
Submitted:
22 April 2024
Posted:
23 April 2024
You are already at the latest version
Abstract
The extraocular muscles (EOMs) possess unique characteristics that set them apart from other skeletal muscles. These muscles, responsible for eye movements, exhibit remarkable resistance to various muscular dystrophies and aging, presenting a significant contrast to the vulnerability of skeletal muscles to these conditions. In this review, we delve into the cellular and molecular underpinnings of the distinct properties of EOMs. We explore their structural complexity, highlighting differences in fiber types, innervation patterns, and developmental origins. Notably, EOM fibers express a diverse array of myosin heavy chain isoforms, retaining embryonic forms into adulthood. Moreover, their motor innervation is characterized by a high ratio of nerve fibers to muscle fibers and the presence of unique neuromuscular junctions. These features contribute to the specialized functions of EOMs, including rapid and precise eye movements. Understanding the mechanisms behind the resilience of EOMs to disease and aging may offer insights into potential therapeutic strategies for treating muscular dystrophies and myopathies affecting other skeletal muscles.
Keywords:
extraocular muscles
; muscular dystrophies
; muscular aging
1. Introduction
The extraocular muscles (EOM) are unique in several aspects: they represent the fastest and most fatigue-resistant muscles within the human body [1,2,3]. Extraocular muscles (EOMs) predominantly exhibit impairment in conditions such as Myasthenia Gravis and mitochondrial myopathies, yet, remarkably, they are spared from various muscular dystrophies, including Duchenne, Becker, limb-girdle, and congenital muscular dystrophies. Furthermore, EOMs demonstrate particular resistance to amyotrophic lateral sclerosis (ALS) [4,5].
The complexity of the actions performed by the extraocular muscles (EOM) is reflected in their anatomical and physiological characteristics. Morphologically and in terms of their molecular composition, they significantly differ from the muscle fibers (MF) of other skeletal muscles. The gene expression profile of the EOM is distinct from that of limb muscles, with differences encompassing over 330 genes involved in metabolic pathways, structural components, development markers, and regenerative processes. Unlike skeletal muscles, the EOM predominantly utilizes an aerobic pathway for carbohydrate metabolism and relies directly on glucose influx from the blood. This metabolic strategy enables them to be among the fastest muscles in the body while also being exceptionally resistant to fatigue [6].
Our review focuses on understanding the cellular and molecular basis of the unique properties of extraocular EOM, which enable them to resist muscular dystrophies and aging. Comprehending how EOM counteract diseases that lead to degeneration and functional loss in skeletal muscles could lay the groundwork for developing gene and cell therapies and medical devices for treating dystrophies and myopathies of various etiologies.
1.1. Structural Characteristics of the EOM
EOM enable the eyeball to perform slow pursuit movements and rapid, instantaneous shifts from one fixation point to another. The EOM comprise six muscles responsible for controlling eyeball movement and one muscle, the levator palpebrae, which governs eyelid motion (Figure 1). Unlike skeletal muscles, the EOM are innervated by cranial nerves, rather than motor neurons located in the spinal cord [7].
The development of EOM originates from the prechordal and cranial paraxial mesoderm [8]. The mesodermal mesenchyme that forms the EOM is interspersed with cells of the cranial neural crest, which contribute to the formation of the sclera and the connective tissue surrounding the muscles [9,10,11,12,13,14,15,16]. Experiments involving the ablation of the neural crest suggest an interactive process in the formation of the EOM between the mesoderm and the neural crest. However, due to variations in experimental models, the involvement of the neural crest in the development and organization of the EOM remains somewhat controversial. Nonetheless, given the contribution of cranial neural crest cells to the development of craniofacial bones, connective tissue, nerves, and the eye, it is plausible that they play a significant role in the organization of the EOM [9,17,18,19,20,21,22].
During embryogenesis, EOMs undergo the same stages of myogenesis as skeletal muscles. However, unlike skeletal muscles, EOMs retain "immature" protein isoforms, including embryonic myosin [23]. It has also been demonstrated that only minor changes occur with age in EOMs, and they are less susceptible to aging compared to skeletal muscles [24,25]. This persistence of "immature" myosins may facilitate the continuous self-renewal and regeneration of EOMs, potentially accounting for their particular resistance to muscular dystrophies and aging.
1.2. Muscle Fibers of Extraocular Muscles
Skeletal muscle fibers (MF) are conventionally classified as slow (type I) and fast (type II with subtypes a, b, and x) to reflect their contractile and metabolic properties [6]. However, this classification is not applicable to the EOM [26,27]. In contrast to skeletal muscles, they possess a distinct classification scheme based on their contractile abilities, anatomical location, and innervation [28].
Each EOM is subdivided into at least two primary layers: the "orbital" layer, featuring thinner MF adjacent to the bony walls of the orbit (forming the central fascicle in the oblique muscles), and the internal "global" layer, containing thicker MFs directly abutting the eyeball and optic nerve (Figure 2) [29,30]. In these layers, the muscle fibers can be either singly or multiply innervated MF [29,30]. The levator palpebrae superioris muscle, uniquely among all EOMs, is not multilayered and comprises solely singly innervated MF [31].
During development, skeletal muscles express both isoforms of myosin heavy chains (MyHC) involved in development: MyHCemb and MyHCneonatal. However, post-development, only the expression of a single adult isoform of MyHC I, IIa, IIb, and IIx is retained. It is the type of expressed MyHC isoform that determines the contraction speed, strength, and ATP consumption by the muscles [32].
The expression of embryonic and neonatal MyHC isoforms is retained in mature EOM, which also express nearly all known MyHC isoforms (Table 1). Additionally, the expression of more than one MyHC isoform in individual muscle fibers has been documented [33,34,35,36,37]. Beyond embryonic forms, two ancient MyHC isoforms, MYH14/7b and MYH15, have been identified in adult EOM [38,39,40].
Nevertheless, the presence of all MyHC isoforms has not always been confirmed by researchers studying EOM. Occasionally, they were unable to identify certain isoforms, referring to such MHC in their studies as unidentified isoforms [41,42,43,44,45,46].
In 1971, Mayr proposed a classification for EOM (Extraocular Muscles) and divided them into six groups: two groups in the "orbital" layer and four in the "global" layer, which remains the standard classification to this day. (Table 1: Types of EOM muscle fibers) [28,47].
- Singly innervated, fast-twitch, and fatigue-resistant in the "orbital" layer.
- Multiply innervated with both fast-twitch and slow-twitch fibers, exhibiting variable fatigue resistance in the "orbital" layer.
- "Red," singly innervated, fast-twitch, and fatigue-resistant in the "global" layer.
- "White," singly innervated, fast-twitch, and exhibiting low fatigue susceptibility in the "global" layer.
- "Intermediate," singly innervated, fast-twitch with moderate fatigue resistance in the "global" layer.
- Multiply innervated, slow-twitch, and fatigue-resistant in the "global" layer.
Despite the established standard, the classification of EOM (extraocular muscles) in the literature is quite heterogeneous. Some studies describe only five types of EOMs in mammals: four in the "global" layer and three in the "orbital" layer, with two identical EOM types present in both layers [48]. In a subsequent study, the same researchers identified six distinct types, with four identical types in each layer [49]. Other studies also classified six different types, but with three distinct EOM types in each layer [30].
Some studies indicate that EOMs are structurally more complex compared to those of other mammals. In 2000, a group of researchers, in their efforts to classify the neuromuscular MF of EOM, identified a third layer known as the marginal zone. Within this zone, they described three types of MF: singly innervated muscle fibers and two types of multiply innervated muscle fibers [50]. However, in later papers by other researchers, there is no mention of a third layer in the human EOM.
Despite extensive research on EOM, scientists have yet to reach a consensus on the distribution and quantity of muscle fiber (MF) types in the EOM. Disagreements among researchers may stem from varying methodologies employed in their studies, the presence of MFs expressing multiple types of MyHC, and the unique and complex structural organization of the EOM. The lack of consistency in classifying EOM MF further underscores their uniqueness. In addition to the multitude of MyHC types found in the EOM, which are not present in other skeletal muscles, these muscles also exhibit distinct motor innervation patterns.
1.3. Motor Innervation
The ratio of nerve fibers to muscle fibers in the EOM is more than tenfold higher (ranging from 1:3 to 1:5 or 1:12) compared to skeletal muscles (ranging from 1:50 to 1:125) [51,52,53]. The architecture of motor endplates depends on the structure, physiology, and histochemistry of the MF [54,55,56,57]. Muscles are categorized into two primary groups based on their contraction capabilities: phasic (twitch) and tonic muscle fiber [51,52,53]. Tonic muscle fibers maintain static tension, or tone. In contrast, phasic muscle fibers can perform rapid contractions but are incapable of sustaining the achieved level of shortening over extended periods [58,59,60].
Muscle fibers characterized by tonic contraction types are prevalent in amphibians and reptiles, and have also been identified in the mammalian EOM [61,62,63,64,65,66]. Across all studied species, these tonic muscle fibers exhibit multiple innervations [66,67,68,69,70]. In contrast, phasic (twitch) muscle fibers possess a single innervation and are broadly classified into two subclasses within skeletal muscles based on their contraction velocity: slow (type I) and fast (type II) [71].
The evolutionarily tonic type of contraction represents an ancient form, as evidenced by the process of muscle development and during embryogenesis, where MF exhibit multiple innervation. During development, motor neurons extend their axons to the prospective sites of skeletal muscle formation coinciding with the emergence of muscle tubes. Multiple axons grow towards the initial contact site, where polyneuronal innervation (or multiple innervation) occurs at this juncture. However, in skeletal muscles, only the most active axon forms the definitive neuromuscular synapse (Figure 3) [58].
Axons innervating phasic (twitch) muscle fibers terminate in a single neuromuscular junction (NMJ), referred to in the literature as an en plaque ending. Conversely, tonic muscle fibers (MyHCsto/I) in the EOM receive multiple innervations, with grape-like NMJ clusters, which are referred to in the literature as en grape endings (Figure 4).
Recent investigations in EOM containing MyHCeom have revealed a novel type of NMJ, characterized by multitendinous endings distributed along the entire length of the MF. Contrary to the en grape NMJ, which are traditionally aligned in a row on one side of the MF, this multitendinous NMJ is situated on both sides of the MF. Furthermore, the study demonstrated that en grappe endings were exclusively identified in myofibrils harboring MyHCsto/I [72].
The distribution of mitochondria within EOM fibers varies based on the type of innervation and the specific EOM. Research from the 1970s to the 1990s documented diverse mitochondrial compositions in EOM fibers, influenced by their contraction patterns. Khairy et al. noted that fibers in the "orbital" layer exhibit a relatively higher mitochondrial content compared to those in the "global" layer [73]. Demer et al. suggested that the differences in mitochondrial content between "orbital" and "global" layers could be attributed to their functional specialization [74]. Other researchers have indicated that EOMs, in general, possess a higher mitochondrial density than skeletal muscles [6].
Singly innervated MF exhibit phasic (twitch) contraction types and contain a high density of mitochondria throughout their length (Figure 5) [75]. In the "global" layer, red, singly innervated MF possess a greater mitochondrial content, whereas pale singly innervated ones have fewer [76]. Conversely, the multi-innervated MF of the "global" layer display a tonic contraction type and house only a scant number of mitochondria. Meanwhile, "orbital" MF can demonstrate phasic (twitch) contractions and NMJ resembling en plaque formations in the muscle's midsection coinciding with en grappe terminations and tonic contraction types in the MF peripheral regions [77]. Consequently, the central portion of the "orbital" multi-innervated MF is also rich in mitochondria, similar to the singly innervated fibers [76].
Based on the type of innervation, three categories of MF can be distinguished [30]:
- 1.
- Singly innervated (twitch) muscle fibers, corresponding to fast skeletal MF. This type constitutes the dominant population in both the "global" (90%) and "orbital" (80%) layers; electrical stimulation of the innervating axon triggers a twitch response based on the "all-or-nothing" principle.
- 2.
- 3.
- Multiply innervated MF in the "orbital" layer (20%) with NMJ en grappe and en plaque, exhibiting corresponding dynamics of contraction in the central and distal parts upon electrical stimulation [80].
In addition to the presence of multiply innervated MF, another distinctive feature of EOM is the preservation of expression in adult muscle of the gamma-subunit of the fetal acetylcholine receptor (γ-AChR), which in skeletal muscles occurs only during the period of intrauterine development or in denervated MF, and within the first two weeks after birth, it is replaced by the ε-subunit (γ/ε switching) [72,81,82].
In EOM, both γ-AChR and ε-AChR subunits are preserved [72,83]. The fetal γ-subunit is essential for the proper maturation of the neuromuscular synapse [84,85,86]. Mutations in γ-AChR can lead to both lethal and non-lethal alterations, such as multiple pterygium syndrome (also known as Escobar Syndrome, a form of congenital myasthenic syndrome) [87,88,89].
The particular composition of AChR subunits in EOM is likely related to the type of MF and their physiological properties. AChRs containing the γ-AChR subunit differ from those containing the ε-AChR subunit in terms of their ionic conductivity and ion channel opening rates [90] The ion channel opening time during limb muscle development decreases from 11 ms to 6 ms when switching from the fetal γ-subunit to the adult ε-subunit of AChR [90]. Interestingly, fast MF expressing MyHCIIa exclusively contain ε-AChR subunits, whereas tonic and slow MyHCsto/I with NMJ en grappe express γ-AChR subunits [72]. The difference in AChR subunit expression corresponds to the difference in contraction speed between these two types of MF.
During evolution, EOM have retained the tonic contraction type, which seems necessary for performing specialized eye movements, such as tracking or sustained fixation on an object. However, maintaining tonic contraction in MF requires multiple innervation, not found in mature skeletal muscles. In addition to having tonic, slow contraction in specific MF, EOM are also the fastest muscles in the body. These functional characteristics of EOM are manifested in the presence of MyHC types not found in skeletal muscles, the retention of the fetal γ-AChR subunit, and differences in the cytoskeletal structure and surrounding connective tissue of the MF.
1.4. Cytoskeleton and Basal Membrane of EOM
Another factor contributing to the disease resistance in EOM might be attributed to the architectural uniqueness of their cytoskeleton. The muscle cytoskeleton comprises a complex network of proteins, organized to interlink and anchor cellular structural components such as myofibrils, nuclei, and sarcolemma, regulating cellular morphology and facilitating the transduction of myofibrillar movements into the surrounding tissues.
Desmin is the first muscle-specific protein discovered during muscle development, and it is such a fundamental component of the muscle fiber that it is used as a marker for muscle tissue. Desmin is considered essential for maintaining the structural and functional integrity of the postsynaptic apparatus [91,92].
Although muscle fibers can develop in the absence of desmin, defects in the genes encoding this protein result in severe myopathy [93]. Nevertheless, it has been discovered that a portion of healthy, undamaged EOM fibers in humans lacks desmin, constituting a true paradigm shift in the understanding that desmin is essential for all muscle fibers. Furthermore, despite the absence of desmin, EOM remain unaffected in desmin-related myopathy [94,9Pl4].
The study conducted by Jing-Xia Liu and Fatima Pedrosa Domellöf confirmed the absence of desmin in many motor endplate of human EOM and further revealed a lack of desmin in the immediate vicinity of the NMJ, in contrast to the abundant presence of desmin at NMJ observed in limb muscles. A concise summary of their findings is presented in Figure 6 [94].
These findings suggest that desmin does not play the same role in EOM as it does in limb muscles. Despite the absence of desmin, these neuromuscular synapses exhibit normal morphology and the normal distribution of many crucial synaptic molecules, including AChR subunits; neurofilaments and synaptophysin; laminin isoforms α2, α4, α5, and β2; and various isoforms of Wnt, neurotrophins, and their receptors in healthy human EOM [96–99]. Consequently, the lack of desmin in neuromuscular synapses is not considered a defect in human EOM.
In the context of these studies, it is particularly noteworthy that mice with desmin deficiency exhibited a shift in MF types towards slow, fatigue-resistant fibers [100,101]. Additionally, one of the functional characteristics of EOM is their high resistance to fatigue.
Another factor contributing to resilience and a significant distinction from skeletal muscle is the composition of the basal membrane, which protects EOM from congenital muscular dystrophy with Lna2 deficiency. The basal membrane is situated on the immediate surface of skeletal muscle fibers, with laminins serving as one of the principal components of the basal membrane. Laminins are associated with receptors that span the thickness of the cell membrane, thereby attaching to the cytoskeleton within the muscle fiber. It has been demonstrated that the basal membrane surrounding the EOM fibers in humans contains not only the typical α-chain laminin isoform present in adult skeletal muscle fibers but also other isoforms that are present in skeletal muscles only during development. Thus, EOM remain unaffected by the congenital Lna2 defect because they typically possess additional isoforms of the same laminin chain [102]. Additionally, EOMs express the Lutheran protein, which is an α5-chain-specific receptor, not found in limb muscles [102]. The presence of laminin isoforms, absent in adult skeletal muscles, is a common feature of all mature EOM, and this may indeed serve as an additional protective mechanism against diseases. In one study, it was shown that the expression of laminin (Lama2) in EOM fibroblasts was increased 17-fold compared to skeletal muscle fibroblasts, consistent with the observation that EOM has much denser connective tissue compared to skeletal muscle overall, including leg muscles [102].
1.5. Connective Tissue
Typically, each MF in the EOM is surrounded by connective tissue and is not in direct contact with adjacent fibers, as is commonly observed in other skeletal muscles. This confers a round contour to the EOM fibers, whereas skeletal muscles exhibit a polygonal shape due to the close proximity of neighboring MF [103].
The connective tissue of the EOM contains fibroblasts that exhibit properties distinct from those in other skeletal muscles, potentially contributing to the unique response of the EOM and their involvement in inflammatory diseases such as Graves' ophthalmopathy (thyroid eye disease) [104–106]. Another feature is the distinct genomic profile of EOM fibroblasts compared to those of skeletal muscle. Orbital fibroblasts develop from neural ectoderm, unlike fibroblasts from other regions, which are of mesenchymal origin [107,108]. Consequently, significant differences in the transcription factors of EOM can be anticipated [108].
Genes associated with a differential immune response have been identified. Specifically, in EOM fibroblasts, there was an increased expression of CD59a, a complement system regulator that impacts the severity of myasthenia [109–111] and IL-6, which is involved in Graves' ophthalmopathy [112,113]. The response of EOM to immune mediators differs from that of other skeletal muscles with fibroblasts playing a significant role in this response [114,115]. Furthermore, the genomic profile of EOM demonstrated elevated expression of calreticulin, which may act as an autoantigen in Graves' ophthalmopathy [116].
1.6. Muscle Fiber Metabolism and Antioxidant Capacity
Enhanced expression of factors such as IL-6 in EOM fibroblasts may increase glucose uptake in MF [109]. Concurrently, the expression profile of MBs themselves suggests that they utilize aerobic carbohydrate metabolism and are dependent on blood glucose levels as an energy source. Support for this theory is evidenced by the reduced expression of phosphoglucomutase (a key enzyme in glycogen synthesis), phosphorylase kinase (a principal regulator of glycogen breakdown or glycogenolysis), and a decreased glycogen content in EOM. The deficiency of these enzymes in skeletal muscles leads to type VIII glycogen storage disease and McArdle's syndrome (type V glycogen storage disease), yet these conditions do not affect EOM [6,7].
In EOM, only a minimal amount of glycogen is synthesized, and these muscles do not rely on it as their primary energy source. Instead, they predominantly oxidize glucose transported through the bloodstream to meet their energy demands. Furthermore, the expression profile of EOM suggests efficient glucose utilization via the Krebs cycle and conversion into energy through oxidative phosphorylation. The reliance of EOM on oxidative phosphorylation accounts for their sensitivity to mitochondrial myopathies [6].
The energetic metabolism based on blood glucose, together with a high mitochondrial content, suggests a mechanistic basis for the common metabolic adaptations utilized by EOM to sustain fatigue-resistant, rapid (∼400 Hz) contractions over relatively prolonged periods [117]. It is anticipated that the high mitochondrial content and oxidative phosphorylation would lead to an increase in the levels of reactive oxygen species (ROS) in EOM, potentially causing significant cellular damage [6]. However, it has been demonstrated that EOM expresses high levels of mRNA for chemoprotective enzymes, glutathione-S-transferase (GST) and UDP-glucuronosyltransferase (UDP-GT), presumably as an adaptation to the elevated ROS, which would be generated during oxidative phosphorylation in EOM. This indicates that EOM has enhanced antioxidant enzyme activity compared to limb muscles [118].
1.7. Protection, regeneration and Aging of EOM
Given the resistance of EOM to multiple myopathies, it is plausible that the progenitor cell niche in EOM markedly differs from that in other muscle groups. The proportion of satellite cells in undamaged adult EOM is quintuple that in limb muscles [4,5]. Additionally, activated satellite cells are observed in intact EOM [119]. A high quantity of progenitor cells is maintained even in the elderly, rendering the EOM more resistant to age-related degeneration or senescence [4,120]. Progenitor cells in EOM exhibit an increased capacity for proliferation, differentiation, and self-renewal [4,121]. Furthermore, these cells display increased expression of trophic factors, including brain-derived neurotrophic factors and nerve growth factors, which may account for their enhanced myogenic activity [122]. These characteristics lead to the hypothesis that EOM are capable of continuous remodeling throughout life [119].
Continuously activated satellite cells may be associated with a unique and/or more abundant progenitor cell subpopulation in adult EOM, which enables EOM to undergo continuous remodeling without stem cell depletion, unlike what is observed in limb skeletal muscles [123]. Similar to limb muscles, EOM contain Pax7-positive satellite cells, and morphometric analysis of histological sections revealed a significantly higher number of Pax7-positive cells compared to limb skeletal muscles [124,125]. Another progenitor cell population is the EOMCD34 cells, characterized as CD34+/Sca1-/CD31-/CD45-. These EOMCD34 cells are myogenic and are maintained during aging in EOM [124,125]. In murine models of DMD (Duchenne Muscular Dystrophy), the EOMCD34 cell population rapidly declines in limb skeletal muscle but is predominantly preserved in EOM. Interestingly, EOMCD34 cells, derived from EOM, exhibit increased proliferative capacity compared to their counterparts from limb skeletal muscles [125].
In the extraocular EOM, there is a complete absence of Pax3-positive myogenic precursor cells, which are typically co-expressed with Pax7 in skeletal muscles and are thought to be responsible for muscle regeneration observed in the absence of Pax7-positive cells [126]. Contrary to the lack of Pax3, it has been demonstrated that there exists a myogenic progenitor cell population expressing the transcription factor Pitx2. Pitx2 is a homeobox transcription factor that plays a pivotal role in myogenesis in the cranial region [127]. Pitx2-positive mononuclear cells are located both in the traditional location of satellite cells and in the interstitial connective tissue of the EOM. Pitx2 progenitor cells represent another significant population involved in the remodeling and regeneration of the EOM. Cells expressing Pitx2 are also predominantly preserved in EOM in murine models of muscular dystrophy and in aging skeletal muscles [128]. Western blotting and immunohistochemistry data showed that 80% of EOMCD34 cells express Pitx2. Knockdown of Pitx2 expression in EOMCD34 cells in vitro reduced the rate of proliferation and disrupted the ability of the cells to fuse into multinucleated muscle tubes [125]. Elevated levels of Pitx2 were maintained in dystrophic and aging EOM cells and EOMCD34 cells compared to limb muscles. Postnatal knockout of Pitx2, specific to skeletal muscles, causes a loss of characteristic expression patterns of myosin heavy chain (MyHC) isoforms in the EOM of transgenic mice, including the loss of EOM-specific (MYH13) and alpha-cardiac (MYH6) MyHC [129,130]. Furthermore, these mice with conditional Pitx2 knockout lose the multiply innervated muscle fibers typically found in the EOM making them more phenotypically similar to limb skeletal muscles [130]. These differential requirements for Pitx2 during development and in adulthood may contribute to the constitutive differences between the EOM and limb skeletal muscles, their preservation in muscular dystrophies, and their resilience to injury and denervation.
The myogenic environment of the EOM, enriched with precursor cells, may be sustained by the expression of several neurotrophic factors, which typically diminish in skeletal muscles [131], including insulin-like growth factor-1 and -2 [132,133], brain-derived neurotrophic factor [134,135], glial cell line-derived neurotrophic factor [136,137] and neurotrophin-3. These neurotrophic factors are crucial for the maintenance and development of ocular motor neurons [138,139].
Consequently, a unique environment enriched with neurotrophic factors and progenitor cells renders the EOM resistant to aging and muscular dystrophies, leading us to the conclusion that the EOM's response to damage or the introduction of myotoxic agents may significantly differ from that of skeletal muscles.
1.8. Response to Acute Damage by Botulinum Toxin in EOM
Injections of botulinum toxin into most other skeletal muscles lead to significant MF atrophy [140,141]. However, animal studies have shown that MF atrophy does not occur in the EOM following botulinum toxin injections [142]. In contrast to the atrophy observed in skeletal muscles, there is evidence describing the hypertrophy of singly innervated MF in EOM as a result of botulinum toxin administration [143]. One potential explanation is that the injection of botulinum toxin into the EOM induces a significant increase in activation, proliferation, and incorporation of progenitor cells into the existing MF [144,145].
2. EOM in Diseases
2.1. The Duchenne Muscular Dystrophy
The cause behind the morphological and functional resilience of EOM to Duchenne muscular dystrophy and related muscular dystrophies has long been a subject of research inquiry [146,147]. Numerous studies support the hypothesis that the remarkable regenerative capability of EOM enables them to remain both morphologically and functionally exempt from many forms of muscular dystrophy As discussed in the preceding section, EOM not only contain a large population of Pax7-positive myogenic precursor cells but also express an abundant population of Pitx2-positive myogenic precursor cells, which is significantly reduced in other skeletal muscles [148].
Hypotheses suggesting why EOM avoid damage in Duchenne muscular dystrophy converge on the notion that EOM-specific properties may aid in preventing or mitigating the deleterious effects of dystrophin deficiency. As an additional compensatory mechanism beyond regeneration, the hyperexpression of the dystrophin-related protein, utrophin (which can functionally replace dystrophin), has been proposed. Utrophin upregulation has been reported in some, but not all, studies conducted on normal and dystrophin-deficient EOM. No increase in utrophin was observed in the EOM of normal dogs or dogs with dystrophin deficiency, nor in the EOM of control humans or patients [149,150].
Another theory of resistance to Duchenne muscular dystrophy involves high antioxidant capacity of EOM [118] and/or the ability to maintain intracellular calcium homeostasis to protect against dystrophin-deficient necrosis. The expression profile of EOM suggests they are capable of maintaining intracellular Ca2+ homeostasis through increased expression of the phospholamban (PLN) gene, which regulates the activity of the sarcoplasmic reticulum Ca2+-ATPase. Decreased PLN levels characterize the abnormal Ca2+ flux observed in cardiac hypertrophy [6].
2.2. Amyotrophic Lateral Sclerosis
ALS is a severe neurodegenerative disease that leads to the death of primarily central and peripheral motor neurons. It is characterized by progressive muscle weakness, paresis, and paralysis. Although the progression of the disease can vary significantly, death usually occurs within 3 to 5 years due to respiratory failure. In ALS, all skeletal muscles are affected, and there are virtually no clinical symptoms from the EOM, even at the late stages of the disease.
Neurons of the oculomotor, trochlear, and abducens nuclei, located in the midbrain and responsible for eye movement, exhibit resistance to degeneration in ALS. This resistance enables patients, even in advanced stages of the disease, to communicate using eye movements, often with the aid of computers [151,152]. Furthermore, there is a vulnerability gradient, where faster motor units are affected earlier than slower ones [153]. Consequently, "fast" muscles, primarily dependent on glycolysis, become paralyzed before slower muscles [154].
Considering the plethora of unique characteristics of EOM, it is not surprising that in ALS, these muscles remain resistant to degeneration even at later stages of the disease. However, in patients with bulbar onset ALS, impairments in EOM functions, such as saccadic speeds and smooth pursuit movements, have been reported [155]. The primary pathoanatomical findings in the EOM of such patients included mild hypertrophy, MF atrophy, and increased connective tissue content [23]. Additionally, a reduction in the levels of myosin heavy chain isoforms MyHCI, MyHCIIa, and MyHCsto in MF was noted [156].
In a study published in the journal Investigative Ophthalmology & Visual Science [40], potential changes in EOM responsible for the preservation of their integrity, were described. It was demonstrated that the quantity of muscle fibers containing MyHCIIa significantly decreases, while the proportion of MyHCeom increases (in the "global" layer) in patients with ALS [40]. A more pronounced shift towards muscle fibers containing MyHCeom was noted in patients with bulbar onset compared to those with spinal onset. These findings correlate with more pronounced clinical manifestations in the EOM of patients with bulbar onset [40].
It is noteworthy that no significant differences in the types of MF containing MyHCIIa or MyHCeom were detected in the "orbital" layer of the EOM, both in donors with spinal and bulbar onset compared to control samples. These findings may indicate a higher resilience of the "orbital" layer MF to pathological processes in comparison with the "global" layer [40].
Intriguingly, the process MF type transition while preserving innervation between MyHCeom and neuronal cells may suggest that the neurons controlling these muscle fibers could also exhibit increased resistance to the pathogenic process in ALS.
3. Conclusions
Highly specialized EOM fundamentally differ from other muscles, warranting their classification as a distinct allotype of MF. They exhibit distinct responses to damage, aging, and diseases specifically affecting skeletal muscles, remaining anatomically and functionally intact even in late stages of such conditions as Duchenne muscular dystrophy or ALS.
Extraocular muscles (EOMs) not only exhibit resilience to individual pathologies but also differ in their anatomical structure, composition, biochemistry, and physiological function compared to other muscles. EOMs are innervated by cranial nerves rather than spinal motor neurons. Some MF within EOM display multiple innervations, forming en grappe endings, a feature not found in any muscle of adult mammals. Additionally, a portion of EOM MF may simultaneously exhibit both phasic and tonic contraction types, and the MyHC expression pattern encompasses the expression of nearly all known MyHC types.
The preservation of embryonic MyHC and fetal γ-AChR, heightened expression of genes associated with growth, development, and regeneration, unique embryology, complex muscle fiber types, diverse innervation patterns, and a metabolism distinct from skeletal muscles — all indicate that studying EOM could aid in the development of potential therapeutic strategies for muscular pathologies linked to diseases, injuries, and aging.
Author Contributions
Conceptualization, A.T., A.B. and S.N.; methodology, N.F., S.B. and D.Sh.; data curation, E.P., V.B. and M.A.; writing—original draft preparation, A.T. and A.B., writing—review and editing, A.T., A.B. and S.N.; supervision, I.Kh., E.Sh., O.G. and N.B.; project administration, O.G. and N.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kjellgren, D.; Thornell, L.E.; Andersen, J.; Pedrosa-Domellöf, F. Myosin heavy chain isoforms in human extraocular muscles. Invest Ophthalmol Vis Sci 2003, 44, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Kjellgren, D.; Ryan, M.; Ohlendieck, K.; Thornell, L.E.; Pedrosa-Domellöf, F. Sarco(endo)plasmic reticulum Ca2+ ATPases (SERCA1 and -2) in human extraocular muscles. Invest Ophthalmol Vis Sci 2003, 44, 5057–5062. [Google Scholar] [CrossRef] [PubMed]
- L, Muralidhar. Amyotrophic Lateral Sclerosis - Recent Advances and Therapeutic Challenges. IntechOpen, 2020. Crossref.
- Stuelsatz, P.; Shearer, A.; Li, Y.; Muir, L.A.; Ieronimakis, N.; Shen, Q.W.; Kirillova, I.; Yablonka-Reuveni, Z. Extraocular muscle satellite cells are high performance myo-engines retaining efficient regenerative capacity in dystrophin deficiency. Dev Biol 2015, 397, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.; Fitzpatrick, K.; McLoon, L.K. Extraocular Muscle Repair and Regeneration. Curr Ophthalmol Rep 2017, 5, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.D.; Gorospe, J.R.; Felder, E.; Bogdanovich, S.; Pedrosa-Domellöf, F.; Ahima, R.S.; Rubinstein, N.A.; Hoffman, E.P.; Khurana, T.S. Expression profiling reveals metabolic and structural components of extraocular muscles. Physiol Genomics 2002, 9, 71–84. [Google Scholar] [CrossRef]
- Sadeh, M. Extraocular muscles. In: ENgel AG, Franzini-Armstrong C, eds. Myology.New York, McGraw-Hill, 1994, 119–127.
- Ziermann, J.M.; Diogo, R.; Noden, D.M. Neural crest and the patterning of vertebrate craniofacial muscles. Genesis 2018, 56, e23097. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, B.L.; Gallina, D.; Thompson, H.; Kasprick, D.S.; Lucarelli, M.J.; Dootz, G.; Nelson, C.; McGonnell, I.M.; Kahana, A. Development of extraocular muscles requires early signals from periocular neural crest and the developing eye. Arch Ophthalmol 2011, 129, 1030–1041. [Google Scholar] [CrossRef]
- Noden, D.M.; Trainor, P.A. Relations and interactions between cranial mesoderm and neural crest populations. J Anat 2005, 207, 575–601. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N., Motlagh, M., Singh G. Anatomy, Head and Neck, Eye Arteries. In: StatPearls. Treasure Island (FL): StatPearls Publishing; July 24, 2023.
- Azzam, D., Cypen, S., Tao J. Anatomy, Head and Neck: Eye Ophthalmic Vein. In: StatPearls. Treasure Island (FL): StatPearls Publishing; July 24, 2023.
- Olitsky, S.E.; Coats, D.K. Complications of Strabismus Surgery. Middle East Afr J Ophthalmol 2015, 22, 271–278. [Google Scholar] [CrossRef]
- Joyce, C., Le, P.H., Peterson, D.C. Neuroanatomy, Cranial Nerve 3 (Oculomotor). In: StatPearls. Treasure Island (FL): StatPearls Publishing; March 27, 2023.
- Zanganeh, T., Legault, G.L. Extraocular Muscle Management With Orbital and Globe Trauma. In: StatPearls. Treasure Island (FL): StatPearls Publishing; July 17, 2023.
- Lacey, H.; Oliphant, H.; Smith, C.; Koenig, M.; Rajak, S. Topographical anatomy of the annulus of Zinn. Sci Rep 2022, 12, 1064. [Google Scholar] [CrossRef]
- Noden, D.M. The embryonic origins of avian cephalic and cervical muscles and associated connective tissues. Am J Anat 1983, 168, 257–276. [Google Scholar] [CrossRef]
- Tzahor, E.; Kempf, H.; Mootoosamy, R.C.; Poon, A.C.; Abzhanov, A.; Tabin, C.J.; Dietrich, S.; Lassar, A.B. Antagonists of Wnt and BMP signaling promote the formation of vertebrate head muscle. Genes Dev 2003, 17, 3087–3099. [Google Scholar] [CrossRef] [PubMed]
- von Scheven, G.; Alvares, L.E.; Mootoosamy, R.C.; Dietrich, S. Neural tube derived signals and Fgf8 act antagonistically to specify eye versus mandibular arch muscles. Development 2006, 133, 2731–2745. [Google Scholar] [CrossRef] [PubMed]
- Gage, P.J.; Suh, H.; Camper, S.A. Dosage requirement of Pitx2 for development of multiple organs. Development 1999, 126, 4643–4651. [Google Scholar] [CrossRef]
- Schilling, T.F.; Walker, C.; Kimmel, C.B. The chinless mutation and neural crest cell interactions in zebrafish jaw development. Development 1996, 122, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Rinon, A.; Lazar, S.; Marshall, H.; Büchmann-Møller, S.; Neufeld, A.; Elhanany-Tamir, H.; Taketo, M.M.; Sommer, L.; Krumlauf, R.; Tzahor, E. Cranial neural crest cells regulate head muscle patterning and differentiation during vertebrate embryogenesis. Development 2007, 134, 3065–3075. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, M.; Liu, J.X.; Brännström, T.; Andersen, P.M.; Stål, P.; Pedrosa-Domellöf, F. Human extraocular muscles in ALS. Invest Ophthalmol Vis Sci 2010, 51, 3494–3501. [Google Scholar] [CrossRef] [PubMed]
- Mühlendyck, H. [Age-dependent changes in transverse sections of muscle fibres from the exterior eye muscles in man]. Z Gerontol 1979, 12, 46–59. [Google Scholar] [PubMed]
- Henson, C.; Staunton, H.; Brett, F.M. Does ageing have an effect on midbrain premotor nuclei for vertical eye movements. Mov Disord 2003, 18, 688–694. [Google Scholar] [CrossRef]
- Stål, P.; Eriksson, P.O.; Schiaffino, S.; Butler-Browne, G.S.; Thornell, L.E. Differences in myosin composition between human oro-facial, masticatory and limb muscles: enzyme-, immunohisto- and biochemical studies. J Muscle Res Cell Motil 1994, 15, 517–534. [Google Scholar] [CrossRef]
- Hoogenraad, T.U.; Jennekens, F.G.; Tan, K.E. Histochemical fibre types in human extraocular muscles, an investigation of inferior oblique muscle. Acta Neuropathol 1979, 45, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Budak, M.T.; Bogdanovich, S.; Wiesen, M.H.; Lozynska, O.; Khurana, T.S.; Rubinstein, N.A. Layer-specific differences of gene expression in extraocular muscles identified by laser-capture microscopy. Physiol Genomics 2004, 20, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Spencer, R.F.; Porter, J.D. Structural organization of the extraocular muscles. Rev Oculomot Res 1988, 2, 33–79. [Google Scholar] [PubMed]
- Spencer, R.F.; Porter, J.D. Biological organization of the extraocular muscles. Prog Brain Res 2006, 151, 43–80. [Google Scholar]
- Porter, J.D.; Burns, L.A.; May, P.J. Morphological substrate for eyelid movements: innervation and structure of primate levator palpebrae superioris and orbicularis oculi muscles. J Comp Neurol 1989, 287, 64–81. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Rossi, A.C.; Smerdu, V.; Leinwand, L.A.; Reggiani, C. Developmental myosins: expression patterns and functional significance. Skelet Muscle 2015, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.; Martin, L.J. Skeletal muscle-restricted expression of human SOD1 causes motor neuron degeneration in transgenic mice. Hum Mol Genet 2010, 19, 2284–2302. [Google Scholar] [CrossRef]
- Wieczorek, D.F.; Periasamy, M.; Butler-Browne, G.S.; Whalen, R.G.; Nadal-Ginard, B. Co-expression of multiple myosin heavy chain genes, in addition to a tissue-specific one, in extraocular musculature. J Cell Biol 1985, 101, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Yu Wai Man, C.Y.; Chinnery, P.F.; Griffiths, P.G. Extraocular muscles have fundamentally distinct properties that make them selectively vulnerable to certain disorders. Neuromuscul Disord 2005, 15, 17–23. [Google Scholar] [CrossRef]
- Briggs, M.M.; Schachat, F. Early specialization of the superfast myosin in extraocular and laryngeal muscles. J Exp Biol 2000, 203, 2485–2494. [Google Scholar] [CrossRef]
- Briggs, M.M.; Schachat, F. The superfast extraocular myosin (MYH13) is localized to the innervation zone in both the global and orbital layers of rabbit extraocular muscle. J Exp Biol 2002, 205, 3133–3142. [Google Scholar] [CrossRef]
- Rossi, A.C.; Mammucari, C.; Argentini, C.; Reggiani, C.; Schiaffino, S. Two novel/ancient myosins in mammalian skeletal muscles: MYH14/7b and MYH15 are expressed in extraocular muscles and muscle spindles. J Physiol 2010, 588, 353–364. [Google Scholar] [CrossRef]
- Lee, L.A.; Karabina, A.; Broadwell, L.J.; Leinwand, L.A. The ancient sarcomeric myosins found in specialized muscles. Skelet Muscle 2019, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Mascarello, F.; Toniolo, L.; Cancellara, P.; Reggiani, C.; Maccatrozzo, L. Expression and identification of 10 sarcomeric MyHC isoforms in human skeletal muscles of different embryological origin. Diversity and similarity in mammalian species. Ann Anat 2016, 207, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Kranjc, B.S.; Sketelj, J.; Albis, A.D.; Ambroz, M.; Erzen, I. Fibre types and myosin heavy chain expression in the ocular medial rectus muscle of the adult rat. J Muscle Res Cell Motil 2000, 21, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Stirn Kranjc, B.; Smerdu, V.; Erzen, I. Histochemical and immunohistochemical profile of human and rat ocular medial rectus muscles. Graefes Arch Clin Exp Ophthalmol 2009, 247, 1505–1515. [Google Scholar] [CrossRef]
- Rushbrook, J.I.; Weiss, C.; Ko, K.; Feuerman, M.H.; Carleton, S.; Ing, A.; Jacoby, J. Identification of alpha-cardiac myosin heavy chain mRNA and protein in extraocular muscle of the adult rabbit. J Muscle Res Cell Motil 1994, 15, 505–515. [Google Scholar] [CrossRef]
- Rubinstein, N.A.; Hoh, J.F. The distribution of myosin heavy chain isoforms among rat extraocular muscle fiber types. Invest Ophthalmol Vis Sci 2000, 41, 3391–3398. [Google Scholar]
- Bicer, S.; Reiser, P.J. Myosin isoform expression in dog rectus muscles: patterns in global and orbital layers and among single fibers. Invest Ophthalmol Vis Sci 2009, 50, 157–167. [Google Scholar] [CrossRef]
- McLoon, L.K., Andrade, F. Craniofacial Muscles: A New Framework for Understanding the Effector Side of Craniofacial Muscle Control. L.K. McLoon and F. Andrade, editors. Springer New York, New York, NY, 2012.
- Mayr, R. Structure and distribution of fibre types in the external eye muscles of the rat. Tissue Cell 1971, 3, 433–462. [Google Scholar] [CrossRef]
- Rashed, R.M.; El-Alfy, S.H.; Mohamed, I.K. Histochemical analysis of muscle fiber types of rat superior rectus extraocular muscle. Acta Histochem 2010, 112, 536–545. [Google Scholar] [CrossRef]
- Rashed, R.M.; El-Alfy, S.H. Ultrastructural organization of muscle fiber types and their distribution in the rat superior rectus extraocular muscle. Acta Histochem 2012, 114, 217–225. [Google Scholar] [CrossRef]
- Wasicky, R.; Ziya-Ghazvini, F.; Blumer, R.; Lukas, J.R.; Mayr, R. Muscle fiber types of human extraocular muscles: a histochemical and immunohistochemical study. Invest Ophthalmol Vis Sci 2000, 41, 980–990. [Google Scholar] [PubMed]
- Shear, T.D.; Martyn, J.A. Physiology and biology of neuromuscular transmission in health and disease. J Crit Care 2009, 24, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Lepore, E.; Casola, I.; Dobrowolny, G.; Musarò, A. Neuromuscular Junction as an Entity of Nerve-Muscle Communication. Cells 2019, 8, 906. [Google Scholar] [CrossRef]
- Hughes, B.W.; Kusner, L.L.; Kaminski, H.J. Molecular architecture of the neuromuscular junction. Muscle Nerve 2006, 33, 445–46. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.E.; Levine, D.N.; Tsairis, P.; Zajac, F.E. Physiological types and histochemical profiles in motor units of the cat gastrocnemius. J Physiol 1973, 234, 723–748. [Google Scholar] [CrossRef]
- Ogata, T.; Yamasaki, Y. Scanning electron-microscopic study on the three-dimensional structure of motor endplates of the slow (tonic) muscle fibers in the frog, Rana n. nigromaculata. Cell Tissue Res 1988, 252, 211–213. [Google Scholar] [CrossRef]
- Boyd-Clark, L.C.; Briggs, C.A.; Galea, M.P. Comparative histochemical composition of muscle fibres in a pre- and a postvertebral muscle of the cervical spine. J Anat 2001, 199, 709–716. [Google Scholar] [CrossRef]
- Callister, R.J.; Callister, R.; Peterson, E.H. Design and control of the head retractor muscle in a turtle, Pseudemys (Trachemys) scripta: I. Architecture and histochemistry of single muscle fibers. J Comp Neurol 1992, 325, 405–421. [Google Scholar] [CrossRef]
- Э Sosnicki, A.A.; Lutz, G.J.; Rome, L.C.; Goble, D.O. Histochemical and molecular determination of fiber types in chemically skinned single equine skeletal muscle fibers. J Histochem Cytochem 1989, 37, 1731–1738. [Google Scholar] [CrossRef] [PubMed]
- Sanchís-Moysi, J.; Idoate, F.; Olmedillas, H.; Guadalupe-Grau, A.; Alayón, S.; Carreras, A.; Dorado, C.; Calbet, J.A. The upper extremity of the professional tennis player: muscle volumes, fiber-type distribution and muscle strength. Scand J Med Sci Sports 2010. [CrossRef] [PubMed]
- Smith M. Neurological Rehabilitation. 5th ed. Darcy A. Umphred St. Louis, MO.; Mosby Elsevier. 2007; ISBN-13: 978-0-323-03306-0.
- Kuffler, S.W.; Vaughan Williams, E.M. Properties of the 'slow' skeletal muscles fibres of the frog. J Physiol 1953, 121, 318–340. [Google Scholar] [CrossRef] [PubMed]
- Kuffler, S.W.; Vaughan Williams, E.M. Small-nerve junctional potentials; the distribution of small motor nerves to frog skeletal muscle, and the membrane characteristics of the fibres they innervate. J Physiol 1953, 121, 289–317. [Google Scholar] [CrossRef] [PubMed]
- Hess, A.; Pilar, G. Slow fibres in the extraocular muscles of the cat. J Physiol 1963, 169, 780–798. [Google Scholar] [CrossRef] [PubMed]
- Hess, A. Two Kinds of Extrafusal Muscle Fibers and Their Nerve Endings in the Garter Snake. Am J Anat 1963, 113, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Hess, A. Further morphological observations of 'en plaque' and 'en grappe' nerve endings on mammalian extrafusal muscle fibers with the cholinesterase technique. Rev Can Biol 1962, 21, 241–248. [Google Scholar] [PubMed]
- Dietert, S.E. The demonstration of different types of muscle fibers in human extraocular muscle fibers in human extraocular muscle by electron microscopy and cholinesterase staining. Invest Ophthalmol 1965, 4, 51–63. [Google Scholar] [PubMed]
- Lichtman, J.W.; Wilkinson, R.S.; Rich, M.M. Multiple innervation of tonic endplates revealed by activity-dependent uptake of fluorescent probes. Nature 1985, 314, 357–359. [Google Scholar] [CrossRef]
- Ridge, R.M. Different types of extrafusal muscle fibres in snake costocutaneous muscles. J Physiol 1971, 217, 393–418. [Google Scholar] [CrossRef]
- Oda, K. Motor innervation and acetylcholine receptor distribution of human extraocular muscle fibres. J Neurol Sci 1986, 74, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Kupfer, C. Motor innervation of extraocular muscle. J Physiol 1960, 153, 522–526. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C. Fiber types in mammalian skeletal muscles. Physiol Rev 2011, 91, 1447–1531. [Google Scholar] [CrossRef]
- Liu, J.X.; Domellöf, F.P. A Novel Type of Multiterminal Motor Endplate in Human Extraocular Muscles. Invest Ophthalmol Vis Sci 2018, 59, 539–548. [Google Scholar] [CrossRef]
- Carry, M.R.; Ringel, S.P.; Starcevich, J.M. Mitochondrial morphometrics of histochemically identified human extraocular muscle fibers. Anat Rec 1986, 214, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Demer, J.L.; Oh, S.Y.; Poukens, V. Evidence for active control of rectus extraocular muscle pulleys. Invest Ophthalmol Vis Sci 2000, 41, 1280–1290. [Google Scholar]
- Porter, J.D.; Baker, R.S. Muscles of a different 'color': the unusual properties of the extraocular muscles may predispose or protect them in neurogenic and myogenic disease. Neurology 1996, 46, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Pachter, B.R. Fiber composition of the superior rectus extraocular muscle of the rhesus macaque. J Morphol 1982, 174, 237–250. [Google Scholar] [CrossRef]
- Chiarandini, D.J.; Davidowitz, J. Structure and function of extraocular muscle fibers. Curr Top Eye Res 1979, 1, 91–142. [Google Scholar]
- Khanna, S.; Porter, J.D. Evidence for rectus extraocular muscle pulleys in rodents. Invest Ophthalmol Vis Sci 2001, 42, 1986–1992. [Google Scholar]
- Chiarandini, D.J.; Stefani, E. Electrophysiological identification of two types of fibres in rat extraocular muscles. J Physiol 1979, 290, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, J.; Chiarandini, D.J.; Stefani, E. Electrical properties and innervation of fibers in the orbital layer of rat extraocular muscles. J Neurophysiol 1989, 61, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, H.J.; Kusner, L.L.; Block, C.H. Expression of acetylcholine receptor isoforms at extraocular muscle endplates. Invest Ophthalmol Vis Sci 1996, 37, 345–351. [Google Scholar]
- Gu, Y.; Hall, Z.W. Immunological evidence for a change in subunits of the acetylcholine receptor in developing and denervated rat muscle. Neuron 1988, 1, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Fraterman, S.; Khurana, T.S.; Rubinstein, N.A. Identification of acetylcholine receptor subunits differentially expressed in singly and multiply innervated fibers of extraocular muscles. Invest Ophthalmol Vis Sci 2006, 47, 3828–3834. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, F.; Vicini, S.; Schuetze, S.M. Embryonic acetylcholine receptors guarantee spontaneous contractions in rat developing muscle. Nature 1988, 335, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Witzemann, V.; Schwarz, H.; Koenen, M.; Berberich, C.; Villarroel, A.; Wernig, A.; Brenner, H.R.; Sakmann, B. Acetylcholine receptor epsilon-subunit deletion causes muscle weakness and atrophy in juvenile and adult mice. Proc Natl Acad Sci U S A 1996, 93, 13286–13291. [Google Scholar] [CrossRef]
- Liu, Y.; Padgett, D.; Takahashi, M.; Li, H.; Sayeed, A.; Teichert, R.W.; Olivera, B.M.; McArdle, J.J.; Green, W.N.; Lin, W. Essential roles of the acetylcholine receptor gamma-subunit in neuromuscular synaptic patterning. Development 2008, 135, 1957–1967. [Google Scholar] [CrossRef]
- Hoffmann, K.; Muller, J.S.; Stricker, S.; Megarbane, A.; Rajab, A.; Lindner, T.H.; Cohen, M.; Chouery, E.; Adaimy, L.; Ghanem, I.; Delague, V.; Boltshauser, E.; Talim, B.; Horvath, R.; Robinson, P.N.; Lochmüller, H.; Hübner, C.; Mundlos, S. Escobar syndrome is a prenatal myasthenia caused by disruption of the acetylcholine receptor fetal gamma subunit. Am J Hum Genet 2006, 79, 303–312. [Google Scholar] [CrossRef]
- Morgan, N.V.; Brueton, L.A.; Cox, P.; Greally, M.T.; Tolmie, J.; Pasha, S.; Aligianis, I.A.; van Bokhoven, H.; Marton, T.; Al-Gazali, L.; Morton, J.E.; Oley, C.; Johnson, C.A.; Trembath, R.C.; Brunner, H.G.; Maher, E.R. Mutations in the embryonal subunit of the acetylcholine receptor (CHRNG) cause lethal and Escobar variants of multiple pterygium syndrome. Am J Hum Genet 2006, 79, 390–395. [Google Scholar] [CrossRef]
- Vogt, J.; Morgan, N.V.; Rehal, P.; Faivre, L.; Brueton, L.A.; Becker, K.; Fryns, J.P.; Holder, S.; Islam, L.; Kivuva, E.; Lynch, S.A.; Touraine, R.; Wilson, L.C.; MacDonald, F.; Maher, E.R. CHRNG genotype-phenotype correlations in the multiple pterygium syndromes. J Med Genet 2012, 49, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Mishina, M.; Takai, T.; Imoto, K.; Noda, M.; Takahashi, T.; Numa, S.; Methfessel, C.; Sakmann, B. Molecular distinction between fetal and adult forms of muscle acetylcholine receptor. Nature 1986, 321, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Bär, H.; Strelkov, S.V.; Sjöberg, G.; Aebi, U.; Herrmann, H. The biology of desmin filaments: how do mutations affect their structure, assembly, and organisation. J Struct Biol 2004, 148, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Askanas, V.; Bornemann, A.; Engel, W.K. Immunocytochemical localization of desmin at human neuromuscular junctions. Neurology 1990, 40, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Clemen, C.S.; Herrmann, H.; Strelkov, S.V.; Schröder, R. Desminopathies: pathology and mechanisms. Acta Neuropathol 2013, 125, 47–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Pedrosa Domellöf, F. Complex Correlations Between Desmin Content, Myofiber Types, and Innervation Patterns in the Human Extraocular Muscles. Invest Ophthalmol Vis Sci 2020, 61, 15. [Google Scholar] [CrossRef] [PubMed]
- Janbaz, A.H.; Lindström, M.; Liu, J.X.; Pedrosa Domellöf, F. Intermediate filaments in the human extraocular muscles. Invest Ophthalmol Vis Sci 2014, 55, 5151–5159. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Brännström, T.; Andersen, P.M.; Pedrosa-Domellöf, F. Different impact of ALS on laminin isoforms in human extraocular muscles versus limb muscles. Invest Ophthalmol Vis Sci 2011, 52, 4842–4852. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Brännström, T.; Andersen, P.M.; Pedrosa-Domellöf, F. Distinct changes in synaptic protein composition at neuromuscular junctions of extraocular muscles versus limb muscles of ALS donors. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- McLoon, L.K.; Harandi, V.M.; Brännström, T.; Andersen, P.M.; Liu, J.X. Wnt and extraocular muscle sparing in amyotrophic lateral sclerosis. Invest Ophthalmol Vis Sci 2014, 55, 5482–5496. [Google Scholar] [CrossRef]
- Harandi, V.M.; Gaied, A.R.; Brännström, T.; Pedrosa Domellöf, F.; Liu, J.X. Unchanged Neurotrophic Factors and Their Receptors Correlate With Sparing in Extraocular Muscles in Amyotrophic Lateral Sclerosis. Invest Ophthalmol Vis Sci 2016, 57, 6831–6842. [Google Scholar] [CrossRef] [PubMed]
- Agbulut, O.; Li, Z.; Mouly, V.; Butler-Browne, G.S. Analysis of skeletal and cardiac muscle from desmin knock-out and normal mice by high resolution separation of myosin heavy-chain isoforms. Biol Cell 1996, 88, 131–135. [Google Scholar]
- Balogh, J.; Merisckay, M.; Li, Z.; Paulin, D.; Arner, A. Hearts from mice lacking desmin have a myopathy with impaired active force generation and unaltered wall compliance. Cardiovasc Res 2002, 53, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Kjellgren, D.; Thornell, L.E.; Virtanen, I.; Pedrosa-Domellöf, F. Laminin isoforms in human extraocular muscles. Invest Ophthalmol Vis Sci 2004, 45, 4233–4239. [Google Scholar] [CrossRef] [PubMed]
- McLoon, L.K.; Vicente, A.; Fitzpatrick, K.R.; Lindström, M.; Pedrosa Domellöf, F. Composition, Architecture, and Functional Implications of the Connective Tissue Network of the Extraocular Muscles. Invest Ophthalmol Vis Sci 2018, 59, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Dennhag, N.; Domellöf, F.P. Understanding the extraocular muscles: connective tissue, motor endplates and the cytoskeleton. Biochem (Lond) 2020, 42, 52–57. [Google Scholar] [CrossRef]
- Smith, T.J. Insights into the role of fibroblasts in human autoimmune diseases. Clin Exp Immunol 2005, 141, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Bednarczuk, T.; Gopinath, B.; Ploski, R.; Wall, J.R. Susceptibility genes in Graves' ophthalmopathy: searching for a needle in a haystack. Clin Endocrinol (Oxf) 2007, 67, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J. Novel aspects of orbital fibroblast pathology. J Endocrinol Invest 2004, 27, 246–253. [Google Scholar] [CrossRef]
- Gage, P.J.; Rhoades, W.; Prucka, S.K.; Hjalt, T. Fate maps of neural crest and mesoderm in the mammalian eye. Invest Ophthalmol Vis Sci 2005, 46, 4200–4208. [Google Scholar] [CrossRef]
- Kusner, L.L.; Young, A.; Tjoe, S.; Leahy, P.; Kaminski, H.J. Perimysial fibroblasts of extraocular muscle, as unique as the muscle fibers. Invest Ophthalmol Vis Sci 2010, 51, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Morgan, B.P.; Chamberlain-Banoub, J.; Neal, J.W.; Song, W.; Mizuno, M.; Harris, C.L. The membrane attack pathway of complement drives pathology in passively induced experimental autoimmune myasthenia gravis in mice. Clin Exp Immunol 2006, 146, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, H.J.; Kusner, L.L.; Richmonds, C.; Medof, M.E.; Lin, F. Deficiency of decay accelerating factor and CD59 leads to crisis in experimental myasthenia. Exp Neurol 2006, 202, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.J.; Afifiyan, N.; Sand, D.; Naik, V.; Said, J.; Pollock, S.J.; Chen, B.; Phipps, R.P.; Goldberg, R.A.; Smith, T.J.; Douglas, R.S. Orbital fibroblasts from patients with thyroid-associated ophthalmopathy overexpress CD40: CD154 hyperinduces IL-6, IL-8, and MCP-1. Invest Ophthalmol Vis Sci 2009, 50, 2262–2268. [Google Scholar] [CrossRef] [PubMed]
- Laban-Guceva, N.; Bogoev, M.; Antova, M. Serum concentrations of interleukin (IL-)1alpha, 1beta, 6 and tumor necrosis factor (TNF-) alpha in patients with thyroid eye disease (TED). Med Arh 2007, 61, 203–206. [Google Scholar] [PubMed]
- Kaminski, H.J.; Li, Z.; Richmonds, C.; Ruff, R.L.; Kusner, L. Susceptibility of ocular tissues to autoimmune diseases. Ann N Y Acad Sci 2003, 998, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.D.; Khanna, S.; Kaminski, H.J.; Rao, J.S.; Merriam, A.P.; Richmonds, C.R.; Leahy, P.; Li, J.; Andrade, F.H. Extraocular muscle is defined by a fundamentally distinct gene expression profile. Proc Natl Acad Sci U S A 2001, 98, 12062–12067. [Google Scholar] [CrossRef]
- Nguyen, B.; Gopinath, B.; Tani, J.; Wescombe, L.; Wall, J.R. Peripheral blood T lymphocyte sensitisation against calsequestrin and flavoprotein in patients with Graves' ophthalmopathy. Autoimmunity 2008, 41, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, R.J.; Chow, C.K.; St Clair, D.K.; Porter, J.D. Extraocular, limb and diaphragm muscle group-specific antioxidant enzyme activity patterns in control and mdx mice. J Neurol Sci 1996, 139, 180–186. [Google Scholar] [CrossRef]
- McLoon, L.K.; Wirtschafter, J. Activated satellite cells in extraocular muscles of normal adult monkeys and humans. Invest Ophthalmol Vis Sci 2003, 44, 1927–1932. [Google Scholar] [CrossRef]
- Formicola, L.; Marazzi, G.; Sassoon, D.A. The extraocular muscle stem cell niche is resistant to ageing and disease. Front Aging Neurosci 2014, 6, 328. [Google Scholar] [CrossRef] [PubMed]
- La Rovere, R.M.; Quattrocelli, M.; Pietrangelo, T.; Di Filippo, E.S.; Maccatrozzo, L.; Cassano, M.; Mascarello, F.; Barthélémy, I.; Blot, S.; Sampaolesi, M.; Fulle, S. Myogenic potential of canine craniofacial satellite cells. Front Aging Neurosci 2014, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Carrero-Rojas, G.; Benítez-Temiño, B.; Pastor, A.M.; Davis López de Carrizosa, M.A. Muscle Progenitors Derived from Extraocular Muscles Express Higher Levels of Neurotrophins and their Receptors than other Cranial and Limb Muscles. Cells 2020, 9, 747. [Google Scholar] [CrossRef] [PubMed]
- Renault, V.; Piron-Hamelin, G.; Forestier, C.; DiDonna, S.; Decary, S.; Hentati, F.; Saillant, G.; Butler-Browne, G.S.; Mouly, V. Skeletal muscle regeneration and the mitotic clock. Exp Gerontol 2000, 35, 711–719. [Google Scholar] [CrossRef]
- Kallestad, K.M.; Hebert, S.L.; McDonald, A.A.; Daniel, M.L.; Cu, S.R.; McLoon, L.K. Sparing of extraocular muscle in aging and muscular dystrophies: a myogenic precursor cell hypothesis. Exp Cell Res 2011, 317, 873–885. [Google Scholar] [CrossRef]
- Hebert, S.L.; Daniel, M.L.; McLoon, L.K. The role of Pitx2 in maintaining the phenotype of myogenic precursor cells in the extraocular muscles. PLoS One 2013, 8, e58405. [Google Scholar] [CrossRef] [PubMed]
- McGeachie, J.K.; Grounds, M.D. The timing between skeletal muscle myoblast replication and fusion into myotubes, and the stability of regenerated dystrophic myofibres: an autoradiographic study in mdx mice. J Anat 1999, 194 (Pt 2), 287–295. [Google Scholar] [CrossRef] [PubMed]
- Heslop, L.; Morgan, J.E.; Partridge, T.A. Evidence for a myogenic stem cell that is exhausted in dystrophic muscle. J Cell Sci 2000, 113 (Pt 12), 2299–2308. [Google Scholar] [CrossRef]
- Girolamo, D.D.; Benavente-Diaz, M.; Murolo, M.; Grimaldi, A.; Lopes, P.T.; Evano, B.; Kuriki, M.; Gioftsidi, S.; Laville, V.; Tinevez, J.Y.; Letort, G.; Mella, S.; Tajbakhsh, S.; Comai, G. Extraocular muscle stem cells exhibit distinct cellular properties associated with non-muscle molecular signatures. Development 2024, 151, dev202144. [Google Scholar] [CrossRef]
- Zhou, Y.; Cheng, G.; Dieter, L.; Hjalt, T.A.; Andrade, F.H.; Stahl, J.S.; Kaminski, H.J. An altered phenotype in a conditional knockout of Pitx2 in extraocular muscle. Invest Ophthalmol Vis Sci 2009, 50, 4531–4541. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, D.; Kaminski, H.J. Pitx2 regulates myosin heavy chain isoform expression and multi-innervation in extraocular muscle. J Physiol 2011, 589, 4601–4614. [Google Scholar] [CrossRef]
- Benítez-Temiño, B.; Davis-López de Carrizosa, M.A.; Morcuende, S.; Matarredona, E.R.; de la Cruz, R.R.; Pastor, A.M. Functional Diversity of Neurotrophin Actions on the Oculomotor System. Int J Mol Sci 2016, 17, 2016. [Google Scholar] [CrossRef] [PubMed]
- McLoon, L.K.; Christiansen, S.P. Increasing extraocular muscle strength with insulin-like growth factor II. Invest Ophthalmol Vis Sci 2003, 44, 3866–3872. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.Y.; von Bartheld, C.S. Expression of insulin-like growth factor 1 isoforms in the rabbit oculomotor system. Growth Horm IGF Res 2011, 21, 228–232. [Google Scholar] [CrossRef]
- Davis-López de Carrizosa, M.A.; Morado-Díaz, C.J.; Tena, J.J.; Benítez-Temiño, B.; Pecero, M.L.; Morcuende, S.R.; de la Cruz, R.R.; Pastor, A.M. Complementary actions of BDNF and neurotrophin-3 on the firing patterns and synaptic composition of motoneurons. J Neurosci 2009, 29, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, C.L.; Fleuriet, J.; Walton, M.M.; Mustari, M.J.; McLoon, L.K. Adaptation of slow myofibers: the effect of sustained BDNF treatment of extraocular muscles in infant nonhuman primates. Invest Ophthalmol Vis Sci 2015, 56, 3467–3483. [Google Scholar] [CrossRef]
- Agarwal, A.B.; Feng, C.Y.; Altick, A.L.; Quilici, D.R.; Wen, D.; Johnson, L.A.; von Bartheld, C.S. Altered Protein Composition and Gene Expression in Strabismic Human Extraocular Muscles and Tendons. Invest Ophthalmol Vis Sci 2016, 57, 5576–5585. [Google Scholar] [CrossRef] [PubMed]
- Harandi, V.M.; Lindquist, S.; Kolan, S.S.; Brännström, T.; Liu, J.X. Analysis of neurotrophic factors in limb and extraocular muscles of mouse model of amyotrophic lateral sclerosis. PLoS One 2014, 9, e109833. [Google Scholar] [CrossRef]
- Steljes, T.P.; Kinoshita, Y.; Wheeler, E.F.; Oppenheim, R.W.; von Bartheld, C.S. Neurotrophic factor regulation of developing avian oculomotor neurons: differential effects of BDNF and GDNF. J Neurobiol 1999, 41, 295–315. [Google Scholar] [CrossRef]
- Chen, J.; von Bartheld, C.S. Role of exogenous and endogenous trophic factors in the regulation of extraocular muscle strength during development. Invest Ophthalmol Vis Sci 2004, 45, 3538–3545. [Google Scholar] [CrossRef]
- Hassan, S.M.; Jennekens, F.G.; Veldman, H. Botulinum toxin-induced myopathy in the rat. Brain 1995, 118 (Pt 2), 533–545. [Google Scholar] [CrossRef] [PubMed]
- Pinter, M.J.; Vanden Noven, S.; Muccio, D.; Wallace, N. Axotomy-like changes in cat motoneuron electrical properties elicited by botulinum toxin depend on the complete elimination of neuromuscular transmission. J Neurosci 1991, 11, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Kranjc, B.S.; Sketelj, J.; D'Albis, A.; Erzen, I. Long-term changes in myosin heavy chain composition after botulinum toxin a injection into rat medial rectus muscle. Invest Ophthalmol Vis Sci 2001, 42, 3158–3164. [Google Scholar] [PubMed]
- Spencer, R.F.; McNeer, K.W. Botulinum toxin paralysis of adult monkey extraocular muscle. Structural alterations in orbital, singly innervated muscle fibers. Arch Ophthalmol 1987, 105, 1703–1711. [Google Scholar] [CrossRef] [PubMed]
- Ugalde, I.; Christiansen, S.P.; McLoon, L.K. Botulinum toxin treatment of extraocular muscles in rabbits results in increased myofiber remodeling. Invest Ophthalmol Vis Sci 2005, 46, 4114–4120. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowski, B.; Pulliam, C.; Betta, N.D.; Kardon, G.; Olwin, B.B. Pervasive satellite cell contribution to uninjured adult muscle fibers. Skelet Muscle 2015, 5, 42. [Google Scholar] [CrossRef]
- Karpati, G.; Carpenter, S. Small-caliber skeletal muscle fibers do not suffer deleterious consequences of dystrophic gene expression. Am J Med Genet 1986, 25, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Andrade, F.H.; Porter, J.D.; Kaminski, H.J. Eye muscle sparing by the muscular dystrophies: lessons to be learned. Microsc Res Tech 2000, 48, 192–203. [Google Scholar] [CrossRef]
- Kuang, S.; Chargé, S.B.; Seale, P.; Huh, M.; Rudnicki, M.A. Distinct roles for Pax7 and Pax3 in adult regenerative myogenesis. J Cell Biol 2006, 172, 103–113. [Google Scholar] [CrossRef]
- Khurana, T.S.; Prendergast, R.A.; Alameddine, H.S.; Tomé, F.M.; Fardeau, M.; Arahata, K.; Sugita, H.; Kunkel, L.M. Absence of extraocular muscle pathology in Duchenne's muscular dystrophy: role for calcium homeostasis in extraocular muscle sparing. J Exp Med 1995, 182, 467–475. [Google Scholar] [CrossRef]
- Matsumura, K.; Ervasti, J.M.; Ohlendieck, K.; Kahl, S.D.; Campbell, K.P. Association of dystrophin-related protein with dystrophin-associated proteins in mdx mouse muscle. Nature 1992, 360, 588–591. [Google Scholar] [CrossRef] [PubMed]
- Caligari, M.; Godi, M.; Guglielmetti, S.; Franchignoni, F.; Nardone, A. Eye tracking communication devices in amyotrophic lateral sclerosis: impact on disability and quality of life. Amyotroph Lateral Scler Frontotemporal Degener 2013, 14, 546–552. [Google Scholar] [CrossRef]
- Kubota, M.; Sakakihara, Y.; Uchiyama, Y.; Nara, A.; Nagata, T.; Nitta, H.; Ishimoto, K.; Oka, A.; Horio, K.; Yanagisawa, M. New ocular movement detector system as a communication tool in ventilator-assisted Werdnig-Hoffmann disease. Dev Med Child Neurol 2000, 42, 61–64. [Google Scholar]
- Pun, S.; Santos, A.F.; Saxena, S.; Xu, L.; Caroni, P. Selective vulnerability and pruning of phasic motoneuron axons in motoneuron disease alleviated by CNTF. Nat Neurosci 2006, 9, 408–419. [Google Scholar] [CrossRef]
- Hegedus, J.; Putman, C.T.; Gordon, T. Time course of preferential motor unit loss in the SOD1 G93A mouse model of amyotrophic lateral sclerosis. Neurobiol Dis 2007, 28, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Donaghy, C.; Pinnock, R.; Abrahams, S.; Cardwell, C.; Hardiman, O.; Patterson, V.; McGivern, R.C.; Gibson, J.M. Slow saccades in bulbar-onset motor neurone disease. J Neurol 2010, 257, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.H.; Kim, J.I.; Lim, Y.M.; Kim, K.K. Abnormal Oculomotor Functions in Amyotrophic Lateral Sclerosis. J Clin Neurol 2018, 14, 464–471. [Google Scholar] [CrossRef]
- Tjust, A.E.; Danielsson, A.; Andersen, P.M.; Brännström, T.; Pedrosa Domellöf, F. Impact of Amyotrophic Lateral Sclerosis on Slow Tonic Myofiber Composition in Human Extraocular Muscles. Invest Ophthalmol Vis Sci 2017, 58, 3708–3715. [Google Scholar] [CrossRef]
Figure 1.
Extraocular muscles.
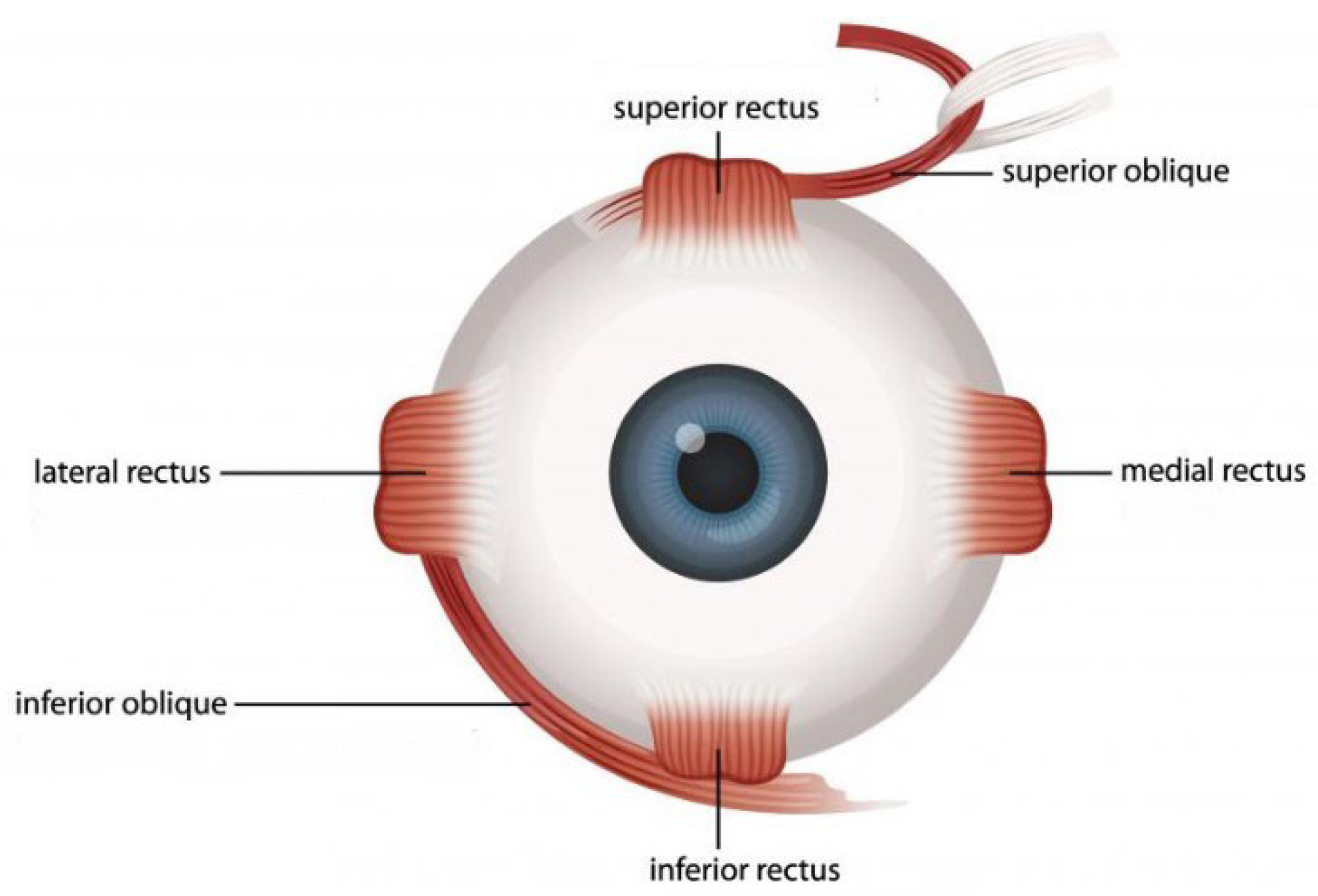
Figure 2.
The division of the EOM into two layers: the "orbital" layer and the "global" layer.
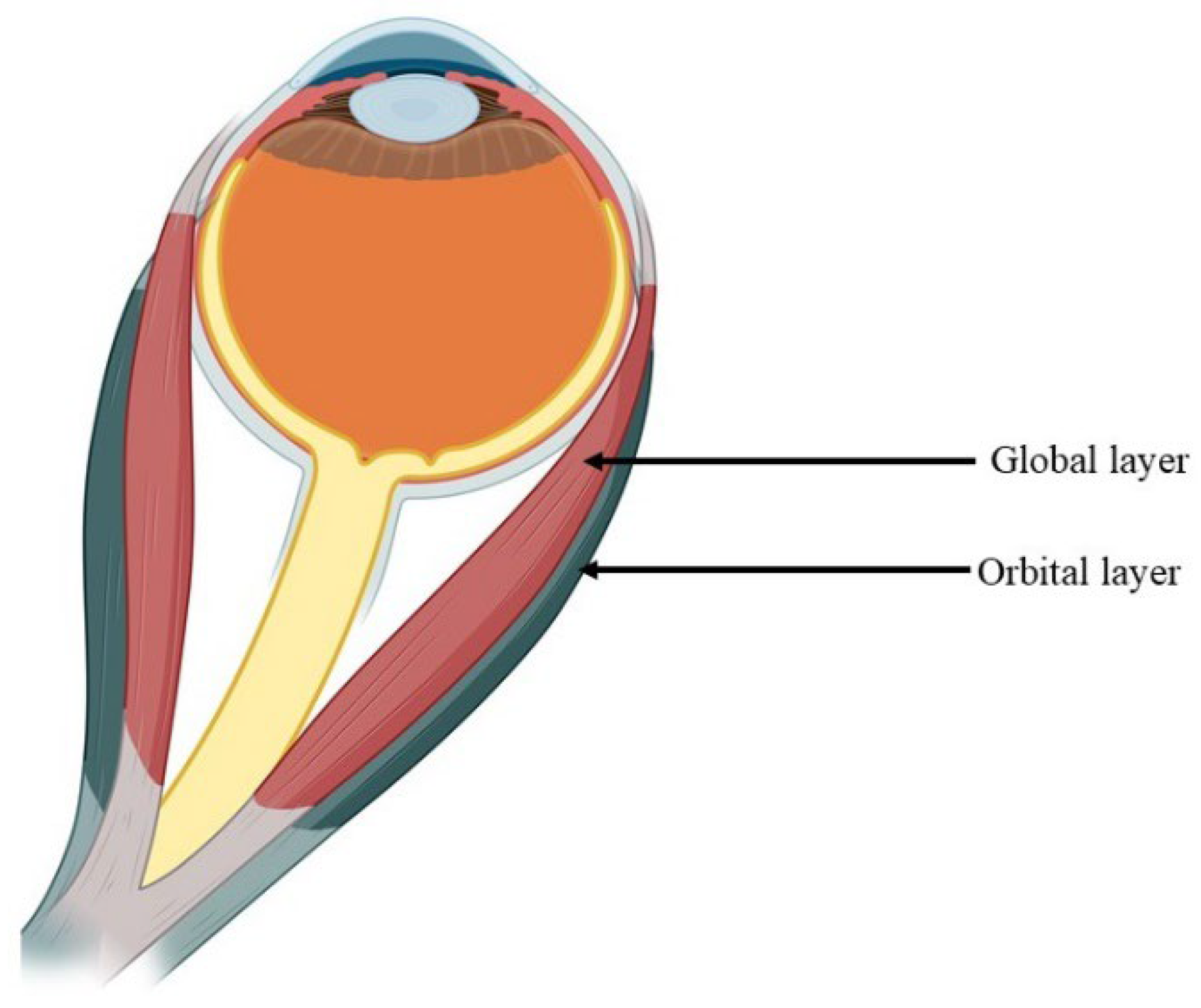
Figure 3.
Elimination of polyneuronal innervation of skeletal muscles.
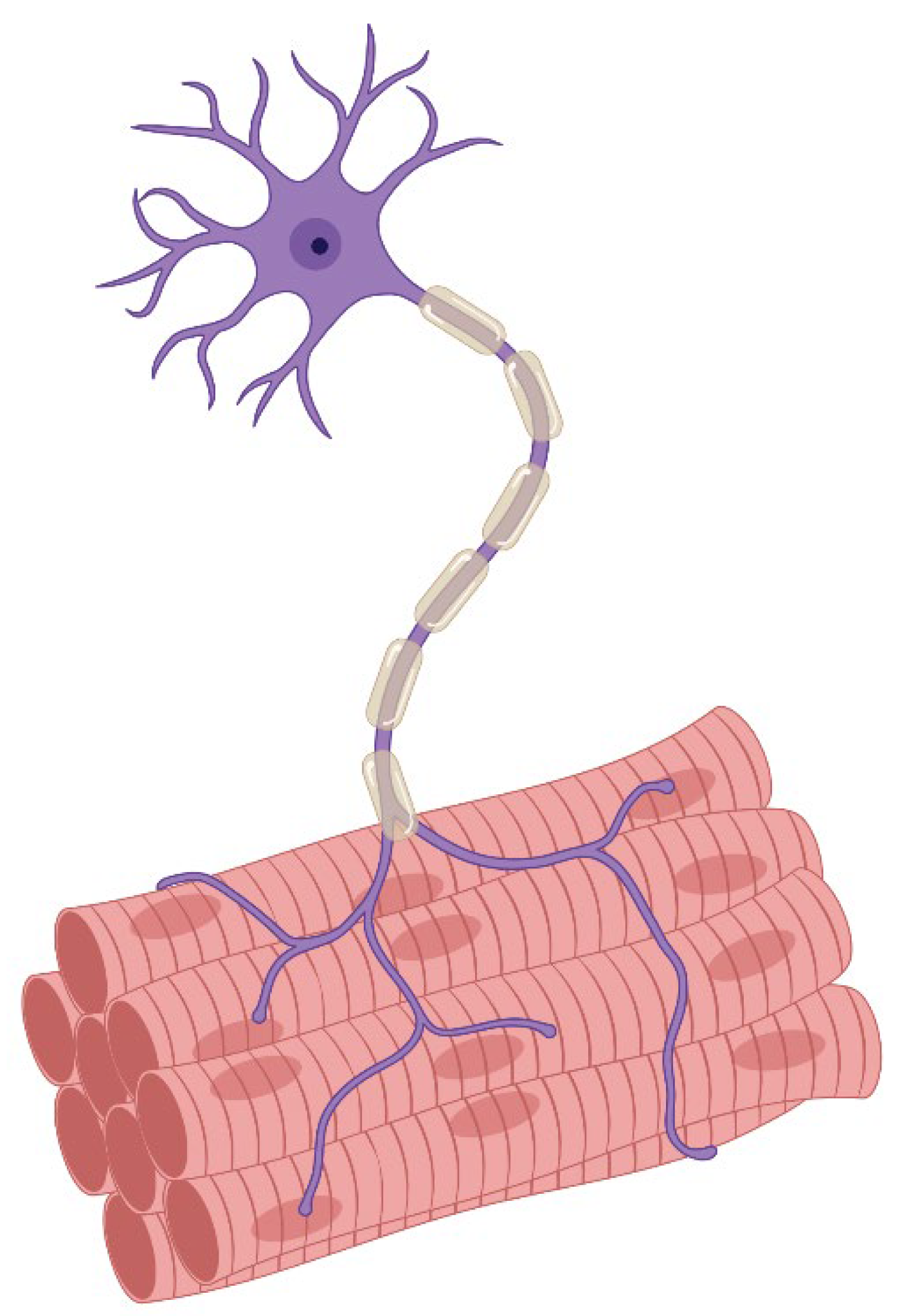
Figure 4.
Schematic morphology of the innervation of the extraocular muscles. III — oculomotor nucleus; IIIn, third cranial nerve. SIF – single innervation fiber, MIF – multiple innervation fibers.
Figure 4.
Schematic morphology of the innervation of the extraocular muscles. III — oculomotor nucleus; IIIn, third cranial nerve. SIF – single innervation fiber, MIF – multiple innervation fibers.
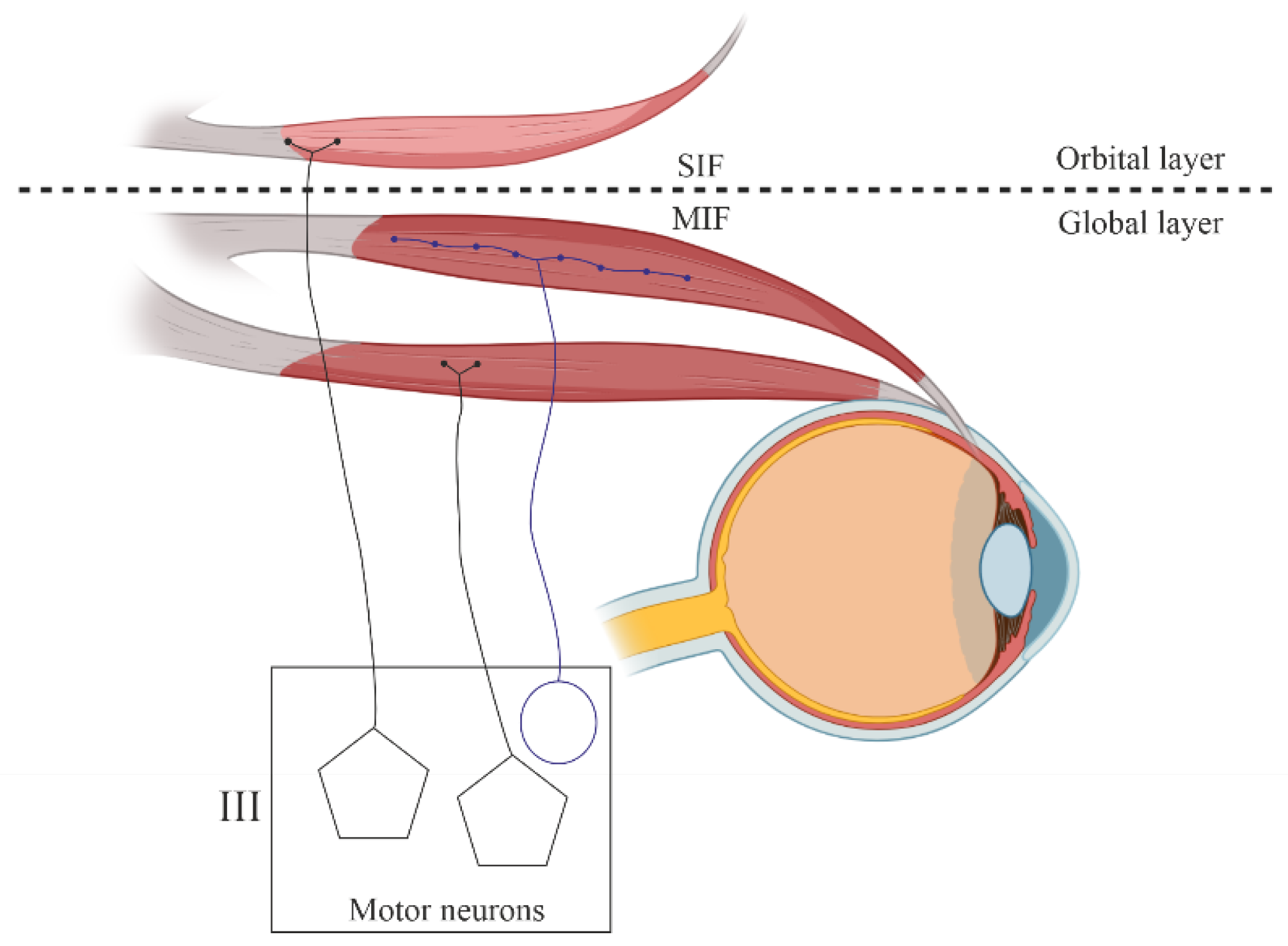
Figure 5.
Schematic representation of various extraocular muscle fibers types contraction mechanisms (SIF — singly innervated fiber; MIF — multiply innervated fiber).
Figure 5.
Schematic representation of various extraocular muscle fibers types contraction mechanisms (SIF — singly innervated fiber; MIF — multiply innervated fiber).
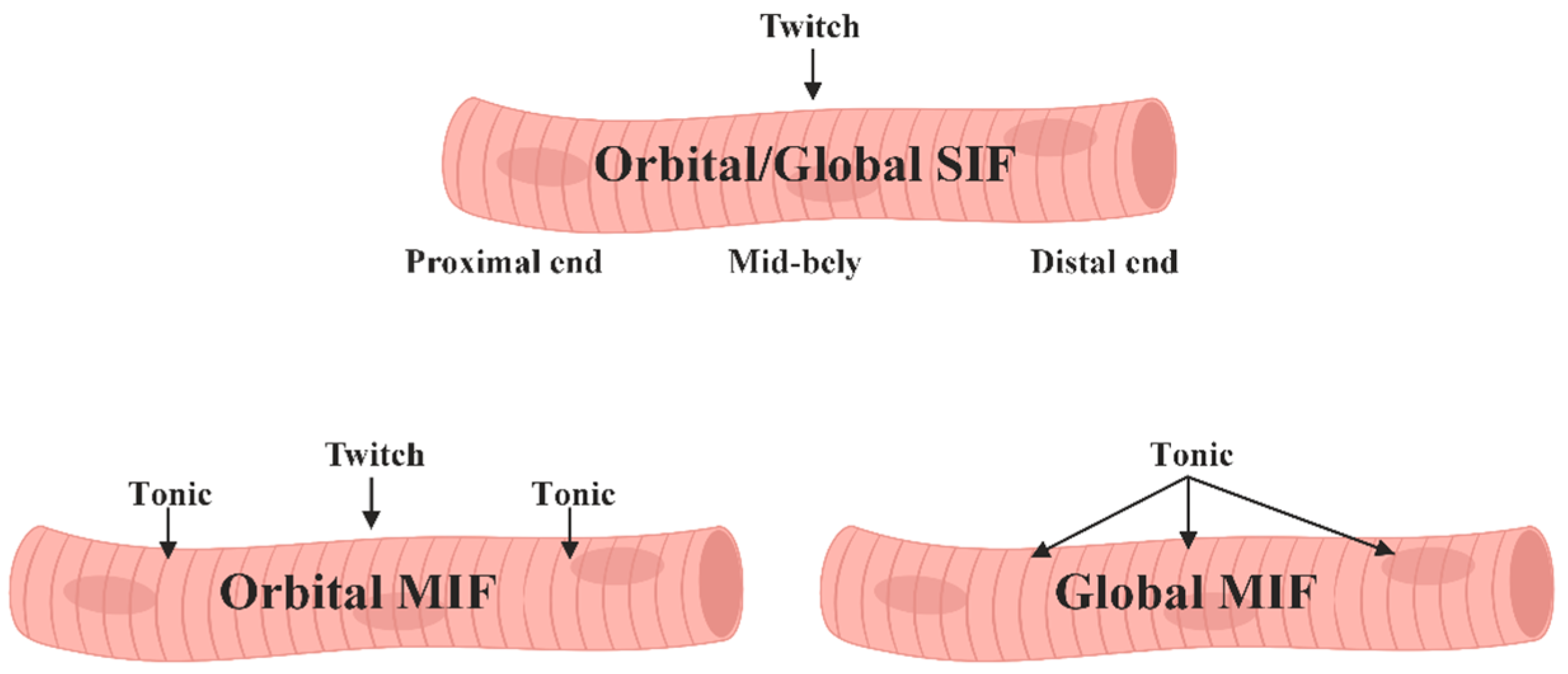
Figure 6.
Schematic illustrations depict the desmin composition patterns in three types of human EOM fibers. (A) demonstrates (A1) uniform desmin staining; (A2) absence of desmin in a confined area; or (A3) absence in a larger area proximal to the NMJ, with uniform staining in the remaining muscle fiber. (B) Desmin was typically present in MyHCsto/I fibers (B1), but absent in a subset of these fibers (B2). (C) Desmin was consistently found in myHCsto/I myofibers with spiral nerve endings. (D) Desmin was present (D1) or absent (D2) in muscle fibers containing MyHCeom.
Figure 6.
Schematic illustrations depict the desmin composition patterns in three types of human EOM fibers. (A) demonstrates (A1) uniform desmin staining; (A2) absence of desmin in a confined area; or (A3) absence in a larger area proximal to the NMJ, with uniform staining in the remaining muscle fiber. (B) Desmin was typically present in MyHCsto/I fibers (B1), but absent in a subset of these fibers (B2). (C) Desmin was consistently found in myHCsto/I myofibers with spiral nerve endings. (D) Desmin was present (D1) or absent (D2) in muscle fibers containing MyHCeom.
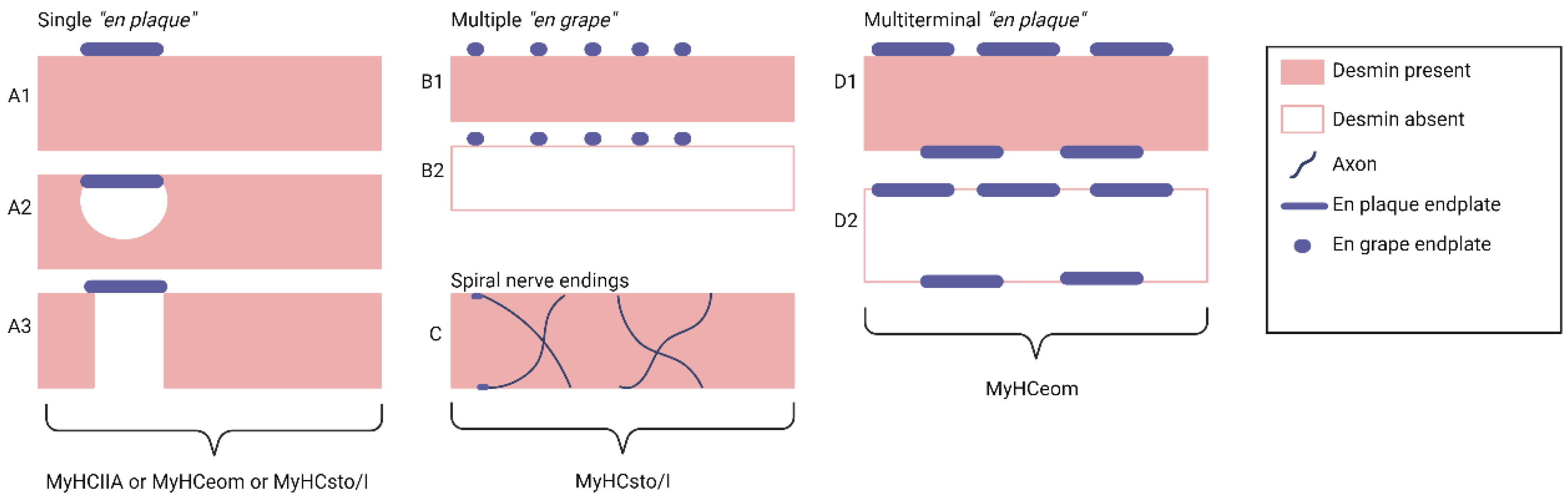

Table 1.
MyHC Isoforms in EOM.
| Type of isoform | Name |
|---|---|
| Fast isoforms of MyHC | MyHCIIa (Myh2); MyHCIIb (Myh4); MyHCIIx (Myh1); MyHCeom (Myh13). |
| Slow isoforms of MyHC | МyHCI-MyHC-B/slow (Myh7); MyHCα-cardiac (Myh6); MyHCsto (MyH7B) |
| MyHC isoforms related to development | MyHCemb (Myh3); MyHCneonatal (Myh8) |
| The ancient sarcomeric MyHC | MYH14/7b (MyH7B), MyH15 (Myh15) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



