
Submitted:
22 April 2024
Posted:
23 April 2024
You are already at the latest version
Abstract
The use of endophytic microbes is increasing in commercial agriculture. This review will describe the endophyte-plant relationship and the fungi and bacteria involved. Their abilities to alleviate biotic (diseases and pests) and abiotic stresses (drought, salt and flooding), remediate pollution and increase photosynthetic capabilities will be described. Their mechanisms of action will be elucidated. These frequently result in increased plant yields. Finally, methods and practices for formulation and commercial use will be described.
Keywords:
holobiont
; plant-microbe interactions
; biocontrol
Introduction
Many microorganisms are becoming important components of agricultural production systems. There are two reasons for this. First, there have been improvements in our understanding of mechanisms of action and the benefits they provide. Second, there have been improvements in formulations and in the ability to integrate biologicals into agricultural systems. Chemical pesticides and fertilizers are usually used, and so biologicals must be integrated into this chemical world.
This chapter will (A) examine the mechanisms of action, (B) describe the benefits to plants and agriculture, and (C) consider integration of biologicals, including microbe-microbe mixtures and integration of biologicals into chemically based agriculture.
Part 1. Holobionts and Benefits to Plants and the Environment
Holobionts and Endophytes
Any organism does not exist in isolation, but instead is an association with numerous other organisms. This aggregation is termed a holobiont [1]. In some cases, bacteria or fungi internally colonize plant roots, and frequently, but not always, are located only there. However, some Rhizobial endophytes ascend into the upper portions of plants [2] and Clonostacys rosea colonize plant leaves when applied topically [3]. Some of these may have little effect on plants, be detrimental (e.g., pathogens) or may have beneficial effects. We have designated plants colonized by beneficial microbes as enhanced plant holobionts (EPHs) [4]. Some of the most useful are endophytes. Endophytes are fungi or bacteria that live within the interior of the plant and cause unapparent and asymptomatic infection [5]. The organisms persist and live for at least the life of an annual crop and are true symbionts—the plants supply nutrients and a habitat while the microbes provide numerous benefits to the plants [6,7]. Trichoderma added as seed treatments grow rapidly from the seed onto the radical and then grow and become established in the interior of the root. They usually are limited to the root and do not become established in the above-ground portions of the plant. Salicycilic acid is apparently required for localization of Trichoderma to roots [8]. Many other fungi and bacteria are endophytic and can potentially be used for seed treatment. These include fungi in the genera Trichoderma [9,10,11], Aspergillus [12], Penicillium [13], Clonostachys [14], Piriformaspora [15], and Yarrowia lipolutoca [16], as well as bacteria in the genera Pseudomonas [17] and Bacillus [17,18], and in the family Rhizobiacae [2], Poveda et al. [19] provides a list of more than 50 endophytic fungi.
Benefits to Plants and the Environment
This section will describe the advantages that microbes provide to plants but will address mechanisms of action later. Many of the mechanisms are similar regardless of the benefits conferred. However, many endophytes cause systemic benefits to plants. For example, many of them colonize only roots, but provide benefits throughout the plant. For example, Trichoderma strains usually colonize only roots but still can protect leaves from powdery mildews and anthracnose.
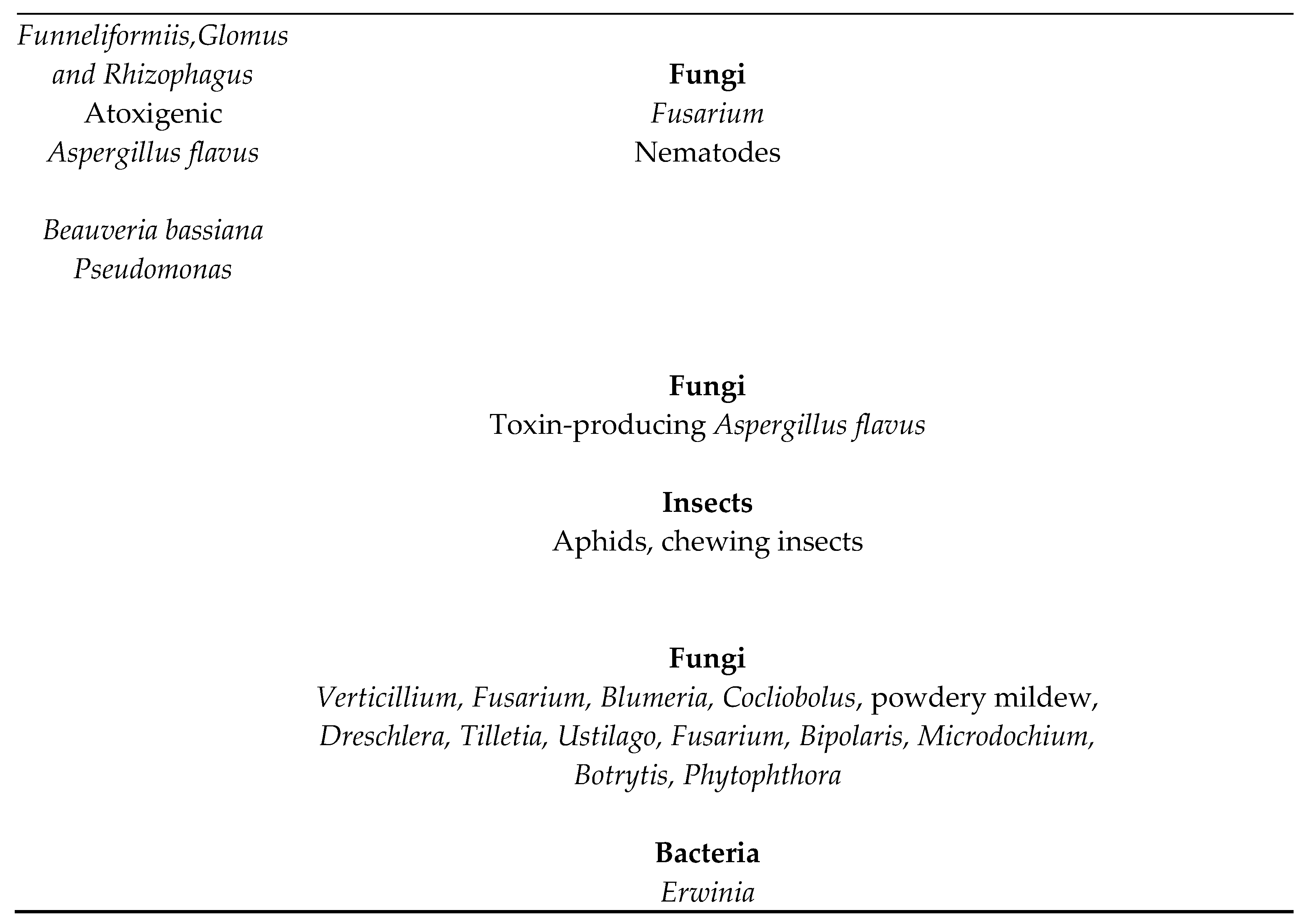
Control of pests, including diseases, insects and even viruses is provided by microorganisms. Many, but not all, are endophytes. For example, there are more than 100 products based on Trichoderma for control of diseases [20]. Nematodes are controlled by specific strains of Trichoderma [21]. While numerous pathogens or pests are controlled, most strains control only a few pests so it is important to determine which products or strains control which pests. Fungi, Oomycetes, nematodes, and insects are controlled by different microbes. Table 1 provides a summary of some of these.
Some fungi, especially Beauveria and Metarhizium control insects, and these fungi may be endophytic [22]. Surprisingly, even Trichoderma have been reported to have insecticidal activity [19,23]. It must be emphasized that not all members of a species or genus have activity against all pests listed—for example, we have screened thousands of Trichoderma strains and found only one that gave effective nematode control. The effective strain was an atrovide, but other strains of that species were ineffective. The author expects that only specific strains would be effective against insects.
However, many, if not all, pathogens and pests can be controlled by microorganisms and the list continues to grow rapidly. Microbial pest control is becoming an integral part of agricultural practice.
Strain Selection
Only some strains of any bacteria or fungus will provide adequate performance in agriculture. The author has screened thousands of Trichoderma strains but only four were advanced to actual products. Strain T22 is a good example. It was originally produced via protoplast fusion [24]. Its parents were a strain with good rhizosphere competence (T95), and is some soils, it provided good protection against seed rots. However, in other soils it was ineffective; it was discovered that in soils where it was ineffective that available iron was limiting. In this situation, bacteria destroyed the hyphae due to their production of iron-chelating compounds (siderophores) [25]. The other parent (T12) was adapted to the low iron conditions, probably by producing its own siderophores. More than 100 individual progeny were produced and T22 was one of them. Analysis of this strain suggested that its genome was primarily of the T12 genome with some segments of T95 [26]. This was recently confirmed, genetic sequence analysis showed that the genome of T22 was primarily composed to the T12 genome but with patches of T95 [27]. This strain is widely used commercially and in research—a Biosis search revealed more than 100 studies that featured this strain.
Evolution of Trichoderma
Trichoderma strains are known for their abilities to produce extracellar enzymes and to parasitize other fungi. In its earliest forms, it was a parasite on wood-decomposing fungi, and then became a soil-inhabiting saprophyte. In the Creataceous-Paloeogene extinction event it acquired the genes for hydrolytic genes by a massive horizontal transfer of genes from other fungi [28,29]. Thus, the events in the natural evolution of these fungi are similar to that which occurred as T22 was developed. These mycoparastic fungi are well suited to gene transfer events since the internal compartments of the prey and parasitic are in close proximity. Gene transfer events are made even more likely because each cell of these fungi contain as many as 50 separate nuclei and so gene transfer becomes likely. This is a tremendous driver of diversity and adaptation since as environmental conditions change different nuclei may contain genes to exploit the new conditions [26].
Mitigation of Abiotic Stresses
Drought
T. afroharzinaum (formerly T. harzianum) reduced effects of drought in greenhouse tomatoes and in field grown corn [33,34] and in wheat [35]. Mycorrhizal fungi had similar effects on watermelons [36]. An endophytic strain of Alternaria provides resistance to drought in tomatoes [37]. Similarly, a strain of Curvularia mitigated effects of drought on rice [38]. Root colonization by a strain of Pseudomonas chlororaphis induced resistance to drought in Arabisopsis [39]; another strain of the bacterium induced drought tolerance in soybean [40]. Similarly, treatment of plants with either Achromobacter piechaudii or Pseudomonas putida reduced drought symptoms in tomato [41].
Salinity
Trichoderma strains alleviated effects of salt damage in tomatoes [34,42]. Similarly, Bacillus [43], Piriformaspora indica [44,45], Dietzia [46] and Yarrowia [47] all minimized the effects of salinity stress on plants. Application of Achromobacter piechaudii minimized effects of salinity as well drought [41].
Flooding
Endophytes can also reduce symptoms of flooding. In our trials, application of a mixture of two strains of Trichoderma reduced damage during flooding conditions. Many of the bacteria and fungi described in the previous two sections also alleviated flood damage.
Enhanced Mineral Nutrition
Arbuscular mycorrhizal fungi are obligate plant symbionts. They form cellular structures between plant cell walls and the plasmalemma. They also form extensive networks of hyphae in the soil and that grow on plant roots. These networks form within roots of the same plants and even between different plants, even between plants of different species. These networks accumulate phosphorus and other nutrients and transfer this to the plants. This enhances mineral nutrition of plants and thereby enhances plant productivity [48].
Trichoderma and other endophytic organisms can also improve plant nutrient status. Some metallic nutrients are poorly soluble and therefore unavailable to plants. Trichoderma strains are able to solubilize essential nutrients [49] and they also can enhance nitrogen uptake [50]. Pseudomods and Trichoderma produce siderophores, which are compounds with high affinity for iron and other metallic plant nutrients [51,52].
Competition
Competition for space or nutrients has been suggested as a mechanism for biocontrol, but usually without good evidence. However, in one case this is the mechanism. Aspergillus flavus infects cotton bolls and other crops. In cotton, it frequently colonizes wounds made in the boll by boll worms. In the boll it produces aflatoxin which is a strong carcinogen. The FDA only allows 20 ppb of the toxin (www.canr.msu.edu/news/trending-aflatoxins. Peter Cotty and his colleagues searched for naturally-occurring stains of A. flavus and used high levels of the atoxigenic strain to Inoculate fields with high levels of the atoxigenic strain. This resulted in good control of aflatoxin by competitively colonizing the infection court. Thus, the atoxigenic form functioned as an endophyte and resulted in good reductions or elimination of aflatoxin production [53].
Alleviation of Environmental Pollutants
Pollution of soils by, for example, heavy metals, organic compounds or other pollutants can adversely affect plant growth and pose health risks to humans. Staplococcus arletta alleviated chromate toxicity in sunflowers by suppressing the uptake of the toxicant and by reducing the hexavalent form to the less toxic trivalent form [35]. However, beneficial effects do not always occur. In a commercial planting of poinsettia, a report was received that said after application of T. afroharzianum all the plants died, while in its absence, the plants were healthy. It was discovered that the plants were grown in sewage sludge with a high level of chromium. T. afroharzianum increased the uptake of the metal to toxic levels.
The ability of a gibberellin-producing strain of Penicillium janthinellumI minimized effects of aluminum toxicity in tomato. Salicylic acid was upregulated in the presence of the fungus and cell membranes were less damaged [54].
Cyanides are frequently present in mine tailings. Endophytic strains of Trichoderma afroharzianum were able to take up and degrade metallocyanins [55]. Water used in olive processing become contaminated with toxic polyphenols. Strains of Trichoderma were able to degrade these toxic compounds [55].
Soils polluted with oily wastes from petroleum drilling or other industrial processes are serious environmental hazards. Plants colonized by Pseudomonas putida were able to grow in soils contaminated by oil and the oil was eventually broken down to less toxic materials. The bacterial strains themselves probably did not break down the oil but they permitted roots to grow by increasing plant stress tolerance and other soil organisms growing on the roots degraded the oil [56].
Enhanced Photosynthesis
All the benefits described require fixed carbon and energy in the form of photosynthate. This must come from photosynthesis [57]. Therefore, for the benefits to EPHs to occur photosynthetic rates need to be increased. Plants that contain endophytes frequently are greener [52]. There are anatomical changes in leaves of plants colonized by endophytes, including higher density of smaller stomata, thicker palisade parenchyma, and larger intercellular spaces in the mesophyll. The modifications in leaf functional anatomical traits affected gas exchanges [58]; in fact, starting from the reproductive phase, the rate of leaf net photosynthesis (NP) was higher in inoculated compared to control plants. These data are consistent with the better maximal PSII photochemical efficiency observed in inoculated plants; conversely no difference in leaf chlorophyll content was found. The PGPM-induced changes in leaf structure and photosynthesis lead to an improvement of plant growth [58].
In plants treated with T. asperellum, many photosynthetic genes were up-regulated [59]. In EPHs, levels of photosynthetic pigments and proteins were increased [60,61,62]. Rhizobia also demonstrates such effects [2]. There are many reports of up-regulation of genes encoding for photosynthetic functions and chlorophyll concentrations were increased. These are summarized in [63].
Enhanced Yield and Plant Growth
Many endophytic microbes enhance plant growth. Rhizobia are used around the world to supply soluble nitrogen to legumes and this increase plant growth, especially under conditions of low nitrogen fertilizer. Piriformasora indica inoculation results in increased plant growth and development, in part through reprogramming genes involved in various physiological processes [44]. In watermelon, mycorrhizae increased plant growth, in part through inactivation of reactive oxygen species (ROS) [36]. There are many reports of the ability of Trichoderma to increase plant growth, including [52] and these advantages across several endophytes has been summarized [4]. A summary of plant growth promotion by mycorrhizae, Trichoderma, Bacillus and Peudomonas, together with the traits associated with them is available [64].
Mechanisms
Antibiosis
In some cases, antibiosis is part of the mechanisms of biocontrol. For example, phenazines have been implicated in the biocontrol of late blight caused by Gaeumannomyces [65]. More than 40 antibiotic compounds have been reported as being produced by Trichoderma [66] but other mechanisms may be more important in biocontrol. It was thought that antibiosis was critical for control of damping off by Trichoderma [67] but in later studies, neither antibiosis nor mycoparaitism were correlated with disease control, instead the ability to control the disease was tightly linked to systemic resistance [7]. However, Trichoderma produces substances that have antibiotic or disease preventing activity including terpenes, non-ribobomal peptides, ketones and terpenes [20].
Parasitism
For many years, mycoparitism was considered an important factor in biological control by Trichoderma. More recently attention has focused on systemic resistance as a more important mechanism [7]. However, Ampelomces quisqualis is an obligate pathogen of powdery mildews and this is the active ingredient is Aq10, a commercial biocontrol product [68] and Coniothrium minitans is a parasite of sclerotia of Sclerotium spp. [69]. In these two cases, parasitism is the sole mechanism of action.
Antifeedants
Control of insect pests may be through the production of microbial metabolites that act as antifeedants. In some cases of insect control by Trichoderma this is the mechanism [19]. Root and seed feeding dipterans detect metabolites of roots, seeds, and volatile exudates from microbial activity on these plant parts and preferentially lay eggs in this area. The emerging larvae feed on the plants, thus severely damaging them. A seed or soil treatment with Cheatomium globosum alters the microbial community so they are no longer attractive to the female dipterans and damage is lessened [70,71].
Systemic Reactions and Differentially Expressed Genes and Proteins
Many, and probably most, endophytes induce systemic reactions in plants. Mention has already been made of control of foliar diseases when the endophyte is located only in the root. Some examples of microbes that induce systemic resistance include Trichoderma [4], Pseudomonas [72], Rhizobium [2], Bacillus [64] and Piraformaspora indica [73].
Elicitors
Endophytic organisms may be located in the outer layers of the root [74,75], and produce signaling compounds that interact usually with receptors in the plasmalemma [4]. There are numerous types of such molecules, which are designated as effectors or affectors also known as microbial associated recognition patterns. They include volatile compounds such as 6-penyl-α-pyrone and 1-octen-3-ol, other small molecular weight molecules such as harzianic acid, heptelic acids, koningic acid, chitoologosaccharides and glucans [4,76]. Proteins may also function as effectors—small protein 1 is produced by T. virens and is essential for induced resistance in maize [77]. Peptabiols are linear peptides that induce plant defense responses [78]. Specific hydrophobins also function as effectors and even sRNAs may play a role [79]. With Trichoderma, release of sRNA leads to upregulation of plant cytoplasmic nucleotide-binding site leucine-rich receptors. After Trichoderma recognition, the plant genome is reprograms genes involved is defense and development and activates antioxidant systems phytohormone that leads to increased systemic resistance [28].
Once affectors interact with cell membranes then systemic signals are produced that are translocated throughout the plant via map kinases [28]. Many, or perhaps most, plant symbionts alter gene regulation. For example, in rice seedlings there were 301 transcripts that were up-regulated, and 420 were related to photosynthetic functions [59]. In P. indica, disease resistance requires reprogramming of genetic components [44], while in plants colonized by Pseudomonas activation of plant defense systems occur [72].
Endophytes have novel methods to induce resistance to biotic and abiotic stresses. As noted earlier, active defense systems involve a cost to the plant, since the production of defense-related compounds and systems are expensive in terms of their energy and fixed carbon requirements. Plants have evolved systems to avoid these requirements. Gene priming is a system whereby genes and proteins involved in defense reactions do not have to be continually produced. Instead of continual production of these substances, changes in the upstream regulatory portions of genes, including alterations of the cellular chromatin, by modification of histones, or by DNA methylation [80,81,82]. Consequently, in the absence of stress, the defensive materials are not produced, but when stressful conditions occur, the modifications in the upstream regulatory regions are activated quickly.
This induced resistance is designated priming, which can be defined as “an adaptive strategy improving plant defense capacity whereby an initial stimulus activates the physiological, transcriptional, metabolomic and epigenetic mechanisms that enable the plant to respond more rapidly and/or efficiently to subsequent exposure to a biotic or abiotic stress” [28]. Since priming occurs via epigentic changes in the DNA, it can be passed to offspring, thus conferring heritable resistance [28,83].
Reactive Oxygen Species (ROS)
When plants (or any other organism) are under stress, deleterious substances are frequently produced. Among the most damaging are reactive oxygen species and they inactivate proteins, damage membranes and nucleic acids. These compounds include free radicals, the superoxide anion (O3+), H2O2 and other highly reactive compounds. To counteract the damage caused by these compounds, plants produce antioxidant molecules. However, once the antioxidants react with ROS they are themselves inactivated [84].
There are various enzymes with reductive abilities that regenerate the active form of the antioxidants [84]. Endophytes have the capability to up-regulate the genes that code for the reductive enzymes. This is a major mechanism of disease, pest, and abiotic stress resistance. Much of the damage to cellular systems is caused by ROS, and the up-regulation of these ameliorates this damage. Endophytes with this ability include Trichoderma [33,85], mycorrhizae [36], Piraformaspora indica [86,87,88].
Acc Deaminase
ROS are not the only metabolites that damage plant cells under biotic or abiotic stresses. Ethylene, like ROS have signaling functions in plants but cause injury to tissues if at high levels. Acc deaminase (1-aminocyclopropane-1-carboxylate) breaks down ACC, which an immediate precursor of ethylene to ammonia and α-ketobuyrate [89]. This enzyme is produced by various bacterial root endophytes, including Pseudomonas spp [90] and fungi such as Trichoderma [91]. When the genes encoding ACC deaminase are produced in plants they ameliorate effects of many kinds of stress.
Thus, a wide range of endophytic bacteria and fungi enhance plant growth through similar mechanisms, even though they are taxonomically diverse. The diverse benefits to plant growth and development are common to numerous endophytes.
Part 2. Integrated Solutions for Commercial Agriculture
The preceding sections have described the scientific underpinnings of the advantages of endophytes. However, endophytes are only useful if they are amenable to commercial agricultural practices. This section will describe practices and systems to deploy endophytes in commercial agriculture.
Large-Scale Production and Formulation Methods
Methods for producing large amounts of endophytes are necessary. The methods used need to be rapid and inexpensive. For bacteria liquid fermentation is widely used, while for fungi semi-solid fermentation is preferable. It is not sufficient just to produce the endophyte, the material produced must have adequate shelf life and the physiological state of the material is important. For example, Trichoderma needs to be acclimated to low moisture levels, otherwise shelf life is inadequate. Adding an osmoticant such as glycerol to the growth medium extended shelf life of the fungal preparations [92].
Formulation methods are required. In many cases, the earliest formulations were powders that were applied to seeds at the time of planting. These were effective but too laborious for most farmers. Liquid formulations are preferred and frequently essential for large-scale agriculture. Liquids can be either water or oil based. Fungal spores frequently are covered with hydrophobins, which are proteins with one hydrophobic and one hydrophilic end. They are small cystine-rich molecules [93]. If fungal spores are dried and mixed with oils, they form a stable suspension. If suspended in oils, they become immiscible with water. However, if suspended in water, they then become immiscible with oils. This is due to the reorientation of the hydrophobins to become either hydrophobic or hydrophilic. This property is useful in formulations to provide concentrated and effective fungal products.
Encapsulations are also useful in formulations. Coating of inoculum with materials such as glucans can frequently provide longer shelf life [94]. Similar improvements in shelf life is possible through the use of invert emulsions. Invert emulsions are mixtures of water, oils and a surfactant (mayonnaise is a good example). They can provide enhanced shelf life, but also increase compatibility with chemical pesticides.
Integrated Biological-Biological and Chemical-Biological Treatments
It seems paradoxical to consider the use of fungi with chemical fungicides, but it is possible. Almost any biological can safely be coated onto seeds if the chemical pesticide is allowed to dry before application of the biological. This permits the use of biologicals in high throughput commercial systems. One method is to treat seeds with liquid slurry treatments and upon exit from the seed treater, application of the biological via a secondary treater. It also is possible to use chemical-biological treatments using alternations of the biological and the chemical. Of course, if endophytes that internally colonize plants, then the living organism does not come into contact with the pesticide. In that case, the strong, but brief, protection is provided by the chemical, while the long-term advantage to plants is provided by the biological. This combination frequently provides better plant protection against biotic abiotic stresses and may increase crop yields. Biological-biological treatments may also be advantageous. Examples are Rhizobium-Trichoderma treatments that both improve plant nutrition and provide the other benefits just described. Other commercial products may include Trichoderma-Trichoderma, Trichoderma-Bacillus. In addition to seed treatments, biologicals can be applied to potting mixes, as drenches when transplanting and usually are fully compatible with any chemical that may be applied. Co-inoculation with Trichoderma and the AMF fungi provided disease control and/or plant yield enhancement [95,96].
References
- Margulis, L.; Fester, R. Symbiosis as a Source of Evolutionary Innovation; MIT Press: Boston, 1991; p. 470. [Google Scholar]
- Chi, F.; Shen, S.-H.; Cheng, H.-P.; Jing, Y.-X.; Yanni, Y.G.; Dazzo, F.B. Ascending migration of endophytic rhizobia, from roots to leaves, inside rice plants and assessment of benefits to rice growth physiology. Appl. Environ. Microbiol. 2015, 71, 7271–7278. [Google Scholar] [CrossRef]
- Sutton, J.C.; Liu, W.; Huang, R.; Owen-Going, N. Ability of Clonostachys rosea to establish and suppress sporulation potential of Botrytis cinerea in deleafed stems of hydroponic greenhouse tomatoes. Biocontrol Sci. Tech. 2002, 12, 413–425. [Google Scholar] [CrossRef]
- Harman, G.E.; Uphoff, N. Advantages and methods of using symbiotic microbes to enhance plant agriculture and the environment. Scientifica 2019. [Google Scholar]
- Wilson, D.O. Endophyte: the evolution of a term, and clarification of its use and defition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Chen, X.-L.; Sun, M.-C.; Chong, S.-I.; Si, Y.-P.; Wu, L.-S. Transcriptome and metabolomic approaches deepen our knowledge of plant-endophyte interactions. Front. Plant Sci. 2022, 12, 700200. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species---opportunistic, avirulent plant symbionts. Nature Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Alonso-Ramirez, A.; Poveda, J.; Martin, I.; Hermosa, R.; Monte, E.; Nicolas, C. Salicylic acid prevents Trichoderma harzianum from entering the vascular system of roots. Moleb. Plant Pathol. 2014, 15, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E. Trichoderma--not just for biocontrol anymore. Phytoparasitica 2011, 39, 103–108. [Google Scholar] [CrossRef]
- Woo, S.J.; Ruocco, M.; Vinale, F.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based and their widespread use in agriculture. Open Mycology J. 2014, 8, 71–126. [Google Scholar] [CrossRef]
- Doni, F.; Zaln, C.R.C.M.; Isahak, A.; Faturrahaman, F.; Anhar, A.; Mohamad, W.; Yusoff, W.M.W.; Uphoff, N. A simple, efficient, and farmer-friendly Trichoderma-based biofertilizer evaluated with the SRI rice management system. Organic Agric. 2017. [Google Scholar] [CrossRef]
- Ismail; Hamayun, M. ; Hussain, A.; Iqbal, A.; Khan, S.A.; Lee, I.-J. Aspergillus niger boosted heat stress tolerance in sunflower and soybean via regulating their metabolic and antioxidant system. J. Plant Interact. 2020, 15, 223–232. [Google Scholar] [CrossRef]
- Ikram, M.; Ali, N.; Jan, G.; Jan, F.G.; Rahman, I.U.; Iqbal, A.; Hamayun, M. IAA producing fungal endophyte Penicillium roqueforti Thom., enhances stress tolerance and nutrients uptake in wheat plants grown on heavy metal contaminated soils. PLoS One 2018, 13, e0208150. [Google Scholar] [CrossRef] [PubMed]
- Shafia, A.; Sutton, J.C.; Yu, H.; Fletcher, R.A. Influence of preinoculation light intensity on development and interactions of Botrytis cinerea and Clonostachys rosea in tomato leaves. Canadian J. Plant Pathol. 2001, 23, 346–357. [Google Scholar] [CrossRef]
- Sherameti, I.; Shahollari, B.; Venus, Y.; Altschmied, L.; Varma, A.; Oelmueller, R. The endophytic fungus Piriformospora indica stimulates the expression of nitrate reductase and the starch-degrading enzyme glucan-water dikinase in tobacco and Arabidopsis roots through a homeodomain transcription factor that binds to a conserved motif in their promoters. J. Biol. Chem. 2005, 280, 26241–26247. [Google Scholar]
- Franza, G.J.; Muhammad, H.; Hussain, A.; Gul, J.; Aman, K.; In-Jung, L. An endophyte isolate of the fungus Yarrowia lipolytica produce metabolite that ameliorate the negative impact of salt stress on the physiology of maiaze. BMC Microbiol. 2019, 19, 3. [Google Scholar]
- Konappa, N.; Krishnamurthy, S.; Arakere, U.C.; Chowdappa, S.; Ramachandrappa, N.S. Efficacy of indigenous plant growth-promoting rhizobacteria and Trichoderma strains in eliciting resistance against bacterial wilt in a tomato. Egyptian J. Biol.l Pest Control 2020, 30, 106. [Google Scholar] [CrossRef]
- Brannen, P.M. , Kenney, D. S. Kodiak: A successful biological-control product for suppression of soil-borne plant pathogens of cotton. J. Industr. Microbiol. Biotechnol. 1997, 19, 169–171. [Google Scholar] [CrossRef]
- Poveda, J.; Eugui, D.; Abtil-Urias, P.; Velasco, P. Endophytic fungi as direct plant growth promoters for sustainable agriculture. Symbiosis 2021, 85, 1–19. [Google Scholar] [CrossRef]
- Woo, S.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef]
- Harman, G.E. Microbial plant symbionts: new tools to improve plant productivity. International Conference of Food Processing and Technology, Paris, France 2018.
- Leger, R.J.S. Studies on adaptations of Metarhizium anisopliae to life in the soil. Journal of Invertebrate Pathology 2008, 98, 271–276. [Google Scholar] [CrossRef]
- Coppola, M.; Diretto, G.; Digilio, M.C.; Woo, S.L.; Giuliano, G.; Molisso, D.; Pennacchio, F.; Lorito, M.; Rao, R. Transcriptome and metabolome reprogramming in tomato plants by Trichoderma harzianum strain T22 primes and enhances defense responses against aphids. Front. Physiol. 2019, 10, 745. [Google Scholar] [CrossRef] [PubMed]
- Stasz, T.E.; Harman, G.E.; Weeden, N.F. Protoplast preparation and fusion in two biocontrol strains of Trichoderma harzianum. Mycologia 1988, 80, 141–150. [Google Scholar] [CrossRef]
- Hubbard, J.P.; Harman, G.E.; Hadar, Y. Effect of soilborne Pseudomonas sp. on the biological control agent, Trichoderma hamatum, on pea seeds. Phytopathology 1983, 73, 655–659. [Google Scholar] [CrossRef]
- Harman, G.E.; Hayes, C.K.; Ondik, K.L. Asexual genetics in Trichoderma and Gliocladium: mechanisms and implications. In Trichoderma and Gliocladium, Vol. 1; Kubicek, C.P., Harman, G.E., Eds.; Taylor and Francis: London, 1998; pp. 243–270. [Google Scholar]
- Stackhouse, T.; Browm, D.; Glenn, A.; Gold, S.E. Fungal imposter syndrome: A forensic analysis of the genomic contributions to the important biological control agent, Trichoderma strain T22. In Proceedings of the Annual meeting Am. Phyopathol. Socirty.
- Woo, S.; Hermosa, R.M.; Lorito, M.; Monte, E. Trichoderma: a multipurpose, plant-beneficial miroorganism for eco-sustainable agriculture. Nature Rev. Microbol. 2023, 21, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Druzhinia, I.S.; Chenthamara, M.S.; Zhang, J.; Atanasove, L.D.Y.; Miao, Y.; Rhami, M.R.; Grujic, M.; Cai, F.; Pourmedhi, M.S.; al, e. Massive lateral transfer of genes encoding cell-wall degrading to the mycoparasitic fungus Trichoderma from its plant-assoxiated hosts. PLOS Genetics 2018.
- Shahid, M.; Pinelli, E.; Dumat, C. Tracing trends in plant physiology and biochemistry: need of databases from genetic to kingdom level. Plant. Physiol. Biochem. 2018, 127, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E. Multifunctional fungal plant symbionts: new tools to enhance plant growth and productivity. New Phytol. 2011, 189, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Rodriquez, R.; Baird, A.S.; Gray, Z.; Groove, R.E.; Harto, R.; et al. Programming plants for climate resilence through symbiotics. In Seed Endophytes; Verma, S., White, J.j., Eds.; Springer: Cham, 2019; pp. 127–137. [Google Scholar]
- Mastouri, F.; Bjorkman, T.; Harman, G.E. Trichoderma harzianum strain T22 enhances antioxidant defense of tomato seedlings and resistance to water deficit. Molec. Plant Microbe Interact. 2012, 25, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- Mastouri, F. Use of Trichoderma spp. to improve plant performance under abiotic stress. PhD, Cornell University, Ithaca, NY, 2010.
- Shukla, V.; Awasthi, R.P.; Rawat, L.; Kumar, J. Regulation of plant growth, photosynthesis and drought tolerance in Triticum aestivum. Ann. Appl. Biol. 2015, 2015. [Google Scholar] [CrossRef]
- Mo, Y.; Wang, R.; Yang, R.; Yang, R.; Jun, Z.; Liu, C.; Li, H. Regulation of plant growth, photosynthesis, and osmosis by an arbuscular fungus in watermelon seedlings. Front. Plant Sci. 2017, 7. [Google Scholar] [CrossRef]
- Azad, K.; Kaminskyj, S. A fungal endophyte strategy for mitigating the effect of salt and drought stress on plant growth. Symbiosis 2016, 68, 73–77. [Google Scholar] [CrossRef]
- Redman, R.S.; Kim, Y.O.; Woodward, C.J.G.C.; Espino, L.; Doty, S.L.; Rodriguez, R.J. Increased fitness of rice plants to abiotic stress via habitat adapted symbiosis: a strategy for mitigating impacts of climate change. PLoS One 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.M.; Kang, B.R.; Kim, Y.C. Transcriptome analysis of induced systemic drought tolerance elicited by Pseudomonas chlororaphis O6 in Arabidopsis thaliana. Plant Pathol J. 2013, 29, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Vaishnav, L.J.; Choudhary, D.K. Regulation of drought responsive in Glycine max is mediated through Pseudomonas simiae strain AU. J. Plant Growth Regul. 2019, 38, 333–342. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria ACC deaminase and the alleviation of plant stress. Adv. Appl. Microbiol. 2004, 56, 297–312. [Google Scholar]
- Mastouri, F.; Bjorkman, T.; Harman, G.E. Seed treatments with Trichoderma harzianum alleviate biotic, abiotic and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, C.S.; Srivastava, S.; Chauhan, P.S.; Seem, K.; Mishra, A.; Sopory, S.K. Plant growth-promoting bacteria Bacillus amyloliquefaciens NBRISN13 modulates gene expression profile of leaf and rhizosphere community in rice during salt stress. Plant Physiol. Biochem. 2013, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Waller, F.; Achatz, B.; Baltruschat, H.; Fodor, J.; Becker, K.; Fischer, M.; Heier, T.; Hueckelhoven, R.; Neumann, C.; von Wettstein, D.; et al. The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. PNAS 2005, 102, 13386–13391. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, M.E.; Abdelsatter, M.; Abdelaym, E.A.; Atia, M.A.M.; Mahmooud, A.W.M.; Saad, M.M.; sl., r. Piriformaspora indica alters Na+/K+ homeostasis, antioxidant enzymes and LeNHX1 expression of greenhouse tomatoes under sakt stress. Scien. Hort. 2019, 256, 108532. [CrossRef]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 1–16. [Google Scholar] [CrossRef]
- Faranza, g.J.; Mahmmad, H.; Hassian, A.; Gul, J.; Ahmad, J.S.; Faranza, G.J.; Mahmood, H.; Hassan, A.; Gul, J.; Amad, I.; et al. An endophytic isolate of Yarrowia lipolytica produce metabolites thst amelorate the negative effects of salt stress on maize. BMC Micorobiol. 2019, 19. [Google Scholar] [CrossRef]
- Harrison, M.J. The arbuscular mycorrhizal symbiosis: an underground association. Trends in Plant Sci. 1997, 2, 54–60. [Google Scholar] [CrossRef]
- Altomare, C.; Norvell, W.A.; Björkman, T.; Harman, G.E. Solubilization of phosphates and micronutrients by the plant-growth-promoting and biocontrol fungus Trichoderma harzianum Rifai 1295-22. Appl. Environ. Microbiol. 1999, 65, 2926–2933. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Mastouri, F. (Eds.) Enhancing nitrogen use efficiency in wheat using Trichoderma seed inoculants; International Society for Plant-Microbe Intereactons: St. Paul, MN, 2010; Volume 7, p. 4. [Google Scholar]
- Bailey, B.A.; Lumsden, R.D. Direct effects of Trichoderma and Gliocladium on plant growth and resistance to plant pathogens. In Trichoderma and Gliocladium, Vol. 2; Harman, G.E., Kubicek, C.P., Eds.; Taylor and Francis: London, 1998; pp. 185–204. [Google Scholar]
- Harman, G.E. Myths and dogmas of biocontrol. Changes in perceptions derived from research on Trichoderma harzianum T-22. Plant Dis. 2000, 84, 377–393. [Google Scholar] [CrossRef] [PubMed]
- Garber, R.K.; Cotty, P.J. Formation of sclerotia and aflatoxins in developing cotton bolls infected by the S strain of Aspergillus flavus and potential for biocontrol with an atoxigenic strain. Phytopathology 1997, 87, 940–945. [Google Scholar] [CrossRef]
- Khan, A.L.; Hussain, J.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.-J. Endophytic fungi: resource for gibberellins and crop abiotic stress resistance. Critical Reviews in Biotechnology 2015, 35, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Lorito, M.; Lynch, J.M. Uses of Trichoderma spp. to remediate soil and water pollution. Adv. Appl. Microbiol. 2004, 56, 313–330. [Google Scholar] [PubMed]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur. J. Plant Pathol. 2007, 119, 329–339. [Google Scholar] [CrossRef]
- Shoresh, M.; Harman, G.E. The relationship between increased growth and resistance induced in plants by root colonizing microbes. Plant Signal. Behavior 2008, 3, 737–739. [Google Scholar] [CrossRef]
- Paradiso, R.; Arena, C.; De Micco, V.; Giordano, M.; Aronne, G.; De Pascale, S. Changes in leaf anatomical traits enhanced photosynthetic activity of soybean Ggrown in hydroponics with plant growth-promoting microorganisms. Front. Plant Science 2017, 8, 674. [Google Scholar] [CrossRef]
- Doni, F.; Fathurrahman, F.; Mispan, M.S.; Suhaimi, M.; Yussof, W.M.W.; Uphoff, N. Transciptome profiling of rice seedlings inoculated with symbiotic fungus Trichoderma asperellum SL2. J. Plant Growth Reg. 2019. [Google Scholar] [CrossRef]
- Vitti, A.; Pellegrini, E.; Nali, C.; Lovelli, S.; Sofo, A.; Valerio, M.; Scopa, A.; Nuzzaci, M. Trichoderma harzianum T-22 induces systemic resistance in tomato Infected by Cucumber mosaic virus. Front. Plant Science 2016, 7, 1520. [Google Scholar] [CrossRef] [PubMed]
- Pelhivan, N.; Yesilyurt, A.M.; Durmas, N.; Karaoglu, S.A. Trichoderma lixii ID11D seed biopriming mitigates dose dependent salt toxicity in maize. Acta Physiol. Plantarum 2017, 39. [Google Scholar] [CrossRef]
- Fu, J.; Wang, W.-F.; LI, Z.-T.; Yang, K.-J. Trichoderma asperellum alleviates the effects of sale-alkaline stress on maize seedling via the regulation of photosynthesis and nitrogen metaolism. Plant Growth Regul. 2018, 85, 363–374. [Google Scholar] [CrossRef]
- Harman, G.E.; Doni, F.; Khada, R.B.; Uphoff, N. Endophytic strains of Trichoderma increase plants’ photosynthetic capability. App. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.; Malfanova, N.; Kamilova, F.; Berg, G. Plant growth promotion by microbes. In Molecular Microbial Ecology of the Rhizosphere; de Bruijn, F., Ed.; J. Wiley and sons, 2013; pp. 561–573. [Google Scholar]
- Thomashow, L.S.; Weller, D.M.; Bonsall, R.F.; Pierson, L.S.I. Production of the antibiotic phenazine-1-carboxylic acid by fluorescent Pseudomonas species in the rhizosphere of wheat. Appl. Environ. Microbiol. 1990, 56, 908–912. [Google Scholar] [CrossRef] [PubMed]
- Sivasithamparam, K.; Ghisalberti, E.L. Secondary metabolism in Trichoderma and Gliocladium. In Trichoderma and Gliocladium, Vol. 1; Kubicek, C.P., Harman, G.E., Eds.; Taylor and Francis: London, 1998; pp. 139–191. [Google Scholar]
- Howell, C.R.; Stipanovic, R.D. Suppression of Phytium ultimum-induced damping-off of cotton seedlings by and its antibiotic, pyoluteorin. Phytopathology 1980, 70, 712–715. [Google Scholar] [CrossRef]
- The Manual of Biocontrol Agents, 4th ed.; Gwynn, R.L., Ed.; British Crop Protection Council: Alton. Hamshire, UK, 2014; p. 277. [Google Scholar]
- Whipps, J.M. Effects of mycoparasites on sclerotia-forming fungi. In Developments In Agricultural And Managed Forest Ecology; Beemster, A. B. R., et al., Eds.; 1990; pp. 0–444. [Google Scholar]
- Eckenrode, C.J.; Harman, G.E.; Webb, D.R. Seed-borne microorganisms stimulate seedcorn maggot egg laying. Nature 1975, 256, 487–488. [Google Scholar] [CrossRef]
- Harman, G.E.; Eckenrode, C.J.; Webb, D.R. Alteration of spermoshere ecosystems affecting ovipoistion of the bean seed fly and attack by soilborne fungi on germinating seeds. Annal. Appl. Biol. 1978, 90, 1–6. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, M.S.C.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Ann. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Waller, F.; Molitor, A.; Pfiffi, S.; Achatz, B.; Kogel, K. The root endophytic fungus Piriformospora indica accelerates host plant development and primes plants for disease resistance. Phytopathology 2008, 98, 6. [Google Scholar]
- Yedidia, I.; Shoresh, M.; Kerem, Z.; Benhamou, N.; Kapulnik, Y.; Chet, I. Concomitant induction of systemic resistance to Pseudomonas syringae pv. lachrymans in cucumber by Trichoderma asperellum (T-203) and accumulation of phytoalexins. Appl. Environ. Microbiol. 2003, 69, 7343–7353. [Google Scholar] [CrossRef]
- Yedidia, I.; Benhamou, N.; Kapulnik, Y.; Chet, I. Induction and accumulation of PR proteins activity during early stages of root colonization by the mycoparasite Trichoderma harzianum strain T-203. Plant Physiol. Biochem. 2000, 38, 863–873. [Google Scholar] [CrossRef]
- Vinale, F.; Ghisalberti, E.L.; Flematti, G.; Marra, R.; Lorito, M.; Sivasithamparam, K. Secondary metabolites produced by a root-inhabiting sterile fungus antagonistic towards pathogenic fungi. Letters in Applied Microbiology 2010, 50, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Djonovic, S.; Vargas, W.A.; Kolomiets, M.V.; Horndeski, M.; Weist, A.; Kenerley, C.M. A proteinaceous elicitor Sm1 from the beneficial fungus Trichoderma virens is required for systemic resistance in maize. Plant Physiol. 2007, 145, 875–889. [Google Scholar] [CrossRef] [PubMed]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational research on Trichoderma: from ‘omics to the field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef]
- Ramirez-Valdespino, C.A.; Casas-Flores, S.; Olmedo-Monfil, V. Trichoderma as a Model to Study Effector-Like Molecules. Front. Microbiol. 2019, 10, 1030. [Google Scholar] [CrossRef]
- Jaskiewicz, M.l.R.; Conrath, U.; Peterhaensel, C. Chromatin modification acts as a memory for systemic acquired resistance in the plant stress response. EMBO Reports 2011, 12, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Jaskiewicz, M.; Peterhansel, C.; Conrath, U. Detection of Histone Modifications in Plant Leaves. Jove-J. Visualized Experi. 2011. [Google Scholar] [CrossRef]
- Conrath, U.; Thulke, O.; Katz, V.; Schwindling, S.; Kohler, A. Priming as a mechanism in induced systemic resistance of plants. European J. Plant Pathol. 2001, 107, 113–119. [Google Scholar] [CrossRef]
- Agripino de Medeiros, H.; de Araujo Filho, J.V.; de Freitas, L.G.; Castillo, P.; Belen Rubio, M.; Hermosa, R.; Monte, E. Tomato progeny inherit resistance to the nematode Meloidogyne javanica linked to plant growth induced by the biocontrol fungus Trichoderma atroviride. Scientific Reports 2017, 7, 40216. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Chepsergon, J.; Mwamburi, L.; Kassem, K.M. Mechanism of drought tolerance using Trichoderma spp. Intern. J, Science Research 2014, 3, 1592–1596. [Google Scholar]
- Sun, C.; Johnson, J.; Cai, D.; Sherameti, I.; Oelmueller, R.; Lou, B. Piriformospora indica confers drought tolerance in Chinese cabbage leaves by stimulating antioxidant enzymes, the expression of drought-related genes and the plastid-localized CAS protein. J Plant Physiol 2010, 167, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Baltruschat, H.; Fodor, J.; Harrach, B.D.; Niemczyk, E.; Barna, B.; Gullner, G.; Janeczko, A.; Kogel, K.-H.; Schaefer, P.; Schwarczinger, I.; et al. Salt tolerance of barley induced by the root endophyte Piriformospora indica is associated with a strong increase in antioxidants. New Phytologist 2008, 180, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Vadassery, J.; Tripathi, S.; Prasad, R.; Varma, A.; Oelmueller, R. Monodehydroascorbate reductase 2 and dehydroascorbate reductase 5 are crucial for a mutualistic interaction between Piriformospora indica and Arabidopsis. Journal of Plant Physiology 2009, 166, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Shelke, G.M.; Kumar, A.; Jha, P.N. Biochemistry and genetics of ACC deaminase: a weapon to “stress ethylene” produced in plants (vol 6, 937, 2015). Front. Microbiol. S 2015, 6, 1255. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiology and Biochemistry (Paris) 2014, 80, 160–167. [Google Scholar] [CrossRef]
- Zhang, S.; Gan, Y.; Xu, B. Mechanisms of the IAA and ACC-deaminase producing strain of Trichoderma longibrachiatum T6 in enhancing wheat seedling tolerance to NaCl stress. BMC Plant Biology 2019, 19, 22. [Google Scholar] [CrossRef]
- Jin, X.; Harman, G.E.; Taylor, A.G. Conidial biomass and desiccation tolerance in Trichoderma harzianum. Biol. Control 1992, 1, 237–243. [Google Scholar] [CrossRef]
- Sunde, M.; Kwan, A.H.T., M. D.; Beever, R.E.; Mackay, J.P. Structual analysis of hydrophobins. Micron 2008, 39, 773–784. [Google Scholar] [CrossRef]
- Harman, G.E.; Custis, D. US Patent 9,090,884. Formulations of viable microorganisms and their method of use 2006.
- Datnoff, L.E.; Nemec, S.; Pernezny, K. Biological control of fusarium crown and root rot of tomato in Florida using Trichoderma harzianum and Glomus intraradices. Biological Control 1995, 5, 427–431. [Google Scholar] [CrossRef]
- Buysens, C.; Cesar, V.; Ferrais, F.; de Boulois, H.; Declerck, S. Inoculation of Medicago sativa cover crop with Rhizophagus irregularis and Trichoderma harzianum increases the yield of subsequently-grown potato under low nutrient conditions. Appl. Soil Ecol. 2016, 105, 137–143. [Google Scholar] [CrossRef]
- Vitti, A.; Pelligrini, E.; Nali, C.; Lovelli, S.; Sofo, A.; Valerio, M.; Scopa, A.; Nuzzaci, M. Trichoderma harzianum T22 induces sytemic restance in tomato infected by cucumber mosaic virus. Front. Plant Sci. 2916, 7, 1520–doi1510. [Google Scholar]
- Poveda, l.; Eudui, D.; Abtil--Urias, P.; Velasco, P. Endophytic fungi as diect plant growth promoters for sustainable agricultue. Symbiosis 2021, 85, 1–19. [Google Scholar] [CrossRef]
- Xue, A.G. Biological control of pathogens causing root rot complex in field pea using Clonostachys rosea strain ACM941. Phytopathology 2003, 93, 329–335. [Google Scholar] [CrossRef]
- Gwynn, R.L. The Manual of Biocontrol Agents, 5th ed.; British Crop Protection Council: Alton, Hamshire UK, 2009; p. 277. [Google Scholar]
- Gill, S.S.; Gill, R.; Trivedi, D.K.; Anjum, N.A.; Sharma, K.K.; Ansari, M.W.; Ansari, A.A.; Johri, A.K.; Prasad, R.; Pereira, E.; et al. Piriformospora indica: Potential and Significance in Plant Stress Tolerance. Front. Microbiol. 2016, 7, 332. [Google Scholar] [CrossRef]
- Hu, J.-L.; Lin, X.-G.; Wang, J.-H.; Shen, W.-S.; Wu, S.; Peng, S.-P.; Mao, T.T. Arbuscular mycorrhizal fungal inoculation enhances suppression of cucumber Fusarium wilt in greenhouse soils. Pedosphere (Beijing) 2010, 20, 586–593. [Google Scholar] [CrossRef]
- Zhang, J.X.; Cotty, P.J. Diversity within communities of L and S strains of Aspergillus flavus in cotton fields in Texas and Arizona. Phytopathology 2007, 97, S172. [Google Scholar]
- Leger, R.J.S. Studies on adaptations of Metarhizium anisopliae to life in the soil. J. Invert. Pathol 2008, 98, 271–276. [Google Scholar] [CrossRef]

Table 1.
Pests and pathogens controlled by different microorganisms.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



