
Submitted:
25 April 2024
Posted:
25 April 2024
You are already at the latest version
Abstract
Currently, the textile industry is among the most rapidly expanding areas of the economy and is an important source of water pollution. There are many efficient chemical and physical methods for treating textile effluent, but they produce secondary pollutants. Therefore, there is a need to manage the textile waste wastewater. The potential EPS-producing and plant growth-promoting rhizobacteria Lysinibacillus capsici PB300 (T) bacteria strain has been isolated from the bark borer-affected Peltophorum pterocarpum plant and has been determined to be effective in > 95 decolorizing Reactive Red 120, and other azo dyes such as AB11385%, orange II 94.62%, Congo red 94.62%, phenol red 94.54%, and mixtures of azo dyes 81.66%. Physico-chemical factors were optimized manually, including Taguchi design. EPS production, PGPR strain, and fabric discoloration of L. capsici were qualitatively studied. The FTIR, GC-MS, and UV absorbance studies also confirmed that the dye had been broken into its amines. Research findings using enzyme assays have shown that the bacteria Lysinibacillus capsici PB300 (T) can utilize laccase and manganese peroxidase and are capable of degrading dyes significantly. According to this work, immobilised L. capsici cells and the studied four bacterial consortiums, Lysinibacillus capsici PB300 (T), Alcaligenes faecalis subsp. Phenolicus DSM 16503 (T), Acinetobacterbaumanni ATCC 19606 (T), and Pseudomonasaeruginosa JCM 5962 may be employed to degrade RR120 effectively, and it is concluded that L. capsici PB300 (T) bacteria is significantly efficient in textile effluent treatment.
Keywords:
RR120
; dye degredation
; biodegradation
; bacterial consortia
; metal-ion tolerant
; immobilized cell
1. Introduction
Colour helps make the world more beautiful and colourful [1]. Dye is an organic chemical substance that is bright, ionizing, and aromatic. Synthetic dyes cause serious environmental pollution. In the textile sector, mostly azo dyes are used to dye clothes, and due to their capacity for oxidative breakdown, synthetic dyes persist in the environment for a long duration. Even in low quantities, the manufacturing of synthetic dyes has the potential to be dangerous, carcinogenic, and mutagenic. Common uses for dyes are in paper, textiles, leather, rubber, plastic cosmetic industries, tannery, printing and dyeing, and food. Humans and nature have been exposed to synthetic toxic dyes for a few million years. Every year, an estimated 700,000 tons of synthetic dyes are produced in about 100,000 types [2]. Azo dyes are the most commonly used category of dyes. Azo dyes are mainly utilized in the textile field [3]. Azo bond (–N = N-) is present in azo dyes and is related to the class of heterocyclic aromatic compounds [4]. Recently, more than 2000 azo dyes have been used in various industries, among which the textile colouration industry is the largest.
According to a previous report, only 10% of dyes are bound in dye applied to fabrics, and 90% of dyes are released into the water resources, which is the root cause of various skin problems [5,6]. Water pollution adversely affects aquatic life and ecological balance, increases chemical and biological oxygen demand (BOD), and alters the pH and chemical composition of the water body are just a few of the detrimental effects of releasing textile effluent into the aqueous ecosystem [6,7]. The entry of dye-contaminated wastewater mixes into the soil and harms the soil nutrients and microbial communities [1,8]. Present in the soil which is directly and indirectly associated with soil health and crop productivity. Besides that the synthetic dyes decrease dissolved oxygen (DO) levels, leading to many harmful impacts on marine flora and fauna. Sometimesthe compounds formed during aerobic degradation of azo dyes are more toxic than the parent azo dyes. These are usually recognised as aromatic amines. As a result, their degradation requires an additional aerobic stage. It has been observed that oxidative enzymes function better under aerobic conditions, and allow potential decolourization and additional detoxification of produced amines, whereas reductive enzymes break down the azo link of azo dyes under aerobic conditions. Consequently, azo dye degradation by successive aerobic processes emerged as a sustainable approach for textile effluent treatment [9].
Different microorganisms in bioreactors are used for the decolourization of textile dyes [10]. The cheapest and most environmentally beneficial technique for managing wastewater-containing dyes is microbial degradation. The bacterial aerobic degradation pathway disturbs the aromatic ring of azo dyes by monooxygenase or dioxygenase, which is then further degraded and mineralized [11]. According to Dixit and Garg [12], Klebsiella pneumoniae bacteria have the potential for toxic azo dye degradation. Previous studies showed that methyl red and MG can be degraded by Klebsiella pneumoniae strain WA-1 [12]. Recently, it has been reported that different Bacillus species have capabilities for textile wastewater decolourization. B. Cohnii RAPT1 significantly decolorizes Reactive Red 120 [13]. But till now, a very small number of aerobic and anaerobic bacterial species have been reported to degrade azo dyes.
Wastewater from textile dyes usually contains a large number of heavy metals. Human beings, plants, and microorganisms are all extremely hazardous to a variety of heavy metals. Among several ions of heavy metals and hexavalent [Cr(VI)] commonly present with azo dyes in effluents of the leather and textile industries [14]. Shewanell decolourization is caused by MBTD16, Paenibacillus glucanlyticus SLM1, and Bacillus amyloliquefaciens, which are examples of ligninolytic bacteria that remove dyes from textile wastewater. These microorganisms represent oxidative enzymes such as lignin peroxidases and laccases. Oxidative enzymes such as laccases, tyrosinases, veratryl alcohol oxidases, peroxidases, and azoreductases biodegrade dyes in wastewater. The mixed bacterial cultures decolourize dye molecules faster and more completely than single cultures do because they include several enzymes that attack the dye molecule at many sites [15].
2. Materials and Methods
2.1. Sample Collection and Bacterial Culture
Standard sample Reactive red 120 (RR120) dye was procured from Sigma Aldrich. All the chemicals, salts, acids, buffers, sugars (carbon source) and nitrogen source were procured from Himedia. Bacteria were isolated from the infected area of the bark borer disease of Peltophorum pterocarpumplant grown at Guru Ghasidas Vishwavidyalaya Bilaspur, Chhattisgarh, India.
To study the bio-decolourization and biodegradation of RR120 dye, the Bushnell Hass medium (BHM) was prepared as the following composition of BHM broth: (gl-1) K2HPO4 1.0, MgSO4 0.2, FeCl3 0.05, CaCl2 0.02, NH4NO3 1.0, supplemented with yeast extract (0.2% w/v), glucose (0.4% w/v), and 1% RR120 dye. To isolate microorganisms that decolourize dyes, 1g of the sample was placed in tubes, and 3 g of the sample was placed in a flask individually into reactive red 120 dye containing BHM broth and incubated for 3-5 days under static and shaking conditions at 35oC. Three consecutive transfers were made in a fresh dye-containing medium until the medium became decolourized. The decolourized samples were serially diluted (10-fold) and 100 µl aliquots of 10-5 to 10-7 dilution were spread and poured on Bushnell Haas agar medium (BHM) agar plates with 100 mg l-1 Reactive Red dye and subsequently incubated for 24-48 hours at 35⸰C. After incubation, the colonies emerged and were repeatedly streaked to purify them. Colony and culture morphology were used as the basis for the primary screening of the isolated bacteria. In secondary screening, the culture was incubated in BHM broth at 35 oC for 24 hours. 1 ml of bacterial suspension was taken from the incubated medium, then it was transferred to 30 ml BHM broth media with dye, incubated in a static condition at 35 oC for 72 hours, and absorbance was measured at every 12-hour interval. The culture having > 95% decolourization with RR120 dye in 12 hours was finally 4 bacterial strains: WR-1, WR-2 (isolated from bark borer sample), TS-1, and TS-3 (isolated from termite mound soil sample) selected. The secondary and final screening of selected cultures from primary screening. The process was carried out using the fast and high degradation efficiency of RR120 dye.
2.2. Identification of RR120 Dye Decolorizing Bacteria
Genomic DNA of 4 bacterial isolates WR-1, WR-2 (isolated from bark borer sample), TS-1, and TS-3 (isolated from termite mound soil sample) were extracted using phenol/chloroform extraction method. After that, the 16S rRNA gene was amplified by PCR using the universal primers 16F27 [5'-CCA GAG TTT GAT CMT GGC TCA G-3'] and 16R1492 [5'-TAC GGY TAC CTT GTT ACG ACT T-3']. The 16S rRNA gene PCR product was amplified and purified using PEG-NaCl precipitation and then sequenced immediately on an ABI® 3730XL automated DNA sequencer (Applied Biosystems, Inc., Foster City, CA) at NCMR sequencing laboratory of the National Centre for Cell Science in Pune, Maharashtra, India according to the manufacturer's instructions. The Laser gene package was used for assembly, while the Ezbiocloud database was used for identification. The neighbor-joining tree method was used in the BLAST and MEGA 11 software to generate a phylogenetic tree (Figure 1).
2.3. Decolourisation Studies
Four identified bacterial isolates TS-1, BOS-24, WR-1, and WR-2 were individually inoculated in BHM broth (gl-1) supplemented with yeast extract (0.2% w/v), glucose (0.4% w/v), and 1% RR120 dye, further culture tubes were kept in the static condition at 37 oC for 72hours incubation for decolourization. A similar situation was applied to the control sample (with dye but without inoculation) and the blank sample (without dye and inoculation). To study the effects of aeration inoculum size and incubation time 5 ml aliquots of the decolourized and control samples were drawn at regular 12 hours intervals and the pallets were removed by centrifugation at 4oC for 10 minutes at 10000 rpm. The RR120 decolorization was studied using the supernatant it was determined using a UV-VS spectrophotometer absorbance at 524 nm. Decolorization % was determined using the formula given below [16].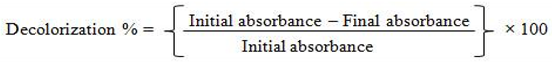
2.4. Aeration, Inoculum Size and Time for Dye Decolorization
Several physical and chemical variables directly impact the decolourization ability of microorganisms during biodegradation processes [7]. Therefore to improve the rate of dye decolorization, decrease the time required for decolorization, and increase the process's affordability various physicochemical parameters were optimized.
Fresh bacterial cultures were cultivated aerobically in BHM broth media and it was incubated at 35-37 oC for 24 hours under anaerobic conditions. 1ml bacterial inoculums were cultivated in 30ml screw-cap tubes having BHM broth (30ml) supplemented with glucose (0.4% w/v), 1% RR120 dye and yeast extract (0.2% w/v), and were incubated at 35-37 oC at 72 hours under shaking (aerobic)condition. 1 ml culture was added in a 100 ml Erlenmeyer flask containing 40 ml BHM broth and incubated at 72 hours in an incubator shaker at 120 rpm at 35-37oC. The absorbance of the culture-inoculated sample and control was measured every 12 hours. The decolourization ability of the WR-1 strain was tested in shaking and static culture at optimum pH 7.0. The cell growth was at 600 nm using the optical density of the culture[16,17].The impact of different inoculum percentages; 1%(0.3ml), 2%(0.6ml), 4%(1.5ml), 6%(1.6ml), 8% (2.4ml), and 10%(3ml) for decolorization of RR 120 dye was studied under anoxic static conditions at 35 ⸰C. The various concentrations of inoculum were added into the 30ml screw cap tubes BHM broth (30ml) media was added with yeast extract (0.2% w/v) glucose (0.4% w/v), and 1% RR120 dye. The effect of various incubation times on the decolourization of Reactive Red 120 in anaerobic static condition at 35 oC, pH 7. 1ml inoculum was added into the BHM medium supplemented with glucose (0.4% w/v), yeast extract (0.2% w/v) and 1% RR120 dye and incubated at 35 oC for 72 hours h under static condition.
2.4.1. Optimization by Taguchi Method
The Taguchi approach for optimization of the pH, temperature, carbon source, nitrogen source, and dye concentration parameters was followed according to Pourbabaee et al., [18]. The factors were optimised using the Taguchi method. The criteria were selected according to their effect on the biodegradation of the RR120 dye. The experiments were designed using the following five variables. Glucose, fructose, sucrose, lactosestarchcarbon and yeast, peptone, ammonium sulphate, ammonium nitrate and urea, and nitrogen sources were added to the BHM culture media. The BHM media with different ranges of pH 4, 5, 6, 7, and 8 and different incubation temperatures of 25-45⸰C. were used. Dye concentrations 0.1, 0.2, 0.3, 0.4 and 0.5 mg/l were added to the BHM culture media [18]. The experiments were set up using the L25 orthogonal array after the factors in the mixed-level design were selected. Minitab 19 statistical software was used for the analysis of data [19].
2.5. Development of Consortium
Compatibility test of Lysinibacillus capsici PB300(T), Alcaligenes faecalis subsp. Phenolicus DSM 16503(T), Acinetobacter baumanni ATCC 19606 (T) and pseudomonas aeruginosa JCM 5962 (T) were observed using the agar well diffusion method. All four bacterial cultures were individually cultivated aerobically in BHM broth, and incubated at 35-37 ⸰C for 24 h under static conditions. These bacterial cultures were developed by mixing in 10 combinations. These cultures were cultivated anaerobically in 30ml BHM broth with 3% RR120 dye-containing medium inoculated according to culture combination and incubated for 12 h at 35-37⸰C under static conditions [20]. The decolorized individually inoculated, consortium sample and control sample (each 3ml) were regularly collected at 2-hour intervals and centrifuged at 4⸰C for 10 minutes at 10000 rpm. The decolourization percentage of RR120 dye was analyzed using a UV-spectrophotometer by calculating the absorbance of the supernatant at 524 nm. Finally, a comparative decolourization percentageby Lysinibacillus capsici, Alcaligenes faecalis sub sp. Phenolicus, Acinetobacter baumanni and Pseudomonas aeruginosa bacterial monocultures and consortium were recorded. The bacterial combination, that showed the highest dye degradation percentage in minimum time, was used as the best combination for further analysis.
2.6. Immobilization of Dye-Degrading Bacteria
The Lysinibacillus capsici PB300 (T) was inoculated in an LB liquid medium and incubated for 24 hours at 35⸰C. After incubation, 5ml media from well-grown bacterial culture media was used as a bacteria source. Sodium alginate (SA) was used as a bio-carrier in this study. The stability of SA in water is very low, due to this problem well well-known synthetic polymer PVA was used in immobilization [21]. The sterile 3% SA and 3% polyvinyl alcohol (PVA) were added. Alginate-alcohol mixture 5ml solution was added to 5 ml bacterial solution and this SA-PVA-bacteria mixed solution was dropped drop by drop using a syringe, into a cold 3% CaCl2 solution. The beads-CaCl2solution was kept in the shaker for 1 hour then it was kept in a freezer at 4⸰C for 24 hours. Beads (0.5mm sized 10 beads) were used for dye decolourization, but first, beads were washed using double distilled water after 24h [22]. The immobilized bacterial beads were several times washed using a phosphate buffer (pH7) then it was added in30ml BHM dye-containing broth media and incubated for 12 hours at 35⸰C.at static condition. Cell-free immobilized beads were added to the control (BHM broth with dye) and blank sample (BHM broth without dye) and incubated under the same conditions. After 12 hours of incubation, the decolourized sample absorbance was measured and the beads were collected. The collected beads were washed with phosphate buffer 3 times, placed in fresh BHM broth media and incubated in the same conditions. The same procedures were repeated 15 times.
2.7. Decolourization of Various Azo Dyes and Mixture of Dyes
The above optimized optimal parameters were used for decolourization of various azo dye viz. RR120, AB113, Orange II, CR, PR and mixed azo dyes. These dyes were mixed at a concentration of 20 mgl-1 of each in the BHM broth medium. The BHM broth was transferred in 30 ml tubes and bacterial cultures were inoculated and then incubated under the above-optimized conditions. After incubation, the decolourization percentage was analysed UV-spectrophotometer by calculating the absorbance of the supernatant at λmax of respective azo dyes and a mixture of dyes.
2.8. Qualitative Estimation of EPS Production by Dye-Degrading Bacteria
The MRS broth culture medium was used to measure the EPS generation of particular bacterial strains that were used in dye decolourization. Selected strains were cultured in MRS broth and incubated for 10 days at 40 ⸰C.
2.9. Dye Decolorization in the Presence of Metal Ions
Decolorizationof RR120 (50 mg/L) and the heavy metals Cu, Pd, Zn, Ni, and Cd (1 mg/l to 6 mg/l) simultaneously under 6 g/L NaCl conditions was examined. Five heavy metals were extracted from the chemical compounds that were the cause of the bacteria's ability to tolerate heavy metals. NaCl was added to a binary mixture including the heavy metals Cu, Pd, Zn, Ni, and Cd. WR1 bacterium strain was inoculated into 30ml of BHM broth media after it had been autoclaved, and various doses of heavy metals (ranging from 1 mg/l to 6 mg/l) and dye had been added separately. The tubes that were inoculated and those that were not incubated for a whole day at 35⸰C under static circumstances. The absorbance of the test and control media samples was measured after incubation to determine the percentage of dye decolourization when heavy metals were present at varying concentrations. Using the previously mentioned technique, the azo dye's decolourization efficacy was determined. Every experiment was conducted in triplicate.
2.10. Decolorization of Textile Fabric
Commercial 100 % cotton 2 set (red and blue fabric) fabric that was purchased from the local textile market market Bilaspur, Chhattisgarh India were cut into small equal sizes. The fabric pieces were packed well and sterilized in an autoclave. Sterilized red and blue fabric pieces were placed in separate tubes in sterilized BHM broth media under aseptic conditions. 1% inoculum of bacteria culture was inoculated in test sample tubes and control media was without culture. All tubes were incubated at 35-37⸰C in anaerobic and static conditions for 5 days. The fabric was extracted from every tube and washed with phosphate buffer 3 times and the fabric was dried normally at room temperature [23].
2.11. Determination of Plant Growth-PromotingTraits
Through the qualitative evaluation of ammonium production and indole acetic acid (IAA), two recognized features, plant growth-promoting traits were investigated.
2.11.1. Qualitative Assessment of IAA Synthesis
The WR-1 bacterial isolate was qualitatively tested for the production of IAA, this bacteria was inoculated into nutrient broth containing 2mg/ml-1 L- tryptophan and it was incubated at 30⸰C for 48 hours on shaking conditions at 125 rpm. 1 ml aliquots of culture media was centrifuged at 10000 rpm for 5 minutes at 4⸰C . 1 ml supernatant was mixed with 2 ml Salkowski’s reagent (150ml con.H2SO4, 250ml distilled water, 7.5ml 0.5M, FeCl3.6H2O) and further it was incubated under the dark condition at 30 ⸰C for 30 minutes. The IAA production was confirmed on the basis development of a pink-red colour [24].
2.11.2. Qualitative Assessment of Ammonium Production
For ammonium (NH3) production, the WR-1 bacteria isolate was grown in peptone broth at 4⸰C for 72 hours on shaking conditions at 125 rpm. After incubation, 10ml culture solution was centrifuged for 15 minutes at 4⸰C at 5000 rpm. 1 ml of Nesseler's reagent was mixed with 9 ml supernatant. The ammonium production was confirmed based on the development of a brown-to-yellow colour [25].
2.12. Enzyme Assay
Bacterial cultures were in LB broth at 24 hours, and after growth culture solution was centrifuged for 10 minutes at 4⸰C at 10,000 rpm. Three times washes were done on the pellets and suspended in 5ml 50 mM potassium phosphate buffer pH 7.0 and the bacterial cells were broken by using a centrifuge for 5 min, 4⸰C at 10000×g. Cell-free supernatants (crude cell extract) were used as a reaction mixture for screening of laccase and manganese peroxidase [26]. Laccase enzyme 0.5 ml guiacol was added to the 0.5 ml reaction mixture as a substrate then incubated for 5 min and the laccase enzyme was screened based on colour change in control and test suspension. To test the presence of manganese peroxidase enzyme 0.5 ml of 50 mM sodium tartrate buffer(pH 5)and 0.5 ml of 20 mM guiacol to 0.5 ml reaction mixture and incubated for 5 minutes. Finally, 0.5 ml of 2 mM H2O2 was added and the brownish-red colour changed. Manganese peroxidase enzyme was present and confirmed based on colour change in control and test suspension.
2.13. Decolorization and Biodegradation Analysis
The decolourization and biodegradation of RR120 were studied in static conditions using FTIR, UV–visible, and GC–MS. During decolourization, the azo bond of the chromophore conjugated in RR120 was degraded due to azo reductase activity and the peak gradually reduced and fully disappeared within 12 hours [27]. For this, 30 ml BHM broth medium with dye after decolourization sample was taken at 12 hours, centrifuged for 10 min, 4⸰C at 10000 × g. to remove pallets, after centrifuge and extraction of dye, metabolites were carried from the supernatant using an equal amount of ethyl acetate. The extracted metabolite was dried over anhydrous Na2SO4 in an oven. During UV-Vis spectral analysis, changes in the absorption spectrum in the decolourized medium (400–800 nm) were recorded in comparison with the decolourization percentage outcomes from the control sample. The biodegraded RR120 was evaluated and compared with the control dye using Fourier Transform Infrared Spectroscopy.
2.13.1. UV-Vis Spectroscopic Study
Decolourization was confirmed by UV-Vis spectroscopic study. UV-Vis analysis was carried out in dye decolourization conformation in which 5ml of incubated decolourized media, control media and blank media were taken in centrifuge tubes and centrifuged for 10min at 10000 rpm, 4⸰C. After the cell biomass was eliminated, the supernatant's absorbance in the centrifuged medium was measured at 524 nm [28].
2.13.2. Analysis of Dye Metabolites Using FTIR and GC-MS
Dried extracted dye metabolites were mixed with kbr in a ratio of 5:95. Mixed samples were pressed using a hydraulic presser to get transparent pellets. Pellets were used for obtaining the IR spectra using FTIR in regions between 4000 and 400 cm−1. The acquired sample spectra were compared between the range of dye metabolites treated by bacteria and the untreated dye control [29]. Spectral library software NIST1.10 (Shimadzu) was used for conformation and match molecular weight, retention times, molecular formula and peak area and height of the metabolites. GC-MS study of dye-degraded metabolites was carried out using a GCMS-TQ8050 NX (Shimadzu). The initial column temperature was 80 ⸰C for 2 min. hold, then increased linearly at 10 ⸰C per minute to 100 ⸰C and hold time at 2 min. The temperature of the injection port was 230⸰C and the GC-MS interface was maintained at 280 ⸰C. The injector temperature was continued maintained at280⸰C. The compounds were identified based on mass spectrum and by using the NIST library[30].
3. Result
3.1. Identification of RR120 Dye Decolorizing Bacteria
RR120 dye decolorizing potent bacterial strains were identified according to biochemical and morphological characteristics and 16S rRNA gene sequence. Four bacterial strains, including Lysinibacilluscapsici PB300 (T), Alcaligenesfaecalissubsp. Phenolicus DSM 16503 (T), Acinetobacterbaumanni ATCC 19606 (T) and Pseudomonasaeruginosa JCM 5962 (T) were identified using 16S rRNA gene sequence. L.Capsici, A.faecalissub sp. Phenolicus, strains isolated from bark borer samples and A.baumanni and P.aeruginosa were isolated from termite mound soil. L.capsici, A.faecalissubsp. Phenolicus (accession no 13436), A.baumanni (accession no 13438) and P. aeruginosa (accession no 13437) bacterial strains were deposited in Microbial Type Culture Collection and Gene Bank (MTCC), Chandigarh, Panjab India.
3.2. Decolourisation Studies
Among four bacterial isolates, Lysinibacillus capsici PB300 (T) showed the highest decolourization percentage (Figure 2). It showed >95 decolourization at 12hours with a 100 mg L−1 dye concentration, at pH 7-8, and temp. 35-37⸰C, in static conditions.
3.3. Optimization of Various Parameters for Decolourizationo of Dye RR120
3.3.1. Effects of Aeration, Incubation Time and Inoculum Size
In the anoxic static and shaking condition, between 12-72 hours, 90% and 83.33%decolourizationand cell growth were observed, but in the shaking condition, cell growth (Figure 3a) was much greater than in the static condition but was less decolourization (less than 80%).In the growth stage of the cell, oxygen has a major impact on its physiological properties. Oxygen can affect the rate at which microbes degrade azo dyes. The findings demonstrated that RR120 needed more than 12 hours in static conditions and 72 hours in shaking conditions to fully decolourize (Figure 3b). Incubation time has a significant impact on the growth and enzymatic activity of microorganisms, which can have an impact on their optimal activity decolourization performance. Following a 12-hour incubation period, a significant amount of decolourization was observed over the temperature range of 35–37 ⸰C. After getting the complete (>99%) decolourization result in 12hours, the effect of time in decolourization was repeated for 12 hours during which decolourization was observed every 2 hours. Figure 3b showed >95% decolourization between 12 to 72 hours. An increase in inoculum size was correlated with an increased rate of decolourization in a limited period. Over 95% of the decolourization occurred in 12 hours at a concentration of 1–10% inoculum, which yielded the highest decolourization. Figure 3c showed that inoculum size significantly affected decolourization. Observation showed that a 1% inoculum size was effective for full decolourization in 12 hours. A faster maximum decolourization was seen by increasing the inoculum size.
3.3.4. Taguchi Statistical Analysis
Taguchi orthogonal array is used to determine the best operating settings, having a beneficial impact on the best combination of several parameter sets (Table 1). Since the average mean response for each level of each factor was estimated first, the delta value for each control factor was also calculated. After that, estimates of the control factors were made using their delta values. In the Taguchi technique, the terms "noise" and "signal" stand for the desired and unwanted values, respectively, for the output characteristic. The Taguchi technique uses the S/N ratio to quantify the quality characteristic that is different from the desired value. According to the type of characteristic, the S/N ratios change. The factor's importance and the delta have a positive correlation with RR120 biodegradation. Hence, the results showed (Table 2) that pH (delta30.5, rank 1) has the highest level of biodegradation of RR120 dye, then dye concentration (delta 15.55, rank 2), temperature (delta 8.4, rank 3), carbon source (delta 7.95, rank 4) and nitrogen source(delta 5.41, rank 5). It was easy to find the best levels of the five control parameters now in levels because Taguchi is adaptable., which was then varied, and the mean response for each run in the Taguchi array was calculated. As a result, process variability was minimized by reducing the influence of noise or uncontrollable factors during operation. A high S/N ratio means that the control factors perform as efficiently as possible and that the noise factors have minimal impact. Hence, the results show (Table 2 S/NRatios) that ph (delta3.68, rank 1) has the highest level of biodegradation of RR120 dye, then dye concentration (delta 2.08, rank 2), temperature (delta1.15, rank 3), carbon source (delta1.08, rank 4) and nitrogen source (delta 0.81, rank 5). Additionally, the Taguchi technique found the best level for each of the 5 factors within the study, which changed accordingly. The biodegradation level of RR120 was determined to be 98.67% and run No. L18 in the Taguchi array was reported to have recovered [19]. Based on the findings presented in Figure 4 and Figure 5, the optimal effective parameters were pH 8, temperature of 40 oC, and dye concentration of 0.2 mg/l. ammonium nitrate as a source of nitrogen and glucose as a carbon source.
3.4. Dye Decolourization by Consortium
For decolourizationof dyes, the selected consortia consisted of four bacterial strains, Lysinibacillus capsici PB300(T), Alcaligenes faecalis subsp. Phenolicus DSM 16503(T), Acinetobacter baumanni ATCC 19606(T) and Pseudomonas aeruginosa JCM 5962(T). All four bacteria survived in the consortium. These four identified bacteria were selected individually which can degrade >95% of the RR120 dye at a pH of 7-8, 40⸰C in 12 hours of incubation. Selected bacteria strains were mixed in 10 combinations, and their degradation ability in the RR120 azo dye was studied. According to the consortium results observation, it was revealed that the complete degradation of dye by the consortium took less than 12 hours as complete to monoculture. The details of the developed consortiums and the result for degradation by a consortium of dye are given in Table 3. The RR120 dye bacterial degradation was confirmed by using UV-visible spectrum spectroscopy of samples inserted with bacterial consortia and untreated control samples. Methyl red and carbon fuchsin dyes showed decolourization percentages of 100% and 96%, respectively, indicating a greater decolourization capacity over their isolates in a consortium of three bacteria (Acinetobacter spp., Proteus spp., and Pseudomonas spp.,). pH 7 and 40 ⸰C conditions, a two-bacterial consortium (Galactomyces geotrichum and Brevibacillus laterosporus) suggested a decolorization percentage of 92% for Remazole Red 198 dye; in contrast, the individual bacteria only demonstrated 58 and 42% decolourization percentages, respectively [31].
3.5. Immobilization of Dye-Degrading Lysinibacillus Capsici PB300(T) Bacteria
The dye decolourization and reusability capacity of immobilized beads were analyzed using a repeated-batch reactor. SA-PVA immobilized bacterial cells showed >96% decolourization of RR 120aliquots (300mg/l) was achieved in 15 cycles in the 12h incubation time. After 15 cycles there was a 10% reduction in decolourization efficiency. As a result, the immobilised bacteria-mediator beads showed good reusability for decolourization of RR120.
3.6. Decolourization of Reactive Dye and Dye Mixtures
Since synthetic dyes with a variety of structures are frequently used in the textile processing industry, the composition of effluents from various industries varies significantly. The decolorization efficacy of various azo dyes used alone, and in combination with BHM broth medium was examined (Figure 6). According to our findings, Lysinibacillus capsici PB300 (T) decolorized azo dyes 95.15% RR120, 85% AB113, 94.62% Orange II, 94.62% CR, 94.54% PR and 81.66% mixed azo dyes at 90.2% after 96 hours under a static condition at 40oC, and pH 8.
3.7. EPS Production
After incubation, the culture media were centrifuged for 10 minutes at 4 ⸰C at 10,000 rpm. The supernatant was centrifuged again and the pellets were removed and the supernatant was centrifuged again. Cold ethanol was added to the supernatant and centrifuged for 5 minutes at 5000 rpm, 4⸰C. The centrifuged sample was kept at 4⸰C for 48 hours and EPS was precipitated in the form of white-brown pellets. From this test, it was determined that this bacteria strain has dye decolourization capacity and also produces EPS.
3.8. Effect of Heavy Mmetals Ions andNaCl on Decolourizationof RR120
By supplementing Cu, Pd, Zn, Ni, and Cd heavy metal concentrations (1 mg/l to 6 mg/l) and 5% NaCl in the decolourization assay medium, we were able to find out the effects of heavy metals ions on Lysinibacillus capsici PB300(T) bacterial decolourization and the WR1 strain's tolerance to heavy metals. The dye decolourization activity result (Figure 7) did not show the same results when all heavy metal ions were utilized. The presence of Cu, Pd, Zn, Ni and Cd heavy metals and the amount of their presence had a significant impact on RR120 dye decolourization, significant difference was found in decolourization %. Increasing concentrations of Cu, Pd, Zn, Ni and Cd heavy metals showed a significant decline in the decolourization. Only a significant amount of considerable level of decolourization was enhanced with Cu2+. When 1, 5 and 10 mm Cu2+ was implemented, decolourization increased growth by 39%, 33% and 6.5%, consequently, over the control. According to this finding, the decolourization efficiency reduces as the heavy metals Cu, Ni, Pd, Zn, and Cd concentrations increase. Previous works have also reported on the decolourization of textile dyes when heavy metal ions are present. When compared to other heavy metals used, the rate of decolourization was reduced when Cd was present. When Zn metal ions various (1mg/L-6mg/L) concentration was 74%, 59%, 56.01%, 41.96%, 36.55% and 29.91% decolourization within 12hours.
3.9. Decolourization of Textile Fabric
To assess the effectiveness of bacteria in coloured fabric, we used red and blue coloured cotton fabric. The fabric pieces were dipped in bacteria-inoculated and uninoculated broth media and the activity of coloured fabric towards dye was analyzed. The change in colour of the fabric was completely observed within the incubation days, resulting in complete fading of the colour of the fabric as shown in Figure 8. The fabric of the infected tubes was significantly lighter in colour than the fabric of the uninoculated tubes after washing and drying. From this study, being in the presence of dye-degrading bacterial enzymes and their enzymatic activity was determined. As per the observation of the result, these bacteria caused the red-coloured fabric to fade completely, whereas the blue-coloured fabric could not fade completely under the same conditions.
3.10. Qualitative Estimation of Plant Growth-Promoting IAA and Ammonium Production
Biosynthesis of plant growth promoting IAA hormone and ammonium was found to be positive in WR-1 isolates. According to qualitative estimation, the formation of indole acetic acid occurred when L-tryptophane was present [25].
3.12. Enzyme Assay
Laccase and manganese peroxidase enzymes were screened during the dye decolourization test. Laccase and manganese peroxidase enzyme screening tests finally showed reddish brown colour and brownish red colour in the test sample. Lysinibacillus capsici PB300 (T) bacterial strain showed positive activity in laccase and manganese peroxidase enzymes. These enzymes convert toxins into less hazardous or non-toxic metabolites, which is beneficial for detoxifying several kinds of industrial substances. In the decolourization experiment, laccase was found to be the protein that is most prevalent and least frequently produced. Our research showed that enzymes were essential in promoting the dye's decolourization [32].
3.13. Decolorization and Biodegradation Analysis
3.13.1. UV-Vis Spectroscopic Analysis
For dye decolourization and degradation analysis, control and test samples were centrifuged every 2h and the UV-Vis absorption of the supernatant was measured. RR120 dye absorbance optical density decreased in the UV-Vis absorption spectrum (524 nm) and was confirmed for decolourization compared to the control samples. The calculated OD and time graph (Figure 9) confirmed that, in the test sample compared to the control, the OD of the samples decreased with increasing time. The peak at 524 nm, which represents the absorbance optical density of RR120 peaks, decreased in comparison to the starting absorbance as the biodegradation was finished (12 hours)[16].
3.13.2. FTIR Analysis of Dye RR120 Metabolites
Essential bands such as aromatic rings, C-N stretching, and the azo chromophore (-N=N-) are considered when evaluating dye degradation. The decolourized compounds' FTIR spectra revealed significant differences in peak positions when compared to the control dye spectrum. The dyes underwent biodegradation as shown by the variation in their FTIR spectra and those of their metabolites obtained during decolorization. The results are displayed in Figure 10a,b. After combining the samples with spectroscopically pure kbr, analyses were performed. FT-IR spectra of the compounds extracted with RR120 azo dye showed a peak for O-H bending at 1381.38 cm-1, along with a supporting medium peak for the C-N stretching group and amine class at 1130.61 cm-1, and another medium peak for the C-N stretching functional group and amine class at 1028.09 cm-1, which indicates the formation of amines[18]. An indication of the presence of the O-H stretching functional group, class alcohol, is the broad and strong peak at 3260.93 cm-1. The strong peaks at 1553.48 cm-1 showed the N-O stretching and nitro class compound. However, other medium peaks at 1397.26 cm-1 and 1078.77 cm-1 confirmed the presence of functional groups O-H bending, class carboxylic acid, C-N stretching group, and amine class compounds, respectively.
3.13.3. GC-MS Analysis of Metabolites Produced by Lysinibacillus capsici PB300(T)
In RR120 dye was treated with L. capsici, and ten peaks were found during GC-MS analysis (Figure 11). Degraded metabolites were identified using NIST1 [33]. 10 library: Butanoic acid (retention time 3.433 min, molecular mass 102 g/mol, area % 49.38), Pentanoic acid(molecular mass 116 g/mol, retention time 4.953 min., area % 3.22), Pentanoic acid(molecular mass 116 g/mol, area % 0.90, retention time 5.345 min.), 2-Piperidinone (molecular mass 99 g/mol, area % 26.72, retention time 13.527 min.), Tridecanoic acid (molecular mass 242 g/mol, area % 0.37, retention time 23.945 min.), 1,4-diazabicyclo[4.3.0] nonan-2,5-dione (molecular mass 168 g/mol, area % 2.27 retention time 24.362 min.,), Cyclo (L-prolyl-L-valine) (molecular mass 196 g/mol, area % 8.77, retention time 25.498 min.,), Pyrrolo[1,2-a]pyrazine-1,4-dione(molecular mass 210 g/mol, area % 1.18, retention time 26.020 min.), L-Leucine(molecular mass 409 g/mol, area % 3.18, retention time 27.036 min.) and Pyrrolo[1,2-a]pyrazine-1,4-dione(molecular mass 210 g/mol, area % 4.01, retention time 27.356 min.) as represented complete details in Table 4.The Pichia kudriavzevii P1 halotolerant yeast can degrade the azo dye acid red B to a percentage of 94.29% at 100 mg/L.
4. Discussion
4.1. Isolation of Dye Decolorizing BacterialStrains
We found that three isolates showed significant RR120 decolourization with efficiency higher than 90%, even if the bacteria which were isolated from the Indarbela tetraonis, bark borer collected from Guru Ghasidas Vishwavidyalaya in Bilaspur, Chhattisgarh, India. In the present study, WR-1 strains showed 99.9% decolourization in the BHM containing carbon: glucose, nitrogen source- ammonium nitrate, 0.2mg/l RR120, at 28 ◦C, pH 7, and after 12 hours of incubation, respectively, under anaerobic static conditions. Likewise, a few PGPR bacteria also remove the colour of azo dye, including Rhizobium meliloti NCIM 2757, Bacillus sp. SR 2-1/1, B. megaterium NCIM 2054, Pseudomonas japonica I-15, and Azotobacter vinelandii MTCC 1241[34]. In the MS media containing 150 mg L−1 of the respective dye at 30oC, pH 6, and 48 hours of incubation, Bacillus sp. MR-1/2, a PGPR strain isolated from the root zone of maize crop, was shown to decolourize 78, 83, 63, and 65% of Reactive black 5, Reactive red 120, Direct blue 1, and Congo red, respectively, under microaerophilic conditions [35]. Moreover, it was found that Bacillus sp. SR-2-1/1, isolated from the root zone of sorghum plants grown in soil contaminated with textile wastewater, could remove 83, 40, 71, and 55% of Congo red, Reactive red 120, Reactive black 5, and Direct blue 1, respectively, from the MS media after 72 hours of incubation in static conditions [36].
4.2. Effects of Various Parameters
In the present investigation isolated bacteria L. capsici strain was able to decolorize the RR120 from BHM in an anaerobic environment. In the presence or absence of oxygen, dyes are susceptible to biological degradation. While the first step of azo-linkage cleavage primarily involves the breakdown of azo dyes in anaerobic conditions, bacteria may mainly destroy the aromatic amines generated in the second step in an aerobic environment. As a result, combining the anaerobic and aerobic phases of a treatment process may accelerate and enhance it [37]. Under static conditions, Kocuria rosea MTCC 1532, a Gram-positive bacterium, has been reported to totally (100%) decolourize the MO (50 mg L− 1). But while the MO is shaking, this strain is unable to decolourize it [38]. pH is one of the environmental factors that control the synthesis of enzymes and the biodegradation of dye molecules into microbial cells [38]. Different organic nitrogen sources, like BE, Pep, and YE, are usually used to enhance the colour degradation of azo dyes by different bacterial strains [39]. The MO was properly decoloured by ESR7, ESR13, ESR20, ESR21, ESD8, and ESB18 at all pH tests conducted under microaerophilic conditions. However, ESD8 was able to decolorize 33.7, 57.0, 32.9, and 31.9% at initial pH 5, pH 6, pH 8, and pH 9 under aerobic conditions [34]. Micrococcus glutamicus NCIM-2168 was used to break down the sulfonated diazo dye C.I. Reactive Green 19A. Under static conditions, this process was faster than under shaking conditions [40]. Similar results using Acinetobacter calcoaceticus showed that azo-dyed amaranth had maximal decolourization while in a static environment. When comparing static culture versus shaking (aerobic) conditions, several studies found that static culture results in more effective dye decolourization. Rhizobium radiobacter was shown to decolourize Direct Black 38 90% of the time under static settings and just 6% of the time under shaking conditions [17]. The Direct Black 38 was decolored in 5 days by Pseudomonas sp. in anaerobic circumstances, and the C. I. Acid Red 88 was decolored in 60 hours by Stenotrophomonas acidaminiphila (BN-3). In a static situation, the Staphylococcus hominis RMLRT03 strain decoloured Acid Orange at 85.52%, while in a shaking condition, decolourization was only 32.47% [41]. The bacterial isolate Acinetobacter baumannii JC359 could decolourize RR120 and showed 96% decolourization at 37 ⸰C, pH 9, within 48 hours [42]. S. haliotis, A. hydrophila, and S. putrefaciensare reported to be 27.71 %, 10.39 % and 27.28% decolourized the RR120 dye azo at pH 8.0, 35 ⸰C [43]. Acinetobacter junii strain FA10 showed maximum 80 % reactive red 120 degradation efficiency at 30 ⸰C, within 72 h of incubation [44].The best inoculum size and time conditions for RR120 decolourization in this study were 1%(0.3ml) inoculums during 12 hours of incubation under both microaerophilic situations. It was found that variation in pH, temperature, carbon-nitrogen, dye con., inoculum size, and aeration condition had a significant impact on RR120 breakdown percentage by L. capsici PB300 (T). The growth rate of L. capsici PB300 (T) was significantly reduced in a static state compared to a shaking condition.
4.3. RR120 Decolorization by Immobilised L. capsici PB300 (T)
A. jandaei strain SCS5 immobilised cells were able to decolourize MR, and when compared to immobilised cells alone, the decolourization rate was greatly increased by immobilised cells combined with mediators like anthraquinone-2,6-disulphonate and magnetic Fe3O4 nanoparticles [40]. The maximum decolourization was observed in the study during the biodegradation of RR120 in a batch continuous bioreactor using an immobilised L. capsici PB300 (T) bacterial cell.
4.4. RR120 Decolorization by Consortium Immobilised L. capsici PB300 (T)
The four bacterial consortiums Lysinibacillus capsici PB300(T), Alcaligenes faecalis subsp. Phenolicus DSM 16503 (T), Acinetobacter baumanni ATCC 19606 (T) and pseudomonas aeruginosa JCM 5962, showed the highest dye degradation percentage in <8 hours. Previous research indicated that the rate of dye degradation increases when multiple cultures are combined in a consortium. When it comes to wastewater treatment and dye bioremediation, microbial consortia work better than pure isolates. Multiple microbial compounds within the consortium co-metabolize, contributing to the expected complete and rapid destruction of the target pollutants [45]. Sphingomonas paucimobilis, Rhizobium radiobacter, and B. Subtilis were the three bacterial species that revealed high decolourization of azo dye decolourization in bacterial consortium throughout a single culture system. In comparison to their isolates, which took 72 hours, a group of 6 bacterial strains demonstrated an 84% decolourization percentage for a dye mixture in just 24 hours [31].
4.5. Decolorization of Many Azo Dyes and Dye Mixtures
In this study, we found that L. capsici PB300 (T) is effective in decolourization of a mixture of azo dyes viz. Reactive Red 120 and other azo dyes (e.g., 85% AB113, 94.62% orange II, 94.62% Congo red, 94.54% phenol red, and 81.66%). Similarly Pseudomonas sp. WS-D/183 is efficient in decolorization of dyes(100 mg/L conc.) RB5 (53.5±1.1%) followed by RR120 (18.3±1.3%), RY2 (17.7±3.2%),Blue Direct (14.7±1.7), Congo Red direct (7.5±2.7) and Orange Direct (3.4±1.1) at 30oC, 24 h[14]. Reactive purple, reactive red M5B, malachite green, and reactive pink MB mixed dyes (200 mg/L) were 70.8% decolorized by Bacillus subtilis and B. Cereus under aerobic conditions at pH 7, 37 oC, for 8 days [46]. Pseudomonas aeruginosa SVM16 decolorizes mixed dyes (50 mg/L) like RR 21 and RO16 at 90.2% after 96 hours under a static condition of 40 oC at pH 8.2, when it was supplemented with jack fruit seed powder [47]. Likewise Clostridium bifermentans, SL186 can decolourize mixed dyes(100 mg/L) reactive black-5, reactive red 3B-A and reactive yellow 3G-P in anaerobic medium at 36 h, pH 10, 35 ⸰C [48]. Brevibacillus laterosporus MTCC 2298 decoloured the mixture (70 mg/L) of structurally distinct dyes, including Remazol red, methyl red, golden yellow HER, rubine GFL, brown 3-REL, brilliant blue GL, scarlet RR, and yeast extract in a static micro-aerobic medium at pH 6–8, 40 oC, in 24 hours [49].
4.6. Heavy Metal Ions Tolerance Activity
Many of the strains linked to P. australis also showed strong resistance to metals like zinc, nickel, and copper, either when added singly or in combination in the growth medium. Since it may improve plant metal uptake and transport to aboveground tissues, metal tolerance is a beneficial characteristic in CW macrophyte plant-related bacteria [50]. When heavy metal ions are present in large concentrations, PGPR Bacillus sp. SR-2-1/1 exhibits excellent potential for decolorizing various azo dyes [36].
4.7. Plant Growth Promotion and EPS Production
We found that isolated bacteria L. capsici PB300 (T) were able to degrade azo dyes and produce IAA hormone and ammonium. Similarly, bacteria such as Pseudomonas, Azospirillum, and Rhizobium species eliminate hazardous chemical contaminants from polluted environments by enzymatic metabolic processes [2]. Besides production of EPS was revealed by plant growth-promoting reported L. capsici PB300 (T) bacteria. Similarly, at pH 6 and 40 °C, Exiguobacterium sp. VK1 97.61% decomposed Malachite Green dye (684.38 mg/g) in 30 minutes through the production of exopolysaccharide [51].
4.8. Enzyme Assay
Reactive red 120 dye is 95–99% decolorized by laccase and manganese peroxidase producing L. capsici PB300 (T), according to our findings. The presence of manganese peroxidase and laccase in the Lysinibacillus capsici PB300 (T) bacterial strain has been proven by the enzymes and laccase. These enzymes contribute to the detoxification of numerous chemicals used in the industry by converting poisons into metabolites that are less dangerous or non-toxic. Laccase is the most prevalent protein and the one that accumulates the least frequently in the decolourization experiment. Our findings demonstrated the critical role enzymes played in expediting the process of dye decolourization. Likewise, a strain of Serratia liquefaciens that generated over 90% of the lip was responsible for decolorizing AB dye, whereas Laccase and a strain of Streptomyces microflavus that produced lip were in charge of textile industrial wastewater decolourization The lip. and laccase producing Aeromonas hydrophila strain decolourize CV dye by 99% [52].
5. Conclusions
In this modern era, every person has a desired colourful life. To fulfil this desire for a colourful life, the use of the dye is growing day by day. One of the most affecting kinds of industrial effluent is thought to originate from the textile industry. The purpose of this study was to decolourize Reactive Red 120 dye using strains of Lysinibacillus capsici PB300 PGPR bacteria that were obtained from the Bark borer sample. In optimized conditions by manual and Taguchi strategy, more than 99% dye decolourization withglucose, ammonium nitrate, 1% inoculum size, dye concentrations of 0.2mg/l under static conditions at pH 8, 40 oC, in 12 hours. Four types of bacterial strains including Lysinibacillus capsici PB300 (T), Alcaligenes faecalis subsp. Phenolicus DSM 16503 (T), Acinetobacter baumanni ATCC 19606 (T) and pseudomonas aeruginosa JCM 5962 (T) were used for consortium development and the consortium result represented that complete decolourization time decreased. As a result, microbial consortia and cell immobilisation bioremediation are emerging as one of the most promising methods because of their ability to break down various kinds of environmental pollutants, strong resistance to complex environments, affordable cost, and environmental friendliness. When heavy metal ions like Zn, Pd, Cu, Ni, and Cd are present, Lysinibacillus capsici PB300 (T)can break the dye. It was reported that the Lysinibacillus capsici strain could decolourize coloured fabric and create EPS. FTIR, UV-Vis and GC-MS analysis confirmed that the dye has been converted to its amines by bacterial metabolism, which is less toxic than the parent dye.
Author Contributions
Kusumlata.; writing—original data preparation, Rajat Pratap Singh, and Balram Ambade.;Data analysis and review, Ashish Kumar.;Data analysis, review, editing and visualization.
Funding
Not Applicable.
Acknowledgments
The authors are thankful to the Central Instrumentation Lab, Central University of Punjab, Bhatinda for GC-MS analysis and National Centre for Microbial Resource, Pune, Maharashtra for Molecular identification of bacterial isolates.
Conflicts of Interest
No conflict of interest.
References
- Ayele, A.; Getachew, D.; Kamaraj, M.; Suresh, A. Phytoremediation of synthetic dyes: an effective and eco-friendly algal technology for the dye abatement. Journal of Chemistry 2021, 1–14. [Google Scholar]
- Rehaman, S.; Aravindan, G.; Karthick, G. Spectral studies of Azo Dye degradation using selected Biofertilizer of Pseudomonas fluorescens. Int. J. Life Sci. Pharma Res, 2021, 11, L73–79. [Google Scholar]
- Florencio, T.D.M.; de Godoi, L.A.; Rocha, V.C.; Oliveira, J.M.S.; Motteran, F.; Gavazza, S.; Damianovic, M.H.R.Z. Anaerobic structured-bed reactor for azo dye decolourization in the presence of sulfate ions. Journal of Chemical Technology & Biotechnology, 2021, 96, 1700–1708. [Google Scholar]
- Carolin, C.F.; Kumar, P.S.; Joshiba, G.J. A sustainable approach to decolourize methyl orange dye from aqueous solution using novel bacterial strain and its metabolites characterization. Clean Technologies and Environmental Policy 2021, 23, 173–181. [Google Scholar] [CrossRef]
- Kour, D.; Khan, S.S.; Kour, H.; Kaur, T.; Devi, R.; Rai, P.K.; Yadav, A.N. Microbe-mediated bioremediation: Current research and future challenges. Journal of Applied Biology and Biotechnology, 2023, 10, 6–24. [Google Scholar] [CrossRef]
- Kusumlata; Ambade, B.; Kumar, A.; Gautam, S. Sustainable Solutions: Reviewing the Future of Textile Dye Contaminant Removal with Emerging Biological Treatments. Limnological Review. 2024, 24, 126–49. [Google Scholar]
- Sreedharan, V.; Saha, P.; Rao, K.V.B. Dye degradation potential of Acinetobacter baumannii strain VITVB against commercial azo dyes. Bioremediation Journal 2021, 25, 347–368. [Google Scholar] [CrossRef]
- Lalnunhlimi, S.; Krishnaswamy, V. Decolorization of azo dyes (Direct Blue 151 and Direct Red 31) by moderately alkaliphilic bacterial consortium. brazilian journal of microbiology, 2016, 47, 39–46. [Google Scholar] [CrossRef]
- Lade, H.; Govindwar, S.; Paul, D. Low-cost biodegradation and detoxification of textile azo dye CI reactive blue 172 by Providencia rettgeri strain HSL1. Journal of Chemistry 2015.
- Chaturvedi, A.; Rai, B.N.; Singh, R.S.; Jaiswal, R.P. A computational approach to incorporate metabolite inhibition in the growth kinetics of indigenous bacterial strain Bacillus subtilis MN372379 in the treatment of wastewater containing Congo red dye. Applied Biochemistry and Biotechnology, 2021, 193, 2128–2144. [Google Scholar] [CrossRef]
- Chen, G.; An, X.; Li, H.; Lai, F.; Yuan, E.; Xia, X.; Zhang, Q. Detoxification of azo dye Direct Black G by thermophilic Anoxybacillus sp. PDR2 and its application potential in bioremediation. Ecotoxicology and Environmental Safety, 2021, 214, 112084. [Google Scholar] [CrossRef] [PubMed]
- Dixit, S.; Garg, S. Enzymatic degradation of sulphonated azo dye using purified azoreductase from facultative Klebsiella pneumoniae. Folia Microbiologica 2021, 66, 79–85. [Google Scholar] [CrossRef]
- Guembri, M.; Neifar, M.; Saidi, M.; Ferjani, R.; Chouchane, H.; Mosbah, A.; Ouzari, H.I. Decolorization of textile azo dye Novacron Red using bacterial monoculture and consortium: Response surface methodology optimization. Water Environment Research 2021, 93, 1346–1360. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Maqbool, Z.; Shahid, M.; Shahzad, T.; Muzammil, S.; Zubair, M.; Mahmood, F. Simultaneous removal of reactive dyes and hexavalent chromium by a metal-tolerant Pseudomonas sp. WS-D/183 harbouring plant growth-promoting traits. Int J Agric Biol 2020, 23, 241–252. [Google Scholar] [CrossRef]
- Kamal, I.M.; Abdeltawab, N.F.; Ragab, Y.M.; Farag, M.A.; Ramadan, M.A. Biodegradation, decolorization, and detoxification of di-azo dye direct Red 81 by halotolerant, alkali-thermo-tolerant bacterial mixed cultures. Microorganisms 2022, 10, 994. [Google Scholar] [CrossRef]
- Ayed, L.; Bekir, K.; Jabeur, C. Modelling and optimization of biodegradation of methylene blue by Staphylococcus aureus through a statistical optimization process: a sustainable approach for waste management. Water Science and Technology 2022, 86, 380–394. [Google Scholar] [CrossRef] [PubMed]
- Telke, A.; Kalyani, D.; Jadhav, J.; Govindwar, S. Kinetics and Mechanism of Reactive Red 141 Degradation by a Bacterial Isolate Rhizobium radiobacter MTCC 8161. Acta Chimica Slovenica 2008, 55. [Google Scholar]
- Pourbabaee, A.A.; Ramezani, S.; Javaheri Daneshmand, H. Biodegradation of malachite green by Klebsiella Terrigenaptcc 1650: the critical parameters were optimized using Taguchi optimization method. J. Bioremed. Biodeg, 2013, 4, 175. [Google Scholar]
- Marvi-Mashhadi, A.; Sharifmoghadam, M.R.; Goharimanesh, M.; Marvi-Mashhadi, M.; Dehghan, H.; Bahreini, M. Methyl red biodegradation based on Taguchi method by two novel bacteria. International Journal of Environmental Science and Technology 2021, 1–12. [Google Scholar] [CrossRef]
- Thiruppathi, K.; Rangasamy, K.; Ramasamy, M.; Muthu, D. Evaluation of textile dye degrading potential of ligninolytic bacterial consortia. Environmental Challenges, 2021, 4, 100078. [Google Scholar] [CrossRef]
- Hameed, B.B.; Ismail, Z.Z. Decolorization, biodegradation and detoxification of reactive red azo dye using non-adapted immobilized mixed cells. Biochemical Engineering Journal, 2018, 137, 71–77. [Google Scholar] [CrossRef]
- Kathiravan, M.N.; Praveen, S.A.; Gim, G.H.; Han, G.H.; Kim, S.W. Biodegradation of Methyl Orange by alginate-immobilized Aeromonas sp. in a packed bed reactor: external mass transfer modelling. Bioprocess and biosystems engineering, 2014, 37, 2149–2162. [Google Scholar] [CrossRef]
- Kaur, J.; Mudgal, G.; Negi, A.; Tamang, J.; Singh, S.; Singh, G.B.; Kesari, K.K. Reactive Black-5, Congo Red and Methyl Orange: Chemical Degradation of Azo-Dyes by Agrobacterium. Water, 2023, 15, 1664. [Google Scholar] [CrossRef]
- Islam, S.; Akanda, A.M.; Prova, A.; Islam, M.T.; Hossain, M.M. Isolation and identification of plant growth promoting rhizobacteria from cucumber rhizosphere and their effect on plant growth promotion and disease suppression. Frontiers in microbiology, 2016, 6, 1360. [Google Scholar] [CrossRef]
- Passari, A.K.; Mishra, V.K.; Leo, V.V.; Gupta, V.K.; Singh, B.P. Phytohormone production endowed with antagonistic potential and plant growth promoting abilities of culturable endophytic bacteria isolated from Clerodendrum colebrookianum Walp. Microbiological research, 2016, 193, 57–73. [Google Scholar] [CrossRef]
- Thanavel, M.; Bankole, P.O.; Selvam, R.; Govindwar, S.P.; Sadasivam, S.K. Synergistic effect of biological and advanced oxidation process treatment in the biodegradation of Remazol yellow RR dye. Scientific reports, 2020, 10, 20234. [Google Scholar] [CrossRef]
- Patel, V.R.; Bhatt, N. Isolation, development and identification of salt-tolerant bacterial consortium from crude-oil-contaminated soil for degradation of di-azo dye Reactive Blue 220. Water Science and Technology, 2015, 72, 311–321. [Google Scholar] [CrossRef]
- Vinayak, A.; Singh, G.B. Synthetic azo dye bio-decolorization by Priestia sp. RA1: process optimization and phytotoxicity assessment. Archives of Microbiology, 2022, 204, 318. [Google Scholar] [CrossRef]
- Singh, G.; Dwivedi, S.K. Decolorization and degradation of Direct Blue-1 (Azo dye) by newly isolated fungus Aspergillus terreus GS28, from sludge of carpet industry. Environmental technology & innovation 2020, 18, 100751. [Google Scholar]
- Abd El-Rahim, W.M.; Moawad, H.; Azeiz, A.Z.; Sadowsky, M.J. Biodegradation of azo dyes by bacterial or fungal consortium and identification of the biodegradation products. The Egyptian Journal of Aquatic Research. 2021, 47, 269–76. [Google Scholar] [CrossRef]
- Henagamage, A.P.; Peries, C.M. Degradation and decolorization of textile azo dyes by effective fungal-bacterial consortium. Molecular Biology Reports, 2023, 50, 8901–8914. [Google Scholar] [CrossRef]
- Thanavel, M.; Bankole, P.O.; Selvam, R.; Govindwar, S.P.; Sadasivam, S.K. Synergistic effect of biological and advanced oxidation process treatment in the biodegradation of Remazol yellow RR dye. Scientific reports, 2020, 10, 20234. [Google Scholar] [CrossRef]
- Si, J.; Peng, F.; Cui, B. Purification, biochemical characterization and dye decolorization capacity of an alkali-resistant and metal-tolerant laccase from Trametes pubescent. Bioresource Technology, 2013, 128, 49–57. [Google Scholar] [CrossRef]
- Haque, MM.; Hossen, MN.; Rahman, A.; Roy, J.; Talukder, MR.; Ahmed, M.; Ahiduzzaman, M.; Haque, MA. Decolorization, degradation and detoxification of mutagenic dye Methyl orange by novel biofilm-producing plant growth-promoting rhizobacteria. Chemosphere 2024, 346, 140568. [Google Scholar] [CrossRef]
- Shahid, M.; Mahmood, F.; Hussain, S.; Shahzad, T.; Haider, M.Z.; Noman, M.; Mustafa, G. Enzymatic detoxification of azo dyes by a multifarious Bacillus sp. strain MR-1/2-bearing plant growth-promoting characteristics. 3 Biotech 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Mahmood, F.; Shahid, M.; Hussain, S.; Shahzad, T.; Tahir, M.; Ijaz, M.; Babar, S.A.K. Potential plant growth-promoting strain Bacillus sp. SR-2-1/1 decolorized azo dyes through NADH-ubiquinone: oxidoreductase activity. Bioresource Technology 2017, 235, 176–184. [Google Scholar] [CrossRef]
- Pham, VH.; Kim, J.; Chang, S.; Bang, D. Investigating bio-inspired degradation of toxic dyes using potential multi-enzyme producing extremophiles. Microorganisms 2023, 11, 1273. [Google Scholar] [CrossRef]
- Parshetti, G.K.; Telke, A.A.; Kalyani, D.C.; Govindwar, S.P. ; Decolorization and detoxification of sulfonated azo dye methyl orange by Kocuria rosea MTCC 1532. J. Hazard Mater. 2010, 176, 503–509. [Google Scholar] [CrossRef]
- Guo, G.; Hao, J.; Tian, F.; Liu, C.; Ding, K.; Xu, J.; Zhou, W.; Guan, Z. Decolorization and detoxification of azo day by halo-alkaliphilic bacterial consortium: systematic investigations of performance, pathway and metagenome. Ecotoxicol. Environ. Saf. 2020, 204, 111073. [Google Scholar] [CrossRef]
- Sharma, S.C.D.; Sun, Q.; Li, J.; Wang, Y.; Suanon, F.; Yang, J.; Yu, C.P. Decolorization of azo dye methyl red by suspended and co-immobilized bacterial cells with mediators anthraquinone-2, 6-disulfonate and Fe3O4 nanoparticles. International Biodeterioration & Biodegradation 2016, 112, 88–97. [Google Scholar]
- Singh, R.P.; Singh, P.K.; Singh, R.L. Bacterial decolorization of textile azo dye acid orange by Staphylococcus hominis RMLRT03. Toxicology international 2014, 21, 160. [Google Scholar] [CrossRef]
- Ameenudeen, S.; Unnikrishnan, S.; Ramalingam, K. Statistical optimization for the efficacious degradation of reactive azo dyes using Acinetobacter baumannii JC359. Journal of Environmental Management 2021, 279, 111512. [Google Scholar] [CrossRef]
- Radhika, B.; Aruna, K. Elucidating a pathway for degradation of azo dye reactive red 120 BY bacterial consortium. Journal of Applied Biological Sciences, 2022, 16, 396–417. [Google Scholar]
- Anwar, F.; Hussain, S.; Ramzan, S.; Hafeez, F.; Arshad, M.; Imran, M.; Abbas, N. Characterization of reactive red-120 decolorizing bacterial strain Acinetobacter junii FA10 capable of simultaneous removal of azo dyes and hexavalent chromium. Water, Air, & Soil Pollution 2014, 225, 1–16. [Google Scholar]
- Agrawal, S.; Tipre, D.; Dave, S.R. Biotreatment of azo dye containing textile industry effluent by a developed bacterial consortium immobilised on brick pieces in an indigenously designed packed bed biofilm reactor. World Journal of Microbiology and Biotechnology 2023, 39, 83. [Google Scholar] [CrossRef]
- Maheswari, N.U.; Sivagami, S. Biological degradation of textile dyes using marine Bacillus species. Int J Pure Appl Biosci 2016, 4, 123–128. [Google Scholar] [CrossRef]
- Mishra, S.; Maiti, A. Optimization of process parameters to enhance the bio-decolorization of Reactive Red 21 by Pseudomonas aeruginosa 23N1. International Journal of Environmental Science and Technology 2019, 16, 6685–6698. [Google Scholar] [CrossRef]
- Joe, M.H.; Lim, S.Y.; Kim, D.H.; Lee, I.S. Decolorization of reactive dyes by Clostridium bifermentans SL186 isolated from contaminated soil. World Journal of Microbiology and Biotechnology 2008, 24, 2221–2226. [Google Scholar] [CrossRef]
- Kurade, M.B.; Waghmode, T.R.; Govindwar, S.P. Preferential biodegradation of structurally dissimilar dyes from a mixture by Brevibacillus laterosporus. Journal of hazardous materials 2011, 192, 1746–1755. [Google Scholar] [CrossRef]
- Zhuang, M.; Sanganyado, E.; Xu, L.; Zhu, J.; Li, P.; Liu, W. High throughput sediment DNA sequencing reveals azo dye degrading bacteria inhabit nearshore sediments. Microorganisms 2020, 8, 233. [Google Scholar] [CrossRef]
- Kalpana, R.; Vignesh, N.S.; Vinothini, K.; Rajan, M.; Ashokkumar, B.; Brindhadevi, K.; Varalakshmi, P. Carbon quantum dots (CQD) fabricated from Exiguobacterium sp. VK2 exopolysaccharide (EPS) using hydrothermal reaction and its biodiesel applications. Fuel 2023, 333, 126426. [Google Scholar] [CrossRef]
- Kishor, R.; Purchase, D.; Saratale, G.D.; Ferreira, L.F.; Bilal, M.; Iqbal, H.M.; Bharagava, R.N. Environment friendly degradation and detoxification of Congo red dye and textile industry wastewater by a newly isolated Bacillus cohnni (RKS9). Environmental Technology & Innovation 2021, 22, 101425. [Google Scholar]
Figure 1.
Nebour Joining three for identified bacterial isolate Lysinibacilluscapsici PB300(T).
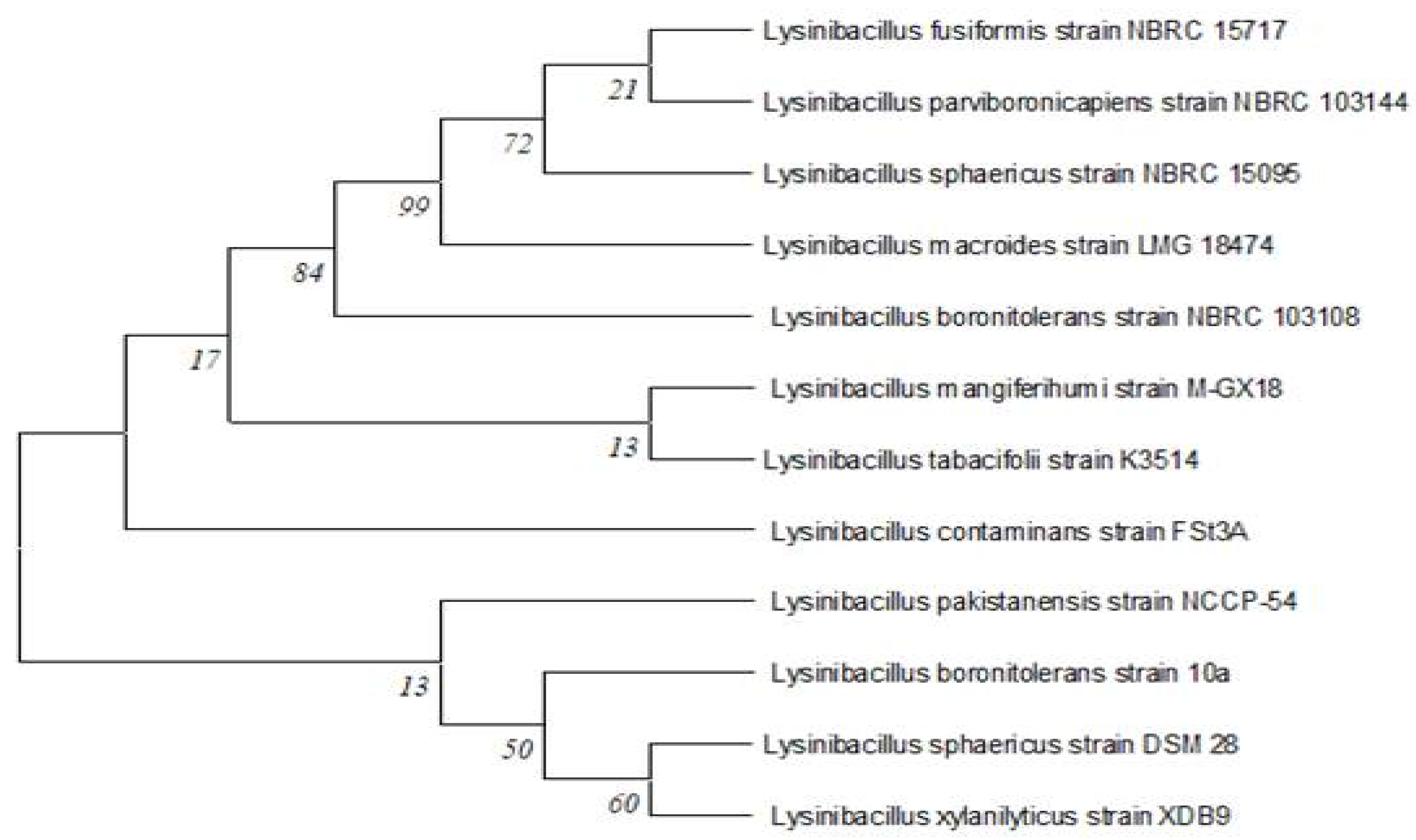
Figure 2.
Decolorized broth media for bacteria isolation.

Figure 3.
a) Effect of cell growth, b) aeration and incubation time (static and shaking) and c) inoculum size on decolourization of RR120 dye.
Figure 3.
a) Effect of cell growth, b) aeration and incubation time (static and shaking) and c) inoculum size on decolourization of RR120 dye.
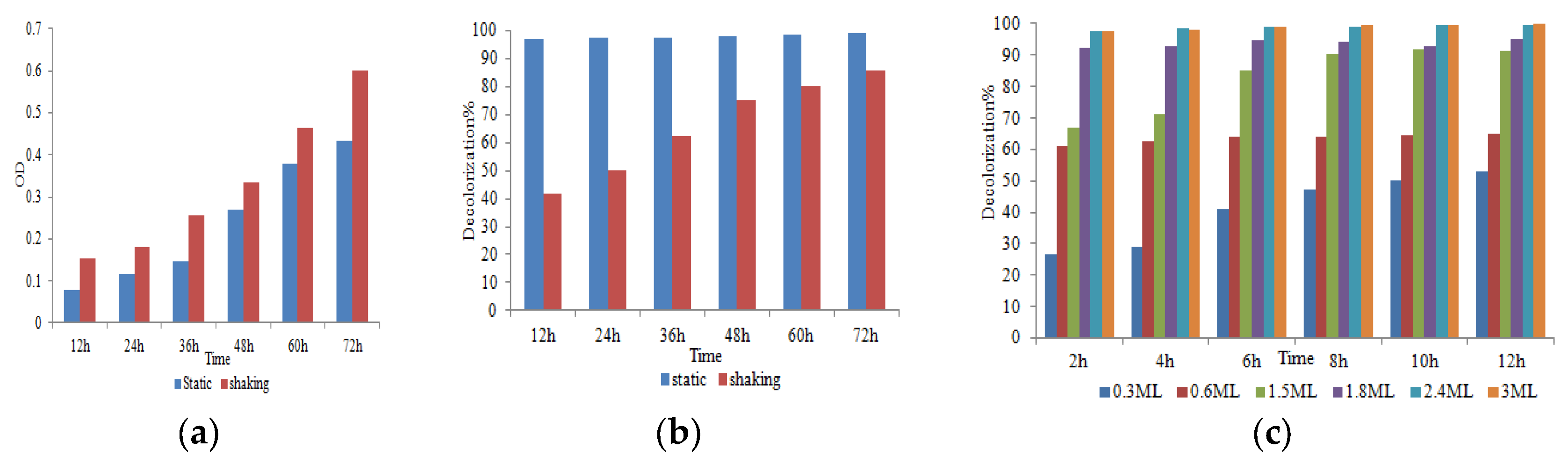
Figure 4.
Main effects plot for data means response.
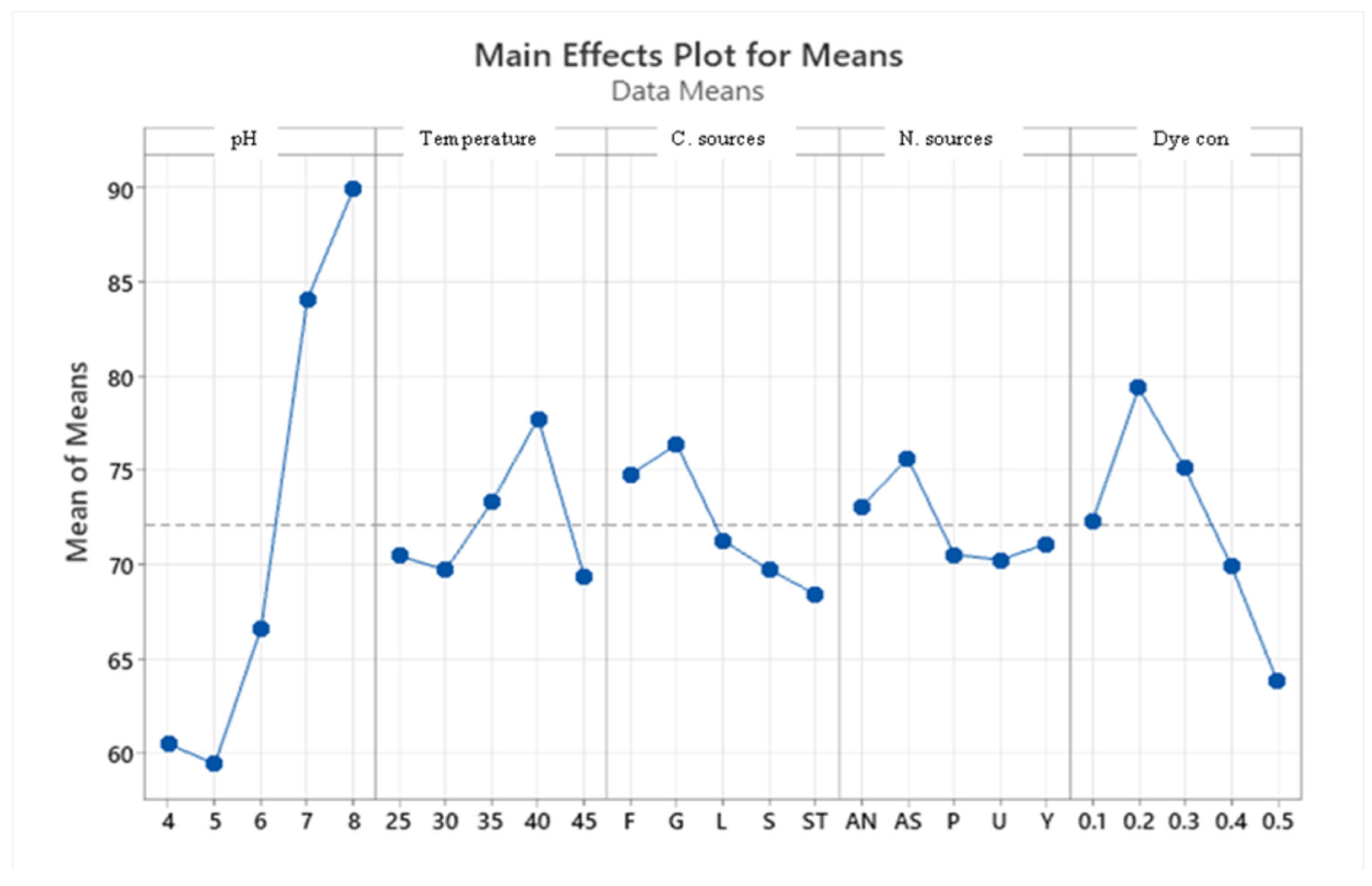
Figure 5.
Main effects plot for data signal-to-noise ratio.
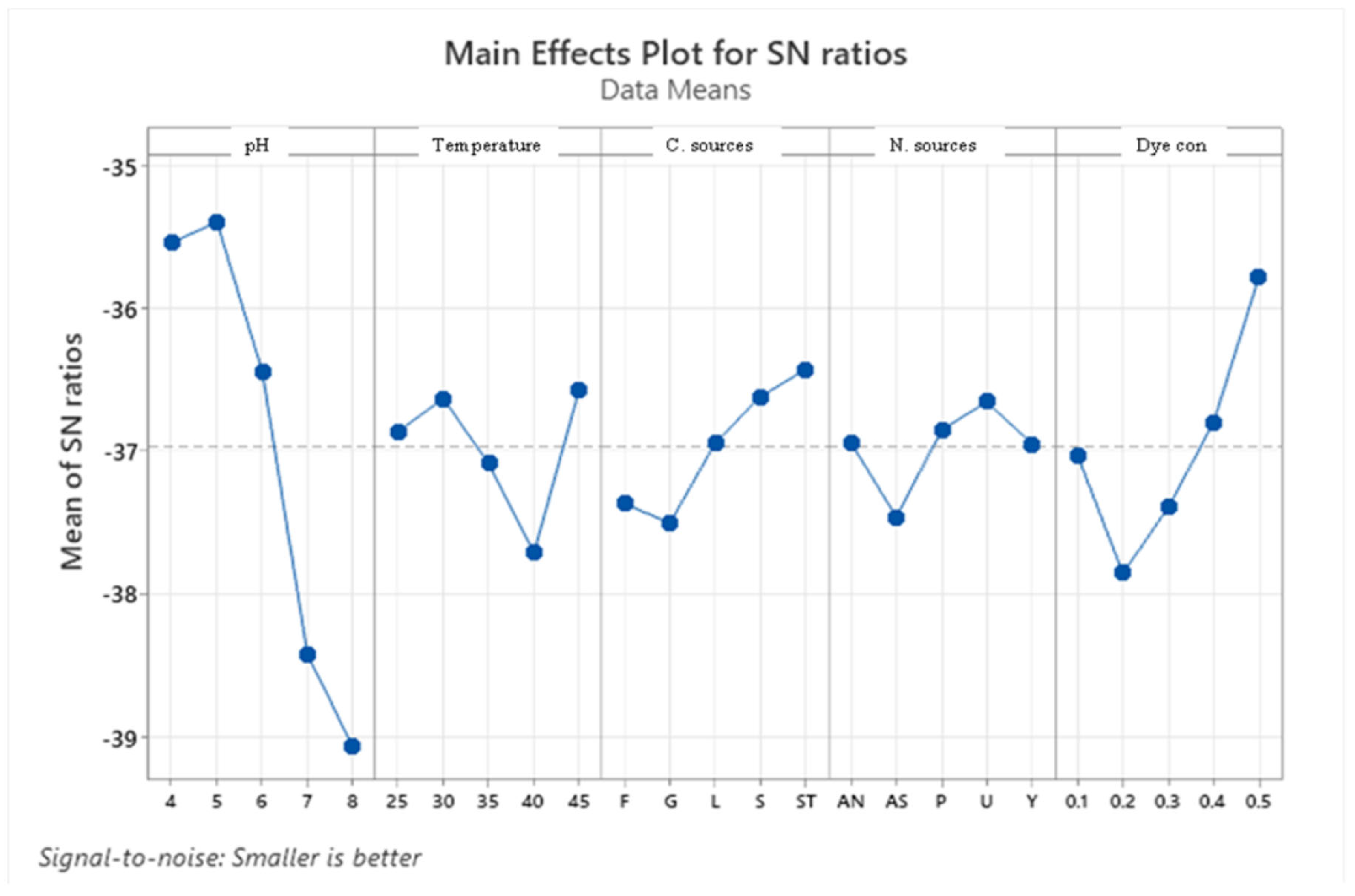
Figure 6.
Decolorization percentage analysis of azo dyes and mixtures of various azo dyes.
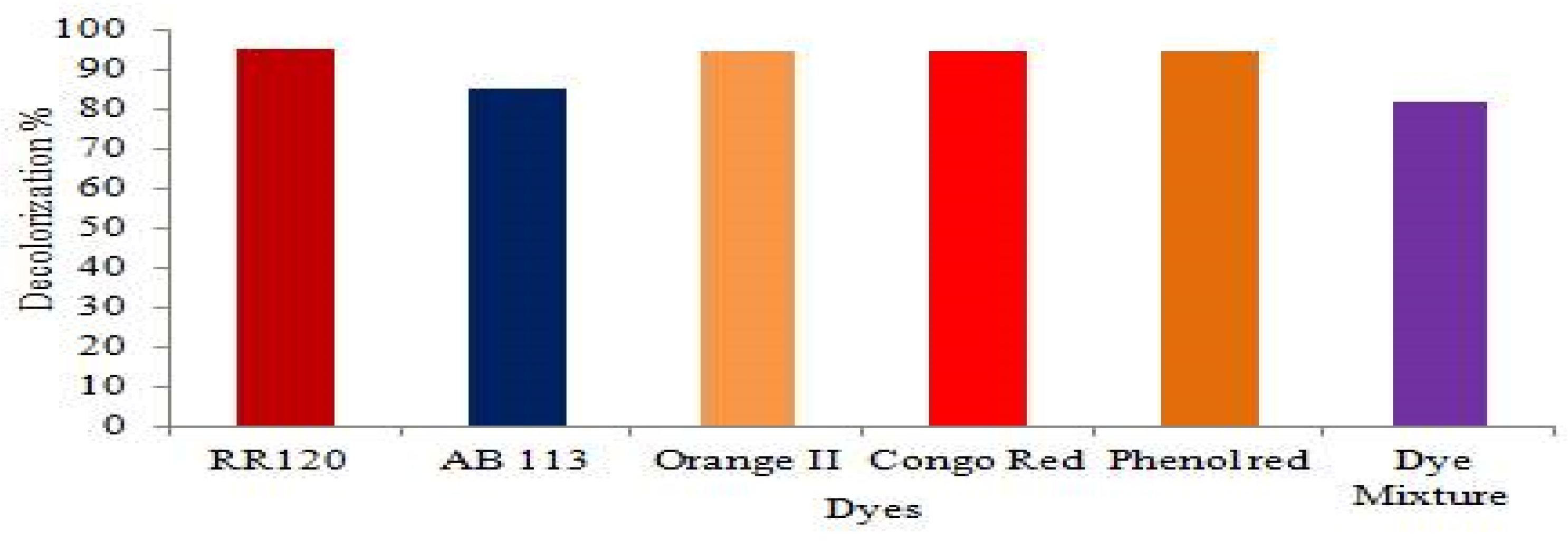
Figure 7.
Effect of heavy metals and NaCl on decolourization of RR120.
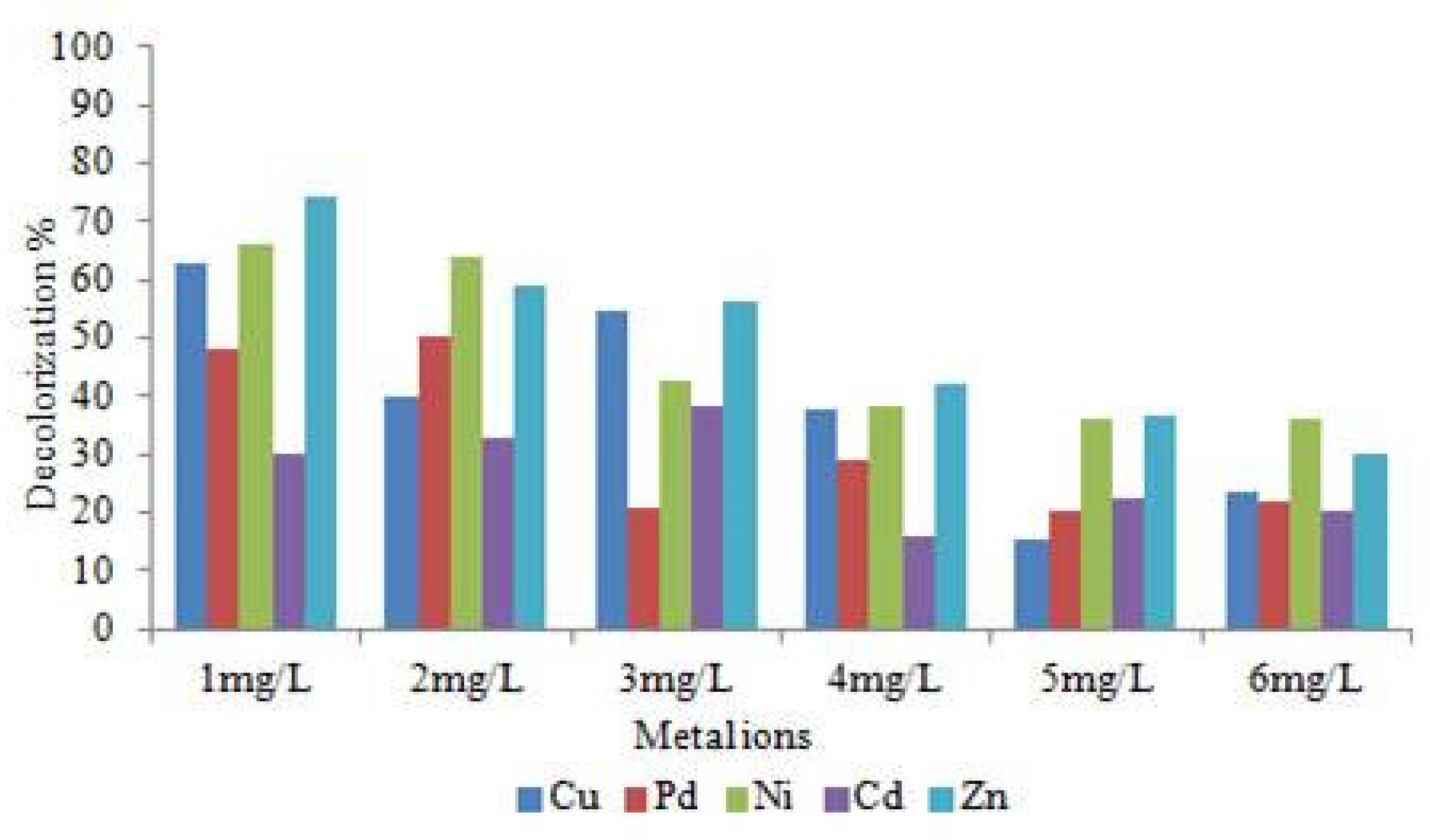
Figure 8.
Decolourization of the colored fabric.

Figure 9.
UV-Vis Spectroscopy analysis for decolorization and biodegradation of RR120 dye.
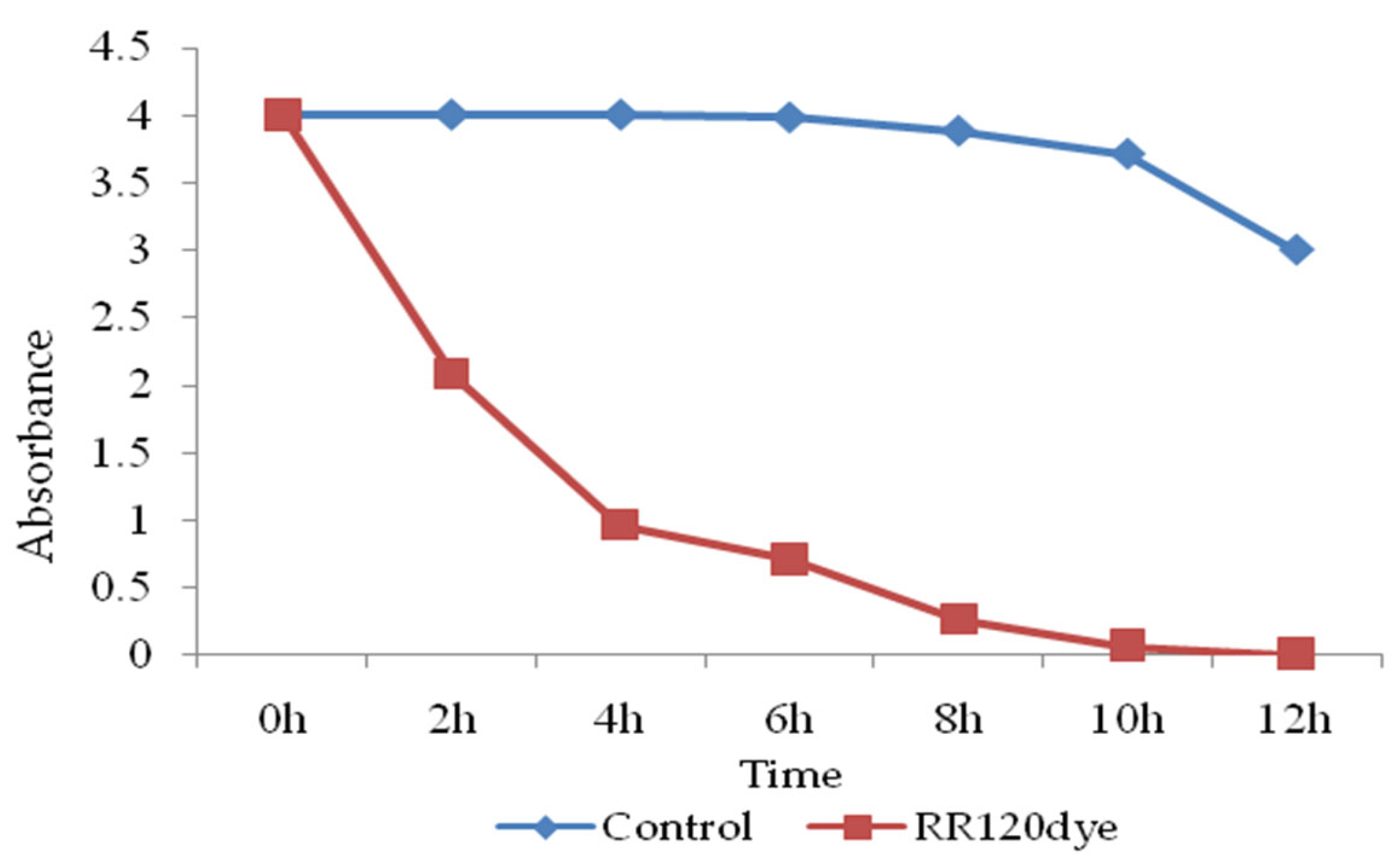
Figure 10.
FTIR analysis of a) RR120 dye, andb) dye metabolites.
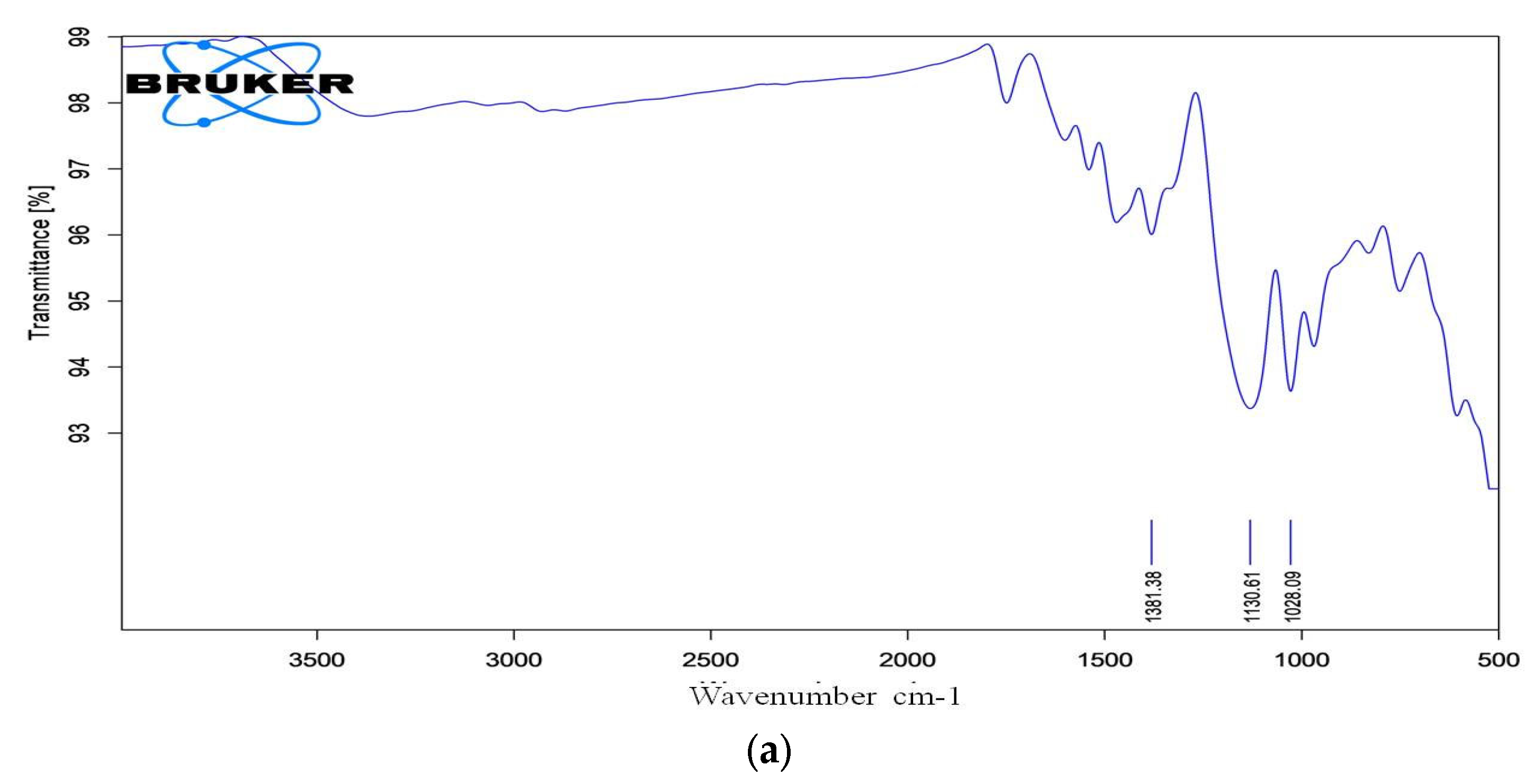
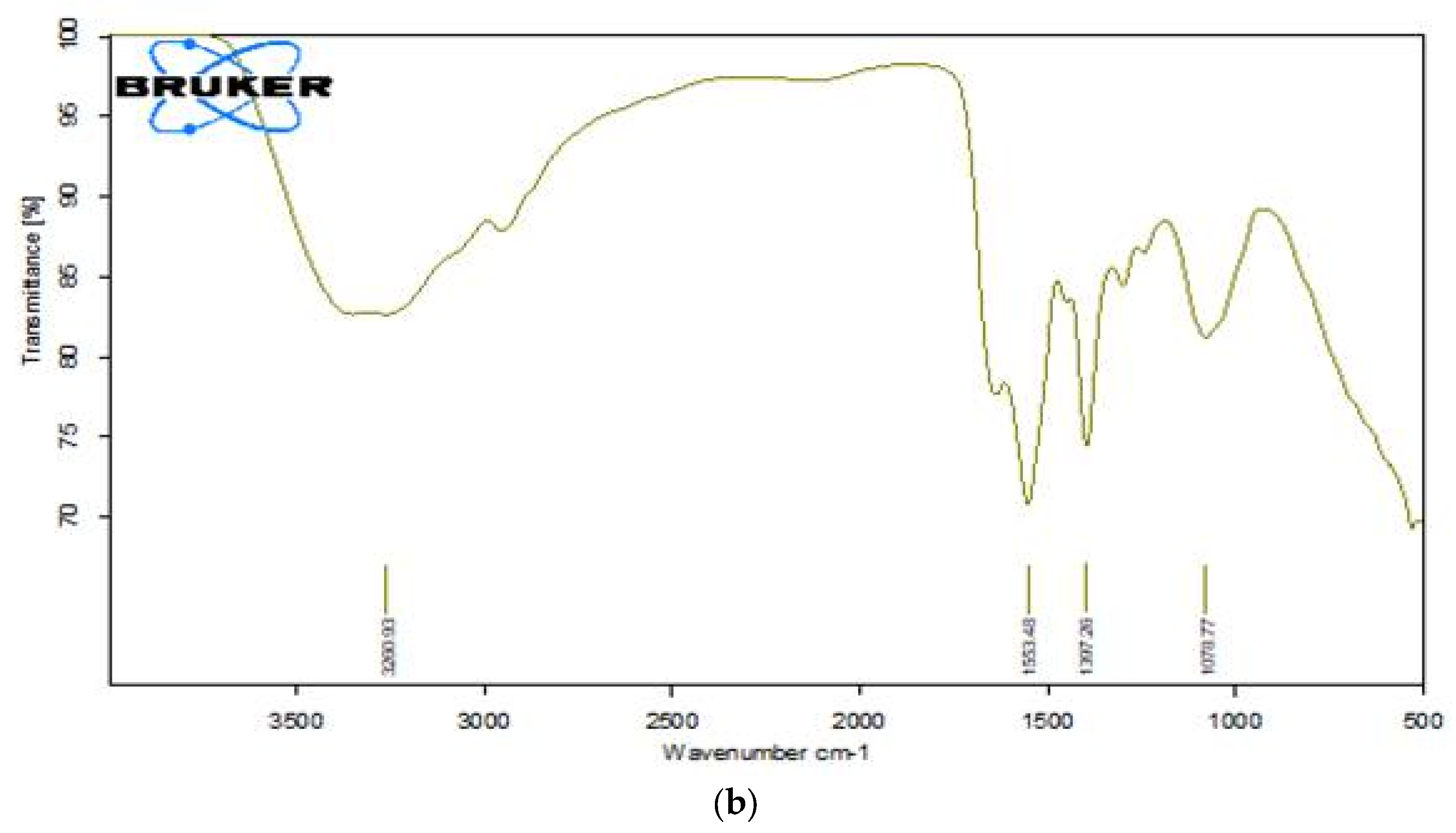
Figure 11.
GC-MS analysis of metabolites produced by Lysinibacillus capsici PB300 (T).
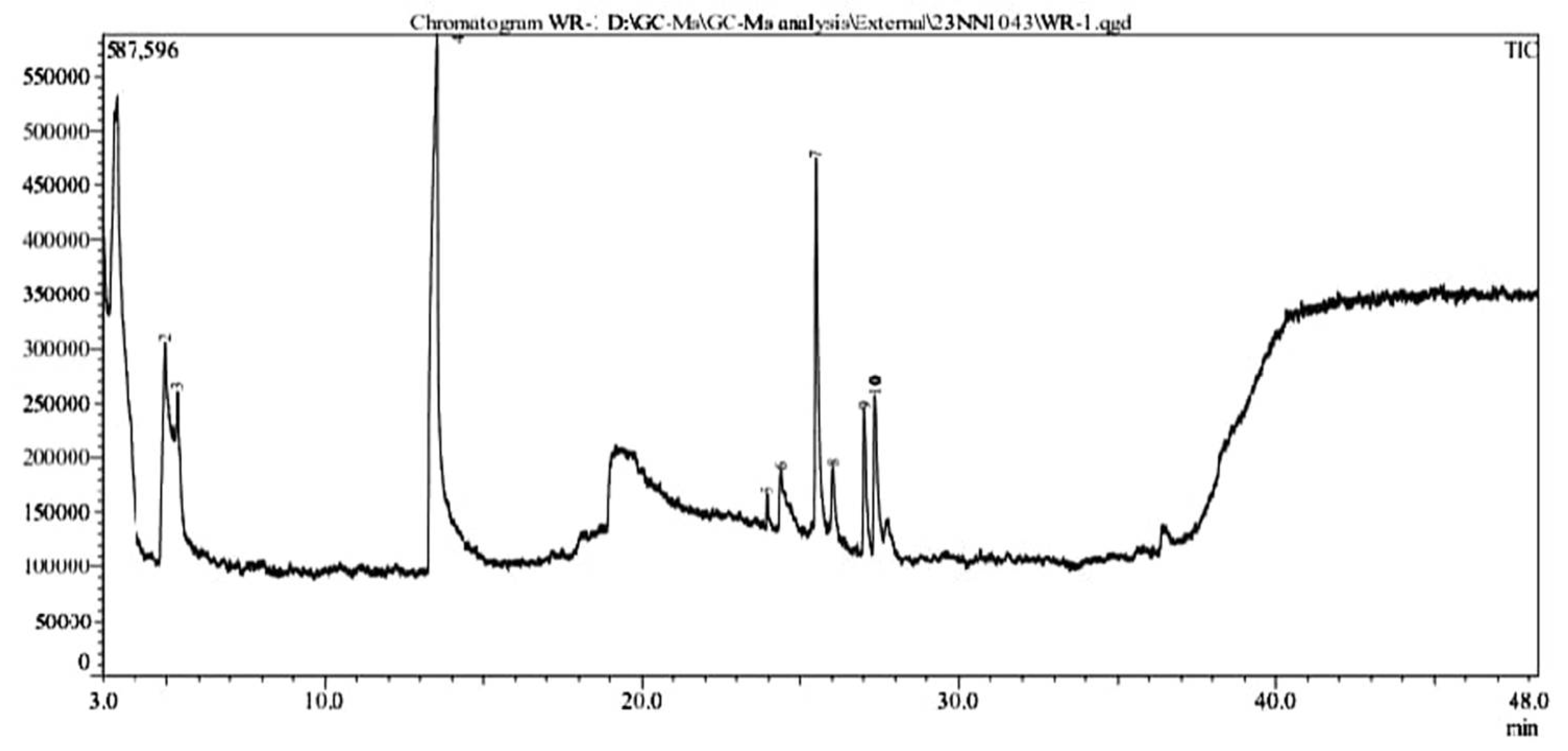

Table 1.
Taguchi’s L25 orthogonal array, decolourization percentage, means value and S/N ratio for optimization of RR120 decolourization, using Lysinibacillu scapsici PB300 (T) bacterial strain.
Table 1.
Taguchi’s L25 orthogonal array, decolourization percentage, means value and S/N ratio for optimization of RR120 decolourization, using Lysinibacillu scapsici PB300 (T) bacterial strain.
| Run | Factors | RR120 Decolourization % |
Mean values |
S/N Ratio | ||||
|---|---|---|---|---|---|---|---|---|
| pH | Temp. | Carbon source |
Nitrogen Source |
Dye concentration |
||||
| L1 | 4 | 25 | G | Y | 0.1 | 59.305 | 59.305 | -35.4618 |
| L2 | 4 | 30 | F | P | 0.2 | 67.63 | 67.63 | -36.6028 |
| L3 | 4 | 35 | S | AS | 0.3 | 64.04 | 64.04 | -36.129 |
| L4 | 4 | 40 | L | AN | 0.4 | 66.62 | 66.62 | -36.4721 |
| L5 | 4 | 45 | ST | U | 0.5 | 44.8 | 44.8 | -33.0256 |
| L6 | 5 | 25 | F | AS | 0.4 | 62.74 | 62.74 | -35.9509 |
| L7 | 5 | 30 | S | AN | 0.5 | 44.32 | 44.32 | -32.932 |
| L8 | 5 | 35 | L | U | 0.1 | 59.335 | 59.335 | -35.4662 |
| L9 | 5 | 40 | ST | Y | 0.2 | 65.825 | 65.825 | -36.3678 |
| L10 | 5 | 45 | G | P | 0.3 | 64.91 | 64.91 | -36.2462 |
| L11 | 6 | 25 | S | U | 0.2 | 70.515 | 70.515 | -36.9656 |
| L12 | 6 | 30 | L | Y | 0.3 | 66.255 | 66.255 | -36.4244 |
| L13 | 6 | 35 | ST | P | 0.4 | 57.39 | 57.39 | -35.1767 |
| L14 | 6 | 40 | G | AS | 0.5 | 72.945 | 72.945 | -37.2599 |
| L15 | 6 | 45 | F | AN | 0.1 | 65.86 | 65.86 | -36.3724 |
| L16 | 7 | 25 | L | P | 0.5 | 69.87 | 69.87 | -36.8858 |
| L17 | 7 | 30 | ST | AS | 0.1 | 84.27 | 84.27 | -38.5135 |
| L18 | 7 | 35 | G | AN | 0.2 | 98.67 | 98.67 | -39.8837 |
| L19 | 7 | 40 | F | U | 0.3 | 90.495 | 90.495 | -39.1325 |
| L20 | 7 | 45 | S | Y | 0.4 | 76.91 | 76.91 | -37.7197 |
| L21 | 8 | 25 | ST | AN | 0.3 | 89.72 | 89.72 | -39.0578 |
| L22 | 8 | 30 | G | U | 0.4 | 85.945 | 85.945 | -38.6844 |
| L23 | 8 | 35 | F | Y | 0.5 | 87.095 | 87.095 | -38.7999 |
| L24 | 8 | 40 | S | P | 0.1 | 92.75 | 92.75 | -39.3463 |
| L25 | 8 | 45 | L | AS | 0.2 | 94.135 | 94.135 | -39.475 |
Table 2.
Mean response and Signal-to-Noise Ratios calculation of Taguchi data for optimization of the physical and nutritional factors and their highest levels of RR120 dye decolourization by the Lysinibacillus capsici PB300 (T).
Table 2.
Mean response and Signal-to-Noise Ratios calculation of Taguchi data for optimization of the physical and nutritional factors and their highest levels of RR120 dye decolourization by the Lysinibacillus capsici PB300 (T).
| Level | pH | Temperature | Carbon source | Nitrogen source | Dye concentration | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | S/N | Mean | S/N | Mean | S/N | Mean | S/N | Mean | S/N | |
| 1 | 60.48 | -35.54 | 70.43 | -36.86 | 74.76 | -37.37 | 73.04 | -36.94 | 72.3 | -37.03 |
| 2 | 59.43 | -35.39 | 69.68 | -36.63 | 76.35 | -37.51 | 75.63 | -37.47 | 79.35 | -37.86 |
| 3 | 66.59 | -36.44 | 73.31 | -37.09 | 71.24 | -36.94 | 70.51 | -36.85 | 75.08 | -37.4 |
| 4 | 84.04 | -38.43 | 77.73 | -37.72 | 69.71 | -36.62 | 70.22 | -36.65 | 69.92 | -36.8 |
| 5 | 89.93 | -39.07 | 69.32 | -36.57 | 68.4 | -36.43 | 71.08 | -36.95 | 63.81 | -35.78 |
| Delta | 30.5 | 3.68 | 8.4 | 1.15 | 7.95 | 1.08 | 5.41 | 0.81 | 15.55 | 2.08 |
| Rank | 1 | 1 | 3 | 3 | 4 | 4 | 5 | 5 | 2 | 2 |
Table 3.
Consortium combination and dye decolourization percentage.
| Bacterial | Code | Combination No. | Combination | Dye degradation% | |||||
| Lysinibacilluscapsici PB300(T) | A | 2h | 4h | 6h | 8h | 10h | 12h | ||
| 1. | A+B | 30.25 | 45.47 | 74.32 | 80.16 | 99.59 | 96.76 | ||
| 2. | A+C | 39 | 51.37 | 69.85 | 87.54 | 89.68 | 99.44 | ||
| 3. | A+D | 47.17 | 65.87 | 78.82 | 79.21 | 89.27 | 99.81 | ||
| Alcaligenesfaecalissubsp. Phenolicus DSM 16503(T) | B | 4. | B+C | 39.62 | 60.37 | 73.95 | 88.95 | 99.15 | 99.94 |
| 5. | B+D | 29.65 | 62.95 | 80.27 | 89.38 | 99.21 | 98.76 | ||
| 6. | C+D | 45.47 | 65.67 | 70.68 | 89.12 | 99.59 | 99.11 | ||
| Acinetobacterbaumanni ATCC 19606(T) | C | 7. | A+B+C | 51.4 | 61.35 | 87.81 | 99.73 | 99.82 | 100 |
| 8. | A+B+D | 40.42 | 65.6 | 87.04 | 99.56 | 99.79 | 99.46 | ||
| Pseudomonasaeruginosa JCM 5962(T) | D | 9. | A+C+D | 33.45 | 65 | 88.96 | 99.76 | 99.14 | 99.48 |
| 10. | A+B+C+D | 49.87 | 72.01 | 87.76 | 99.91 | 100 | 100 | ||
Table 4.
Compound, Molecular formula, R. Time and Molecular Weight of Dye RR120 degraded metabolites studied using GC-MS.
Table 4.
Compound, Molecular formula, R. Time and Molecular Weight of Dye RR120 degraded metabolites studied using GC-MS.
| No. | Compound | Molecular formula | R.Time (min.) | Molecular Weight (g/mol) | Peak Area % | Peak Height % |
|---|---|---|---|---|---|---|
| 1. | Butanoic acid | C5H10O2 | 3.433 | 102 | 49.38 | 23.88 |
| 2. | Pentanoic acid | C6H12O2 | 4.953 | 116 | 3.22 | 5.54 |
| 3. | Pentanoic acid | C6H12O2 | 5.345 | 116 | 0.90 | 2.85 |
| 4. | 2-Piperidinone | C5H9NO | 13.527 | 99 | 26.72 | 25.43 |
| 5. | Tridecanoic acid | C15H30O2 | 23.945 | 242 | 0.37 | 1.64 |
| 6. | 1,4-diazabicyclo[4.3.0]nonan-2,5-dione | C8H12N2O2 | 24.362 | 168 | 2.27 | 3.13 |
| 7. | Cyclo(L-prolyl-L-valine) | C10H16N2O2 | 25.498 | 196 | 8.77 | 19.45 |
| 8. | Pyrrolo[1,2-a]pyrazine-1,4-dione | C11H18N2O2 | 26.020 | 210 | 1.18 | 2.91 |
| 9. | L-Leucine | C25H47NO3 | 27.036 | 409 | 3.18 | 7.31 |
| 10. | Pyrrolo[1,2-a]pyrazine-1,4-dione | C11H18N2O2 | 27.356 | 210 | 4.01 | 7.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



