Submitted:
22 April 2024
Posted:
23 April 2024
You are already at the latest version
Abstract
The growing demand in industrial and biotechnological settings for more efficient enzymes with enhanced biochemical features, particularly thermostability and thermotolerance, necessitates a timely response. Renowned for their versatility, thermostable enzymes, offer significant promise across a range of applications, including agricultural, medicinal, and biotechnological domains. This comprehensive review summarizes the structural attributes, catalytic mechanisms, and the relationships between structure and function of two major classes of thermostable enzymes: α-amylases and laccases. These enzymes serve as valuable models for understanding the structural basis of protein thermostability. Commercial significance of these enzymes and researchers' interest in further optimization and innovation, this article can greatly contribute to ongoing research on thermostable enzymes and aiding industries in optimizing production processes. It also give insights to the exploration of suitable strategies and factors for enhancing thermostability, resulting in heightened multipronged stability and notable enhancements in the enzymes’ industrial applicability.
Keywords:
thermostable enzymes
; -amylases
; laccases
; industrial applications
Introduction
The enduring significance of thermostable enzymes stems from their diverse applications across industries, including food, pharmaceuticals, and biotechnology. In the realm of industrial biocatalysis, enzymes face stringent demands, necessitating robust and thermostable biocatalysts to meet high industrial standards. Enzymes characterized as thermostable possess intrinsic stability, enabling them to endure elevated temperatures well beyond 50 oC, reaching as high as 80 oC, 90 oC or even more [1,2]. These enzymes maintain their structural integrity and distinctive features under such extreme conditions. This inherent stability provides significant biotechnological advantages compared to mesophilic enzymes (active optimally at 25 to 50 oC) or psychrophilic enzymes (active optimally at 5 to 25 oC). Thermostable enzymes are more easily purified through heat treatment (as one of the initial step), exhibit increased resilience and resistance to chemical denaturants, allow for higher substrate concentrations, contribute to lower viscosity, pose fewer risks of microbial contamination, and often lead to higher reaction rates [1]. This heightened stability allows them to thrive in harsh environments, including applications like enzymatic bioremediation of xenobiotics and innovative green processes [3]. The demand for industrially relevant thermostable enzymes has created a pressing need to identify easily accessible, cost-effective, and process-friendly sources. This review is motivated by the rapid strides in discovering novel thermostable enzymes from unconventional sources. Among these enzymes, α-amylases and laccases stand out as crucial players in industries such as pulp and paper, starch processing, textiles, detergents, fuels, alcohols, and pharmaceuticals, constituting the major consumers of these thermostable enzymes. Alpha-amylase patents distributed across different categories: in biofuels, beverages, pharmaceuticals, detergents, food, animal feed, and textiles, were found deposited in intellectual property databases [4]. Besides being widespread in plants, animals, fungi, unicellular eukaryotes like eubacteria and archaea, ‘Dictyo-type’ α-amylase is also found to be widespread and may be ancestral in the Unikonts, a clade comprising animals, fungi (Opisthokonts) and amoebozoa [5]. The activity of α-amylase has been observed to be associated with the neurodegenerative diseases such as Alzheimer’s disease. Studies also show that α-amylase synthesis is linked with mTOR (Mechanical Target of Rapamycin) signaling pathways [6]. mTOR serves as a key regulator of celluar growth by controlling the both anabolic and catabolic processes [7,8].
Another dimension of the profound interest in thermostable enzymes lies in the exploration of the thermodynamic stability of proteins. Investigating the interplay between stability, flexibility, or plasticity and catalytic efficiency adds a layer of understanding to these enzymes' properties. The heightened interest in thermostable enzymes has catalyzed a focus on developing enzymes with enhanced thermostability or thermotolerance through genetic engineering or site-directed mutagenesis, revolutionizing the attainment of desired enzyme properties. The preference for enzymatic processes, particularly thermostable ones, over conventional methods in various industries is attributed to their rapid and specific action, along with advantages in energy, time, raw material, and chemical savings. Crucially, their environmentally friendly nature further underscores their appeal. Moreover, conducting processes at elevated temperatures using thermostable enzymes not only diminishes the threat of microbial contamination, a notable advantage, but also serves to lower substrate viscosity, enhance transfer rates, and augment solubility in the course of reaction procedures [2].
Thermostable enzymes, beyond their inherent thermostability, exhibit favorable characteristics such as a wide pH tolerance and resistance to organic solvents, positioning them as superior to other enzyme groups. This underscores the need for continued efforts in screening and isolating novel sources, developing innovative purification approaches to enhance yield and purity, and ultimately harnessing thermostable enzymes for diverse industrial applications. The present review article is an effort to address thermostable α-amylases and laccases indicating their varied sources of origin, structural characteristics and catalytic mechanism, structure-function relationships as well as factors attributing and various strategies resulting thermostability. Besides, the prevailing challenges that exist in order to cope up with aiming for further research and advances in the field of thermostable enzymes providing insights into future direction in the field of thermostable enzymes, have been focused on.
Thermostable α-Amylases
Thermostable α-amylases, constituting the largest share of industrial enzyme sales at approximately 25%, play a pivotal role across diverse sectors such as fermentation, textile, food, detergent, brewing, biorefinery, paper, and pharmaceutical industries [9,10], as illustrated in Figure 1 and also listed in Table 1. Particularly crucial in starch liquefaction processes, these enzymes serve as valuable models for studying thermal adaptation in proteins [11,12]. Widely distributed in nature, α-amylases originate from various sources, including microbes, animals, and plants, with a notable presence in germinating seeds where they contribute significantly to carbohydrate metabolism [13,14].
Established in 1998, the CAZy Carbohydrate-Active Enzymes (CAZymes) offers users online access [15] (http://www.cazy.org) that is regularly updated to a sequence-based family classification linking sequences with the specificities and three-dimensional structures of enzymes that are involved in the assembly alteration, and degradation of oligo-and polysaccharides [16]. The enzymes that are currently covered in the CAZy database that catalyze the breakdown, biosynthesis or modification of carbohydrates and glycoconjugates are:
a) Glycosyl Hydrolases (GHs): hydrolysis and/or rearrangement of glycosidic bonds.
b) Glycosyl Transferases (GTs): formation of glycosidic bonds.
c) Polysaccharide Lyases (PLs): non-hydrolytic cleavage of glycosidic bonds.
d) Carbohydrate Esterases (CEs): hydrolysis of carbohydrate esters.
e) Auxiliary Activities (AAs): redox enzymes that act in conjunction with CAZymes.
In recent years, there has been significant growth in the CAZy classification system, including the introduction of new families and the establishment of subfamilies within existing ones [17]. Currently, of the 189 class glycoside hydrolases (GH) families in total, four are considered α-amylase families: i) GH13? proven to be the most abundant and largest α-amylase family composed of a (β/α)8 barrel structure; (ii) GH57? the second and smaller α-amylase family composed of a (β/α)7 barrel structure; (iii) GH119? a very small family related to GH57; (IV) GH126? composed of (α/α)6 barrel structure [18]. Families GH13, GH57 and GH119 employ the α-retaining mechanism of α-amylase involving two catalytic residues in the active site; a glutamic acid/base catalyst and an aspartate as the nucleophile and the anomeric carbon remains in the same position as it is mediated by the double-displacement mechanism. In contrast, GH126 employ the inverting reaction mechanism in which an anomeric carbon position is shifted from β to α through a single-displacement mechanism [19]. The α-amylase family GH13 has already been divided into 47 subfamilies, with additional subfamilies still emerging. Nature doesn’t always provide enzymes with the desired properties; hence protein engineering has been proposed as an alternative method to enhance the physicochemical traits of enzymes [20]. Leveraging solved crystal structures, a structure-guided consensus approach is recognized as an effective and dependable method for refining enzyme properties [21]. Employing this approach to enhance the thermostability of α-amylase without additional Ca2+ would benefit process efficiency and reduce the cost of the starch liquefaction process [22,23] This on other hand, requires both a starting enzyme with which to work and adequate structural information to guide the modifications. In the related efforts, Li and coworkers [21] showed from their experimental observations that malto-hexaose-forming α-amylsae from Bacillus stearothermophilus (AmyMH) is an adequate starting point from which to design a more thermostable α-amylase without added Ca2+. Previously, attention was directed towards a loop situated in domain B across various bacterial α-amylases as a means to enhance their thermostability. Suzuki and coworkers [24] suggested that eliminating the equivalent loop formed by R176-G177 (according to BAA numbering) could significantly boost the thermostability of Bacillus amyloliquefaciens α-amylase (BAA). This depiction has been replicated in several other bacterial α-amylases from different species, yielding similar improvements in thermostability [25,26]. Furthermore, the enhanced thermostability of Bacillus licheniformis α-amylase (BLA) was achieved by removing amide-containing side chains through the mutation of N190F (according to BLA numbering [27].
Despite the escalating demand for thermostable enzymes in multiple industries, the production and properties of α-amylases have been restricted by their susceptibility to extremes of pH, temperature, external conditions, and catalytic efficiency [28,29]. In this context, thermostable enzymes play a critical role in withstanding the high temperatures inherent in industrial processes [10]. Microbial sources, especially fungi and bacteria, particularly those belonging to the Bacillus genus (e.g., B. subtilis, B. stearothermophilus, B. licheniformis, B. amyloliquefaciens), are often favored for industrial production due to their cost-effectiveness, consistency, efficient use of time and space, and ease of process modification and optimization [11].
The ongoing quest for novel thermostable α-amylases has extended to plant sources, seeking alternatives that meet the standards set by microbial sources. Plant-derived sources such as barley [30], mung bean [31], potato [32,33], soybean [12], radish [34], red pitaya peel [35], wheat [36], broad bean [14], and sword bean [37] have been explored. Reports of thermostable α-amylases have emerged from plant sources cultivated in local areas, including Vicia faba (65 °C), red pitaya (Hylocereus polyrhizus) peel (70 °C), wheat (Triticum aestivum) seeds (68 °C), soybean (Glycine max) seeds (70 °C), mung bean (Vigna radiata) seeds (65 °C), and sword bean (Canavalia gladiate) seeds (70 °C). Some sources of thermostable α-amylases and their respective industrial applications is listed in Table 2.
Structural Characteristics and Catalytic Mechanism of α-Amylases
The architecture of α-amylases consists of a single polypeptide chain that folds into three distinctive domains (Figure 1): a) Domain A, the catalytic domain characterized by an N-terminal (β/α)8 barrel (also known as TIM barrel) structure, featuring eight parallel β-strands forming a barrel shape surrounded by eight α-helices; b) Domain B, an irregular β-rich structure that constitutes a significant portion of the substrate binding cleft, responsible for notable variations in size, structure, and substrate specificity among different α-amylases; and c) Domain C, forming the C-terminal segment of the sequence [38,39,40]. Some maltogenic amylases exhibit an additional D-domain after the C-domain, the function of which remains currently unknown [41].
The catalytic mechanism of α-amylases (α-1,4-glucan-4-glucanohydrolase, EC 3.2.1.1) classifies them as endozymes within the glycosyl hydrolase (GH13 family), targeting internal α-glycosidic bonds in polysaccharides like starch, glycogen, and others, leading to the hydrolysis of these bonds and the production of α-anomeric mono- or oligosaccharides [42], as depicted in Figure 1. In plants, α-amylases play a crucial role in the degradation of stored starch in germinating seeds, releasing sugars essential for proper plant growth [43]. Unlike many other amylases exclusive to hydrolyzing α-D-(1,4)-glycosidic bonds, maltogenic amylases exhibit remarkable catalytic versatility, hydrolyzing both α-D-(1,4)- and α-D-(1,6)-glycosidic bonds and participating in transglycosylation reactions, transferring glycosyl units to the C3, C4, or C6 hydroxyl groups of diverse acceptor mono- or disaccharides [44].
With only a few exceptions, nearly all identified α-amylases exhibit structural stability attributed to the presence of a calcium ion located at the interface between domains A and B. This stability is disrupted upon the removal of the calcium ion, leading to a noticeable decline in catalytic activity. A prominent illustration of this phenomenon is the α-amylase from Bacillus licheniformis (BLA), a hyperthermostable enzyme extensively utilized in biotechnology for starch and complex carbohydrate breakdown at temperatures reaching 110 oC, and also serving as a crucial component in detergents and baking additives [45]. The elucidation of BLA’s structure in the metal-containing state, coupled with cmalliomparisons to the apo-enzyme (calcium-depleted form), has provided insights into how metal ions regulate enzyme activity. Studies have revealed that the addition of calcium chelators significantly reduces the stability of BLA [46,47], resulting in a heightened susceptibility to proteolysis [48]. This serves as compelling evidence of the indispensable stabilizing role played by the calcium ion. Consequently, α-amylases are proposed as a novel category of metallo-enzymes distinguished by a prosthetic group—an alkaline-earth metal instead of a transition element. This group primarily serves a structural role, akin to disulfide bridges [49]. Calcium ions are believed to contribute structurally, positioned too distantly from the active site to directly participate in catalysis [50,51,52]. The heightened thermostability of the enzyme, attributed to calcium ions, is explained by their salting-out effect on hydrophobic residues within the protein, inducing a more compact structural conformation [53]. In certain α-amylases, the presence of a chloride ion in the active site enhances catalytic efficiency by inducing conformational changes around the active site, potentially by elevating the pKa (acid dissociation constant) of a hydrogen-donating residue in the active site.
Factors Contributing to Stability in Thermostable α-Amylases
Thermostable enzymes serve as valuable models for comprehending the physico-chemical factors governing protein thermostability. Identifying the structural characteristics implicated in thermal stability facilitates the engineering and design of more robust enzymes for diverse industrial applications. Over the past two decades, α-amylase has emerged as a crucial model system for investigating the thermal adaptation of enzymes [40].
Studies based on amino acid sequence prediction, including the distribution of amino acids and dipeptide composition, help discern factors contributing to thermostability, distinguishing thermophilic and mesophilic proteins. Despite sequence identity and structural similarity, thermophilic proteins exhibit higher frequencies of charged, hydrophobic, and aromatic amino acids compared to mesophilic counterparts [54]. The presence of fewer cysteines in thermophilic proteins is associated with their oxidation at higher temperatures. Sequence analysis proves valuable in predicting protein thermostability when structural information is lacking. Comparing dynamic features of mesophilic proteins and their thermophilic homologs with increased sequence identity and structural similarity but distinct thermostability offers insights into the physical basis of protein stability [55]. Calcium ions play crucial roles in determining the structure, function, and stability of thermophilic α-amylases, providing resistance or tolerance to thermal inactivation by maintaining correct protein conformation [56,57,58,59]. The removal of calcium ions irreversibly inactivates barley α-amylase, while calcium ion addition restores activity in certain bacterial α-amylases [60]. Mostly, all α-amylases are Ca2+-dependent with some exceptions of Ca2+-independent α-amylases [61,62,63,64]; and also, some α-amylases that are inhibited by Ca2+ [65,66]. The role of α-amylases is mainly structural because their catalytic sites are far away from the calcium-binding sites [52,53]. Several studies have been made on the effect of calcium ions on the activity and stability of α-amylases from thermophiles which may help in determining the mechanism of Ca2+-binding proteins in the presence of extreme thermal environment as was investigated by Liao and coworkers in which they studied the influence of calcium ions on the structure and thermal characterization of α-amylase (AGXA) from thermophilic Anoxybacillus sp.GXS-BL [67].
Multiple factors contribute to thermostability, such as increased hydrogen bonds, ionic and electrostatic interactions, hydrophobic interactions, disulfide bonds, metal binding, salt bridges, ion pairs, aromatic clusters, sidechain-sidechain interactions, shorter surface loops, GC-rich codons, charged amino acid ratios, amino acid preferences, post-translational modifications, and solute accumulation [11]. Thermophilic proteins are characterized by more rigid and compact packing density, lower thermal motion, decreased flexibility, shorter surface loops, stabilization by heat-stable chaperones, reduced water-accessible hydrophobic surface, decreased entropy difference between folded and unfolded states, increased proline frequency, and decreased thermolabile residue occurrence compared to mesophilic counterparts [11,12].
In terms of industrial applications, enzyme stability is crucial, with a focus on thermodynamic and long-term stability. Various strategies for stability enhancement, including immobilization, addition of stabilizing agents, chemical modification, protein engineering, and genetic engineering through cloning and expression of thermostable α-amylase genes, have been explored. Site-directed mutagenesis and the revolutionary approach of directed evolution have emerged as promising strategies for thermostabilization [68,69]. Economic considerations in starch processing industries drive the need for α-amylases active at higher temperatures, and ongoing research has shifted the focus from stability engineering to pH activity profile and substrate specificity engineering, resulting in the development of α-amylases with novel and improved properties.
Thermostable Laccases
Laccases are recognized as environmentally friendly proteins and green biocatalysts, setting them apart from other oxidases. Unlike certain oxidases, laccases do not depend on toxic H2O2 or any mediator for the reduction reaction, and they exclusively generate water as the end product by reducing molecular oxygen. These glycoproteins exist in monomeric, dimeric, and tetrameric forms, displaying the ability to oxidize a wide range of inorganic substrates, aromatic compounds, and organic compounds. Primarily employed for the breakdown of chemical contaminants, laccase's low substrate specificity and monoelectronic oxidation of substrates in various complexes make it versatile [70]. Although laccase effectively degrades emerging contaminants, its application on a large scale necessitates features like reusability, thermostability, and operational stability. Achieving these characteristics often involves techniques such as immobilization and the production or isolation of robust laccase variants with desired attributes.
Laccase, also identified as benzenediol:oxygen oxidoreductase (EC 1.10.3.2), stands as a crucial enzyme with a central role in diverse biological processes. This copper-containing enzyme serves as a catalyst for oxidizing a wide array of organic and inorganic compounds, facilitating the reduction of oxygen molecules to produce water as a byproduct. Its versatility in mediating oxidative reactions has sparked considerable interest in scientific research and industrial applications across various fields, including biotechnology, environmental science, and agriculture (Figure 2). In this context, there is a pressing need to delve deeper into the properties, functions, and applications of laccase, exploring its significance in different domains.
Laccase exhibits a broad distribution in nature, being present in bacteria, fungi, plants, and insects. This widespread occurrence underscores the fundamental importance of laccase in nature and its relevance in various biological and ecological contexts. The diverse functions of laccases are contingent upon their source organism, contributing to processes such as lignin degradation, pigmentation, fruiting body formation, fungal morphogenesis, detoxification, sporulation, and pathogenesis in fungi; melanin formation, and endospore coat protein synthesis in bacteria; and lignification, wound healing, and iron oxidation in plants [71,72,73,74,75,76]. Thermostable laccases are advantageous for the eco-friendly remediation of hazardous synthetic dyes, particularly in the treatment of high-temperature dyeing wastewater. Table 3 lists some commercially available laccases with their industrial applications.
Examples of laccase sources reported from bacteria include Azospirrullum lipoferum [77], Anabaena azollae [78], Bacillus subtilis [79], Streptomyces cyaneus [80], S. lavendulae [81], and Marinomonas mediterranea [82]. In fungi, laccases are widely distributed in ascomycetes, basidiomycetes, and deuteromycetes, encompassing various species such as Trametes (Coriolus) versicolor, T. hirsute, T. ochracea, T. villosa, T. gallica, Cerrena maxima, Phlebia radiata, Coriolopsis polyzona, Lentinus tigrinus, Pleurotus eryngii, Myceliophthora thermophila, Aspergillus, Curvularia, Penicillium, Chaetomium thermophile, Mycelia sterlia, Cantharellus cibarius, Lactarius piperatus, Russula delica, Thelephora terestris, Agaricus, Marasmius, Tricholoma, and Volvariella [83].
In contrast to fungal laccases, bacterial laccases exhibit higher activity and greater stability at elevated temperatures, high pH, and high concentrations of chloride and copper ions [84,85,86]. Laccases have also been reported in various plants, including Japanese lacquer tree (Rhus vernicifera), lacquer, mango, mung bean, peach, pine, prune, sycamore, Pinus taeda, Populus trichocarpa, Acer pseudoplatanus, Liriodendron tulipifera, Zinnia elegans, tobacco (Nicotiana tabacum), Zea mays, Lolium perenne, Leucaena leucocephala, and Carica papaya. Plant laccases generally exhibit a higher molecular mass compared to fungal laccases, attributed to the increased glycosylation in plant laccases (22-45%) compared to fungal counterparts (10-25%) [87]. Glycosylation significantly influences various aspects of laccase functionality, including copper retention, thermal stability, and enzymatic activity. Some sources of thermostable laccases with industrial applications is listed in Table 4.
Laccase has gained attention for its versatile applications in biotechnological fields, including dye decolorization, biopulping, biobleaching, xenobiotic degradation, food processing, biopolymer modification, ethanol production, biosensor development, drug and organic synthesis, among others [83]. However, a common challenge with many isolated laccases is their relatively low enzyme activity yield and sensitivity to extreme conditions like temperature, pH, and metal ions. These limitations can impede their widespread use in large-scale commercial and industrial applications. Addressing these challenges, thermostability emerges as a crucial attribute. Thermostable laccases not only facilitate enzyme reactions at elevated temperatures with enhanced rates but also mitigate the risk of microbial contamination. They are particularly valuable in applications such as biobleaching of pulp and the treatment of colored industrial effluents. Thermostable laccases have been predominantly reported from thermophilic bacteria and fungi. Numerous approaches are currently under investigation to enhance laccase activity and mitigate thermal enzyme inactivation. While common strategies involve chemical modifications and enzyme immobilization on solid supports, these methods often pose challenges in terms of synthetic complexity and sustainability, leading to high costs. Recent studies explore the use of co-solvents like polyethylene glycol and ionic liquids, showing promise in improving reaction kinetics for enzymes like alcohol dehydrogenase and α-chymotrypsin. Additionally, deep eutectic solvents (DESs) have emerged as environmentally friendly alternatives for biotechnological applications, owing to their compatibility with enzymes [88]. DESs typically consist of a hydrogen bond acceptor (HBA), such as a salt, and a hydrogen bond donor (HBD), like polyols and sugars. The interaction between HBA and HBD forms hydrogen bonds, resulting in DESs with a significantly lower melting temperature compared to their individual components. Noteworthy properties associated with DESs include low flammability, low vapor pressure, high solvability, low volatility, thermal and chemical stability, and broad polarity. A specific category within DESs, referred to as natural deep eutectic solvents (NADESs), is exclusively prepared using raw materials derived from natural sources, such as sugars and amino acids [89]. This natural origin makes NADESs suitable for various food-related applications, including applications in food packaging. Multiple patents have been filed predominately from fungi between 2008 and 2019, which are related to various applications and obtained from multiple sources.
Structural Characteristics and Catalytic Mechanism of Laccases
The three-dimensional structure of most of the laccases from bacteria, fungi, and plants reveals a polypeptide chain organized into three cupredoxin-like domains, domain 1, 2 and 3 arranged sequentially, as depicted in Figure 2 [83]. The function of domain 2 is to join and position domains 1 and 3, and a trinuclear cluster (TNC) is formed at the interface between domains 1 and 3. Laccases, being copper-containing glycoproteins, exist in either dimeric or tetrameric forms, with each monomer housing four copper atoms. These copper sites in laccases fall into three distinct groups: Type-1 (blue copper center), Type-2 (normal copper), and Type-3 (coupled binuclear copper centers) [90,91], each exhibiting characteristic electronic paramagnetic resonance (EPR) signals.
Type-1 copper centers coordinate with two histidines, one cysteine, and one methionine as ligands, imparting the deep blue color to laccases with an intense electronic absorption band near 600 nm (ε = 5000 M−1 cm−1). However, certain laccases, like the "white laccase" in Pleurotus ostreatus or 'yellow laccases,' exhibit variations in absorption due to altered oxidation states or the presence of additional elements [92,93]. Type-2 copper, coordinated by two histidines and a water molecule, lacks visible spectrum absorption and is typically positioned near Type-3 copper. Type-3 copper, coordinated by three histidines and a hydroxyl bridge, displays electron absorption around 330 nm. The structure and properties of these copper centers classify laccases into low-redox potential (bacteria and plants) and high-redox potential (basidiomycetes, especially white-rot fungi) categories, influencing their suitability for diverse applications.
The catalytic mechanism of laccase relies on the distributed copper atoms across three distinct centers (Figure 2). Three key steps characterize laccase-mediated catalysis:
- Type-1 Copper Reduction by Reducing Substrate: Laccase initiates the reaction by accepting electrons from the substrate, reducing the Type-1 copper center.
- Internal Electron Transfer: Electron transfer occurs from Type-1 to Type-2 and Type-3 copper centers, forming a trinuclear cluster.
- Reduction of Oxygen to Water: The trinuclear copper cluster reduces molecular oxygen to water, concluding the catalytic cycle.
The overall reaction for laccase catalysis can be summarized as: 4RH + O2 → 4R• + 2H2O (Equation 1) Here, RH represents the substrate molecules, and laccase oxidizes these substrates (4RH), generating free radicals (4R•) and reducing molecular oxygen (O2) to water (2H2O). This underscores laccases' pivotal role in generating free radicals for diverse reactions, including polymerization. The oxidative capabilities of laccases can be enhanced in the presence of specific low-molecular-weight compounds functioning as redox mediators, for example ABTS, 1-hydroxybenzotriazole (HBT), violuric acid, 2,2,6,6,-tetramethyl-1-piperidinyloxy free radical (TEMPO), etc. The combination of laccases with these compounds in ‘laccase-mediator systems’ has proven to significantly broaden the substrate range of the enzymes and enhance their efficiency in oxidizing resistant molecules or complex polymers [94].
Structure-Function Relationship among Laccases
Laccases, renowned for their capacity to catalyze the oxidation of various compounds, demonstrate versatility in function and industrial applications. Among their diverse functions, lignification/delignification stands out as particularly significant due to its involvement in various industrial processes such as pulp and paper manufacturing, biobleaching, bioenergy production, biomass conversion, biofuel production, and environmental pollutant removal. Laccases also play roles in polymerization/depolymerization of lignin, fungal pathogenesis, wound healing, sclerotization, morphogenesis, sporulation, pigmentation, fruiting body formation, melanin formation, and endospore coat protein synthesis [83,95,96,97,98,99].
In the plant lignification process, monolignols undergo polymerization through dehydrogenation, facilitated by enzymes including laccases found in the cell wall. Experimental studies reveal that laccases from various plant species efficiently oxidize monolignols, contributing to dehydrogenative polymer formation [100,101,102]. Laccase expression, mainly in the secondary xylem, has been reported in trees like Populus trichocarpa and Pinus taeda, suggesting involvement in plant lignin biosynthesis. Plant transformation studies using laccase gene constructs further support this involvement [103,104,105]. In fungi, laccases mediate lignin biodegradation, breaking down lignin polymer through oxidative processes, releasing phenolic compounds [106]. As discussed before, directed evolution stands as a potent protein engineering strategy, employing iterative cycles of random mutagenesis and selection under specific selective pressures. This approach aims to fine-tune the inherent characteristics of native enzymes, adapting them to the rigorous conditions of industrial operations or instilling them with new properties. Thus, by employing a combination of enzyme-directed evolution and rational design, successfully engineered fungal laccases, produced in yeast, has been investigated to function effectively under alkaline pH and high temperatures. These optimized conditions align with the requirements commonly found in the kraft process and the manufacture of fiberboard.
Despite sharing similar molecular architecture, plant and fungal laccases exhibit wide phylogenetic, physicochemical, and functional diversity [83,107,108]. Plant laccases participate in lignin biosynthesis, while fungal laccases are involved in lignin degradation or depolymerization. The redox potential of laccase is crucial, with fungal laccases (higher redox potential) capable of acting on both phenolic and non-phenolic subunits, contributing to lignin degradation. Plant laccases, with a lower redox potential, polymerize lignin by facilitating the coupling of phenoxy radicals [109].
pH dependence of fungal and plant laccases is proposed as a factor influencing their dual role in lignin degradation or synthesis [110,111]. Fungal laccases typically have low pH optima, adapted to acidic growth conditions, while plant laccases, being intracellular, have pH optima closer to the physiological range. Differences in pH optima may be linked to the dual function of laccases. Hakulinen and coworkers [112] have reported that the architectural differences at the C-terminal end of M. albomyces and T. versicolor laccases might be responsible for their role in lignification and delignification, respectively.
The three-dimensional structure of laccases, leading to an altered microenvironment at the enzyme's active site, is suggested as the basis for their dual action in lignin biosynthesis and degradation. Structural distinctions in the C-terminal region have been reported, contributing to the role of laccases in lignification and delignification. Computational studies using bioinformatic tools have provided insights into the molecular basis of lignin synthesis and degradation, offering valuable information for future strategies aiming to modify laccase structure in plants and fungi to improve lignin biosynthesis and biodegradability [113].
Major Strategies to Enhance Thermostability
α-Amylase serves as a crucial industrial biocatalyst in the process of starch liquefaction and also stands as a significant model enzyme for exploring thermal adaptation in proteins. Presently, there's a surge in demand for enzymes, particularly those adaptable to industrial applications, prompting researchers to delve into diverse sources like metagenomes [114]. Despite this, bacterial sources continue to dominate the industrial landscape due to their diversity and requisite properties, notably stability and functionality at high temperatures commonly encountered in industrial processes [115]. Enhancing the thermostability of an enzyme primarily involves three strategies. The first entails sourcing extremophiles in hopes of enzyme behavior mirroring that of its host. The second strategy involves shielding the enzyme structure through immobilization on suitable matrices like cloisites or via the addition of certain cations, crowding agents, and deep eutectic solvents [116,117,118]. However, both strategies have inherent limitations, such as the rarity of finding natural sources with desired industrial-grade properties and the inability of many enzymes to be stabilized using additives. Thus, consideration of a third strategy involving protein structure design or protein engineering to meet thermal stability demands arises [119]. This involves modifying key features of protein structure critical for thermal adaptation, including enhancing rigidity, reducing loop length, optimizing core packing, intensifying surface hydration, and designing stabilizing interactions like salt bridges and hydrophobic interactions.
Numerous studies have explored the enhancement of enzyme thermostability through protein engineering. Protein engineering on hotspot residues is recognized as a highly effective strategy for enhancing both the stability and activity of enzymes. Computer modeling was employed to delve deeper into the structural underpinnings of the variance in thermostability between the wild-type enzyme and its variants. Thus, suggesting that augmenting the number of salt bridges and hydrophobic interactions surrounding Lys209 serves as the primary mechanism driving the enhanced compactness of the enzyme's protein structure [120]. Yuan’s group [121] studies on improving thermostability of Bacillus amyloliquefaciens alpha-amylase by multipoint mutations. Rational protein design, a top-down approach, aids in identifying necessary modifications for achieving thermal stability but is hindered by the extensive information required for reliable predictions for each protein. An alternative approach to circumvent these limitations involves random alteration of protein structure, such as random mutagenesis, recombination, and targeted mutagenesis combined with computational biology, termed Computer-Aided Directed Evolution of Enzymes (CADEE). For instance, Suzuki’s group [24] utilized site-directed mutagenesis of the BAA gene, deleting Arg176 and Gly177 while substituting Ala for Lys269, to engineer a thermostable mutation.
Wang’s group [122] endeavored to enhance the thermostability of α-amylase through combinatorial coevolving-site saturation mutagenesis, a pivotal strategy in directed protein evolution. Similarly, a directed evolution approach using a combination of random and site-directed mutagenesis was adopted to enhance the laccase activity of Caldalkalibacillus thermarum strain TA2.A1 for its application in lignin degradation [123]. Li’s group [21] employed structure-based rational design to enhance the thermostability of AmyMH, the maltohexaose-forming α-amylase from Bacillus stearothermophilus, in the absence of added Ca2+. Thermostability of α-amylase is enhanced upon mutatiting S187D/N188T, A269K/S187D, and A269K/S187D/N188T via site-directed mutagenesis in B. licheniformis [124]. Through a systematic approach to enzyme engineering, which combined enzyme-directed evolution and rational design, Rodríguez-Escribano and coworkers [125] successfully altered the optimal pH of the laccase for lignin phenol oxidation from acidic to basic with an objective to cultivate laccases capable of operating under extreme conditions of high temperature and pH, characteristic of industrial wood conversion processes into kraft pulp and fiberboard. From a structural perspective, integrative strategies like enhancing substrate affinity, introducing electrostatic interactions, alleviating steric hindrance, increasing flexibility of the active site, N- and C-terminal engineering, and augmenting intramolecular and intermolecular hydrophobic interactions are well-established for improving both activity and thermostability [126].
Current Challenges, Research Aims and Recent Advances in the Field of Thermostable α-Amylases and Laccases
Among the various commercially available enzymes, α-amylases seem to be the most versatile enzymes in the industrial enzyme sector no doubt due to the abundance of starch, with the applications ranging from the conversion of starch to sugar syrups, and the production of cyclodextrins for the pharmaceutical industry. With increase in their application spectrum, the research is focused on developing new α-amylases with more thermophilic, thermotolerant and pH tolerant characteristics to improve starch gelatinization, decrease media viscosity, accelerate catalytic reactions and decrease the risks of bacterial contamination. The most thermostable α-amylase currently used in industrial processes is from Bacillus licheniformis [127]. It remains active for several hours at 90 °C. One extracellular enzyme from Pyrococcus woesei was isolated that is active between 40 °C and 130 °C with an optimum at 100 °C and pH 5.5 [128]. In spite of this, retaining high α-amylase activity at pH around 4.0 is still desired for industrial starch processing. But there seemed to be no great progress in essence, and it needs some huge technological advances. Nevertheless, the structural and dynamic features of amylase may give some inspiration to understand or improve other enzymes’ thermostability, as the heat resistance is always a subject of unfailing interest [27]. In spite of the great significance of α-amylase in biotechnology, the greatest challenge of ensuring its stability for economic viability has to be taken into account. To address this challenge, recent attention has been directed towards enhancing both the functionality and stability of α-amylase. Various emerging technologies, including irradiation, pulsed electric field, high pressure, and sonication, have been employed to improve its secondary structure, thermal stability, and overall performance thereby resulting in economic benefits. These latest technologies stated in a recent review article, offer potential avenues for enhancing the stability and efficiency of α-amylase, thereby contributing to its utility in various industrial processes [129]. By optimizing these techniques, researchers aim to overcome the limitations associated with α-amylase stability and unlock its full potential for applications in biotechnology and related fields. Similarly, the current utilization of laccases in industry appears to be restricted in comparison to their potential. Efforts in research should prioritize reducing production costs and enhancing the tools for precise control of reactions on specific polyphenols and other substrates targeted by these enzymes. Primary challenges related to the industrial deployment of laccases involve production expenses and the broad range of substrates they can act upon. While the extensive substrate diversity of laccases offers advantages for biodegradation purposes, it also presents hurdles in their commercial utilization within biocatalysis due to by-product formation resulting from free radical chemistry. Recent developments have introduced new areas of application such as plastic degradation, diagnostic tool development, among others. Therefore, it is reasonable to anticipate a rise in the number of patented innovations in the foreseeable future. Consequently, laccases are poised to penetrate a broader range of industrial sectors, potentially supplanting conventional methods with more environmentally sustainable production routes.
Certainly, addressing the significant expenses associated with laccase manufacturing and purification stands as a paramount challenge that needs resolution to facilitate the enzyme's widespread utilization [130]. Various strategies have been investigated to mitigate enzyme production costs, such as on-site manufacturing, utilizing economical raw materials for enzyme synthesis, exploring novel enzymes with improved activity rates and versatile characteristics, employing cost-effective purification methods, and immobilizing enzymes. In order to fully envision the utilization of laccase on an industrial scale, its activity must be accurately quantifiable on complex substrates and within intricate matrices. Therefore, it becomes imperative to reliably quantify laccase activity on such substrates. Industrial applications often involve substrates much more intricate than those typically assessed using spectrophotometry (by monitoring the appearance of a coloured oxidation product), such as lignin, effluents, and textile dyes. Moreover, the complexity of matrices and mixtures necessitates the development of alternative analytical methods. A recent study [131] outlined several methods, including FTIR, fluorimetry, calorimetry, electrochemistry, and electron paramagnetic resonance, which could address this challenge. However, substantial optimization efforts are essential in the upcoming years before these methods can be considered routine analyses.
Future Directions in the Field of Thermostable Enzymes
In order to utilize thermostable enzymes for industrial purposes, it’s crucial to produce the enzyme on a large scale at a low cost. However, the traditional approach to enzyme production and purification is time-consuming and inefficient. Therefore, employing cloning, purification, and over-expression techniques for such enzymes using a suitable expression system can effectively address this issue. It’s evident that further research is necessary in upcoming studies on thermostable enzymes to fully harness its industrial potential. There is significant potential for enhancing the thermal stability of enzymes. Studies that delve into assessing the thermostability of the mutant enzymes by examining disparities in model structures between the wild-type and mutant ones. Such analysis offers theoretical benchmarks for refining and developing thermostable enzymes. Additionally, introducing tailor-made approaches through systemic enzyme engineering, combining enzyme-directed evolution and rational design is an adapted endeavor nowadays aiming to furnish extremophilic biocatalysts capable of industrial applications. The generic techniques of protein purification and recovery that includes filtration followed by membrane ultrafiltration, precipitation followed by dialysis, freeze and thaw followed by centrifugation and chromatographic techniques demands several steps which are expensive and demand considerable time and energy [132]. Thus, there is an ample opportunity for enhancement in this connection. Nonetheless, encountering challenges persists in discovering a new enzyme with verified activity, largely due to the fact that many proteins are forecasted solely on sequence similarity, leaving their functions hypothetical. It is imperative to experimentally characterize predicted proteins to ascertain sequence-to-function correlations.
Given the biotechnological significance of α-amylase, its substantial stability represents a paramount challenge for ensuring its economic feasibility. Hence, there has been a surge in interest in enhancing both its functionality and stability. In pursuit of this objective, a recent review has outlined the utilization of a blend of emerging technologies alongside traditional approaches on α-amylases from diverse sources.
Environmental and health concerns in chemical hair dyeing, such as, laccases have attracted considerable interest due to their capability for cross-coupling polymerization of phenolic monomers and their high oxidation potential. For instance, a thermostable bacterial laccase derived from Brevibacillus agri (LacT) has demonstrated significant potential for widespread utilization in the hair dyeing industry as a substitute for conventional chemical hair dyes [133].
With the emerging advances in thermostable laccase and its current application in lignin-first, in future research should focus more on the interaction between thermostable laccases and lignin substrates. At present, the utilization of thermostable laccases exhibiting exceptional characteristics in various environments, their applications have mainly been confined to textile industry, paper industry and the oxidation of small molecular substrates. To date, only a handful of thermostable laccases have been employed in reactions involving macromolecular lignin, with thorough investigations into their mechanisms still lacking, thus demanding more extensive research on this. Moreover, for the specialized discovery of laccases, there is a necessity for further exploration and enhancement of methods in involving metagenomic DNA extraction or enrichment from thermal environments.
Concluding Remarks
Thermostable enzymes not only provide valuable insights into the thermodynamic stability of proteins from a fundamental standpoint but also contribute to understanding the intricate relationship between stability, flexibility, and catalytic efficiency. α-Amylases and laccases, considered as exemplary representatives of thermostable enzymes, hold immense significance in biotechnological and industrial applications. The heightened interest in these enzymes has sparked intensified research endeavors aiming to discover new and improved variants of α-amylases and laccases. Consequently, the current scenario emphasizes the need to explore additional sources of thermostable enzymes or enhance the thermotolerance of existing enzymes through genetic modifications or site-directed mutagenesis, with the goal of achieving specific and desired properties in these enzymes.
Acknowledgements
The guidance and mentorship of my supervisors and mentors, and the support from University Grants Commission, New Delhi; Dr. D.S. Kothari Postdoctoral Program, Pune; and Department of Science and Technology-Science and Engineering Research Board, New Delhi, India, is gratefully acknowledged.
Compliance with ethical standards
Disclosure of potential conflicts of interest:
he authors declare that they have no conflict of interest in the publication.
Research involving human participants and/or animals
This article does not contain any studies with human participants or animals performed by any of the authors.
Informed consent
Not applicable.
References
- Vieille, C.; Zeikus Gregory, J. Hyperthermophilic Enzymes: Sources, Uses, and Molecular Mechanisms for Thermostability. Microbiology and Molecular Biology Reviews 2001, 65, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Che Hussian, C.H.A.; Leong, W.Y. Thermostable enzyme research advances: a bibliometric analysis. Journal of Genetic Engineering and Biotechnology 2023, 21, 37. [Google Scholar] [CrossRef] [PubMed]
- Mateljak, I.; Alcalde, M. Engineering a Highly Thermostable High-Redox Potential Laccase. ACS Sustainable Chemistry & Engineering 2021, 9, 9632–9637. [Google Scholar] [CrossRef]
- Ferreira, A.V.F.; Silva, F.F.; Silva, A.A.M.; Azevedo, L.S.; da Fonseca, S.T.D.; Camilo, N.H.; Dos Santos, K.P.E.; de Carvalho, L.C.; Tarabal, V.S.; da Silva, J.O.; et al. Recent Patents on the Industrial Application of Alpha-amylases. Recent Pat Biotechnol 2020, 14, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Da Lage, J.L.; Danchin, E.G.; Casane, D. Where do animal alpha-amylases come from? An interkingdom trip. FEBS Lett 2007, 581, 3927–3935. [Google Scholar] [CrossRef]
- Guo, L.; Liang, Z.; Zheng, C.; Liu, B.; Yin, Q.; Cao, Y.; Yao, J. Leucine Affects α-Amylase Synthesis through PI3K/Akt-mTOR Signaling Pathways in Pancreatic Acinar Cells of Dairy Calves. J Agric Food Chem 2018, 66, 5149–5156. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, P.; Kimmel, A.R. mTORC1/AMPK responses define a core gene set for developmental cell fate switching. BMC Biol 2019, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, P.; Majithia, A.R.; Rosel, D.; Liao, X.H.; Khurana, T.; Kimmel, A.R. Integrated actions of mTOR complexes 1 and 2 for growth and development of Dictyostelium. Int J Dev Biol 2019, 63, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Gigras, P.; Mohapatra, H.; Goswami, V.K.; Chauhan, B. Microbial α-amylases: a biotechnological perspective. Process Biochemistry 2003, 38, 1599–1616. [Google Scholar] [CrossRef]
- Sindhu, R.; Binod, P.; Madhavan, A.; Beevi, U.S.; Mathew, A.K.; Abraham, A.; Pandey, A.; Kumar, V. Molecular improvements in microbial α-amylases for enhanced stability and catalytic efficiency. Bioresour Technol 2017, 245, 1740–1748. [Google Scholar] [CrossRef]
- Prakash, O.; Jaiswal, N. alpha-Amylase: an ideal representative of thermostable enzymes. Appl Biochem Biotechnol 2010, 160, 2401–2414. [Google Scholar] [CrossRef] [PubMed]
- Prakash, O.; Jaiswal, N. A highly efficient and thermostable α-amylase from soya bean seeds. Biotechnol Appl Biochem 2010, 57, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Muralikrishna, G.; Nirmala, M. Cereal α-amylases—an overview. Carbohydrate Polymers 2005, 60, 163–173. [Google Scholar] [CrossRef]
- Singh, K.; Ahmad, F.; Singh, V.K.; Kayastha, K.; Kayastha, A.M. Purification, biochemical characterization and Insilico modeling of α-amylase from Vicia faba. Journal of Molecular Liquids 2017, 234, 133–141. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: functions and literature. Nucleic Acids Res 2022, 50, D571–d577. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Research 2014, 42, D490–D495. [Google Scholar] [CrossRef]
- Mareček, F.; Janeček, Š. A Novel Subfamily GH13_46 of the α-Amylase Family GH13 Represented by the Cyclomaltodextrinase from Flavobacterium sp. No. 92. Molecules 2022, 27, 8735. [Google Scholar] [CrossRef]
- Janeček, Š. Advances in Amylases—What’s Going on? Molecules 2023, 28, 7268. [Google Scholar] [CrossRef]
- Janeček, Š.; Svensson, B. How many α-amylase GH families are there in the CAZy database? 6, -10. [CrossRef]
- Ben Mabrouk, S.; Aghajari, N.; Ben Ali, M.; Ben Messaoud, E.; Juy, M.; Haser, R.; Bejar, S. Enhancement of the thermostability of the maltogenic amylase MAUS149 by Gly312Ala and Lys436Arg substitutions. Bioresour Technol 2011, 102, 1740–1746. [Google Scholar] [CrossRef]
- Li, Z.; Duan, X.; Wu, J. Improving the thermostability and enhancing the Ca(2+) binding of the maltohexaose-forming α-amylase from Bacillus stearothermophilus. J Biotechnol 2016, 222, 65–72. [Google Scholar] [CrossRef]
- Desse Haki, G.; Anceno, A.; Rakshit, S. Atypical Ca2+-independent, raw-starch hydrolysing α-amylase from Bacillus sp. GRE1: Characterization and gene isolation. World Journal of Microbiology and Biotechnology 2008, 24, 2517–2524. [Google Scholar] [CrossRef]
- Haki, G.D.; Rakshit, S.K. Developments in industrially important thermostable enzymes: a review. Bioresour Technol 2003, 89, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Ito, N.; Yuuki, T.; Yamagata, H.; Udaka, S. Amino Acid Residues Stabilizing a Bacillus α-Amylase against Irreversible Thermoinactivation*. Journal of Biological Chemistry 1989, 264, 18933–18938. [Google Scholar] [CrossRef] [PubMed]
- Khemakhem, B.; Ali, M.B.; Aghajari, N.; Juy, M.; Haser, R.; Bejar, S. The importance of an extra loop in the B-domain of an α-amylase from B. stearothermophilus US100. Biochemical and Biophysical Research Communications 2009, 385, 78–83. [Google Scholar] [CrossRef]
- Ben Ali, M.; Khemakhem, B.; Robert, X.; Haser, R.; Bejar, S. Thermostability enhancement and change in starch hydrolysis profile of the maltohexaose-forming amylase of Bacillus stearothermophilus US100 strain. Biochem J 2006, 394, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Declerck, N.; Machius, M.; Wiegand, G.; Huber, R.; Gaillardin, C. Probing structural determinants specifying high thermostability in Bacillus licheniformis alpha-amylase. J Mol Biol 2000, 301, 1041–1057. [Google Scholar] [CrossRef]
- Choi, J.M.; Han, S.S.; Kim, H.S. Industrial applications of enzyme biocatalysis: Current status and future aspects. Biotechnol Adv 2015, 33, 1443–1454. [Google Scholar] [CrossRef]
- Sivaramakrishnan, S.; Gangadharan, D.; Nampoothiri, K.M.; Soccol, C.R.; Pandey, A. α-Amylases from Microbial Sources -- An Overview on Recent Developments. Food Technology & Biotechnology 2006, 44, 173–184. [Google Scholar]
- Greenwood, C.T.; MacGregor, A.W. THE ISOLATION OF α-AMYLASE FROM BARLEY AND MALTED BARLEY, AND A STUDY OF THE PROPERTIES AND ACTION-PATTERNS OF THE ENZYMES. Journal of the Institute of Brewing 1965, 71, 405–417. [Google Scholar] [CrossRef]
- Tripathi, P.; Lo Leggio, L.; Mansfeld, J.; Ulbrich-Hofmann, R.; Kayastha, A.M. Alpha-amylase from mung beans (Vigna radiata)--correlation of biochemical properties and tertiary structure by homology modelling. Phytochemistry 2007, 68, 1623–1631. [Google Scholar] [CrossRef]
- Nakamura, Y.; Koizumi, R.; Uchino, M.; Sato, H.; Takano, K. Purification and Characterization of α-Amylase from Potato Tuber. Food Preservation Science 2010, 36, 271–276. [Google Scholar] [CrossRef]
- Sarker, G.K.; Hasan, S.; Nikkon, F.; Mosaddik, A.; Sana, N.K.; Rahman, H.; Park, S.; Lee, D.-S.; Cho, S.K. Purification, characterization, and biochemical properties of α-amylase from potato. Journal of the Korean Society for Applied Biological Chemistry 2010, 53, 8–14. [Google Scholar] [CrossRef]
- Khare, S.; Prakash, O. Purification and Biochemical Characterization of α-Amylase from Radish (Raphanus sativus L.) Seeds Using Response surface methodology. Proceedings of the National Academy of Sciences, India Section B: Biological Sciences 2019, 89, 79–88. [Google Scholar] [CrossRef]
- Amid, M.; Manap, M.Y.A. Purification and characterisation of a novel amylase enzyme from red pitaya (Hylocereus polyrhizus) peel. Food Chemistry 2014, 165, 412–418. [Google Scholar] [CrossRef]
- Singh, K.; Kayastha, A.M. A-amylase from wheat (Triticum aestivum) seeds: its purification, biochemical attributes and active site studies. Food Chem 2014, 162, 1–9. [Google Scholar] [CrossRef]
- Posoongnoen, S.; Thummavongsa, T. Purification and characterization of thermostable α-amylase from germinating Sword bean (Canavalia gladiata (Jacq.) DC.) seeds. Plant Biotechnol (Tokyo) 2020, 37, 31–38. [Google Scholar] [CrossRef]
- Banner, D.W.; Bloomer, A.C.; Petsko, G.A.; Phillips, D.C.; Pogson, C.I.; Wilson, I.A.; Corran, P.H.; Furth, A.J.; Milman, J.D.; Offord, R.E.; et al. Structure of chicken muscle triose phosphate isomerase determined crystallographically at 2.5Å resolution: using amino acid sequence data. Nature 1975, 255, 609–614. [Google Scholar] [CrossRef]
- Svensson, B.; Søgaard, M. Protein engineering of amylases. Biochem Soc Trans 1992, 20, 34–42. [Google Scholar] [CrossRef]
- MacGregor, E.A. α-Amylase structure and activity. Journal of Protein Chemistry 1988, 7, 399–415. [Google Scholar] [CrossRef]
- Nielsen, J.E.; Borchert, T.V. Protein engineering of bacterial α-amylases. Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology 2000, 1543, 253–274. [Google Scholar] [CrossRef]
- van der Maarel, M.J.; van der Veen, B.; Uitdehaag, J.C.; Leemhuis, H.; Dijkhuizen, L. Properties and applications of starch-converting enzymes of the alpha-amylase family. J Biotechnol 2002, 94, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Zhang, Z.-L.; Hanzlik, S.; Cook, E.; Shen, Q.J. Salicylic acid inhibits gibberellin-induced alpha-amylase expression and seed germination via a pathway involving an abscisic-acid-inducible WRKY gene. Plant Molecular Biology 2007, 64, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Kuriki, T.; Imanaka, T. The concept of the alpha-amylase family: structural similarity and common catalytic mechanism. J Biosci Bioeng 1999, 87, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Vihinen, M.; Mäntsälä, P. Microbial amylolytic enzymes. Crit Rev Biochem Mol Biol 1989, 24, 329–418. [Google Scholar] [CrossRef] [PubMed]
- Violet, M.; Meunier, J.C. Kinetic study of the irreversible thermal denaturation of Bacillus licheniformis alpha-amylase. Biochem J 1989, 263, 665–670. [Google Scholar] [CrossRef]
- Hwang, K.Y.; Song, H.K.; Chang, C.; Lee, J.; Lee, S.Y.; Kim, K.K.; Choe, S.; Sweet, R.M.; Suh, S.W. Crystal structure of thermostable alpha-amylase from Bacillus licheniformis refined at 1.7 A resolution. Mol Cells 1997, 7, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Machius, M.; Wiegand, G.; Huber, R. Crystal structure of calcium-depleted Bacillus licheniformis alpha-amylase at 2.2 A resolution. J Mol Biol 1995, 246, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Stein, E.A.; Sumerwell, W.N.; Fischer, E.H. Metal content of alpha-amylases of various origins. J Biol Chem 1959, 234, 2901–2905. [Google Scholar] [CrossRef]
- Boel, E.; Brady, L.; Brzozowski, A.M.; Derewenda, Z.; Dodson, G.G.; Jensen, V.J.; Petersen, S.B.; Swift, H.; Thim, L.; Woldike, H.F. Calcium binding in alpha-amylases: an X-ray diffraction study at 2.1-A resolution of two enzymes from Aspergillus. Biochemistry 1990, 29, 6244–6249. [Google Scholar] [CrossRef]
- Hsiu, J.; Fischer, E.H.; Stein, E.A. Alpha-Amylases as Calcium-Metalloenzymes. II. Calcium and the Catalytic Activity. Biochemistry 1964, 3, 61–66. [Google Scholar] [CrossRef]
- Larson, S.B.; Greenwood, A.; Cascio, D.; Day, J.; McPherson, A. Refined molecular structure of pig pancreatic alpha-amylase at 2.1 A resolution. J Mol Biol 1994, 235, 1560–1584. [Google Scholar] [CrossRef] [PubMed]
- Buisson, G.; Duée, E.; Haser, R.; Payan, F. Three dimensional structure of porcine pancreatic alpha-amylase at 2.9 A resolution. Role of calcium in structure and activity. The EMBO Journal 1987, 6, 3909–3916. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.X.; Wang, Y.B.; Pan, Y.J.; Li, W.F. Differences in amino acids composition and coupling patterns between mesophilic and thermophilic proteins. Amino Acids 2008, 34, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.A.; Bort, B.R.; Martínez, J.; Randez-Gil, F.; Buesa, C.; Sanz, P. Purification and characterization of a new alpha-amylase of intermediate thermal stability from the yeast Lipomyces kononenkoae. Biochem Cell Biol 1995, 73, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Declerck, N.; Machius, M.; Joyet, P.; Wiegand, G.; Huber, R.; Gaillardin, C. Hyperthermostabilization of Bacillus licheniformis alpha-amylase and modulation of its stability over a 50 degrees C temperature range. Protein Eng 2003, 16, 287–293. [Google Scholar] [CrossRef]
- Torrance, J.W.; MacArthur, M.W.; Thornton, J.M. Evolution of binding sites for zinc and calcium ions playing structural roles. Proteins: Structure, Function, and Bioinformatics 2008, 71, 813–830. [Google Scholar] [CrossRef] [PubMed]
- Goyal, N.; Gupta, J.K.; Soni, S.K. A novel raw starch digesting thermostable α-amylase from Bacillus sp. I-3 and its use in the direct hydrolysis of raw potato starch. Enzyme and Microbial Technology 2005, 37, 723–734. [Google Scholar] [CrossRef]
- Khajeh, K.; Ranjbar, B.; Naderi-Manesh, H.; Ebrahim Habibi, A.; Nemat-Gorgani, M. Chemical modification of bacterial alpha-amylases: changes in tertiary structures and the effect of additional calcium. Biochim Biophys Acta 2001, 1548, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Bush, D.S.; Sticher, L.; van Huystee, R.; Wagner, D.; Jones, R.L. The calcium requirement for stability and enzymatic activity of two isoforms of barley aleurone alpha-amylase. J Biol Chem 1989, 264, 19392–19398. [Google Scholar] [CrossRef] [PubMed]
- Hmidet, N.; Bayoudh, A.; Berrin, J.G.; Kanoun, S.; Juge, N.; Nasri, M. Purification and biochemical characterization of a novel α-amylase from Bacillus licheniformis NH1: Cloning, nucleotide sequence and expression of amyN gene in Escherichia coli. Process Biochemistry 2008, 43, 499–510. [Google Scholar] [CrossRef]
- Asoodeh, A.; Chamani, J.; Lagzian, M. A novel thermostable, acidophilic α-amylase from a new thermophilic “Bacillus sp. Ferdowsicous” isolated from Ferdows hot mineral spring in Iran: Purification and biochemical characterization. International Journal of Biological Macromolecules 2010, 46, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Satyanarayana, T. High maltose-forming, Ca2+-independent and acid stable α-amylase from a novel acidophilic bacterium, Bacillus acidicola. Biotechnol Lett 2010, 32, 1503–1507. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, R.; Noorwez, S.M.; Satyanarayana, T. Production and partial characterization of thermostable and calcium-independent alpha-amylase of an extreme thermophile Bacillus thermooleovorans NP54. Lett Appl Microbiol 2000, 31, 378–384. [Google Scholar] [CrossRef]
- Tanaka, A.; Hoshino, E. Secondary calcium-binding parameter of Bacillus amyloliquefaciens alpha-amylase obtained from inhibition kinetics. J Biosci Bioeng 2003, 96, 262–267. [Google Scholar] [CrossRef]
- Mehta, D.; Satyanarayana, T. Biochemical and molecular characterization of recombinant acidic and thermostable raw-starch hydrolysing α-amylase from an extreme thermophile Geobacillus thermoleovorans. Journal of Molecular Catalysis B: Enzymatic 2013, 85-86, 229–238. [Google Scholar] [CrossRef]
- Liao, S.M.; Liang, G.; Zhu, J.; Lu, B.; Peng, L.X.; Wang, Q.Y.; Wei, Y.T.; Zhou, G.P.; Huang, R.B. Influence of Calcium Ions on the Thermal Characteristics of α-amylase from Thermophilic Anoxybacillus sp. GXS-BL. Protein Pept Lett 2019, 26, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.; Wyss, M. Engineering proteins for thermostability: the use of sequence alignments versus rational design and directed evolution. Current Opinion in Biotechnology 2001, 12, 371–375. [Google Scholar] [CrossRef]
- Arnold, F.H. Engineering proteins for nonnatural environments. The FASEB Journal 1993, 7, 744–749. [Google Scholar] [CrossRef]
- Bibi, M.; Yasmin, A.; Blanford, C.; Safdar, N. Production and characterization of a highly stable laccase from extreme thermophile Geobacillus stearothermophilus MB600 isolated from hot spring of Gilgit Balitistan (Pakistan). Journal of Taibah University for Science 2023, 17, 2268903. [Google Scholar] [CrossRef]
- Hoopes, J.T.; Dean, J.F.D. Ferroxidase activity in a laccase-like multicopper oxidase from Liriodendron tulipifera. Plant Physiology and Biochemistry 2004, 42, 27–33. [Google Scholar] [CrossRef]
- McCaig, B.C.; Meagher, R.B.; Dean, J.F. Gene structure and molecular analysis of the laccase-like multicopper oxidase (LMCO) gene family in Arabidopsis thaliana. Planta 2005, 221, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Leatham, G.F.; Stahmann, M.A. Studies on the Laccase of Lentinus edodes: Specificity, Localization and Association with the Development of Fruiting Bodies. Microbiology 1981, 125, 147–157. [Google Scholar] [CrossRef]
- Thurston, C.F. The structure and function of fungal laccases. Microbiology 1994, 140, 19–26. [Google Scholar] [CrossRef]
- Nagai, M.; Kawata, M.; Watanabe, H.; Ogawa, M.; Saito, K.; Takesawa, T.; Kanda, K.; Sato, T. Important role of fungal intracellular laccase for melanin synthesis: purification and characterization of an intracellular laccase from Lentinula edodes fruit bodies. Microbiology 2003, 149, 2455–2462. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, K.; Streibel, M.; Jahn, B.; Haase, G.; Brakhage, A.A. Biosynthesis of fungal melanins and their importance for human pathogenic fungi. Fungal Genetics and Biology 2003, 38, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Givaudan, A.; Effosse, A.; Faure, D.; Potier, P.; Bouillant, M.-L.; Bally, R. Polyphenol oxidase in Azospirillum lipoferum isolated from rice rhizosphere: Evidence for laccase activity in non-motile strains of Azospirillum lipoferum. FEMS Microbiology Letters 1993, 108, 205–210. [Google Scholar] [CrossRef]
- Malliga, P.; Uma, L.; Subramanian, G. Lignolytic activity of the cyanobacterium Anabaena azollae ML2 and the value of coir waste as a carrier for BGA biofertilizer. Microbios 86.
- Martins, L.O.; Soares, C.M.; Pereira, M.M.; Teixeira, M.; Costa, T.; Jones, G.H.; Henriques, A.O. Molecular and biochemical characterization of a highly stable bacterial laccase that occurs as a structural component of the Bacillus subtilis endospore coat. J Biol Chem 2002, 277, 18849–18859. [Google Scholar] [CrossRef]
- Arias, M.E.; Arenas, M.; Rodríguez, J.; Soliveri, J.; Ball Andrew, S.; Hernández, M. Kraft Pulp Biobleaching and Mediated Oxidation of a Nonphenolic Substrate by Laccase from Streptomyces cyaneus CECT 3335. Applied and Environmental Microbiology 2003, 69, 1953–1958. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Endo, K.; Ito, M.; Tsujibo, H.; Miyamoto, K.; Inamori, Y. A thermostable laccase from Streptomyces lavendulae REN-7: purification, characterization, nucleotide sequence, and expression. Biosci Biotechnol Biochem 2003, 67, 2167–2175. [Google Scholar] [CrossRef]
- Solano, F.; Lucas-Elío, P.; Fernández, E.; Sanchez-Amat, A. Marinomonas mediterranea MMB-1 Transposon Mutagenesis: Isolation of a Multipotent Polyphenol Oxidase Mutant. Journal of Bacteriology 2000, 182, 3754–3760. [Google Scholar] [CrossRef]
- Dwivedi, U.N.; Singh, P.; Pandey, V.P.; Kumar, A. Structure–function relationship among bacterial, fungal and plant laccases. Journal of Molecular Catalysis B: Enzymatic 2011, 68, 117–128. [Google Scholar] [CrossRef]
- Held, C.; Kandelbauer, A.; Schroeder, M.; Cavaco-Paulo, A.; Guebitz, G.M. Biotransformation of phenolics with laccase containing bacterial spores. Environmental Chemistry Letters 2005, 3, 74–77. [Google Scholar] [CrossRef]
- Rosconi, F.; Fraguas, L.F.; Martínez-Drets, G.; Castro-Sowinski, S. Purification and characterization of a periplasmic laccase produced by Sinorhizobium meliloti. Enzyme and Microbial Technology 2005, 36, 800–807. [Google Scholar] [CrossRef]
- McMahon, A.M.; Doyle, E.M.; Brooks, S.; O’Connor, K.E. Biochemical characterisation of the coexisting tyrosinase and laccase in the soil bacterium Pseudomonas putida F6. Enzyme and Microbial Technology 2007, 40, 1435–1441. [Google Scholar] [CrossRef]
- Singh Arora, D.; Kumar Sharma, R. Ligninolytic fungal laccases and their biotechnological applications. Appl Biochem Biotechnol 2010, 160, 1760–1788. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.E.; Andanson, J.M.; Verney, V. Improving laccase thermostability with aqueous natural deep eutectic solvents. Int J Biol Macromol 2020, 163, 919–926. [Google Scholar] [CrossRef]
- Dai, Y.; van Spronsen, J.; Witkamp, G.J.; Verpoorte, R.; Choi, Y.H. Natural deep eutectic solvents as new potential media for green technology. Anal Chim Acta 2013, 766, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Messerschmidt, A.; Huber, R. The blue oxidases, ascorbate oxidase, laccase and ceruloplasmin Modelling and structural relationships. European Journal of Biochemistry 1990, 187, 341–352. [Google Scholar] [CrossRef]
- Solomon, E.I.; Baldwin, M.J.; Lowery, M.D. Electronic structures of active sites in copper proteins: contributions to reactivity. Chemical Reviews 1992, 92, 521–542. [Google Scholar] [CrossRef]
- Palmieri, G.; Cennamo, G.; Faraco, V.; Amoresano, A.; Sannia, G.; Giardina, P. Atypical laccase isoenzymes from copper supplemented Pleurotus ostreatus cultures. Enzyme and Microbial Technology 2003, 33, 220–230. [Google Scholar] [CrossRef]
- Leontievsky, A.A.; Vares, T.; Lankinen, P.; Shergill, J.K.; Pozdnyakova, N.N.; Myasoedova, N.M.; Kalkkinen, N.; Golovleva, L.A.; Cammack, R.; Thurston, C.F.; et al. Blue and yellow laccases of ligninolytic fungi. FEMS Microbiol Lett 1997, 156, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Aza, P.; Camarero, S. Fungal Laccases: Fundamentals, Engineering and Classification Update. Biomolecules 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Piontek, K.; Antorini, M.; Choinowski, T. Crystal structure of a laccase from the fungus Trametes versicolor at 1.90-A resolution containing a full complement of coppers. J Biol Chem 2002, 277, 37663–37669. [Google Scholar] [CrossRef] [PubMed]
- Gianfreda, L.; Xu, F.; Bollag, J.-M. Laccases: A Useful Group of Oxidoreductive Enzymes. Bioremediation Journal 1999, 3, 1–26. [Google Scholar] [CrossRef]
- Jaiswal, N.; Pandey, V.P.; Dwivedi, U.N. Purification of a thermostable laccase from Leucaena leucocephala using a copper alginate entrapment approach and the application of the laccase in dye decolorization. Process Biochemistry 2014, 49, 1196–1204. [Google Scholar] [CrossRef]
- Jaiswal, N.; Pandey, V.P.; Dwivedi, U.N. Purification of a thermostable alkaline laccase from papaya (Carica papaya) using affinity chromatography. Int J Biol Macromol 2015, 72, 326–332. [Google Scholar] [CrossRef]
- Jaiswal, N.; Pandey, V.P.; Dwivedi, U.N. Immobilization of papaya laccase in chitosan led to improved multipronged stability and dye discoloration. Int J Biol Macromol 2016, 86, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; O'Malley, D.M.; Whetten, R.; Sederoff, R.R. A Laccase Associated with Lignification in Loblolly Pine Xylem. Science 1993, 260, 672–674. [Google Scholar] [CrossRef]
- Liu, L.; Dean, J.F.D.; Friedman, W.E.; Eriksson, K.-E.L. A laccase-like phenoloxidase is correlated with lignin biosynthesis in Zinnia elegans stem tissues. The Plant Journal 1994, 6, 213–224. [Google Scholar] [CrossRef]
- Sterjiades, R.; Dean, J.F.; Eriksson, K.E. Laccase from Sycamore Maple (Acer pseudoplatanus) Polymerizes Monolignols. Plant Physiol 1992, 99, 1162–1168. [Google Scholar] [CrossRef]
- Ranocha, P.; McDougall, G.; Hawkins, S.; Sterjiades, R.; Borderies, G.; Stewart, D.; Cabanes-Macheteau, M.; Boudet, A.-M.; Goffner, D. Biochemical characterization, molecular cloning and expression of laccases – a divergent gene family – in poplar. European Journal of Biochemistry 1999, 259, 485–495. [Google Scholar] [CrossRef]
- Sato, Y.; Wuli, B.; Sederoff, R.; Whetten, R. Molecular Cloning and Expression of Eight Laccase cDNAs in Loblolly Pine (Pinus taeda)*. Journal of Plant Research 2001, 114, 147–155. [Google Scholar] [CrossRef]
- Ranocha, P.; Chabannes, M.; Chamayou, S.; Danoun, S.; Jauneau, A.; Boudet, A.M.; Goffner, D. Laccase down-regulation causes alterations in phenolic metabolism and cell wall structure in poplar. Plant Physiol 2002, 129, 145–155. [Google Scholar] [CrossRef]
- Camarero, S.; Galletti, G.C.; Martínez, A.T. Preferential degradation of phenolic lignin units by two white rot fungi. Appl Environ Microbiol 1994, 60, 4509–4516. [Google Scholar] [CrossRef] [PubMed]
- Colaneri, M.J.; Vitali, J. Copper dynamics in doped metal-bis(histidine) complexes. J Phys Chem A 2014, 118, 4688–4694. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, R.; Behera, R.; Padhi, S.K.; Guru, R.K. Computational Phylogenetic Study and Data Mining Approach to Laccase Enzyme Sequences. 2013.
- Li, K.; Xu, F.; Eriksson, K.E. Comparison of fungal laccases and redox mediators in oxidation of a nonphenolic lignin model compound. Appl Environ Microbiol 1999, 65, 2654–2660. [Google Scholar] [CrossRef] [PubMed]
- Gorbacheva, M.A.; Shumakovich, G.P.; Morozova, O.V.; Strel’tsov, A.V.; Zaitseva, E.A.; Shleev, S.V. Comparative study of biocatalytic reactions of high and low redox potential fungal and plant laccases in homogeneous and heterogeneous reactions. Moscow University Chemistry Bulletin 2008, 63, 94–98. [Google Scholar] [CrossRef]
- Madhavi; Lele, S. Laccase: Properties and Applications. BioResources 2009, 4. [Google Scholar] [CrossRef]
- Hakulinen, N.; Kiiskinen, L.-L.; Kruus, K.; Saloheimo, M.; Paananen, A.; Koivula, A.; Rouvinen, J. Crystal structure of a laccase from Melanocarpus albomyces with an intact trinuclear copper site. Nature Structural Biology 2002, 9, 601–605. [Google Scholar] [CrossRef]
- Awasthi, M.; Jaiswal, N.; Singh, S.; Pandey, V.P.; Dwivedi, U.N. Molecular docking and dynamics simulation analyses unraveling the differential enzymatic catalysis by plant and fungal laccases with respect to lignin biosynthesis and degradation. J Biomol Struct Dyn 2015, 33, 1835–1849. [Google Scholar] [CrossRef]
- Yun, J.; Kang, S.; Park, S.; Yoon, H.; Kim, M.-J.; Heu, S.; Ryu, S. Characterization of a Novel Amylolytic Enzyme Encoded by a Gene from a Soil-Derived Metagenomic Library. Applied and Environmental Microbiology 2004, 70, 7229–7235. [Google Scholar] [CrossRef] [PubMed]
- Mehta, D.; Satyanarayana, T. Bacterial and Archaeal α-Amylases: Diversity and Amelioration of the Desirable Characteristics for Industrial Applications. Frontiers in Microbiology 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Khozeymeh Nezhad, M.; Aghaei, H. Tosylated cloisite as a new heterofunctional carrier for covalent immobilization of lipase and its utilization for production of biodiesel from waste frying oil. Renewable Energy 2021, 164, 876–888. [Google Scholar] [CrossRef]
- Varriale, S.; Delorme, A.E.; Andanson, J.-M.; Devemy, J.; Malfreyt, P.; Verney, V.; Pezzella, C. Enhancing the Thermostability of Engineered Laccases in Aqueous Betaine-Based Natural Deep Eutectic Solvents. ACS Sustainable Chemistry & Engineering 2022, 10, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.E.; Andanson, J.-M.; Verney, V. Improving laccase thermostability with aqueous natural deep eutectic solvents. International Journal of Biological Macromolecules 2020, 163, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Pouyan, S.; Lagzian, M.; Sangtarash, M.H. Enhancing thermostabilization of a newly discovered α-amylase from Bacillus cereus GL96 by combining computer-aided directed evolution and site-directed mutagenesis. Int J Biol Macromol 2022, 197, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, L.; Xiao, Y.; Li, J.; Long, L.; Wang, F.; Zhang, S. Identification and thermoadaptation engineering of thermostability conferring residue of deep sea bacterial α-amylase AMY121. Journal of Molecular Catalysis B: Enzymatic 2016, 126, 56–63. [Google Scholar] [CrossRef]
- Yuan, S.; Yan, R.; Lin, B.; Li, R.; Ye, X. Improving thermostability of Bacillus amyloliquefaciens alpha-amylase by multipoint mutations. Biochem Biophys Res Commun 2023, 653, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Huang, R.; He, B.; Du, Q. Improving the thermostability of alpha-amylase by combinatorial coevolving-site saturation mutagenesis. BMC Bioinformatics 2012, 13, 263. [Google Scholar] [CrossRef]
- Yang, Y.; Ghatge, S.; Hur, H.-G. Improvement of thermoalkaliphilic laccase (CtLac) by a directed evolution and application to lignin degradation. Applied Microbiology and Biotechnology 2023, 107, 273–286. [Google Scholar] [CrossRef]
- Li, Z.; Duan, X.; Chen, S.; Wu, J. Improving the reversibility of thermal denaturation and catalytic efficiency of Bacillus licheniformis α-amylase through stabilizing a long loop in domain B. PLOS ONE 2017, 12, e0173187. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Escribano, D.; Pliego-Magán, R.; de Salas, F.; Aza, P.; Gentili, P.; Ihalainen, P.; Levée, T.; Meyer, V.; Petit-Conil, M.; Tapin-Lingua, S.; et al. Tailor-made alkaliphilic and thermostable fungal laccases for industrial wood processing. Biotechnology for Biofuels and Bioproducts 2022, 15, 149. [Google Scholar] [CrossRef] [PubMed]
- Nezhad, N.G.; Rahman, R.; Normi, Y.M.; Oslan, S.N.; Shariff, F.M.; Leow, T.C. Recent advances in simultaneous thermostability-activity improvement of industrial enzymes through structure modification. Int J Biol Macromol 2023, 232, 123440. [Google Scholar] [CrossRef] [PubMed]
- Declerck, N.; Machius, M.; Joyet, P.; Wiegand, G.; Huber, R.; Gaillardin, C. Hyperthermostabilization of Bacillus licheniformis α-amylase and modulation of its stability over a 50°C temperature range. Protein Engineering, Design and Selection 2003, 16, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Grzybowska, B.; Szweda, P.; Synowiecki, J. Cloning of the thermostable alpha-amylase gene from Pyrococcus woesei in Escherichia coli: isolation and some properties of the enzyme. Mol Biotechnol 2004, 26, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Abedi, E.; Kaveh, S.; Mohammad Bagher Hashemi, S. Structure-based modification of a-amylase by conventional and emerging technologies: Comparative study on the secondary structure, activity, thermal stability and amylolysis efficiency. Food Chem 2024, 437, 137903. [Google Scholar] [CrossRef] [PubMed]
- Idris, A.S.O.; Pandey, A.; Rao, S.S.; Sukumaran, R.K. Cellulase production through solid-state tray fermentation, and its use for bioethanol from sorghum stover. Bioresource Technology 2017, 242, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Dubessay, P.; Record, E.; Audonnet, F.; Michaud, P. Recent advances in laccase activity assays: A crucial challenge for applications on complex substrates. Enzyme Microb Technol 2024, 173, 110373. [Google Scholar] [CrossRef] [PubMed]
- Nadar, S.S.; Pawar, R.G.; Rathod, V.K. Recent advances in enzyme extraction strategies: A comprehensive review. Int J Biol Macromol 2017, 101, 931–957. [Google Scholar] [CrossRef]
- Panwar, V.; Dey, B.; Sheikh, J.N.; Dutta, T. Thermostable bacterial laccase for sustainable dyeing using plant phenols. RSC Advances 2022, 12, 18168–18180. [Google Scholar] [CrossRef]
- Chai, K.P.; Othman, N.F.B.; Teh, A.-H.; Ho, K.L.; Chan, K.-G.; Shamsir, M.S.; Goh, K.M.; Ng, C.L. Crystal structure of Anoxybacillus α-amylase provides insights into maltose binding of a new glycosyl hydrolase subclass. Scientific Reports 2016, 6, 23126. [Google Scholar] [CrossRef]
- Abdella, M.A.A.; El-Sherbiny, G.M.; El-Shamy, A.R.; Atalla, S.M.M.; Ahmed, S.A. Statistical optimization of chemical modification of chitosan-magnetic nano-particles beads to promote Bacillus subtilis MK1 α-amylase immobilization and its application. Bulletin of the National Research Centre 2020, 44, 40. [Google Scholar] [CrossRef]
- Enguita, F.J.; Martins, L.O.; Henriques, A.O.; Carrondo, M.A. Crystal structure of a bacterial endospore coat component. A laccase with enhanced thermostability properties. J Biol Chem 2003, 278, 19416–19425. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Solomon, E.I. Electron transfer and reaction mechanism of laccases. Cellular and Molecular Life Sciences 2015, 72, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Kherouf, M.; Habbeche, A.; Benamia, F.; Saoudi, B.; Kerouaz, B.; Ladjama, A. Statistical optimization of a novel extracellular alkaline and thermostable amylase production from thermophilic Actinomadura keratinilytica sp. Cpt29 and its potential application in detergent industry. Biocatalysis and Agricultural Biotechnology 2021, 35, 102068. [Google Scholar] [CrossRef]
- Timilsina, P.M.; Pandey, G.R.; Shrestha, A.; Ojha, M.; Karki, T.B. Purification and characterization of a noble thermostable algal starch liquefying alpha-amylase from Aeribacillus pallidus BTPS-2 isolated from geothermal spring of Nepal. Biotechnology Reports 2020, 28, e00551. [Google Scholar] [CrossRef] [PubMed]
- Slavić, M.Š.; Kojić, M.; Margetić, A.; Stanisavljević, N.; Gardijan, L.; Božić, N.; Vujčić, Z. Highly stable and versatile α-amylase from Anoxybacillus vranjensis ST4 suitable for various applications. International Journal of Biological Macromolecules 2023, 249, 126055. [Google Scholar] [CrossRef]
- Du, R.; Song, Q.; Zhang, Q.; Zhao, F.; Kim, R.C.; Zhou, Z.; Han, Y. Purification and characterization of novel thermostable and Ca-independent α-amylase produced by Bacillus amyloliquefaciens BH072. Int J Biol Macromol 2018, 115, 1151–1156. [Google Scholar] [CrossRef]
- Priyadarshini, S.; Pradhan, S.K.; Ray, P. Production, characterization and application of thermostable, alkaline α-amylase (AA11) from Bacillus cereus strain SP-CH11 isolated from Chilika Lake. Int J Biol Macromol 2020, 145, 804–812. [Google Scholar] [CrossRef]
- Afrisham, S.; Badoei-Dalfard, A.; Namaki-Shoushtari, A.; Karami, Z. Characterization of a thermostable, CaCl2-activated and raw-starch hydrolyzing alpha-amylase from Bacillus licheniformis AT70: Production under solid state fermentation by utilizing agricultural wastes. Journal of Molecular Catalysis B: Enzymatic 2016, 132, 98–106. [Google Scholar] [CrossRef]
- Hmidet, N.; El-Hadj Ali, N.; Haddar, A.; Kanoun, S.; Alya, S.-K.; Nasri, M. Alkaline proteases and thermostable α-amylase co-produced by Bacillus licheniformis NH1: Characterization and potential application as detergent additive. Biochemical Engineering Journal 2009, 47, 71–79. [Google Scholar] [CrossRef]
- Fincan, S.A.; Özdemir, S.; Karakaya, A.; Enez, B.; Mustafov, S.D.; Ulutaş, M.S.; Şen, F. Purification and characterization of thermostable α-amylase produced from Bacillus licheniformis So-B3 and its potential in hydrolyzing raw starch. Life Sciences 2021, 264, 118639. [Google Scholar] [CrossRef]
- Arikan, B. Highly thermostable, thermophilic, alkaline, SDS and chelator resistant amylase from a thermophilic Bacillus sp. isolate A3-15. Bioresour Technol 2008, 99, 3071–3076. [Google Scholar] [CrossRef]
- Gupta, N.; Paul, J.S.; Jadhav, S.K. Biovalorizing agro-waste 'de-oiled rice bran' for thermostable, alkalophilic and detergent stable α-amylase production with its application as laundry detergent additive and textile desizer. Int J Biol Macromol 2024, 256, 128470. [Google Scholar] [CrossRef]
- Prakash, B.; Vidyasagar, M.; Madhukumar, M.S.; Muralikrishna, G.; Sreeramulu, K. Production, purification, and characterization of two extremely halotolerant, thermostable, and alkali-stable α-amylases from Chromohalobacter sp. TVSP 101. Process Biochemistry 2009, 44, 210–215. [Google Scholar] [CrossRef]
- Shukla, R.J.; Singh, S.P. Characteristics and thermodynamics of α-amylase from thermophilic actinobacterium, Laceyella sacchari TSI-2. Process Biochemistry 2015, 50, 2128–2136. [Google Scholar] [CrossRef]
- Santorelli, M.; Maurelli, L.; Pocsfalvi, G.; Fiume, I.; Squillaci, G.; La Cara, F.; Del Monaco, G.; Morana, A. Isolation and characterisation of a novel alpha-amylase from the extreme haloarchaeon Haloterrigena turkmenica. Int J Biol Macromol 2016, 92, 174–184. [Google Scholar] [CrossRef]
- Michelin, M.; Silva, T.M.; Benassi, V.M.; Peixoto-Nogueira, S.C.; Moraes, L.A.B.; Leão, J.M.; Jorge, J.A.; Terenzi, H.F.; Polizeli, M.d.L.T.M. Purification and characterization of a thermostable α-amylase produced by the fungus Paecilomyces variotii. Carbohydrate Research 2010, 345, 2348–2353. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-c.; Hu, H.-f.; Ma, J.-w.; Yan, Q.-j.; Liu, H.-j.; Jiang, Z.-q. A novel high maltose-forming α-amylase from Rhizomucor miehei and its application in the food industry. Food Chemistry 2020, 305, 125447. [Google Scholar] [CrossRef]
- Edoamodu, C.E.; Nwodo, U.U. Enterobacter sp. AI1 produced a thermo-acidic-tolerant laccase with a high potential for textile dyes degradation. Biocatalysis and Agricultural Biotechnology 2021, 38, 102206. [Google Scholar] [CrossRef]
- Allala, F.; Bouacem, K.; Boucherba, N.; Azzouz, Z.; Mechri, S.; Sahnoun, M.; Benallaoua, S.; Hacene, H.; Jaouadi, B.; Bouanane-Darenfed, A. Purification, biochemical, and molecular characterization of a novel extracellular thermostable and alkaline α-amylase from Tepidimonas fonticaldi strain HB23. Int J Biol Macromol 2019, 132, 558–574. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Zhao, N.; Ma, J.W.; Liu, J.; Yan, Q.J.; Jiang, Z.Q. High-level expression of a novel α-amylase from Thermomyces dupontii in Pichia pastoris and its application in maltose syrup production. Int J Biol Macromol 2019, 127, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Dror, A.; Fishman, A. ENGINEERING NON-HEME MONO- AND DIOXYGENASES FOR BIOCATALYSIS. Computational and Structural Biotechnology Journal 2012, 2, e201209011. [Google Scholar] [CrossRef] [PubMed]
- Maphupha, M.; Juma, W.P.; de Koning, C.B.; Brady, D. A modern and practical laccase-catalysed route suitable for the synthesis of 2-arylbenzimidazoles and 2-arylbenzothiazoles. RSC Adv 2018, 8, 39496–39510. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Couto, S.; Lardizábal, P.M.d. Laccases for Denim Bleaching: An Eco-Friendly Alternative. The Open Textile Journal 2012, 5, 1–7. [Google Scholar] [CrossRef]
- de Salas, F.; Pardo, I.; Salavagione, H.J.; Aza, P.; Amougi, E.; Vind, J.; Martínez, A.T.; Camarero, S. Advanced Synthesis of Conductive Polyaniline Using Laccase as Biocatalyst. PLoS One 2016, 11, e0164958. [Google Scholar] [CrossRef]
- Call, H.P.; Mücke, I. History, overview and applications of mediated lignolytic systems, especially laccase-mediator-systems (Lignozym®-process). Journal of Biotechnology 1997, 53, 163–202. [Google Scholar] [CrossRef]
- Cindy; Mendoza, D. M.; Konadu, K.T.; Ichinose, H.; Sasaki, K. Multiple laccase-mediator system treatments for carbonaceous matter degradation in double refractory gold ore. Hydrometallurgy 2023, 221, 106129. [Google Scholar] [CrossRef]
- Wellington, K.W.; Govindjee, V.P.; Steenkamp, P. A laccase-catalysed synthesis of triaminated cyclohexa-2,4-dienones from catechol. Journal of Catalysis 2018, 368, 306–314. [Google Scholar] [CrossRef]
- Hämäläinen, V.; Grönroos, T.; Suonpää, A.; Heikkilä, M.W.; Romein, B.; Ihalainen, P.; Malandra, S.; Birikh, K.R. Enzymatic Processes to Unlock the Lignin Value. Front Bioeng Biotechnol 2018, 6, 20. [Google Scholar] [CrossRef]
- Othman, A.M.; Elsayed, M.A.; Elshafei, A.M.; Hassan, M.M. Purification and biochemical characterization of two isolated laccase isoforms from Agaricus bisporus CU13 and their potency in dye decolorization. Int J Biol Macromol 2018, 113, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Mehandia, S.; Sharma, S.C.; Arya, S.K. Isolation and characterization of an alkali and thermostable laccase from a novel Alcaligenes faecalis and its application in decolorization of synthetic dyes. Biotechnology Reports 2020, 25, e00413. [Google Scholar] [CrossRef] [PubMed]
- Diamantidis, G.; Effosse, A.; Potier, P.; Bally, R. Purification and characterization of the first bacterial laccase in the rhizospheric bacterium Azospirillum lipoferum. Soil Biology and Biochemistry 2000, 32, 919–927. [Google Scholar] [CrossRef]
- Khan, S.I.; Sahinkaya, M.; Colak, D.N.; Zada, N.S.; Uzuner, U.; Belduz, A.O.; Çanakçi, S.; Khan, A.Z.; Khan, S.; Badshah, M.; et al. Production and characterization of novel thermostable CotA-laccase from Bacillus altitudinis SL7 and its application for lignin degradation. Enzyme and Microbial Technology 2024, 172, 110329. [Google Scholar] [CrossRef]
- Sondhi, S.; Kaur, R.; Madan, J. Purification and characterization of a novel white highly thermo stable laccase from a novel Bacillus sp. MSK-01 having potential to be used as anticancer agent. Int J Biol Macromol 2021, 170, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Leung, I.K.H. Novel Thermophilic Bacterial Laccase for the Degradation of Aromatic Organic Pollutants. Frontiers in Chemistry 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Ayothiraman, S.; Dhakshinamoorthy, V. Production of highly thermo-tolerant laccase from novel thermophilic bacterium Bacillus sp. PC-3 and its application in functionalization of chitosan film. J Biosci Bioeng 2019, 127, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.M.; Patel, A.K.; Singhania, R.R.; Tsai, C.H.; Chen, S.Y.; Chen, C.W.; Dong, C.D. Heterologous expression of bacterial CotA-laccase, characterization and its application for biodegradation of malachite green. Bioresour Technol 2021, 340, 125708. [Google Scholar] [CrossRef] [PubMed]
- Basheer, S.; Rashid, N.; Akram, M.S.; Akhtar, M. A highly stable laccase from Bacillus subtilis strain R5: gene cloning and characterization. Biosci Biotechnol Biochem 2019, 83, 436–445. [Google Scholar] [CrossRef]
- Xu, G.; Wang, J.; Yin, Q.; Fang, W.; Xiao, Y.; Fang, Z. Expression of a thermo- and alkali-philic fungal laccase in Pichia pastoris and its application. Protein Expr Purif 2019, 154, 16–24. [Google Scholar] [CrossRef]
- Ibrahim, V.; Mendoza, L.; Mamo, G.; Hatti-Kaul, R. Blue laccase from Galerina sp.: Properties and potential for Kraft lignin demethylation. Process Biochemistry 2011, 46, 379–384. [Google Scholar] [CrossRef]
- Murugesan, K.; Nam, I.-H.; Kim, Y.-M.; Chang, Y.-S. Decolorization of reactive dyes by a thermostable laccase produced by Ganoderma lucidum in solid state culture. Enzyme and Microbial Technology 2007, 40, 1662–1672. [Google Scholar] [CrossRef]
- Alshiekheid, M.A.; Umar, A.; Ameen, F.; Alyahya, S.A.; Dufossé, L. Biodegradation of chromium by laccase action of Ganoderma multipileum. Journal of King Saud University - Science 2023, 35, 102948. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, L.; Guo, W.; Jia, L.; Fu, Y.; Gui, S.; Lu, F. Cloning, expression, and characterization of a thermostable and pH-stable laccase from Klebsiella pneumoniae and its application to dye decolorization. Process Biochemistry 2017, 53, 125–134. [Google Scholar] [CrossRef]
- Callejón, S.; Sendra, R.; Ferrer, S.; Pardo, I. Cloning and characterization of a new laccase from Lactobacillus plantarum J16 CECT 8944 catalyzing biogenic amines degradation. Applied Microbiology and Biotechnology 2016, 100, 3113–3124. [Google Scholar] [CrossRef] [PubMed]
- da Rocha, A.B.; de Aquino Saraiva, R.; de Siqueira, V.M.; Yogui, G.T.; de Souza Bezerra, R.; de Assis, C.R.D.; Sousa, M.S.B.; de Souza Buarque, D. Shrimp laccase degrades polycyclic aromatic hydrocarbons from an oil spill disaster in Brazil: A tool for marine environmental bioremediation. Marine Pollution Bulletin 2023, 194, 115445. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, B.; Xu, W.; Zhang, W.; Guang, C.; Mu, W. Efficient removal of sulfonamides and tetracyclines residues by the laccase-mediator system employing a novel laccase from Lysinibacillus fusiformis. Journal of Environmental Chemical Engineering 2022, 10, 108809. [Google Scholar] [CrossRef]
- Liu, N.; Shen, S.; Jia, H.; Yang, B.; Guo, X.; Si, H.; Cao, Z.; Dong, J. Heterologous expression of Stlac2, a laccase isozyme of Setosphearia turcica, and the ability of decolorization of malachite green. Int J Biol Macromol 2019, 138, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, D.; Wu, Z.; Li, D.; Cai, Y.; Lu, Y.; Zhao, X.; Xue, H. Multiple Tolerance and Dye Decolorization Ability of a Novel Laccase Identified from Staphylococcus Haemolyticus. J Microbiol Biotechnol 2020, 30, 615–621. [Google Scholar] [CrossRef]
- Blánquez, A.; Ball, A.S.; González-Pérez, J.A.; Jiménez-Morillo, N.T.; González-Vila, F.; Arias, M.E.; Hernández, M. Laccase SilA from Streptomyces ipomoeae CECT 3341, a key enzyme for the degradation of lignin from agricultural residues? PLoS One 2017, 12, e0187649. [Google Scholar] [CrossRef]
- Brander, S.; Mikkelsen, J.D.; Kepp, K.P. TtMCO: A highly thermostable laccase-like multicopper oxidase from the thermophilic Thermobaculum terrenum. Journal of Molecular Catalysis B: Enzymatic 2015, 112, 59–65. [Google Scholar] [CrossRef]
- Navas, L.E.; Martínez, F.D.; Taverna, M.E.; Fetherolf, M.M.; Eltis, L.D.; Nicolau, V.; Estenoz, D.; Campos, E.; Benintende, G.B.; Berretta, M.F. A thermostable laccase from Thermus sp. 2.9 and its potential for delignification of Eucalyptus biomass. AMB Express 2019, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Suman, S.K.; Khatri, M.; Dhawaria, M.; Kurmi, A.; Pandey, D.; Ghosh, S.; Jain, S.l. Potential of Trametes maxima IIPLC-32 derived laccase for the detoxification of phenolic inhibitors in lignocellulosic biomass prehydrolysate. International Biodeterioration & Biodegradation 2018, 133, 1–8. [Google Scholar] [CrossRef]
- Zheng, F.; An, Q.; Meng, G.; Wu, X.J.; Dai, Y.C.; Si, J.; Cui, B.K. A novel laccase from white rot fungus Trametes orientalis: Purification, characterization, and application. Int J Biol Macromol 2017, 102, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Ai, M.Q.; Wang, F.F.; Huang, F. Purification and Characterization of a Thermostable Laccase from Trametes trogii and Its Ability in Modification of Kraft Lignin. J Microbiol Biotechnol 2015, 25, 1361–1370. [Google Scholar] [CrossRef]
- Jaiswal, N.; Pandey, V.P.; Dwivedi, U.N. Immobilization of papaya laccase in chitosan led to improved multipronged stability and dye discoloration. International Journal of Biological Macromolecules 2016, 86, 288–295. [Google Scholar] [CrossRef]
Figure 1.
Schematic representations of -amylases industrial applications, structural relation with thermostablilty and mechanism of catalytic functions. A. An illustration model of the -amylases uses in the diverse commercial applications. B. Structural depiction of the crystal structure of -amylase from Anoxybacillus species (TASKA, PDB 5A2A [134]) showing a single polypeptide chain folding with the relative positions of the three structural domains: a) Domain A, the catalytic domain, b) Domain B, that constitutes a significant portion of the substrate binding cleft, responsible for notable variations in size, structure, and substrate specificity among different α-amylases, and c) Domain C, forming the C-terminal segment of the sequence. Domains A, B and C are shown in green, orange, and blue, respectively and the calcium ions in magenta. C. Illustration depicting the chemical structure of starch (comprising amylose and amylopectin) and the enzymatic conversion into sugar units [135]. The catalytic mechanism of α-amylases involves the cleavage of the internal α-glycosidic bonds in polysaccharides like starch, glycogen, and others, leading to the hydrolysis of these bonds and the production of α-anomeric mono- or oligosaccharides.
Figure 1.
Schematic representations of -amylases industrial applications, structural relation with thermostablilty and mechanism of catalytic functions. A. An illustration model of the -amylases uses in the diverse commercial applications. B. Structural depiction of the crystal structure of -amylase from Anoxybacillus species (TASKA, PDB 5A2A [134]) showing a single polypeptide chain folding with the relative positions of the three structural domains: a) Domain A, the catalytic domain, b) Domain B, that constitutes a significant portion of the substrate binding cleft, responsible for notable variations in size, structure, and substrate specificity among different α-amylases, and c) Domain C, forming the C-terminal segment of the sequence. Domains A, B and C are shown in green, orange, and blue, respectively and the calcium ions in magenta. C. Illustration depicting the chemical structure of starch (comprising amylose and amylopectin) and the enzymatic conversion into sugar units [135]. The catalytic mechanism of α-amylases involves the cleavage of the internal α-glycosidic bonds in polysaccharides like starch, glycogen, and others, leading to the hydrolysis of these bonds and the production of α-anomeric mono- or oligosaccharides.
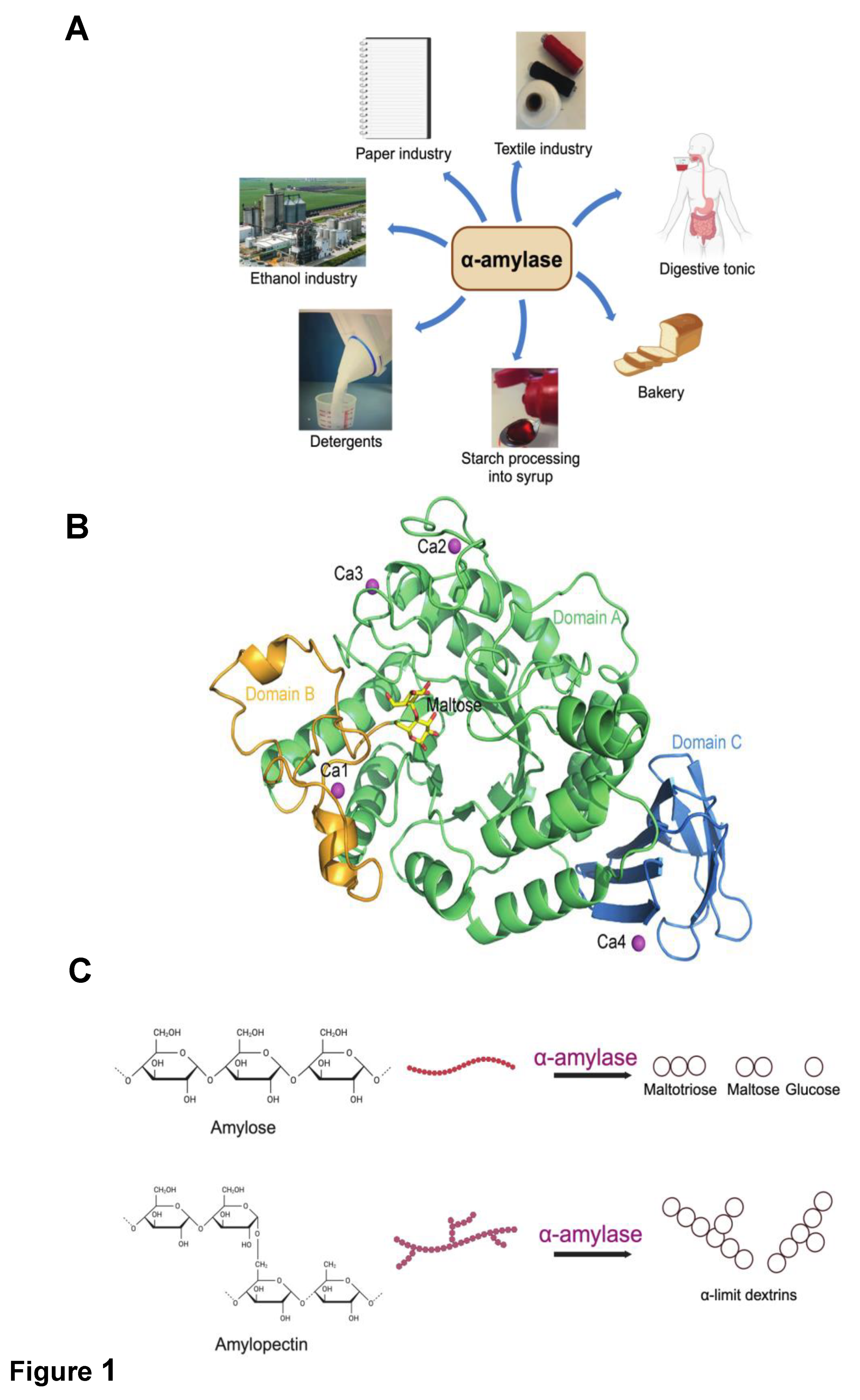
Figure 2.
Schematic representations of laccases industrial applications, structural relation with thermostablilty and mechanism of catalytic functions. A. A comprehensive model of laccases uses in the diverse industrial applications. B. Structural representation of the crystal structure of the laccase from Bacillus subtilis (PDB 1GSK, [136]) showing the polypeptide chain with relative postioning of the three homologous cupredoxin domains, domain 1, 2, and 3 arranged, sequentially. Domain 2 joins and positions domains 1 and 3. Domain 1, 2, and 3 are depicted in red, blue, and green. C. Illustration representing the catalytic mechanism of laccases: the substrate-induced reduction of the T1 copper, followed by the transfer of electrons to the TNC and subsequent reduction of O2 (adapted from [94,137]. The first step is the oxidation of the substrate by mononuclear copper T1, which serves as an electron acceptor, converting Cu2+ to Cu+ oxidation state. After removing an electron from the substrate, an unstable cationic radical is formed which is oxidized by a second enzymatic reaction or undergo non-enzymatic reactions, such as hydration or polymerization. The electrons removed from the substrate at the T1 site are transferred to the T2/T3 center for conversion of O2 to H2O. Complete reduction of molecular oxygen to water requires four molecules of reducing substrate. Thus, the stoichiometry of the enzymatic reaction of the catalytic mechanism of laccases is represented by the equation: 4RH + O2 → 4R + 2H2O, where RH signifies the substrate.
Figure 2.
Schematic representations of laccases industrial applications, structural relation with thermostablilty and mechanism of catalytic functions. A. A comprehensive model of laccases uses in the diverse industrial applications. B. Structural representation of the crystal structure of the laccase from Bacillus subtilis (PDB 1GSK, [136]) showing the polypeptide chain with relative postioning of the three homologous cupredoxin domains, domain 1, 2, and 3 arranged, sequentially. Domain 2 joins and positions domains 1 and 3. Domain 1, 2, and 3 are depicted in red, blue, and green. C. Illustration representing the catalytic mechanism of laccases: the substrate-induced reduction of the T1 copper, followed by the transfer of electrons to the TNC and subsequent reduction of O2 (adapted from [94,137]. The first step is the oxidation of the substrate by mononuclear copper T1, which serves as an electron acceptor, converting Cu2+ to Cu+ oxidation state. After removing an electron from the substrate, an unstable cationic radical is formed which is oxidized by a second enzymatic reaction or undergo non-enzymatic reactions, such as hydration or polymerization. The electrons removed from the substrate at the T1 site are transferred to the T2/T3 center for conversion of O2 to H2O. Complete reduction of molecular oxygen to water requires four molecules of reducing substrate. Thus, the stoichiometry of the enzymatic reaction of the catalytic mechanism of laccases is represented by the equation: 4RH + O2 → 4R + 2H2O, where RH signifies the substrate.
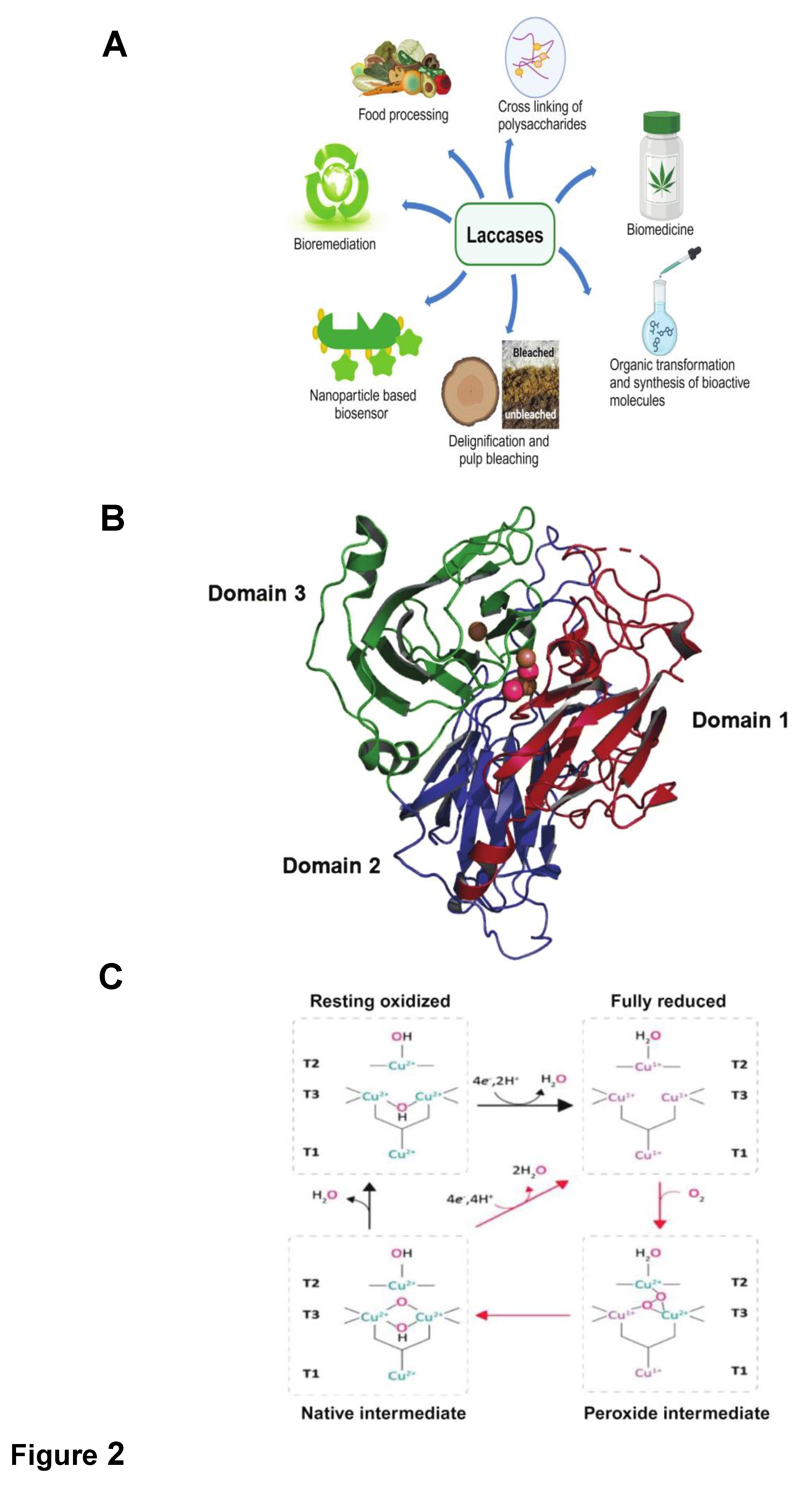

Table 1.
Commercially available α-amylases and their industrial applications.
| Microbial source of α amylases |
Commercial Name of α- amylase |
Manufacturer | Industrial applications |
|---|---|---|---|
| Aspergillus oryzae | Fructamyl® FHT | Erbslöh Geisenheim AG |
Beverage industry |
| Bacillus licheniformis | Liquozyme® SC DC | Novozymes | Liquefaction for ethanol production |
| Bacillus amyloliquefaciens | BAN® | Novozymes | Oat starch liquefaction |
| Bacillus licheniformis | Termamyl® | Novozymes | Adjunct liquefaction |
| Aspergillus oryzae | Fungamyl | Novozymes | Baking |
| Bacillus subtilis | Validase BAA | IMCD Germany | Food and Feed |
| Bacillus subtilis | ZylozymeTM AA | Kemin Industries | Biofuel |
| Bacillus licheniformis | Bioconvert ALKA | Noor Enzymes | Biofuel |
| Genetically modified microorganism | Stainzyme® Plus Evity® 48 T | Novozymes | Detergent |
| Genetically modified microorganism | Aquazym® | Novozymes | Textile |
Listed are commercial α-amylases, each derived from specific microbial sources, offering various industrial purposes. Sources: https://erbsloeh.com/; https://www.novozymes.com/en/products/ethanol/liquefaction/liquozyme; https://www.novozymes.com/en/products/baking/freshness/ban?utm_source; https://www.novozymes.com/en/products/plant-based-foods/plant-based-dairy/termamyl; https://www.novozymes.com/en/products/baking/flour-correction/fungamyl?utm_source; https://www.imcd.de/en/trade-names/food-and-nutrition-BG3/validase-a5u690000000fr0AAA/; https://www.kemin.com/na/en-us/markets/biofuels/products/zylozyme-aa; http://www.enzyme-india.com/amylase-enzymes-biofuel.html; https://www.novozymes.com/en/products/laundry/starch-stains/stainzyme-plus-evity-48-t; https://www.novozymes.com/en/products/pulp-paper/aquazym-480-l.
Table 2.
Microbial Sources of thermostable α-amylases.
| Source of α-amylases | Optimum Temperature | Industrial applications | References |
|---|---|---|---|
| Actinomadura keratinilytica sp. Cpt29 | 70 °C | Laundry detergent additive | [138] |
| Aeribacillus pallidus BTPS- 2 | 70 °C | Starch liquefcation | [139] |
| Anoxybacillus vranjensis ST4 | 60–80 °C | Starch hydrolysis | [140] |
| Bacillus amyloliquefaciens BH072 | 60 °C | Food processing | [141] |
| Bacillus cereus SP- CH11 | 65 °C | Food processing | [142] |
| Bacillus licheniformis AT70 | 60 °C | Starch degradation | [143] |
| Bacillus licheniformis NH1 strain | 70 °C | Laundry detergent additive | [144] |
| Bacillus licheniformis So-B3 | 70 °C | Hydrolyzing raw starch | [145] |
| Bacillus sp. isolate A3-15 | 100 °C | Textile industry | [146] |
| Bacillus tequilensis TB5 | 60 °C | Textile de-sizer | [147] |
| Chromohalobacter sp. TVSP 101 | 65 °C | Starch hydrolysis | [148] |
| Geobacillus thermoleovorans | 80 °C | Improvement of washing efficiency of detergents | [149] |
| Germinated wheat seeds (Triticum aestivum) |
68 °C | Starch processing | [36] |
| Haloterrigena turkmenica | 55 °C | Agricultural residues treatment | [150] |
| Paecilomyces variotii | 60 °C | Starch degradation | [151] |
| Rhizomucor miehei | 75 °C | Food processing | [152] |
| Rhizopus oligosporus | 60 °C | Laundry detergent additive | [153] |
| Soybean (Glycine max) seeds | 75 °C | Starch liquefaction | [12] |
| Tepidimonas fonticaldi strain HB23 | 80 °C | Laundry detergent additive | [154] |
| Thermomyces dupontii | 60 °C | Maltose syrup production | [155] |
Listed are sources of α-amylases from different microbes with their optimum temperature which can be useful in the industrial purposes.
Table 3.
Commercially available Laccases and their industrial applications.
| Source of Laccases | Commercial Name of Laccase | Manufacturer | Industrial applications |
|---|---|---|---|
| Myceliophthora thermophila laccase expressed in Aspergillus oryzae | Denilite™ I Denilite™ II |
Novozymes [156] Novozymes |
|
| Zylite | Zytex Biotech Private Limited [156] | Textile | |
| Ecostone LC10 | AB Enzymes GmbH | ||
| IndiStar | Genencor International Inc. | ||
| Novoprime Base 268 | Novozymes [157] | ||
| Primagreen Ecofade LT100 | Genencor International Inc. [158] | ||
| Novozym® 51,003 | Novozymes [159] | ||
| White-rot fungi (Phanerochaete chrysosporium, Trametes versicolor) | Lignozym® Process Laccase Y120 Novozym® 51,003 |
IBB Netzwerk GmbH [160] Amano Enzyme [161] Novozymes [159] |
Paper Food processing |
| Filamentous fungi and yeasts | Suberase® | Novozymes [162] | Brewing |
| Genetically engineered bacterial laccase | MetZyme® LIGNO™ | MetZen [163] | Bio-refinery |
Listed are commercial α-Laccases with their specific microbial sources, serving different industrial purposes. Sources: https://www.novozymes.com/en; https://www.zytex.com/; https://www.abenzymes.com/en/your-industry/textiles/; https://www.textileweb.com/doc/genencor-launches-the-indistar-coloradjust-sy-0001.
Table 4.
Microbial Sources of thermostable Laccases.
| Source of Laccases | Optimum Temperature | Industrial applications | References |
|---|---|---|---|
| Agaricus bisporus CU13 | 55 °C | Decolorization of synthetic dyes | [164] |
| Alcaligenes faecalis XF1 | 80 °C | Decolorization of synthetic dyes | [165] |
| Azospirillum lipoferum | 70 °C | Ecological role in the process of root colonization | [166] |
| Bacillus altitudinis SL7 | 55 °C | Bioremediation of lignin contaminated wastewater from pulp and paper industries | [167] |
| Bacillus sp. MSK-01 | 75 °C | Proposed as an anti-proliferative agent to cancer cells | [168] |
| Bacillus sp. PC- 3 | 60 °C | Functionalization of chitosan film for antimicrobial activity | [169,170] |
| Bacillus subtilis | 60 °C | Biodegradation of the fungicide | [171] |
| Bacillus subtilis strain R5 | 55 °C | Degradation of synthetic dyes | [172] |
|
Caldalkalibacillus thermarum TA2.A1 |
70 °C | Lignin degradation | [123] |
| Coprinopsis cinerea | 70 °C | Wastewater treatment | [173] |
| Enterobactersp. AI1 | 60 °C | Degradation and detoxification of synthetic dyes | [153] |
| Galerina sp. HC1 | 60 °C | Demethylation of lignin | [174] |
| Ganoderma lucidum KMK2 | 60 °C | Decolorization of reactive dyes | [175] |
| Ganoderma multipileum | 70 °C | Biodegradation of chromium | [176] |
| Geobacillus stearothermophilus MB600 | 90 ℃ | Biodegradation of pollutants | [70] |
| Geobacillus yumthangensis | 60 °C | Degradation of organic pollutants | [169] |
| Klebsiella pneumoniae | 70 ℃ | Decolorization of synthetic dyes | [177] |
| Lactobacillus plantarumJ16 CECT 8944 | 60 °C | Eliminating toxic compounds present in fermented food and beverages | [178] |
| Litopenaeus vannamei | >90 °C | Marine bioremediation | [179] |
| Lysinibacillus fusiformis | 80 °C | Removal of sulfonamides and tetracyclines residues | [180] |
| Setosphearia turcica | 60 °C | Decolorization of malachite green | [181] |
| Staphylococcus haemolyticus | 60 °C | Textile finishing | [182] |
| Streptomyces ipomoeae CECT 3341 | 60 ± 6 °C | Decolorization and detoxification of textile dyes | [183] |
| Thermobaculum terrenum | 80 °C | Protein engineering studies | [184] |
| Thermus sp. 2.9 | 70 °C | Delignification of Eucalyptus biomass | [185] |
| Trametes maxima IIPLC- 32 | 50–70 °C | Detoxification of phenolic inhibitors in lignocellulosic biomass | [186] |
| Trametes orientalis | 80 °C | Decolorization and bioremediation of synthetic dyes | [187] |
| Trametes trogii | 70 °C | Modification of kraft lignin | [188] |
| Leucaena leucocephala | 80 °C | Decolorization of synthetic dyes | [98,189] |
| Carica papaya | 70 °C | Dye decolorization | [98,189] |
Listed are sources of laccases from diverse microbes with specific optimal temperatures which are proven to be useful in industrial applications.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



