
Submitted:
25 April 2024
Posted:
26 April 2024
You are already at the latest version
Abstract
Globally, habitat fragmentation has increased the proximity between wildlife, humans, and emerging predators such as free-ranging dogs. In these fragmented landscapes, encounters between primates and dogs are escalating, with primates often falling victim to dog attacks while navigating patchy landscapes and fragmented forests. We aim to investigate how these primates deal with the simultaneous threats posed by humans and predators, specifically focusing on the adaptive strategies of Central Himalayan Langur (CHL) in the landscape of fear. To address this, we conducted a behavioral study on the CHL in an agro-forest landscape, studying them for a total of 3912 hours over two consecutive years. Our results indicate that, compared to their most common resting behavior, CHL allocate more time to feeding and locomotion, and less time to socializing in the presence of humans and predatory dogs. Additionally, they exhibit increased feeding and locomotion and reduced social behavior in agro-forest or open habitats. These behavioral patterns reflect adaptive responses to the landscape of fear, where the presence of predators significantly influences their behavior and resource utilization. This study suggests measures to promote coexistence between humans and wildlife through the integration of effective management strategies that incorporate both ecological and social dimensions of human-wildlife interactions.
Keywords:
human-induced rapid environmental change
; human-wildlife interactions
; landscape of fear
; predator-prey systems
; resource availability
1. Introduction
Dogs (Canis familiaris) are globally the most widespread human-introduced carnivores [1]. Originating roughly 14,000 years ago [2], the dog population grew alongside the human population, with estimates nearing a billion worldwide [3]. Through their long-term association with humans, dogs have become integral members of human society, fulfilling various roles, such as pets, protectors, and guards for agriculture and livestock [4,5]. Across the globe, dogs exhibit varying degrees of free-ranging behavior, bringing them into interactions with wildlife on multiple levels [1]. Particularly in developing nations, the threat of free-ranging dogs escalates as human populations expand into natural areas in search of suitable agricultural land and natural resources [6]. Despite their long history as human companions and workers, the impact of dogs on wildlife hinges on their management practices, whether they are fully domesticated and leashed, roam freely, rely on human-provided food or lead independent lives. Additionally, the proximity of dogs to natural reserves or urban pockets of wildlife plays a significant role in the frequency and nature of interactions with wildlife [6,7].
Despite their morphological differences from wolves due to domestication, dogs retain characteristics of the carnivore guild [1,4]. In certain regions, dogs have emerged as the most abundant carnivores, disrupting ecosystems [8] and negatively impacting wildlife. Dog behaviors, including direct predation, harassment, and competition can lead to fear-induced behavioral changes in wildlife and affect their resource base and ultimately their fitness. In addition, the mere presence of dogs can lead to disease transmission [9,10,11]. Prominent review articles provide insightful analyses of the complex dynamics of human-dog-wildlife interactions, highlighting the need for effective management strategies to mitigate adverse effects on biodiversity [1,4,7,14,15]. However, there remains a need to understand how dogs are increasingly impacting individual species of wildlife as humans expand their agricultural areas.
Primate species represent one of the most diverse groups of mammals, totaling 504 species [14]. Unfortunately, approximately 65% of these species are currently under threat of extinction due to habitat loss and resource depletion [15]. Primate habitats are becoming significantly fragmented as a result of rising human demand for agricultural land, industrial growth, and the extraction of natural resources (e.g., mining, fuelwood), [16,17]. Tropical and subtropical forests, which are home to a large diversity of primates worldwide, have over 50% of their habitats fragmented [16]. In these fragmented landscapes, encounters between primates and invasive species, notably dogs, are growing, as primates often fall victim to dog attacks while navigating patchy landscapes and fragmented forests [19,20]. For example, dogs have been hunting and eating white-tailed small-eared Galagos (Otolemur garnettii lasiotis) in Kenya's fragmented landscapes [19]. Similarly, in Madagascar, dogs have also been observed preying on wild ring-tailed lemurs (Lemur catta) [20], and similar incidents involving langurs and macaques have been reported from comparable landscapes in Asia [21,22,23,24]. The presence of predators, including dogs, profoundly influences primate behavior and activity patterns. Primates, like many other species, have adapted to minimize the risk of predation through behavioral strategies such as alarm calls, vigilance, and counter-attacking [26,27]. Studies conducted in various ecosystems have demonstrated how the perceived risk of predation alters vigilance behavior [27], movement patterns [28], foraging strategies [29], and social dynamics [30].
Anthropogenic impacts on animal behavior result from both direct human disturbances, like fragmentation, and indirect disturbances, such as the introduction of alien species [31]. The initial reactions of animals to these changes are often behavioral, helping them adapt to new conditions [32,33,34]. Primates living in anthropogenically disturbed areas regularly show changes in habitat use, foraging, and activity budget [36,37]. For instance, chimpanzees (Pan troglodytes verus) in Guinea, West Africa, demonstrated a preference for traveling, resting, and socializing in habitat types with lower levels of human-induced pressure [37]. Compared to natural processes, anthropogenic pressures expose animals to environments they have not experienced in their evolutionary history, and behavioral changes are occurring at faster rates [38]. Even though these behavioral adaptations might have short-term benefits, they may also disrupt other long-evolved, fitness-related behaviors (e.g., social structure, reproductive strategies), which could negatively impact species persistence and have cascading effects on ecosystems [35,40].
Shared agroforest environments offer a complex web of opportunities and costs to wildlife. Our study species, Central Himalayan Langurs (CHL) inhabit Himalayan high-altitude agroforest landscapes. CHL dwell within fragmented natural oak forests and rely on seasonal crops (May-June and September-October) as well as on the leaves of fodder trees that grow along the edges of agricultural fields year-round [41,42]. However, foraging within agricultural fields exposes the langurs to their main predator, feral dogs [20,43], and increases the likelihood of frequent negative encounters with local people [40,43]. Predator presence can create a ‘landscape of fear’, and can influence the behavior and distribution of prey species within an ecosystem. Thus, ecological dynamics for CHL in such agroforest landscapes are not solely driven by the availability of resources, but also by the fear of predation and the strategies animals employ to mitigate that risk [45,46,47]. By understanding how animals adapt their behavior to minimize risks while maximizing opportunities in shared landscapes, we can develop more effective approaches for managing human-wildlife interactions and fostering sustainable coexistence.
While a growing number of case studies describe behavioral responses to human-induced rapid environmental change in specific systems [46], our understanding of such changes due to predatory feral dogs remains limited. We know that the presence of dogs can impact the activity of wild animals [3,7,13,49], but how animals respond to such changes has not been previously addressed, to our knowledge. We investigate how the fear landscape created by humans and predators affects the activity patterns of CHL in agroforest environments. By investigating how these primates deal with the simultaneous threats of humans and predators, we aim to understand more about the complex structure of fear behavior and how it impacts interactions between humans and wildlife. We hypothesized that H1: The predator-induced landscape of fear will have an impact on the activity patterns of CHL; H2: The human-induced landscape of fear will have an impact on the activity patterns of CHL; H3: CHL will modify their behavior in the habitat associated with these landscapes of fear. We predicted that in the landscapes of fear induced by predators and human activities, CHL will prioritize moving and feeding behaviors over socializing to maximize their fitness.
2. Materials and Methods
2.1. Study site and Population
The study was conducted from May 2022 to December 2023 in Mandal Valley, Chamoli, Uttarakhand, Garhwal Himalayas, India (coordinates: 30°28'04"N 79°16'31"E, elevation: 1500–1800 meters above sea level). The habituation process for the study group "S Group" began in 2014. Since then, this group has remained the subject of continuous study up to the present day. The total home range of the S group encompasses both natural forests and agricultural fields, frequently bringing them into proximity with humans and predatory domestic dogs (Figure 1). Every member of the group, as well as each newcomer, was individually identified and monitored throughout the duration of the study. During the observation time for this study (May 2022 to December 2023), the group consisted of 64 individuals, comprising 12 infants, nine juveniles, 16 subadults, 20 adult females, and seven adult males. For further information about the study site, refer to [40,42,48,49]
2.2. Behavioral Data Collection
Data were collected on 43 adult and subadult (age more than 3 years) langurs of both sexes for two consecutive years. Data collection was carried out over eight months in 2022 (May to December) and ten months in 2023 (January to December). The S Group was tracked from 06:00 to 18:00 by three observers (the authors, HN, VM, and a field assistant) for a total of 326 days, averaging 20 days per month. Three time each observation hour (i.e., every 20-minutes), we conducted an instantaneous scan sample that could last up to 15 minutes [50], for a total of 3912 hours of observation—240 hours on average per month (2,052 hours in 2022 and 1,860 hours in 2023). All state behaviors of the visible S Group individuals were recorded during the 15-minute scan, noting the activity that individuals were engaged in when first spotted. The dogs were fully habituated to the presence of researchers, and we documented both dog and human presence with each scan. If dogs and humans were within approximately 10 meters of any S Group individual during a particular scan, they were noted as present; otherwise, they were considered absent. Habitat-type codes were recorded for each scan sample, and further details of these codes are presented in the Supplementary Materials.
2.3. Behavioral Definitions
Using the comprehensive ethogram outlined by Dolhinow [51], we categorized langur behavior into four broad state behaviors, as detailed below:
- Feeding: When an animal is ingesting or masticating food.
- Locomotion: An animal moves, at any pace, from one location to another, over a distance of more than 1 meter.
- Resting: An animal remains stationary, hindquarters on the supporting surface, either asleep (i.e., eyes closed) or awake (i.e., alert, with eyes open). It may simultaneously engage in l vigilance behavior.
- Social: Social interactions like grooming, play, or sexual activity.
2.4. Study Variables
Presence of predatory dog(s) In all the models, a binary value of this variable is used to denote whether one or more dogs is present within a 10-meter radius of the S Group or not. These free-roaming dogs may belong to villagers or be stray dogs within the S Group's territory. Presence of human(s): In all the models, a binary value of this variable is used to denote whether one or more humans is present within a 10-meter radius of the S Group or not. Humans usually include villagers or occasional tourists within the S Group's territory. Habitat: The study group's habitat encompasses various areas, including dense forest patches, open patches near human settlements, and agricultural fields (Figure 1). Initially, the entire study area was categorized based on location and vegetation, which were later combined into two forest types: open forests and dense forests across all models. Further details regarding these classifications are presented in Supplementary Tables S1 and S2.
2.5. Model Formulation and Statistical Analysis
To investigate the potential impact of our study variables on the four major behaviors of CHL, we conducted a Multinomial Logistic Regression (MLR) analysis using the nnet package [54,55] in R version 4.2.2 [54]. In MLR, the log odds provide an estimate of the relationship between predictor variables (habitat, presence of dogs, presence of humans) and the probabilities of each categorical outcome variable (the four types of behavior) in comparison to the reference category. Specifically, we used the log odds of feeding, locomotion, and social behavior relative to resting behavior as the baseline for all models. Selection of the reference category should be driven by theoretical or substantive considerations [55]. Thus, we use resting behavior as the reference category, given it’s a dominant behavior in colobines for digesting their folivorous diet [56]. Additionally, selecting a reference category with a substantial number of observations can enhance the precision of estimates [57].
MLR serves as an extension of binary logistic regression, accommodating more than two categories of dependent variables. MLR, like binary logistic regression, models the log odds of the outcomes as a linear combination of the predictor variables and employs maximum likelihood estimation to assess the probability of categorical membership, generating a unique set of parameters for each level of the dependent variable, referred to as regression coefficients (log odds) [58]. To understand how different factors affect the likelihood of each category of the outcome variable, we identified three predictor variables, Presence of Dog, Presence of Human, and Habitat. We constructed eight potential models based on these variables and compared them based on Akaike's information criteria (AIC), we applied information theory (IT) to identify the best model [59,60,61,62]. This method's fundamental principle is to use ΔAICc values to rank the model among the candidate set of models (Δi = AICc(i) - AICc (min)) [59]. Standardized model weights express the probability that a given model is the best among those in the set of models (Table 1). With the "aictab" function from the AICcmodavg package, we assessed the Akaike's weight, or relative likelihood, and the cumulative weight of each model [63]. These metrics indicate the degree to which one model is more likely than another in explaining the variance in the data. AIC is the preferred method for model selection in ecology and evolution due to its effectiveness in addressing methodological challenges [64,65,66]. This method has been widely adopted in primate behavior studies across various species, including Japanese macaques [67] (Macaca fuscata fuscata), vervet monkeys [68] (Chlorocebus pygerythrus), chimpanzees [69] (Pan troglodytes schweinfurthii), black howler monkeys [70] (Alouatta pigra), and mountain gorillas [71] (Gorilla beringei beringei).
3. Results
3.1. Selection of Activity Pattern
Among the eight models that were tested, the integrated model emerged as the most accurate in explaining the factors influencing the activity patterns of CHL. This model demonstrated the lowest AICc value and the highest model probability, with a weight of 1 (as shown in Table 1). The integrated model incorporates all independent variables (dog presence, human presence, and habitat type). In contrast, the remaining seven models, including the null model (as presented in Table 1), collectively exhibited a probability of less than 0.01.
In the presence of predatory dogs, the log odds of adult CHL engaging in feeding activities over resting activities was 0.016, whereas for locomotion and social activities, it was 0.156 and -0.386, respectively (see Table 2). Free ranging dogs significantly influenced the activity patterns of adult langurs, resulting increased feeding and locomotion but decreased social activity when dogs were present in their habitat (see Figure 2a, Table 2, z=0.601, z=6.135, z=-11.963; p<0.01).
Similarly, in the presence of humans, the log odds of adult CHL engaging in feeding activities over resting activities was 0.288, whereas for locomotion and social activities, it was 0.202 and -0.163, respectively (see Table 2). Adult langurs exhibited increased feeding and locomotion but decreased social activity when humans were present (Figure 2b, Table 2, z=11.25, z=8.122, z=-5.407; p<0.01).
Furthermore, in open habitats, the log odds of adult CHL engaging in feeding activities over resting activities was 0.585, whereas for locomotion and social activities, it was 0.106 and -0.492, respectively (see Table 2). Habitat type significantly influenced the activity patterns of adult langurs, resulting in increased feeding and locomotion but decreased social activity in open habitat (Figure 2c, Table 2, z=25.641, z=4.717, z=-17.872; p<0.01).
4. Discussion
4.1. Behavior Modifications in the Predator-Induced Landscape of Fear
Our results demonstrate that the presence of predatory dogs causes CHL to alter their behavior, increasing their feeding and movement activity while lowering their social interaction. Colobine populations can differ in feeding and resting patterns and these differences are more likely to be a result of behavioral adaptations than a by-product of their shared evolutionary history [72]. By allocating maximum time to resting, colobines optimize their digestive efficiency and maximize the extraction of nutrients from their folivorous diet [56]. Under the landscape of fear, the presence of predators can shape the behavior and distribution of prey species even when predation itself doesn't occur [73,74,75]. Adjusting the allocation of foraging time is a common strategy for animals to mitigate the risk of predation [76]. Because animals respond to predators through complex, compensatory behavioral strategies, it is difficult to predict how these behaviors will ultimately impact fitness [77]. Prey species facing higher levels of predation risk may allocate more energy toward developing and maintaining anti-predator defences or reducing other energetically expensive activities [56,57].
The natural Banj oak (Quercus leucotrichophora) forest, which is the preferred habitat for langurs [41], has become fragmented, due to the high dependency of local people on the surrounding natural forest [52,53]. Langurs are increasingly dedicating more time to feeding on fodder crops in agricultural land, primarily due to the fragmentation of their preferred habitat [41,42,44]. Our results suggest that the differences in resource availability in modified agricultural landscapes relative to their preferred oak forests, leads to greater feeding and moving, requiring them to expend more energy to forage and navigate their surroundings. These areas also do not provide many safe areas to rest and socialize. Based on 312 langur-dog interactions, all successful predatory attacks by dogs on S Group individuals occurred in agricultural fields, where langurs are notably vulnerable due to the lack of connected tree cover, providing a low chance of escape [20,43]. Our results suggest that despite the threat of predation, CHL increase their feeding time in predator-rich environments because these areas provide crucial resources for them. Similar behavior is also observed among herbivores in Africa, due to the distribution of grasslands and waterholes, grazers often have no choice but to utilize habitats located near the few available waterholes and grasslands, despite the high predation risk associated with these areas [82]. This behavioral adjustment reflects their prioritization of foraging and movement over social activities to maximize their fitness. Additionally, CHL employ high-cost anti-predator strategies such as giving alarm calls and engaging in fights with predators to mitigate the risk of predation [18]. Overall, our findings highlight the complex interactions between habitat fragmentation, foraging behavior, and predation risk in langurs, emphasizing their adaptive responses to changes in their environment to achieve optimal fitness.
4.2. Behavior Modifications in the Human-Induced Landscape of Fear
Similar to the presence of predatory dogs, the presence of humans caused CHL to alter their behavior, increasing their feeding and movement activity while lowering their social interaction. This effect is partially spatial, as langurs are attempting to forage on the crops, and moving into agricultural areas where they are likely to encounter humans. Human presence can have significant effects on the behavior of various animals, for example changing the foraging and vigilance behavior of elk (Cervus elephus) [83], and Nubian ibex (Capra nubiana) [84]. Human activities such as hunting, habitat modification, and disturbance can create perceived threats for wildlife, for example, in the case of red deer (Cervus elaphus) and wild boar (Sus scrofa), human presence can influence their use of habitat and behavior [44]. The livelihood of local villagers in this region depends on agro-forestry crops [85] such as Grewia optiva, Celtis australis, and Ficus virens, which are also important parts of the CHL diet [48]. In response to the perceived threat posed by langurs to their crops and livelihoods, villagers engage in various negative interactions with the langurs, including throwing stones, using loud noises and firecrackers to chase them away, and even resorting to poisoning [41]. Human behavior and tolerance toward wildlife play crucial roles in shaping the landscape of fear in human-influenced environments [86]. In agroforest contexts in Africa, the economic value of cultivated food crops and the extent of damage caused by chimpanzees (Pan troglodytes) can significantly influence human attitudes, tolerance, and behavior toward them [87].
Human tolerance toward CHL can also influence the effectiveness of management and conservation efforts [41]. The negative attitude of the local community towards langurs is due to the langurs' frequent consumption of their agricultural and fodder crops. To promote tolerance, it is critical to address the livelihood challenges faced by the local community. By integrating the livelihood needs of local communities into mitigation models, we can enhance their value perception of wildlife. This, in turn, facilitates community support for mitigating human-wildlife interactions through educational initiatives, habitat restoration, and promotion of coexistence. Our findings contribute towards the understanding of how human behavior and tolerance affect the spatial distribution and CHL behavior (i.e., the landscape of fear). This evidence can inform land-use planning and management strategies, such as the identification and establishment of corridors within the territory of the S Group. Consequently, we recommend integrating effective management strategies incorporating ecological and social dimensions of human-wildlife interactions. We can develop comprehensive and socially acceptable interventions by considering factors such as human behavior and tolerance alongside ecological variables like species ecology, population dynamics, and habitat preferences.
4.3. Behavior Modifications in the Habitats Associated with the Landscape of Fear
We found that CHL exhibits an increase in feeding and locomotion activity and a decrease in social interaction and resting in open forest habitats. Primates in fragmented landscapes spend more time feeding to compensate for limited resources [88]. The home range of S Group includes fragmented patches of dense forest, agricultural land, and human settlements, and they often move between the forest and agricultural fields for crop foraging [41]. Interactions with dogs typically occur in open forest patches or agricultural fields where continuous canopy cover is absent [18,42]. The fragmentation and loss of habitats is a major factor leading to the decline of many species, partly due to increases in predation by human-introduced predators [33]. Primates show changes in their activity budget, foraging and ranging in anthropogenically disturbed environments; for example, chimpanzees (Pan troglodytes verus) preferred to rest, travel, and socialize in habitats with less human activity [89]. In anthropogenically disturbed areas, vervet monkeys spend significantly less time socializing and more time engaged in movement [90]. Similarly, toque macaques (Macaca sinica) in human-modified habitats allocated more time moving and vigilance and less time resting [91]. Based on the evidence presented here, we propose that the availability of food resources in open habitats plays a crucial role in shaping the behavior and ecology of CHL; specifically, by increasing feeding and locomotion in decreasing resting and social interactions. Future studies focusing on the relationship between food availability and primate behavior in agroforest landscapes have the potential to generate valuable insights that can contribute to conservation, management, and coexistence strategies. Identifying tree species and landscape features that sustain primate food sources while preserving crop yields can help agroforestry management practices establish a balance between agricultural productivity and biodiversity conservation efforts.
5. Conclusion
Our findings highlight that CHL allocates more time to feeding and locomotion and less time to socializing in the presence of humans and predatory dogs. This behavioral shift is anticipated as an adaptive strategy aimed at increasing individual fitness in environments where predation risk and human disturbances are prevalent. These behaviors demonstrate their adaptive responses to the landscape of fear, where the mere presence of predators influences their behavior and resource utilization. Overall, our findings provide valuable insights into how CHL adapt their behavior to deal with threats from humans and predators, ultimately contributing to our understanding of wildlife responses to anthropogenic pressures in shared habitats. In the long run, this adaptation may lead to a shift in CHL habitat use patterns, social dynamics, and foraging behavior. Therefore, we propose the following practices. 1- Create safe corridors and protected areas where CHL can access essential resources without facing significant threats from humans or dogs. 2- Conduct comprehensive research to gain deeper insights into CHL social structures, behavior, and dynamics. 3- Engage local stakeholders in collaborative wildlife management and conflict resolution efforts. By implementing these measures, we may promote coexistence between humans and wildlife while aiming to mitigate the long-term impact of human disturbances and predation risk on wildlife populations. Overall, by establishing partnerships with communities, conservation organizations can promote sustainable practices, raise conservation awareness, and empower local communities to take ownership of conservation initiatives.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, H.N., VN., and K.H.G.; methodology, H.N.; software, H.N., and K.H.G; formal analysis, H.N.; investigation, H.N., and V.M; resources, H.N., and J.T; data curation, H.N., VN., and K.H.G.; writing—original draft preparation, H.N., K.H.G.; writing—review and editing, H.N., VN., K.H.G., J.T., D.S., and R.D; visualization, H.N., V.M., and D.S.; supervision, J.T., D.S., and R.D.; funding acquisition, H.N., V.M., and J.T. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was funded by Rufford Small Grant, grant number 37193-B and University of Toronto field research funding.
Institutional Review Board Statement
Our study adhered to Indian wildlife protection laws, complied with the principles for the Ethical Treatment of Non-human Primates set forth by the American Society of Primatologists, and followed the Code of Best Practices for Field Primatology as outlined by the American Society of Primatologists. The entire data collection process was noninvasive, and we conducted our research in close collaboration with Indian Forest authorities and the National Biodiversity Board.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. The datasets analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We acknowledge Dr. Anindya Sinha and the National Institute of Advanced Studies, Bengaluru, India, for their constant support and assistance in acquiring the necessary permits to conduct the study in India. We are grateful to the Forest Department of Uttarakhand for permitting us to do research in the Kedarnath Wildlife Sanctuary. Lastly, we warmly thank our field assistants, Mr. Suraj, Mr. Harish, Mr. Mohit and research assistant Mr. Diganta, Mr. Soumalya, Ms. Simran, and Mr. Ryan for their unwavering support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vanak, A.T.; Gompper, M.E. Dogs Canis Familiaris as Carnivores: Their Role and Function in Intraguild Competition. Mammal Rev. 2009, 39, 265–283. [Google Scholar] [CrossRef]
- Vila, C. Phylogenetic Relationships, Evolution, and Genetic Diversity of the Domestic Dog. J. Hered. 1999, 90, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Gompper, M.E. Free-Ranging Dogs and Wildlife Conservation; 1st ed.; Oxford university press: Oxford (GB), 2014; ISBN 978-0-19-966321-7. [Google Scholar]
- Hughes, J.; Macdonald, D.W. A Review of the Interactions between Free-Roaming Domestic Dogs and Wildlife. Biol. Conserv. 2013, 157, 341–351. [Google Scholar] [CrossRef]
- Driscoll, C.A.; Macdonald, D.W. Top Dogs: Wolf Domestication and Wealth. J. Biol. 2010, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, M.A.; Singer, R.S.; Silva-Rodríguez, E.; Stowhas, P.; Pelican, K. Domestic Dogs in Rural Communities around Protected Areas: Conservation Problem or Conflict Solution? PLOS ONE 2014, 9, e86152. [Google Scholar] [CrossRef] [PubMed]
- Home, C.; Bhatnagar, Y.V.; Vanak, A.T. Canine Conundrum: Domestic Dogs as an Invasive Species and Their Impacts on Wildlife in India. Anim. Conserv. 2018, 21, 275–282. [Google Scholar] [CrossRef]
- Feldmann, B.M. The Problem of Urban Dogs. Science 1974, 185, 903–903. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, E.G.; Dickman, C.R.; Letnic, M.; Vanak, A.T. Dogs as Predators and Trophic Regulators. In Free-Ranging Dogs and Wildlife Conservation; Gompper, M.E., Ed.; Oxford University Press, 2013; pp. 55–68 ISBN 978-0-19-966321-7.
- Banks, P.B.; Bryant, J.V. Four-Legged Friend or Foe? Dog Walking Displaces Native Birds from Natural Areas. Biol. Lett. 2007, 3, 611–613. [Google Scholar] [CrossRef]
- Doherty, T.S.; Dickman, C.R.; Glen, A.S.; Newsome, T.M.; Nimmo, D.G.; Ritchie, E.G.; Vanak, A.T.; Wirsing, A.J. The Global Impacts of Domestic Dogs on Threatened Vertebrates. Biol. Conserv. 2017, 210, 56–59. [Google Scholar] [CrossRef]
- Young, J.K.; Olson, K.A.; Reading, R.P.; Amgalanbaatar, S.; Berger, J. Is Wildlife Going to the Dogs? Impacts of Feral and Free-Roaming Dogs on Wildlife Populations. BioScience 2011, 61, 125–132. [Google Scholar] [CrossRef]
- Gompper, M.E. Adding Nuance to Our Understanding of Dog–Wildlife Interactions and the Need for Management. Integr. Comp. Biol. 2021, 61, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Fernández, D.; Kerhoas, D.; Dempsey, A.; Billany, J.; McCabe, G.; Argirova, E. The Current Status of the World’s Primates: Mapping Threats to Understand Priorities for Primate Conservation. Int. J. Primatol. 2022, 43, 15–39. [Google Scholar] [CrossRef] [PubMed]
- Estrada, A.; Garber, P.A.; Chaudhary, A. Current and Future Trends in Socio-Economic, Demographic and Governance Factors Affecting Global Primate Conservation. PeerJ 2020, 8, e9816. [Google Scholar] [CrossRef] [PubMed]
- Estrada, A.; Garber, P.A.; Rylands, A.B.; Roos, C.; Fernandez-Duque, E.; Di Fiore, A.; Nekaris, K.A.-I.; Nijman, V.; Heymann, E.W.; Lambert, J.E.; et al. Impending Extinction Crisis of the World’s Primates: Why Primates Matter. Sci. Adv. 2017, 3, e1600946. [Google Scholar] [CrossRef] [PubMed]
- Waters, S.; Watson, T.; Farris, Z.J.; Bornbusch, S.; Valenta, K.; Clarke, T.A.; Chetry, D.; Randriana, Z.; Owen, J.R.; El Harrad, A.; et al. Dogs, Primates, and People: A Review. In Primates in Anthropogenic Landscapes; McKinney, T., Waters, S., Rodrigues, M.A., Eds.; Developments in Primatology: Progress and Prospects; Springer International Publishing: Cham, 2023; ISBN 978-3-031-11735-0. [Google Scholar]
- Nautiyal, H.; Tanaka, H.; Huffman, M.A. Anti-Predator Strategies of Adult Male Central Himalayan Langurs (Semnopithecus schistaceus) in Response to Domestic Dogs in a Human-Dominated Landscape. Primates 2023, 64, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Pihlström, H.; Rosti, H.; Lombo, B.M.; Pellikka, P. Domestic Dog Predation on White-Tailed Small-Eared Galago (Otolemur garnettii lasiotis) in the Taita Hills, Kenya.
- Moresco, A.; Larsen, R.S.; Sauther, M.L.; Cuozzo, F.P.; Jacky, A.Y.; Millette, J.B. Survival of a Wild Ring-Tailed Lemur (Lemur catta) with Abdominal Trauma in an Anthropogenically Disturbed Habitat. Madag. Conserv. Dev. 2012, 7, 49–52. [Google Scholar] [CrossRef]
- Knight, J. Herding Monkeys to Paradise: How Macaque Troops Are Managed for Tourism in Japan. In Herding Monkeys to Paradise; Brill, 2011.
- Najmuddin, M.F.; Haris, H.; Norazlimi, N.; Md-Zain, M.; Mohd-Ridwan, A.R.; Shahrool-Anuar, R.; Husna, H.A.; Abdul-Latiff, M.A.B. Predation of Domestic Dogs (Canis lupus familiaris) on Schlegel’s Banded Langur (Presbytis neglectus) and Crested Hawk-Eagle (Nisaetus cirrhatus) on Dusky Leaf Monkey (Trachypithecus obscurus) In Malaysia. J. S.S.M. 2019, 6, 39–50. [Google Scholar]
- Bishop, N.; Hrdy, S.B.; Teas, J.; Moore, J. Measures of Human Influence in Habitats of South Asian Monkeys. Int. J. Primatol. 1981, 2, 153–167. [Google Scholar] [CrossRef]
- Chakraborty, B.; Pithva, K.; Mohanty, S.; McCowan, B. Lethal Dog Attacks on Adult Rhesus Macaques (Macaca Mulatta) in an Anthropogenic Landscape. Primates 2024, 65, 151–157. [Google Scholar] [CrossRef]
- Isbell, L.A. Predation on Primates: Ecological Patterns and Evolutionary Consequences. Evol. Anthropol. Issues News Rev. 1994, 3, 61–71. [Google Scholar] [CrossRef]
- Hart, D. Predation on Primates: A Biogeographical Analysis. In Primate Anti-Predator Strategies; Gursky, S.L., Nekaris, K.A.I., Eds.; Springer US: Boston, MA, 2007; ISBN 978-0-387-34810-0. [Google Scholar]
- Campos, F.A.; Fedigan, L.M. Spatial Ecology of Perceived Predation Risk and Vigilance Behavior in White-Faced Capuchins. Behav. Ecol. 2014, 25, 477–486. [Google Scholar] [CrossRef]
- Willems, E.P.; Hill, R.A. Predator-Specific Landscapes of Fear and Resource Distribution: Effects on Spatial Range Use. Ecology 2009, 90, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Cowlishaw, G. Trade-Offs between Foraging and Predation Risk Determine Habitat Use in a Desert Baboon Population. Anim. Behav. 1997, 53, 667–686. [Google Scholar] [CrossRef]
- Isbell, L.A. Predation on Primates: Ecological Patterns and Evolutionary Consequences. Evol. Anthropol. Issues News Rev. 1994, 3, 61–71. [Google Scholar] [CrossRef]
- Robertson, B.A.; Hutto, R.L. A Framework for Understanding Ecological Traps and an Evaluation of Existing Evidence. Ecology 2006, 87, 1075–1085. [Google Scholar] [CrossRef] [PubMed]
- Wong, B.B.M.; Candolin, U. Behavioral Responses to Changing Environments. Behav. Ecol. 2015, 26, 665–673. [Google Scholar] [CrossRef]
- Sih, A.; Ferrari, M.C.O.; Harris, D.J. Evolution and Behavioural Responses to Human-Induced Rapid Environmental Change. Evol. Appl. 2011, 4, 367–387. [Google Scholar] [CrossRef] [PubMed]
- Tuomainen, U.; Candolin, U. Behavioural Responses to Human-Induced Environmental Change. Biol. Rev. 2011, 86, 640–657. [Google Scholar] [CrossRef]
- Bryson-Morrison, N.; Tzanopoulos, J.; Matsuzawa, T.; Humle, T. Activity and Habitat Use of Chimpanzees (Pan troglodytes verus) in the Anthropogenic Landscape of Bossou, Guinea, West Africa. Int. J. Primatol. 2017, 38, 282–302. [Google Scholar] [CrossRef]
- Negrín, A.R.; Fuentes, A.C.; Espinosa, D.C.; Dias, P.A.D. The Loss of Behavioral Diversity as a Consequence of Anthropogenic Habitat Disturbance: The Social Interactions of Black Howler Monkeys. Primates 2016, 57, 9–15. [Google Scholar] [CrossRef]
- McKinney, T. A Classification System for Describing Anthropogenic Influence on Nonhuman Primate Populations. Am. J. Primatol. 2015, 77, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Sih, A. Understanding Variation in Behavioural Responses to Human-Induced Rapid Environmental Change: A Conceptual Overview. Anim. Behav. 2013, 85, 1077–1088. [Google Scholar] [CrossRef]
- Ehrlich, P.R. Intervening in Evolution: Ethics and Actions. Proc. Natl. Acad. Sci. 2001, 98, 5477–5480. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, H. Behavioral Ecology of the Central Himalayan Langur (Semnopithecus schistaceus) in a Human Dominated Landscape: Multi-Species Interactions and Conservation Implications, Kyoto University, 2020.
- Nautiyal, H.; Mathur, V.; Sinha, A.; Huffman, M.A. The Banj Oak Quercus leucotrichophora as a Potential Mitigating Factor for Human-Langur Interactions in the Garhwal Himalayas, India: People’s Perceptions and Ecological Importance. Glob. Ecol. Conserv. 2020, 22, e00985. [Google Scholar] [CrossRef]
- Gajare, K.H. Interaction between Feral Dogs and Central Himalayan Langur (Semnopithecus schistaceus ), Mumbai university, 2023.
- Mathur, V.; Teichroeb, J.; Nautiyal, H. Understanding Variability In Crop Foraging By Himalayan Langurs (Semnopithecus schistacesus) And Other Wildlife Species Using Direct Observation And Camera Traps In An Anthropogenic Landscape. In American Socity Of Primatology Conference, Reno, USA, 21/06/2023.
- Theuerkauf, J.; Rouys, S. Habitat Selection by Ungulates in Relation to Predation Risk by Wolves and Humans in the Białowieża Forest, Poland. For. Ecol. Manag. 2008, 256, 1325–1332. [Google Scholar] [CrossRef]
- Nowak, K.; Wimberger, K.; Richards, S.A.; Hill, R.A.; le Roux, A. Samango Monkeys (Cercopithecus albogularis labiatus) Manage Risk in a Highly Seasonal, Human-Modified Landscape in Amathole Mountains, South Africa. Int. J. Primatol. 2017, 38, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Candolin, U.; Wong, B.B.M. Behavioural Responses to a Changing World: Mechanisms and Consequences; Oxford University Press, 2012; ISBN 978-0-19-960256-8.
- Lenth, B.; Knight, R.; Brennan, M. The Effects of Dogs on Wildlife Communities. Nat. Areas J. 2008, 28, 218–227. [Google Scholar] [CrossRef]
- Nautiyal, H. Life in the Extreme: Time-Activity Budgets and Foraging Ecology of Central Himalayan Langur (Semnopithecus schistaceus) in the Kedarnath Wildlife Sanctuary, Uttarakhand India, Bharathidasan University, 2015.
- Mathur, V. Choice and Characteristics of Sleeping Sites in a Troop of Central Himalayan Langurs (Semnopithecus schistaceus), Indian Institute of Science Education and Research Mohali, 2019.
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef]
- Dolhinow, P. A Behavior Repertoire for the Indian Langur Monkey (Presbytis entellus). Primates 1978, 19, 449–472. [Google Scholar] [CrossRef]
- William Venables; Brian Ripley Modern Applied Statistics with S; New York.
- Ripley, B.; Venables, W. Nnet: Feed-Forward Neural Networks and Multinomial Log-Linear Models 2023.
- R A Language and Environment for Statistical Computing. 2022.
- Long, J.S.; Freese, J. Regression Models for Categorical Dependent Variables Using Stata, Second Edition; Stata Press, 2006; ISBN 978-1-59718-011-5.
- Davies, G.; Oates, J. Colobine Monkeys: Their Ecology, Behaviour and Evolution; Cambridge University Press, 1994; ISBN 978-0-521-33153-1.
- Agresti, A. Categorical Data Analysis; John Wiley & Sons, 2012; ISBN 978-0-470-46363-5.
- Anderson, C.J.; Rutkowski, L. Multinomial Logistic Regression. In Best Practices in Quantitative Methods; Osborne, J., Ed.; SAGE Publishing, 2008; pp. 390–409 ISBN 978-1-4129-4065-8.
- Advanced Issues and Deeper Insights. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Burnham, K.P., Anderson, D.R., Eds.; Springer: New York, NY, 2002; pp. 267–351. ISBN 978-0-387-22456-5. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC Model Selection and Multimodel Inference in Behavioral Ecology: Some Background, Observations, and Comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Symonds, M.R.E.; Moussalli, A. A Brief Guide to Model Selection, Multimodel Inference and Model Averaging in Behavioural Ecology Using Akaike’s Information Criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- Richards, S.A.; Whittingham, M.J.; Stephens, P.A. Model Selection and Model Averaging in Behavioural Ecology: The Utility of the IT-AIC Framework. Behav. Ecol. Sociobiol. 2011, 65, 77–89. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c) 2023.
- Rushton, S.P.; Ormerod, S.J.; Kerby, G. New Paradigms for Modelling Species Distributions? J. Appl. Ecol. 2004, 41, 193–200. [Google Scholar] [CrossRef]
- Whittingham, M.J.; Stephens, P.A.; Bradbury, R.B.; Freckleton, R.P. Why Do We Still Use Stepwise Modelling in Ecology and Behaviour? J. Anim. Ecol. 2006, 75, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.B.; Omland, K.S. Model Selection in Ecology and Evolution. Trends Ecol. Evol. 2004, 19, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Duboscq, J.; Romano, V.; Sueur, C.; MacIntosh, A.J.J. Scratch That Itch: Revisiting Links between Self-Directed Behaviour and Parasitological, Social and Environmental Factors in a Free-Ranging Primate. R. Soc. Open Sci. 2016, 3, 160571. [Google Scholar] [CrossRef] [PubMed]
- Mercier, S.; Neumann, C.; van de Waal, E.; Chollet, E.; Meric de Bellefon, J.; Zuberbühler, K. Vervet Monkeys Greet Adult Males during High-Risk Situations. Anim. Behav. 2017, 132, 229–245. [Google Scholar] [CrossRef]
- Lonsdorf, E.V.; Wilson, M.L.; Boehm, E.; Delaney-Soesman, J.; Grebey, T.; Murray, C.; Wellens, K.; Pusey, A.E. Why Chimpanzees Carry Dead Infants: An Empirical Assessment of Existing Hypotheses. R. Soc. Open Sci. 2020, 7, 200931. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Negrín, A.; Coyohua-Fuentes, A.; Chavira, R.; Canales-Espinosa, D.; Dias, P.A.D. Primates Living Outside Protected Habitats Are More Stressed: The Case of Black Howler Monkeys in the Yucatán Peninsula. PLOS ONE 2014, 9, e112329. [Google Scholar] [CrossRef]
- Galbany, J.; Muhire, T.; Vecellio, V.; Mudakikwa, A.; Nyiramana, A.; Cranfield, M.R.; Stoinski, T.S.; McFarlin, S.C. Incisor Tooth Wear and Age Determination in Mountain Gorillas from Volcanoes National Park, Rwanda. Am. J. Phys. Anthropol. 2018, 167, 930–935. [Google Scholar] [CrossRef]
- Kraus, J.B.; Strier, K.B. Geographic, Climatic, and Phylogenetic Drivers of Variation in Colobine Activity Budgets. Primates 2022, 63, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Khater, M.; Murariu, D.; Gras, R. Predation Risk Tradeoffs in Prey: Effects on Energy and Behaviour. Theor. Ecol. 2016, 9, 251–268. [Google Scholar] [CrossRef]
- Brown, J.S.; Laundré, J.W.; Gurung, M. The Ecology of Fear: Optimal Foraging, Game Theory, and Trophic Interactions. J. Mammal. 1999, 80, 385–399. [Google Scholar] [CrossRef]
- Gaynor, K.M.; Brown, J.S.; Middleton, A.D.; Power, M.E.; Brashares, J.S. Landscapes of Fear: Spatial Patterns of Risk Perception and Response. Trends Ecol. Evol. 2019, 34, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Kotler, B.P.; Brown, J.S.; Hasson, O. Factors Affecting Gerbil Foraging Behavior and Rates of Owl Predation. Ecology 1991, 72, 2249–2260. [Google Scholar] [CrossRef]
- Lind, J.; Cresswell, W. Determining the Fitness Consequences of Antipredation Behavior. Behav. Ecol. 2005, 16, 945–956. [Google Scholar] [CrossRef]
- Trussell, G.C.; Matassa, C.M.; Luttbeg, B. The Effects of Variable Predation Risk on Foraging and Growth: Less Risk Is Not Necessarily Better. Ecology 2011, 92, 1799–1806. [Google Scholar] [CrossRef]
- Wisenden, B.D. Female Convict Cichlids Adjust Gonadal Investment in Current Reproduction in Response to Relative Risk of Brood Predation. Can. J. Zool. 1993, 71, 252–256. [Google Scholar] [CrossRef]
- Sharma, C.M.; Gairola, S.; Ghildiyal, S.K.; Suyal, S. Forest Resource Use Patterns in Relation to Socioeconomic Status. Mt. Res. Dev. 2009, 29, 308–319. [Google Scholar] [CrossRef]
- Makino, Y. Lopping of Oaks in Central Himalaya, India. Mt. Res. Dev. 2011, 31, 35–44. [Google Scholar] [CrossRef]
- Valeix, M.; Loveridge, A.J.; Chamaillé-Jammes, S.; Davidson, Z.; Murindagomo, F.; Fritz, H.; Macdonald, D.W. Behavioral Adjustments of African Herbivores to Predation Risk by Lions: Spatiotemporal Variations Influence Habitat Use. Ecology 2009, 90, 23–30. [Google Scholar] [CrossRef]
- Ciuti, S.; Northrup, J.M.; Muhly, T.B.; Simi, S.; Musiani, M.; Pitt, J.A.; Boyce, M.S. Effects of Humans on Behaviour of Wildlife Exceed Those of Natural Predators in a Landscape of Fear. PLOS ONE 2012, 7, e50611. [Google Scholar] [CrossRef]
- Tadesse, S.A.; Kotler, B.P. Impact of Tourism on Nubian Ibex (Capra nubiana) Revealed through Assessment of Behavioral Indicators. Behav. Ecol. 2012, 23, 1257–1262. [Google Scholar] [CrossRef]
- Mahato, S.; Dasgupta, S.; Todaria, N.P.; Singh, V.P. Agroforestry Mapping and Characterization in Four Districts of Garhwal Himalaya. Energy Ecol. Environ. 2016, 1, 86–97. [Google Scholar] [CrossRef]
- Bersacola, E.; Hill, C.M.; Hockings, K.J. Chimpanzees Balance Resources and Risk in an Anthropogenic Landscape of Fear. Sci. Rep. 2021, 11, 4569. [Google Scholar] [CrossRef] [PubMed]
- McLennan, M.R.; Hill, C.M. Troublesome Neighbours: Changing Attitudes towards Chimpanzees (Pan troglodytes) in a Human-Dominated Landscape in Uganda. J. Nat. Conserv. 2012, 20, 219–227. [Google Scholar] [CrossRef]
- Huang, Z.; Yuan, P.; Huang, H.; Tang, X.; Xu, W.; Huang, C.; Zhou, Q. Effect of Habitat Fragmentation on Ranging Behavior of White-Headed Langurs in Limestone Forests in Southwest China. Primates 2017, 58, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Bryson-Morrison, N.; Tzanopoulos, J.; Matsuzawa, T.; Humle, T. Activity and Habitat Use of Chimpanzees (Pan troglodytes verus) in the Anthropogenic Landscape of Bossou, Guinea, West Africa. Int. J. Primatol. 2017, 38, 282–302. [Google Scholar] [CrossRef]
- Thatcher, H.R.; Downs, C.T.; Koyama, N.F. Anthropogenic Influences on the Time Budgets of Urban Vervet Monkeys. Landsc. Urban Plan. 2019, 181, 38–44. [Google Scholar] [CrossRef]
- Jayapali, U.; Perera, P.; Cresswell, J.; Dayawansa, N. Does Anthropogenic Influence on Habitats Alter the Activity Budget and Home Range Size of Toque Macaques (Macaca sinica)? Insight into the Human-Macaque Conflict. Trees For. People 2023, 13, 100412. [Google Scholar] [CrossRef]
Figure 1.
A study area map illustrating the spatial distribution of forested areas, villages, and agricultural land within the habitat of the S Group.
Figure 1.
A study area map illustrating the spatial distribution of forested areas, villages, and agricultural land within the habitat of the S Group.
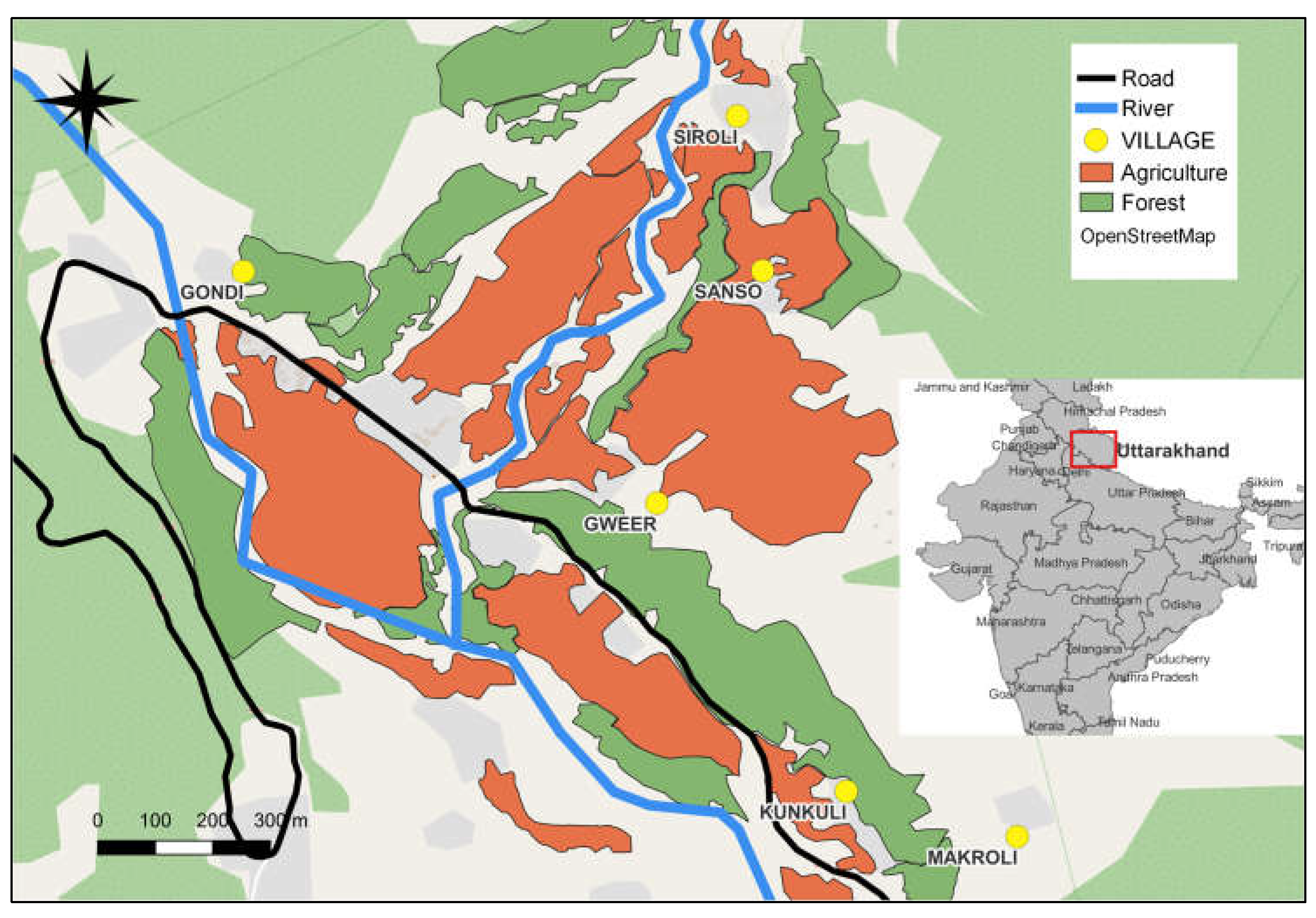
Figure 2.
Predictor variables a) presence of dog, b) presence of human, c) habitat type, in relation to activity pattern. .
Figure 2.
Predictor variables a) presence of dog, b) presence of human, c) habitat type, in relation to activity pattern. .
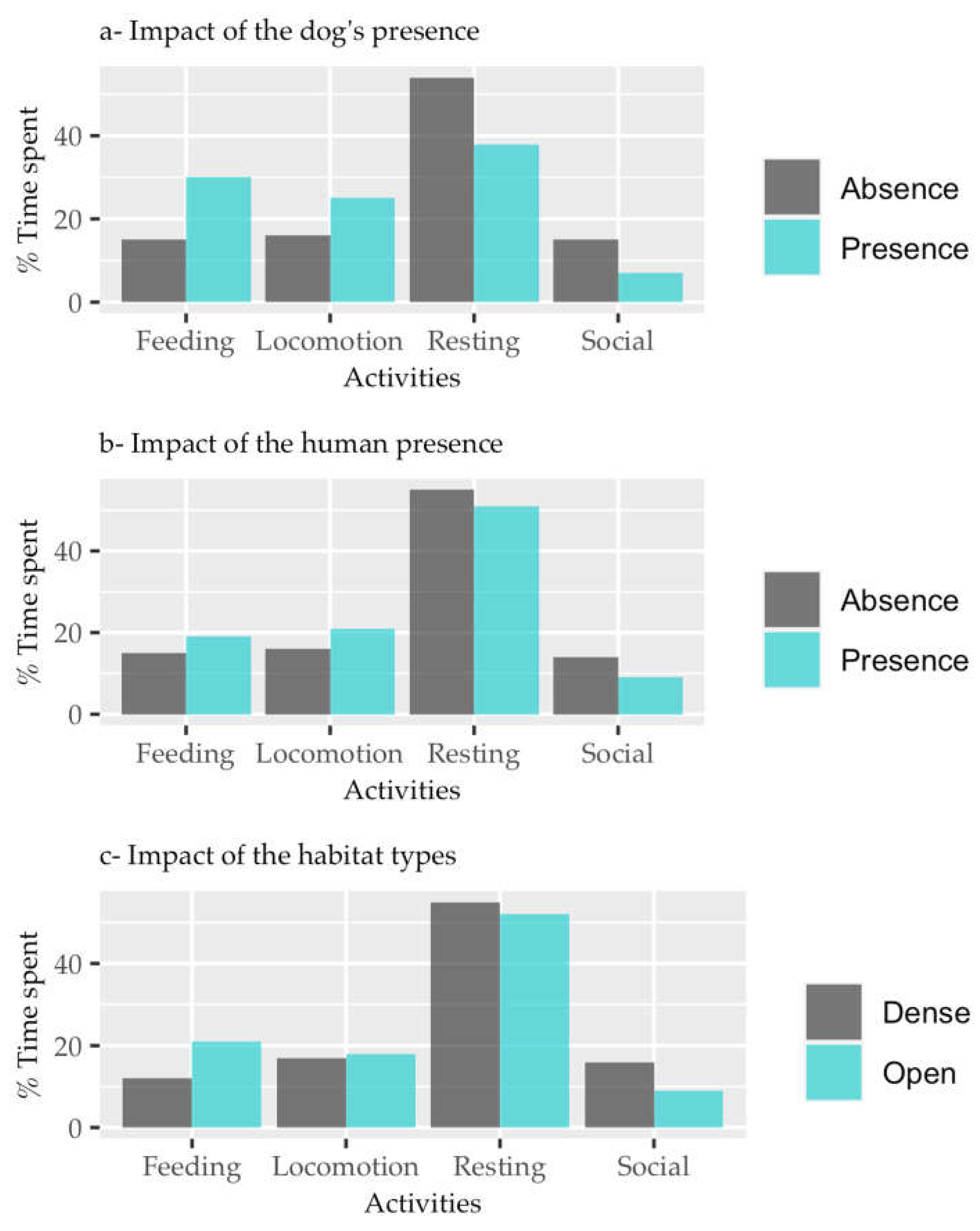

Table 1.
Model selection based on AIC.
| MLR Models | K | AICc | ΔAICc | Weight |
|---|---|---|---|---|
| PD+PH+H-OF+( 1 Year) | 12 | 154340.4 | 0 | 1 |
| PH+H-OF+( 1| Year) | 9 | 154564.4 | 223.96 | 0 |
| PD+H-OF+( 1| Year) | 9 | 154569.3 | 228.96 | 0 |
| H-OF+( 1| Year) | 6 | 154959.4 | 618.97 | 0 |
| PD+PH+( 1| Year) | 9 | 155579.5 | 1239.11 | 0 |
| PD+( 1| Year) | 6 | 155833.3 | 1492.9 | 0 |
| PH+( 1| Year) | 6 | 155888.1 | 1547.76 | 0 |
| MLR~1+( 1| Year) | 3 | 156371.4 | 2030.96 | 0 |
ΔAICc: difference in the AICc between the model with the lowest AICc and the following one; AICc: Akaike's information criterion corrected for small sample size; Cum. weight: Cumulative weight; H-OF: Habitat Type-Open Forest; K: number of free parameters in the model; PD: Presence of Dog; PH: Presence of Human; Weight: model probabilities.
Table 2.
Results from a multinomial logistic regression model.
| Predictor variable | Presence of Dog | Presence of Human | Habitat- Open Forest | |
|---|---|---|---|---|
| Feeding | z value | 0.601 | 11.255 | 25.641 |
| p value | <0.01 | <0.01 | <0.01 | |
| *Coef (SE) | 0.016 (0.027) | 0.288 (0.026) | 0.585 (0.023) | |
| Locomotion | z value | 6.135 | 8.122 | 4.717 |
| p value | <0.01 | <0.01 | <0.01 | |
| Coef (SE) | 0.156 (0.025) | 0.202 (0.025) | 0.106 (0.022) | |
| Social | z value | 11.963 | 5.407 | 17.872 |
| p value | <0.01 | <0.01 | <0.01 | |
| Coef (SE) | 0.386 (0.032) | 0.163 (0.030) | 0.492 (0.028) | |
*Coef (SE): Regression coefficient (standard error).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



