, Ceratocephala testiculata, Descurainia Sophia. Along the lower gentle parts of the slopes and the trails, small numbers of annual and perennial herbs are common - Allium sabulosum, Gypsophila alsinoides, Petrosimonia glaucescens, Androsace maxima, Scorzonera pusilla, Sisimbrium loeselii, Camelina sylvestris, Nonnea caspia, Juncus bufonius, Teucrium polium, Sinaps arvensis, in very small quantities, rare specimens include Asparagus persicus, Galium aparine, Malva pusila, Marrubium vulgare, Equisetum ramosissima, Potentilla supine. Analysis of the systematic composition of the flora shows that the first 8 leading families contain 58.1% of the total species composition of the flora. The richest species are from the families Brassicaceae (7 species), Poaceae (4 species), Asteraceae (3 species) and Chenopodiaceae (3 species). It includes species of the following main life forms: trees – 5 (11.6%), shrubs – 2 (4.7%), subshrubs – 6 (14.0%), perennial herbs – 13 (30.2%), and young children – 17 (39.5%).
Population 2 Karakozaiym in the Karakozaiym Gorge (Tyubkaragan Peninsula). The Karakozayim Gorge is located on the Tyubkaragan peninsula, 25 km west of the village of Fort Shevchenko, and occupies ridges with heavily eroded slopes, along which limestone-shell rocks are exposed. Ridges with soft slopes and plateau-like peaks are composed of sandstones. Ridges alternate with complex plains of different hypsometric levels. In the Karakozayim gorge, the nature of the vegetation cover is a forb-hawthorn (Crataegus anbigua – Herba varia) community. The total projective coverage is 45-55%. The landscape is natural, the relief is mountainous. The tree layer is formed by single trees Morus alba (up to 4.5 m tall) and the dominant Crataegus ambigua. The shrub layer on rocky ridge tops with limestone outcrops is a characteristic element of landscapes. On the steep upper parts of slopes and flat peaks with underdeveloped rocky-gravelly soils, the most common shrubs up to 500-800 cm in height are Rhamnus sintenesii, Caragana grandiflora; near the springs there are single specimens of Rubus caesius and Reaumuria fruticosa. On the plains and along the lower gentle parts of the slopes there is a complex of subshrubs Nanophyton erinaceum, Artemisia gurganica, Atraphaxis replicata, Teucrium polium. On plains with clay soils and rocky ridges, Onopordum acanthium, Allium sabulosum, Lagochilus acutilobus, Haplophyllum obtusifolim, Onosma staminea, Alopecurus arundinaceus, Ceratocephala testiculata, Cramde edentula are usually developed. On rocky bottoms of sais and ravines with flat limestone outcrops, along ravines in undulating relief, the ephemerides Poa bulbosa, Eremopyrum orientale, Alyssum desertorum, A. turkestanicum are abundant, and there is a variety of annual and perennial herbs - Androsace maxima, Tragopogon ruber, Chorispora tenella, Salsola australis, Nonea caspica, Gagea peticulata, Lepidium ruderale, Bromus squarrosus, Minuartia regeliana, scattered spots of Fumaria parviflora, Veronica amoema, Lappula sinaica, in the rocks near the springs there are thickets of Mentha longifolia, single specimens of Potentilla transcaspia, in shady places in the bushes of Equisetum ramosissima and Galium humifusum, as well as on the lower parts of the slopes and trails of ridges, Cardaria draba, Descurainia Sophia, Glaucium corniculatum are abundant. Analysis of the systematic composition of the flora shows that the first 7 leading families contain 58.5% of the total species composition of the flora. The richest species in the family are Brassicaceae (7 species), Poaceae (4 species), Boraginaceae (3 species), Lamiaceae (3 species), Rosaceae (3 species), Asteraceae (2 species) and Chenopodiaceae (2 species). This circumstance gives reason to consider the flora of the Karakozay gorges to be quite monotonous. It includes species of the following main life forms: trees – 2 (4.9%), shrubs – 4 (9.8%), subshrubs – 4 (9.8%), perennial herbs – 15 (36.5%), and young children – 16 (39.0%).
Population 3 Emdikorgan in the Emdikorgan Gorge (Northern Aktau Ridge). Emdikorgan Gorge - located in the Northern Aktau ridge, a small gorge about 300-350 m long and about 50 m wide, with steep slopes on all sides. The northern walls of the gorge are steep, and rocky, falling to the foothill plains at a height of 100-200 m with a foot strewn with steep debris - rocks, boulders, stones. The soil cover on the mountain slopes is thin and gravelly. Soils of light mechanical composition predominate - light loamy, sandy loam, sandy. Individuals of hawthorns are located in the hollows of the gorge, between the rocks, where more humid growing conditions are formed. On the gentle slopes of the gorge, wormwood-grass and catnip-mint communities are formed, along the bottom of the gorge - forb-shrub communities, and in rocky areas - ephedra populations. Hawthorn in this gorge forms a hawthorn-joster-forb (Crataegus ambigua – Rhamnus sintensii – Herba varia) community. The dominant species in the community is the dubious hawthorn, the codominant is Sintensa’s jockstrap, and the remaining species are classified as components of the community. Plants in this community are also arranged in 3 tiers: woody (up to 250 cm in height), shrub (100-120 cm in height), and herbaceous (from 10 to 50 cm in height). The tree layer is formed by mature hawthorn trees. The shrub layer is formed by Sintensa’s jockstrap, tamarix, and young specimens of dubious hawthorn. The herbaceous layer, the most numerous in species composition, is formed by Falcaria vulgaris, Galium aparine, Haplophyllum bungee, Ixiolirion tataricum, Linaria leptoceras, Marrubium vulgare, Meniocus linifolius, Lepidium perfoliatum, Mentha longifolia, Onosma stamineum, Poa bulbosa, Prangos odontalgica, Scandix stellata and others. The hawthorns in the Emdikorgan gorge are in good condition, with larger leaf blades, the degree of fruiting is high - more than 65% and no damage to the plants was detected. The species composition of the community is represented by 58-60 species from 41 genera and 23 families. Despite the harsh and arid conditions and the lack of available moisture throughout the growing season, the number of species in this gorge is higher than in previous ones in Western Karatau. Among the identified species, the predominant number belongs to the families Asteraceae, Poaceae, Chenopodiaceae, and Lamiaceae - about 65%. The remaining families contain 1-2 genera and species. Among the life forms, herbaceous perennials predominate - 32 species or 53.3%. The second position is occupied by young plants - 16 species or 29.3%, woody forms are in last place - 11 species or 18.4%. Hawthorn in this gorge has a small number of specimens, but both old trees and young individuals are present.
Population 4 Samal in the Samal Gorge (Western Karatau Ridge). The Samal Gorge is located in the northern part of the Western Karatau ridge, a deep, highly winding gorge, about 2 km long, with a width ranging from 30 to 100 m. The southern slopes are steep, more dissected by a dense network of canyon-like ravines than the northern ones. The height of the slopes ranges from 100 to 350 m. The slopes of the mountains are rocky and gravelly, with numerous outcrops of bedrock. Western Karatau is composed of Permian and Triassic sandstones, shales, and siltstones with interlayers of conglomerates and limestones. At the foot of the ridge, Jurassic variegated deposits come to the surface - clays, siltstones, sandstones, and sands are less common. An abundant stream of fresh water flows along the bottom of the gorge for about a kilometer. Along the bottom of the gorge along the stream grow dense thickets of mint with Dubrovnik, catnip, and horehound (Mentha longifolia, Teucrium polium, Marrubium vulgare, Nepeta cataria), around which there are areas of tree and shrub vegetation of Crataegus ambigua, Rhamnus sintensii, less often Caragana grandiflora. The lower parts of the slopes are occupied by mesophytic and mesoxerophytic communities with the participation of Centaurea squarossa, Cousinia onopordioides, Verbascum songaricum, Verbascum blattaria, Plantago lanceolata, Inula britanica, Medicago caerulea, Malva pusila, Equisetum ramosissimum, Rubus caesius, Stellaria media, Phragmites communis and others. On the mountain slopes between the rocks along the banks of a flowing spring rivulet, single trees grow - Elaeagnus angustifolia and Ulmus pumila. The upper and lower parts of the gentle slopes are occupied by sparse white-earth wormwood and Gurgaon-wormwood communities with the participation of Artemisia terrae-albae, Artemisia gurganica, Tanacetum santolina, Ephedra distachya, Echinops ritro, Alhagi pseudalhagi Acanthophyllum pungens, Meristotropis triphylla; grasses are abundant: Stipa caspia, S. caucasica, Agropyron fragile, Poa bulbosa. Due to the inaccessibility and steepness of the slopes, the gorge is not used for grazing livestock. Hawthorn doubtful forms hawthorn-mint-forb (Crataegus ambigua – Mentha longifolia – Herba varia) communities. The appearance of the hawthorns in the Samal Gorge is at a good level, the plants have higher vitality, and there are no signs of damage to the plants. The dominant species in the community is dubious hawthorn, the codominant is longleaf mint. The remaining species are classified as components of the plant community. The species composition is quite abundant, including both mesophytic and xerophytic elements. Based on taxonomic analysis, it was revealed that 55 species from 49 genera and 25 families grow on the territory of the Samal Gorge. The predominant number of species belong to the families Asteraceae, Brassicaceae, Lamiaceae, Poaceae, Rosaceae, and Fabaceae. Species from these 6 leading families make up 58.2% of the total number of plant species in the gorge. Analysis of the nature of life forms showed that herbaceous perennials dominate in the Samal gorge - 26 species (47.3% of the total number of species); in second place are herbaceous young plants - 13 species (23.6%), third place is occupied by trees and shrubs - 5 species each (18.2%), subshrubs - 3 species (5.5%), subshrubs - 2 species (3. 6%) and shrub – 1 species (1.8%).
3.6. Age Composition of C. ambigua in Populations
Growing in different conditions, hawthorn is distinguished by various indicators in the growing season, productivity, and development. Identifying the characteristics of growth and development, as well as the variability of individuals, allows for targeted selection of forms for introduction into culture and selective selection.
When inventorying the population, 4 groups of hawthorns of different ages were taken into account: juveniles (of root sucker or seed origin); virginal (large individuals, but not reaching the generative period); young generative; adults are generative (Table 3).
The data obtained show that the age composition of C. ambigua in the gorges is not the same. Thus, the largest number of specimens was noted for the Sultan Epe Gorge, followed by the Karakozayim Gorge. In third place is the Samal Gorge, the smallest number of individuals is in the Emdikorgan Gorge.
It is worth noting that the low number of hawthorn individuals in the Emdikorgan Gorge is associated with its small size. The size of the Sultan Epe Gorge, as the largest, explains the largest number of specimens of the species under study.
A more important parameter is the ratio of the main age groups of plants, which reflects the state of the populations. Thus, the predominance of young regenerative individuals in the population structure indicates a young state and the possibility of further development; middle-aged individuals – about population stability; old individuals - about the degradation of the population with the prospect of extinction.
3.7. Morphological Structure of C. ambigua Populations
Populations of C. ambigua in the territory of Western Karatau (Mangistau region, Kazakhstan) are characterized as low trees or shrubs. These studies involved reference plants from four populations of C. ambigua from different age groups in vegetative and generative organs. Age states: virgin (V); young generative (YG); adult generative (AG). The parameters of the vegetative organs of plants were studied: plant height, cm (PH); plant crown diameter, cm (CD); trunk height, cm (TH); trunk diameter, cm (TD); spike size, cm (SS); leaf length, cm (LL); leaf width, cm (SW); leaf petiole length, cm (LPL); leaf area, cm (LA). The parameters of generative organs were also studied: inflorescence diameter, cm (ID); number of flowers on 1 inflorescence, pcs (NFI); number of inflorescences on 1 branch, pcs (NIB); pedicel length, mm (PL); flower diameter, mm (FD); fruit weight (FM); fruit length (FL); fruit width(FW); number of seeds (NS); seed mass (SM). The results for the main parameters are presented in tables (Table 4 and Table 5).
A correlation matrix was also generated for the three age groups (Figure 3, Figure 4 and Figure 5). The four-population factor was also included in the correlation matrix.
A positive significant correlation was recorded in the generative organs of young generative (YG) and adult generative (AG) plants. It has been determined that one of the main indicators is plant height (PH) according to age characteristics in various populations. A box plot analysis of variables by plant height was generated (Figure 6).
The results of the analysis of morphological indicators were also analyzed by the PCoA method (principal component analysis) according to two parameters (indicators of vegetative and generative organs of three age groups from four populations). The results are presented below (Figure 7 and Figure 8).
PCoA grouping (principal component analysis) showed an obvious difference in the fourth population according to the generative characteristics of the organs. The indicators of the vegetative organs do not reveal this difference so significantly. To ensure a more accurate delineation of the four populations, the method of constructing dendrograms according to two parameters (vegetative and generative organs) using the UPGMA method was applied (Figure 9).
The comparative figure shows two dendrograms for vegetative and generative traits. The differences between the fourth population from the rest are shown in 100 values.
3.8. Genetic Analysis of C. ambigua Populations
Sequencing results were obtained using six genetic markers atpF-atpH, ITS, matK, psbK-psbI, rbcL, and trnH-psbA. The raw sequence data is attached to this article in additional files in FASTA format (Table S1). Table 6 presents the statistical parameters of the sequences (length, conserved sites, variable sites, parsimony informative sites, singleton).
The shortest sequence was recorded in psbK-psbI 193 bp. The longest sequence is in matK and is 570 bp. Table 7 shows the frequency of occurrence of nucleotides in four populations and six main markers.
An assessment was made of the informativeness of certain genetic markers for the process of dating the genetic diversity of the studied populations. Figure 10 shows the results of constructing six UPGMA dendrograms for various genetic markers.
Various figures show different phylogenetic dendrograms giving different scenarios for the relationship of the studied populations. A special pattern is also observed when constructing dendrograms.
4. Discussion
Research on the status of C. ambigua populations using morphological and phylogenetic methods on the territory of Kazakhstan and in the world is not significant. Populations of C. ambigua have characteristic traits that are manifested in morphological and genetic states. Studies are confirming the significance of differences between populations in vegetative and generative parameters of organs, as well as the significance of genetic studies [66,67].
Floristic analysis showed differences among the studied populations. Population 1 Sultan Epe is the richest in species of the families Brassicaceae (7 species), Poaceae (4 species), Asteraceae (3 species), and Chenopodiaceae (3 species). It includes species of the following main life forms: trees – 5 (11.6%), shrubs – 2 (4.7%), subshrubs – 6 (14.0%), perennial herbs – 13 (30.2%), and young children – 17 (39.5%). Population 2 Karakozaiym is richest in species of the families Brassicaceae (7 species), Poaceae (4 species), Boraginaceae (3 species), Lamiaceae (3 species), Rosaceae (3 species), Asteraceae (2 species), and Chenopodiaceae (2 species). This circumstance gives reason to consider the flora of the Karakozay gorges to be quite monotonous. It includes species of the following main life forms: trees – 2 (4.9%), shrubs – 4 (9.8%), subshrubs – 4 (9.8%), perennial herbs – 15 (36.5%), and young children – 16 (39.0%). Population 3 Emdikorgan is represented by 58-60 species from 41 genera and 23 families. Despite the harsh and arid conditions and the lack of available moisture throughout the growing season, the number of species in this gorge is higher than in previous ones in Western Karatau. Among the identified species, the predominant number belongs to the families Asteraceae, Poaceae, Chenopodiaceae, and Lamiaceae - about 65%. The remaining families contain 1-2 genera and species. Population 4 Samal grows together with 55 species from 49 genera and 25 families. The predominant number of species belong to the families Asteraceae, Brassicaceae, Lamiaceae, Poaceae, Rosaceae, and Fabaceae. Species from these 6 leading families make up 58.2% of the total number of plant species in the gorge. Analysis of the nature of life forms showed that herbaceous perennials dominate in the Samal gorge - 26 species (47.3% of the total number of species); in second place are herbaceous young plants - 13 species (23.6%), third place is occupied by trees and shrubs - 5 species each (18.2%), subshrubs - 3 species (5.5%), subshrubs - 2 species (3. 6%) and shrub – 1 species (1.8%). Such an abundance of species diversity was also recorded in this territory over a long period, which confirms the stability of plant communities [68].
The age parameters of C. ambigua populations vary and are not homogeneous. In the Samal population, a predominance of generative individuals of dubious hawthorn was noted. The share of generative specimens was 73.2%, pregenerative specimens – 26.8% of the total number of specimens. Thus, the population of dubious hawthorns in the Samal Gorge can be characterized as middle-aged stable, developing with a predominance of generative individuals. In the population of Sultan Epe, there is a predominance of generative plants of dubious hawthorn. The share of young generative individuals was 42.2% of the total number of specimens, adult generative individuals - 28.2%. Young and virgin individuals accounted for about 30%. Thus, the population of dubious hawthorn in the Sultan Epe Gorge can be characterized as middle-aged, developing with a predominance of generative individuals. In the Karakozayim population, 2 peaks of age groups of doubtful hawthorn plants were noted. The first peak occurs in young generative plants (67.7%), and the second peak in young plants (26.9%). At the same time, the proportion of virginal plants and adult generative ones remains very low. This ratio is formed in the population when experiencing extremely unfavorable conditions in the past period. Part of the population was destroyed, probably as a result of many years of drought, but is currently recovering. Thus, we can characterize the population of dubious hawthorn in the Karakozayim Gorge as unstable and stopping. In the Emdikorgan population, a predominance of generative individuals of dubious hawthorn was noted. Thus, the share of generative specimens was 52.7%, and pregenerative specimens – was 47.3%, that is, different age groups of plants are evenly represented. Thus, the hawthorn population in the Emdikorgan Gorge can be characterized as middle-aged, stable, and developing with the dominance of young generative plants. This structure is also consistent with previous studies [69].
Morphological parameters of vegetative and generative organs show significance in differences between C. ambigua populations. The fourth population, Samal, differs from other populations in coordinates and forms an auxiliary group according to the generative trait, which is presented in Figure 8. These results are also confirmed in similar studies and can be used in determining key populations of C. ambigua [70].
Sequencing results using six genetic markers atpF-atpH, ITS, matK, psbK-psbI, rbcL, and trnH-psbA showed positive results. Genetic differences between populations were identified for four out of six markers, which shows the informativeness of these genetic markers for populations of C. ambigua. Genetic markers are one of the main tools for identifying key populations [71,72,73,74,75]. The fourth population of Samal, based on the genetic marker psbK-psbI, stood out as the root group of the others. The results of this study can be used to preserve the biological diversity of the region [76,77].
5. Conclusions
Research on C. ambigua populations in the western Karatu region of the Mangystau region of Kazakhstan has shown significant results. The positive application of morphological analysis and genetic dating methods has been proven. Also, the obtained data on floristic analysis and age structure of C. ambigua populations confirm the differences between populations. Populations according to age structure, the fourth population of Samal has a high concentration of 30.9% of adult generative plants. This also proves the significant indicators of the generative organs of young generative and adult generative plants. This group of plants from the fourth Samal population forms a separate group according to these indicators. Also, this population has a high percentage of young generative plants of 42.3%. According to the results of floristic analysis, it was shown that the populations in the Samal gorge are dominated by herbaceous perennials - 26 species (47.3% of the total number of species); in second place are herbaceous young plants - 13 species (23.6%), third place is occupied by trees and shrubs - 5 species each (18.2%), subshrubs - 3 species (5.5%), subshrubs - 2 species (3.6%) and shrub – 1 species (1.8%). It can be judged that this territory is especially rich in species diversity. The comparative dendrogram of UPGMA shows the informativeness of the use of genetic markers. The psbK-psbI region can be used to judge the difference between the fourth Samal population and the other three. Based on the above conclusions from the application of methods for studying population materials, it can be judged that the populations of the western Karatau of the Mangystau region.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Results of sequencing materials of four populations of C. ambigua using genetic markers atpF-atpH, ITS, matK, psbK-psbI, rbcL and trnH-psbA in FASTA format.
Author Contributions
Conceptualization, I.A. and D.N.; methodology, O.A.; software, O.A.; validation, S.M. and B.I.; formal analysis, O.A.; investigation, T.A.; resources, I.A.; data curation, O.A.; writing—original draft preparation, O.A.; writing—review and editing, O.A.; visualization, O.A.; supervision, I.A.; project administration, I.A.; funding acquisition, I.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by «Scientific and practical basis of reproduction, conservation, use of fruit-berry plants of the natural flora of Western, Eastern, Central and Northern Kazakhstan to ensure food security BR21882166». Committee of Science of the Ministry of Science and Higher Education of the Republic of Kazakhstan.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Letukhova, V.; Potapenko, I.; Fedoronchuk, M. Taxonomic analysis of some species of the genus Crataegus (Rosaceae) from the flora of Crimea. Ukr. Bot. J. 2014, 71, 182–187. [Google Scholar] [CrossRef]
- Poletiko, O.M. Hawthorn - Crataegus L // In the book: Trees and shrubs of the USSR.-M.; L.: USSR Academy of Sciences, 1954. [Google Scholar]
- Rusanov, F. N. (1965). Introduced hawthorns of the botanical garden of the Academy of Sciences of the UzSSR. Dendrology of Uzbekistan, 1, 8-254 (In Russian).
- Chang, Q. , Zuo, Z. Hawthorn. The Journal of Clinical Pharmacology 42(6), 605–612.
- Kristafovich, A.N. , Paleobotany. - L.: Science, 1957. -594s.
- Alekhin, V.V. , Kudryasheva L.V., Govorukhin V.S. Geography of plants with basics of botany. - M.: Uchpedgiz, 1957. -519 pp.
- Zhukovsky, P.M. Cultivated plants and their relatives. -L.: Kolos, 1964. -792 p.
- Wulf, E.V. Introduction to the historical geography of plants. - M.; L.: USSR Academy of Sciences, 1933. [Google Scholar]
- Rusanov, F.N. History of the development of the river Crataegus L // B book: Introduction and acclimation of plants. -Tashkent: Fan, 1970.- Issue 6.- pp. 20-28.
- Tsinovskis, R.E. Hawthorns of the Balkhash region. - Riga: Zinatne, 1971.-384 p.
- Boboreko, E.Z. Hawthorn. -Minsk: Science and Technology, 1974. -222 s.
- Baytenov, M.S. Flora of Kazakhstan. – Almaty: Gylym, 2001.- 280 p.
- Sokolov, S.Ya. , Svyazeva O.A. Geography of woody plants USSR.-M.; L.: Nauka, 1965. [Google Scholar]
- Kriissmann, G. Die Laubgeholze Aiifgabe. - Berlin: Verlag Parey, 1951.-С.118-125.
- Komorov, VL. 1939. Flora SSSR. M. 9. 416-468 (in Russian).
- Flora of Kazakhstan. – Alma-Ata, 1961. - T. 4. – 410 p.
- Baytenov, M.S. Flora of Kazakhstan. – Almaty: Gylym, 2001.- 280 p.
- Kubentayev, S.A.; Alibekov, D.T.; Perezhogin, Y.V.; Lazkov, G.A.; Kupriyanov, A.N.; Ebel, A.L.; Izbastina, K.S.; Borodulina, O.V.; Kubentayeva, B.B. Revised checklist of endemic vascular plants of Kazakhstan. PhytoKeys 2024, 238, 241–279. [Google Scholar] [CrossRef] [PubMed]
- Red Book of Kazakhstan. – Almaty: Science, 1981. – T. 2. – 99 p.
- Aralbay, N.K. , Kudabaeva G.M., Imanbaeva A.A. and others. Catalog of rare and endangered plant species of the Mangystau region (Red Book). – Aktau, 2006. -32 p.
- Safronova, I.N. Deserts of Mangyshlak (essay on vegetation) // Proceedings of Bot. Inta RAS. - 1996. – Issue 18. – 211 p.
- Kisykov, U.K. Materials on the flora of mountain Mangyshlak // Tr. Institute of Botany An KazSSR. - 1955. – T.1. – pp. 84-117.
- Sumbembayev, A.; Abugalieva, S.; Danilova, A.; Matveyeva, E.; Szlachetko, D. Flower morphometry of members of the genus Dactylorhiza Necker ex Nevski (Orchidaceae) from the Altai Mountains of Kazakhstan. Biodiversitas J. Biol. Divers. 2021, 22. [Google Scholar] [CrossRef]
- Romanovich, V.V. Towards the use of elements of wild flora for landscaping industrial centers and settlements of the Mangyshlak Peninsula. // Tr. Institute of Regional Pathology. - Alma-Ata: Science, 1969. – T.18. – 187 p.
- Cherepanov, S.K. Vascular plants of the USSR. - Leningrad: Science, 1981. – 509 p.
- Lyubimov, V.B. On the question of taxonomy of Сrataegus trancaspica A.Pojark. // Bulletin of the Main bot. garden – M.: Nauka, 1989. – Issue 151. – P. 47-50.
- Imanbaeva. A.; Garden, R.O.K.M.E.B.; Ishmuratova, M.Y.; Tuyakova, A.T. Screening of Mangystau Flora for Wild Relatives of Cultivated Plants. Central Eur. J. Bot. 2015, 1, 12–20. [Google Scholar] [CrossRef]
- Pavlov, N.B. Flora Kazahstana [Flora of Kazakhstan]. 1956. Vol 1. Alma-Ata: AS Kazakh SSR, 347 p.
- Duysenova N., I. et al. The age composition of populations of Crataegus ambigua in the natural conditions of Mangyshlak //Вестник Карагандинскoгo университета. Серия: Биoлoгия. Медицина. Геoграфия. – 2017. – №. 4. – С. 29-34.
- DÖNMEZ, A. A. (2004). The genus Crataegus L. (Rosaceae) with special reference to hybridisation and biodiversity in Turkey. Turkish Journal of Botany, 28(1), 29-37.
- Zargar, M.; Dyussibayeva, E.; Orazov, A.; Zeinullina, A.; Zhirnova, I.; Yessenbekova, G.; Rysbekova, A. Microsatellite-Based Genetic Diversity Analysis and Population Structure of Proso Millet (Panicum miliaceum L.) in Kazakhstan. Agronomy 2023, 13, 2514. [Google Scholar] [CrossRef]
- Zeinullina, A. , Zargar, M., Dyussibayeva, E., Orazov, A., Zhirnova, I., Yessenbekova, G.,... & Hu, Y. G. (2023). Agro-Morphological Traits and Molecular Diversity of Proso Millet (Panicum miliaceum L.) Affected by Various Colchicine Treatments. Agronomy, 13(12), 2973.
- Orazov, A. , Yermagambetova, M., Myrzagaliyeva, A., Mukhitdinov, N., Tustubayeva, S., Turuspekov, Y., & Almerekova, S. (2024). Plant height variation and genetic diversity between Prunus ledebouriana (Schlecht.) YY Yao and Prunus tenella Batsch based on using SSR markers in East Kazakhstan. PeerJ, 12, e16735.
- Hebert P., D. , Cywinska A., Ball S. L., deWaard J. R. Biological identifications through DNA barcodes // Proc Biol Sci. ‒ 2003. ‒ Vol. 270, № 1512. ‒ C. 313-321.
- Kress W., J. , Erickson D. L. A two-locus global DNA barcode for land plants: the coding rbcL gene complements the non-coding trnH-psbA spacer region // PLoS One. ‒ 2007. ‒ Vol. 2, № 6. ‒ C. e508.
- Kress W., J. , Erickson D. L. DNA barcodes: methods and protocols // Methods Mol Biol. ‒ 2012. ‒ Vol. 858. ‒ C. 3-8.
- Chase M., W. , Salamin N., Wilkinson M., Dunwell J. M., Kesanakurthi R. P., Haider N., Savolainen V. Land plants and DNA barcodes: short-term and long-term goals // Philos Trans R Soc Lond B Biol Sci. ‒ 2005. ‒ Vol. 360, № 1462. ‒ C. 1889-95.
- Lahaye, R. , van der Bank M., Bogarin D., Warner J., Pupulin F., Gigot G., Maurin O., Duthoit S., Barraclough T. G., Savolainen V. DNA barcoding the floras of biodiversity hotspots // Proc Natl Acad Sci U S A. ‒ 2008. ‒ Vol. 105, № 8. ‒ C. 2923-8.
- Chen, S. , Yao H., Han J., Liu C., Song J., Shi L., Zhu Y., Ma X., Gao T., Pang X., Luo K., Li Y., Li X., Jia X., Lin Y., Leon C. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species // PLoS One. ‒ 2010. ‒ Vol. 5, № 1. ‒ C. e8613.
- Vol. 9, №1. – С.32-43.
- Nieto Feliner, G. , Rosselló J. A. Better the devil you know? Guidelines for insightful utilization of nrDNA ITS in species-level evolutionary studies in plants // Mol Phylogenet Evol. ‒ 2007. ‒ Vol. 44, № 2. ‒ C. 911-9.
- Schultz, J. , Müller T., Achtziger M., Seibel P. N., Dandekar T., Wolf M. The internal transcribed spacer 2 database--a web server for (not only) low level phylogenetic analyses // Nucleic Acids Res. ‒ 2006. ‒ Vol. 34, № Web Server issue. ‒ C. W704-7.
- Patwardhan, A. , Ray S., Roy A. Molecular Markers in Phylogenetic Studies-A Review // J. Phylogen. Evolution. Biol. – 2014, – Vol.2, №2. - Р.1-9.
- Orazov, A.; Myrzagaliyeva, A.; Mukhitdinov, N.; Tustubayeva, S. Callus induction with 6-BAP and IBA as a way to preserve Prunus ledebouriana (Rosaceae), and endemic plant of Altai and Tarbagatai, East Kazakhstan. Biodiversitas J. Biol. Divers. 2022, 23. [Google Scholar] [CrossRef]
- Кoshim, A.G.; Sergeyeva, A.M.; Bexeitova, R.T.; Aktymbayeva, A.S. LANDSCAPE OF THE MANGYSTAU REGION IN KAZAKHSTAN AS A GEOMORPHOTOURISM DESTINATION: A GEOGRAPHICAL REVIEW. Geoj. Tour. Geosites 2020, 29, 385–397. [Google Scholar] [CrossRef]
- Sagyndykova, M.; Imanbayeva, A.; Gassanova, G.; Ishmuratova, M. Current Status and Resources of Alhagi pseudalhagi (Fabaceae) in the Atyrau Region, Western Kazakhstan. Diversity 2024, 16, 219. [Google Scholar] [CrossRef]
- Smirnova, O.V.; Zaugolnova, L.B.; Ermakova, I.M. Cenopopulation of Plants; Science Publishing House: Moscow, Russia, 1976; p. 217. [Google Scholar]
- Pavlov, N.B. Flora Kazahstana [Flora of Kazakhstan]; Publishing House of the Kazakh Academy of Sciences: Almaty, Kazakhstan, 1956; Volume 1, p. 347. [Google Scholar]
- Goloskokov, V.P. (Ed.) Illiustrirovannyi Opredelitel Rastenii Kazakhstana [Illustrated Determinant of Plants of Kazakhstan]; Nauka: Almaty, Kazakhstan, 1972; Volume 2. (In Russian) [Google Scholar]
- Kamelin, R.V. Key to Plants of Central Asia. A Critical Abstract of Flora; Science Publishing House: Leningrad, Russia, 2015; Volume 11. [Google Scholar]
- International Plant Names Index. Available online: www.ipni.org (accessed on 30 January 2024).
- Serebryakov, I.G. Ecological Morphology of Plants. Life Forms of the Overgrowths and Conifers; High School: Moscow, Russia, 1982; 380p. (In Russian) [Google Scholar]
- Shay, J.E.; Pennington, L.K.; Montiel-Molina, J.A.M.; Toews, D.J.; Hendrickson, B.T.; Sexton, J.P. Rules of Plant Species Ranges: Applications for Conservation Strategies. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Kew Royal Botanical Garden. Plants of the World Online. Available online: www.powo.science.kew.org (accessed on 30 January 2024).
- Komarov, A.; Palenova, M.; Smirnova, O. The concept of discrete description of plant ontogenesis and cellular automata models of plant populations. Ecol. Model. 2003, 170, 427–439. [Google Scholar] [CrossRef]
- Fedorova, S.V. Methodological Approaches in Population Botany and Plant Ecology. Am. J. Biosci. 2020, 8, 73. [Google Scholar] [CrossRef]
- Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin 19:11–15.
- Maniatis, T. , Fritsch E.E., Sambrook J. Molecular cloning. A laboratory manual. – New York: Cold Spring Harbor Laboratory, 1982. – P.545.
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed]
- Available online:. Available online: http://blast.ncbi.nlm.nih.gov (accessed on 7 November 2021).
- Kuziev, R.K.; Sektimenko, V.E. Soils of Uzbekistan; Extremum Press Publishing House: Tashkent, Uzbekistan, 2009; p. 351. (In Russian) [Google Scholar]
- Kuziev, R.K.; Yuldashev, G.Y.U.; Akramov, I.A. Bonitization of Soils; The Way of Science Publishing House: Tashkent, Uzbekistan, 2004; p. 127. [Google Scholar]
- Joshi, S.P.; Gupta, V.S.; Aggarwal, R.K.; Ranjekar, P.K.; Brar, D.S. Genetic diversity and phylogenetic relationship revealed by intersimple sequence repeat (ISSR) polymorphism in the genus Oryza. Theor. Appl. Genet. 2000, 100, 1311–1320. [Google Scholar] [CrossRef]
- Rohlf, F. NTSYSpc: Numerical Taxonomy and Multivariate Analysis System, Version 2.02; Exeter Software: Setauket, NY, USA, 1998. [Google Scholar]
- Tuyakova, A. T. , Imanbayeva, A. A., Duysenova, N. I., & Ishmuratova, M. Y. (2016). Anatomical structure study of aerial organs of Crataegus ambigua CA Mey. Biosciences Biotechnology Research Asia, 13(3), 1303-1309. [CrossRef]
- Phipps, J.B. Biogeographic, Taxonomic, and Cladistic Relationships Between East Asiatic and North American Crataegus. Ann. Mo. Bot. Gard. 1983, 70, 667. [Google Scholar] [CrossRef]
- Duysenova, N. I. , Imanbaeva, А. А., Tuyakova, А. Т., & Kopbaeva, G. B. (2017). The age composition of populations of Crataegus ambigua in the natural conditions of Mangyshlak. Вестник Карагандинскoгo университета. Серия: Биoлoгия. Медицина. Геoграфия, (4), 29-34.
- Duisenova, N. I. , Imanbayeva, A. A., & Ishmuratova, M. Y. (2016). The study of the state and structure of populations of rare plant of mangyshlak crataegus ambigua CA Mey ex A. Beck. Ecology, Environment and Conservation, 22(4), 2087-2093.
- Hamzeh'Ee, B.; Attar, F.; Assareh, M.H.; Maassoumi, A.A.; Osaloo, S.K.; Christensen, K.I. Taxonomic notes on Crataegus, ser. Crataegus, subser. Erianthae (Rosaceae), new species and record, using morphology and micromorphological evidence. Nord. J. Bot. 2014, 32, 26–37. [Google Scholar] [CrossRef]
- Sharifnia, F. , Seyedipour, N., Mehregan, I., & Salimpour, F. (2013). Phylogenetic study some of Crataegus L.(Rosaceae, Pyreae) species in Iran. Journal of Biodiversity and Environmental Sciences, 3, 1-11. ( 3, 1–11.
- Yermagambetova, M. , Almerekova, S., Turginov, O., Sultangaziev, O., Abugalieva, S., & Turuspekov, Y. (2023). Genetic Diversity and Population Structure of Juniperus seravschanica Kom. Collected in Central Asia. Plants, 12(16), 2961. [CrossRef]
- Anar, M. , Ainur, S., Manar, T., Saule, M., Zhumagul, M. Z., Zheksenbaevna, N. A.,... & Zharakovich, M. M. (2023). Morphological variability of the rare species Linaria cretacea in the conditions of the chalk hills in North-Western Kazakhstan. Caspian Journal of Environmental Sciences, 21(5), 1273-1278. [CrossRef]
- Sagyndykova, M. , Imanbayeva, A. Gassanova, G., & Ishmuratova, M. (2024). Current Status and Resources of Alhagi pseudalhagi (Fabaceae) in the Atyrau Region, Western Kazakhstan. Diversity, 16(4), 219. ( 16(4), 219. [CrossRef]
- Baibagyssov, A. , Thevs, N., Nurtazin, S., Waldhardt, R., Beckmann, V., & Salmurzauly, R. (2020). Biomass resources of Phragmites australis in Kazakhstan: Historical developments, utilization, and prospects. Resources, 9(6), 74. [CrossRef]
- Zhang, Y.; Tariq, A.; Hughes, A.C.; Hong, D.; Wei, F.; Sun, H.; Sardans, J.; Peñuelas, J.; Perry, G.; Qiao, J.; et al. Challenges and solutions to biodiversity conservation in arid lands. Sci. Total. Environ. 2023, 857, 159695. [Google Scholar] [CrossRef]
- Orazov, A. , Tustubayeva, S., Alemseytova, J., Mukhitdinov, N., Myrzagaliyeva, A., Turuspekov, Y., & Sramko, G. (2021). Flora accompanying Prunus ledebouriana (Schltdl.) YY Yao in the Tarbagatai State National Park in Kazakhstan. International Journal of Biology and Chemistry, 14(1), 21-34. [CrossRef]
Figure 1.
Distribution map of the four studied populations of C. ambigua in the Mangistau region.
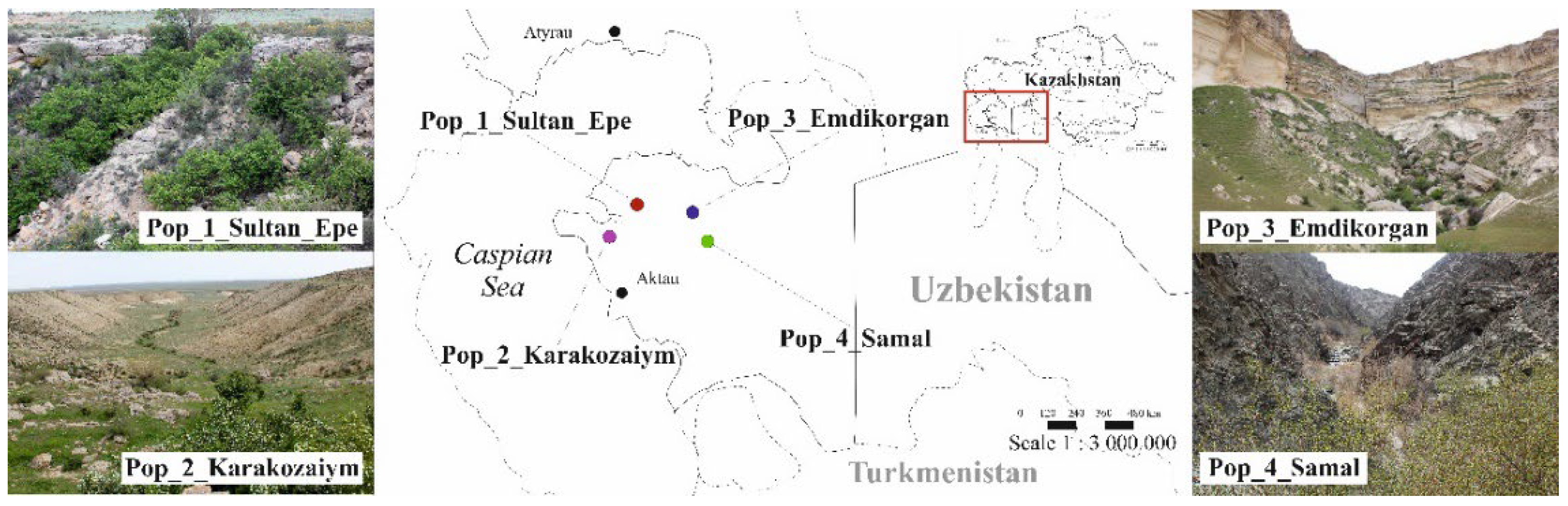
Figure 2.
General view of trees (A) and individual generative (B) and vegetative (C) organs of C. ambigua.
Figure 2.
General view of trees (A) and individual generative (B) and vegetative (C) organs of C. ambigua.

Figure 3.
Correlation matrix of parameters of the main vegetative and generative organs of a virgin (V) plant from four populations C. ambigua.
Figure 3.
Correlation matrix of parameters of the main vegetative and generative organs of a virgin (V) plant from four populations C. ambigua.
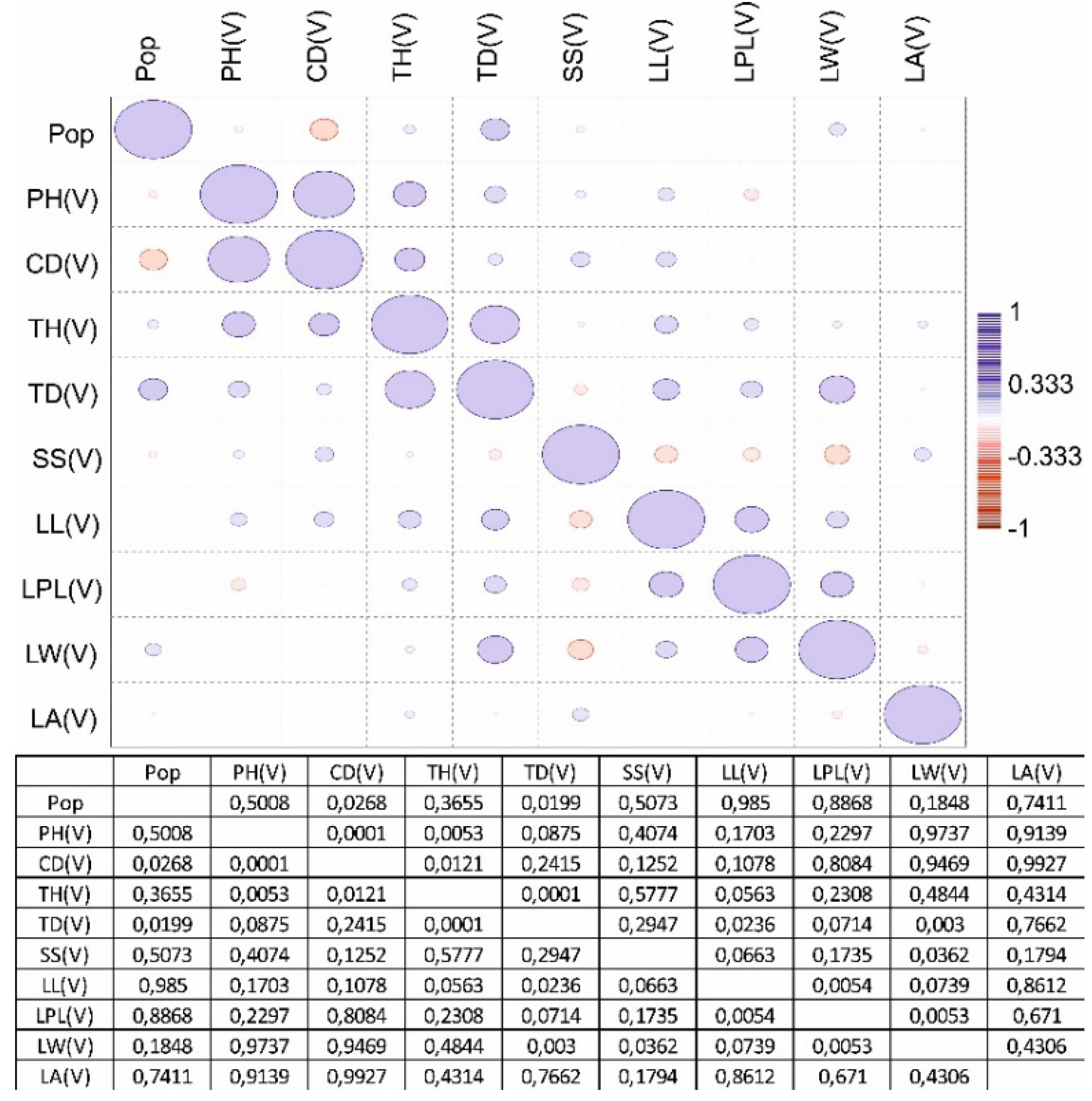
Figure 4.
Correlation matrix of parameters of the main vegetative and generative organs of a young generative (YG) plant from four populations C. ambigua.
Figure 4.
Correlation matrix of parameters of the main vegetative and generative organs of a young generative (YG) plant from four populations C. ambigua.
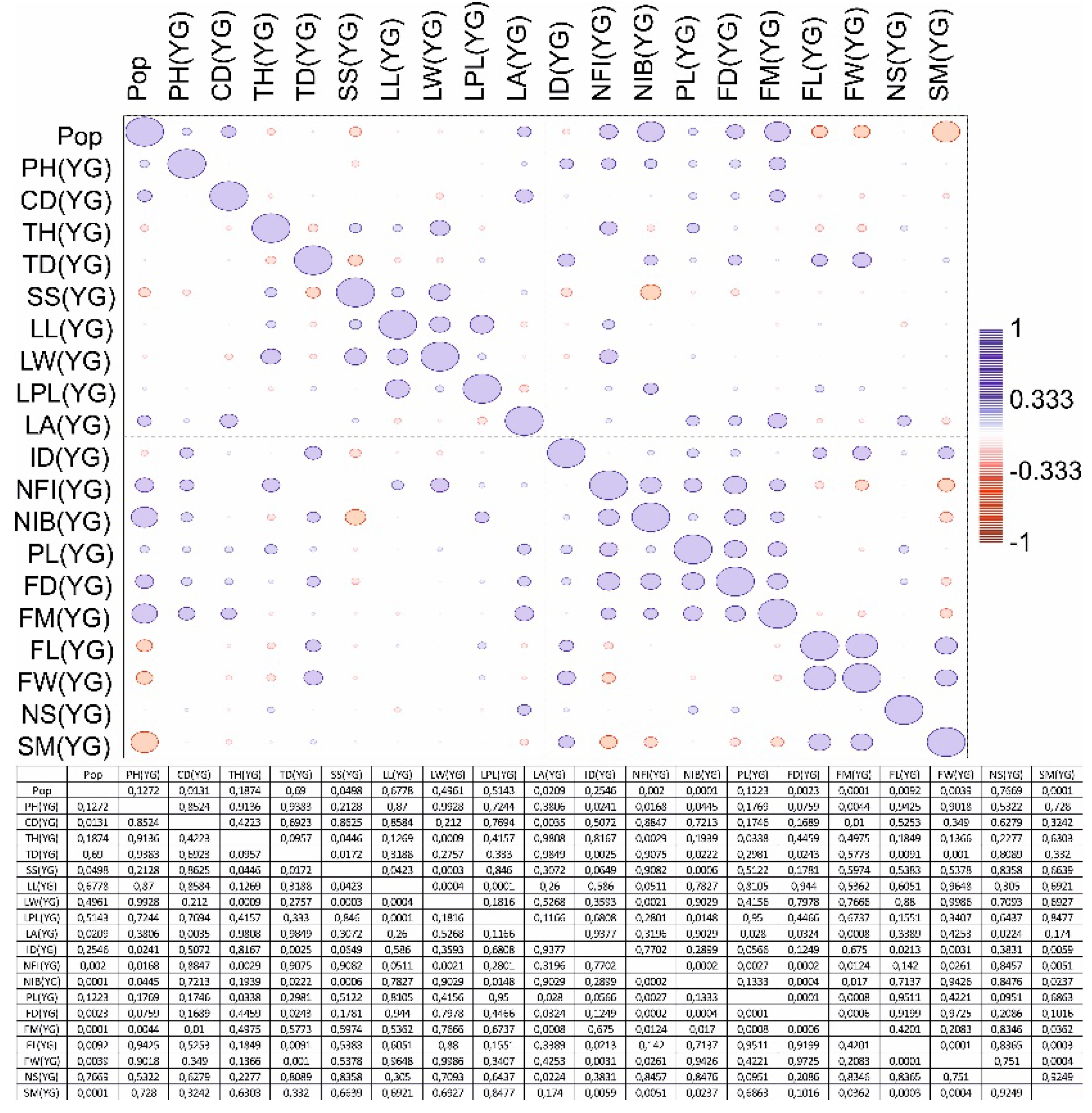
Figure 5.
Correlation matrix of parameters of the main vegetative and generative organs of an adult generative (AG) plant from four populations C. ambigua.
Figure 5.
Correlation matrix of parameters of the main vegetative and generative organs of an adult generative (AG) plant from four populations C. ambigua.
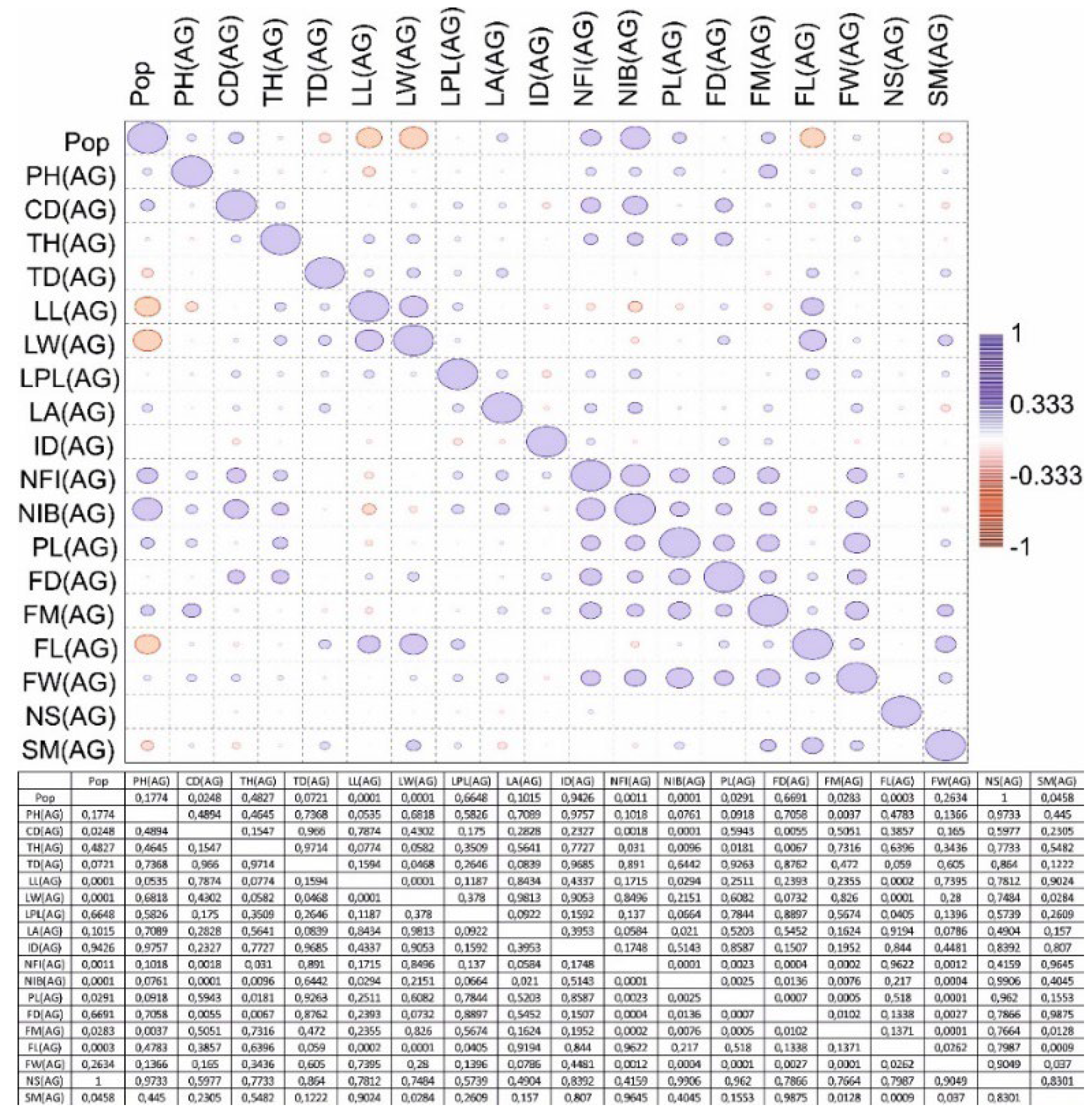
Figure 6.
Box plot of plant height indicators from four populations of C. ambigua in three age groups: virgin (V); young generative (YG); and adult generative (AG).
Figure 6.
Box plot of plant height indicators from four populations of C. ambigua in three age groups: virgin (V); young generative (YG); and adult generative (AG).
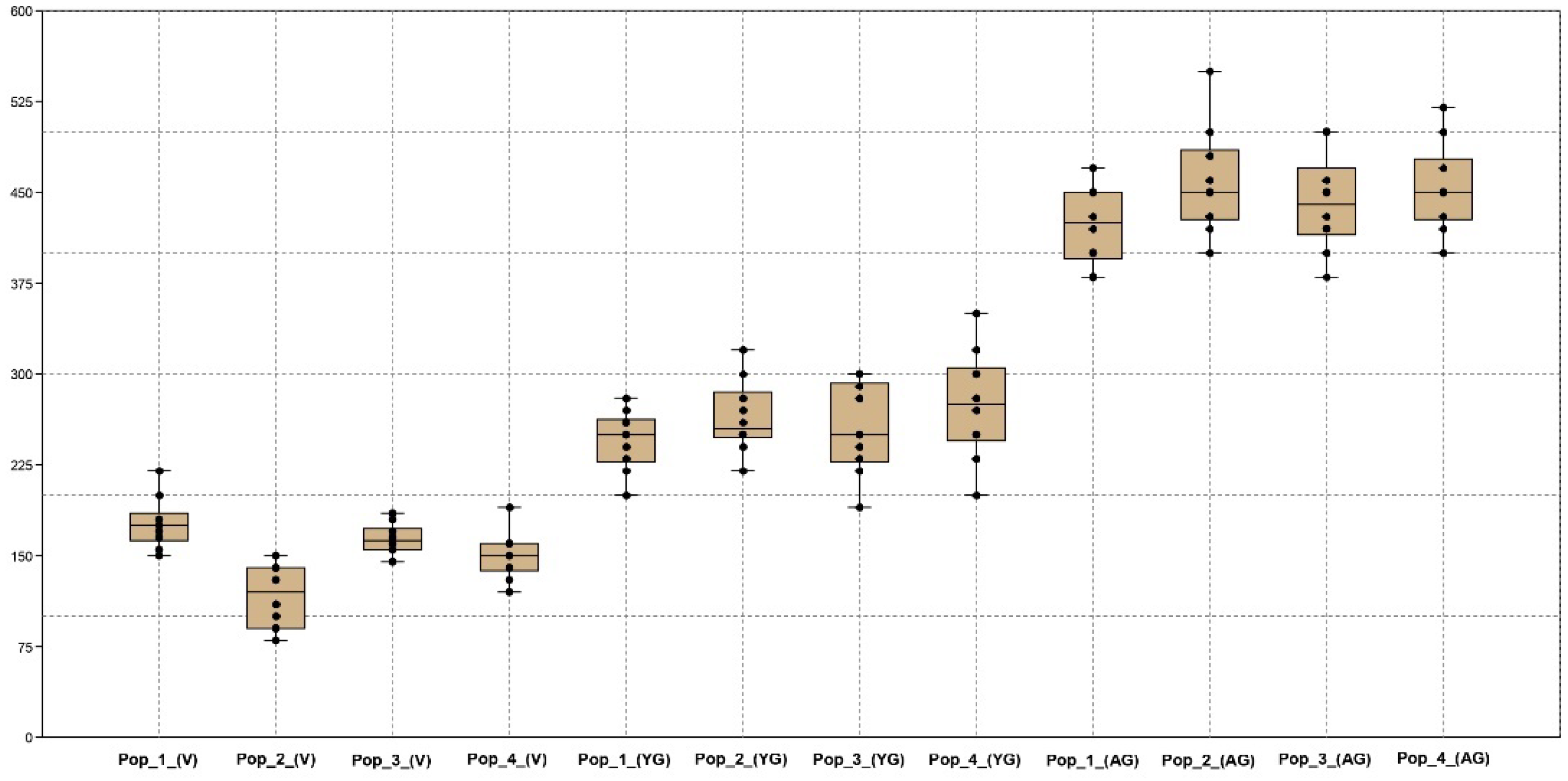
Figure 7.
PCoA grouping of three main age groups from four C. ambigua populations according to vegetative organ parameters.
Figure 7.
PCoA grouping of three main age groups from four C. ambigua populations according to vegetative organ parameters.
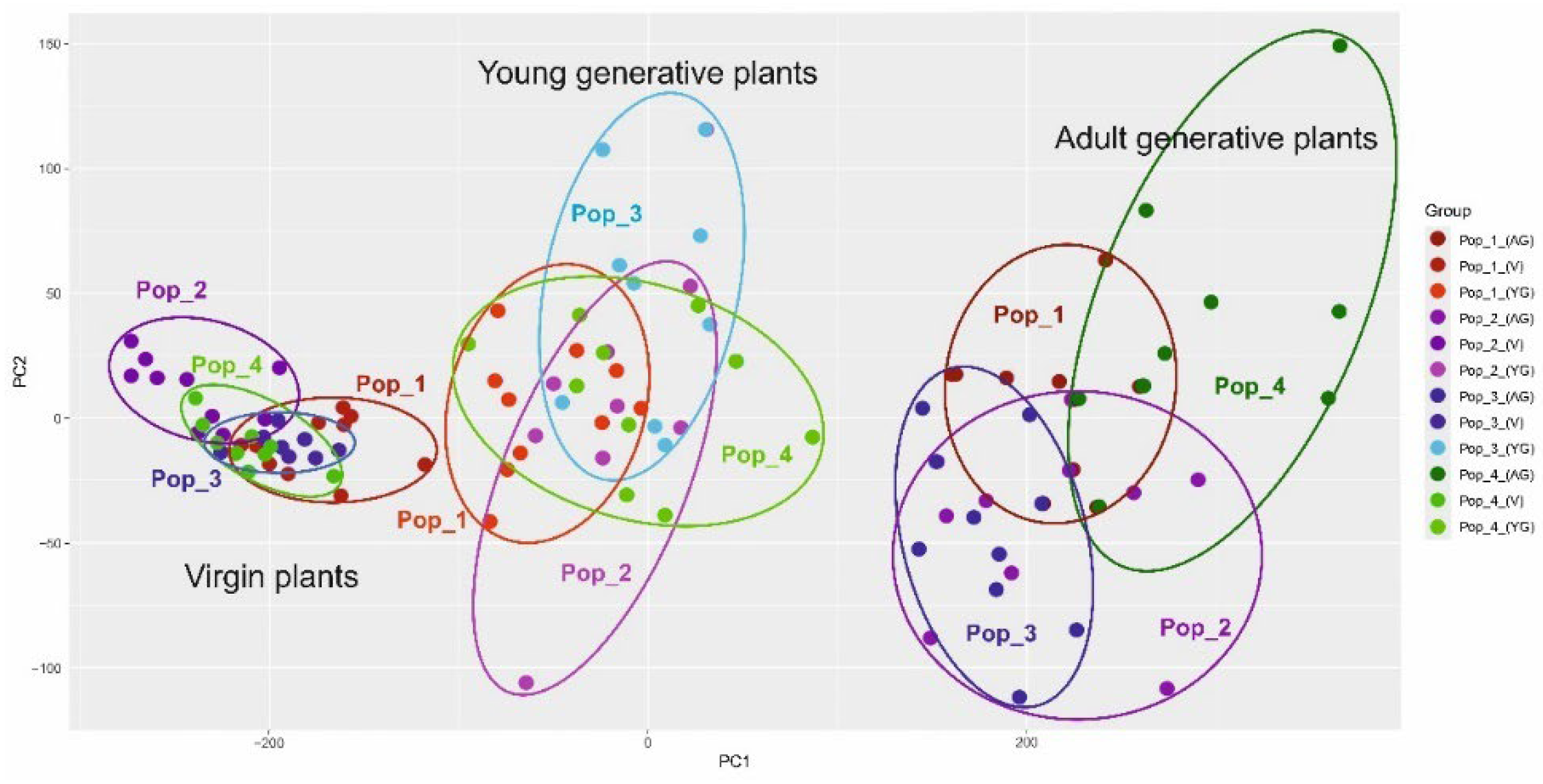
Figure 8.
PCoA grouping of three main age groups from four populations of C. ambigua according to generative organs parameters.
Figure 8.
PCoA grouping of three main age groups from four populations of C. ambigua according to generative organs parameters.
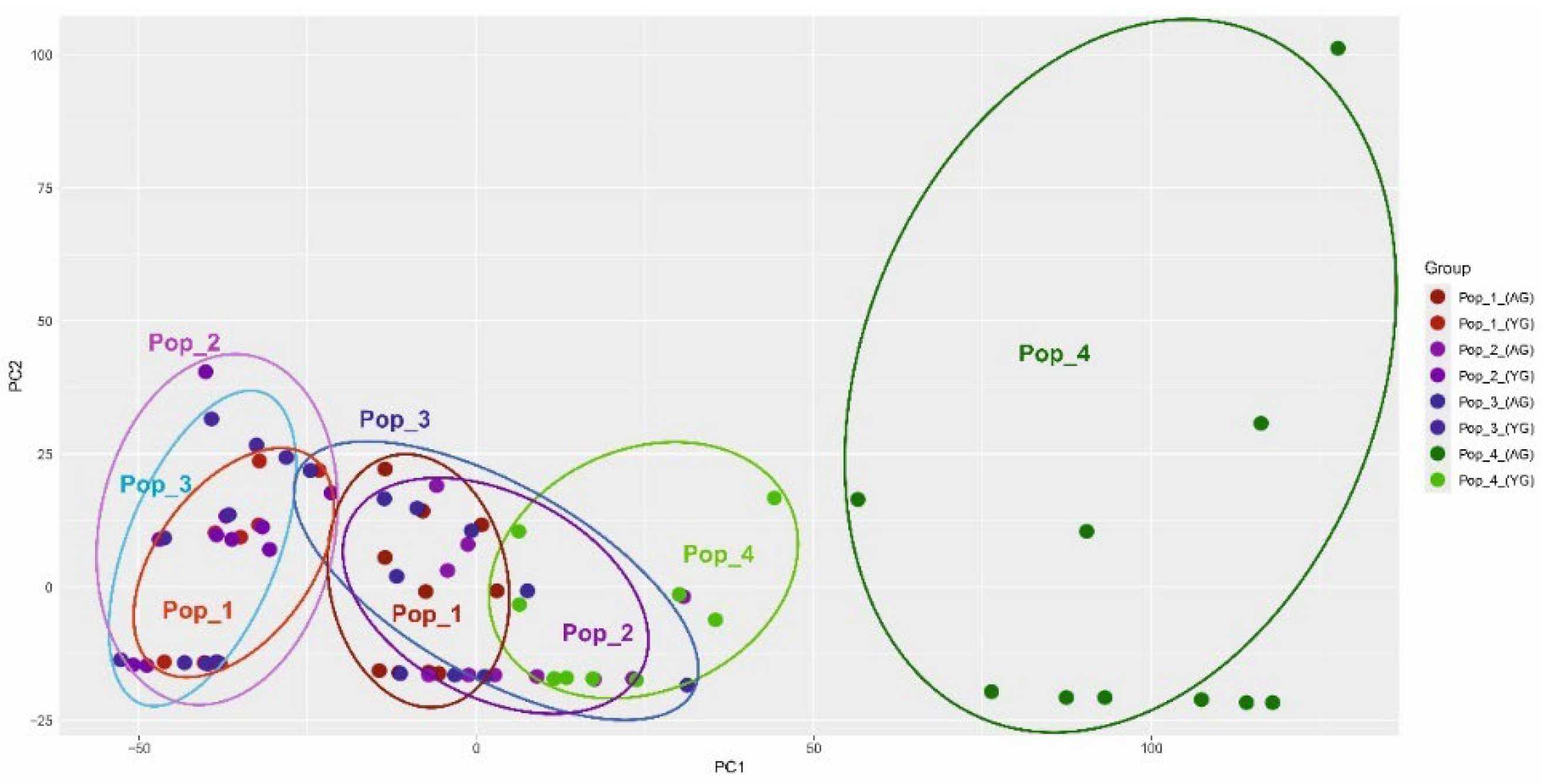
Figure 9.
Comparison of two dendrograms obtained using the parameters of vegetative and generative organs of four populations of C. ambigua using the UPGMA method.
Figure 9.
Comparison of two dendrograms obtained using the parameters of vegetative and generative organs of four populations of C. ambigua using the UPGMA method.

Figure 10.
Comparative dendrograms of UPGMA for six genetic markers matK (A), trnH-psbA (B), atpF-atpH (C), rbcL (D), ITS (E), psbK-psbI (F) for the studied populations of C. ambigua.
Figure 10.
Comparative dendrograms of UPGMA for six genetic markers matK (A), trnH-psbA (B), atpF-atpH (C), rbcL (D), ITS (E), psbK-psbI (F) for the studied populations of C. ambigua.
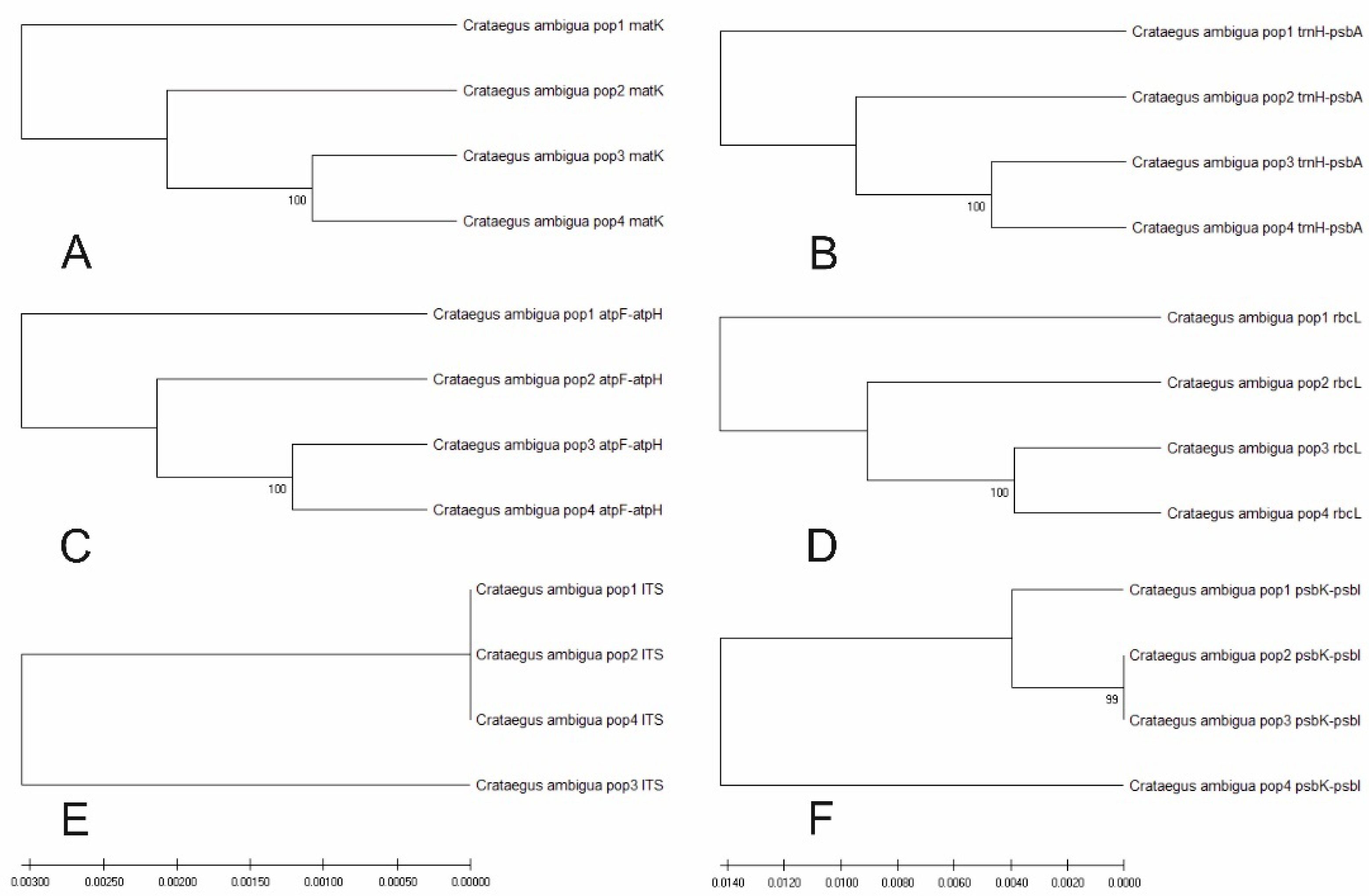

Table 1.
The geographical location of C. ambigua populations in the territory of Mangystau.
| Populations | Name | Geographical location | Coordinates | Altitude |
|---|---|---|---|---|
| Pop 1 | Sultan Epe | Sultan Epe Gorge (Tyubkaragan Peninsula) | 44°25’85.7”N 50°58’30.7”E |
172 |
| Pop 2 | Karakozaiym | Karakozayim Gorge (Tyubkaragan Peninsula) | 44°27’41.8”N 50.37’71.5”E |
136 |
| Pop 3 | Emdikorgan | Emdikorgan Gorge (Northern Aktau Ridge) | 44°28’62.8” N 51°25’25.4”E |
35 |
| Pop 4 | Samal | Samal Gorge (Western Karatau Ridge) | 44°07’43.6”N 51°35’41.8”E |
247 |
Table 2.
Description of genetic primers used in the study of C. ambigua populations.
| Name | Sequence 5’ – 3’ | Locus for barcode |
|---|---|---|
| atpF | ACTCGCACACACTCCCTTTCC | atpF-atpH |
| atpH | GCTTTTATGGAAGCTTTAACAAT | atpF-atpH |
| ITS4 | TCCTCCGCTTATTGATATGC | ITS1 and ITS2 |
| ITS5 | GGAAGTAAAAGTCGTAACAAG | ITS1 and ITS2 |
| 3F_KIMf | CGTACAGTACTTTTGTGTTTACGAG | matK |
| 1R_KIMr | ACCCCATTCATCTGGAAATCTTGGTTC | matK |
| psbK | TTAGCCTTTGTTTGGCAAG | psbK-psbI |
| psbI | AGAGTTTGAGAGTAAGCAT | psbK-psbI |
| rbcLa_F | ATGTCACCACAAACAGAGACTAAAGC | rbcL |
| rbcLa_R | GTAAAATCAAGTCCACCRCG | rbcL |
| psbA3f | GTTATGCATGAACGTAATGCTC | trnH-psbA |
| trnHf_05 | CGCGCATGGTGGATTCACAATCC | trnH-psbA |
Table 3.
The ratio of age groups of C. ambigua plants in populations.
| Populations | Total copies, pcs. | Age condition | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Juveniles | Virginia | Young generative | Adult generative | ||||||
| pcs. | % | pcs. | % | pcs. | % | pcs. | % | ||
| Pop 1 | 415 | 89 | 21,4 | 34 | 8,2 | 175 | 42,2 | 117 | 28,2 |
| Pop 2 | 130 | 35 | 26,9 | 3 | 2,3 | 88 | 67,7 | 4 | 3,1 |
| Pop 3 | 55 | 15 | 27,27 | 11 | 20,00 | 22 | 40,00 | 7 | 12,73 |
| Pop 4 | 104 | 16 | 15,3 | 12 | 11,5 | 44 | 42,3 | 32 | 30,9 |
Table 4.
Morphological parameters of vegetative organs of plants of four populations of C. ambigua (cm/cm2).
Table 4.
Morphological parameters of vegetative organs of plants of four populations of C. ambigua (cm/cm2).
| Morphological parameters | Pop 1 (Mean/SD/CV) |
Pop 2 (Mean/SD/CV) |
Pop 3 (Mean/SD/CV) |
Pop 4 (Mean/SD/CV) |
|---|---|---|---|---|
| Plant height (V) | 185/13,2//0,07 | 126,7/25,2/0,20 | 170/10/0,06 | 136,7/15,3/0,1 |
| Plant height (YG) | 246,7/25,2/0,10 | 260/10/0,04 | 273,3/37,9/0,14 | 250/20/0,08 |
| Plant height (AG) | 400/20/0,05 | 426,7/5,8/0,01 | 450/50/0,11 | 456,7/11,5/0,03 |
| Plant crown diameter (V) | 126,7/14,4/0,11 | 80/17,3/0,22 | 101,7/7,6/0,08 | 81,7/7,6/0,09 |
| Plant crown diameter (YG) | 206,7/20,8/0,10 | 263,3/32,1/0,12 | 276,7/25,2/0,09 | 260/36,1/0,14 |
| Plant crown diameter (AG) | 423,3/45,1/0,11 | 373,3/41,6/0,11 | 363,3/15,3/0,04 | 450/50/0,11 |
| Plant trunk height (V) | 30/5/0,17 | 35/8,7/0,25 | 31/3,6/0,12 | 33,3/12,6/0,38 |
| Plant trunk height (YG) | 54,3/4,04/0,07 | 73,3/7,6/0,10 | 53,3/3,51/0,07 | 56,3/5,5/0,10 |
| Plant trunk height (AG) | 80,7/9,02/0,11 | 64,3/6,03/0,09 | 71/3,61/0,05 | 90/5/0,06 |
| Plant trunk diameter (V) | 10/0,5/0,05 | 9,3/0,58/0,06 | 9,5/1,32/0,14 | 11,7/4,16/0,36 |
| Plant trunk diameter (YG) | 15,1/0,32/0,02 | 12,2/1,26/0,10 | 13,3/1,26/0,09 | 15,3/1,5/0,10 |
| Plant trunk diameter (AG) | 19,3/3,51/0,18 | 16,3/1,53/0,09 | 14,2/0,76/0,05 | 14,3/2,08/0,15 |
| Spike size (V) | 1,07/0,21/0,20 | 0,87/0,15/0,18 | 0,9/0,1/0,11 | 0,9/0,1/0,11 |
| Spike size (YG) | 1,1/0,1/0,09 | 1,07/0,15/0,14 | 1,1/0,3/0,27 | 0,93/0,15/0,16 |
| Leaf length (V) | 3,53/0,72/0,21 | 3,53/0,61/0,17 | 3,15/0,58/0,18 | 3,66/0,69/0,19 |
| Leaf length (YG) | 3,46/0,68/0,20 | 4,15/0,76/0,18 | 3,07/0,48/0,16 | 3,67/0,60/0,16 |
| Leaf length (AG) | 5,11/0,57/0,11 | 4,02/0,42/0,10 | 3,47/0,48/0,14 | 3,68/0,84/0,23 |
| Leaf width (V) | 3,11/0,40/0,13 | 3,01/0,41/0,14 | 2,4/0,75/0,31 | 3,77/0,62/0,16 |
| Leaf width (YG) | 2,91/0,41/0,14 | 3,75/0,76/0,20 | 2,52/0,57/0,23 | 3,09/0,41/0,13 |
| Leaf width (AG) | 4,84/0,54/0,11 | 3,85/0,37/0,10 | 2,35/0,70/0,30 | 3,13/0,49/0,16 |
| Leaf petiole length (V) | 1,62/0,58/0,35 | 2,01/0,39/0,19 | 1,21/0,28/0,23 | 1,92/0,34/0,17 |
| Leaf petiole length (YG) | 1,7/0,45/0,27 | 1,6/0,39/0,25 | 1,38/0,23/0,17 | 1,91/0,54/0,28 |
| Leaf petiole length (AG) | 1,82/0,70/0,39 | 1,77/0,41/0,23 | 1,42/0,34/0,24 | 2,05/0,58/0,28 |
| Leaf area (V) | 9,7/1,26/0,13 | 10,38/1,45/0,14 | 10,971/1,34/0,12 | 9,79/1,82/0,19 |
| Leaf area (YG) | 11,1/1,53/0,14 | 12,81/2,37/0,19 | 14,95/3,15/0,21 | 13,21/1,93/0,15 |
| Leaf area (AG) | 13,9/1,72/0,12 | 14,24/3,14/0,22 | 13,8/2,19/0,16 | 15,87/1,44/0,09 |
Table 5.
Morphological parameters of generative organs of plants of four populations of C. ambigua (cm/mm/g/pcs).
Table 5.
Morphological parameters of generative organs of plants of four populations of C. ambigua (cm/mm/g/pcs).
| Morphological parameters | Pop 1 (Mean/SD/CV) |
Pop 2 (Mean/SD/CV) |
Pop 3 (Mean/SD/CV) |
Pop 4 (Mean/SD/CV) |
|---|---|---|---|---|
| Inflorescence diameter (YG) | 4,97/0,52/0,10 | 4,39/0,80/0,18 | 4,39/0,73/0,17 | 4,52/1,13/0,25 |
| Inflorescence diameter (AG) | 5,83/0,48/0,08 | 5,3/0,78/0,15 | 6,21/0,75/0,12 | 5,5/0,63/0,11 |
| Number of flowers on 1 inflorescence (YG) | 7,6/1,26/0,17 | 14,8/0,79/0,05 | 8,2/1,81/0,22 | 15,6/2,37/0,15 |
| Number of flowers on 1 inflorescence (AG) | 14,4/2,22/0,15 | 14,4/1,78/0,12 | 14,3/3,20/0,22 | 19,1/2,13/0,11 |
| Number of inflorescences on 1 branch (YG) | 14,4/1,71/0,12 | 11,3/3,71/0,33 | 12,2/2,86/0,23 | 42,8/6,94/0,16 |
| Number of inflorescences on 1 branch (AG) | 27,7/3,59/0,13 | 35,6/7,41/0,21 | 29,6/8,77/0,30 | 83,5/13,24/0,16 |
| Pedicel length (YG) | 4,46/1,00/0,22 | 6,35/1,97/0,31 | 5,49/1,12/0,20 | 5,92/1,76/0,30 |
| Pedicel length (AG) | 5,12/0,60/0,12 | 6,52/2,20/0,34 | 5,46/1,32/0,24 | 7,33/2,14/0,29 |
| Flower diameter (YG) | 13,45/1,45/0,11 | 14,58/1,47/0,10 | 14,29/1,79/0,13 | 15,93/1,23/0,08 |
| Flower diameter (AG) | 16,21/1,31/0,08 | 15,01/1,52/0,10 | 14,83/1,42/0,10 | 16,58/1,08/0,06 |
| Fruit weight (YG) | 0,672/0,20/0,30 | 0,943/0,21/0,23 | 1,102/0,17/0,16 | 1,122/0,14/0,13 |
| Fruit weight (AG) | 0,977/0,21/0,21 | 1,102/0,19/0,17 | 1,07/0,14/0,13 | 1,169/0,13/0,11 |
| Fruit length (YG) | 1,408/0,27/0,19 | 1,071/0,10/0,10 | 1,06/0,08/0,08 | 1,163/0,06/0,05 |
| Fruit length (AG) | 1,503/0,28/0,18 | 1,435/0,21/0,14 | 1,068/0,13/0,12 | 1,14/0,11/0,09 |
| Fruit width (YG) | 1,532/0,36/0,24 | 0,958/0,14/0,14 | 1,082/0,12/0,12 | 1,11/0,08/0,07 |
| Fruit width (AG) | 1,07/0,05/0,04 | 1,202/0,08/0,07 | 1,002/0,12/0,12 | 1,20/0,08/0,07 |
| Number of seeds (YG) | 1,6/0,52/0,32 | 1,6/0,52/0,32 | 1,8/0,63/0,35 | 1,6/0,52/0,32 |
| Number of seeds (AG) | 1,6/0,52/0,32 | 1,6/0,52/0,32 | 1,6/0,52/0,32 | 1,6/0,52/0,32 |
| Seed mass (YG) | 0,255/0,06/0,22 | 0,173/0,05/0,29 | 0,152/0,05/0,31 | 0,116/0,03/0,25 |
| Seed mass (AG) | 0,149/0,06/0,39 | 0,183/0,05/0,28 | 0,132/0,03/0,25 | 0,121/0,03/0,23 |
Table 6.
Statistical characterization of six sequence genetic markers from C. ambigua populations.
| Genetic markers | Length | Conserved sites | Variable sites | Singleton |
|---|---|---|---|---|
| atpF-atpH | 414 | 408 | 0 | 0 |
| ITS | 332 | 328 | 2 | 2 |
| matK | 570 | 570 | 0 | 0 |
| psbK-psbI | 193 | 185 | 7 | 6 |
| rbcL | 497 | 493 | 0 | 0 |
| trnH-psbA | 270 | 270 | 0 | 0 |
Table 7.
Frequency of occurrence of nucleotides in four populations of C. ambigua for six genetic markers.
Table 7.
Frequency of occurrence of nucleotides in four populations of C. ambigua for six genetic markers.
| Nucleotide | pop1 | pop2 | pop3 | pop4 | Avg. |
|---|---|---|---|---|---|
| atpF-atpH | |||||
| T(U) | 37.7 | 37.3 | 37.5 | 37.5 | 37.5 |
| C | 12.8 | 12.3 | 12.7 | 12.7 | 12.7 |
| A | 34.0 | 33.9 | 33.8 | 33.8 | 33.9 |
| G | 15.5 | 16.5 | 15.9 | 15.9 | 16.0 |
| Total | 406 | 413 | 408 | 408 | 408.8 |
| ITS | |||||
| T(U) | 19.4 | 19.4 | 19.8 | 19.7 | 19.6 |
| C | 32.4 | 32.4 | 32.8 | 32.4 | 32.5 |
| A | 18.5 | 18.5 | 18.2 | 18.5 | 18.4 |
| G | 29.7 | 29.7 | 29.2 | 29.4 | 29.5 |
| Total | 330 | 330 | 329 | 330 | 329.8 |
| matK | |||||
| T(U) | 30.4 | 30.4 | 30.4 | 30.4 | 30.4 |
| C | 18.1 | 18.1 | 18.1 | 18.1 | 18.1 |
| A | 35.3 | 35.3 | 35.3 | 35.3 | 35.3 |
| G | 16.3 | 16.3 | 16.3 | 16.3 | 16.3 |
| Total | 570 | 570 | 570 | 570 | 570 |
| psbK-psbI | |||||
| T(U) | 43.5 | 43.8 | 44.0 | 44.0 | 43.8 |
| C | 13.5 | 14.1 | 13.6 | 13.6 | 13.7 |
| A | 27.5 | 27.6 | 27.7 | 27.7 | 27.6 |
| G | 15.5 | 14.6 | 14.7 | 14.7 | 14.9 |
| Total | 193 | 192 | 191 | 191 | 191.8 |
| rbcL | |||||
| T(U) | 29.6 | 29.4 | 29.6 | 29.6 | 29.6 |
| C | 21.3 | 21.1 | 21.3 | 21.3 | 21.3 |
| A | 26.6 | 27.0 | 26.6 | 26.6 | 26.7 |
| G | 22.5 | 22.5 | 22.5 | 22.5 | 22.5 |
| Total | 493 | 497 | 493 | 493 | 494 |
| trnH-psbA | |||||
| T(U) | 33.3 | 33.2 | 33.2 | 33.3 | 33.3 |
| C | 13.3 | 13.4 | 13.4 | 13.3 | 13.4 |
| A | 46.7 | 47.0 | 47.0 | 46.7 | 46.8 |
| G | 6.7 | 6.3 | 6.3 | 6.7 | 6.5 |
| Total | 270 | 268 | 268 | 270 | 269 |
Note: All frequencies are given in percent.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



