
Submitted:
25 April 2024
Posted:
26 April 2024
You are already at the latest version
Abstract
The colpodellids consist of free-living predatory protists phylogenetically related to pathogenic Apicomplexan parasites such as Plasmodium species, Cryptosporidium sp. and Toxoplasma gondii. Among the colpodellids, Colpodella species prey on bodonids, algae, and ciliates. Voromonas pontica preys on Percolomonas species. In the past decade, Colpodella species have been reported to cause opportunistic infections in humans and animals, and having the potential for zoonotic infections in humans due to the presence of Colpodella species in ticks. In the reported cases, transmission and pathogenic stages of Col-podella were not identified by light microscopy. In the colpodellid V. pontica, cyst stages of both the pred-ator and its prey Percolomonas cosmopolitus have not been identified by light microscopy. In this study we evaluated the dye components used for Sam-Yellowe’s trichrome staining, from different major vendors to determine the consistency and reproducibility of the staining protocol to aid the use of the staining proto-col for diagnosis. We show that although slight variations in the color of the stained cells are obtained, life cycle stages of Colpodella ATCC 50594 could be clearly identified, regardless of the vendor used. For the first time, Sam-Yellowe’s trichrome staining series, could identify cysts of V. pontica and its prey, demonstrating that the staining protocol can identify life cycle stages of other colpodellids. Sam-Yellowe's trichrome staining will aid molecular diagnosis of infections caused by colpodellids in human and ani-mals, identify colpodellids in ticks and flies and from environmental samples, when used for staining.
Keywords:
Apicomplexa
; Colpodella species
; colpodellids
; Colpodella sp. 50594
; life cycle
; myzocytosis
; Sam-Yellowe’s trichrome stains
; Voromonas cysts
; Voromonas pontica
1. Introduction
Colpodellids are free-living protists related phylogenetically to the pathogenic apicomplexans such as Toxoplasma gondii, Plasmodium sp., Babesia sp., and Cryptosporidium sp. [1]. The colpodellids which include Colpodella species, Voromonas pontica (previously Colpodella pontica) and Alphamonas are biflagellated and possess trophozoite and cyst stages in their life cycles [2]. Colpodellids prey on bodonids, ciliates and algae using the process of myzocytosis where cytoplasmic contents of the prey are aspirated into the cytoplasm of the predator. Ectoparasitic colpodellids such as Colpodella gonderi and C. tetrahymenae attach to ciliates for prolonged feeding in contrast to the predatory attacks observed with bodonid or algal prey [3,4]. In cyst forming species, the food vacuole associated with remnants of the cytoplasm including the nucleus, differentiates into a cyst [5,6,7,8]. Some cyst forming species also divide by longitudinal fission [7]. Colpodella unguis and C. edax do not form cysts [7,9]. Colpodella species have been identified in two human infections and in animal infections suggesting that these free-living protists are opportunistic pathogens and capable of infecting human and animal hosts [10,11]. A case of anemia in a relapsing infection caused by Colpodella sp. strain HEP, with DNA sequence homology to C. tetrahymenae was described in a patient from Yunan Province, Southwestern China infecting erythrocytes and causing a Babesia-like infection [10]. A second human case reported from Heilonjiang Province, Northeast China, was tick-borne, with the patient exhibiting neurological symptoms [11]. Polymerase chain reaction (PCR) of cerebrospinal fluid, identified DNA with sequence homology to Colpodella species and is designated Colpodella spp. HLJ. Colpodella sp. DNA was also identified in Ixodes persulatus ticks in woodlands around the patient’s living area [11]. Life cycle stages causing infection were not described, stages in the ticks were also not described and no staining was performed to identify life cycle stages of Colpodella spp. HLJ [10,11]. Colpodella gonderi was identified in the urine of a human case of urinary tract infection with an uncertain etiology, using Giemsa staining [12]. Colpodella species have been reported in various animals including in cattle and Rhipicephalus microplus ticks infesting cattle with 100% sequence identity to Colpodella sequences identified from a raccoon dog in Poland [13]. Colpodella sp. have also been identified in raccoons [14], cat [15], dog [16], goats and attached Haemaphysali longicornis ticks [17], and horses [18]. Colpodella species DNA sequences identified in the blood of a South China Tiger that died of babesiosis-like symptoms after a tick bite had 90.1% sequence identity to Colpodella sp. strain HEP and 90.4 % sequence identity to Colpodella sp. strain HLJ [19]. Ticks within the tiger enclosure and grass around the enclosure also contained Colpodella species [19]. Oligonucleotide primers targeting Cryptosporidium species identified Colpodella species from fecal samples of zoo felines in North East China [20]. For the human and animal cases reported, polymerase chain reaction (PCR) was used to identify Colpodella sp. DNA. However microscopic identify of life cycle stage morphology of Colpodella species in the human and animal hosts, ticks and flies [21,22] were not described. In two of the human cases, Giemsa stain was used to identify protists in blood and urine [10,12]. However, these were not identified as transmission or pathogenic stages and life cycle stages in erythrocytes were not easily distinguished [10]. In previous studies we described the life cycle of Colpodella sp. ATCC 50594, investigated the process of myzocytosis and identified previously undocumented life cycle stages using Sam-Yellowe’s trichrome staining protocols [23,24,25,26]. Sam-Yellowe’s trichrome staining protocol was developed to identify and distinguish cyst stages of Colpodella sp. ATCC 50594 and its prey Parabodo caudatus. Both protists are biflagellated and form cysts of similar size, indistinguishable by Giemsa staining [25]. We identified the trophozoites of V. pontica and P. cosmopolitus in earlier studies using Giemsa staining, but were unable to differentiate the cyst stages [27]. In this study our first goal was to evaluate the consistency and reproducibility of the dyes used for Sam-Yellowe’s trichrome staining. The dyes neutral red, brilliant green and methylene blue were obtained from different major vendors, with a focus on identifying young trophozoites and cysts of Colpodella sp. ATCC 50594 since these stages can be obscured by bacteria or other debris in samples and in cultures. In previous staining studies, we obtained the dyes neutral red, brilliant green and methylene blue from the same vendors [23,24,25]. The vendor supplying neutral red closed operations resulting in the purchase of neutral red from a different vendor. Initial use of neutral red from the new vendor resulted in variations in the final colors on stained cells. To ensure consistency and reproducibility of the staining protocol, we also evaluated methylene blue and brialliant green from different vendors. We show that dyes obtained from different vendors produce minor color variations in stained cells without affecting the identification of life cycle stages of Colpodella sp. ATCC 50594. Our second goal was to identify cyst stages of V. pontica and P. cosmopolitus using Sam-Yellowe’s trichrome staining series. We show for the first time that cysts of V. pontica and P. cospmopolitus in culture can be distinguished. Routine light microscopy to identify colpodellids can be achieved with specimen staining in less than ten minutes using Sam-Yellowe’s trichrome stains.
2. Materials and Methods
Voromonas pontica Cavalier-Smith (ATCC 50640) and Colpodella sp. (ATCC 50594) (American Type Culture Collection, Manassas, Virginia, USA) were maintained in diprotist cultures with prey species; Percolomonas cosmopolitus and Parabodo caudatus, respectively, as described previously [26]. Enterobacter aerogenes bacterized Hay medium (Ward’s Science, Rochester, New York, USA) was used for culture. Voromonas pontica was cultured in Hay medium (Ward’s Science) mixed with Carolina Seawater (Carolina Biological Supply Company, Burlington, North Carolina, USA). Diprotist cultures in 25 cm2 tissue culture flasks capped tightly with plug-seal caps were incubated at 22-24⁰C. Colpodella sp. was cultured as described previously [26]. Cultures were examined using an inverted microscope with phase contrast and Sam-Yellowe’s trichrome staining was performed to monitor cell growth. All cultures were maintained aseptically without antibiotics or filtration for bacteria elimination. Plasmodium falciparum (strain FCR3) was cultured as described previously and ring-infected erythrocytes fixed in absolute methanol for staining [28]. Cryptosporidium parvum oocysts were obtained from AlphaTec (https://www.alphatecsystems.com/).
Staining and Light Microscopy
Voromonas pontica ATCC 50640 and Colpodella sp. ATCC 50594 cultures were processed for formalin fixation and staining as described previously [26,27]. Smears were prepared on glass slides from the cell suspension, air-dried and either stained with Giemsa stain or Sam-Yellowe’s trichrome stain. Briefly, an equal volume of 10% formalin was added to the cell cultures of Colpodella sp. or cell suspensions of V. pontica following centrifugation of culture and resuspension of the pellet in 1X PBS. Fixed cells were centrifuged, formalin supernatants were discarded and pellets washed in 1X PBS. The formalin fixed cells were resuspended in 80 µl 1X PBS and smears prepared on glass slides for Giemsa and Sam-Yellowe’s trichrome staining.
Sam-Yellowe’s trichrome stain A with 0.3 % Methylene blue, 1 min; 1% Brilliant green, 5 min and 1% Neutral Red (Electron Microscopy Sciences, EMS), 1 min, Sam-Yellowe’s trichrome D with 1 % crystal violet, 30 seconds; 1 % Brilliant green, 2 minutes; 1% Neutral red, 1minute and Sam-Yellowe’s trichrome I with 1 % crystal violet, 1 min; 1% Brilliant green, 2 min and 1 % Safranin, 2 min [24,25] and Sam-Yellowe’s trichrome K with 0.3 % Methylene blue, 1 min; 1% Fast green (aqueous), 5 min and 1% Neutral Red [29], Table 1.
Neutral red, brilliant green and methylene blue purchased from different vendors were used in the staining protocol to evaluate differences in the final color of stained cells following staining (Table 2). The original neutral red dye obtained form ScholAR Chemistry can no longer be obtained due to the closing of the company. We therefore tested neutral red from other vendors (Table 1 and Table 2). All stained smears were examined under oil immersion at x1000 magnification and images were captured using an Olympus BX43 compound microscope attached to an Infinity HD Lumenera digital camera and Olympus U-TV0.35xc-2 adapter using Infinity HD Capture software. Images were adjusted to 300 dpi using the CMYK color mode on Adobe photo shop (CS6).
3. Results
Figure 1 shows cysts and juvenile trophozoite stages of Colpodella sp. ATCC 50594 from a diprotist culture formalin-fixed and smeared on glass slides for staining.
The life cycle stages of the protists could be detected among the background of bacteria in the diprotist culture. Young cysts of Colpodella sp. were detected (panel A, black arrows) and could be distinguished from mature cysts (panel B, white arrows). Clusters of P. caudatus cysts were also detected (red arrows, panels B and C). Free swimming juvenile trophozoite stages of Colpodella sp. ATCC 50594 with a fusiform morphology were also detected (yellow arrows). Colpodella sp. ATCC 50594 (yellow arrow) attached to prey P. caudatus (red arrow) in myzocytosis were also detected among the cysts (Figure 2A).
Colpodella sp. young trophozoites unobscured by bacteria were detected with the flagella visible (Figure 2B, black arrowheads). Voromonas pontica stained using 1% methylene blue show that trophozoite stages of V. pontica (Figure 3A–C) and its prey P. cosmopolitus (Figure 3D) can be distinguished morphologically and identified based on the number of flagella. Voromonas pontica (yellow arrow) attached to P. cosmopolitus (red arrow) (Figure 3E) can also be identified and the tubular tether (black arrow) between predator and prey identified.
However, similar to previous studies with Colpodella sp. ATCC 50594 using Giemsa staining, trophozoites of V. pontica could be distinguished from those of P. cosmopolitus but cysts could not be distinguished since both cysts stained uniformly blue. We therfore performed Sam-Yellowe’s trichrome staining on formalin-fixed V. pontica (formerly Colpodella pontica) from a diprotist culture containing the prey protist P. cosmopolitus using Sam-Yellowe’s trichrome I, shown in Figure 4.
Trophozoite stages of V. pontica and P. cosmopolitus were detected and could be distinguished by the number of flagella identified on each protist. Percolomonas cosmopolitus are tetraflagellate with three short flagella and a fourth long flagellum (Figure 4, black box), V. pontica are biflagellate with heterodynamic flagella (Figure 4, red box). Voromonas pontica (yellow arrow) and P. cosmopolitus (red arrow) attached in myzocytosis were also identified (Figure 4, green box). Voromonas pontica uses the process of myzocytosis to aspirate the cytoplasmic contents of the prey through the tubular tether. Using Sam-Yellowe’s trichrome staining, cysts of V. pontica were identified for the first time as shown in Figure 5A–D (black arrows). The cysts bear morphological resemblance to the young demilune cysts of Colpodella sp. ATCC 50594, with the partially white-blue-purple color described for Colpodella cysts. The cysts of V. pontica were identified and found mostly in clusters. Evenly stained, round cysts identified in the culture are cysts of P. cosmopolitus (white arrow).
In order to determine if Sam-Yellowe’s trichrome staining series could stain and distinguish life cycle stages of pathogenic apicompexans, blood stages of methanol-fixed P. falciparum were stained with Sam-Yellowe’s trichrome A. Ring-infected (black arrows) and uninfected erythrocytes were detected (Figure 6, panel A). Additionally, formalin-fixed Cryptosporidium parvum oocysts were also stained with Sam-Yellowe’s trichrome A (Figure 6, panel C) and oocysts (red arrows) were detected. Giemsa staining routinely used for staining methanol-fixed smears of P. falciparum and C. parvum was also used in this study for comparison. Both P. falciparum and C. parvum stained with Giemsa stain (Figure 6, panels B and D, respectively) identified ring stages of P. falciparum (panel B) and oocysts of C. parvum (panel D).
In order to evaluate dyes obtained from different vendors for consistency and reproducibility in staining protocols, we used neutral red, brilliant green and methylene blue combinations from different vendors to stain Colpodella sp. ATCC 50594 (Table 1). Figure 7A–C shows the type of variations in color obtained. Slight color variations were obtained when neutral red from different vendors was used. However trophozoites and cyst stages of predator and prey were clearly identified (Figure 7A–C).
4. Discussion
In previous studies we developed staining protocols to aid routine light microscopic examination of Colpodella sp. ATCC 50594 and its prey P. caudatus, and to distinguish cysts of both protists in a diprotist culture [25]. Light microscope visualization of stained life cycle stages allowed for clearer interpretations of transmission electron micrographs, led to the identification of previously undocumented stages in the life cycle of Colpodella sp. ATCC 50594 and aided the description of the life cycle of Colpodella sp. ATCC 50594 in cluture [23,24,25]. Previous investigations of staining protocols for Colpodella sp. ATCC 50594 included the use of monochrome and dichrome staining using individual basic and acidic dyes or the use of dyes in combinations, and the use of commercial PROTO-FIX Trichrome stain (Wheatley’s modification) [25]. Poor differentiation of stages with these and more acidic dye combinations led to the development of the three color dye protocols used in the present study [25]. The following dyes; Aqueous Fast green or Fast green in 0.5 % alcohol, Methyl green, malachite green and Janus green were substituted for brilliant green resulting in an overall reddish hue in the final color. However, life cycle stages were still identified regardlessof the combination.
Two major goals of the current study were firstly, to demonstrate that dyes obtained from different vendors used in Sam-Yellowe’s trichrome staining protocols could identify life cycle stages of Colpodella sp. ATCC 50594, particularly young trophozoite and cyst stages. Secondly, we wanted to identify and distinguish cyst stages of V. pontica (formerly Colpodella pontica) and P. cosmopolitus using Sam-Yellowe’s trichrome staining series. Cyst stages of V. pontica and P. cosmopolitus have not be described using light microscopy. In the last decade, human and animal infections by novel strains of Colpodella species have been reported and increased reports of the detection of Colpodella sp. in ticks, flies and pet dogs and cats have been made [11,19,30,31,32]. Furthermore, Colpodella sp. have been identified in ticks during surveillance studies to identify the presence and distribution of pathogens in the ticks [30,31]. The identification of Colpodella sp. in ticks and the association of tick bites in animal and human infection suggests the potential for zoonotic infections with ticks transmitting Colpodella sp. from animals to humans [11,19]. In the reported human and animal cases, molecular techniques using PCR and DNA sequencing identified sequences homologous to strains and species of Colpodella sp. identified in human infections [10,11]. DNA obtained from animal specimens, ticks and soil was also found to be homologous to the strains identified in human infections [18,19,30,31]. However, in all the cases reported, the life cycle stages of Colpodella sp. transmitting infection or responsible for pathogenesis was not described [11,12,18,19,30,31], and the mechanism of infection is unknown. The staining protocols described in this study can aid morphological identification of life cycle stages in human and animal specimens. Bi-flagellated trophozoites of Colpodella sp. can be identified as fusiform shaped cells. Immature and mature cysts are identified as irregularly shaped spheres, with the characteristic demilune colors of the young cyst stage [25]. Both stages are easily identified and distinguished from the life cycle stages of the prey and in the background of bacteria present in the culture. If attachment of Colpodella sp. to human and animal cells occurs during infection, the staining protocols will identify such attachments. In previous studies from our lab we have used Sam-Yellowe’s trichrome staining to identify and characterize predator and prey attached in myzocytosis [8,23,24]. Knowledge of transmission and pathogenic stages of Colpodella will aid proper diagnosis of infections caused by Colpodella species and avoid the confusion of infections caused by Colpodella sp. with Babesia sp. or Plasmodium sp. Yuan et al. [10] and Neculicioiu et al. [12] reported the use of Giemsa staining to identify Colpodella sp. However, details of the life cycle stages observed were not described. The search for accessible and rapid staining protocols to identify life cycle stages of pathogens and to aid diagnosis is an area of intense investigations as reported for cyst stage detection in Acanthamoeba sp. [32]. Distinct features of V. pontica morphology including the heterodynamic flagella and cell shape, can be identified with the use of Sam-Yellowe’s trichrome stains. Due to the identification of Colpodella sp. in diverse geographic locations and the confusion of symptoms in the reported cases with symptoms of babesiosis and malaria [10,11,19,30], we wanted to demonstrate that the staining protocol is robust enough to clearly differentiate other protists if present, in different tissue specimens from hosts in different geographic environments. In this study we show that Sam-Yellowe’s trichrome staining protocols developed to identify trophozoite and cyst stages of Colpodella sp. [25] can be used to identify life cycle stages of the pathogenic apicomplexans P. falciparum and C. parvum. Plasmodium falciparum ring-infected erythrocytes in the blood stage and oocysts, of C. parvum were identified by the staining protocols. The oocysts were easily recognized with Sam-Yellowe’s trichrome staining when compared to Giemsa staining. In mixed infections of piroplasms and Colpodella sp. in blood, life cycle stages of each protist can be distinguished. Similarly, in mixed Cryptosporidium sp. and Colpodella infections, protists can be identified in fecal samples, and specific life cycle stages identified. Sam-Yellowe’s trichrome staining protocol is completed in less than 10 minutes resulting in vibrant colors that clearly distinguish life cycle stages providing good contrast and differentiation of developmental stages of trophozoites and cysts [23,24,25] in the colpodellids. The use of the staining protocol to identify new Colpodella species will provide a clearer understanding of morphological similarities and differences among Colpdella species. Since neutral red can no longer be purchased from ScholAR Chemistry, we have evaluated neutral red from different vendors and find consistent and reproducible final colors using EMS neutral red. In this study we report for the first time the identification of the cyst stages of V. pontica by light microscopy and demonstrate that Sam-Yellowe’s trichrome staining protocols are robust enough to identify Colpodella sp. trophozoite and cyst stages in human and animal specimens, in ticks, flies and environmental samples such as in soil and water.
Author Contributions
Conceptualization, T.S.Y.; methodology, M.S; O.E.A; software, T.S.Y.; validation, T.S.Y, M.S., and O.E.A., ; formal analysis, T.S.Y., M.S., and O.E.A..; investigation, T.S.Y., M.S., and O.E.A.; resources, T.S.Y.; data curation, T.S.Y.; writing—original draft preparation, T.S.Y, M.S., and O.E.A.; writing—review and editing, T.S.Y.; visualization, T.S.Y., M.S., and O.E.A.; supervision, T.S.Y..; project administration, T.S.Y.; funding acquisition, T.S.Y. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by funds from the Cleveland State University Undergraduate Summer Research Award 2019 and NIH Bridges to Baccalaureate Program, Cuyahoga Community College, Cleveland, Ohio.
Acknowledgments
We gratefully acknowledge Mary M. Asraf for excellent technical assistance.
Conflicts of Interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
References
- Kuvardina, O.N.; Leander, B.S.; Aleshin, V.V.; Myl’nikov, A.P.; Keeling, P.J.; Simdyanov, T.G. The phylogeny of colpodellids (Alveolata) using small subunit rRNA gene sequences suggests they are the free-living sister group to apicomplexans. J Eukaryot Microbiol. 2002, 49:498-504. [CrossRef]
- Gile, G.H.; Slamovits, C.H. Transcriptomic analysis reveals evidence for a cryptic plastid in the colpodellid Voromonas pontica, a close relative of chromerids and apicomplexan parasites. PLoS One, 2014, 9(5):e96258. [CrossRef]
- Olmo, J.L.; Esteban, G.F.; Finlay, B.J. New records of the ectoparasitic flagellate Colpodella gonderi on non-Colpoda ciliates. J Int Microbiol, 2011, 14:207-211. [CrossRef]
- Cavalier-Smith, T.; Chao, E.E. Protalveolate phylogeny and systematics and the origins of Sporozoa and dinoflagellates (phylum Myzozoa nom. Nov.). Eur J Protistol, 2004, 40:185-212. [CrossRef]
- Simpson, A.G.B.; Patterson, D.J. Ultrastructure and identification of the predatory flagellate Colpodella pugnax Cienkowski (Apicomplexa) with a description of Colpodella turpis n. sp. and a review of the genus. Syst Parasitol, 1996, 33:187-198.
- Brugerolle, G. Colpodella vorax: Ultrastructure, predation, life-cycle, mitosis, and phylogenetic relationships. Europ. J. Protistol., 2002, 38:113-125. [CrossRef]
- Mylnikov, A.P.; Mylnikova, Z.M. Feeding spectra and pseudoconoid structure in predatory alveolate flagellates. Inland Water Biol. 2008, 1:210-216. [CrossRef]
- Sam-Yellowe, T.Y.; Fujioka, H.; Peterson, J.W. Ultrastructure of Myzocytosis and Cyst Formation, and the Role of Actin in Tubular Tether Formation in Colpodella sp. (ATCC 50594). Pathogens, 2022, 11:455.
- Mylnikov, A.P. Ultrastructure and phylogeny of colpodellids (Colpodellida, Alveolata). Biol Bulletin, 2009, 36:582-590. [CrossRef]
- Yuan, C.L.; Keeling, P.J.; Krause, P.J. et al. Colpodella spp.–like Parasite Infection in Woman, China. Emerg Infect Dis. 2012, 18:125-127.
- Jiang, J-F.; Jiang, R-R.; Chang, Q-C.; Zheng, Y-C.; Jiang, B-G.; Sun, Y.; Jia, N.; Wei, R.; Bo, H-B.; Huo, Q-B.; Wang, H.; von Fricken, M. E.; Cao, W-C. Potential novel tick-borne Colpodella species parasite infection in patient with neurological symptoms. PLOS Negl. Trop. Dis., 2018, 12(8):e0006546. [CrossRef]
- Neculicioiu, V. S.; Colosi, I. A.; Toc, D. A.; Lesan, A.; Costache, C. When a ciliate meets a flagellate: A rare case of Colpoda spp. and Colpodella spp. isolated from the urine of a human patient. Case report and brief review of the literature. Biology (Basel) 2021 Jun; 10 (6):476.
- Squarre, D.; Nakamura, Y.; Hayashida, K.; Kawai, N.; Chambaro, H.; Namangala, B.; Sugimoto, C.; Yamagishi, J. Investigation of the piroplasm diversity circulating in wildlife and cattle of the greater Kafue ecosystem, Zambia. Parasit Vectors. 2020,13:599. [CrossRef]
- Solarz, W.; Najberek, K; Wilk-Wozniak, E.; Biedrzycka, A. Raccoons foster the spread of freshwater and terrestrial microorganisms-mammals as source of microbial eDNA. Divers. Distrib. 2020, 26 453-459. [CrossRef]
- Wheatley, M.A.; Shamoun, J.; Maggi, R.; Breitschwerdt, E.B.; Sommer, S.L.; Cullen, J.M.; Stowe, D.M. Eosinophilic pericardial effusion and pericarditis in a cat. JFMS Open Rep., 2023 9:20551169231213498. [CrossRef]
- Huggins, L. G.; Colella, V.; Koehler, A. V.; Schunack, B.; Traub, R. J. A multipronged next-generation sequencing metabarcoding approach unearths hyperdiverse and abundant dog pathogen communities in Cambodia. Transbound Emerg Dis. 2022, 69:1933-1950. [CrossRef]
- Qi, Y.; Wang, J.; Lu, N.; Qi, X.; Yang, C.; Liu, B.; Lu, Y.; Gu, Y.; Tan, W.; Zhu, C.; Ai, L.; Rao, J.; Mao, Y.; Yi, H.; Li, Y.; Yue, M. Potential novel Colpodella spp. (phylum Apicomplexa) and high prevalence of Colpodella spp. in goat-attached Haemaphysalis longicornis ticks in Shandong province, China. Ticks Tick Borne Dis. 2024, 15:102328. [CrossRef]
- Xu, M.; Hu, Y.; Qiu, H.; Wang, J.; Jiang, J. Colpodella sp. (Phylum Apicomplexa) Identified in Horses Shed Light on Its Potential Transmission and Zoonotic Pathogenicity. Front Microbiol. 2022, 13:857752. [CrossRef]
- Chiu, H.C.; Sun, X.; Bao, Y.; Fu, W.; Lin, K.; Chen, T.; Zheng, C.; Li, S.; Chen, W.; Huang, C. Molecular identification of Colpodella sp. of South China tiger Panthera tigris amoyensis (Hilzheimer) in the Meihua Mountains, Fujian, China. Folia Parasitol (Praha). 2022, 69:2022.019. [CrossRef]
- Hussein, S.; Li, X.; Bukharr, S. M.; Zhou, M.; Ahmad, S.; Amhad, S.; Javid, A.; Guan, C.; Hussain, A.; Ali, W.; Khalid, N,;Ahmad, U, Tian, L.; Hou, Z. Cross-genera amplification and identification of Colpodella sp. with Cryptosporidium primers in fecal samples of zoo felids from northeast China. Braz J Biol. 2021 Sep 6; 83:e247181.doi.10.1590/1519-6984.247181 eCollection 2021. [CrossRef]
- Neupane, S.; Saski, C.; Nayduch, D. House fly larval grazing alters dairy cattle manure microbial communities. BMC Microbiol. 2021, 21:346. [CrossRef]
- Phetkarl, T.; Fungwithaya, P.; Udompornprasith, S.; Amendt, J. Sontigun, N. Preliminary study on prevalence of hemoprotozoan parasites harbored by Stomoxys (Diptera: Muscidae) and tabanid flies (Diptera: Tabanidae) in horse farms in Nakhon Si Thammarat province, Southern Thailand. Vet World. 2023, 16:2128-2134. [CrossRef]
- Getty, T.A.; Peterson, J.W.; Fujioka, H.; Walsh, A.M.; Sam-Yellowe, T.Y. Colpodella sp. (ATCC 50594) Life Cycle: Myzocytosis and Possible Links to the Origin of Intracellular Parasitism. Trop Med Infect Dis., 2021, 6:127. [CrossRef]
- Sam-Yellowe, T.Y.; Getty, T.A.; Addepalli, K.; Walsh, A.M.; Williams-Medina, A.R.; Fujioka, H.; Peterson, J.W. Novel life cycle stages of Colpodella sp. (Apicomplexa) identified using Sam-Yellowe’s trichrome stains and confocal and electron microscopy. Int Microbiol., 2022, 25:669-678.
- Sam-Yellowe, T.Y.; Addepalli, K.; Yadavalli, R.; Peterson, J.W. New trichrome stains identify cysts of Colpodella sp. (Apicomplexa) and Bodo caudatus. J Int Microbiol, 2019, 23:303-311. [CrossRef]
- Yadavalli, R.; Sam-Yellowe, T.Y. Developmental stages identified in the trophozoite of the free-living Alveolate flagellate Colpodella sp. (Apicomplexa). J Int Microbiol, 2017, 20:178-183.
- Sam-Yellowe, T.Y.; Yadavalli, R. Voromonas pontica Identified by Giemsa Staining and Anti-RhopH3 Protein Reactivity. J Microbiol Modern Tech, 2019, 4(1): 103.
- Yadavalli, R.; Peterson, J.W.; Drazba, J.A.; Sam-Yellowe, T.Y. Trafficking and Association of Plasmodium falciparum MC-2TM with the Maurer’s Clefts. Pathogens, 2021, 10:431. [CrossRef]
- Sam-Yellowe, T. Y.; Asraf, M. M.; Peterson, J. W.; Fujioka, H. Fluorescent Nanoparticle Uptake by Myzocytosis and Endocytosis in Colpodella sp. ATCC 50594. Microorganisms. 2023, 11:1945.
- Matsimbe, A. M.; Magaia, V.; Sanchez, G. S.; Neves, L.; Noormahomed, E.; Antunes, S.; Domingos, A. Molecular detection of pathogens in ticks infesting cattle in Nampula province, Mozambique. Exp. Appl. Acarol. 2017, 73, 91-102. [CrossRef]
- Zhao, G.P., Wang, Y.X.; Fan ZW, Ji Y, Liu MJ, Zhang WH, Li XL, Zhou SX, Li H, Liang S, Liu W, Yang Y, Fang LQ. Mapping ticks and tick-borne pathogens in China. Nat Commun. 2021 Feb 17;12(1):1075.
- Wu, S. Meng, J.; Yu, F.; Zhou, C.; Yang, B.; Chen, X.; Yang, G.; Sun, Y.; Cao, W.; Jiang, J.; Wu, J.; Zhan, L. Molecular epidemiological investigation of piroplasms carried by pet cats and dogs in an animal hospital in Guiyang, China. Front Microbiol. 2023,14:1266583.
- El-Sayed, N.M.; Hikal, W.M. Several staining techniques to enhance the visibility of Acanthamoeba cysts. Parasitol Res, 2015, 114:823-830. [CrossRef]
Figure 1.
Sam-Yellowe’s trichrome A staining of the predator Colpodella sp. ATCC 50594 cysts and young trophozoite stages (yellow arrows). A. Young dual-colored demilune cysts of Colpodella sp. ATCC 50594 showing a partially light blue-white and dark blue-purple staining pattern (black arrows). B. Mature Colpodella sp. ATCC 50594 cysts are shown in panel B (white arrows) showing the initiation of division with the appearance of two to three “lobes” with an even dark blue-brownish appearance. Free swimming young trophozoites of Colpodella sp. ATCC 50594 are shown in panels C and D (yellow arrows, fusiform-shaped trophozoites). Clusters of Parabodo caudatus (prey) cysts are indicated by the red arrows (panels B and C). Background of cells shown are bacteria from the diprotist culture. Anti-bacterial zones were observed around the young and mature cysts of Colpodella sp. 50594 (black and white arrows). Two different lots of neutral purchased at different times from ScholAR Chemistry were used for staining. 1% neutral red used in panel A was an older lot. In panels B-D a second dye lot from ScholAR Chemistry before the company closed. Cells from diprotist culture were formalin-fixed. Scale bars, 20 µm.
Figure 1.
Sam-Yellowe’s trichrome A staining of the predator Colpodella sp. ATCC 50594 cysts and young trophozoite stages (yellow arrows). A. Young dual-colored demilune cysts of Colpodella sp. ATCC 50594 showing a partially light blue-white and dark blue-purple staining pattern (black arrows). B. Mature Colpodella sp. ATCC 50594 cysts are shown in panel B (white arrows) showing the initiation of division with the appearance of two to three “lobes” with an even dark blue-brownish appearance. Free swimming young trophozoites of Colpodella sp. ATCC 50594 are shown in panels C and D (yellow arrows, fusiform-shaped trophozoites). Clusters of Parabodo caudatus (prey) cysts are indicated by the red arrows (panels B and C). Background of cells shown are bacteria from the diprotist culture. Anti-bacterial zones were observed around the young and mature cysts of Colpodella sp. 50594 (black and white arrows). Two different lots of neutral purchased at different times from ScholAR Chemistry were used for staining. 1% neutral red used in panel A was an older lot. In panels B-D a second dye lot from ScholAR Chemistry before the company closed. Cells from diprotist culture were formalin-fixed. Scale bars, 20 µm.
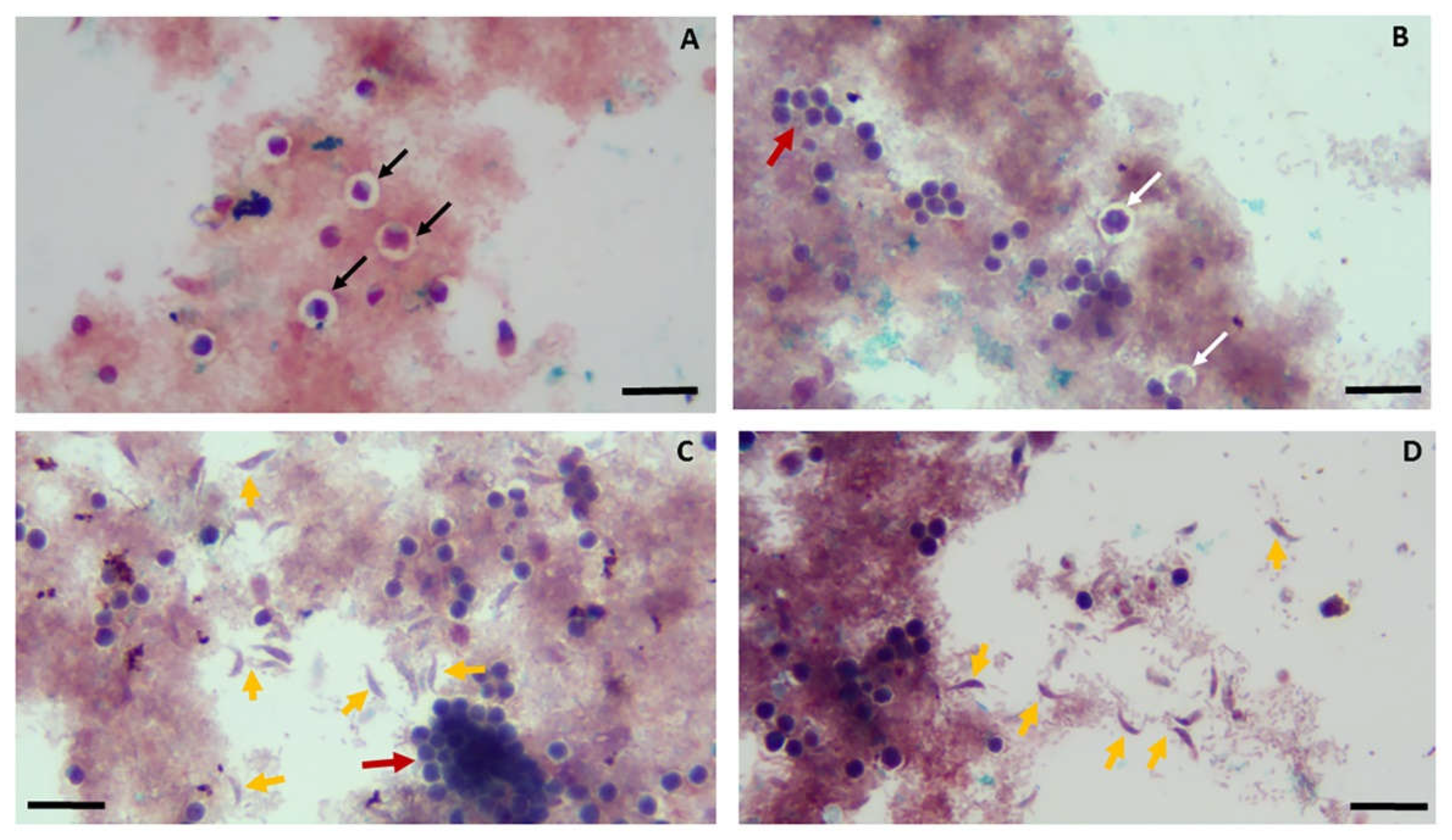
Figure 2.
Sam-Yellowe’s trichrome A staining of Colpodella sp. ATCC 50594 cysts, trophozoites, young trophozoite stages (yellow arrows) and predator and prey in myzocytosis. Panel shows young Colpodella cysts (black arrows), and the predator Colpodella sp. ATCC 50594 (yellow arrows) and the prey P. caudatus (red arrows) attached in myzocytosis. Panels B and C show young free-swimming trophozoites of Colpodella sp. 50594. 1% neutral red purchased from EMS was used in panels A-C. Cells from diprotist culture were formalin-fixed. Scale bars, 20 µm.
Figure 2.
Sam-Yellowe’s trichrome A staining of Colpodella sp. ATCC 50594 cysts, trophozoites, young trophozoite stages (yellow arrows) and predator and prey in myzocytosis. Panel shows young Colpodella cysts (black arrows), and the predator Colpodella sp. ATCC 50594 (yellow arrows) and the prey P. caudatus (red arrows) attached in myzocytosis. Panels B and C show young free-swimming trophozoites of Colpodella sp. 50594. 1% neutral red purchased from EMS was used in panels A-C. Cells from diprotist culture were formalin-fixed. Scale bars, 20 µm.
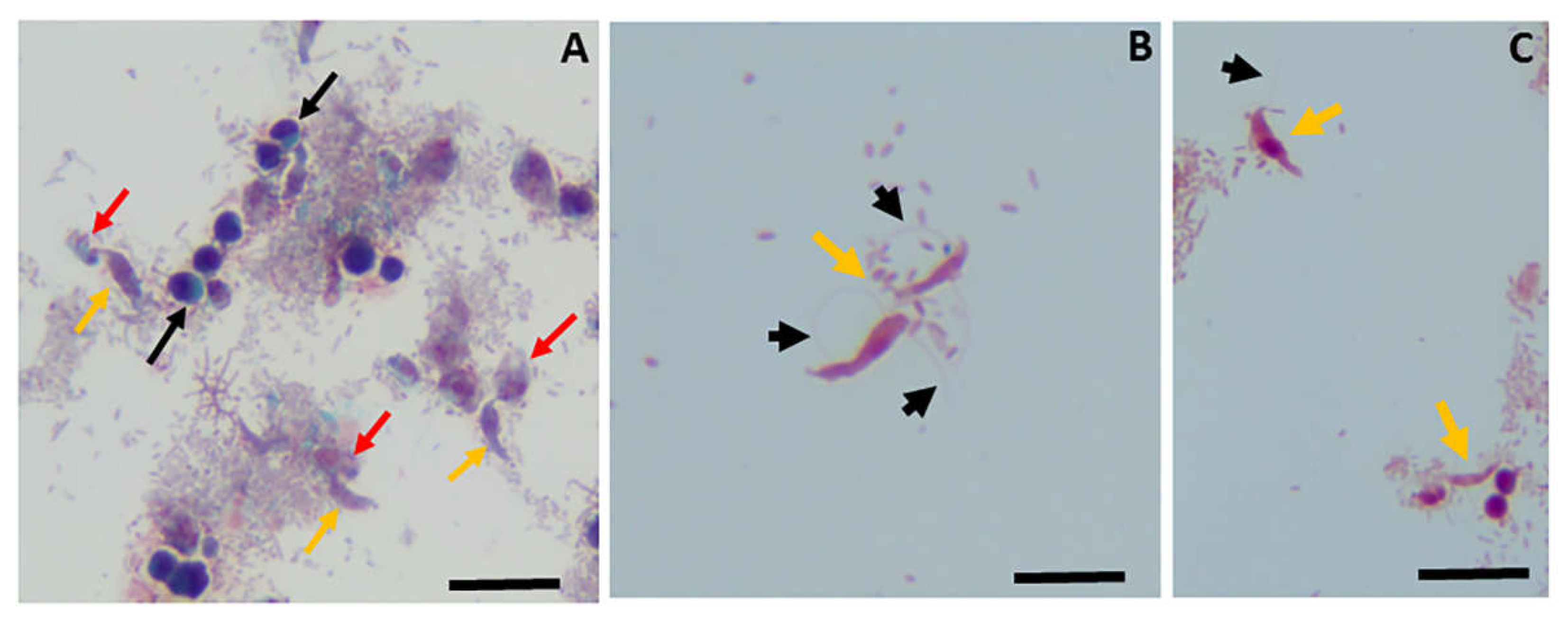
Figure 3.
1% Methylene blue staining of Voromonas pontica and Percolomonas cosmopolitus. Panels A-C, biflagellate predator V. pontica; panel D, tetraflagellate prey P. cosmopolitus; panel E, V. pontica (yellow arrow) and P. cosmopolitus (red arrow) attached in myzocytosis. Black arrow shows tubular tether formed between predator and prey during myzocytosis and used to aspirate cytoplasmic contents of the prey. Cells from diprotist culture were formalin-fixed. Scale bars panels A-D, 20 µm; panel E, 30µm.
Figure 3.
1% Methylene blue staining of Voromonas pontica and Percolomonas cosmopolitus. Panels A-C, biflagellate predator V. pontica; panel D, tetraflagellate prey P. cosmopolitus; panel E, V. pontica (yellow arrow) and P. cosmopolitus (red arrow) attached in myzocytosis. Black arrow shows tubular tether formed between predator and prey during myzocytosis and used to aspirate cytoplasmic contents of the prey. Cells from diprotist culture were formalin-fixed. Scale bars panels A-D, 20 µm; panel E, 30µm.
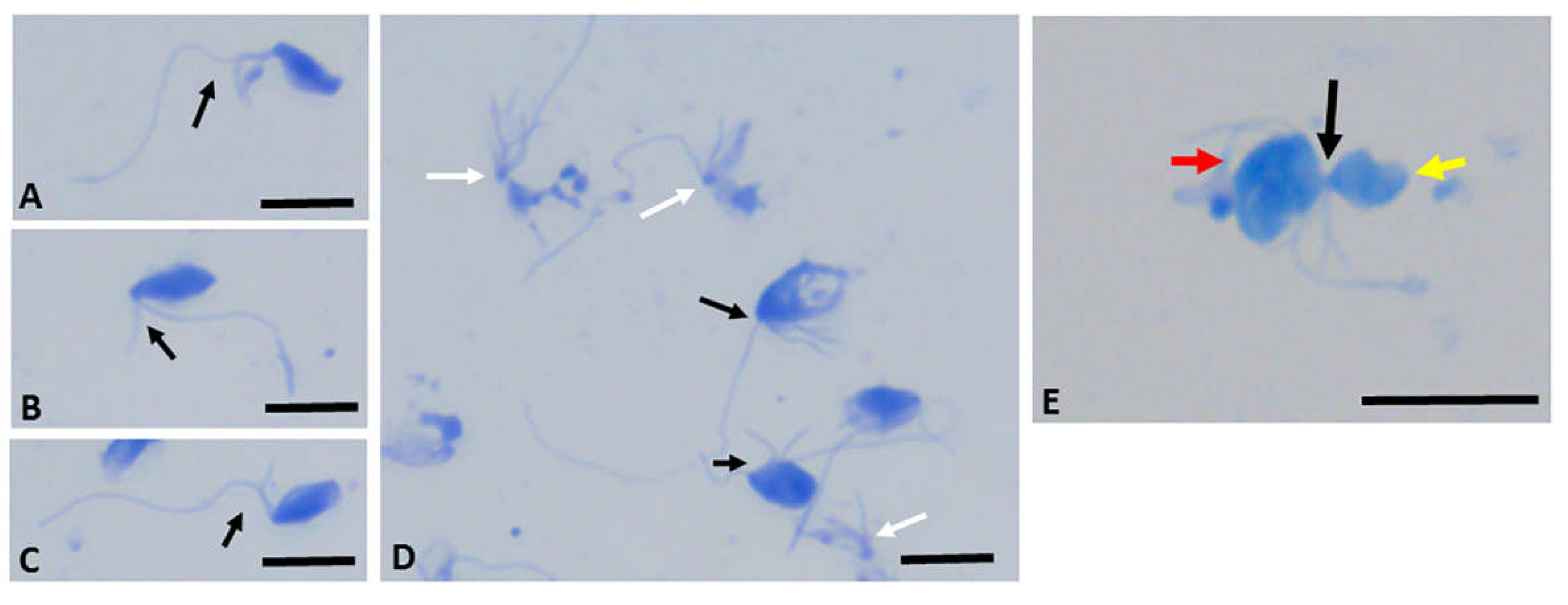
Figure 4.
Sam-Yellowe’s trichrome I staining of a diprotist culture of the marine predator Voromonas pontica (bi-flagellate) and its prey Percolomonas cosmopolitus (tetraflagellate). Percolomonas cosmopolitus trophozoites (black box), V. pontica (red box) and cells in myzocytosis (green box), V. pontica (yellow arrow) and P. cosmopolitus (red arrow) in attached in the process of myzocytosis. Cells from diprotist culture were formalin-fixed. Scale bars, 20 µm.
Figure 4.
Sam-Yellowe’s trichrome I staining of a diprotist culture of the marine predator Voromonas pontica (bi-flagellate) and its prey Percolomonas cosmopolitus (tetraflagellate). Percolomonas cosmopolitus trophozoites (black box), V. pontica (red box) and cells in myzocytosis (green box), V. pontica (yellow arrow) and P. cosmopolitus (red arrow) in attached in the process of myzocytosis. Cells from diprotist culture were formalin-fixed. Scale bars, 20 µm.
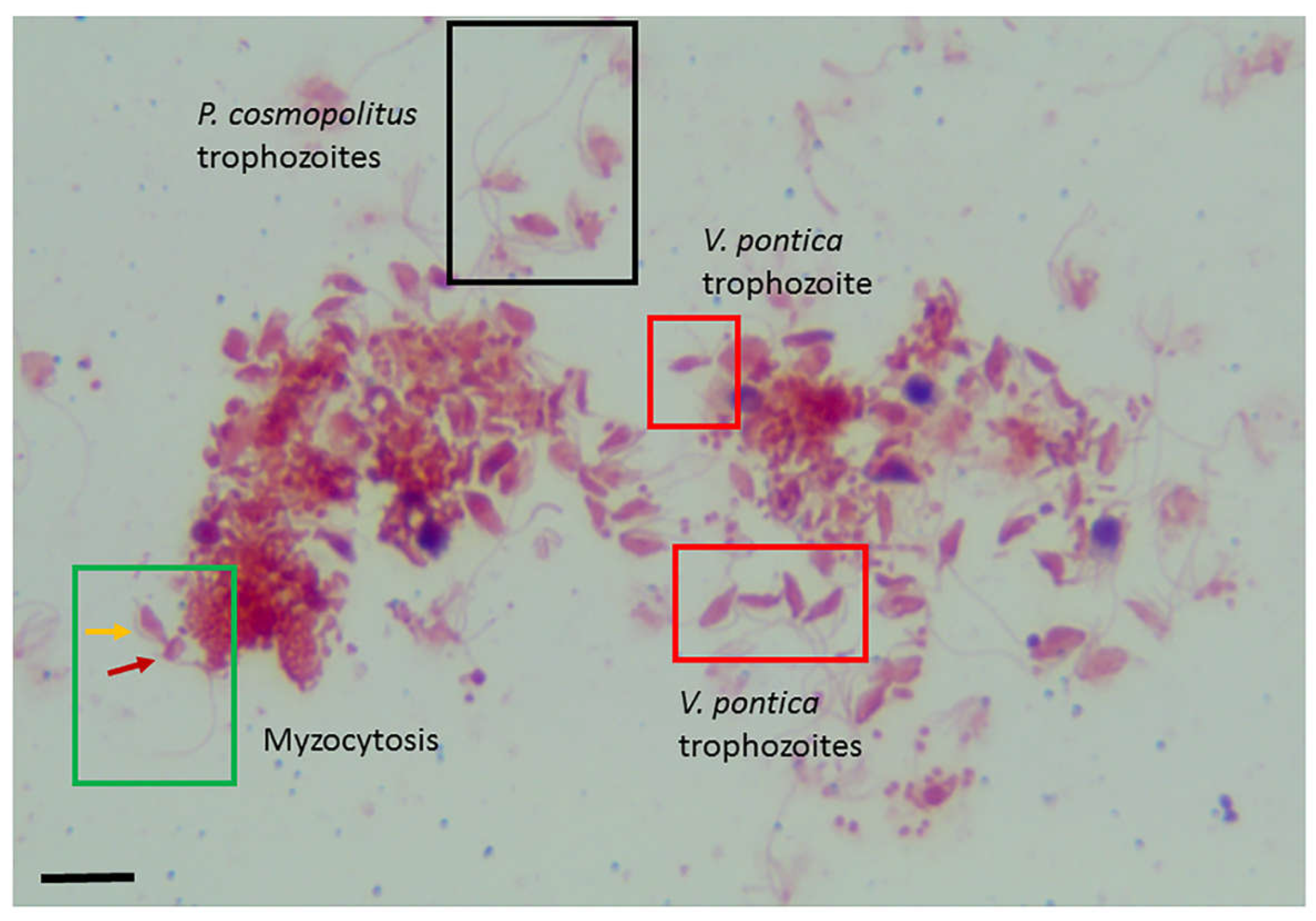
Figure 5.
Sam-Yellowe’s trichrome I staining of a diprotist culture of Voromonas pontica and P. cosmopolitus showing cyst stages of V. pontica (black arrows, panels A-D) and P. cosmopolitus (white arrow, panel D). Cells from diprotist culture were formalin-fixed. Scale bars, 20 µm.
Figure 5.
Sam-Yellowe’s trichrome I staining of a diprotist culture of Voromonas pontica and P. cosmopolitus showing cyst stages of V. pontica (black arrows, panels A-D) and P. cosmopolitus (white arrow, panel D). Cells from diprotist culture were formalin-fixed. Scale bars, 20 µm.
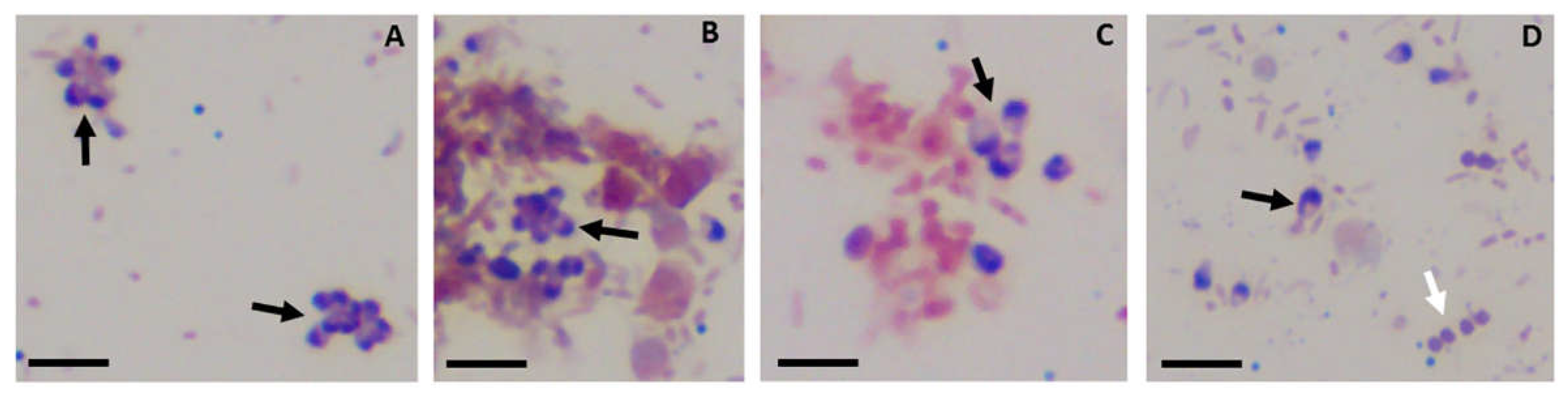
Figure 6.
Sam-Yellowe’s trichrome A and Giemsa staining of Plasmodium falciparum and Cryptosporidium parvum. Ring and trophozoite stages of methanol-fixed P. falciparum were stained by Sam-Yellowe’s trichrome A (panel A) and Giemsa stain (panel B). Black arrows show ring stages of P. falciparum. Oocysts of C. parvum were stained with Sam-Yellowe’s trichrome A (panel C) and Giemsa stain (panel D). Red arrows show oocysts of C. parvum.
Figure 6.
Sam-Yellowe’s trichrome A and Giemsa staining of Plasmodium falciparum and Cryptosporidium parvum. Ring and trophozoite stages of methanol-fixed P. falciparum were stained by Sam-Yellowe’s trichrome A (panel A) and Giemsa stain (panel B). Black arrows show ring stages of P. falciparum. Oocysts of C. parvum were stained with Sam-Yellowe’s trichrome A (panel C) and Giemsa stain (panel D). Red arrows show oocysts of C. parvum.
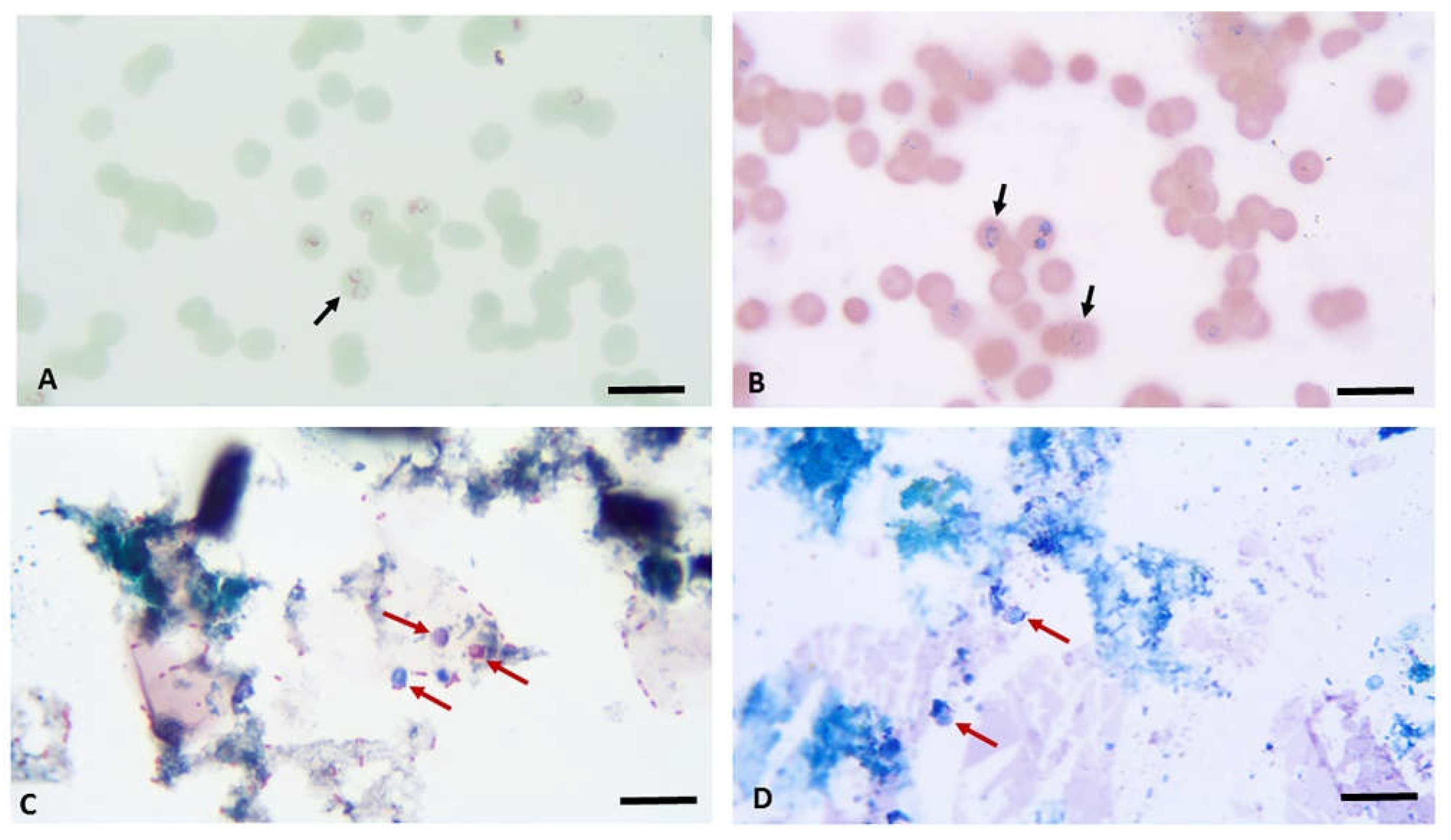
Figure 7.
Sam-Yellowe’s trichrome A performed using neutral red dye from three diferent major vendors. A, Carolina neutral red; B, ScholARs neutral red and C, Ward’s neutral red. Brilliant green from ACROS and methylene blue from EMD were used for the staining protocol. Yellow arrows indicate Colpodella trophizoites, red arrow (panel A) shows Parabodo caudatus trophozoite in myzocytosis with Colpodella trophozoite. Red arrow heads indicate P. caudatus trophozoites. Black arrows indicate young demilune Colpodella cysts and blue arrow head (panel C) shows a cluster of P. caudatus cysts. 1% neutral red used in panel B was from the last lot purchased from ScholAR Chemistry before the company closed. Scale bar, 10 µm.
Figure 7.
Sam-Yellowe’s trichrome A performed using neutral red dye from three diferent major vendors. A, Carolina neutral red; B, ScholARs neutral red and C, Ward’s neutral red. Brilliant green from ACROS and methylene blue from EMD were used for the staining protocol. Yellow arrows indicate Colpodella trophizoites, red arrow (panel A) shows Parabodo caudatus trophozoite in myzocytosis with Colpodella trophozoite. Red arrow heads indicate P. caudatus trophozoites. Black arrows indicate young demilune Colpodella cysts and blue arrow head (panel C) shows a cluster of P. caudatus cysts. 1% neutral red used in panel B was from the last lot purchased from ScholAR Chemistry before the company closed. Scale bar, 10 µm.
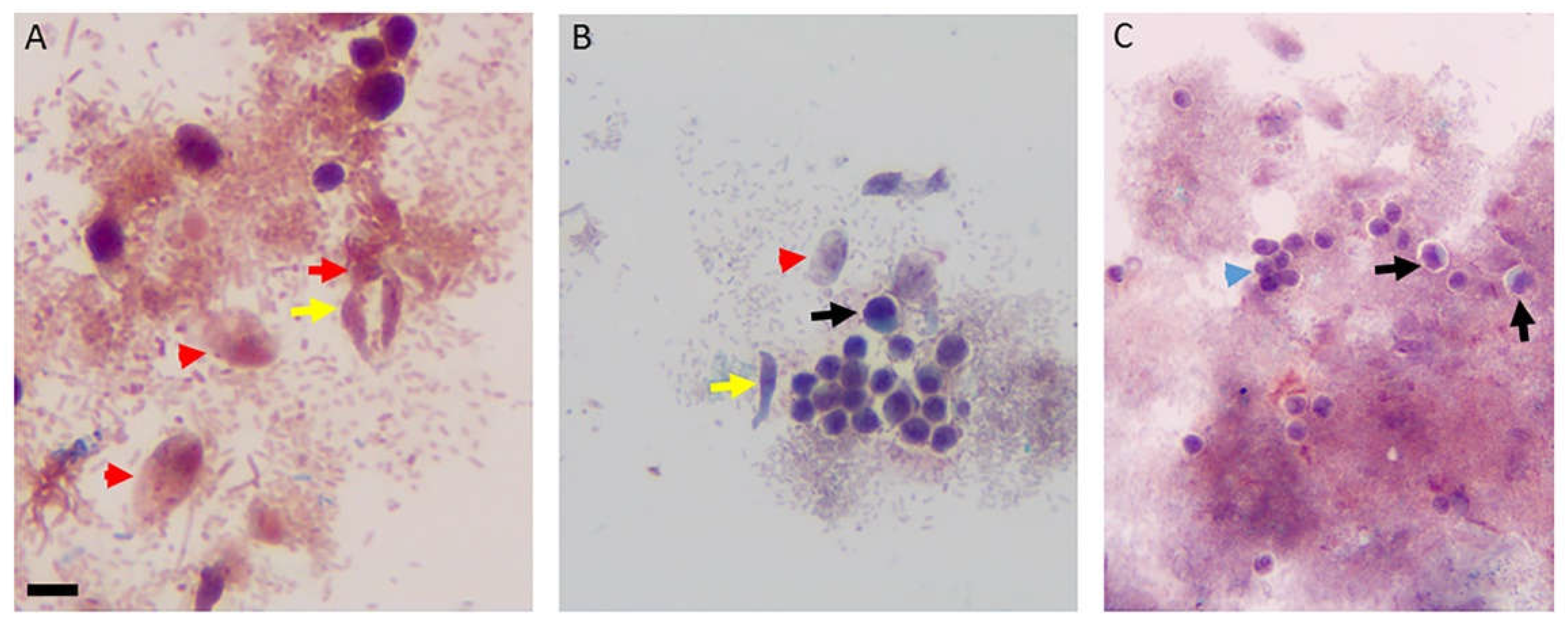

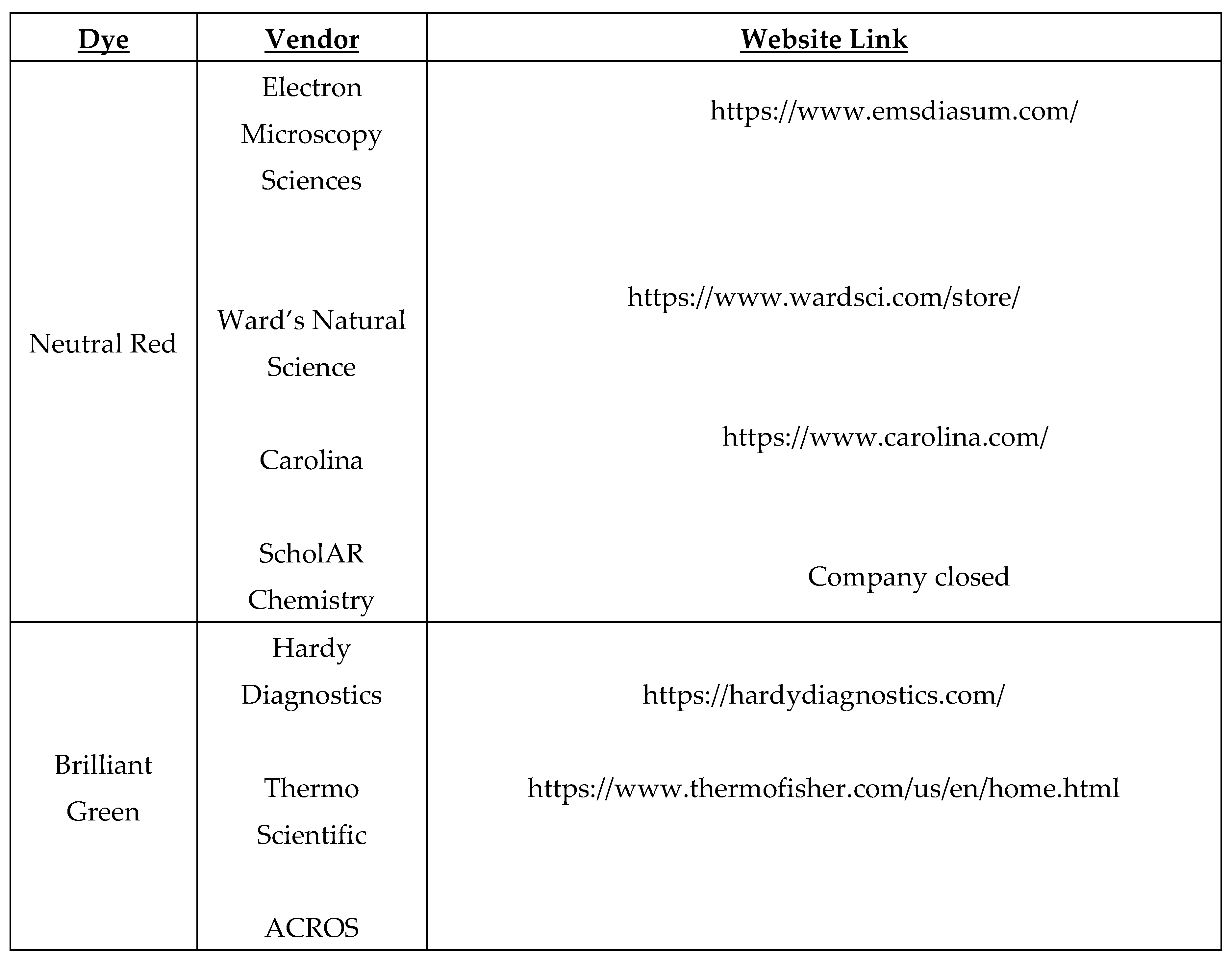
Table 1.
Sam-Yellowe’s trichrome staining protocol with dyes from different vendors.
| 1 Minute | 5 Minutes | 1 Minute |
|---|---|---|
| Fisher Chemical 0.3% Methylene Blue | ACROS 1% Brilliant Green | EMS 1% Neutral Red |
| EMD 0.3% Methylene Blue | ACROS 1% Brilliant Green | EMS 1% Neutral Red |
| EMD 0.3% Methylene Blue | ACROS 1% Brilliant Green | Ward’s Science 1% Neutral Red |
| EMD 0.3% Methylene Blue | ACROS 1% Brilliant Green | Ward’s Science 1% Neutral Red |
| EMD 0.3% Methylene Blue | ACROS 1% Brilliant Green | ScholAR Chemistry 1% Neutral Red |
| EMD 0.3% Methylene Blue | ACROS 1% Brilliant Green | Carolina 1% Neutral Red |
| EMD 0.3% Methylene Blue | Thermo Scientific 1% Brilliant Green | EMS 1% Neutral Red |
| EMD 0.3% Methylene Blue | Thermo Scientific 1% Brilliant Green | ScholAR Chemistry 1% Neutral Red |
| EMD 0.3% Methylene Blue | Hardy Diagnostic 1% Brilliant Green | EMS 1% Neutral Red |
| EMD 0.3% Methylene Blue | Hardy Diagnostic 1% Brilliant Green | SchoAR Chemistry 1% Neutral Red |
| EMD 0.3% Methylene Blue | Sigma 1% Fast Green | EMS 1% Neutral Red |
| EMD 0.3% Methylene Blue | Sigma 1% Fast Green | ScholAR Chemistry 1% Neutral Red |
| EMD 0.3% Methylene Blue | Carolina 1% Methyl Green | EMS 1% Neutral Red |
| EMD 0.3% Methylene Blue | Carolina 1% Methyl Green | ScholAR Chemistry 1% Neutral Red |
| EMD 0.3% Methylene Blue | Ward’s Science 1% Malachite Green | EMS 1% Neutral Red |
| EMD 0.3% Methylene Blue | Ward’s Science 1% Janus Green | EMS 1% Neutral Red |
| EMD 0.3% Methylene Blue | Ward’s Science 1% Janus Green | ScholAR Chemistry 1% Neutral Red |
| EMD 0.3% Methylene Blue | Carolina 0.5% Fast Green Alcohol | EMS 1% Neutral Red |
| EMD 0.3% Methylene Blue | Carolina 0.5% Fast Green Alcohol | ScholAR Chemistry 1% Neutral Red |
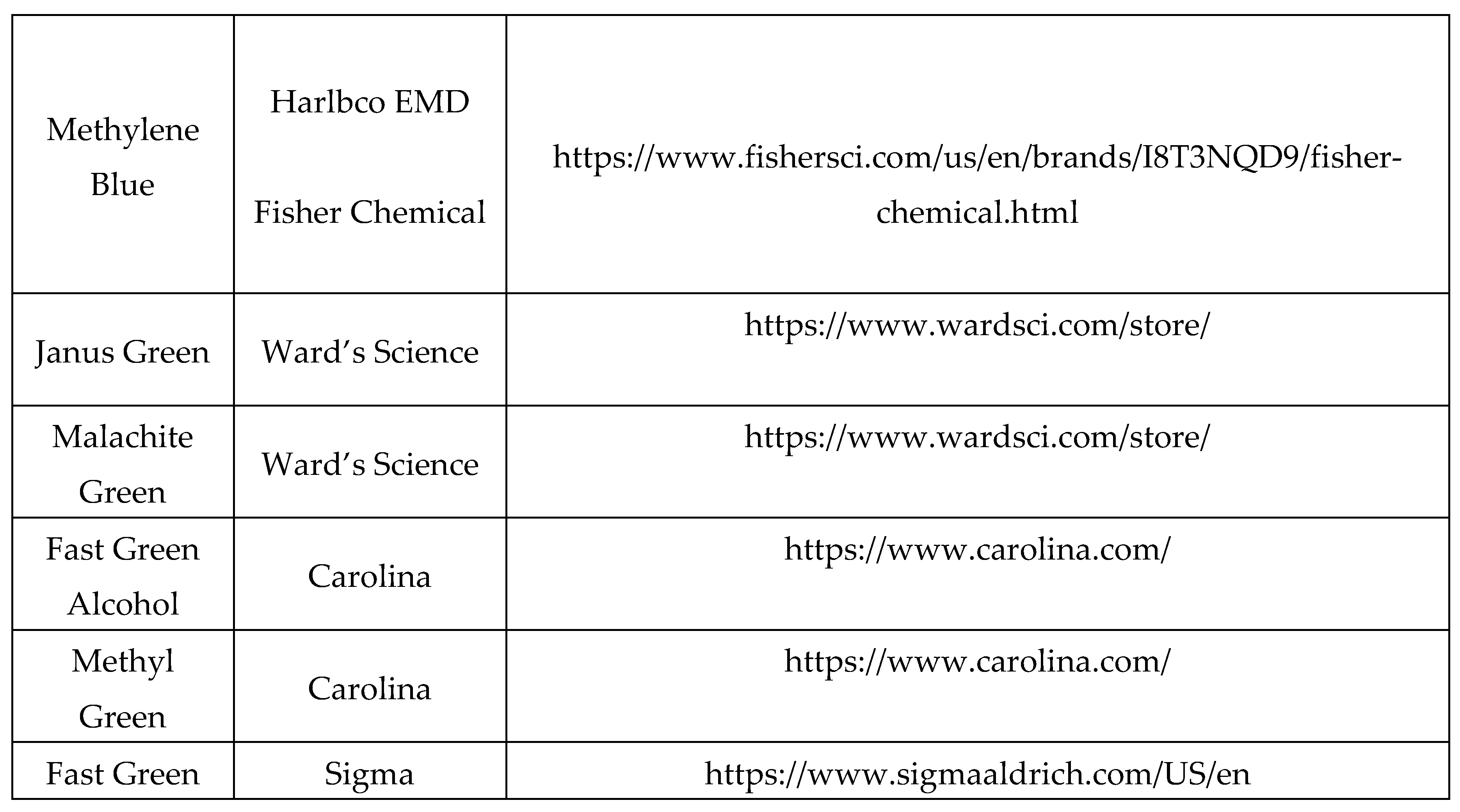
Table 2.
Dyes from Major Vendors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



