
Submitted:
26 April 2024
Posted:
28 April 2024
You are already at the latest version
Abstract
Dissemination of resistant pathogens via food supply chains is a notable public health danger from farm to fork. The proposed work analysed the distribution of E. coli among various sources of animal-based food supply chain. Overall, 500 samples were heaped up from livestock, poultry sectors, environment, fisheries, and dairy. Standard microbiological procedures were used to isolate and identify E. coli. Further, isolates were confirmed by using MALDI-TOF and virulence associated genes (VAGs). The phenotypic resistance pattern of the isolates was determined by molecular identification of antibiotic resistance genes (ARGs) through PCR using specific primers. Shiga toxin producing E. coli (STEC) were subjected to PCR based O typing using specific primers against different O types. Overall results revealed that 154 (30.5%) samples were confirmed as E. coli, in which77 (50%) isolates were MDR E. coli. Among these, total 52 (67.53%) isolates showed array of VAGs in which 21 (40.38%) were confirmed as STEC based on stx1 and stx2. Furthermore, 12/52 (23.07%) isolates were detected as Non-O157 STEC co-harboring mcr-1 and blaNMD-1. Overall, O26 STEC was found to be the most prevalent among different non-O157 types. Herein, results proposed that detection of such STEC MDR E. coli causing serious public health consequences in developing countries.
Keywords:
AMR
; Food supply chain
; Non-O157
; MDR
; STEC
1. Introduction
The aberrant antimicrobial usage in food producing animals (FPAs) either for growth enhancement or as precautionary measure resulted in selection pressure steers the evolution of resistant bacterial strains. Such resistant bacteria further spread via food supply chain to humans via consumption of contaminated food, direct contact with FPAs or indirectly by animal waste that contaminated the animal production systems, agriculture, aquaculture and environment [1]. Animal based food chains are very complicated due to great diversity in food production, processing, packaging, and transportation which mostly resulted in reoccurrence of microbes responsible for foodborne illnesses [2].
Foodborne infections (FBIs) are serious global health concerns result in substantial socioeconomic impact and are associated with notable morbidity and high death rate among various age groups across the globe. Largely, FBIs are caused by specific microbial pathogens or other food contaminant e.g. bacteria, fungi, and mycotoxins. Addiotnally, environmental contaminants may also be associated with such infections, which may contaminate food during production or transportation [3]. Bacteria remain one of the most common causes of FBIs around the world, such as Shiga toxin producing E. coli (STEC), Salmonella, Campylobacter and Listeria etc [4].
STEC is one of the recognized zoonotic foodborne bacterial isolate, responsible for causing, that madiarrhea, hemorrhagic colitis, and hemolytic uremic syndrome. Nonetheless, the emergence of Non-O157-STEC in recent times has been reported sporadically and considered as a potential health threat [5]. Several serogroups such as O26, O103, and O145 etc. are among prevalent O types during different recorded outbreaks of STEC. Shiga toxin (Stx) produced by STEC contains two distinct immunodominant epitopes i.e. Stx1 and Stx2. These Stx toxins play a critical role in the virulence of STEC inside the host for the onset of infection. The Shiga toxins serve as a ribotoxin which stalls synthesis of proteins inside the eukaryotic host cells and results in apoptosis [6].
Identifying specific research gaps in the context of STEC MDR E. coli transmission in food supply chains is critical for developing targeted interventions and mitigating the associated risks. In recent past, various studies showing public health significance of Non-O157 STEC occurance from various parts of the world [7]. FPAs are recognized reservoirs of STECs, characterization of STEC from a range of domesticated animals has been reported by now. Previously, we have reported MDR E. coli from poultry, fresh and frozen meat [8,9]. Keeping this idea, the present study is proposed to investigate MDR STEC is range of sources comprised food supply chain, as there is limited data available on STEC from farm to fork due to improper surveillance and monitoring programms.
2. Materials and Methods
2.1. Ethical Approval
The present study was conducted after proper approval from the Institutional Review Board (IRB) and Ethical Review Committee (ERC) Ref No. GCUF/ERC/137, Date: 3-02-2023, Government College University Faisalabad. Samples were collected with prior permission from stakeholders on written consent. All the experiments were performed in the “One Health AMR laboratory (OH-AMR lab)” at the Institute of Microbiology, Government College University Faisalabad. The matrix-assisted Laser Deionization-TOF experiment was conducted at the National Institutes of Health, M4QP+GW7 Islamabad, Pakistan
2.2. Collection, Processing and Transportation of Sample
Overall, N = 500 samples were collected in containers under sterile condition from different food and related sources of animal origin, including livestock samples (n= 75) chicken, meat and veal, poultry samples (n=75) chicken meat, cloacal/anal swabs and droppings, environmental samples(n=125) containing slaughter house, open market, dairy and poultry waste and various transport means, fisheries(n= 100) including fish, shrimps, market waste and transport means and dairy samples(n= 125)including raw milk, yoghurt, dairy cream, cheese and outlet waste. Samples were enriched in peptone water and transported in icebags, shifted it to the laboratory, Institute of Microbiology, for future investigation (Figure 1).
2.3. Isolation and Purification of E. coli
Enriched samples were streaked on MacConkey agar and EMB agar (OXOID®, Basingstoke, UK), and incubation was done for 24 h at 37 oC. Samples were proceed for cultural and morphological characteristics. Fuerthermore, isolates were subjected for Biochemical identification using an API 20E kit (Biomeurex, Craponne, France) according to the manufacturer protocol.
2.4. MALDI-TOF
All the isolates were confirmed through Matrix assisted laser deionization- time of flight-based VITEK® MS V3.2 (Biomeurex, Craponne, France). All the procedure was carried out as per guidelines and recommendations available in the manufacturer manual and protocol. The cyanohyrodxycinamic acid (CHCA @ 0.01 µL) was used as matrix for the isolates. Whereas ATCC™ E. coli (8739) was kept as control. The slides for VITEK® MS were prepared by using Vitek® PICKME NIBS (Biomeurex, Craponne, France), ATCC™ control and the isolates were inoculated and directed circles, correspondingly. Subsequently, CHCA (0.01 µL). was added to make the tested sample ready to be interpreted through MYLA® software (Biomeurex, Craponne, France).
2.5. VAGs-Based Detection
Detection of different VAGs was done through PCR, using specific primers in addition to the marker VAGs for E. coli, e.g., stx1, stx2, ompT, hylF, iutA, fimH and iss etc. The confirmation of isolates was done after observing the presence of a minimal three E. coli specific VAGs.To achieve this, DNA extraction using the GeneJET Genomic DNA purification kit K0722 was used (Thermo-Scientific™), according to the given protocol.
Follwinf that, for VAGs, PCR (T3000, Thermo-cycler:48 Biomerta™, Göttingen, Germany) was carried out with specific conditions , along with their respective annealing temperature. The reaction setup was contained initial denaturation of template DNA at 94 C for 5min, followed by 30 reaction cycles of DNA enaturation at 94 oC for 1min, specific primer annealing for 55s, and extension at 72 oC for 60s. Finally, a 10min extension was done at 72 oC.
The PCR reaction mixture (25 µL) compromised: 5 µL template DNA, F&R primers (100 pM) 1 µL each, DreamTaq (Thermo-Scientific™, Waltham, MA, USA) 8 µL, and SuperQ water (Ambion-AM9932, Thermo-Scientific™, Waltham, MA, USA) 10 µL. For the observation of PCR amplicons, agarose (CSL-AG500; CLEAVER SCIENTIFIC®, Rugby, UK) gel electrophoresis was conducted and visualized under a gel trans-illuminator (BioRad, Hercules, CA, USA).
2.6. Antibiotic Susceptibility Testing
Antibiotic resistance profiling of E. coli isolates (n = 52) was conducted by Kirby–Bauer disc diffusion assay according to the 2021 CLSI guidelines. Antibiotics involved in the study include Ampicillin, cefepime, ciprofloxacin, levofloxacin, chloramphenicol, trimethoprim, imipenem, meropenem, colistin, tetracycline, and tigecycline. E. coli ATCC™8739 was used as quality control during the process. According to CLSI guidelines the Broth microdilution method (BMD) was used to estimate the minimum inhibitory concentration (MIC), while colistin and tigecycline were estimated according to the EUCAST-CLSI and FDA recommendations.
2.7. Phenotypic Confirmation of Colistin Resistance
The Rapid Polymyxin Test (RPT) was carried out for the phenotypic confirmation of colistin resistant isolates as described previously (Nordmann et al., 2016). Polymyxin stock solution was prepared in Muller Hinton broth (MHB) with colistin (Oxoid™), as a final concentration of 0.2 mg/mL was reached. Afterwards, rapid polymyxin NP solution (RPS) was prepared by adding 6.25g of MHB and .0125g phenol red, the pH of the RPS was adjusted at 6.7. Moreover, sterilized filtered D + glucose (1 %) was also added in RPS. At the start of the experiment colistin was dispensed in RPS to get a solution having colistin @ 5 μg/150μL. Bacterial inoculum was prepared from freshly grown E. coli culture; bacterial colonies were resuspended to obtain 3.5 McFarland standard for further procedure. The procedure was performed in 96 well plates, EUCAST recommendations were followed for the preparation of bacterial suspension and usage i.e. 15 minutes post preparation and no longer than 60 minutes. All the isolates that grew in the presence of colistin were considered as colistin resistant isolates
2.8. CarbaNPCLSI Test
CLSI recommended CarbNP assay was carried out to confirm carbapenem resistant isolates (CLSI 2018). Two Eppendorf tubes were prepared and labeled separately by adding 20 mM Tris-HCl-buffer (100 µL) in each. Moreover, two solutions were prepared named solution A and Solution B. The solution A was prepared with 0.1 mm/L ZnSO4 and 0.5% phenol red indicator, pH of solution A was kept at 7.8. Whereas, to get solution B imipenem @ 6 mg/mL was mixed in solution A. Afterwards, solution A was dispensed in tube 1 and solution B was added in tube 2. Both tubes were incubated for 2. +15 hours at 37 oC, appearance of yellow color in tube B was interpreted as positive for CarbNP test.
2.9. Molecular Characterization of ARGs
Along with phenotypic-resistance profiling, various ARGs were identified with the help of PCR using specific primers. The DNA was extracted using a Genomic DNA purification kit K0722 (Thermo-Scientific™, Waltham, MA, USA). A total of 25 µL reaction mixture for PCR was comprised of 5 µL DNA template, 10 µL Green DreamTaq mix (Thermo Fisher Scientific, Waltham, MA, USA), and F&R primers (100 pM) 1 µL each. A total of 8 µL SuperQ nuclease-free water was added to get the desired volume (25 µL). Lastly, 1.5% agarose (CSL-AG500; CLEAVER SCIENTIFIC®, Rugby, UK) gel electrophoresis was used to observe the PCR products.
2.10. O Typing
Shiga toxin (stx1, stx2) producing E. coli (STEC; N=21) were subjected to O typing through PCR using specific primers for various O types i.e. O26,103,111,121, and O145 [10]. Conventional PCR was performed as described previously [11]. Briefly, total 25- µL PCR mix composed of 4 µL template DNA, 8 µL Green DreamTaq mix (Thermo Fisher Scientific, USA), 1 µL each primer (F&R), SuperQ water was added to get the required volume. PCR Reaction was conducted with following conditions: 30 cycles of denaturation at 94°C for 35 s, annealing at 55°C for 35 s, and extension at 72°C for 1 minute followed by final extension for 10 minutes. Furthermore, agarose (CSL-AG500; CLEAVER SCIENTIFIC®, Rugby, UK) gel (1.5 % with 1 mg/ml ethidium bromide) was prepared for electrophoresis to examine the PCR products.
2.11. Statistical Analysis
The particulars were combined in a Excel (Microsoft Office 365) spreadsheets for various different statistical analysis. The relationship between variables from different sample sources was studied through correlation and linear regression. The strength of existing relationship among variables was quantified. For comparison of means data was subjected to analysis of variance (ANOVA), to find out if the associated source means are significantly different and p-value < 0.05 was set as significant.
3. Results
3.1.1. Distribution of E. coli from Various Food Origin
A total of 154/500 (30.8%) of food samples were tested positive for E. coli, sample wise distribution showed that among several groups, poultry and fisheries acquired significantly higher number of E. coli. The highest prevalence of E. coli was observed in poultry 38.67% followed by Fisheries 32%. Additionally, environmental and dairy samples exhibited 29.6%, while 25.37% E. coli was detected in livestock samples. Further category-wise distribution of samples and E. coli prevalence is given in Table 1.
3.1.2. Distribution of NON-O157 STEC among Various Sample Sources
Overall, out VAGs confirmed 21/52 (40.38%) isolates were observed as STEC. According to distribution, the highest distribution of STEC was 75%, observed in beef samples followed by mutton, cloacal/anal swabs, chicken meat, veal, and poultry droppings i.e., 42.85%, 40%, 27.27% and 25% respectively. Furthermore, among STEC, a total of 12(57.14%) were found to be Non-0157 STEC co harboring blaNDM-1 & mcr-1. Likewise, various sample sources exhibited results as in Non-0157 STEC co harboring blaNDM-1 & mcr-1. (Table 2)
A total of 21(40.38%) of STEC samples were observed for their respective O types, among them 11(52.38%) isolates belong to O26 including 4(36.36%) from beef sample, 2(18.18%) from chicken meat, cloacal /anal swabs and poultry droppings. While 1(33.33%) and 2(50%) of Non-0157 STEC co harboring blaNDM-1 & mcr-1 isolates were detected with O103, O121 and O111, O145 respectively in poultry, livestock category and slaughterhouse samples. (Table 3)
3.1.3. Distribution of Non-O157 STEC Co-Harboring blaNMD-1-1 & mcr-1 among Various Sample Sources
Overall, out of STEC a total of 12/21(57.14%) were distinguished as Non-0157 STEC co harboring blaNDM-1 & mcr-1, where sample wise division was shown the higher number 3 (25%) in chicken meat followed by 2(16.67%) in beef samples, cloacal/anal swabs and poultry droppings, while 1(8.33%) was observed in mutton and slaughterhouse samples, details are given in Table 2.
3.1.4. VAGs Detection
Overall, 52 (67.53%) isolates have shown the presence of range of VAGs (Figure 2b). Inclusively, 11(14.28%) of the isolates carry eight VRGs. Additionally, 8(10.38%) of the isolates carry seven ARGs followed by 7(9.09%) having five VRGs. Moreover, 4(5.19%), 8(10.38%) and 10(12.98%) of the isolates carried four VRGs respectively. Furthermore, within all the isolates 23(29.87%), three marker genes i.e., ompT, hylF and iss, were identified. In addition to these most common VAGs among E. coli was stx-1, stx-2 followed by eae, papC and traT (Figure 3).
- (a)
- Heat map showing the distribution of five major categories of food specimen from various food origin showing in coloured boxes on the right side, along with Antibiotic resistance genes including ESBLs, MBLs, qnrs, sul, tet, mcr (1,11) and fos showing in colored boxes on right size. These genes were grouped as low, intermediate and high frequency.
- (b)
- Heat map showing the placement of virulence genes associated with Fimbrae, mobility, toxin and iron uptake etc. in E. coli isolates from various food groups mainly livestock (mutton, beef, veal), poultry (chicken meat, cloacal/nasal swabs & dropping), environment (slaughter house & transport means), and from dairy (raw milk & cheese) shown as low, intermediate and high on the right side.
- (c)
- Heat map showing Co-existence of NDM and mcr-1 with VRGs based E. coli detection as in this case VRGs detected mainly from livestock (mutton, beef, veal), poultry (chicken meat, cloacal/nasal swabs & dropping) and one from environmental sample (slaughter house) show high, low and intermediate values along with co-occurrence of ARGs (NDM & mcr-1).
Furthermore, in addition to the stx1 and stx2 all Non-O157 STEC co-harboring blaNDM-1 and mcr-1 isolates have shown the presence of range of VAGs including hylA, eae, iss, papC, papA, papG, tsh, ibeA, iuOD, fimH, traT, OmpT, AmpC etc. (Table 4)
3.1.5. Resistance Profiling of the Isolates
Resistance pattern of the Non-O157 STEC co-harboring blaNDM-1 and mcr-1 isolates, showed 100% resistant to ampicillin followed by cefepime. Additionally, chloramphenicol showed 75% tetracycline, 72%, trimethoprim, 65%, ciprofloxacin and levofloxacin 51%, colistin 45%, fosfomycin 40%, imipenem, and meropenem, 10% and least resistance shown in case of tigecycline i.e., 4% respectively. (Figure 4; Table 5). Moreover, all Non-O157 STEC co harboring blaNDM-1 and mcr-1 have shown positive for CarbaNP and rapid polyminx confirmation.
- (a)
- Scatter plot showing the presence of ESBLs genes in various food sources. The high number of blaCTX-M was detected in fisheries via transport means whereas blaSHV is more in beef. Among blaTEM, blaOXA, blaCMY highest prevalence observed in raw milk, poultry waste and slaughter housing sample respectively.
- (b)
- Scatter plot in MBLs showcasing highest prevalence of blaNDM-1 and blaOXA in poultry droppings and lowest in poultry farm waste while blaIMP in fisheries market waste.
- (c)
- Scatter plot describing qnrs, among which qnrS was highest in poultry farm waste followed by qnrA which is more in open market waste. Along with qnrB which is more in raw milk. Between gyrs, gyrA is more prevalent than gyrB and detected more in open market waste.
- (d)
- Scatter plot elaborating sul and tet genes. Here sul1 and sul2 are more prevalent in environmental waste samples while tetA is more in number in dairy waste as compared to tetB that is more in open market waste.
3.1.6. ARGs
In general, a total of 77/154 (50%) isolates have shown the presence of different ARGs (Figure 2a). Among various ESBLs the following detection pattern was observed i.e., CTX-M (52.94%), SHV (66.67%), TEM (83.33%), OXA (77.78%) and CMY (75%). Regarding detection rate of MBLs showed blaNDM (62.5%) followed by blaIMP (71.42%), blaOXA (62.50%), blaKPC (25%) and lowest in blaVMP (11.11%). Regarding Qnr genes, qnrS was observed in 66.67% isolates followed by qnrA (50%) and qnrB (28.57%) (Figure 5). Likewise, detection rate of gyrA was 80% and gyrB 70%, sul180% and sul2 50% respectively. Furthermore, tetB and tetA were observed in 80% and 66.67% respectively.
4. Discussion
E. coli emerges not only as a key protagonist due to its ubiquity in various ecosystems, ability to readily exchange genetic material containing resistance genes but also show how antimicrobial use and resistance development in one sector can swiftly disseminate and manifest in other domains [12]. Therefore, more studies required that depict the pathogenicity and resistance paradigm of E. coil from food origin may play a significant role to improve the health impact caused by such pathogens.
In this regard, the present study focused on the Non-O157 STEC co-harboring blaNDM and mcr-1 in different sources which make food supply chain a complicated unit (Figure 6). Overall, 2.4 % (12/500) of Non-O157 STEC co harboring mcr-1 and blaNDM were found in all studied sources, estimated distribution of Non-O157 STEC in food supply chain is worrying. Though most often STEC O157 are linked with outbreaks but in recent time it is reported that non-O157 STEC strains displayed the capability to cause illness like HUS [13]. Now the research focused on distribution, incidence, and pathogenicity of Non-157 STEC is extensively reported because of the recent outbreaks caused by these serogroups. In a very recent time, a study was reported the AMR of Non-O157 STEC from human source and domesticated animals, results showed significant incidence of Non-O157 STEC co-harboring mcr-1 gene [5] that corroborate the findings of the present investigation. Likewise, current findings correlate with documented work of [14] that showed the detection of STEC from poultry in Nigeria. Similarly, [15] reported the detection of STEC i.e. stx1, stx2 among poultry dropping and cloacal swabs. These results aligned with the findings reported from different low- and middle-income countries (LMICs) and China. [16-18].
Overall, a total of 21/500 (4.2 %) isolates were confirmed as STEC, both stx1 and stx2 were detected in these isolates. Previously, reported data on Non-O157 STEC revealed incidence rate up to from 5 to 10% [19, 20], but due to geographical differences and variation in food categories, processing, transportation and STEC identification protocols, direct comparison of the findings is difficult. Even then, the current findings affirm the previous data and show relative distribution of STEC among studied sources. Furthermore, available literature suggests that among various Non-O157 STEC O types O26 is the most abundant serogroup and has been reported in various studies. Likewise, the current findings displayed that O26 type was the most prevalent (52%) serogroup among studied samples. In recent past, a study reported the presence of O26 type in Non-O157 STEC isolated from retail foods in China [21]. Similarly, detection of Non-O157 STEC in the present investigations validates the previously described particulars from different parts of the world, which stated that FBIs due to non-O157 STEC are exceeding [22].
All STEC isolates displayed array of VAGs with stx1 and stx2. Findings are consistent with former reports showing the distribution of stx1 and stx2 among STEC isolated from different food sources. Epidemiological and clinical investigations suggested that presence of stx2 is essential to cause HUS, and such strains that exhibit additional VAG can cause severe illnesses [23]. Furthermore, findings of the present study agree with published findings that food originated Non-O157 STEC displayed a range of VAGs [24]. In Recent past different studies from Pakistan have also shown the most common VAG among STEC isolated from food animal origin was stx2 (24.81%) gene [25], another study in USA determined the prevalence of stx (16.6%) from STEC retrieved from food animals [26]. These findings are correlate with studies conducted in several European countries (Hungary, Poland, Switzerland, Austria etc) reported similar VAG pattern in various meat samples [27].
The isolates of the study presented substantial resistance against the studied antibiotics. Statistically significant resistance pattern (Figure 7) of the isolates is supported by previous reported findings in reference to the STEC from food samples [28]. Antibiotic resistance especially against quinolones may aid in virulence of STEC by enabling the pathogen to induce stx1 or stx2. Significant resistance among isolates advocates the notion that unofficial and irrational antimicrobial use is a predisposing factor in the emergence and dissemination of AMR and resistant superbugs.
- (a)
- Linear regression/correlation (multiple R = 0.72) showed the co-existence of blaNDM-1 and mcr-1 in VRGs based confirmed E. coli. Here in chicken meat showing the highest percentage in marker VAGs (stx1, stx2, eae, hylA, fimH) along with blaNDM-1 and mcr-1 co harboring E. coli followed by beef, mutton, poultry droppings and cloacal /anal swabs. As same increase pattern was observed in both cases.
- (b)
- Linear regression of Non O157, blaNDM-1 and mcr-1 harboring STEC isolates revealed the correlation among MICs of the isolates from chicken meat and chicken droppings.
- (c)
- Linear regression shows correlation of different selected sources and various Serotypes. In livestock category (beef samples) exhibited high percentage and same sample sources showing the high level of O26 in STEC cohobring blaNDM-1 and mcr-1 followed by chicken meat, cloacal/anal swabs and poultry dropping. Along with other Otypes, O103, O121 were also present along with O111 and O145 among selected food sources showing same rise pattern.
A range of ARGs were detected in the studied isolates, which aligned with work of [16] [29-32]. (Figure 4). Similarly, MBLs were prevalent upto 68/500 (13.8%) which correlate with the work of [33], but here clinical isolates were used for the study. Another study from Nigeria showing the occurrence of specially blaVIM in E. coli from food illness based diarrheal samples [34]. The current research work indicated the multiple number of ARGs, showing significant correlation between antibiotic resistance paradigm of the STEC isolates and dispensation of ARGs (Figure 7). For example, the highest resistance among E. coli was observed against Ampicillin, and blaTEM was recorded in 83% of the isolates. Similarly, 40% of the isolates were found to be resistant to colistin, and blaIMP was observed in 40% [14] % of the resistant strains. These findings are correlate with work of [14], showing high resistance pattern and detection of various ARGs among STEC isolated from food producing animals. In poultry and livestock sector, a common malpractice of not only hasty antibiotics usage as growth booster but also in food production chains and animals mainly fed and discharged in same environment, carry a huge amount of ARGs contaminated the surroundings can cause major hazards in ecosystem health [30]. This practice benefaction exploring the antibiotic-resistant strains, such as mcr-1- and blaNDM harboring E. coli. The current research endorsed the idea, as almost 40% of the STEC isolates exhibited resistance against colistin, which serves as a resort antibiotic for Gram-negative bacilli (GNB). Likewise, same observation was recorded in Malaysia where colistin showed resistance pattern with a 100% detection rate of mcr-1 among E. coli isolated from poultry and poultry meat [35]. These findings are coincide with current observations from Bangladesh, which elaborated more colistin percentage (55.77%) of mcr-1-harboring E. coli among commercial poultry meat [36]. A research from Czech Republic also exhibited 27.7% colistin resistance from meat samples [37]. Similarly, a study from China reported (27.9%) of E. coli co-harboring mcr-1 and blaNDM-1 from river water which serve as reservoir for transmission [38]. Furthermore, in developing countries, reports showing high colistin and blaNDM resistance mainly in poultry, livestock sector where environment serve as the leading cause of transmission and this issue need to address on urgent basis. There should be complete ban on the imprudent antibiotics’ usage as growth booster in poultry and livestock production to fend off the surfacing of the various antibiotic-resistant strains.
AMR and foodborne pathogens monitoring and surveillance in the food supply chain is a multi-faceted and complex phenomenon. Owing to the fact, the present study has some limitations, for instance, geographical distribution of the sample was biased. In future, a comprehensive study with samples from various geographical locations with different socioeconomic standards and food production means may be conducted. Moreover, the present study lacks the detailed genomics of STEC isolates e.g. whole genome sequencing, sequence typing, functional genomics etc. Such studies may be planned to illustrate the molecular insights of STEC resistance and pathogenesis.
5. Conclusions
In conclusion, the study contributes valuable insights into the molecular characteristics of Non-O157 STEC from various food sources. These findings have given us proof that abrupt antibiotic usage in poultry, livestock farms not only effect the food supply chain but also play as major role in transfer of various resistant superbugs such as STEC harboring mcr-1 and blaNDM-1 to the community through contaminated environment. The widespread presence of such resistant E. coli strains among food supply chain is worrisome that needs immediate action to bring safe and healthy food from farm to fork.
6. Patents
Not applicable
Author Contributions
AS: PhD scholar performed all the research work, manuscript writing. BA, MHR Study layout, supervision, write-up. MS & JS technical check and manuscript revision.
Funding
Funds for the study: Project Contract No.25099 in Coordinated Research Project No. D52044 from International Atomic Energy Agency (IAEA), Vienna Austria.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSPD2024R986), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
Authors Declared no conflict of interest.
References
- Gompel, L.V.; Luiken, R.E.C; Hansen, R. B.; Munk, P.; Bouwknegt, M.; Heres, L.; Greve, G.D.; Scherpenisse, P.; Jongerius-Gortemaker, B.G.M.; Tersteeg-Zijderveld, M.H.G.; GarciaCobos, S.; Dohmen, W.; Dorado-Gracienaar, J.A.; Urlings, B.A.P.; Wagenaar, J.P.; Heederik, D.J.J.; Schmitt, H.; Bossers, A.; Smit, L.A. M.; et al.Description and determinants of the faecal resistome and microbiome of farmers and slaughterhouse workers: a metagenome-wide cross-sectional study. Environ.Int. 2020, 143, 105939. [Google Scholar] [CrossRef]
- Pokharel, S; Priyanka, S. ; Bipin, A. Antimicrobial use in food animals and human health: time to implement ‘One Health’approach. Antimicrob Resist Infect Control. 2020, 9, 181. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, J.; Zhang, R.; Chen, L.; Zhang, H.; Qi, X.; Chen, J.; et al. Epidemiology of foodborne diseases caused by Salmonella in Zhejiang Province, China, between 2010 and 2021. Public Health Front. 2023, 11, 1127925. [Google Scholar]
- Gallo, M.; Ferrara, L.; Calogero, A.; Montesano, D.; Naviiglio, D.; et al. Relationships between food and diseases: What to know to ensure food safety. Food Res.Int. 2020, 137, 109414. [Google Scholar] [CrossRef]
- Pan, Y.; Hu, B.; Bai, X.; Yang, X.; Cao, L.; Liu, Q.; Sun, H.; Li, J.; Zhang, J.; Jin, D.; Xiong, Y.; et al. Antimicrobial Resistance of Non-O157 Shiga Toxin Producing Escherichia coli Isolated from Humans and Domestic Animals. Antibiotics. 2021, 10, 1. [Google Scholar] [CrossRef]
- Joseph, A.; Cointe, A.; kurkdijan, P. M.; Rafat, C.; Hertig, A.; et al. Shiga Toxin-Associated Hemolytic Uremic Syndrome: A Narrative Review. Toxins. 2020, 12, 2. [Google Scholar] [CrossRef]
- Tiseo, K.; Huber, L.; Gilbert, M.; Robinson, T.P.; Van Boeckel, T. P.; et al. Global trends in antimicrobial use in food animals from 2017 to 2030. Antibiot. 2020, 9, 918. [Google Scholar] [CrossRef]
- Nawaz, Z.; Aslam, B.; Zahoor, M.A.; Siddique, A.B.; Rafique, A.; Aslam, R.; Qamar, M.U.; Ali, S.; Mubeen, M.; et al. Frequency of Extended Spectrum Beta Lactamase Producing Escherichia coli in Fresh and Frozen Meat. Pak. Vet. J. 2020, 41, 1. [Google Scholar]
- Usman, M.; Rasool, M.H.; Khurshid, M.; Aslam, B.; Baloch, Z.; et al. Co-Occurrence of mcr-1 and Carbapenem Resistance in Avian Pathogenic E. coli Serogroups O78 ST95 from Colibacillosis-Infected Broiler Chickens. Antibiotics. 2023, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Bording-Jorgensen, M.; Parsons, B.; Szelewicki, J.; Lioyd, C.; Chui, L.; et al. Molecular Detection of Non-O157 Shiga Toxin Producing Escherichia coli (STEC) Directly from Stool Using Multiplex qPCR Assays. Microorganism. 2022, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Iguchi, A.; Iyoda, S.; Seto, K.; Taguchi, M. ; Kumeda, Y.; et al. Multiplex Real-Time PCR Assays for Screening of Shiga Toxin 1 and 2 Genes, Including All Known Subtypes, and Escherichia coli O26-, O111-, and O157-Specific Genes in Beef and Sprout Enrichment Cultures. JFP. 2015, 78, 10.
- Silva, V.; Correia, S.; Pereira, J.E.; Igrejas, G.; Poeta, P.; et al. Surveillance and environmental risk assessment of antibiotics and amr/args related with mrsa: one health perspective. AMR. 2020, 271–295. [Google Scholar]
- Kinnula, S.; hemminki, K.; Kotilainen, H.; Ruotsalainen, E.; Tarkka, E.; Salmenlinna, S.; Hallanvuo, S.; Leinonen, E.; Jukka, O.; Rimhanen-Finne, R.; et al. Outbreak of multiple strains of non-O157 Shiga toxin-producing and enteropathogenic Escherichia coli associated with rocket salad, Finland, autumn 2016. Euro. Surveill. 2018, 23, 35. [Google Scholar] [CrossRef] [PubMed]
- Al-Mustapha, A. I.; Raufu, I.A.; Ogundijo, Q.A.; Brouwer, M.S.M.; Adetunji, V. ; Heikinheimo, A.; et al. Antibiotic resistance genes, mobile elements, virulence genes, and phages in cultivated ESBL-producing Escherichia coli of poultry origin in Kwara State, North Central Nigeria. Int. J. Food. Microbiol, 2023, 389, 110086. [Google Scholar]
- Hasona, I.F.; helmy, S.M.; and El Gamal, A. D.; et al.Prevalence, virulence factors, and antimicrobial resistance profiles of Shiga toxin-producing Escherichia coli isolated from broiler chickens in Egypt. Vet. Res. Forum. 2023, 14, 131–138. [Google Scholar] [PubMed]
- Anyanwu, M.U.; Jaj, F.I.; R. Okpala, C.O.; Njoga, E.O.; Okafor, N.A.; Oguttu, J.W.; et al. Mobile colistin resistance (mcr) gene-containing organisms in poultry sector in low-and middle-income countries: Epidemiology, characteristics, and one health control strategies. Antibiotics. 2023, 12, 1117.
- Sadek, J.; Rosa. J.M.O.; Maky, M. A.; Dandrawy, M.K.; Nordman, P.; Poirel, L.; et al. Genomic Features of MCR-1 and Extended-Spectrum β-Lactamase Producing Enterobacterales from Retail Raw Chicken in Egypt Mustafa. Microorganisms. 2021, 9, 195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, L.; Wang, J.; Yassin, A.K.; Butaye, P.; Kelly, P.; Gong, J.; Guo, W.; Li, J.; Li, M.; Yang, F.; Feng, Z.; Jiang, P.; Song, C.; Wang, Y.; You. J.; Price, S.; Qi, K.; Kang, Y.; Wang, C.; et al. Molecular detection of colistin resistance genes (mcr-1, mcr-2 and mcr-3) in nasal/oropharyngeal and anal/cloacal swabs from pigs and poultry. Sci. Rep. 2018, 8, 3705. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Mondol, A.S.; Azmi, I.J.; Boer, E.D.; beumer, R.R.; Zwietering, M.H.; Heuvelink, A.E.; Talukder, K. A.; et al. Occurrence and Characterization of Shiga Toxin–Producing Escherichia coli in Raw Meat, Raw Milk, and Street Vended Juices in Bangladesh. Foodborne Pathog. Dis. 2010, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Ju, W.; Shen, J.; Li, Y.; Toro, M.A.; Zahoo, S.; Ayers, S.; Najjar, M.B.; Meng, J.; et al. Non-O157 Shiga toxin-producing Escherichia coli in retail ground beef and pork in the Washington D.C. area. Food Microbiol. 2012, 32, 2. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Sun, H.; Fan, R.; Fu, S.; Zhang, J.; Matussek, A.; Xiong, Y.; Bai, X.; et al. Genetic diversity of the intimin gene (eae) in non-O157 Shiga toxin-producing Escherichia coli strains in China. Sci. Rep. 2020, 10, 3275. [Google Scholar] [CrossRef] [PubMed]
- Lee, JB.et al. , (2018)." Pathogenic and phylogenetic characteristics of non-O157 Shiga toxin-producing Escherichia coli isolates from retail meats in South Korea. J. Vet. Sci. 2017, 19, 2. [Google Scholar]
- Byrne, L.; Adams, N.; Jenkins, C.; et al. Association between Shiga Toxin–Producing Escherichia coli O157:H7 stx Gene Subtype and Disease Severity, England, 2009–2019. Emerge. Infect. Dis. 2020, 26, 10. [Google Scholar] [CrossRef]
- Li, Y.; Huang, T.Y.; Ye, C.; Chen, L.; Liang, Y.; Wang, K. ; Liu, J.; et al. Formation and Control of the Viable but Non-culturable State of Foodborne Pathogen Escherichia coli O157:H7. Front. Microbiol. 2020, 11, 1202. [Google Scholar]
- Shah, I. H. Molecular Characterization and Virulence Gene Profile of Entero-pathogenic and Shiga-Toxin Producing Escherichia coli from Food of Animal Origin and Environmental Sources. SKUAST Kashmir. 2023.
- Alotaibi, K.; and Khan, A.A. Prevalence and Molecular Characterization of Shiga Toxin Producing Escherichia coli from Food and Clinical Samples. Pathogens. 2023, 12, 1302. [Google Scholar] [CrossRef] [PubMed]
- Nüesch-Inderbinen, M.; Treier, A.; Stevens, M.J.A.; Stephen, R.; et al. Whole genome sequence-based characterisation of Shiga toxin-producing Escherichia coli isolated from game meat originating from several European countries. Sci. Rep. 2023, 13, 3247. [Google Scholar] [CrossRef]
- Shakerian, A.; rahimi, E. ; Emad, P.; et al. Vegetables and Restaurant Salads as a Reservoir for Shiga Toxigenic Escherichia coli: Distribution of Virulence Factors, O-Serogroups, and Antibiotic Resistance Properties. JFP. 2016, 79, 7.
- Sakthikarthikeyan, S.; Sivakumar, M.; Manikandan, R.; Senthkumar, P.; Sureskumar, V.; Malmarugan, S.; Prabhu, M.; Ramakrishnan, V.; et al. Prevalence and Molecular Characterization of Multidrug-resistant ESBL-producing E. coli in Commercial Poultry. Indian J. Anim. Res. 2024, 1, 6. [Google Scholar] [CrossRef]
- Tiedje, J. M.; Fu, Y.; Mei, Z.; Schaffer, A.; Dou, Q.; Amelung, W.; Elsner, M.; Adu-Gyamfi, J.; Heng, L.; Virta, M.; Jiang, X.; Smidt, H.; Topp, E.; Wang, F.; et al. Antibiotic resistance genes in food production systems support One Health opinions. Curr. Opin. Environ. Sci. Health. 2023, 43, 100492. [Google Scholar] [CrossRef]
- Hinthong, W.; Thaotumpitak. ; Sripradite, J.; Indrrawttana, N.; Srisook, T.; Kongngoen, T.; Atwill, E.R.; Jeamsripong, S.; et al. Antimicrobial resistance, virulence profile, and genetic analysis of ESBL-producing Escherichia coli isolated from Nile tilapia in fresh markets and supermarkets in Thailand. Plos one. 2024, 19, e0296857. [Google Scholar]
- Zhang, S.; Huang, Y.; Yang, G.; Wu, Q.; Zhang, J.; Wang, J.; Ding, Y.; Ye, Q.; Wu, S.; Gu, Q.; Zhang, Y.; et al. High prevalence of multidrug-resistant Escherichia coli in retail aquatic products in China and the first report of mcr-1-positive extended-spectrum β-lactamase-producing E. coli ST2705 and ST10 in fish. Int. J. Food Microbiol. 2024, 408, 110449. [Google Scholar] [CrossRef]
- Ilham, D.; Souad, L.; Asmae, L.H.; kawtar, N.; Mohammed, T. ; Nabila, S.; et al. Prevalence, antibiotic resistance profile, MBLs encoding genes, and biofilm formation among clinical carbapenem-resistant Enterobacterales isolated from patients in Mohammed VI University Hospital Centre, Morocco. Lett Appl Microbiol. 2023, 76, ovad107.
- Nwike, I. E.; Ugwu, M.C.; Ejikeugwu, P.C.; Ujam, N.T.; Iroha, I.R.; Esimone, C. O.; et al. Phenotypic and molecular characterization of enteropathogenic Escherichia coli and Salmonella spp. causing childhood diarrhoea in Awka, South-Eastern Nigeria. BNRC. 2023, 47, 97. [Google Scholar]
- Karim, M. R.; Zakaria, Z.; Hassan, L.; Faiz, N.K.; Ahmad, N.I.; et al. Antimicrobial Resistance Profiles and Co-Existence of Multiple Antimicrobial Resistance Genes in mcr-Harbouring Colistin-Resistant Enterobacteriaceae Isolates Recovered from Poultry and Poultry Meats in Malaysia. Antibiotics. 2023, 12, 6–1060. [Google Scholar] [CrossRef]
- Ali, M.W.; Utsho, K.S.; Karmakar, S.; Nazmul Hoque, Md.; Rahman, Md.T.; Hassan, J.; et al. et First report on the molecular characteristics of mcr-1 colistin resistant E. coli isolated from retail broiler meat in Bangladesh. Int. J. Food Microbiol. 2023, 388, 110065. [Google Scholar] [CrossRef] [PubMed]
- Sismova, P.; Sukar, I.; Kolidentsev, N.; Chytilova, I.; Bardon, J.; Dolejska, M.; Nesporova, K.; et al. Plasmid-mediated colistin resistance from fresh meat and slaughtered animals in the Czech Republic: nation-wide surveillance 2020–2021. Microbiol. spectr. 2023, 11, e00609–00623. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.H.; Liu, Y.Q.; Duan, X.X.; Yang, T.Y.; Li, F.Y.; Zou, M.; Liu, B. T.; et al. et al. High prevalence and genomic characteristics of carbapenem-resistant Enterobacteriaceae and colistin-resistant Enterobacteriaceae from large-scale rivers in China. Environ Pollut. 2023, 331, 121869. [Google Scholar]
Figure 1.
Distribution of E. coli in various food origin which depicted by colorful parts of pie chart showed the percentage of E. coli in five major categories which then further subdivided according to sample source. The colorful parts of pie chart also showed the percentage from different categories.
Figure 1.
Distribution of E. coli in various food origin which depicted by colorful parts of pie chart showed the percentage of E. coli in five major categories which then further subdivided according to sample source. The colorful parts of pie chart also showed the percentage from different categories.
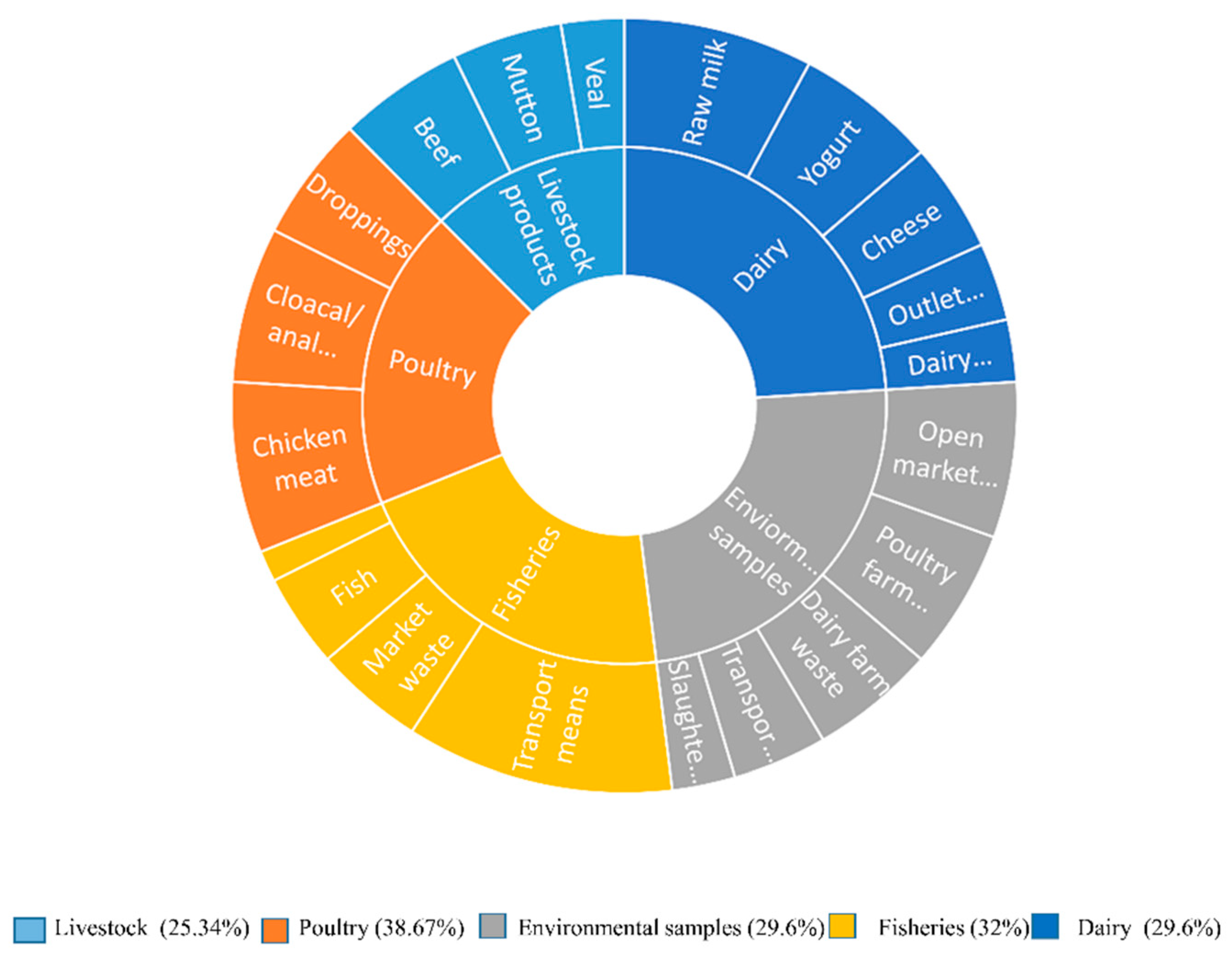
Figure 2.
Heat map showing distribution of antibiotic resistance gene(ARGs), virulence resistance genes (VRGs) and co-existence of NDM & mcr-1 associated with VRGs based E. coli detection from various sources.
Figure 2.
Heat map showing distribution of antibiotic resistance gene(ARGs), virulence resistance genes (VRGs) and co-existence of NDM & mcr-1 associated with VRGs based E. coli detection from various sources.

Figure 3.
Showing the circular cluster heat map illustrated the Resistance pattern of various standard antibiotics against VRGs based confirmed E. coli isolates harboring NDM & mcr-1. The resistance pattern among E. coli isolates harboring NDM and mcr-1 represented by color and cluster showing overall resistance percentages against antibiotics (AMP, CMP, COL, LVX, MEM, IMI, CIP, TRI, TOC, FOS).
Figure 3.
Showing the circular cluster heat map illustrated the Resistance pattern of various standard antibiotics against VRGs based confirmed E. coli isolates harboring NDM & mcr-1. The resistance pattern among E. coli isolates harboring NDM and mcr-1 represented by color and cluster showing overall resistance percentages against antibiotics (AMP, CMP, COL, LVX, MEM, IMI, CIP, TRI, TOC, FOS).
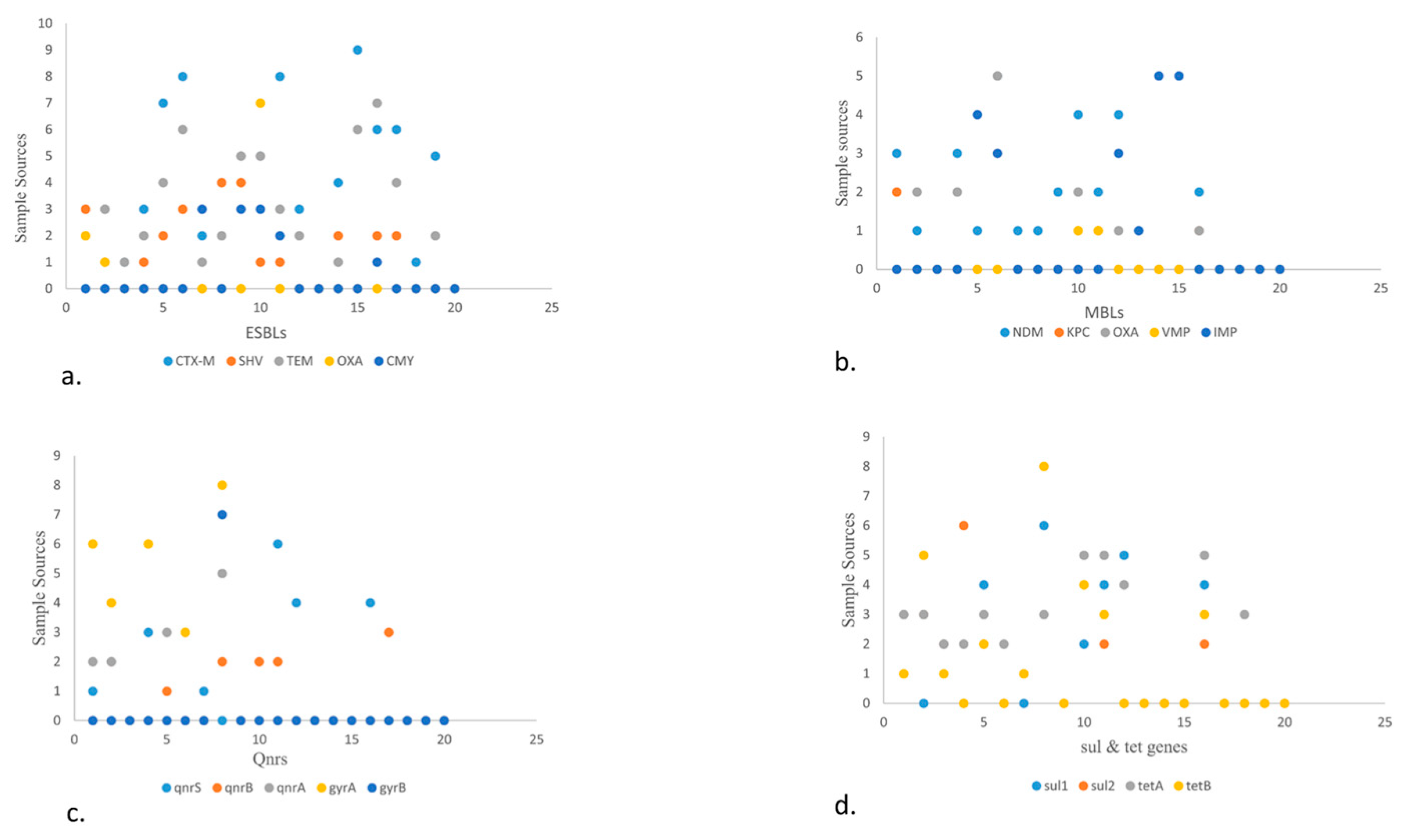
Figure 4.
Scatter plot describing the sample sources against different VRGs from different food origin. Scatter plot showing the sample sources and ARGs in the form of numerical value. The Y-axis showing the sample source like (0-9) depicts (beef, mutton, veal, chicken meat, cloacal/anal swabs, dropping, environmental samples (slaughter house, open market & transport waste, dairy & poultry waste), fish, shrimps, market waste, transport means, and dairy (raw milk, yoghurt, dairy cream, cheese, outlet waste) and x-axis showing ARGs (ESBLs, MBLs, qnrs, tet, sul).
Figure 4.
Scatter plot describing the sample sources against different VRGs from different food origin. Scatter plot showing the sample sources and ARGs in the form of numerical value. The Y-axis showing the sample source like (0-9) depicts (beef, mutton, veal, chicken meat, cloacal/anal swabs, dropping, environmental samples (slaughter house, open market & transport waste, dairy & poultry waste), fish, shrimps, market waste, transport means, and dairy (raw milk, yoghurt, dairy cream, cheese, outlet waste) and x-axis showing ARGs (ESBLs, MBLs, qnrs, tet, sul).
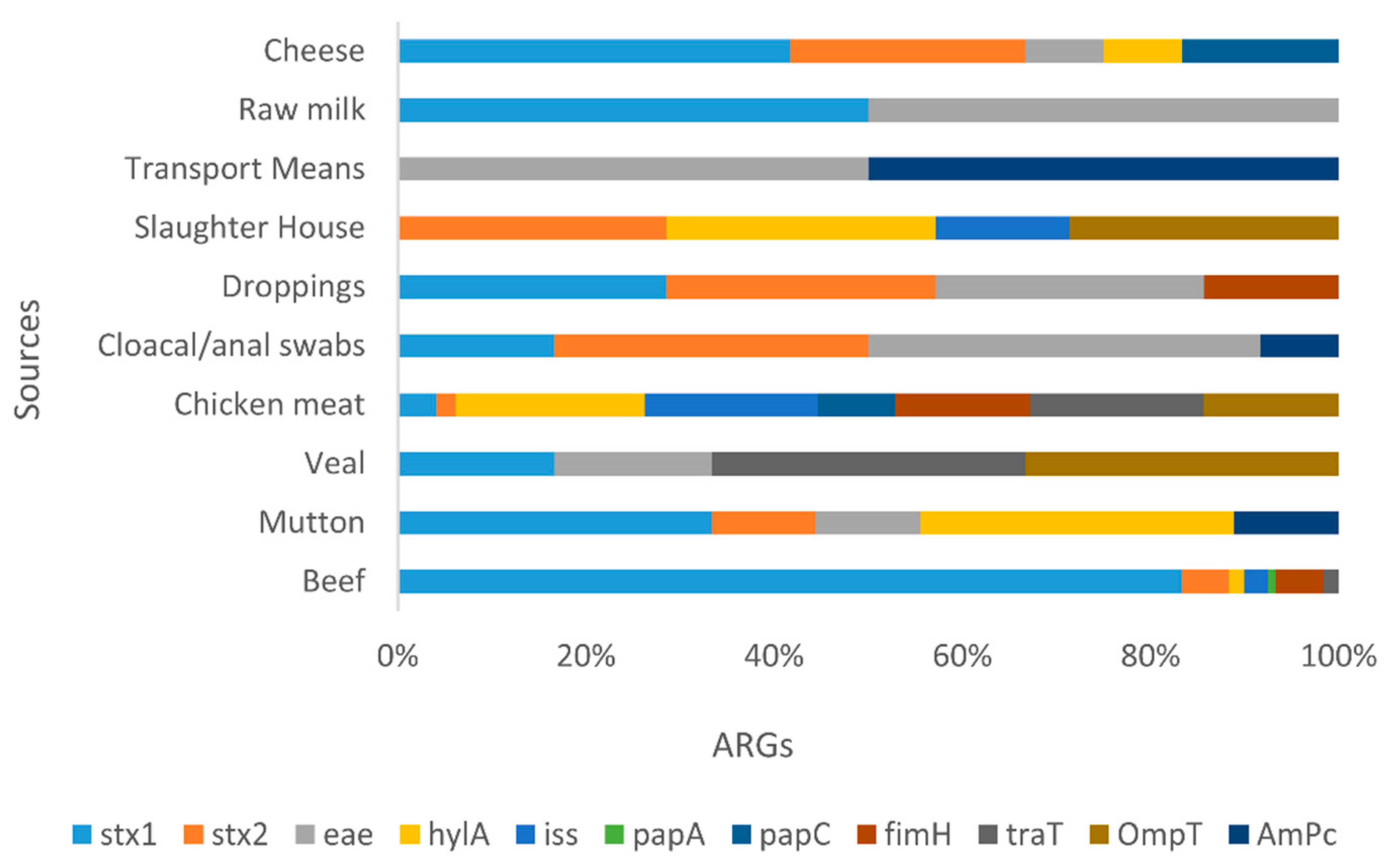
Figure 5.
Stacked bar showcasing prevalence of various VRGs in E. coli isolates from various food sources. This stalked bar describing the detection of various VRGs (stx1, stx2, eae, HylA, iss, pap A. papC, papG, fimH, traT, OmpT and AmpC) shown in various colors against selected food sources from livestock. Poultry, Dairy and environmental sample categories. The highest prevalence in case of stx1 was found in cheese, followed by stx2 more in slaughter house samples. The chicken meat carried more HylA, iss, traT, OmpT and papC whereas papA and fimH was more in beef. Followed by AmpC which was high in various transport means from environment samples.
Figure 5.
Stacked bar showcasing prevalence of various VRGs in E. coli isolates from various food sources. This stalked bar describing the detection of various VRGs (stx1, stx2, eae, HylA, iss, pap A. papC, papG, fimH, traT, OmpT and AmpC) shown in various colors against selected food sources from livestock. Poultry, Dairy and environmental sample categories. The highest prevalence in case of stx1 was found in cheese, followed by stx2 more in slaughter house samples. The chicken meat carried more HylA, iss, traT, OmpT and papC whereas papA and fimH was more in beef. Followed by AmpC which was high in various transport means from environment samples.
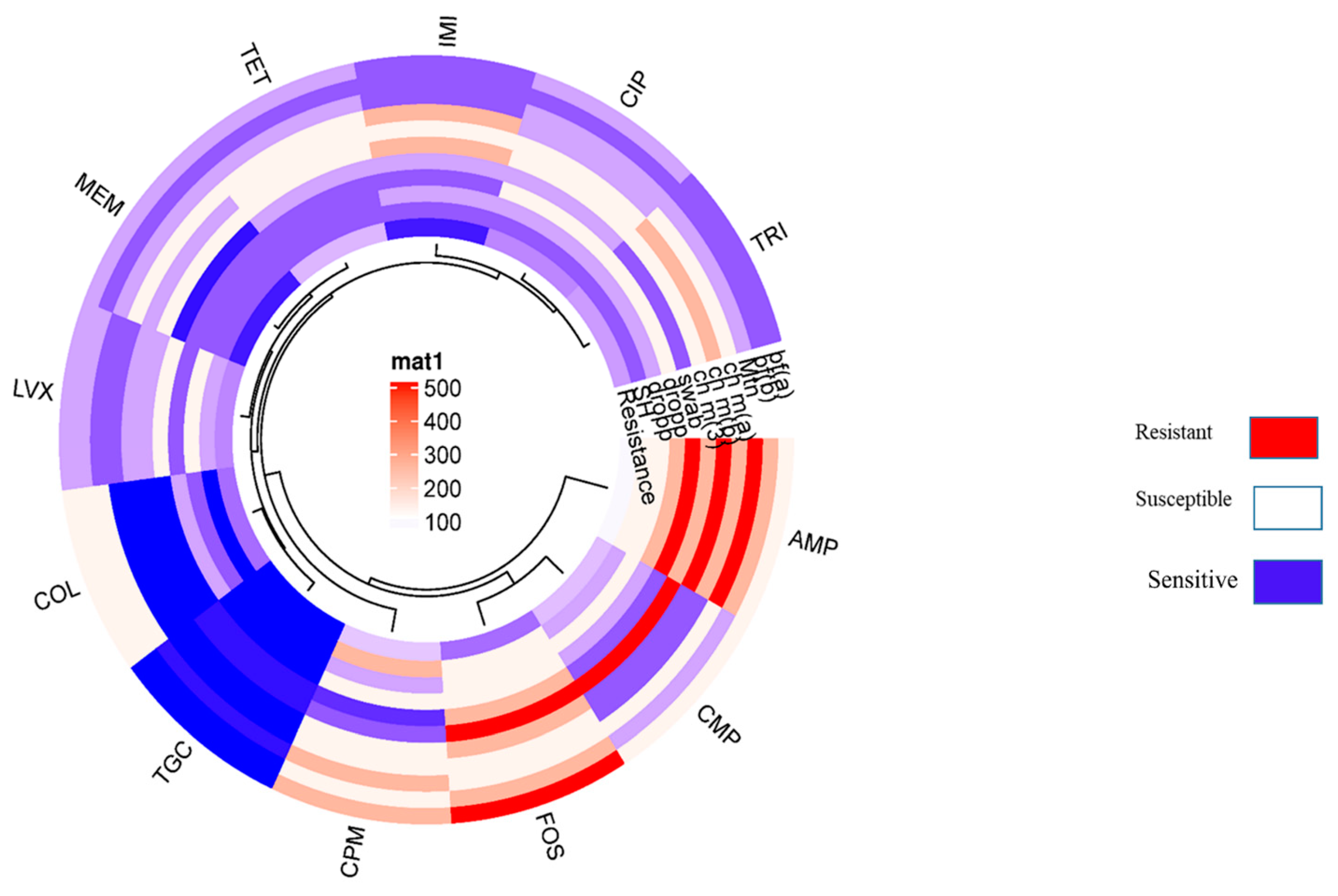
Figure 6.
Chord diagram (https://www.everviz.com/) represented flow between various studied samples sources i.e. nodes. Each source is displayed with an alphabet on outer circle and arcs between different sources showing the connection and relevance with adjacent source.
Figure 6.
Chord diagram (https://www.everviz.com/) represented flow between various studied samples sources i.e. nodes. Each source is displayed with an alphabet on outer circle and arcs between different sources showing the connection and relevance with adjacent source.

Figure 7.
Linear regression analysis from different food sources.
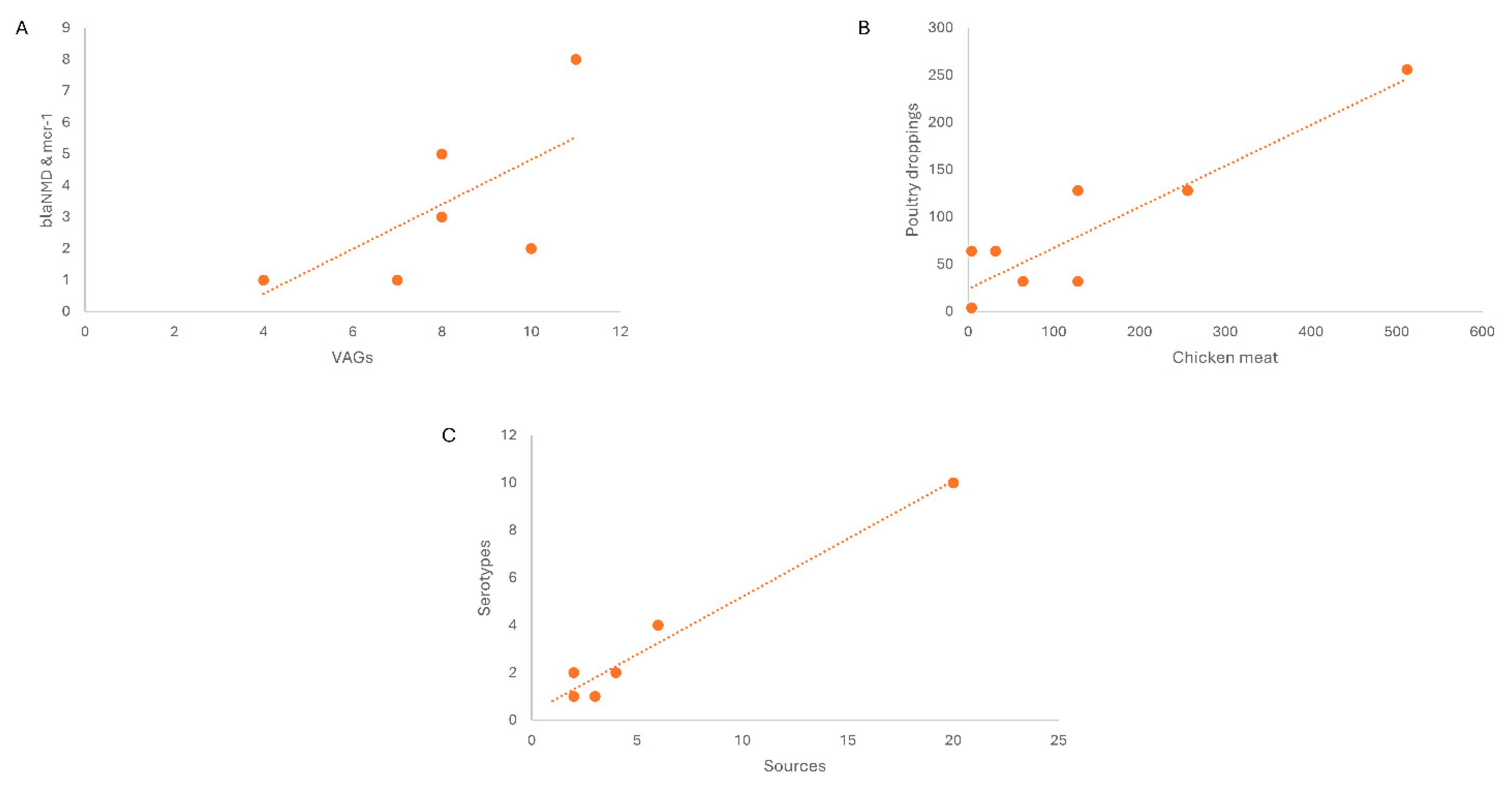

Table 1.
Distribution of E. coli from various food origin.
| Specimen Category | Sources | Collected Samples |
Total Samples |
Positive Sample |
Distribution of E. coli positive Samples | Distribution of E. coli from selected categories |
P-value | ||
|---|---|---|---|---|---|---|---|---|---|
| Livestock products | Beef | 25 | 75 | 8 | 32% | 19/75 (25.34%) |
|||
| Mutton | 25 | 7 | 28% | .001* | |||||
| Veal | 25 | 4 | 16% | ||||||
| Poultry | Chicken meat | 25 | 75 | 11 | 44% | 29/75 (38.67%) |
|||
| Cloacal/anal swabs | 25 | 10 | 40% | ||||||
| Droppings | 25 | 8 | 32% | ||||||
| Environmental samples | Slaughterhouse | 25 | 125 | 4 | 16% | 37/125 (29.6%) |
|||
| Open market waste | 25 | 10 | 40% | ||||||
| Transport Means | 25 | 6 | 24% | ||||||
| Dairy farm waste | 25 | 8 | 32% | ||||||
| Poultry farm waste | 25 | 9 | 36% | ||||||
| Fisheries | Fish | 25 | 100 | 6 | 24% | 32/100 (32%) |
|||
| Shrimps | 25 | 2 | 8% | ||||||
| Market waste | 25 | 7 | 28% | ||||||
| Transport means | 25 | 17 | 68% | ||||||
| Dairy | Raw milk | 25 | 125 | 12 | 48% | 37/125 (29.6%) |
|||
| Yogurt | 25 | 9 | 36% | ||||||
| Dairy cream | 25 | 4 | 16% | ||||||
| Cheese | 25 | 7 | 28% | ||||||
| Outlet waste | 25 | 5 | 20% | ||||||
| Grand Total | 500 | 154 | 154/500 (30.8%) |
||||||
*Highly significant results among isolates from various sample sources. Summary: Multiple R = 0.87; R Square = 0.76; Adjusted R Square = 0.68; SE = 4.15.
Table 2.
Non O157 STEC co harboring blaNDM-1 & mcr-1 among various sample sources along with detected O types.
Table 2.
Non O157 STEC co harboring blaNDM-1 & mcr-1 among various sample sources along with detected O types.
| Specimen category | VAGs based confirmed E. coli |
STEC % out of E. coli | Non O157 STEC co harboring blaNDM-1 & mcr-1 | O Type | Statistical analysis |
|---|---|---|---|---|---|
| Beef | 8(32%) | 6/8(75%) | 2/6(33.33%) | O26 , O103 & O121 | 0.05* |
| Mutton | 7(28%) | 3/7(42.85%) | 1/3(33.33%) | O26, O103 & O111 | |
| Veal | 4(16%) | 1/4(25%) | 1/1(100%) | O121 | |
| Chicken meat | 11(44%) | 3/11(27.27%) | 3/3(100%) | O26 & O145 |
|
| Cloacal/anal swabs | 10(40%) | 4/10(40%) | 2/4(50%) | O26, O111 & O145 |
|
| Droppings | 8(32%) | 2/8(25%) | 2/2(100%) | O26 |
|
| Slaughterhouse | 4(16%) | 2/4(50%) | 1/1(100%) | O103& O12 |
|
| Overall | 52(29.71%) | 21/52(40.38%) | 12/21(57.14%) |
*Statistical significance among Non-O157 O types.
Table 3.
Non-0157 O types among STEC isolated from various sources along with co-existence of ARGs. *Results are statistically significant in reference to STEC from beef and poultry droppings with O26
Table 3.
Non-0157 O types among STEC isolated from various sources along with co-existence of ARGs. *Results are statistically significant in reference to STEC from beef and poultry droppings with O26
| Sources | STEC | Non-O157 O types | Co-existence of ARGs | P value | ||||
| O26 | O103 | O111 | O121 | O145 |
0.05* |
|||
| Beef | 6 | 4 | 1 | -- | 1 | -- | blaNDM,mcr-1, blaTEM,blaOXA, blaKPC, blaqnrA, | |
| Mutton | 3 | 1 | 1 | 1 | -- | blaNDM,mcr-1, blaSHV, blaOXA, | ||
| Veal | 1 | -- | -- | -- | 1 | -- | blaNDM,mcr-1, blaCTX-M, blaTEM, blatetB | |
| Chicken meat | 3 | 2 | -- | -- | -- | 1 | blaNDM,mcr-1, blaCTX-M,blaqnrS, | |
| Cloacal/anal swabs | 4 | 2 | -- | 1 | -- | 1 | blaNDM,mcr-1, blaqnrB, | |
| Droppings | 2 | 2 | -- | -- | -- | -- | blaNDM,mcr-1, blatetA | |
| Slaughterhouse |
2 |
-- | 1 | -- | 1 | -- | blaNDM,mcr-1, blaTEM,blaqnrS,blatetA,blaSul2,blatetB | |
| Total | 21 | 11 | 3 | 2 | 3 | 2 | ||
Table 4.
Co-existence of blaNDM-1 and mcr-1 in VAGs based confirmed E. coli .
| Sample Source |
Positive VAGs in E. coli |
NDM | mcr-1 | Stx | Pap | P value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| stx1 | stx2 | Eae | hylA | Iss | papA | papC | fimH | TraT | OmpT | AmpC | |||||
| Beef | 8 | 3 37.5% |
2 25% |
1 12.5% |
6 75% |
ND | 2 25% |
3 37.50% |
1 12.5% |
ND |
6 75% |
2 25% |
ND | ND |
.001* |
| Mutton | 7 |
1 14.28% |
1 14.28% |
3 42.85% |
1 14.28% |
1 14.28% |
3 42.85% |
ND | ND | ND | ND | ND | ND | 1 14.28% |
|
| Veal | 4 | ND | 1 25% |
1 25% |
ND | 1 25% |
ND | ND | ND | ND | ND | 2 50% |
2 50% |
ND | |
| Chicken Meat |
11 | 3 27.27% |
8 72.72% |
2 18.18% |
1 9.09% |
ND | 8 88.89% |
9 81.81% |
ND | 4 36.36% |
7 63.63.% |
9 81.81% |
7 63.63% |
ND | |
| Cloacal /anal Swabs |
10 | 1 10% |
2 20% |
2 20% |
4 40% |
5 50% |
ND | ND | ND | ND | ND | ND | ND | 1 10% |
|
|
Droppings |
8 | 5 62.5% |
2 25% |
2 25% |
2 25% |
2 25% |
ND | ND | ND | ND | 1 12.5% |
ND | ND | ND | |
| Slaughter House |
4 | 1 25% |
1 25% |
ND | 2 50% |
ND | 2 50% |
1 25% |
ND | ND | ND | ND | 2 50% |
ND | |
| Total |
52 |
14 | 17 | 11 | 16 | 9 | 15 | 13 | 1 | 4 | 14 | 13 | 11 | 2 | |
*Highly significant results among VAGs in isolates from various sample sources
Table 5.
Antibiotic susceptibility testing of Non O157, blaNDM-1 and mcr-1 harboring STEC Isolates.
| Antibiotics | Conc. |
CLSI EUCAST/FDA Resistance Breakpoint |
Beef samples (n=2) |
Mutton sample (n=1) |
Veal sample (n=1) |
Chicken meat (n=3) |
Poultry Cloacal /anal swabs (n=2) |
Poultry droppings (n=2) |
Slaughter house (n=1) |
|||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ampicillin | 10 µg | ≥32 | 128 | 256 |
512 |
256 | 256 | 512 | 256 | 512 | 512 | 256 | 128 | 128 |
| Cefepime | 30µg | ≥16 | 256 | 128 |
256 |
256 | 128 | 128 | 32 | 32 | 16 | 128 | 64 | 256 |
| Ciprofloxacin | 5µg | ≥4 | 64 | 32 |
64 |
128 | 64 | 128 | 128 | 64 | 64 | 128 | 64 | 32 |
| Levofloxacin | 5µg | ≥4 | 64 | 64 |
32 |
32 | 32 | 64 | 64 | 64 | 128 | 32 | 128 | 64 |
| Chloramphenicol | 30µg | ≥32 | 128 | 64 |
128 |
64 | 32 |
32 |
512 | 32 | 32 | 64 | 128 | 64 |
| Trimethoprim | 5µg | ≥16 | 32 | 32 |
64 |
32 | 128 |
256 |
128 | 64 | 32 | 128 | 64 | 32 |
| Imipenem | 10µg | ≥4 | 32 | 32 |
32 |
32 | 256 | 128 |
256 |
128 |
64 | 32 | 64 | 32 |
| Meropenem | 10µg | ≥4 | 64 | 32 |
64 |
64 | 128 | 64 | 128 | 8 | 8 | 32 | 32 | 32 |
| Colistin | 10µg | ≥8 | 128 | 128 |
128 |
32 | 4 | 4 | 4 | 4 | 4 | 64 | 32 | 4 |
| Tetracycline | 30µg | ≥16 | 64 | 32 |
64 |
64 | 128 | 128 | 128 | 32 | 64 | 32 | 32 | 32 |
| Tigecycline | 15µg | ≥8 | 4 | 4 |
8 |
8 | 4 | 4 | 8 | 4 | 8 | 4 | 4 | 4 |
| Fisfomycin | 200 μg | ≥64 | 512 | 256 |
128 |
128 | 128 | 256 | 512 | 128 | 256 | 128 | 128 | 128 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



