
Submitted:
25 April 2024
Posted:
26 April 2024
You are already at the latest version
Abstract
It has been widely established that the characterization of extracellular vesicles (EVs), particularly small EVs (sEVs), shed by different cell types into biofluids, helps identify biomarkers and therapeutic targets in neurological and neurodegenerative diseases. Recent studies are also exploring the efficacy of mesenchymal stem cell-derived extracellular vesicles naturally enriched with therapeutic microRNAs and proteins for treating various diseases. Besides, EVs released by various neural cells play a crucial function in the modulation of signal transmission in the brain in physiological conditions. However, in pathological conditions, such EVs can facilitate the spread of pathological proteins from one brain region to the other. On the other hand, analysis of EVs in biofluids can identify sensitive biomarkers for diagnosis, prognosis, and disease progression. This review discusses the potential therapeutic use of stem cell-derived EVs in several central nervous system diseases. It lists their differences and similarities and confers various studies exploring EVs as biomarkers. Further advances in EV research in the coming years will likely lead to the routine use of EVs in therapeutic settings.
Keywords:
exosomes
; extracellular vesicles
; diagnostic. sequencing
; biomarkers
1. Background
Extracellular vesicles (EVs) constitute a heterogeneous range of membrane-bound vesicles, broadly classified as exosomes and microvesicles. Exosomes originate from endosomes, whereas microvesicles are formed by the direct budding of the plasma membrane [1]. The formation of exosomes depends directly on the biogenesis of multivesicular bodies (MVBs) [2]. These membrane structures are highly dynamic and directly involved in internalizing extracellular ligands or cellular components, promoting their recycling or degradation [2,3]. Endosome formation occurs directly from the invagination of the plasma membrane [1,4]. While most endosomes undergo lysosomal degradation, some will mature and become late endosomes [1,5], which generate intraluminal vesicles (ILVs) through the internal budding of the endosomal membrane itself. During this event, nucleic acids, lipids, and cytosolic proteins are incorporated into the vesicles formed [1]. The ILVs within late endosomes range in size from 30-140 nm and mature as exosomes [1,3,6,7]. This process begins with the reorganization of the endosome membrane so that it is highly enriched in tetraspanins [8]. Among tetraspanins, CD9 and CD63 are critical in exosome biogenesis [1]. Subsequently, the endosomal sorting complexes necessary for transport (ESCRTs) are recruited to the ILV formation site [9]. It is known that the generation of ILVs is facilitated by the interaction between syntenin and ALIX and the availability of heparan sulfate, syndecans, ALIX, and ESCRTs [10]. Although the ESCRT pathway is considered the primary mode of generation of exosomes, different studies have shown other ESCRT-independent mechanisms for exosome biogenesis [11,12]. Eventually, the direct fusion of the MVB with the plasma membrane promotes the release of exosomes into the extracellular space [1]. In facilitating exosome release, RAB11 and RAB35 play an auxiliary role in the fusion of the MVB with the plasma membrane [13,14]. However, another mode of exosome release may be via the budding of the plasma membrane independently of Rab GTPases [1]. Another way is through SNARE proteins [15,16], typically involved in the membrane fusion process between two organelles [17,18]. In conclusion, exosomes have varied biogenesis and release of different endosome subtypes through various mechanisms and may contain different internalized molecules depending on the cell type and physiological state.
The formation processes of microvesicles [19] (MVs) are less known than exosomes [1,20]. It is believed to involve vertical molecular traffic directly to the plasma membrane, associated with a redistribution of membrane lipids and the use of machinery on the surface to allow compression and formation of the vesicle [20,21]. Therefore, a series of processes result in the biogenesis of MVs, with redistribution of phospholipids, including repositioning phosphatidylserine in the plasma membrane and contraction of the actin-myosin machinery [4]. For example, external factors may contribute to the release of MVs, such as the influx of calcium inducing the redistribution of membrane phospholipids, ultimately resulting in the increased release of MVs [22]. It is important to note that EVs cannot be classified into exosomes or microvesicles based on their size or cargo composition. Therefore, the International Society for Extracellular Vesicles guidelines suggest the EV scientists refrain from using the phrase “exosomes” unless there is evidence they have been released from endosomes [23]. Instead, the use of phrases, small EVs (sEVs) and large EVs is recommended based on the size of EVs.
2. Potential and Applications of EVs-Derived Mesenchymal Stem Cells
Recent studies are exploring the therapeutic value of mesenchymal stem cells-derived extracellular vesicles (MSC-EVs) for treating infectious diseases [24]. These results suggest that the MSC-EVs can eliminate pathogens, regulate immunity, and secrete antimicrobial factors, repairing tissue injuries, inhibiting the replication of pathogens and activating macrophages [24]. Previously, the therapeutic efficiency of MSCs was attributed to their capability to migrate and engraft in damaged tissues [25]. However, studies showed that administered MSCs typically do not reach the target tissue in sufficient numbers. On the other hand, the secretome of MSCs, including EVs, growth factors, and cytokines, likely reaches the various organ systems [25]. Such observations have led to the theory that the efficacy of MSC treatments mainly comes from the paracrine effects mediated by the secreted factors of MSCs [24]. MSCs have gained much importance in the past decades due to their therapeutic and clinical roles [25,26]. However, recent studies have highlighted the importance and advantages of using MSC-EVs because of their ability to cross biological barriers, higher safety profile and lower risks for tumorigenesis [25].
Several studies demonstrated the deep involvement of EVs in the physiology of the
Central nervous system (CNS), including processes such as synaptic activity, neuron-glia communication, and immune system response [27]. As well known, since MSC-EVs can cross biological barriers, such as the blood-brain barrier (BBB), to modulate the immune response, repair damaged tissue, and reduce inflammation, MSC-EVs seem attractive for treating CNS diseases.
MSC-EVs can transport RNA, proteins, and other biologically relevant molecules from cell to cell; EVs can also transport viruses, viral proteins, and nucleic acids [28]. A study conducted by Wenshuo Zhou [29] showed that MSC-EVs can carry the ZIKA virus, suggesting the possibility of disease spreading in CNS by EVs. In the context of ZIKA virus infection and, perhaps, other infections, studies in this area are fundamental to understanding the importance of EVs and developing strategies to treat CNS infections using EV-based drugs [29].
Besides their excellent utility in infection treatments, EVs may be useful as disease biomarkers. EVs of variable cells can be isolated from peripheral circulation using different methods [30]. In a study conducted by Pulliam [30], brain-derived EVs purified by a precipitation/immunoaffinity approach using antibodies against neuronal cell adhesion molecule L1CAM demonstrated the composition of EVs in CNS disorders, including HIV-associated neurological disorders (HAND) and Alzheimer’s disease (AD) [30]. Thus, the characterization of EVs is a valuable tool to identify biomarkers for CNS diseases, including infections. Therefore, exploring the possibilities of treating CNS infections with MSC-EVs is vital. The advantages of using EVs over the use of MSCs are apparent. Due to the properties of crossing physiological barriers, such as the BBB, reduced risks of immune response, not replicating after intravenous or intranasal administration, and being less likely to form clots, they seem ideal for treating brain disorders [31].
3. Central Nervous System as a Target and Niche for EVs
The importance of intercellular communication in the body's physiology is well established. Recent research has revealed that EVs can modulate and coordinate signal transmission in the brain [32]. According to studies, practically every cell in the CNS may release EVs, allowing them to play various roles in the brain [33], such as mediating communication between neurons and glial cells [34,35]. They can also facilitate proximal and distant communication between cells in the CNS and periphery [36].
The brain's microvascular system is necessary for maintaining homeostasis. EVs generated by vascular endothelial cells (ECs) have been reported to increase the survival, proliferation, and migration of oligodendrocyte precursor cells [37]. An essential component of the brain is the neuron, an electrically excitable cell that communicates with other cells via synapses. Additionally, research has revealed that neurons can communicate with other cells by producing EVs. These neuron-derived EVs (nEVs) can carry miRNA into ECs, specifically miR-132, to regulate the integrity of the vasculature by targeting the eukaryotic elongation factor-2 kinase (eek2) [38]. Furthermore, growth factors such as vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF) may be present in these nEVs, which can be released into the extracellular matrix (ECM) and contribute to protection [39]. nEVs can also be shed from glutamatergic synapses, which are likely to bind to nearby neurons and cause interneuronal communication [40,41]. An intriguing study discovered that nEVs can be released in situ, enhancing neuronal activity and modulating astrocyte function [42]. Secretion of nEVs can also be controlled via depolarization [43,44,45,46]. In addition, they can increase excitatory transmission at the presynaptic location by increasing the lipid fraction, such as sphingolipid metabolism in neurons [47]. Therefore, nEVs can modulate neuron function, but neurons can modulate EV function [42], demonstrating that communication can go both ways [48]. Furthermore, these nEVs can also deliver miRNA into astrocytes, causing the glutamate transporter-1 (GLT1) protein to be regulated [49]. nEVs can also communicate with microglia by increasing the expression of the C3 protein, which causes neurite degeneration [50]. This suggests that the nEVS can regulate synaptic function by inducing microglia phagocytosis, causing weaker synapses to be removed while solid synapses are retained [50]. Furthermore, EVs can stimulate neurite development [51] as well as contribute to cell survival by lowering apoptosis-induced cell death [52].
Glia cells in the CNS are responsible for the inflammatory response and neurotropic support [53]. More specifically, research indicates that microglia – a subset of glial cells – are primarily responsible for communication with neurons [48]. Furthermore, microglia play an important role in the initial response to pathological processes such as injury and infection [54]. Aside from their role in brain physiology, reactive microglia can release a miR-146a-5p that inhibits two synaptic function proteins through EVs, which can propagate the degenerative process in a neuronal culture, resulting in a reduction in dendritic spine and synapse function [55].
The release of EVs into the cerebrospinal fluid (CSF) has been found to be influenced by age [56]. This research adds to an in vitro investigation that found astrocyte-derived EVs (ADEVs) could transport oligodendrocyte senescence-related factors, including arginase-1 and tyrosine phosphatase zeta [57]. Astrocytes form a thick arborization that supports CNS cells and helps in regulating chemical impulses. As a result, ADEVs can carry miRNAs, which regulate neuronal protein expression and alter dendritic arborization in hippocampus neurons [58]. Also, it can transmit Neuroglobin (NGB) to neurons, which reduces degeneration and hence contributes to protection [59]. They can also produce growth factors that help cells in the brain differentiate [60]. The ADEVs can impact the activity of neural stem progenitor cells (NSPC), which produce many cell types in the brain. They can attach to NSPCs and induce differentiation into a specific phenotype, such as astrocytes. As a result, these EVs can control the crossover between neurogenic and gliogenic phases during development [61]. EVs containing inflammatory factors like tumor necrosis factor-alpha (TNF- α) are not limited to microglia [62]. Astrocytes can also release pro-inflammatory molecules like IL-1b in the brain [63]. ADEVs have a distinct signature when released by reactive astrocytes. ADEVs comprise a protein associated with target selectivity, such as integrins, as well as a cytokine associated with neuroinflammation [64]. Neurotoxic proteins may be present, resulting in neurodegeneration [65]. Reactive astrocytes can shed increased quantity of EVs to aid tissue healing [66].
Oligodendrocytes are CNS cells that support, maintain, and mediate chemical signals that can change information. In addition, myelin is produced in the axons of mature oligodendrocytes, which is required for action potential transmission [67] Studies have shown that EVs play a part in all of this communication by maintaining axonal integrity [68] and sending non-useful cellular material to microglia, which helps to retain myelin [69]. Furthermore, by lowering oxidative stress, oligodendrocyte-derived EVs can improve neuronal metabolism and protect neurons [45]. Increases in Ca2+ in the microenvironment can help preserve myelin by stimulating the release of myelin-associated proteins such as myelin proteolipid protein (PLP), 2',3'-Cyclic nucleotide 3'-phosphodiesterase (CNP), and myelin basic protein (MBP) [70]. The fusion of EVs generated from oligodendrocytes in the axons can be improved by increasing Ca2+ [70].
Overall, EVs can play an important role in maintaining brain homeostasis, but they can also be shed by injured cells, contributing to developing pathological conditions in the CNS [71]. As a result, the question arises as to whether such EVs are involved in CNS disorders.
4. EVs as Biomarkers for CNS Diseases
EVs, secreted by virtually all cells [72], can protect their encapsulated material from the extracellular environment [73] and cross the blood-brain barrier in either direction [74]. As a result, EVs released by CNS cells during a disease condition may reflect the altered physiology of the secretory cell [75]. These EVs, detected in biological fluids such as CSF, blood, urine, and saliva, could be biomarkers [76] as showed in Figure 1.
Despite advances in finding CNS disease biomarkers based on blood and CSF samples and neuroimaging techniques, some limitations remain. In this context, EVs and their cargo, such as proteins and miRNAs, are being investigated as potential biomarkers in CNS diseases, particularly in peripheral samples. Such investigations can contribute to developing sensitive and non-invasive biomarkers for diagnosis, prognosis, and disease progression [77,78,79]. The studies characteristics and main findings are summarized in Table 1.
5. Alzheimer’s Disease
Alzheimer's disease (AD) is a progressive neurodegenerative disease and the most common cause of dementia. Amyloid-β and tau protein aggregates are the principal neuropathological hallmarks of AD [80]. However, there is no correlation between the quantities of these proteins measured in CSF and blood. In patients with AD, blood levels of the brain-derived exosomal proteins amyloid-β42 (Aβ-42), total-tau (t-tau), tau phosphorylated at threonine 181 (p-T181-tau), and tau phosphorylated at serine 396 (p-S396-tau) are greater than in control patients. These findings are consistent with what is seen in CSF, indicating that they could be used in clinical practice as non-invasive premortem AD biomarkers [81]. Still, in patients with AD and mild cognitive impairment, neurogranin levels are higher in the CSF and plasma but lower in the blood plasma neuronal-derived EVs [82]. However, combining the EV-associated synaptic proteins neurogranin, growth-associated protein 43 (GAP43), synaptosome-associated protein 25 (SNAP25), and synaptotagmin 1 could potentially predict AD 5 to 7 years before cognitive impairment [83].
Numerous miRNAs transported by EVs associated with AD have been described in the literature [84,85,86,87]. The fact that the let-7e miRNA is enriched in nEVs when detected in plasma suggests that it could be a biomarker for AD diagnosis [88]. Yang et al. found that serum EVs enriched with miR-135a and miR-384 and depleted with miR-193b are characteristics of AD patients compared to controls. EV-associated miR-384 was the most effective in distinguishing between other dementia diseases among the three miRNAs [89]. Whether or not these isolated findings are promising, the profile of miRNAs carried by EVs in different biochemical fluids in AD patients allows for a systemic assessment of the process [90,91,92].
As a biomarker discovery tool, EVs can be used to track the prognostic and progression of AD. According to a pilot study conducted by Li et al., hsa-miR-1306-5p, hsa-miR-93-5p, hsa-miR-424-5p, and hsa-miR-3065-5p, as well as expression of P-S396-tau in plasma-derived EVs, provide a protein and miRNA signature to differentiate AD patients from other dementia disorders and healthy patients. Therefore, it could afford a better prognostic capacity [93]. Aharon et al. investigated the role of EVs in the progression of AD. As the disease progressed, neural markers MOG and CD171 in serum EVs increased proportionally. EVs inflammatory cytokines were elevated in AD patients. Furthermore, a link between the severity of the disease and the lowering of particular miRNAs was discovered. Thus, this conjunction of alterations could be employed as a diagnostic for AD severity [94].
6. Parkinson’s Disease
Parkinson's disease (PD) is a neurological disease that causes motor dysfunction [95]. The etiology and spread of the disease are attributed to the aggregation of alpha-synuclein (α-syn), which in part may be mediated by EVs [96]. Although α-syn levels in EVs have been investigated as potential biomarkers for the premature diagnosis and progression of PD, many findings are conflicting. Patients with PD had a greater α-syn concentration in CNS-derived plasma EVs, which is associated with disease severity [97]. A larger concentration of α-syn in plasma EVs could distinguish PD from control patients and other illnesses that can produce α-syn aggregation [98], corroborating the previous conclusion. In individuals with early-stage PD, levels of α-syn in plasma neuronal exosomes are increased and longitudinally greater than baseline levels [99]. In addition, the rise in α-syn in nEVs also occurs before the diagnosis of PD and continues to increase with disease progression [100]. These findings could help promote early diagnostic biomarkers and prognostic for PD progression [99,100].
Patients with mild and severe PD, on the other hand, have much lower plasma levels of α-syn EVs than patients without PD [101]. Furthermore, in patients with PD, the level of α-syn in CNS-derived serum exosomes is smaller than in patients with essential tremor and healthy people. When patients with PD were subdivided, the decline was more significant in non-tremor-dominant (NTD) patients than in tremor-dominant (TD) patients. Although inconsistent with other findings, this could help promote PD diagnostic biomarkers and distinguish NTD from TD [102]. These discrepancies could be due to variances in extraction processes, types, and markers from particular CNS cells from EVs. Therefore, α-syn deserves additional studies to develop as a useful clinical biomarker. In addition, an increase in α-synuclein (α-synOlig) e α-synOlig/total α-synuclein ratio in EVs was seen in alternative fluids such as saliva. Therefore, this fluid can be used as an alternative to promote α-syn-associated PD biomarkers [103]. In plasma-derived EVs from PD patients, the expression of hsa-miR-30c-2-3p and miR-331-5p was increased, while the expression of hsa-miR-15b-5p, hsa-miR-138-5p, hsa-miR-338-3p, hsa-miR-106b-3p, and hsa-miR-431-5p and miR-505 was decreased [104]. While miR-24 and miR-195 had increased expression in PD patients' serum EVs, miR-19b had lower expression [105]. Furthermore, a profile of six serum EV-derived miRNAs expressed differently in PD patients can be employed as early PD diagnostic and progression biomarkers [106]. The miR-34a-5p, which is involved in neurobiology, was up-regulated in plasma small EVs and associated with disease duration [107].
Proteomic analysis of EVs in serum, urine, plasma possibly derived from neurons, revealed that several proteins are differentially present between PD patients and controls [108,109,110,111]. DJ-1 protein is significantly elevated in EVs derived from plasma neurons as well as exosomes isolated from urine in PD patients compared to controls [112,113]. Furthermore, Ser(P)-1292 LRRK2 levels are high in idiopathic PD patients' urine exosomes [114]. The significantly higher plasma EV pro-IL-1 and TNF- levels in PD patients compared to controls could indicate that inflammation is involved in the disease's development and progression [115].
7. Huntington’s Disease
Huntington's disease (HD) is a neurodegenerative hereditary condition that usually results in motor, cognitive, and psychiatric disorders [116]. It is caused by the repeat expansion of the CAG in the huntingtin gene (HTT). The mutant HTT (mHTT) protein includes an expanded polyglutamine (polyQ) repeat and can form aggregates, which is neurotoxic [116]. In vitro, EVs were found to integrate both extended repeat RNA and polyQ protein. Despite this, no evidence of neurotoxicity has been found, most likely due to experimental limitations [117]. Platelets have a higher concentration of mHTT than other blood cells, with an increase proportional to the course of HD [118]. In the case of EV platelets, however, mHTT was undetectable. Additionally, activated platelets no longer release EVs, indicating that mHTT-mediated procoagulant contribution is absent in HD [119]. So far, no study has looked at the miRNA profile carried by EVs, which could be useful in understanding and promoting HD biomarkers [120].
8. Multiple Sclerosis
Multiple sclerosis (MS) is a chronic autoimmune disease characterized by inflammation, demyelination, and neuro-axonal damage. It is frequently linked to progressive disability and a variety of clinical outcomes and prognoses [121]. Recently, researchers identified the miR-150-5p and let-7b-5p myeloid EVs as potential biomarkers for cognitive deficits in MS [122]. In relapsing-remitting MS (RRMS), the miR-326 is upregulated in EVs derived from conventional T cells. It is associated with immunopathogenesis and may serve as a biomarker for the diagnosis and progression of MS [123]. The concentration of EVs and their miRNA load in plasma, serum or specific cells can be used to predict response and monitor therapies [124,125,126,127].
Serum EVs miRNAs are differentially expressed in isolated syndrome and RRMS, possibly contributing as stage-specific biomarker potential [128]. During RRMS relapse, the hsa-miR-122-5p, hsa-miR-196b-5p, hsa-miR-301a-3p, and hsa-miR-532-5p in serum EVs [129], as well as hsa-miR-148-5p in erythrocyte-derived EVs [130], compared to the control group were found differentially expressed. Moreover, differentially expressed serum exosomal miRNAs in RRMS and progressive MS can predict disease subtypes [131].
Other EV cargo may be associated with different pathways during MS pathology, providing potential non-invasive biomarkers based on fluids detected. Myelin proteins, synaptopodin, and synaptophysin, as well as the complement cascade and sulfatide, can be altered and found in plasma, serum, neuronal, and astrocytic-plasma EVs during the pathophysiology of MS [132,133,134]. Plasma-derived EVs released by T cells, B cells, and monocytes were elevated in untreated RRMS cases with low disability, allowing for a better understanding of immune system modifications when noticeable changes in disability or demyelinating events are not yet visible [132]. Increased levels of CNS endothelial cell-derived EVs in plasma could be a biomarker of BBB permeability and active illness in MS [135].
9. Epilepsy
Epilepsy, a brain disorder that involves the epileptogenic processes, is characterized by a predisposition to produce epileptic seizures [136]. EVs have the potential to be diagnostic and prognostic biomarkers for epileptogenesis and epilepsy [137]. In patients with temporal lobe epilepsy, alterations in CSF miRNAs, including exosomal miRNAs, were demonstrated in a pioneering study [138]. Another study demonstrated that EV-associated miR3613-5p, miR-4668-5p, miR-8071, and miR-197-5p isolated from plasma may distinguish people with mesial temporal lobe epilepsy and hippocampal sclerosis from healthy controls. Furthermore, miR-8071 was linked to seizure severity [139]. In an experimental approach, exosomal downregulation of miR-346 and miR-331-3p was seen in the forebrain, resulting in epileptogenesis in a mouse model of chronic temporal lobe epilepsy (TLE) [140].
In TLE patients, plasma sEVs derived miRNA panel (hsa-miR-342-5p, hsa-miR-584-5p, hsamiR-125b-5p, hsa-miR-199a-3p, and hsa-miR-199a-5p) can predict the probability of drug-resistant children [141]. The expression of miR194-2-5p, miR15a-5p, miR-132-3p, and miR-145-5p from serum exosomes may be linked to Focal Cortical Dysplasia (FCD) and refractory epilepsy via the signaling pathways mTOR, PI3K-Akt, p53, TGF-beta, and cell cycle control [142]. In addition, coagulation factor IX and thrombospondin-1, which may be implicated in various epileptic physiological processes, were shown to be differentially expressed in serum-derived EVs in clinical samples and chronic epilepsy models [143].
10. Traumatic Brain Injury
A traumatic brain injury (TBI) is a brain injury induced by an external force that occurs suddenly [144]. The profile of proteins from EVs circulating can help promote potential biomarkers for diagnosis, identify the temporal profile and predict outcomes in patients with TBI [145,146]. In EVs isolated from CSF, several proteins were associated with severe TBI. The injured human brain released more EVs, which could contain cell-to-cell communication components in both cell death signaling as well as neurodegenerative pathways [147]. Most neurofunctional proteins increased in neuron-derived EVs isolated from plasma in acute TBI, while neuropathological proteins increased in chronic and acute TBI, possibly indicating phase specificity [148]. The genes CDC2, CSNK1A1, and CTSD were upregulated in salivary EVs, which could be used as biomarkers in mild TBI (mTBI) [149].
In the mTBI, tau, A42, and IL-10 concentrations in neuronal-derived EVs isolated from plasma were higher than in controls. Post-concussive symptoms and post-traumatic stress disorder were linked to tau and IL-10 levels, respectively [150]. Sleep quality was linked to EV levels of IL-10 and TNF in mTBI patients. mTBI patients with a significant risk of sleep difficulties have higher levels of inflammatory cytokines in their blood [151]. EV concentrations of neurofilament light chain (NfL) from plasma are related to repetitive mTBIs, even years after the injury, with the highest levels in those with unfavorable chronic symptoms [152]. Corroborating these findings, increased GFAP and NfL concentrations in blood-derived EVs one year after injury are higher in TBI patients than controls. They are also associated with injury severity and poor recovery outcomes [153].
In TBI, distinct miRNA profiles in EVs involve different disease-related pathways. A predictive blood test for TBI can be created using miR-203b-5p, miR-203a-3p, miR-206, and miR-185-5p EVs from plasma [154]. In the chronic mTBI cohort, 45 miRNAs in plasma EVs were substantially different from control groups, and they were mostly related to pathways involved in neuronal function, vascular remodeling, blood-brain barrier integrity, and neuroinflammation [155]. The hsa-miR-139-5p and hsa-miR-18a-5p EVs from plasma were significantly differentially expressed in the repetitive TBI and could be used to predict chronic TBI symptom persistence [156]. In TBI cases associated with disturbed consciousness, elevated GFAP and eleven miRNAs were seen in EVs from plasma.These data, taken together, could represent a valuable source of TBI biomarkers [157].
11. EVs as Treatment for CNS diseases
After Raposo and coauthors had shown in 1996 that EVs could stimulate adaptative immune responses, several studies were undertaken to understand their role and how and if they could be used as treatment tools. At first, it was known that EVs were closely linked to the pathogenesis of some diseases, including neurological diseases such as Alzheimer’s and Parkinson’s, becoming new therapeutic targets [158,159,160]. Further, they were exploited directly as therapeutic agents, not only because they can stimulate tissue repair and act as immunotherapy agents but also because of the possibility of their use as vehicles for drug delivery [158,159,160,161].
Almost a decade after the initial understanding of the role of EVs, their ability to transfer exogenous nucleic acids and proteins between cells was brought to light [160,161]. Because they are secreted by most cells and can be acquired from the patients, they are a more biocompatible vehicle for drug delivery than other available vehicles [160]. The techniques of loading EVs with therapeutic agents are divided into two groups: exogenous loading (when the molecule of interest is incorporated into or onto the isolated EVs and endogenous loading (when the molecules are provided so the parental cell can incorporate them into the EVs they will secrete) [159,160,161]. EVs also have the innate ability to cross the BBB, and if specific surface ligands are employed, they become a valuable treatment tool for CNS diseases (158-161) as showed in Figure 1. The studies characteristics and main findings are summarized in Table 2.
12. Alzheimer Disease (AD)
Known as the most prevalent kind of dementia, AD patients present neurodegeneration, and the pathogenesis of this disorder is complex and multifactorial. Fewer studies are testing EVs in AD models compared to other pathologies. Among these studies, it was shown in vitro that MSC-EVs derived from human adipose tissue contained neprilysin, an enzyme able to lyse the beta-amyloid peptide, a peptide strongly associated with the pathogenesis of AD [162,163,164].
In vivo, the administration of EVs can also be helpful in AD because of their scavenger role. They can trap the beta-amyloid peptide and deliver it to microglia, where it can be degraded, increasing its clearance [162,163]. Numerous pre-clinical studies also demonstrate that stem cell-derived EVs are an emerging therapeutic tool for several CNS disorders [165]. They can be loaded with many different agents and present the ability to cross the BBB, becoming an important drug vehicle [158,161,166,167]. Depending on what is loaded into the EVs, they can block gene defects, replace missing genes, modulate the downstream effects of diseases and change cell phenotypes [167]. So far, studies suggest that EVs may have to be administrated several times once they are biologically degraded [167]. They also demonstrated that EVs are an alternative to cell therapy because of their lesser side effects such as oncogenic transformation, undesirable cellular differentiation, and embolic risk [161]. EVs can also target both neurons and glia following intranasal administration, as demonstrated in a mouse model of AD [165] implying that EVs can potentially induce transcriptomic changes in both neurons and glia that could slow down disease pathogenesis and cognitive problems.
Despite promising prospects, there needs to be more quantitative data on the amount of EVs or drugs loaded into EVs that reach the CNS after intravenous or intranasal administration [167]. In addition, the endogenous cargo from the parental cell may cause adverse effects in some diseases or even interact with the loaded material [166]. Therefore, a better characterization of chosen EVs in pre-clinical models is critical for understanding their properties before employing them in treating AD [158,166].
13. Parkinson Disease (PD)
PD is also a common neurodegenerative disorder in which patients present death of dopaminergic neurons in the substantia nigra of the brain due to elevated alpha-synuclein, brain inflammation, and oxidative stress [158,159,161,167]. One of the main goals of developing new therapeutics for PD and other neurodegenerative disorders is delivering drugs across the BBB to show their effects in the CNS [167]. Studies already achieved down-regulation of alpha-synuclein in animal models through systemic injection of EVs from dendritic cells (DCs). These EVs displayed rabies viral glycoprotein to target brain cells and were loaded with siRNA to alpha-synuclein [159,161,167,168,169].
EVs were also used to deliver Catalase, an antioxidant that is diminished in PD patients' brains, as a means to cross the BBB. They were secreted from genetically modified macrophages that overexpressed Catalase, and the results were reduced inflammation and neuroprotection in a mouse model. Studies also proved that EVs loaded with Catalase and delivered to mice intranasally also showed a reduction of brain inflammation, when compared to free Catalase administration [161,166,170,171,172].
The primary drug available for PD is dopamine, but its exclusive peripheric effects are well known once it cannot cross the BBB. However, when loaded in EVs, an improved functional recovery in a murine PD model without toxicity in the hippocampus, liver, spleen, or lungs was observed [161,173]. In vitro studies showed that EVs from deciduous teeth-derived stem cells present neuroprotective potential and prevent apoptosis in 80% of dopaminergic neurons when EVs from stem cells maintained in 3D cultures were employed. However, such effects were absent with EVs from stem cells maintained in standard 2D culture conditions, revealing that even the same type of stem cell can release different EVs based on the medium and culture conditions employed [166,170,172].
14. Huntington Disease (HD)
HD is a neurodegenerative disorder caused by a specific alteration in the Huntington gene (HTT). It is dominantly inherited, and patients present with progressive cognitive impairment, involuntary movements, and psychiatric changes [159,161]. The onset is between 35-45 years, and death occurs approximately 15 years later, with no therapy available to treat or slow down disease progression, only to mitigate symptoms [166].
In preclinical studies, incorporating a hydrophobically modified small interfering RNA (hsiRNA) capable of enhancing stability and promoting cellular internalization into EVs targeted the wild and mutant huntingtin mRNA. It led to dose-dependent silencing of huntingtin mRNA and protein both in vitro (primary cortical neurons culture) and in vivo (after infusion in the cerebral spinal fluid in an HD mouse model) [158,159,161,166,167]. However, the EVs were isolated from U87 glioblastoma cells, and researchers recognize that this cell line can provoke tumor formation, highlighting the importance of more pre-clinical studies [158,159,161].
Further investigations exploit the use of EVs loaded with microRNAs (miRNAs) in an HD mouse model. More specifically, the miR-124, whose expression is down-regulated in HD [158,159,161,168]. The miR-214 is highly and specifically expressed in all brain regions but the pituitary gland and has a regulatory role in CNS development and diseases, supporting adult neurogenesis [166,168]. It has been reported to slow down HD progression in transgenic mice by promoting neuronal differentiation. However, after the injection of EVs into the mouse striatum, Lee and colleagues showed modest therapeutic efficacy and no behavioral improvement, with a reduction of REST protein expression [159,161,166,170].
Additionally, Wu and coauthors tested the therapeutic potential of small interfering RNA (siRNA), molecules capable of using RNA-induced silencing complex (RISC) to provide gene silencing. EVs expressing neuron-specific rabies viral glycoprotein (RVG) and loaded with siRNA targeting Huntington transcript were injected intravenously to control and transgenic mice. The association with RVG and siRNA significantly reduced the transcript expression by up to 46% and 54%, depending on the transgenic mouse line (BACHD and N171-18Q, respectively) [166].
15. Multiple Sclerosis (MS)
MS is the most common non-traumatic cause of neurological disorders of CNS. Patients present multifocal inflammation, demyelination, and neuronal loss. Studies have used MSC-EVs and showed improvements in motor deficits and reduced brain atrophy, following the expected reductions in plasma proinflammatory cytokines in animal models. In vivo and ex vivo, it was also proven that increased myelin levels and decreased oxidative stress could be obtained by employing EVs isolated from IFNy-stimulated DC. Administration of these IFNy-stimulated EVs to mice with experimental autoimmune encephalomyelitis (EAE) led to decreased disease evolution and spinal cord demyelination [158,159]. Similar results were seen with both intravenous and intranasal administrations [174,175].
Recent data showed positive results of using MSC-EVs in the EAE mouse model, a preclinical model of MS. Researchers injected EVs from MSCs and MSCs stimulated with IFN-ɣ and employed PBS solution and MSCs as controls. Mice were treated at day 18, when the disease peaked, and were clinically evaluated. It was shown that intravenous administration of EVs from MSCs stimulated by IFN-ɣ reduced the mean clinical score, demyelination, and neuroinflammation compared to the PBS control group. Also, an upregulation of CD4+ CD25+ FOXP3+regulatory T cells within the spinal cords of mice was observed [169].
The same group showed that in vitro co-culture of EVs from MSCs stimulated by IFN-ɣ with activated peripheral blood mononuclear cells (PBMCs) reduced their proliferation and levels of proinflammatory cytokines while increasing immunosuppressive components. Further, they sequenced RNA from IFN-ɣ EVs, which suggested they contained antiinflammatory RNA responsible for partial inhibiting Tregs induction. These IFN-ɣ EVs also carry multiple antiinflammatory and neuroprotective proteins, enlightening the possibility of a free-cell therapeutic.
Additional research groups have studied the effects of IV administration of MSCs EVs in a Theiler’s murine encephalomyelitis virus-induced demyelinating disease, and their results also showed a positive effect in improving motor deficits, reducing brain atrophy, increasing cell proliferation, decreasing inflammatory infiltration, and inflammation [172,173]. As the therapeutic pre-clinical properties of EVs for MS have been well explained, their preventive role is a subject of new studies. In 2021, Fleming developed in a mouse model a “vaccine-like treatment” for the antigens most related to MS. Since then, EVs have been studied as novel vehicles for vaccine deliveries, and the researchers aim to facilitate antigen delivery to monocytes, DC, macrophages, and microglia, bettering the immune tolerance [173,175].
16. Epilepsy
Characterized by recurrent seizures, epilepsy is a chronic disorder that may cause hippocampal injury, BBB disruption, and neurodegeneration, especially when patients evolve from a seizure episode to status epilepticus (SE). SE leads to acute neuroinflammation, which progresses into chronic neuroinflammation. EVs may modulate this pathological process, improving the cognitive function of the patients and preventing new spontaneous seizures [158,159,170,171].
Studies have used EVs from human bone marrow-derived MSCs in a mouse model of induced status epilepticus. EVs were applied intranasally (IN) twice within 24 hours after SE, and the results showed a reduction of proinflammatory cytokines in the acute phase after SE and better cognitive and memory functions with neuroprotection and maintenance of normal neurogenesis in the hippocampus [176]. However, additional studies are needed to better understand the role of EVs as a treatment against spontaneous recurrent seizures that emerge weeks or months after SE. The long-term effects of EV treatment after SE are unknown, particularly on chronic epilepsy development, or whether intermittent administrations after SE are critical to restrain chronic epilepsy development. Also, the potential of EV treatment to prevent psychiatric comorbidities is yet to be investigated. Despite these limitations, the overall results suggest that hMSC-EVs have the potential as an antiepileptic therapy [171].
17. Traumatic Brain Injury (TBI)
Traumatic Brain Injury (TBI) may be caused by an ischemic insult or trauma, among other insults. Studies showed that murine bone marrow-derived MSC-EVs injected intravenously into rats and mice with traumatic brain injury resulted in improved spatial learning, reduced neurological deficits and inflammation, and improved angiogenesis and neurogenesis [158,159,177]. EVs derived from MSCs were also applied intraperitoneally in animal models of preterm brain injury, and the results showed long-lasting cognitive improvement and restoration of short-term myelination deficits [178].
Studies have also investigated the role of human MSC-derived EVs in a controlled cortical impact (CCI) injury model. Intravenous administration 1 hour after injury suppressed proinflammatory cytokines (IL-1b) and tumor necrosis factor-alpha (TNF-a), improving spatial learning, promoting sensorimotor function recovery, and increasing vascular density after trauma. However, the cortical lesion volume was not altered. The effect was proven to be dose-dependent [170,171]. A recent study demonstrated that intranasal administration of MSC-EVs ninety minutes after a CCI is an efficient approach to target EVs into neurons and microglia into hemispheres on ipsilateral and contralateral sides of CCI [179]. In this study, dose-dependent effects of MSC-EVs were seen on both acute and chronic neuroinflammation and cognitive and mood impairments after TBI. An optical intranasal dose of MSC-EVs in this study restrained NLRP3 inflammasome activation in the acute phase of TBI, which was sustained in the chronic phase, leading to reduced chronic neuroinflammation. Notably, such EV treatment prevented the overactivation of p38/MAPK signaling in the chronic phase, providing a mechanistic insight into reduced chronic neuroinflammation mediated by MSC-EVs. Furthermore, the study demonstrated better hippocampal neurogenesis on the injured cerebral hemisphere with the maintenance of standard brain-derived neurotrophic factor (BDNF)-phosphorylated cyclic AMP response element binding protein (cCREB) signaling [180]
Additional studies have shown that endothelial colony-forming cell-derived EVs can promote recovery in mice subjected to CCI. These EVs reduced brain edema and the degradation of tight-junction proteins. Such results have led to the belief that EVs have an essential role in restoring the BBB integrity [137,170,171]. EVs derived from other stem cells, such as the adipose tissue MSC-derived EVs, were shown to have a beneficial role in the CCI-induced TBI in rats through the noncoding RNA MALAT1. When analyzing the brain and spleen transcriptome, this RNA-regulated gene was found to be involved in inflammation. The study suggested that EVs first migrate to the spleen, liver, and lungs (within 1 hour), and only after three days could the impact of EVs be observed on the brain. The study also implied that EVs work by inhibiting the release of peripheral macrophages and monocytes from the spleen, with fewer immune cells trespassing the damaged BBB [181].
18. EVs and CRISPR-Based Gene Editing Systems
Approximately 90% of human pathogenic genetic variants are single-base mutations or insertions and deletions of fewer than a dozen base pairs [182], underscoring the relevance of genetic editing tools in addressing these genetic aberrations. By leveraging CRISPR-based approaches, researchers can generate cellular and animal models harboring disease-associated mutations, facilitating the elucidation of disease mechanisms and the screening of potential therapeutic interventions. Moreover, the advent of precise genome editing techniques, such as base editing and prime editing (reviewed in REF. [183,184]), holds promise for correcting pathogenic mutations at the molecular level, offering potential avenues for therapeutic intervention.
In order for genome editors to fulfill their therapeutic potential within living organisms, they need to be effectively delivered to specific cells without triggering any undesirable reactions. Enhancing the delivery efficiency of CRISPR/Cas9 and gene editing is achievable through the use of viral vectors. However, the practical application of viral vectors is hindered by inherent drawbacks, including safety concerns, high expenses, complexity in handling, and limited cargo capacity [185,186,187]. Nonviral vectors, on the other hand, rely on physical methods and synthetic or natural materials, including microinjection, electroporation, liposomes, nanoparticles, and gold nanoparticles [188,189]. While these alternatives offer advantages such as simplicity, cost-effectiveness, and versatility, they are hindered by issues such as cytotoxicity, limited tissue penetration, and imprecise targeting delivery, constraining their further advancement [189,190].
Considering their capacity to carry diverse biomacromolecules, target specific cells, modulate immune responses, and undergo engineering modifications, EVs emerge as promising candidates for delivering CRISPR-based gene editors. In general, the delivery mechanism for the CRISPR/Cas9 system can be categorized into three main forms: DNA, RNA, or ribonucleoprotein complexes (RNPs).
Studies have initially demonstrated the capacity of tumor EVs to be engineered to deliver CRISPR/Cas9 plasmid DNA and RNP complexes both in vitro and in vivo [191,192,193]. Kim et al. demonstrated that SKOV3 cancer cell-derived exosomes carrying CRISPR-Cas9 plasmid and sgRNA to target the PARP1 gene in SKOV3 xenograft mice [10]. The researchers demonstrated that the treatment introduced indels in PARP1-positive SKOV3 cells, leading to apoptosis of these cells in mice with ovarian cancer. Furthermore, they illustrated that cancer cells exhibit a preference for internalizing exosomes derived from cancer cells over those produced by epithelial cells. This finding underscores how cell tropism can impact the targeting capability of EVs towards particular cell types [191].
EVs can be engineered to facilitate the encapsulation of CRISPR-Cas9 ribonucleoprotein (RNP) complexes. One approach involves employing protein tethering, wherein the Cas9 enzyme is fused with protein domains or peptides that interact with complementary domains within EVs, thus promoting encapsulation. For instance, the ARRDC1 domain, known for its involvement in plasma membrane budding and EV generation, can be utilized [194,195,196]. ARRDC1 interacts specifically with proteins containing WW domains, such as itchy E3 ubiquitin protein ligase (ITCH) [194]. To achieve encapsulation, the WW domains of ITCH are fused to the N-terminus of Cas9 [196]. Consequently, the genetically modified Cas9 is encapsulated within EVs through interactions between the WW domains and ARRDC1. This engineered Cas9 RNP, when encapsulated, demonstrated a GFP knockdown efficiency of up to 13.4% in U2OS cells [196].
Encapsulation of Cas9 ribonucleoprotein (RNP) complexes can also be facilitated through the fusion of a myristoylated peptide, such as Src kinase or Basp1 [197,198]. This peptide guides the fused protein to associate with the cytoplasmic membrane, and subsequently, the inner membrane of extracellular vesicles (EVs) [198]. For instance, a peptide (MGGKLSKKKKGYNVNDEKAKEKDKKAEGAA) was fused to the N-terminus of spCas9, resulting in increased levels of Cas9 within EVs [198]. Similarly, another peptide (GSNKSKPK), derived from the N-terminus of Src kinase, when fused to the N-terminus of Cas9, led to its myristoylation and enrichment within EVs [199]. EVs engineered with VSV-G at their membrane and encapsulating Cas9 RNP induced knockout of the eGFP reporter gene in recipient cells with an efficiency of approximately 45% in vitro [199].
Duchenne muscular dystrophy (DMD), arising from diverse mutations in the Dmd gene, stands as one of the prevalent genetic disorders, capable of diminishing the expression of the dystrophin protein [200]. Majeau et al. showed that in mdx mice with a nonsense mutation in exon 23 of the Dmd gene, injection of serum EVs containing CRISPR/Cas9 RNPs led to the deletion of exons 23 and 24. This resulted in effective exon deletion and restoration of dystrophin protein expression in muscle tissues compared to the control group [201]. Furthermore, in hDMD/mdx mice expressing the human Dmd gene, the Dmd genes were also susceptible to modification [201]. Another group integrated an RNA aptamer sequence (Com) into the sgRNA loop and then fused the Com-binding protein into both terminals of CD63 [202]. This Com/com interaction facilitated the enrichment of CRISPR-Cas9 RNPs into extracellular vesicles EVs. They further showed that these EVs were capable of introducing indels in DMD/mdx mice when injected into the tibialis anterior muscle [21]. Similarly, Gee et al. introduced a novel technique named "NanoMEDIC," where they encapsulated RNP complexes within EVs utilizing an HIV-derived Gag protein. To facilitate the recruitment of the RNP into EVs, the researchers employed a chemical-induced dimerization system known as FKBP12 and FRB. They fused the Cas9 and Gag proteins to a heterodimerizer, enabling conditional dimerization upon the addition of an inducible chemical ligand to attract the Cas9 protein into the EVs [203]. Their findings demonstrated that NanoMEDIC achieved over 90% exon-skipping efficiency in skeletal muscle cells derived from iPS cells of DMD patients.
In summary, using EVs to deliver CRISPR-based gene editing tools offers a promising therapeutic approach for genetic disorders. EVs possess natural advantages like cell targeting and low immunogenicity, enabling precise editing of disease-causing mutations. Preclinical studies have shown efficacy in various models, as discussed above, though challenges like scalability and optimization remain. With ongoing advancements in EV engineering and CRISPR technology, EV-mediated CRISPR gene editing holds great potential for clinical translation, offering hope for improved outcomes in genetic disorder treatment.
19. Conclusion and future perspectives
EVs found in biological fluids are attractive candidates for biomarker discovery. On the other hand, EVs naturally shed by various stem/progenitor cells are increasingly considered excellent biologics for treating neurological and neurodegenerative diseases. As discussed in this review, different cell types can release these nanosized vesicles into body fluids, providing information about parental cells' physiological and pathological status. More studies are critical in attaining deep knowledge regarding the biological potential of EVs for diagnosing and treating neurological and neurodegenerative diseases. The points discussed in this review highlight a great potential in further extending EV analysis for future clinical applications. Particularly, brain-derived EVs in body fluids such as plasma could aid precision medicine approaches as analysis of the composition of EVs from specific brain cell types in individual patients could help assess the extent of neuronal dysfunction or astrocyte and microglia-mediated neuroinflammatory signaling mechanisms in various brain disorders. Such an approach could revolutionize the diagnosis, prognosis, and treatment of various pathologies in hard-to-treat neurological and neurodegenerative diseases.
Author Contributions
Conceptualization, G.Z., J.I.B.G., D.M., A.K.S..; methodology, G.Z., D.M., F.B.; software and figures, G.Z., I.M.G., V.W.; validation, I.M.G.; formal analysis, A.M.A.; investigation, G.Z., J.I.B.G., D.M., P.G.d.S., A.M.A., I.M.G., F.B., V.W.; resources, J.C.C.; data curation, G.Z., P.G.d.S., J.I.B.G., F.A.C.X.; writing—original draft preparation, G.Z., J.I.B.G., D.M., P.G.d.S., A.M.A., I.M.G., V.W.; writing—review and editing, G.Z., J.I.B.G., F.A.C.X., A.K.S., J.C.C.; supervision, A.K.S., J.C.C.; project administration, A.K.S., J.C.C.; funding acquisition, J.C.C. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
Data sharing is not applicable.
Acknowledgments
P.G.S.: A.M.A., F.B. and I.G.M are supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brazil (CAPES)—Finance Code 001. G.Z., D.M. and J.C.C are supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS). A.K.S. is supported by grants from the National Institutes of Health (National Institute for Aging grants, R01AG075440–01 and 1RF1AG074256–01).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell Mol Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Klumperman, J.; Raposo, G. The complex ultrastructure of the endolysosomal system. Cold Spring Harb Perspect Biol. 2014, 6, a016857. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Thery, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol. 2013, 113, 1–11. [Google Scholar] [CrossRef]
- Stoorvogel, W.; Strous, G.J.; Geuze, H.J.; Oorschot, V.; Schwartz, A.L. Late endosomes derive from early endosomes by maturation. Cell 1991, 65, 417–427. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science. 2020, 367. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef]
- Pols, M.S.; Klumperman, J. Trafficking and function of the tetraspanin CD63. Exp Cell Res. 2009, 315, 1584–1592. [Google Scholar] [CrossRef]
- Colombo, M.; Moita, C.; van Niel, G.; Kowal, J.; Vigneron, J.; Benaroch, P.; et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci. 2013, 126 Pt 24, 5553–5565. [Google Scholar] [CrossRef]
- Baietti, M.F.; Zhang, Z.; Mortier, E.; Melchior, A.; Degeest, G.; Geeraerts, A.; et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol. 2012, 14, 677–685. [Google Scholar] [CrossRef]
- Wei, D.; Zhan, W.; Gao, Y.; Huang, L.; Gong, R.; Wang, W.; et al. RAB31 marks and controls an ESCRT-independent exosome pathway. Cell Res. 2021, 31, 157–177. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ahn, J.S.; Kim, S.; Kim, H.J.; Kim, S.H.; Kang, J.S. The potential theragnostic (diagnostic+therapeutic) application of exosomes in diverse biomedical fields. Korean J Physiol Pharmacol. 2018, 22, 113–125. [Google Scholar] [CrossRef]
- Jin, H.; Tang, Y.; Yang, L.; Peng, X.; Li, B.; Fan, Q.; et al. Rab GTPases: Central Coordinators of Membrane Trafficking in Cancer. Front Cell Dev Biol. 2021, 9, 648384. [Google Scholar] [CrossRef]
- Hsu, C.; Morohashi, Y.; Yoshimura, S.; Manrique-Hoyos, N.; Jung, S.; Lauterbach, M.A.; et al. Regulation of exosome secretion by Rab35 and its GTPase-activating proteins TBC1D10A-C. J Cell Biol. 2010, 189, 223–232. [Google Scholar] [CrossRef]
- Holz, R.W.; Zimmerberg, J. Dynamic Relationship of the SNARE Complex with a Membrane. Biophys J. 2019, 117, 627–630. [Google Scholar] [CrossRef]
- Verweij, F.J.; Bebelman, M.P.; Jimenez, C.R.; Garcia-Vallejo, J.J.; Janssen, H.; Neefjes, J.; et al. Quantifying exosome secretion from single cells reveals a modulatory role for GPCR signaling. J Cell Biol. 2018, 217, 1129–1142. [Google Scholar] [CrossRef]
- Logan, M.R.; Lacy, P.; Odemuyiwa, S.O.; Steward, M.; Davoine, F.; Kita, H.; et al. A critical role for vesicle-associated membrane protein-7 in exocytosis from human eosinophils and neutrophils. Allergy. 2006, 61, 777–784. [Google Scholar] [CrossRef]
- Puri, N.; Roche, P.A. Mast cells possess distinct secretory granule subsets whose exocytosis is regulated by different SNARE isoforms. Proc Natl Acad Sci U S A. 2008, 105, 2580–2585. [Google Scholar] [CrossRef]
- Sun, M.; Xue, X.; Li, L.; Xu, D.; Li, S.; Li, S.C.; et al. Ectosome biogenesis and release processes observed by using live-cell dynamic imaging in mammalian glial cells. Quant Imaging Med Surg. 2021, 11, 4604–4616. [Google Scholar] [CrossRef]
- Tricarico, C.; Clancy, J.; D'Souza-Schorey, C. Biology and biogenesis of shed microvesicles. Small GTPases. 2017, 8, 220–232. [Google Scholar] [CrossRef]
- D'Souza-Schorey, C.; Clancy, J.W. Tumor-derived microvesicles: shedding light on novel microenvironment modulators and prospective cancer biomarkers. Genes Dev. 2012, 26, 1287–1299. [Google Scholar] [CrossRef]
- Bucki, R.; Bachelot-Loza, C.; Zachowski, A.; Giraud, F.; Sulpice, J.C. Calcium induces phospholipid redistribution and microvesicle release in human erythrocyte membranes by independent pathways. Biochemistry. 1998, 37, 15383–15391. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018, 7, 1535750. [Google Scholar] [CrossRef]
- You, J.; Fu, Z.; Zou, L. Mechanism and Potential of Extracellular Vesicles Derived From Mesenchymal Stem Cells for the Treatment of Infectious Diseases. Front Microbiol. 2021, 12, 761338. [Google Scholar] [CrossRef]
- Gowen, A.; Shahjin, F.; Chand, S.; Odegaard, K.E.; Yelamanchili, S.V. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Challenges in Clinical Applications. Front Cell Dev Biol. 2020, 8, 149. [Google Scholar] [CrossRef]
- Yin, K.; Wang, S.; Zhao, R.C. Exosomes from mesenchymal stem/stromal cells: a new therapeutic paradigm. Biomark Res. 2019, 7, 8. [Google Scholar] [CrossRef]
- Lopez-Guerrero, J.A.; Ripa, I.; Andreu, S.; Bello-Morales, R. The Role of Extracellular Vesicles in Demyelination of the Central Nervous System. Int J Mol Sci. 2020, 21. [Google Scholar] [CrossRef]
- Horn, M.D.; MacLean, A.G. Extracellular Vesicles as a Means of Viral Immune Evasion, CNS Invasion, and Glia-Induced Neurodegeneration. Front Cell Neurosci. 2021, 15, 695899. [Google Scholar] [CrossRef]
- Zhou, W.; Woodson, M.; Sherman, M.B.; Neelakanta, G.; Sultana, H. Exosomes mediate Zika virus transmission through SMPD3 neutral Sphingomyelinase in cortical neurons. Emerg Microbes Infect. 2019, 8, 307–326. [Google Scholar] [CrossRef]
- Pulliam, L.; Sun, B.; Mustapic, M.; Chawla, S.; Kapogiannis, D. Plasma neuronal exosomes serve as biomarkers of cognitive impairment in HIV infection and Alzheimer's disease. J Neurovirol. 2019, 25, 702–709. [Google Scholar] [CrossRef]
- Koniusz, S.; Andrzejewska, A.; Muraca, M.; Srivastava, A.K.; Janowski, M.; Lukomska, B. Extracellular Vesicles in Physiology, Pathology, and Therapy of the Immune and Central Nervous System, with Focus on Extracellular Vesicles Derived from Mesenchymal Stem Cells as Therapeutic Tools. Front Cell Neurosci. 2016, 10, 109. [Google Scholar] [CrossRef]
- Schiera, G.; Di Liegro, C.M.; Di Liegro, I. Extracellular Membrane Vesicles as Vehicles for Brain Cell-to-Cell Interactions in Physiological as well as Pathological Conditions. Biomed Res Int. 2015, 2015, 152926. [Google Scholar] [CrossRef]
- Gassama, Y.; Favereaux, A. Emerging Roles of Extracellular Vesicles in the Central Nervous System: Physiology, Pathology, and Therapeutic Perspectives. Front Cell Neurosci. 2021, 15, 626043. [Google Scholar] [CrossRef]
- Andras, I.E.; Toborek, M. Extracellular vesicles of the blood-brain barrier. Tissue Barriers. 2016, 4, e1131804. [Google Scholar] [CrossRef]
- Fruhbeis, C.; Frohlich, D.; Kramer-Albers, E.M. Emerging roles of exosomes in neuron-glia communication. Front Physiol. 2012, 3, 119. [Google Scholar] [CrossRef]
- Delpech, J.C.; Herron, S.; Botros, M.B.; Ikezu, T. Neuroimmune Crosstalk through Extracellular Vesicles in Health and Disease. Trends Neurosci. 2019, 42, 361–372. [Google Scholar] [CrossRef]
- Kurachi, M.; Mikuni, M.; Ishizaki, Y. Extracellular Vesicles from Vascular Endothelial Cells Promote Survival, Proliferation and Motility of Oligodendrocyte Precursor Cells. Plos One. 2016, 11, e0159158. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, Y.; Du, X.F.; Li, J.; Zi, H.X.; Bu, J.W.; et al. Neurons secrete miR-132-containing exosomes to regulate brain vascular integrity. Cell Res. 2017, 27, 882–897. [Google Scholar] [CrossRef]
- Schiera, G.; Proia, P.; Alberti, C.; Mineo, M.; Savettieri, G.; Di Liegro, I. Neurons produce FGF2 and VEGF and secrete them at least in part by shedding extracellular vesicles. J Cell Mol Med. 2007, 11, 1384–1394. [Google Scholar] [CrossRef] [PubMed]
- Chivet, M.; Javalet, C.; Laulagnier, K.; Blot, B.; Hemming, F.J.; Sadoul, R. Exosomes secreted by cortical neurons upon glutamatergic synapse activation specifically interact with neurons. J Extracell Vesicles. 2014, 3, 24722. [Google Scholar] [CrossRef] [PubMed]
- Lachenal, G.; Pernet-Gallay, K.; Chivet, M.; Hemming, F.J.; Belly, A.; Bodon, G.; et al. Release of exosomes from differentiated neurons and its regulation by synaptic glutamatergic activity. Mol Cell Neurosci. 2011, 46, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Men, Y.; Yelick, J.; Jin, S.; Tian, Y.; Chiang, M.S.R.; Higashimori, H.; et al. Exosome reporter mice reveal the involvement of exosomes in mediating neuron to astroglia communication in the CNS. Nat Commun. 2019, 10, 4136. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Korner, R.; Gaitanos, L.; Klein, R. Exosomes mediate cell contact-independent ephrin-Eph signaling during axon guidance. J Cell Biol. 2016, 214, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Goldie, B.J.; Dun, M.D.; Lin, M.; Smith, N.D.; Verrills, N.M.; Dayas, C.V.; et al. Activity-associated miRNA are packaged in Map1b-enriched exosomes released from depolarized neurons. Nucleic Acids Res. 2014, 42, 9195–9208. [Google Scholar] [CrossRef] [PubMed]
- Fruhbeis, C.; Frohlich, D.; Kuo, W.P.; Amphornrat, J.; Thilemann, S.; Saab, A.S.; et al. Neurotransmitter-triggered transfer of exosomes mediates oligodendrocyte-neuron communication. PLoS Biol. 2013, 11, e1001604. [Google Scholar] [CrossRef]
- Faure, J.; Lachenal, G.; Court, M.; Hirrlinger, J.; Chatellard-Causse, C.; Blot, B.; et al. Exosomes are released by cultured cortical neurones. Mol Cell Neurosci. 2006, 31, 642–648. [Google Scholar] [CrossRef]
- Antonucci, F.; Turola, E.; Riganti, L.; Caleo, M.; Gabrielli, M.; Perrotta, C.; et al. Microvesicles released from microglia stimulate synaptic activity via enhanced sphingolipid metabolism. EMBO J. 2012, 31, 1231–1240. [Google Scholar] [CrossRef]
- Eyo, U.B.; Wu, L.J. Bidirectional microglia-neuron communication in the healthy brain. Neural Plast. 2013, 2013, 456857. [Google Scholar] [CrossRef]
- Morel, L.; Regan, M.; Higashimori, H.; Ng, S.K.; Esau, C.; Vidensky, S.; et al. Neuronal exosomal miRNA-dependent translational regulation of astroglial glutamate transporter GLT1. J Biol Chem. 2013, 288, 7105–7116. [Google Scholar] [CrossRef]
- Bahrini, I.; Song, J.H.; Diez, D.; Hanayama, R. Neuronal exosomes facilitate synaptic pruning by up-regulating complement factors in microglia. Sci Rep. 2015, 5, 7989. [Google Scholar] [CrossRef]
- Lemaire, Q.; Raffo-Romero, A.; Arab, T.; Van Camp, C.; Drago, F.; Forte, S.; et al. Isolation of microglia-derived extracellular vesicles: towards miRNA signatures and neuroprotection. J Nanobiotechnology. 2019, 17, 119. [Google Scholar] [CrossRef] [PubMed]
- Van den Broek, B.; Pintelon, I.; Hamad, I.; Kessels, S.; Haidar, M.; Hellings, N.; et al. Microglial derived extracellular vesicles activate autophagy and mediate multi-target signaling to maintain cellular homeostasis. J Extracell Vesicles. 2020, 10, e12022. [Google Scholar] [CrossRef] [PubMed]
- Paolicelli, R.C.; Bergamini, G.; Rajendran, L. Cell-to-cell Communication by Extracellular Vesicles: Focus on Microglia. Neuroscience. 2019, 405, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Murgoci, A.N.; Duhamel, M.; Raffo-Romero, A.; Mallah, K.; Aboulouard, S.; Lefebvre, C.; et al. Location of neonatal microglia drives small extracellular vesicles content and biological functions in vitro. J Extracell Vesicles. 2020, 9, 1727637. [Google Scholar] [CrossRef] [PubMed]
- Prada, I.; Gabrielli, M.; Turola, E.; Iorio, A.; D'Arrigo, G.; Parolisi, R.; et al. Glia-to-neuron transfer of miRNAs via extracellular vesicles: a new mechanism underlying inflammation-induced synaptic alterations. Acta Neuropathol. 2018, 135, 529–550. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Sun, H.; Usuki, S.; Sakai, S.; Hanamatsu, H.; Mioka, T.; et al. A potential function for neuronal exosomes: sequestering intracerebral amyloid-beta peptide. FEBS Lett. 2015, 589, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Willis, C.M.; Nicaise, A.M.; Bongarzone, E.R.; Givogri, M.; Reiter, C.R.; Heintz, O.; et al. Astrocyte Support for Oligodendrocyte Differentiation can be Conveyed via Extracellular Vesicles but Diminishes with Age. Sci Rep. 2020, 10, 828. [Google Scholar] [CrossRef] [PubMed]
- Luarte, A.; Henzi, R.; Fernandez, A.; Gaete, D.; Cisternas, P.; Pizarro, M.; et al. Astrocyte-Derived Small Extracellular Vesicles Regulate Dendritic Complexity through miR-26a-5p Activity. Cells. 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Venturini, A.; Passalacqua, M.; Pelassa, S.; Pastorino, F.; Tedesco, M.; Cortese, K.; et al. Exosomes From Astrocyte Processes: Signaling to Neurons. Front Pharmacol. 2019, 10, 1452. [Google Scholar] [CrossRef]
- Proia, P.; Schiera, G.; Mineo, M.; Ingrassia, A.M.; Santoro, G.; Savettieri, G.; et al. Astrocytes shed extracellular vesicles that contain fibroblast growth factor-2 and vascular endothelial growth factor. Int J Mol Med. 2008, 21, 63–67. [Google Scholar] [CrossRef]
- Stronati, E.; Conti, R.; Cacci, E.; Cardarelli, S.; Biagioni, S.; Poiana, G. Extracellular Vesicle-Induced Differentiation of Neural Stem Progenitor Cells. Int J Mol Sci. 2019, 20. [Google Scholar] [CrossRef]
- Raffaele, S.; Lombardi, M.; Verderio, C.; Fumagalli, M. TNF Production and Release from Microglia via Extracellular Vesicles: Impact on Brain Functions. Cells. 2020, 9. [Google Scholar] [CrossRef]
- Bianco, F.; Perrotta, C.; Novellino, L.; Francolini, M.; Riganti, L.; Menna, E.; et al. Acid sphingomyelinase activity triggers microparticle release from glial cells. EMBO J. 2009, 28, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Borgmann, K.; Edara, V.V.; Stacy, S.; Ghorpade, A.; Ikezu, T. Activated human astrocyte-derived extracellular vesicles modulate neuronal uptake, differentiation and firing. J Extracell Vesicles. 2020, 9, 1706801. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Sheng, S.; Wang, Y.; Ding, L.; Xu, X.; Xia, X.; et al. Astrocyte-derived extracellular vesicles: A double-edged sword in central nervous system disorders. Neurosci Biobehav Rev. 2021, 125, 148–159. [Google Scholar] [CrossRef]
- Sun, H.; Cao, X.; Gong, A.; Huang, Y.; Xu, Y.; Zhang, J.; et al. Extracellular vesicles derived from astrocytes facilitated neurite elongation by activating the Hippo pathway. Exp Cell Res. 2022, 411, 112937. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; Knosche, T.R. Action potential propagation and synchronisation in myelinated axons. PLoS Comput Biol. 2019, 15, e1007004. [Google Scholar] [CrossRef] [PubMed]
- Fruhbeis, C.; Kuo-Elsner, W.P.; Muller, C.; Barth, K.; Peris, L.; Tenzer, S.; et al. Oligodendrocytes support axonal transport and maintenance via exosome secretion. PLoS Biol. 2020, 18, e3000621. [Google Scholar] [CrossRef] [PubMed]
- Kramer-Albers, E.M. Extracellular vesicles in the oligodendrocyte microenvironment. Neurosci Lett. 2020, 725, 134915. [Google Scholar] [CrossRef]
- Kramer-Albers, E.M.; Bretz, N.; Tenzer, S.; Winterstein, C.; Mobius, W.; Berger, H.; et al. Oligodendrocytes secrete exosomes containing major myelin and stress-protective proteins: Trophic support for axons? Proteomics Clin Appl. 2007, 1, 1446–1461. [Google Scholar] [CrossRef]
- Lizarraga-Valderrama, L.R.; Sheridan, G.K. Extracellular vesicles and intercellular communication in the central nervous system. FEBS Lett. 2021, 595, 1391–1410. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Saint-Pol, J.; Gosselet, F.; Duban-Deweer, S.; Pottiez, G.; Karamanos, Y. Targeting and Crossing the Blood-Brain Barrier with Extracellular Vesicles. Cells. 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Mustapic, M.; Eitan, E.; Werner, J.K., Jr; Berkowitz, S.T.; Lazaropoulos, M.P.; Tran, J.; et al. Plasma Extracellular Vesicles Enriched for Neuronal Origin: A Potential Window into Brain Pathologic Processes. Front Neurosci. 2017, 11, 278. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.D.; Gercel-Taylor, C. The origin, function, and diagnostic potential of RNA within extracellular vesicles present in human biological fluids. Front Genet. 2013, 4, 142. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, Y.; Huang, Y.; Zhang, H.; Lu, H.; Zheng, J.C. Exosomal miRNAs in central nervous system diseases: biomarkers, pathological mediators, protective factors and therapeutic agents. Prog Neurobiol. 2019, 183, 101694. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.F.; Wu, N.; Wang, L.; Li, J. Circulating microRNAs: a novel class of potential biomarkers for diagnosing and prognosing central nervous system diseases. Cell Mol Neurobiol. 2013, 33, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Hansson, O. Biomarkers for neurodegenerative diseases. Nat Med. 2021, 27, 954–963. [Google Scholar] [CrossRef]
- DeTure, M.A.; Dickson, D.W. The neuropathological diagnosis of Alzheimer's disease. Mol Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef]
- Kim, K.Y.; Shin, K.Y.; Chang, K.A. Brain-Derived Exosomal Proteins as Effective Biomarkers for Alzheimer's Disease: A Systematic Review and Meta-Analysis. Biomolecules 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Lin, H.; He, X.; Chen, L.; Dai, Y.; Jia, W.; et al. Neurogranin as a cognitive biomarker in cerebrospinal fluid and blood exosomes for Alzheimer's disease and mild cognitive impairment. Transl Psychiatry. 2020, 10, 125. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Zhu, M.; Kong, C.; Pang, Y.; Zhang, H.; Qiu, Q.; et al. Blood neuro-exosomal synaptic proteins predict Alzheimer's disease at the asymptomatic stage. Alzheimers Dement. 2021, 17, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Yang, G.; Yan, Q.Q.; Zhao, J.; Li, S. Exosome-encapsulated microRNAs as promising biomarkers for Alzheimer's disease. Rev Neurosci. 2019, 31, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.H.; Zhu, J.; Wang, Y.C.; Wu, J.; Liu, J.R.; Guo, H.D. Effects of exosomal miRNAs in the diagnosis and treatment of Alzheimer's disease. Mech Ageing Dev. 2021, 200, 111593. [Google Scholar] [CrossRef] [PubMed]
- Van Giau, V.; An, S.S. Emergence of exosomal miRNAs as a diagnostic biomarker for Alzheimer's disease. J Neurol Sci. 2016, 360, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Zhao, B.; Zhao, J.; Li, S. Potential Roles of Exosomal MicroRNAs as Diagnostic Biomarkers and Therapeutic Application in Alzheimer's Disease. Neural Plast. 2017, 2017, 7027380. [Google Scholar] [CrossRef] [PubMed]
- Durur, D.Y.; Tastan, B.; Ugur Tufekci, K.; Olcum, M.; Uzuner, H.; Karakulah, G.; et al. Alteration of miRNAs in Small Neuron-Derived Extracellular Vesicles of Alzheimer's Disease Patients and the Effect of Extracellular Vesicles on Microglial Immune Responses. J Mol Neurosci. 2022, 72, 1182–1194. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.T.; Liu, C.G.; Gao, S.C.; Zhang, Y.; Wang, P.C. The Serum Exosome Derived MicroRNA-135a, -193b, and -384 Were Potential Alzheimer's Disease Biomarkers. Biomed Environ Sci. 2018, 31, 87–96. [Google Scholar]
- Vogrinc, D.; Goricar, K.; Kunej, T.; Dolzan, V. Systematic Search for Novel Circulating Biomarkers Associated with Extracellular Vesicles in Alzheimer's Disease: Combining Literature Screening and Database Mining Approaches. J Pers Med. 2021, 11. [Google Scholar] [CrossRef]
- Serpente, M.; Fenoglio, C.; D'Anca, M.; Arcaro, M.; Sorrentino, F.; Visconte, C.; et al. MiRNA Profiling in Plasma Neural-Derived Small Extracellular Vesicles from Patients with Alzheimer's Disease. Cells. 2020, 9. [Google Scholar] [CrossRef]
- Dong, Z.; Gu, H.; Guo, Q.; Liang, S.; Xue, J.; Yao, F.; et al. Profiling of Serum Exosome MiRNA Reveals the Potential of a MiRNA Panel as Diagnostic Biomarker for Alzheimer's Disease. Mol Neurobiol. 2021, 58, 3084–3094. [Google Scholar] [CrossRef]
- Li, F.; Xie, X.Y.; Sui, X.F.; Wang, P.; Chen, Z.; Zhang, J.B. Profile of Pathogenic Proteins and MicroRNAs in Plasma-derived Extracellular Vesicles in Alzheimer's Disease: A Pilot Study. Neuroscience. 2020, 432, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Aharon, A.; Spector, P.; Ahmad, R.S.; Horrany, N.; Sabbach, A.; Brenner, B.; et al. Extracellular Vesicles of Alzheimer's Disease Patients as a Biomarker for Disease Progression. Mol Neurobiol. 2020, 57, 4156–4169. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; et al. Parkinson disease. Nat Rev Dis Primers. 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, G.; Loov, C.; Persson, E.; Lazaro, D.F.; Takeda, S.; Bergstrom, J.; et al. Secretion and Uptake of alpha-Synuclein Via Extracellular Vesicles in Cultured Cells. Cell Mol Neurobiol. 2018, 38, 1539–1550. [Google Scholar] [CrossRef]
- Shi, M.; Liu, C.; Cook, T.J.; Bullock, K.M.; Zhao, Y.; Ginghina, C.; et al. Plasma exosomal alpha-synuclein is likely CNS-derived and increased in Parkinson's disease. Acta Neuropathol. 2014, 128, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Stuendl, A.; Kraus, T.; Chatterjee, M.; Zapke, B.; Sadowski, B.; Moebius, W.; et al. alpha-Synuclein in Plasma-Derived Extracellular Vesicles Is a Potential Biomarker of Parkinson's Disease. Mov Disord. 2021, 36, 2508–2518. [Google Scholar] [CrossRef]
- Niu, M.; Li, Y.; Li, G.; Zhou, L.; Luo, N.; Yao, M.; et al. A longitudinal study on alpha-synuclein in plasma neuronal exosomes as a biomarker for Parkinson's disease development and progression. Eur J Neurol. 2020, 27, 967–974. [Google Scholar] [CrossRef]
- Jiang, C.; Hopfner, F.; Katsikoudi, A.; Hein, R.; Catli, C.; Evetts, S.; et al. Serum neuronal exosomes predict and differentiate Parkinson's disease from atypical parkinsonism. J Neurol Neurosurg Psychiatry. 2020, 91, 720–729. [Google Scholar] [CrossRef]
- Hong, Z.; Tian, C.; Stewart, T.; Aro, P.; Soltys, D.; Bercow, M.; et al. Development of a Sensitive Diagnostic Assay for Parkinson Disease Quantifying alpha-Synuclein-Containing Extracellular Vesicles. Neurology. 2021, 96, e2332–e45. [Google Scholar] [CrossRef] [PubMed]
- Si, X.; Tian, J.; Chen, Y.; Yan, Y.; Pu, J.; Zhang, B. Central Nervous System-Derived Exosomal Alpha-Synuclein in Serum May Be a Biomarker in Parkinson's Disease. Neuroscience. 2019, 413, 308–316. [Google Scholar] [CrossRef]
- Cao, Z.; Wu, Y.; Liu, G.; Jiang, Y.; Wang, X.; Wang, Z.; et al. alpha-Synuclein in salivary extracellular vesicles as a potential biomarker of Parkinson's disease. Neurosci Lett. 2019, 696, 114–120. [Google Scholar] [CrossRef]
- Xie, S.; Niu, W.; Xu, F.; Wang, Y.; Hu, S.; Niu, C. Differential expression and significance of miRNAs in plasma extracellular vesicles of patients with Parkinson's disease. Int J Neurosci. 2022, 132, 673–688. [Google Scholar] [CrossRef]
- Cao, X.Y.; Lu, J.M.; Zhao, Z.Q.; Li, M.C.; Lu, T.; An, X.S.; et al. MicroRNA biomarkers of Parkinson's disease in serum exosome-like microvesicles. Neurosci Lett. 2017, 644, 94–99. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Huang, L.; Shao, C.; Nie, T.; Xia, L.; Cui, B.; et al. Several miRNAs derived from serum extracellular vesicles are potential biomarkers for early diagnosis and progression of Parkinson's disease. Transl Neurodegener. 2021, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Grossi, I.; Radeghieri, A.; Paolini, L.; Porrini, V.; Pilotto, A.; Padovani, A.; et al. MicroRNA-34a-5p expression in the plasma and in its extracellular vesicle fractions in subjects with Parkinson's disease: An exploratory study. Int J Mol Med. 2021, 47, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Anastasi, F.; Masciandaro, S.M.; Carratore, R.D.; Dell'Anno, M.T.; Signore, G.; Falleni, A.; et al. Proteomics Profiling of Neuron-Derived Small Extracellular Vesicles from Human Plasma: Enabling Single-Subject Analysis. Int J Mol Sci. 2021, 22. [Google Scholar] [CrossRef]
- Kitamura, Y.; Kojima, M.; Kurosawa, T.; Sasaki, R.; Ichihara, S.; Hiraku, Y.; et al. Proteomic Profiling of Exosomal Proteins for Blood-based Biomarkers in Parkinson's Disease. Neuroscience. 2018, 392, 121–128. [Google Scholar] [CrossRef]
- Jiang, R.; Rong, C.; Ke, R.; Meng, S.; Yan, X.; Ke, H.; et al. Differential proteomic analysis of serum exosomes reveals alterations in progression of Parkinson disease. Medicine (Baltimore). 2019, 98, e17478. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kojima, K.; Mobley, J.A.; West, A.B. Proteomic analysis of urinary extracellular vesicles reveal biomarkers for neurologic disease. EBioMedicine. 2019, 45, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.H.; Chen, Z.T.; Zhou, R.L.; Zhang, X.; Ye, Q.Y.; Wang, Y.Z. Increased DJ-1 and alpha-Synuclein in Plasma Neural-Derived Exosomes as Potential Markers for Parkinson's Disease. Front Aging Neurosci. 2018, 10, 438. [Google Scholar] [CrossRef]
- Ho, D.H.; Yi, S.; Seo, H.; Son, I.; Seol, W. Increased DJ-1 in urine exosome of Korean males with Parkinson's disease. Biomed Res Int. 2014, 2014, 704678. [Google Scholar] [CrossRef] [PubMed]
- Fraser, K.B.; Rawlins, A.B.; Clark, R.G.; Alcalay, R.N.; Standaert, D.G.; Liu, N.; et al. Ser(P)-1292 LRRK2 in urinary exosomes is elevated in idiopathic Parkinson's disease. Mov Disord. 2016, 31, 1543–1550. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.; Chung, C.C.; Chen, J.H.; Yu, R.C.; Hong, C.T. Cytokine Profile in Plasma Extracellular Vesicles of Parkinson's Disease and the Association with Cognitive Function. Cells. 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- McColgan, P.; Tabrizi, S.J. Huntington's disease: a clinical review. Eur J Neurol. 2018, 25, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Abels, E.R.; Redzic, J.S.; Margulis, J.; Finkbeiner, S.; Breakefield, X.O. Potential Transfer of Polyglutamine and CAG-Repeat RNA in Extracellular Vesicles in Huntington's Disease: Background and Evaluation in Cell Culture. Cell Mol Neurobiol. 2016, 36, 459–470. [Google Scholar] [CrossRef]
- Denis, H.L.; Lamontagne-Proulx, J.; St-Amour, I.; Mason, S.L.; Rowley, J.W.; Cloutier, N.; et al. Platelet abnormalities in Huntington's disease. J Neurol Neurosurg Psychiatry. 2019, 90, 272–283. [Google Scholar] [CrossRef]
- Denis, H.L.; Lamontagne-Proulx, J.; St-Amour, I.; Mason, S.L.; Weiss, A.; Chouinard, S.; et al. Platelet-derived extracellular vesicles in Huntington's disease. J Neurol. 2018, 265, 2704–2712. [Google Scholar] [CrossRef]
- Ciregia, F.; Urbani, A.; Palmisano, G. Extracellular Vesicles in Brain Tumors and Neurodegenerative Diseases. Front Mol Neurosci. 2017, 10, 276. [Google Scholar] [CrossRef]
- Dobson, R.; Giovannoni, G. Multiple sclerosis - a review. Eur J Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Scaroni, F.; Visconte, C.; Serpente, M.; Golia, M.T.; Gabrielli, M.; Huiskamp, M.; et al. miR-150-5p and let-7b-5p in Blood Myeloid Extracellular Vesicles Track Cognitive Symptoms in Patients with Multiple Sclerosis. Cells. 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Azimi, M.; Ghabaee, M.; Naser Moghadasi, A.; Izad, M. Altered Expression of miR-326 in T Cell-derived Exosomes of Patients with Relapsing-remitting Multiple Sclerosis. Iran J Allergy Asthma Immunol. 2019, 18, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Saenz-Cuesta, M.; Alberro, A.; Munoz-Culla, M.; Osorio-Querejeta, I.; Fernandez-Mercado, M.; Lopetegui, I.; et al. The First Dose of Fingolimod Affects Circulating Extracellular Vesicles in Multiple Sclerosis Patients. Int J Mol Sci. 2018, 19. [Google Scholar] [CrossRef]
- Blonda, M.; Amoruso, A.; Grasso, R.; Di Francescantonio, V.; Avolio, C. Multiple Sclerosis Treatments Affect Monocyte-Derived Microvesicle Production. Front Neurol. 2017, 8, 422. [Google Scholar] [CrossRef]
- Amoruso, A.; Blonda, M.; D'Arrigo, G.; Grasso, R.; Di Francescantonio, V.; Verderio, C.; et al. Effect of fingolimod action on the release of monocyte-derived microvesicles in multiple sclerosis patients. J Neuroimmunol. 2018, 323, 43–48. [Google Scholar] [CrossRef]
- Ebrahimkhani, S.; Beadnall, H.N.; Wang, C.; Suter, C.M.; Barnett, M.H.; Buckland, M.E.; et al. Serum Exosome MicroRNAs Predict Multiple Sclerosis Disease Activity after Fingolimod Treatment. Mol Neurobiol. 2020, 57, 1245–1258. [Google Scholar] [CrossRef]
- Cuomo-Haymour, N.; Bergamini, G.; Russo, G.; Kulic, L.; Knuesel, I.; Martin, R.; et al. Differential Expression of Serum Extracellular Vesicle miRNAs in Multiple Sclerosis: Disease-Stage Specificity and Relevance to Pathophysiology. Int J Mol Sci. 2022, 23. [Google Scholar] [CrossRef]
- Selmaj, I.; Cichalewska, M.; Namiecinska, M.; Galazka, G.; Horzelski, W.; Selmaj, K.W.; et al. Global exosome transcriptome profiling reveals biomarkers for multiple sclerosis. Ann Neurol. 2017, 81, 703–717. [Google Scholar] [CrossRef]
- Groen, K.; Maltby, V.E.; Scott, R.J.; Tajouri, L.; Lechner-Scott, J. Erythrocyte microRNAs show biomarker potential and implicate multiple sclerosis susceptibility genes. Clin Transl Med. 2020, 10, 74–90. [Google Scholar] [CrossRef]
- Ebrahimkhani, S.; Vafaee, F.; Young, P.E.; Hur, S.S.J.; Hawke, S.; Devenney, E.; et al. Exosomal microRNA signatures in multiple sclerosis reflect disease status. Sci Rep. 2017, 7, 14293. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, P.; Nogueras-Ortiz, C.; Kim, S.; Delgado-Peraza, F.; Calabresi, P.A.; Kapogiannis, D. Synaptic and complement markers in extracellular vesicles in multiple sclerosis. Mult Scler. 2021, 27, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Moyano, A.L.; Li, G.; Boullerne, A.I.; Feinstein, D.L.; Hartman, E.; Skias, D.; et al. Sulfatides in extracellular vesicles isolated from plasma of multiple sclerosis patients. J Neurosci Res. 2016, 94, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Galazka, G.; Mycko, M.P.; Selmaj, I.; Raine, C.S.; Selmaj, K.W. Multiple sclerosis: Serum-derived exosomes express myelin proteins. Mult Scler. 2018, 24, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Mazzucco, M.; Mannheim, W.; Shetty, S.V.; Linden, J.R. CNS endothelial derived extracellular vesicles are biomarkers of active disease in multiple sclerosis. Fluids Barriers CNS. 2022, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.S.; Acevedo, C.; Arzimanoglou, A.; Bogacz, A.; Cross, J.H.; Elger, C.E.; et al. ILAE official report: a practical clinical definition of epilepsy. Epilepsia. 2014, 55, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Karttunen, J.; Heiskanen, M.; Lipponen, A.; Poulsen, D.; Pitkanen, A. Extracellular Vesicles as Diagnostics and Therapeutics for Structural Epilepsies. Int J Mol Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Raoof, R.; Jimenez-Mateos, E.M.; Bauer, S.; Tackenberg, B.; Rosenow, F.; Lang, J.; et al. Cerebrospinal fluid microRNAs are potential biomarkers of temporal lobe epilepsy and status epilepticus. Sci Rep. 2017, 7, 3328. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Zhang, H.; Xie, W.; Meng, F.; Zhang, K.; Jiang, Y.; et al. Altered microRNA profiles in plasma exosomes from mesial temporal lobe epilepsy with hippocampal sclerosis. Oncotarget. 2017, 8, 4136–4146. [Google Scholar] [CrossRef]
- Gitai, D.L.G.; Dos Santos, Y.D.R.; Upadhya, R.; Kodali, M.; Madhu, L.N.; Shetty, A.K. Extracellular Vesicles in the Forebrain Display Reduced miR-346 and miR-331-3p in a Rat Model of Chronic Temporal Lobe Epilepsy. Mol Neurobiol. 2020, 57, 1674–1687. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Chen, Y.; Hua, Y.; Xu, L.; Zhu, M.; et al. Circulating MicroRNAs From Plasma Small Extracellular Vesicles as Potential Diagnostic Biomarkers in Pediatric Epilepsy and Drug-Resistant Epilepsy. Front Mol Neurosci. 2022, 15, 823802. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.D.; Pan, H.Y.; Huang, J.B.; Liu, X.P.; Li, J.H.; Ho, C.J.; et al. Circulating MicroRNAs from Serum Exosomes May Serve as a Putative Biomarker in the Diagnosis and Treatment of Patients with Focal Cortical Dysplasia. Cells. 2020, 9. [Google Scholar] [CrossRef]
- Lin, Z.; Gu, Y.; Zhou, R.; Wang, M.; Guo, Y.; Chen, Y.; et al. Serum Exosomal Proteins F9 and TSP-1 as Potential Diagnostic Biomarkers for Newly Diagnosed Epilepsy. Front Neurosci. 2020, 14, 737. [Google Scholar] [CrossRef] [PubMed]
- Blennow, K.; Brody, D.L.; Kochanek, P.M.; Levin, H.; McKee, A.; Ribbers, G.M.; et al. Traumatic brain injuries. Nat Rev Dis Primers. 2016, 2, 16084. [Google Scholar] [CrossRef] [PubMed]
- Beard, K.; Yang, Z.; Haber, M.; Flamholz, M.; Diaz-Arrastia, R.; Sandsmark, D.; et al. Extracellular vesicles as distinct biomarker reservoirs for mild traumatic brain injury diagnosis. Brain Commun. 2021, 3, fcab151. [Google Scholar] [CrossRef] [PubMed]
- Mondello, S.; Guedes, V.A.; Lai, C.; Czeiter, E.; Amrein, K.; Kobeissy, F.; et al. Circulating Brain Injury Exosomal Proteins following Moderate-To-Severe Traumatic Brain Injury: Temporal Profile, Outcome Prediction and Therapy Implications. Cells. 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Manek, R.; Moghieb, A.; Yang, Z.; Kumar, D.; Kobessiy, F.; Sarkis, G.A.; et al. Protein Biomarkers and Neuroproteomics Characterization of Microvesicles/Exosomes from Human Cerebrospinal Fluid Following Traumatic Brain Injury. Mol Neurobiol. 2018, 55, 6112–6128. [Google Scholar] [CrossRef] [PubMed]
- Goetzl, E.J.; Elahi, F.M.; Mustapic, M.; Kapogiannis, D.; Pryhoda, M.; Gilmore, A.; et al. Altered levels of plasma neuron-derived exosomes and their cargo proteins characterize acute and chronic mild traumatic brain injury. FASEB J. 2019, 33, 5082–5088. [Google Scholar] [CrossRef]
- Cheng, Y.; Pereira, M.; Raukar, N.; Reagan, J.L.; Queseneberry, M.; Goldberg, L.; et al. Potential biomarkers to detect traumatic brain injury by the profiling of salivary extracellular vesicles. J Cell Physiol. 2019, 234, 14377–14388. [Google Scholar] [CrossRef]
- Gill, J.; Mustapic, M.; Diaz-Arrastia, R.; Lange, R.; Gulyani, S.; Diehl, T.; et al. Higher exosomal tau, amyloid-beta 42 and IL-10 are associated with mild TBIs and chronic symptoms in military personnel. Brain Inj. 2018, 32, 1277–1284. [Google Scholar] [CrossRef]
- Gottshall, J.L.; Guedes, V.A.; Pucci, J.U.; Brooks, D.; Watson, N.; Sheth, P.; et al. Poor Sleep Quality is Linked to Elevated Extracellular Vesicle-Associated Inflammatory Cytokines in Warfighters With Chronic Mild Traumatic Brain Injuries. Front Pharmacol. 2021, 12, 762077. [Google Scholar] [CrossRef] [PubMed]
- Guedes, V.A.; Kenney, K.; Shahim, P.; Qu, B.X.; Lai, C.; Devoto, C.; et al. Exosomal neurofilament light: A prognostic biomarker for remote symptoms after mild traumatic brain injury? Neurology 2020, 94, e2412–e2423. [Google Scholar] [CrossRef] [PubMed]
- Flynn, S.; Leete, J.; Shahim, P.; Pattinson, C.; Guedes, V.A.; Lai, C.; et al. Extracellular vesicle concentrations of glial fibrillary acidic protein and neurofilament light measured 1 year after traumatic brain injury. Sci Rep. 2021, 11, 3896. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.; Hemphill, M.; Yang, Z.; Beard, K.; Sewell, E.; Shallcross, J.; et al. Multi-Dimensional Mapping of Brain-Derived Extracellular Vesicle MicroRNA Biomarker for Traumatic Brain Injury Diagnostics. J Neurotrauma. 2020, 37, 2424–2434. [Google Scholar] [CrossRef] [PubMed]
- Ghai, V.; Fallen, S.; Baxter, D.; Scherler, K.; Kim, T.K.; Zhou, Y.; et al. Alterations in Plasma microRNA and Protein Levels in War Veterans with Chronic Mild Traumatic Brain Injury. J Neurotrauma. 2020, 37, 1418–1430. [Google Scholar] [CrossRef] [PubMed]
- Devoto, C.; Lai, C.; Qu, B.X.; Guedes, V.A.; Leete, J.; Wilde, E.; et al. Exosomal MicroRNAs in Military Personnel with Mild Traumatic Brain Injury: Preliminary Results from the Chronic Effects of Neurotrauma Consortium Biomarker Discovery Project. J Neurotrauma. 2020, 37, 2482–2492. [Google Scholar] [CrossRef] [PubMed]
- Puffer, R.C.; Cumba Garcia, L.M.; Himes, B.T.; Jung, M.Y.; Meyer, F.B.; Okonkwo, D.O.; et al. Plasma extracellular vesicles as a source of biomarkers in traumatic brain injury. J Neurosurg. 2020, 134, 1921–1928. [Google Scholar] [CrossRef] [PubMed]
- Shaimardanova, A.A.; Solovyeva, V.V.; Chulpanova, D.S.; James, V.; Kitaeva, K.V.; Rizvanov, A.A. Extracellular vesicles in the diagnosis and treatment of central nervous system diseases. Neural Regen Res. 2020, 15, 586–596. [Google Scholar] [PubMed]
- Gratpain, V.; Mwema, A.; Labrak, Y.; Muccioli, G.G.; van Pesch, V.; des Rieux, A. Extracellular vesicles for the treatment of central nervous system diseases. Adv Drug Deliv Rev. 2021, 174, 535–552. [Google Scholar] [CrossRef]
- Mager, I.; Breakefield, X.O.; Wood, M.J.; SELA. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discov. 2013, 12, 347–357. [Google Scholar]
- Wiklander, O.P.B.; Brennan, M.A.; Lotvall, J.; Breakefield, X.O.; El Andaloussi, S. Advances in therapeutic applications of extracellular vesicles. Sci Transl Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- An, K.; Klyubin, I.; Kim, Y.; Jung, J.H.; Mably, A.J.; O'Dowd, S.T.; et al. Exosomes neutralize synaptic-plasticity-disrupting activity of Abeta assemblies in vivo. Mol Brain. 2013, 6, 47. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Sun, H.; Sakai, S.; Mitsutake, S.; Okada, M.; Tahara, H.; et al. Decreased amyloid-beta pathologies by intracerebral loading of glycosphingolipid-enriched exosomes in Alzheimer model mice. J Biol Chem. 2014, 289, 24488–24498. [Google Scholar] [CrossRef] [PubMed]
- Katsuda, T.; Tsuchiya, R.; Kosaka, N.; Yoshioka, Y.; Takagaki, K.; Oki, K.; et al. Human adipose tissue-derived mesenchymal stem cells secrete functional neprilysin-bound exosomes. Sci Rep. 2013, 3, 1197. [Google Scholar] [CrossRef] [PubMed]
- Attaluri, S.; Jaimes Gonzalez, J.; Kirmani, M.; Vogel, A.D.; Upadhya, R.; Kodali, M.; et al. Intranasally administered extracellular vesicles from human induced pluripotent stem cell-derived neural stem cells quickly incorporate into neurons and microglia in 5xFAD mice. Front Aging Neurosci. 2023, 15, 1200445. [Google Scholar] [CrossRef] [PubMed]
- Ananbeh, H.; Vodicka, P.; Kupcova Skalnikova, H. Emerging Roles of Exosomes in Huntington's Disease. Int J Mol Sci. 2021, 22. [Google Scholar] [CrossRef]
- Didiot, M.C.; Hall, L.M.; Coles, A.H.; Haraszti, R.A.; Godinho, B.M.; Chase, K.; et al. Exosome-mediated Delivery of Hydrophobically Modified siRNA for Huntingtin mRNA Silencing. Mol Ther. 2016, 24, 1836–1847. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Im, W.; Mook-Jung, I.; Kim, M. MicroRNA-124 slows down the progression of Huntington's disease by promoting neurogenesis in the striatum. Neural Regen Res. 2015, 10, 786–791. [Google Scholar] [PubMed]
- Cooper, J.M.; Wiklander, P.B.; Nordin, J.Z.; Al-Shawi, R.; Wood, M.J.; Vithlani, M.; et al. Systemic exosomal siRNA delivery reduced alpha-synuclein aggregates in brains of transgenic mice. Mov Disord. 2014, 29, 1476–1485. [Google Scholar] [CrossRef]
- Lee, S.T.; Im, W.; Ban, J.J.; Lee, M.; Jung, K.H.; Lee, S.K.; et al. Exosome-Based Delivery of miR-124 in a Huntington's Disease Model. J Mov Disord. 2017, 10, 45–52. [Google Scholar] [CrossRef]
- Upadhya, R.; Shetty, A.K. Extracellular Vesicles for the Diagnosis and Treatment of Parkinson's Disease. Aging Dis. 2021, 12, 1438–1450. [Google Scholar] [CrossRef]
- Kojima, R.; Bojar, D.; Rizzi, G.; Hamri, G.C.; El-Baba, M.D.; Saxena, P.; et al. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson's disease treatment. Nat Commun. 2018, 9, 1305. [Google Scholar] [CrossRef]
- Qu, M.; Lin, Q.; Huang, L.; Fu, Y.; Wang, L.; He, S.; et al. Dopamine-loaded blood exosomes targeted to brain for better treatment of Parkinson's disease. J Control Release. 2018, 287, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Pusic, A.D.; Pusic, K.M.; Clayton, B.L.; Kraig, R.P. IFNgamma-stimulated dendritic cell exosomes as a potential therapeutic for remyelination. J Neuroimmunol. 2014, 266, 12–23. [Google Scholar] [CrossRef]
- Leggio, L.; Paterno, G.; Vivarelli, S.; L'Episcopo, F.; Tirolo, C.; Raciti, G.; et al. Extracellular Vesicles as Nanotherapeutics for Parkinson's Disease. Biomolecules. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Upadhya, D.; Hattiangady, B.; Kim, D.K.; An, S.Y.; Shuai, B.; et al. Intranasal MSC-derived A1-exosomes ease inflammation, and prevent abnormal neurogenesis and memory dysfunction after status epilepticus. Proc Natl Acad Sci U S A. 2017, 114, E3536–E3545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chopp, M.; Meng, Y.; Katakowski, M.; Xin, H.; Mahmood, A.; et al. Effect of exosomes derived from multipluripotent mesenchymal stromal cells on functional recovery and neurovascular plasticity in rats after traumatic brain injury. J Neurosurg. 2015, 122, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Drommelschmidt, K.; Serdar, M.; Bendix, I.; Herz, J.; Bertling, F.; Prager, S.; et al. Mesenchymal stem cell-derived extracellular vesicles ameliorate inflammation-induced preterm brain injury. Brain Behav Immun. 2017, 60, 220–232. [Google Scholar] [CrossRef]
- Kodali, M.; Madhu, L.N.; Reger, R.L.; Milutinovic, B.; Upadhya, R.; Gonzalez, J.J.; et al. Intranasally administered human MSC-derived extracellular vesicles inhibit NLRP3-p38/MAPK signaling after TBI and prevent chronic brain dysfunction. Brain Behav Immun. 2023, 108, 118–134. [Google Scholar] [CrossRef]
- Kodali, M.; Madhu, L.N.; Reger, R.L.; Milutinovic, B.; Upadhya, R.; Attaluri, S.; et al. A single intranasal dose of human mesenchymal stem cell-derived extracellular vesicles after traumatic brain injury eases neurogenesis decline, synapse loss, and BDNF-ERK-CREB signaling. Front Mol Neurosci. 2023, 16, 1185883. [Google Scholar] [CrossRef]
- Patel, N.A.; Moss, L.D.; Lee, J.Y.; Tajiri, N.; Acosta, S.; Hudson, C.; et al. Long noncoding RNA MALAT1 in exosomes drives regenerative function and modulates inflammation-linked networks following traumatic brain injury. J Neuroinflammation. 2018, 15, 204. [Google Scholar] [CrossRef]
- Landrum, M.J.; Lee, J.M.; Benson, M.; et al. ClinVar: public archive of interpretations of clinically relevant variants. Nucleic Acids Res. 2016, 44, D862–D868. [Google Scholar] [CrossRef] [PubMed]
- Villiger, L.; Joung, J.; Koblan, L.; Weissman, J.; Abudayyeh, O.O.; Gootenberg, J.S. CRISPR technologies for genome, epigenome and transcriptome editing. Nat Rev Mol Cell Biol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.J.; Liu, D.R. Prime editing for precise and highly versatile genome manipulation. Nat Rev Genet. 2023, 24, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.C.; Zhang, T.; Gao, J.Q. The in vivo fate and targeting engineering of crossover vesicle-based gene delivery system. Adv Drug Deliv Rev. 2022, 187, 114324. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Liu, C.; Wang, N.; et al. Delivery of CRISPR/Cas systems for cancer gene therapy and immunotherapy. Adv Drug Deliv Rev. 2021, 168, 158–180. [Google Scholar] [CrossRef] [PubMed]
- Kostyushev, D.; Kostyusheva, A.; Brezgin, S.; et al. Gene Editing by Extracellular Vesicles. Int J Mol Sci. 2020, 21, 7362. [Google Scholar] [CrossRef] [PubMed]
- Taha, E.A.; Lee, J.; Hotta, A. Delivery of CRISPR-Cas tools for in vivo genome editing therapy: Trends and challenges. J Control Release. 2022, 342, 345–361. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hu, S.; Chen, X. Non-viral delivery systems for CRISPR/Cas9-based genome editing: Challenges and opportunities. Biomaterials. 2018, 171, 207–218. [Google Scholar] [CrossRef]
- Madigan, V.; Zhang, F.; Dahlman, J.E. Drug delivery systems for CRISPR-based genome editors. Nat Rev Drug Discov. 2023, 22, 875–894. [Google Scholar] [CrossRef]
- Kim, S.M.; Yang, Y.; Oh, S.J.; Hong, Y.; Seo, M.; Jang, M. Cancer-derived exosomes as a delivery platform of CRISPR/Cas9 confer cancer cell tropism-dependent targeting. J Control Release. 2017, 266, 8–16. [Google Scholar] [CrossRef] [PubMed]
- McAndrews, K.M.; Xiao, F.; Chronopoulos, A.; LeBleu, V.S.; Kugeratski, F.G.; Kalluri, R. Exosome-mediated delivery of CRISPR/Cas9 for targeting of oncogenic KrasG12D in pancreatic cancer. Life Sci Alliance. 2021, 4, e202000875. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Shi, Q.; Yang, T.; et al. In Vivo Visualized Tracking of Tumor-Derived Extracellular Vesicles Using CRISPR-Cas9 System. Technol Cancer Res Treat. 2022, 21, 15330338221085370. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Foot, N.; Ang, C.S.; et al. Arrestin-Domain Containing Protein 1 (Arrdc1) Regulates the Protein Cargo and Release of Extracellular Vesicles. Proteomics. 2018, 18, e1800266. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.; Freed, E.O. ARRDC1 as a mediator of microvesicle budding. Proc Natl Acad Sci U S A. 2012, 109, 4025–4026. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yu, J.; Kadungure, T.; Beyene, J.; Zhang, H.; Lu, Q. ARMMs as a versatile platform for intracellular delivery of macromolecules. Nat Commun. 2018, 9, 960. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Yang, X.; Li, Q.; et al. Myristoylation of Src kinase mediates Src-induced and high-fat diet-accelerated prostate tumor progression in mice. J Biol Chem. 2017, 292, 18422–18433. [Google Scholar] [CrossRef] [PubMed]
- Dooley, K.; McConnell, R.E.; Xu, K.; et al. A versatile platform for generating engineered extracellular vesicles with defined therapeutic properties. Mol Ther. 2021, 29, 1729–1743. [Google Scholar] [CrossRef] [PubMed]
- Whitley, J.A.; Kim, S.; Lou, L.; et al. Encapsulating Cas9 into extracellular vesicles by protein myristoylation. J Extracell Vesicles. 2022, 11, e12196. [Google Scholar] [CrossRef]
- Duan, D.; Goemans, N.; Takeda, S.; Mercuri, E.; Aartsma-Rus, A. Duchenne muscular dystrophy. Nat Rev Dis Primers. 2021, 7, 13. [Google Scholar] [CrossRef]
- Majeau, N.; Fortin-Archambault, A.; Gérard, C.; Rousseau, J.; Yaméogo, P.; Tremblay, J.P. Serum extracellular vesicles for delivery of CRISPR-CAS9 ribonucleoproteins to modify the dystrophin gene. Mol Ther. 2022, 30, 2429–2442. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Lyu, P.; Yoo, K.; et al. Engineered extracellular vesicles as versatile ribonucleoprotein delivery vehicles for efficient and safe CRISPR genome editing. J Extracell Vesicles. 2021, 10, e12076. [Google Scholar] [CrossRef] [PubMed]
- Gee, P.; Lung, M.S.Y.; Okuzaki, Y.; et al. Extracellular nanovesicles for packaging of CRISPR-Cas9 protein and sgRNA to induce therapeutic exon skipping. Nat Commun. 2020, 11, 1334. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Both sides of EVs application: Therapeutics and Biomarkers. Isolated stem cells from different sources may release extracellular vesicles (EVs) with anti-inflammatory and immunomodulatory characteristics, which are significant candidates for treatment of several neurological diseases. After EVs isolation, purification, and characterization by specific methods, they can be prepared for administration by intranasal (I.N.) or intravenous (I.V.) injection and the improvement of different cerebral functions can be measured. Also, EVs have the capacity of crossing the blood-brain barrier (BBB), which is a fundamental characteristic for therapeutics of central nervous system (CNS) diseases. On the other side, EVs are promising biomarkers for several neurodegenerative and neurological pathologies since they are secreted by cells from CNS and can be detected in biological fluids such as saliva, blood, and urine. After EVs isolation, purification and characterization and analysis of their cargo, such as DNA, RNA, proteins or/and lipids can be identified as biomarkers and used for different clinical applications.
Figure 1.
Both sides of EVs application: Therapeutics and Biomarkers. Isolated stem cells from different sources may release extracellular vesicles (EVs) with anti-inflammatory and immunomodulatory characteristics, which are significant candidates for treatment of several neurological diseases. After EVs isolation, purification, and characterization by specific methods, they can be prepared for administration by intranasal (I.N.) or intravenous (I.V.) injection and the improvement of different cerebral functions can be measured. Also, EVs have the capacity of crossing the blood-brain barrier (BBB), which is a fundamental characteristic for therapeutics of central nervous system (CNS) diseases. On the other side, EVs are promising biomarkers for several neurodegenerative and neurological pathologies since they are secreted by cells from CNS and can be detected in biological fluids such as saliva, blood, and urine. After EVs isolation, purification and characterization and analysis of their cargo, such as DNA, RNA, proteins or/and lipids can be identified as biomarkers and used for different clinical applications.
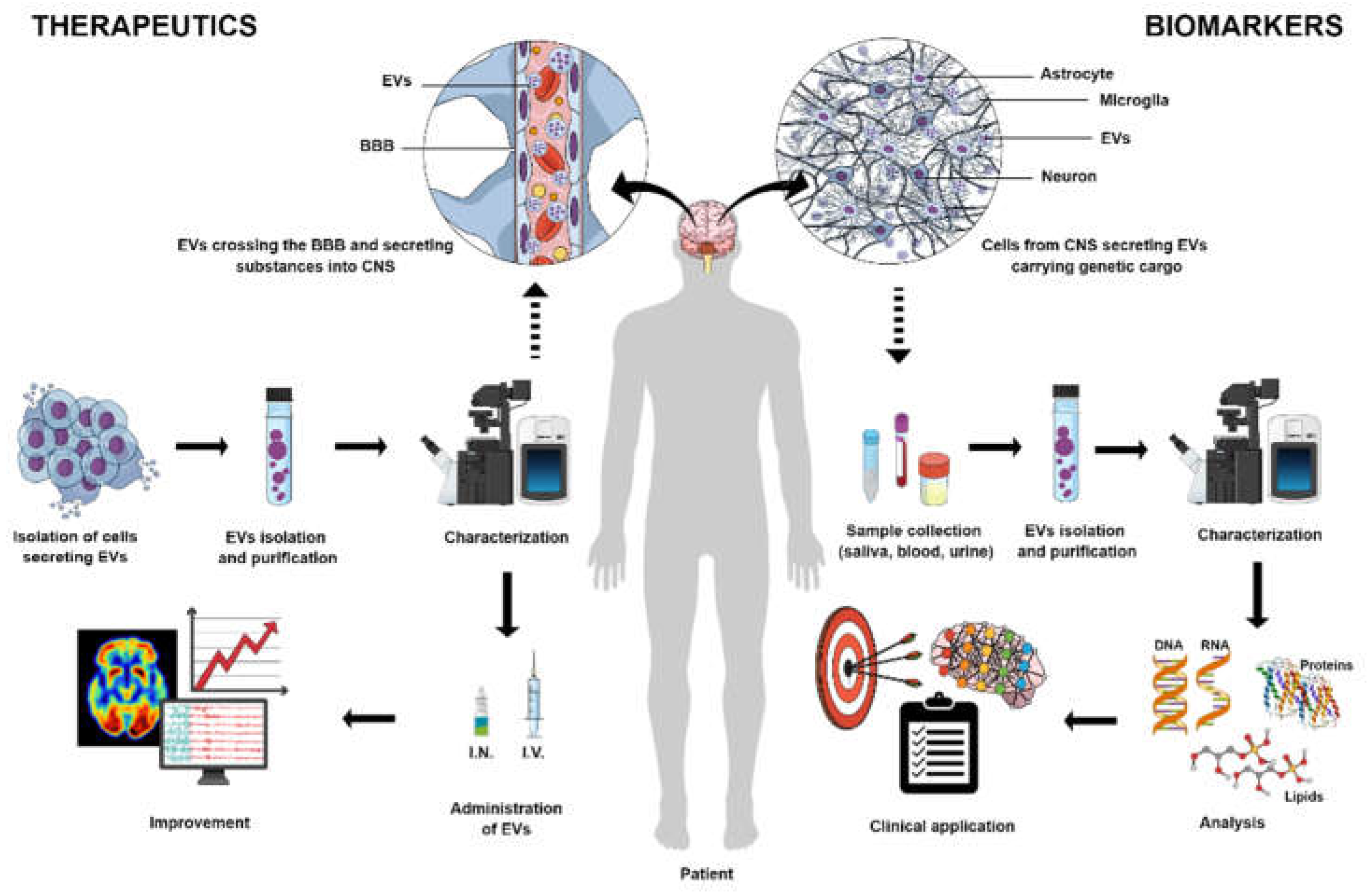

Table 1.
Main findings of extracellular vesicle biomarkers in non-invasive samples of neurological diseases.
Table 1.
Main findings of extracellular vesicle biomarkers in non-invasive samples of neurological diseases.
| Disease | Author | Year | Sample type | EVs type reported | Main findings - Biomarkers | ||
|---|---|---|---|---|---|---|---|
| Alzheimer’s disease | Kim, K. Y.; et al | 2021 | Plasma/Serum | Brain-derived exosomes | Protein | ↑ | Aβ-42 |
| t-tau | |||||||
| p-T181-tau | |||||||
| p-S396-tau | |||||||
| Liu, W.; et al | 2020 | Plasma | Neuronally derived exosomes | Protein | ↓ | Ng | |
| Jia, L.; et al | 2021 | Plasma | Neuronal-derived exosomes | Protein | ↓ | Ng | |
| GAP43 | |||||||
| SNAP25 | |||||||
| SYT1 | |||||||
| Li, F.; et al | 2020 | Serum | Extracellular vesicles | Protein | ↑ | Aβ1-42 | |
| total-tau | |||||||
| P-T181-tau | |||||||
| P-S396-tau | |||||||
| miRNA | ↑ | hsa- miR-424-5p | |||||
| hsa-miR-3065-5p | |||||||
| hsa-miR-93-5p | |||||||
| ↓ | hsa-miR-1306-5p | ||||||
| Durur, D. Y.; et al | 2022 | Plasma | Small neuron-derived extracellular vesicles (sNDEVs) | miRNA | ↑ | Let-7e | |
| Yang, T. T.; et al | 2018 | Serum | Exosomes | miRNA | ↑ | miR-135a | |
| miR-384 | |||||||
| ↓ | miR-193b | ||||||
| Aharon, A.; et al | 2020 | Plasma (platelet-poor plasma) | Extracellular vesicles | miRNA | ↓ | has-let-7g-5p | |
| has-miR126-3p | |||||||
| has-miR142-3p | |||||||
| has-miR146a-5p | |||||||
| has-mir26b-5p | |||||||
| has-mir223-3p | |||||||
| Cell marker | ↑ | CD171/L1CAM - axonal | |||||
| MOG - glial | |||||||
| CD31+ CD41-%, %CD144 - endothelial | |||||||
| Cytokine | ↑ | IFN-γ | |||||
| RANTES | |||||||
| GRO | |||||||
| IL-2 | |||||||
| IL-8 | |||||||
| AgRP | |||||||
| Growth Factor Content | ↑ | PDGF-BB | |||||
| TPO | |||||||
| ↓ | UPAR | ||||||
| VEGF-D | |||||||
| Receptors VEGFR-2 and 3 | |||||||
| FGF-4 | |||||||
| EGF | |||||||
| ANG-1 | |||||||
| Parkinson’s Disease | Shi, M.; et al | 2014 | Plasma | Exosomes | Protein | ↑ | α-syn |
| Stuendl, A.; et al | 2021 | Plasma | Extracellular vesicles | Protein | ↑ | α-syn | |
| Niu, M.; et al | 2020 | Plasma | Neuronal-derived exosomes | Protein | ↑ | α-syn | |
| Jiang, G.; et al | 2020 | Serum | Neuron-derived exosomes | Protein | ↑ | α-syn | |
| Chung, C-C.; et al | 2021 | Plasma | Extracellular vesicles | Protein | ↓ | α-syn | |
| Si, X.; et al | 2019 | Serum | CNS-derived exosomes | Protein | ↓ | α-syn | |
| Cao, Z.; et al | 2019 | Saliva | Exosomes | Protein | ↑ | α-synOlig | |
| α-synOlig/α-syn total ratio | |||||||
| Ho, D. H.; et al | 2014 | Urine | Exosomes | Protein | ↑ | Protein deglycase DJ-1 | |
| Zhao, Z-H.; et al | 2019 | Plasma | Neural-derived exosomes | Protein | ↑ | Protein deglycase DJ-1 | |
| Exosomes/total Protein deglycase DJ-1 ratio | |||||||
| α-syn | |||||||
| Fraser, K. B.; et al | 2016 | Urine | Exosomes | Protein | ↑ | Ser(P)-1292 LRRK2 | |
| Yao, Y-F.; et al | 2018 | Plasma | Exosomes | miRNA | ↑ | miR-331-5p | |
| ↓ | miR-505 | ||||||
| Xie, S.; et al | 2020 | Plasma | Extracellular vesicles | miRNA | ↑ | hsa-miR-30c-2-3p | |
| ↓ | hsa-miR-15b-5p | ||||||
| hsa-miR-138-5p | |||||||
| hsa-miR-338-3p | |||||||
| hsa-miR-106b-3p | |||||||
| hsa-miR-431-5p | |||||||
| Cao, X-Y.; et al | 2017 | Serum | Exosomes | miRNA | ↑ | miR-195 | |
| miR-24 | |||||||
| ↓ | miR-19b | ||||||
| He, S.; et al | 2021 | Serum | Extracellular vesicles | miRNA | ≠ | hsa-miR-374a-5p | |
| hsa-miR-374b-5p | |||||||
| hsa-miR-199a-3p | |||||||
| hsa-miR-28-5p | |||||||
| hsa-miR-22-5p | |||||||
| hsa-miR-151a-5p | |||||||
| Grossi, I.; et al | 2021 | Plasma | Pure small extracellular vesicles | miRNA | ↑ | miR-34a-5p | |
| Chan, L.; et al | 2021 | Plasma | Extracellular vesicles | Cytokine | ↑ | pro-IL-1β (pro-interleukin) | |
| TNF-α (tumor necrosis factor) | |||||||
| Multiple sclerosis | Scaroni, F.; et al | 2022 | Plasma | Myeloid Evs | miRNA | ↑ | miR-150-5p |
| ↓ | let-7b-5p | ||||||
| Azimi, M.; et al | 2019 | Whole blood | T Cell-derived Exosomes | miRNA | ↑ | miR-326 | |
| Selmaj, I.; et al | 2017 | Serum | Exosomes | miRNA | ↓ | hsa-miR-122-5p | |
| hsa-miR-196b-5p | |||||||
| hsa-miR-301a-3p | |||||||
| hsa-miR-532-5p | |||||||
| Groen, K.; et al | 2020 | Whole blood | Erythrocyte-derived EVs | miRNA | ↑ | hsa-miR-148-5p | |
| Ebrahimkhani, S.; et al | 2017 | Serum | Exosomes | miRNA | ↑ | miR-15b-5p | |
| miR-451a | |||||||
| miR-30b-5p | |||||||
| miR-342-3p | |||||||
| miR-127-3p | |||||||
| miR-370-3p | |||||||
| miR-409-3p | |||||||
| miR-432-5p | |||||||
| Moyano, A.; et al | 2016 | Plasma | Small Evs | Glycolipid | ↑ | C16:0 sulfatide | |
| Galazka, G.; et al | 2018 | Serum | Exosomes | Protein | ↑ | MOG | |
| Bhargava, P.; et al | 2020 | Plasma | Neuronal-enriched EVs | Protein | ↓ | Synaptopodin | |
| Synaptophysin | |||||||
| Astrocytic-enriched EVs | Complement cascade component | ↑ | C1q | ||||
| C3 | |||||||
| C3b/iC3b | |||||||
| C5 | |||||||
| C5a | |||||||
| Factor H | |||||||
| Blandford, S.; et al | 2022 | Plasma | Extracellular vesicles | EV concentration (cell-specific) | ↑ | CD3+ (T cells) | |
| CD4+ T cells and monocytes | |||||||
| CD8+ T cells | |||||||
| CD14+ (Monocytes) | |||||||
| CD19+ (B cells) | |||||||
| Mazzucco, M.; et al | 2022 | Plasma | CNS endothelial derived Evs | EV concentration | ↑ | CD3/CD4- pETX/MAL+ CD31+ | |
| CD3/CD4- pETX/MAL+ CD105+ | |||||||
| CD3/CD4- pETX/MAL+ CD144+ | |||||||
| Epilepsy | Yan, S.; et al | 2017 | Plasma | Exosomes | miRNA | ↓ | miR-4668-5p |
| miR-4322 | |||||||
| miR-8071 | |||||||
| miR-6781-5p | |||||||
| miR-197-5p | |||||||
| ↑ | miR3613-5p | ||||||
| Wang, Y.; et al | 2022 | Plasma | Small EVs | miRNA | ↑ | hsa-miR-584a-5p | |
| hsa-miR-342a-5p | |||||||
| hsa-miR-150-3p | |||||||
| hsa-miR-125b-5p | |||||||
| Chen, S-D.; et al | 2020 | Serum | Exosomes | miRNA | ↑ | miR-194-2 | |
| miR-15a | |||||||
| miR-132 | |||||||
| Lin, Z.; et al | 2020 | Serum | Exosomes | Protein | ↑ | F9 | |
| ↓ | TSP-1 | ||||||
| Traumatic brain injury | Kryshawna, B.; et al | 2021 | Plasma | Extracellular vesicles | Protein | ↑ | GFAP |
| IL6 | |||||||
| Flynn, S.; et al | 2021 | Serum | Extracellular vesicles | Protein | ↑ | GFAP | |
| Goetzl, E. J.; et al | 2019 | Plasma | Neuron-derived exosomes | Protein | ↓ | CD81 (Acute) | |
| RAB10 (Acute) | |||||||
| ↑ | Annexin VII (Acute) | ||||||
| UCH-L1 (Acute) | |||||||
| All spectrin fragments (Acute) | |||||||
| Claudin-5 (Acute) | |||||||
| Occludion (Acute) | |||||||
| NKCC-1 (Acute) | |||||||
| Aquaporin 4 (Acute/Chronic) | |||||||
| Synaptogyrin-3 (Acute/Chronic) | |||||||
| Aβ42 (Acute/Chronic) | |||||||
| P-T181-tau (Acute/Chronic) | |||||||
| P-S396 (Chronic) | |||||||
| total tau (Acute/Chronic) | |||||||
| PRPc (Acute/Chronic) | |||||||
| Cytokine | ↑ | IL-6 (Acute/Chronic) | |||||
| Gill, J.; et al | 2018 | Plasma | Neuronal-derived exosomes | Protein | ↑ | tau | |
| Aβ42 | |||||||
| Cytokine | ↑ | IL-10 | |||||
| Gottshall, J.; et al | 2022 | Plasma | Extracellular vesicles | Cytokine | ↑ | IL-6 | |
| Cheng, Y.; et al | 2019 | Saliva | Extracellular vesicles | Gene | ↑ | CDC2 | |
| CSNK1A1 | |||||||
| CTSD | |||||||
| Ko, J.; et al | 2020 | Plasma | Brain-derived Evs | miRNA | ≠ | miR-203b-5p | |
| miR-203a-3p | |||||||
| miR-206 | |||||||
| miR-185-5p | |||||||
| Ghai, V.; et al | 2020 | Plasma | Extracellular vesicles | miRNA | ↓ | miR-139-5p | |
| miR-143-3p | |||||||
| miR-146a-5p | |||||||
| miR-192-5p | |||||||
| miR-203a-3p | |||||||
| miR-21-5p | |||||||
| miR-423-5p | |||||||
| miR-483-5p | |||||||
Abbreviations: AgRP (agouti related peptide); ANG-1 (angiopoietin 1); Aβ-42 (amyloid-β42); CNS (central nervous system); EGF (epidermal growth factor); EVs (extracellular vesicles); FGF-4 (fibroblast growth factor); F9 (coagulation factor IX); GAP43 (growth associated protein 43); GFAP (glial fibrillary acidic protein); GRO (growth-regulated oncogene-alpha, CXCL1); IFN-γ (IFN-gamma); IL-2 (interleukin-2); IL-8 (interleukin-8); L1CAM (L1 cell adhesion molecule); miRNA (microRNA); MOG (myelin oligodendrocyte glycoprotein); Ng (neurogranin); PDGF-BB (platelet-derived growth factor BB); p-S396-tau (tau phosphorylated at serine 396); p-T181-tau (tau phosphorylated at threonine 181); RANTES (chemokine (C-C motif) ligand 5, CCL5); Ser(P)-1292 LRRK2 (leucine-rich repeat kinase 2); SNAP25 (synaptosome associated protein 25); SYT1 (synaptotagmin 1); TPO (thrombopoietin); TSP-1 (thrombospondin-1); t-tau (total-tau); UPAR (growth factor urokinase receptor); VEGF-D (vascular endothelial growth factor); α-syn (alpha-synuclein).
Table 2.
Extracellular vesicles as therapeutic options.
| Disease | Author | Year | Material | Model | Via | Main Findings |
|---|---|---|---|---|---|---|
| Alzheimer’s disease | Katsuda, T.; et al | 2013 | MSC-EVs derived from human adipose tissue containing neprilysin | in vitro | NA | Suggests the lyse of beta amyloid peptide |
| Yuyama, K.; et al | 2014 | Neuroblastoma-derived exosomes | Heterozygotic transgenic mice that express the human APP | Stereotaxic injection | Increased clearance of beta amyloid peptite by trapping it inside the EVs and deliviering it to microglica, where it can be degraded | |
| Parkinson’s Disease | Jarmalavicuite, A.; et al | 2015 | EVs from stem cells from human exfoliated deciduous teeth | in vitro | NA | Neuroprotective potential and preventention of apoptosis in 80% of dopaminergic neurons when maintained in 3D culture |
| Narbute, K.; et al | 2019 | EVs from stem cells from human exfoliated deciduous teeth | Rat model | IN | Improvement of the rat gait parameters and increased tyrosine hydroxylase expression in substantia nigra | |
| Qu, M.; et al | 2018 | Blood EVs loaded with dopamine | Murine model | IV | Improvement of functional recovery without toxicity in hippocampus, liver, spleen or lungs | |
| Cooper, J. M.; et al | 2014 | Exosomes expressing RVG loaded with α-Syn siRNA | Transgenic mice expressing the human phosphorylation-mimic α-Syn | IV | Down-regulation of alpha synuclein | |
| Kojima, R.; et al | 2018 | HEK-293 T cells with a modified plasmid coding for catalase | Mice | IN | Reduction of brain inflammation, when compared to free Catalase administration | |
| Huntington’s Disease | Didiot, M-C.; et al | 2016 | Hydrophobic siRNA targeting Huntintinton mRNA in Glioblastoma EVs | Mice | Stereotactic injection in mouse striatum | Decrease of protein level in vitro and in vivo with no clinical improvement |
| Liu, T.; et al | 2015 | EVs loaded with miR-124 | R6/2 HD transgenic mice | Stereotactic injection in mouse striatum | Slowed down progression of HD, promotesd neuronal differentiation and survival | |
| Lee, S-T.; et al | 2017 | miR-124 expressing HEK293 cell line | R6/2 transgenic HD mice | Stereotactic injection in mouse striatum | Reduction of REST protein expression, no behavior improvement | |
| Wu, T.; et al | 2018 | Exosomes expressing the neuron-specific RVG peptide, loaded with siRNA targeting human huntingtin exon 1 | BACHD and N171-82Q transgenic mice | IV | Reduced HTT expression up to 46% and 54% | |
| Multiple sclerosis | Laso-Garcia, F.; et al | 2018 | MSC-EVs from human adipose tissue | Theiler's murine encephalomyelitis virus induced demyelinating disease | IV | Improvement in motor deficits, reducion of brain atrophy, increasion in cell proliferation in the subventricular zone and decreasion of inflammatory infiltrates in the spinal cord |
| Pusic, A. D.; et al | 2013 | IFNy-stimulated dendritic cells | Ex vivo (mature hippocampal slice cultures) / In vivo | NA / IN | Increase of myelin levels | |
| Riazifar, M.; et al | 2019 | human MSCs EVs | Experimental autoimmune encephalomyelitis mouse model | IV | Stimulation by IFNγ reduced the mean clinical score of EAE mice compared control, reduced demyelination, decreased neuroinflammation, and upregulated the number of CD4+CD25+FOXP3+ regulatory T cells (Tregs) within the spinal cords of EAE mice. Co-culture of IFNγ-Exo with activated peripheral blood mononuclear cells cells in vitro reduced levels of pro-inflammatory Th1 and Th17 cytokines | |
| Epilepsy | Long, Q.; et al | 2017 | MSC-EVs from human bone marrow | Mouse model of induced status epilepticus | IN | Reduction of proinflammatory cytokines, better cognitive and memory functions. Neuroprotection, reducing neural loss, preserving GABAergic intraneurons |
| Traumatic brain injury | Doeppner, T. R.; et al | 2015 | Human bone marrow-derived MSC | Mice after focal cerebral ischemia | IV | Angiogenesis, improvement of neurological impairment, long-term neuroprotection |
| Zhang, Y.; et al | 2015 | Murine bone marrow derived MSC-EVs | Traumatic brain injured rats and mice | IV | Improvement of spatial learning, reduction of neurological deficits, better angiogenesis and neurogenesis, reduction of inflammation | |
| Kim, D-K.; et al | 2016 | Human MSCs EVs | Traumatic brain injuried mice | IV | Improvement of spatial learning and pattern separation ability, decrease of neuroinflammation | |
| Drommelschmidt. K.; et al | 2017 | Human bone marrow-derived MSC | Rodent model of inflammation-induced brain injury | IP | Improvement of long-lasting cognitive functions, amelioration of inflammation, restoration of short-term myelination deficits | |
| Gao, W.; et al | 2018 | ECFCs EVs | Traumatic Brain Injured mice | Stereotactic injection after trauma | Inhibition of PTEN expression and increasion of AKT expression, changes accompanied by reductions in Evans blue dye extravasation, brain edema and tight junction degradation | |
| Patel, N. A.; et al | 2018 | Adipose-derived stem cell EVs containing MALAT1 | Mild controlled cortical impact -induced traumatic brain injury in rat | IV | Recovery of function on motor behavior and reduction in cortical brain injury |
Abbreviations: APP (amyloid-β precursor protein); AKT (Protein kinase B); BACHD (transgenic mice expressing a neuropathogenic, full-length human mutant Huntingtin gene); EAE (experimental autoimmune encephalomyelitis); ECFCs (endothelial colony-forming cells); EVs (extracellular vesicles); HD (Huntington disease); HEK293 (immortalized human embryonic kidney cells); HTT gene that provides instructions for making a protein called huntingtin; IFNy (interferon gama); IN (intranasal injection); IP (intraperitoneally injection); IV: intravenous injection; MALAT1 (metastasis-associated lung adenocarcinoma transcript 1); miR-124 (most abundant microRNA in the adult brain); mRNA (messenger RNA); MSCs (mesenchymal stem cells); NA (not applicable); N171-82Q (transgenic line expressing an N-terminally truncated human huntingtin cDNA); PTEN (protein that inhibits cell growth and promotes cellular apoptosis); REST protein (RE1-Silencing Transcription Factor); RVG (rabies viral glycoprotein); R6/2 HD (mice model expressing a portion of human Huntington’s disease gene under human gene promoter elements); siRNA (small interfering RNA); α-Syn (α-Synuclein).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



