
Submitted:
29 April 2024
Posted:
29 April 2024
You are already at the latest version
Abstract
The rapid spread of non-indigenous species (NIS) poses a significant threat to biodiversity globally, with the Mediterranean region being particularly susceptible due to increased human activities and its status as a marine biodiversity hotspot. In this study, we focus on the introduction and distribution of Fistularia petimba, a member of the Fistulariidae family, in the eastern Mediterranean Sea, specifically on the coasts of Amorgos Island, Greece. Through a baseline fisheries study conducted over 12 months, utilizing experimental sampling with gillnets, trammel nets, and longlines, one individual of F.petimba was captured off the coast of Katapola Bay. Morphological examination confirmed its identity, with measurements on meristic characteristics obtained and stomach content analyzed. This finding represents a significant addition to the documented distribution of F. petimba in the Mediterranean Sea, particularly in the Aegean Sea, underscoring the importance of ongoing research in uncovering new occurrences and expanding our understanding of marine biodiversity and ecosystem changes. Further investigation into the ecological preferences and population dynamics of F.petimba in the Aegean Sea is crucial for informed conservation and management efforts.
Keywords:
Fistularia petimba
; Red cornetfish
; Mediterranean Sea
; Invasive species
; NIS
; Lessepsian species
1. Introduction
One of the main threats that biodiversity is currently facing is the rapid spread of non-indigenous species (NIS). NIS are defined as the array of species spreading outside their natural or native distribution range [1]. Different areas worldwide have been experiencing vast impacts from the introduction of non-indigenous species, oftenly related to increased human activities such as, opening of canals, the continuous growth of the shipping industry across biogeographic barriers [2,3], a wide range of changes in water temperature due to climate change [4,5,6], fishing pressure [7,8,9] and habitat degradation or loss of species [10,11,12]. In the studied area of the Mediterranean Sea, recognized as one of the main hotspots of marine biodiversity [13,14], the effects of NIS are apparent both in terms of introduction rate [15] and number of introduced species [16], leading to the global acknowledgment of the Mediterranean region as hotspot area for NIS [17]. Additionally, recent findings during the past decades, points Mediterranean ports as a major hotspot for the introduction of NIS in this basin [18,19].
A region where visible changes in aquatic biodiversity have occurred is the eastern Mediterranean Sea where a rapid introduction of fish species with Indo-pacific origin are observed, i.e., the Levantine Sea [20,21,22,23], raising significantly the overall amount of fish biomass up to 90% in specific habitats [24,25]. Though, Indo-Pacific fish species could potentially arrive through various ways in the Levantine basin, most likely by immigration path through the Suez Canal which opened to shorten the commercial shipping ways between the Indian Ocean and the Mediterranean Sea in 1869 [26]. It is assumed that species that normally resided in the Red Sea and the Indian Ocean traversed through the Suez Canal, and proceeded northwards along the Levant coast actively or passively aided by human activity [27]. These species were named Lessepsian after the name of the constructor of the canal Engineer and Diplomat Ferdinand de Lesseps [28].
The aforementioned group of Lessepsian species established in the Mediterranean Sea, currently includes Fistularia commersonii [29] and Fistularia petimba, which belong to the Fistulariidae (order of Syngnathiformes), also called cornetfishes or flutemouths. There is only one genus in this family, Fistularia, with four different species: Fistularia commersonii Rüppell, 1838, Fistularia corneta Gilbert & Starks, 1904, Fistularia petimba Lacepède, 1803 and Fistularia tabacaria Linnaeus, 1758 [30]. The species F.tabacaria inhabits the tropical Atlantic, while its closest relative, F. commersonii, inhabits the Pacific and Indian Oceans. F. petimba spans the tropical Atlantic and Indo-West Pacific Oceans, whereas F. corneta is confined to the tropical eastern Pacific [31]. Fistularidae species are predators, inhabiting shallow waters of tropical and subtropical areas [31]. Although F. commersonii originated from the Indo-Pacific region [32] a widespread geographical distribution pattern has been observed in the eastern part of the Mediterranean Sea [33] with multiple sightings of this Lessepsian immigrant as a result of the opening of the Suez Canal in 1869. Due to its rapid growth and reproduction cycle, it has successfully formed large populations in the areas where it has been observed, with notable ecological impacts on the native species [34]. Fistularia commersonii is a piscivorous species mainly feeding on smaller fish and complements its diet with some Crustacea species. As the size of the species increases, there is a corresponding increase in the size of the prey consumed [34].
Another member of the Fistulariidae family is the red cornetfish, F. petimba which is naturally distributed in the Indo-West Pacific, the tropical Atlantic [32], the East Atlantic Ocean [35] and with a time lag of 20 years of its first citing in the western Mediterranean Sea (1996) it has the last ten years reported from several locations in the eastern Mediterranean Sea [36,37]. In this study, we show the immigration path of F.petimba in the eastern Mediterranean Sea through a stepping stone process of establishment through the Suez Canal [38,39].
2. Materials and Methods
Sampling Methodology
A monthly experimental fishery sampling was performed in Amorgos Island, Central Aegean Sea, between September 2022 and August 2023 (Figure 1).
The sampling method used was three types of gears: Gillnets (GNS), Trammel nets (GTR), and Long Lines (LLS). The gears used were designed to study the population dynamics of targeted fisheries species.
For GNS and GTR, nine different mesh sizes (20, 22, 24, 26, 28, 30, 32, 36, 45) and for LLS six different hook sizes (9, 10, 11, 12, 13, 14) were used to reflect the most commonly used mesh and hook sizes in small-scale fisheries of the Aegean Sea.
The nets used in this study had a total length of 900 meters, consisting of 100-meter panels for each of the nine mesh sizes. The height of each compartment was 100 meshes. The arrangement of mesh sizes for both GNS and GTR were randomly selected and illustrated in Figure 2 and Figure 3. For longlines, each compartment had a length of 200 meters and equipped with 200 hooks (100 hooks per hook size). One compartment comprised of the hook sizes 9 and 10, and two more compartments comprised of the combinations of hook sizes 11 – 12, and 13 – 14, respectively.
All samples were stored in a freezer (-20oC) until transported to the Laboratory of Applied Hydrobiology of the Agricultural University of Athens, Greece, for further examination.
Identification of the Species
Fistularia petimba was identified based on its morphological characteristics [31] following the genus description given by Fritzsche (1976): Fistularia species can be identified by their elongated body. Juveniles of this genus display a specific form, known as the "villosa Form" described by Liitken in 1880, characterized by rows of small hooked spinules on their bodies, which may or may not persist into adulthood. Notably, these fish may have elongated bony plates along the midline of their body anterior to the dorsal or anal fins. Their lateral line curves anteriorly, almost reaching the middle of their back, and extends posteriorly into a filament produced by the middle two caudal rays. Occasionally, the posterior lateral line ossifications bear sharp retrorse spines. They possess a small terminal mouth at the end of their elongated snout, and the first four vertebrae are fused, with a total vertebra count ranging from 76 to 87. Their teeth, found on the premaxillae, dentaries, and pharyngobranchials, are small. Notably, they lack a spinous dorsal fin, and both their dorsal and anal fins are short-based and opposed, each typically having 13-20 rays. Their caudal fin features 14 rays (7+7), and their branchiostegals are usually arranged in a pattern of 3 + 2 or 2 + 3.
Fistularia petimba is distinguished from its confamilial species by its specific morphological features [31]: number of rays on the dorsal fin (13-17), the anal fin (13-16) elongated bony plates embedded in the skin along the midline of the back, with posterior lateral line ossifications terminating in a retrorse spine. These distinctive traits served to distinguish F. petimba from other species of the same genus (Figure 4).
The following morphometric characteristics were measured to the nearest second decimal in mm or g: Total Length without filament (TL), Total Length with filament (TLf), Filament Length (fL), Standard Length (SL), Fork Length (FL), Body Deth (BD), Head Length (HL), Eye Diameter (ED), Snout Length (SN), Dorsal Fin Length (DFL), Dorsal Fin Height (DFH), Pectoral Fin Length (PFL), Pelvic Fin Height (PFH), Dorsal Fin Length (PvFL), Pelvic Fin Height (PvFH), Caudal Fin Length (CFL), Caudal Fin Height (CFH), Anal Fin Length (AFL), Anal Fin Height (AFH), Pre-dorsal Fin Length (pDFL), Pre-dorsal Fin Height (pDFH), Pre-pectoral Fin Length (pPFL), Pre-pectoral Fin Height (pPFH), Pre-pelvic Fin Length (pPvFL), Pre-pelvic Fin Height (pPvFH), Pre-anal Fin Length (pAFL), Pre-anal Fin Height (pAFH) and Total Wet Weight (TWW).
Stomach Content and Sex Maturity
Distibution of F.petimba in the Mediterranean Sea
In order to, compile a map with records of F.petimba in the Mediterranean Sea, a literature review (until February 2024) was performed using Google Scholar. This review used two main keywords, namely “Fistularia petimba” and “Red cornetfish” together with additional keywords to extend and maximize the capture probability of F.petimba records: “Mediterranean Sea”, “Invasive species”, “NIS”, “ecology”, “habitat” and “lessepsian”.
To visualize the species' geographical distribution in the Mediterranean Sea, a map illustrating the occurrences of F.petimba was generated using ArcGIS [46] and by integrating data from this study and published records from scientific journals.
3. Results
An individual of F.petimba (Figure 4) was captured with GTR with a mesh size of 26 mm, deployed between 24.5 m and 30.6 m depth on the 27th of May 2023 at 7:50 PM and hauled on the 28th of May 2023 at 6:40 AM (soak time 11 hours and 50 minutes), off the coast of Katapola Bay, Amorgos Island, Greece (lon="25.85879922" lat="36.82760281"). The measurements for each of the morphometric characteristics of the species are presented in Table 1.
The stomach of the individual was empty. Following macroscopic examination no gonads were identified and thus sex or maturity stage could not be determined.
4. Discussion
The expansion of global trade and travel has increased the chances for species to migrate beyond their native ranges [47,48]. This immigration often reflects the traffic patterns within regional or worldwide transportation systems. When an area is invaded, it becomes a source for the subsequent spread of the organism to other locations in the basin [49].
The Suez Canal, since its completion in the late 19th century, has served as a major conduit for the immigration of marine organisms between the Red Sea and the Mediterranean Sea. This artificial connection has facilitated the establishment of numerous NIS in the Mediterranean basin, reshaping the region's biodiversity and ecological dynamics [50].
Marinas in the Mediterranean Sea are significant areas, not only for the initial introductions of NIS, but also for subsequent secondary invasions, acting as stepping stones in the spread of NIS [48,51].
In the marine environment, NIS can become invasive, resulting in the displacement of indigenous species and thus leading to a range of negative consequences. These effects encompass the loss of native genetic diversity, alterations to habitats, shifts in community composition, changes to food web dynamics and ecosystem functions, disruptions to the provision of ecosystem services, threats to human health, and significant economic damages [52].
The capture of Fistularia petimba specimen in Amorgos Island, Central Aegean Sea represents a significant addition to the existing knowledge of the species' distribution in the Mediterranean region. Prior to this finding, there were 31 citings in the Mediterranean Sea, out of them only one was documented in the Aegean Sea, specifically in Samos (Figure 5, Table 2).
The first record in Cadiz, Spain appears to be an incidental catch from the endemic habitat in the Atlantic Ocean. The following documented occurrences of the Lessepsian immigrant, F. petimba, within the Mediterranean basin reveal a pattern of its progressive invasion into novel habitats. Having originated in the Red Sea, this species initiated a significant dispersal path along the coastlines of Syria, Egypt and Turkey. The species' course is validated with documented sightings in the area of Cyprus, concluding its eventual establishment within the Aegean Sea. Each documented occurrence represents a critical point in the species' biogeographic spread throughout the Mediterranean Sea.
This interpretation of chronological milestones offers valuable insights into the stepping-stone spreading pattern characterizing F. petimba's invasion in the Mediterranean Sea. Tracing its cross-border path from Red Sea to the Aegean Sea, provides researchers with a deeper understanding of the mechanisms of invasive populations' establishment within marine ecosystems.
According to Papageorgiou et al., 2023 the mean total length for gonad maturity is 440 mm for females and 410 mm. The specimen of this study (390 mm) also agrees with those results as no gonads were visible macroscopically. There was no histological examination performed.
The occurrence of this species in Amorgos Island suggests a wider presence within the Aegean Sea than previously recognized. This finding underscores the importance of ongoing fisheries research in uncovering new occurrences and expanding our understanding of marine biodiversity in the region. Further investigations into the ecological preferences, population dynamics, and potential impact of Fistularia petimba in the Aegean Sea and the Mediterranean basin are important to understand succession rates and ecological impacts to enhance conservation efforts and inform sustainable management practices.
Funding
This research was co-funded by Blue Marine Foundation (BMF) and Cyclades Preservation Fund (CPF) – Conservation Collective (CC) under the project entitled “Base line study for fisheries management on coastal areas based on local ecological knowledge in Amorgos Island”.
Acknowledgments
The authors thank the Blue Marine Foundation (BMF) and Cyclades Preservation Fund (CPF) for their support, as well as, the Professional Fishing Association of Amorgos and Ioannis Vekris for their expertise and assistance throughout all aspects of our study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Agency, E. e. Climate change, impacts and vulnerability in Europe EEA 2012. https://www.eea.europa.eu/publications/climate-impacts-and-vulnerability-2012.
- Lewis, S. L.; Maslin, M. A. Defining the Anthropocene. Nature 2015, 519, 171–180. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P. E.; Simberloff, D.; Bacher, S.; Blackburn, T. M.; Carlton, J. T.; Dawson, W.; Essl, F.; Foxcroft, L. C.; Genovesi, P.; et al. Scientists' warning on invasive alien species. Biol Rev Camb Philos Soc 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Genner, M. J.; Sims, D. W.; Wearmouth, V. J.; Southall, E. J.; Southward, A. J.; Henderson, P. A.; Hawkins, S. J. Regional climatic warming drives long-term community changes of British marine fish. Proc Biol Sci 2004, 271, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Munday, P. L.; Jones, G. P.; Pratchett, M. S.; Williams, A. J. Climate change and the future for coral reef fishes. Fish and Fisheries 2008, 9, 261–285. [Google Scholar] [CrossRef]
- Simpson, Stephen D. ; Jennings, S.; Johnson, Mark P.; Blanchard, Julia L.; Schön, P.-J.; Sims, David W.; Genner, Martin J. Continental Shelf-Wide Response of a Fish Assemblage to Rapid Warming of the Sea. Current Biology 2011, 21, 1565–1570. [Google Scholar] [CrossRef]
- Pauly, D.; Christensen, V.; Dalsgaard, J.; Froese, R.; Torres, F. Fishing Down Marine Food Webs. Science 1998, 279, 860–863. [Google Scholar] [CrossRef]
- Jennings, S.; Alvsvåg, J.; Cotter, A. J. R.; Ehrich, S.; Greenstreet, S. P. R.; Jarre-Teichmann, A.; Mergardt, N.; Rijnsdorp, A. D.; Smedstad, O. Fishing effects in northeast Atlantic shelf seas: patterns in fishing effort, diversity and community structure. III. International trawling effort in the North Sea: an analysis of spatial and temporal trends. Fisheries Research 1999, 40, 125–134. [Google Scholar] [CrossRef]
- Daan, N.; Gislason, H.; G. Pope, J.; C. Rice, J. Changes in the North Sea fish community: evidence of indirect effects of fishing? ICES Journal of Marine Science 2005, 62, 177–188. [Google Scholar] [CrossRef]
- Jones, G. P.; McCormick, M. I.; Srinivasan, M.; Eagle, J. V. Coral decline threatens fish biodiversity in marine reserves. Proceedings of the National Academy of Sciences 2004, 101, 8251–8253. [Google Scholar] [CrossRef]
- Airoldi, L.; Balata, D.; Beck, M. W. The Gray Zone: Relationships between habitat loss and marine diversity and their applications in conservation. Journal of Experimental Marine Biology and Ecology 2008, 366, 8–15. [Google Scholar] [CrossRef]
- WILSON, S. K.; FISHER, R.; PRATCHETT, M. S.; GRAHAM, N. A. J.; DULVY, N. K.; TURNER, R. A.; CAKACAKA, A.; POLUNIN, N. V. C.; RUSHTON, S. P. Exploitation and habitat degradation as agents of change within coral reef fish communities. Global Change Biology 2008, 14, 2796–2809. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R. A.; Mittermeier, C. G.; da Fonseca, G. A. B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C. N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PLoS One 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Cinar, M. E.; Garcia Raso, J. E.; Bianchi, C. N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterranean Marine Science 2010, 11. [Google Scholar] [CrossRef]
- Costello, M. J.; Coll, M.; Danovaro, R.; Halpin, P.; Ojaveer, H.; Miloslavich, P. A Census of Marine Biodiversity Knowledge, Resources, and Future Challenges. PLOS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Ambrogi, A. O. Biotic Invasions in a Mediterranean Lagoon. Biological Invasions 2000, 2, 165–176. [Google Scholar] [CrossRef]
- Occhipinti-Ambrogi, A.; Marchini, A.; Cantone, G.; Castelli, A.; Chimenz, C.; Cormaci, M.; Froglia, C.; Furnari, G.; Gambi, M. C.; Giaccone, G.; et al. Alien species along the Italian coasts: an overview. Biological Invasions 2011, 13, 215–237. [Google Scholar] [CrossRef]
- Romeo, T.; Pietro, B.; Pedà, C.; Consoli, P.; Andaloro, F.; Fossi, M. C. First evidence of presence of plastic debris in stomach of large pelagic fish in the Mediterranean Sea. Mar Pollut Bull 2015, 95, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Goren, M.; Galil, B. S. A review of changes in the fish assemblages of Levantine inland and marine ecosystems following the introduction of non-native fishes. Journal of Applied Ichthyology 2005, 21, 364–370. [Google Scholar] [CrossRef]
- Harmelin, M.; Bitar, G.; Harmelin, J.-G.; Monestiez, P. The littoral fish community of the Lebanese rocky coast (eastern Mediterranean Sea) with emphasis on Red Sea immigrants. Biological Invasions 2005, 7, 625–637. [Google Scholar] [CrossRef]
- Parravicini, V.; Azzurro, E.; Kulbicki, M.; Belmaker, J. Niche shift can impair the ability to predict invasion risk in the marine realm: an illustration using Mediterranean fish invaders. Ecology Letters 2015, 18, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Raitsos, D.; Beaugrand, G.; Georgopoulos, D.; Zenetos, A.; Pancucci, A.; Theocharis, A.; Papathanassiou, E. Global climate change amplifies the entry of tropical species into the Eastern Mediterranean Sea. Limnology and oceanography 2010, 55, 1478–1484. [Google Scholar] [CrossRef]
- Kalogirou, S.; Azzurro, E.; Bariche, M. The Ongoing Shift of Mediterranean Coastal Fish Assemblages and the Spread of Non-Indigenous Species. 2012.
- Cinar, M. E.; Bilecenoglu, M.; Ozturk, Β.; Katagan, Τ.; Yokes, Μ. Β.; Aysel, V.; Dagli, E.; Acik, S.; Ozcan, T.; Erdogan, H. An updated review of alien species on the coasts of Turkey. Mediterranean Marine Science 2011, 12, 257–315. [Google Scholar] [CrossRef]
- Golani, D. The Sandy Shore of the Red Sea-Launching Pad for Lessepsian (Suez Canal) Migrant Fish Colonizers of the Eastern Mediterranean. Journal of Biogeography 1993, 20, 579–585. [Google Scholar] [CrossRef]
- Galil, B. S.; Mienis, H. K.; Hoffman, R.; Goren, M. Non-indigenous species along the Israeli Mediterranean coast: tally, policy, outlook. Hydrobiologia 2021, 848, 2011–2029. [Google Scholar] [CrossRef]
- Por, F. D. Lessepsian Migration: The Influx of Red Sea Biota into the Mediterranean by Way of the Suez Canal; Springer, 1978. [CrossRef]
- Stern, N. I. R.; Paz, G. U. Y.; Yudkovsky, Y.; Lubinevsky, H.; Rinkevich, B. The arrival of a second ‘Lessepsian sprinter’? A first record of the red cornetfish Fistularia petimba in the Eastern Mediterranean. Mediterranean Marine Science 2018, 18, 524–528. [Google Scholar] [CrossRef]
- Ragheb, E. Morphometric and meristic characteristics of the first record Fistularia petimba (Lacepède, 1803) and Fistularia commersonii (Rüppell, 1838) (Piscès: Fistulariidae) from the Egyptian Mediterranean waters (West Alexandria). The Egyptian Journal of Aquatic Research 2022, 48, 143–150. [Google Scholar] [CrossRef]
- Fritzsche, R. A. A REVIEW OF THE CORNETFISHES, GENUS FISTULARIA (FISTULARIIDAE), WITH A DISCUSSION OF INTRAGENERIC RELATIONSHIPS AND ZOOGEOGRAPHY. BULLETIN OF MARINE SCIENCE 1976, 26, 106–204. [Google Scholar]
- Fischer, W. B.; G., *!!! REPLACE !!!* (Eds.) ;Fisheries and Aquaculture Management Division. FAO Species identification sheets for fishery purposes. Western Indian Ocean: fishing area 51. 1984.
- Sánchez-Tocino, L.; Hidalgo, F.; Pontes, M. Primera cita de Fistularia commersonii Ruppell, 1838 (Osteichtyes: Fistulariidae) en aguas mediterráneas de la Península Ibérica. Zoologica Baetica 2007, 18, 79–84. [Google Scholar]
- Kalogirou, S.; Corsini-Foka, M.; Kondylatos, G.; Wennhage, H. Diet of the invasive piscivorous fish Fistularia commersonii in a recently colonized area of eastern Mediterranean. Biological Invasions 2007, 9, 887–896. [Google Scholar] [CrossRef]
- Bañón, R.; Sande, C. First record of the red cornetfish Fistularia petimba (Syngnathiformes: Fistularidae) in Galician waters: a northernmost occurrence in the eastern Atlantic. Journal of Applied Ichthyology 2008, 24, 106–107. [Google Scholar] [CrossRef]
- Cárdenas, S.; Alvarez-Berastegui, D.; Ortiz, J. M. First record of Fistularia petimba Laccepéde, 1803 (Pisces FIstulariidae) off the coast of Cadiz (southern Iberian Peninsula). Boletín del Instituto Español de Oceanografía 1997, 13, 83–86. [Google Scholar]
- Ünlüoğlu, A.; Akalın, S.; Dal, İ.; Tıraşın, E.; Aydın, C. First record of red cornetfish Fistularia petimba (Syngnathiformes: Fistulariidae) from Antalya and İskenderun Bays along Turkish coasts of the Mediterranean Sea. Journal of Applied Ichthyology 2018, 34, 977–980. [Google Scholar] [CrossRef]
- Soydemir Çiftçi, N.; Ayas, D.; Doğangün, M. New locality record for Fistularia petimba Lacepède, 1803 from the Northeastern Mediterranean Sea (Mersin Bay). 2019.
- Karan, S.; Uyan, A.; Doğdu, S.; Gürlek, M.; Ergüden, D.; Turan, C. Genetic confirmation of Red cornetfish, Fistularia petimba (Syngnathiformes: Fistularidae) occurrence in Turkish marine waters. 2019, 4, 125-129.
- Frost, W. E. Observations on the Food of Eels (Anguilla anguilla) from the Windermere Catchment Area. Journal of Animal Ecology 1946, 15, 43–53. [Google Scholar] [CrossRef]
- Hyslop, E. J. Stomach contents analysis—a review of methods and their application. Journal of Fish Biology 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Hynes, H. B. N. THE FOOD OF THE FRESHWATER STICKLEBACKS (GASTEROSTEUS ACULIEATUS AND PYGOSTEUS PUNGITIUS) WITH A REVIEW OF METHODS USED N THE STUDIES OF THE FOOD OF FISHES. Journal of Animal Ecology 1950, 19, 36–58. [Google Scholar] [CrossRef]
- Brown-Peterson, N. J.; Wyanski, D. M.; Saborido-Rey, F.; Macewicz, B. J.; Lowerre-Barbieri, S. K. A Standardized Terminology for Describing Reproductive Development in Fishes. Marine and Coastal Fisheries 2011, 3, 52–70. [Google Scholar] [CrossRef]
- Ungaro, N. Field manual on macroscopic identification of maturity stages for the Mediterranean fishery resources.; Rome, 2008. https://www.faomedsudmed.org/pdf/publications/TD21.pdf.
- Follesa, C. , Carbonara, P. Atlas of the maturity stages of Mediterranean fishery resources, /: Italy, 2019. https, 2019. [Google Scholar]
- Esri. ArcGIS Online | Cloud-Based GIS Mapping Platform. 2018. Available online: https://www.esri.com/en-us/arcgis/products/arcgis-online/overview (accessed on 26 March 2024).
- Floerl, O.; Inglis, G. J. Starting the invasion pathway: the interaction between source populations and human transport vectors. Biological Invasions 2005, 7, 589–606. [Google Scholar] [CrossRef]
- Ferrario, J.; Caronni, S.; Occhipinti-Ambrogi, A.; Marchini, A. Role of commercial harbours and recreational marinas in the spread of non-indigenous fouling species. Biofouling 2017, 33, 651–660. [Google Scholar] [CrossRef]
- Drake, J. M.; Lodge, D. M. Global hot spots of biological invasions: evaluating options for ballast–water management. Proceedings of the Royal Society of London. Series B: Biological Sciences 2004, 271, 575–580. [Google Scholar] [CrossRef]
- Galil, B. S.; Boero, F.; Campbell, M. L.; Carlton, J. T.; Cook, E.; Fraschetti, S.; Gollasch, S.; Hewitt, C. L.; Jelmert, A.; Macpherson, E.; et al. ‘Double trouble’: the expansion of the Suez Canal and marine bioinvasions in the Mediterranean Sea. Biological Invasions 2015, 17, 973–976. [Google Scholar] [CrossRef]
- Ulman, A.; Ferrario, J.; Occhipinti, A.; Arvanitidis, C.; Bandi, A.; Bertolino, M.; Bogi, C.; Chatzigeorgiou, G.; Çiçek, B.; Içek, C.; et al. A massive update of non-indigenous species records in Mediterranean marinas. PeerJ 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Katsanevakis, S. , Wallentinus, I. , Zenetos,A., Leppäkoski,E., Çinar,M.E., Oztürk,B., Grabowski,M., Golani, D., Cardoso,A. C. Impacts of invasive alien marine species on ecosystem services and biodiversity: a pan-European review Aquatic Invasions 2014, 9, 391–423. [Google Scholar]
- Crocetta, F.; Al Mabruk, S. A. A.; Azzurro, E.; Bakiu, R.; Bariche, M.; Batjakas, I. E.; Bejaoui, T.; Ben Souissi, J.; Cauchi, J.; Corsini-Foka, M.; et al. “New Alien Mediterranean Biodiversity Records” (November 2021). Mediterranean Marine Science 2021, 22, 724–746. [Google Scholar] [CrossRef]
- Chirine, H.; Ibrahim, A.; Alshawy, F. First record of Red cornetfish, Fistularia petimba Lacepède, 1803 (Actinopterygii: Fistulariidae) from the Syrian coast. International Journal of Aquatic Biology 2019, 7, 175–179. [Google Scholar] [CrossRef]
- Ibrahim, A.; Alshawy, F.; Hussein, C. Confirmation records and new distribution of the red cornet fish fistularia petimba lacepède, 1803 (actinopterygii: fistulariidae) in the Syrian Marine Waters (Eastern Mediterranean). Species 2020, 21, 95–100. [Google Scholar]
- Dragicevic, B.; Anadoli, O.; Angel, D.; Benabdi, M.; Bitar, G.; Castriota, L.; Crocetta, F.; Deidun, A.; Dulčić, J.; Edelist, D. New Mediterranean Biodiversity Records (19). 2019. 20 December.
- Cerim, H.; Yapıcı, S.; Gülşahin, A.; Soykan, O.; Bilge, G. The First Record of the Red Cornetfısh (Fistularia petimba Lacepède, 1803) in the Aegean Sea. Düzce Üniversitesi Bilim ve Teknoloji Dergisi 2021, 9, 607–615. [Google Scholar] [CrossRef]
- Papageorgiou, M.; Resaikos, V.; Petrou, A. A preliminary assessment of Fistularia petimba (Lacepède, 1803) migration in the Mediterranean Sea: historical and new data from Cyprus (Levantine Sea) with biological notes. Mediterranean Marine Science 2023, 24, 446–453. [Google Scholar] [CrossRef]
Figure 1.
Map of the study area in Amorgos Island, Greece. The red line marks the proposed seasonal Fisheries Restricted Areas (FRAs), while the yellow areas indicate the proposed permanent FRAs.
Figure 1.
Map of the study area in Amorgos Island, Greece. The red line marks the proposed seasonal Fisheries Restricted Areas (FRAs), while the yellow areas indicate the proposed permanent FRAs.
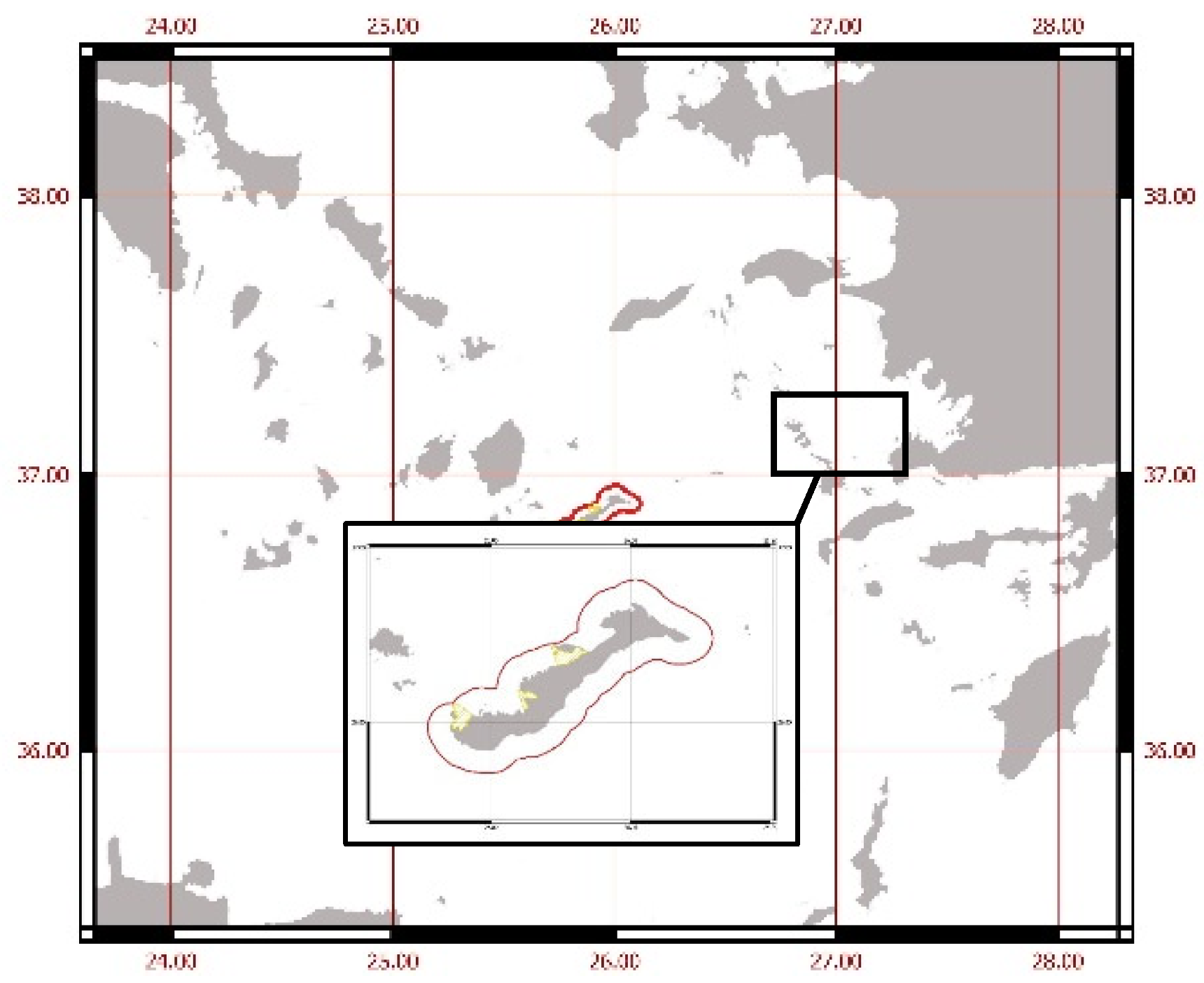
Figure 2.
GNS mesh size arrangement on this study.
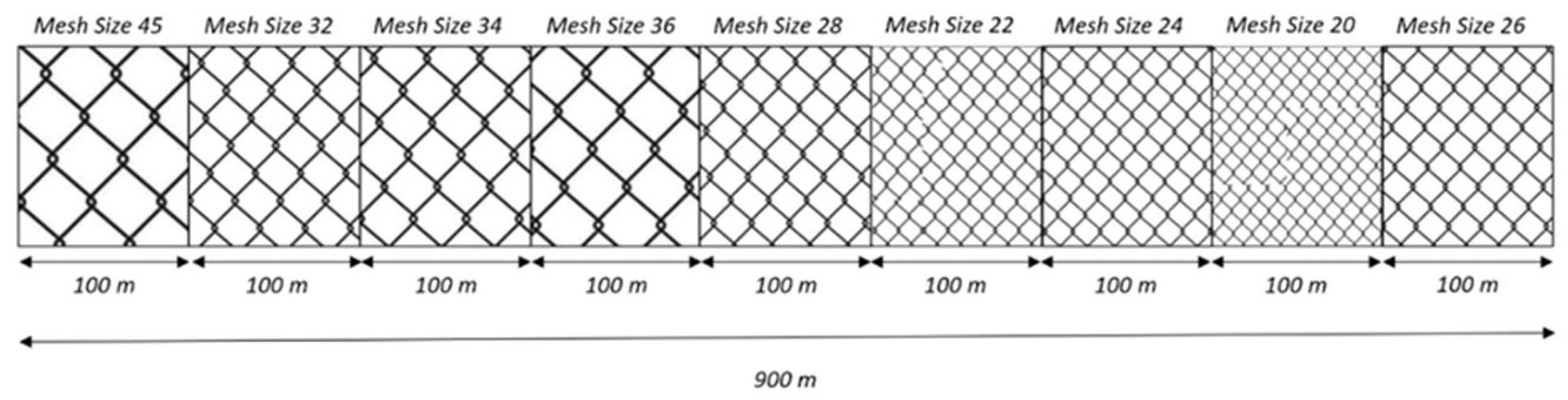
Figure 3.
GTR mesh size arrangement on this study.
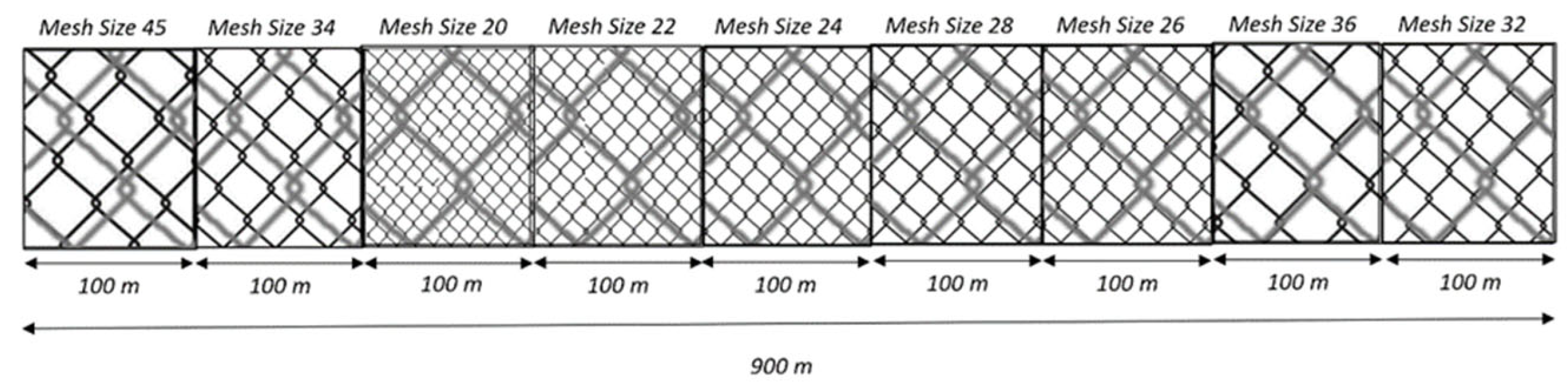
Figure 4.
Fistularia petimba individual from Amorgos Island and its identification characteristics: (a) reddish colour and (b) elongate bony plates embedded in skin.
Figure 4.
Fistularia petimba individual from Amorgos Island and its identification characteristics: (a) reddish colour and (b) elongate bony plates embedded in skin.

Figure 5.
Citings of Fistularia petimba in the Mediterranean Sea [46].
Figure 5.
Citings of Fistularia petimba in the Mediterranean Sea [46].
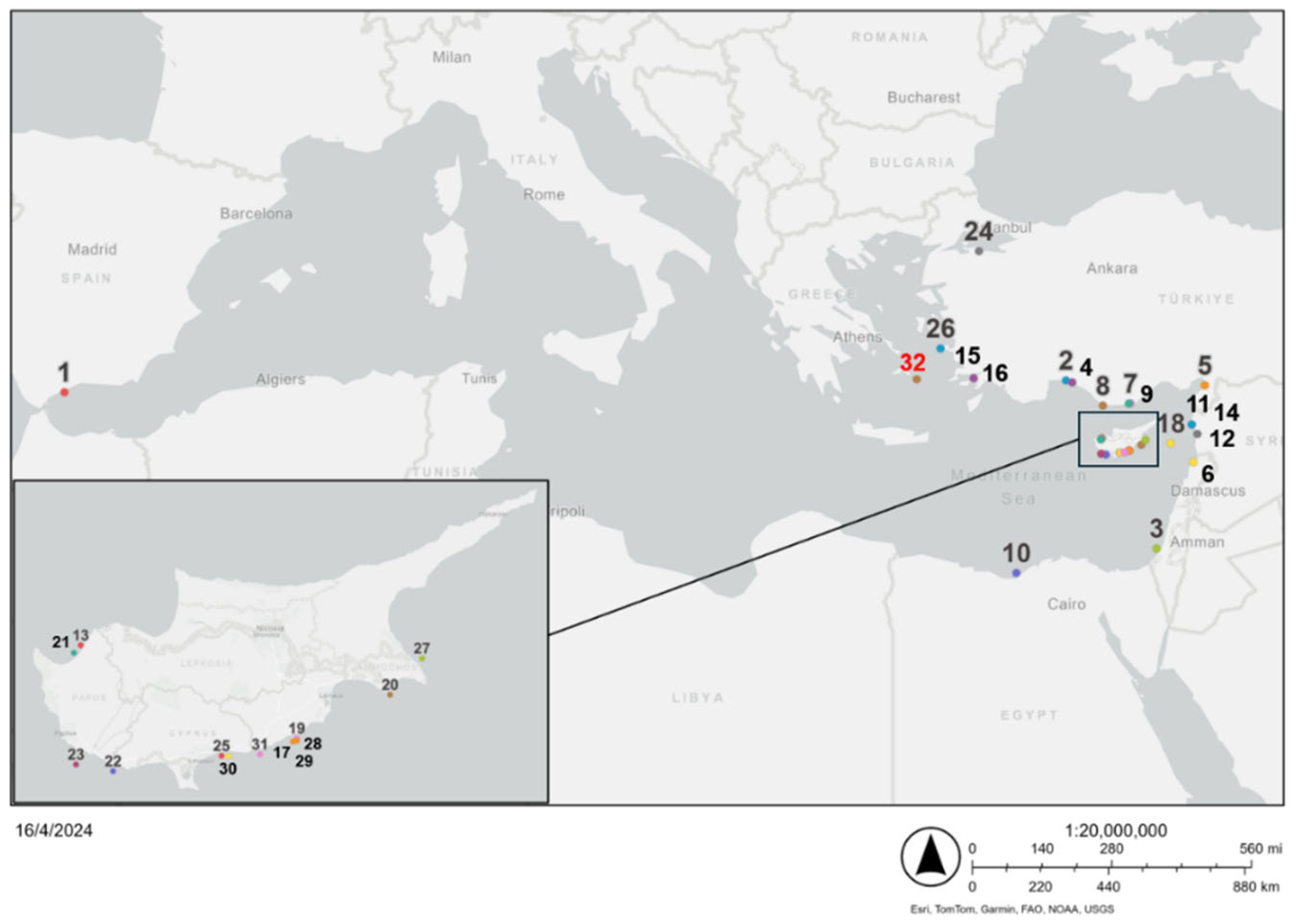

Table 1.
Morphometric characheristics and measurements of Fistularia petimba from Amorgos Island, Greece.
Table 1.
Morphometric characheristics and measurements of Fistularia petimba from Amorgos Island, Greece.
| Morphometrics | Measurement (mm or g) |
|---|---|
| Total Length without filament (TL) | 395.00 |
| Total Length with filament (TLf) | 530.00 |
| Filament Length (fL) | 124.23 |
| Standard Length (SL) | 378.00 |
| Fork Length (FL) | 383.00 |
| Body Deth (BD) | 7.98 |
| Head Length (HL) | 142.00 |
| Eye Diameter (ED) | 10.97 |
| Snout Length (SN) | 114.00 |
| Dorsal Fin Length (DFL) | 15.41 |
| Dorsal Fin Height (DFH) | 28.89 |
| Pectoral Fin Length (PFL) | 6.66 |
| Pelvic Fin Height (PFH) | 16.81 |
| Dorsal Fin Length (PvFL) | 2.54 |
| Pelvic Fin Height (PvFH) | 6.83 |
| Caudal Fin Length (CFL) | 19.01 |
| Caudal Fin Height (CFH) | 5.10 |
| Anal Fin Length (AFL) | 15.16 |
| Anal Fin Height (AFH) | 27.37 |
| Pre-dorsal Fin Length (pDFL) | 67.00 |
| Pre-pectoral Fin Length (pPFL) | 319.00 |
| Pre-pelvic Fin Length (pPvFL) | 190.00 |
| Pre-anal Fin Length (pAFL) | 310.00 |
| Total Wet Weight (TWW) | 34.00 |
Table 2.
Data of the 32 citings of Fistularia petimba in the Mediterranean Sea.
| No | Location | Latitude | Longitude | Date (capture) | Depth (m) | Gear Type | Sample size |
|---|---|---|---|---|---|---|---|
| 1 | Cadiz, Spain [36] | 36.455097 | -4.703372 | 23/06/1996 | 50 | Gillnet | 1 |
| 2 | Antalya Bay, Turkey [37] | 36.793556 | 31.209167 | 28/10/2016 | 35 – 43 | Bottom trawl | 1 |
| 3 | Ashdod, Israel [29] | 31.813950 | 34.459717 | 12/11/2016 | 80 | Bottom trawl | 1 |
| 4 | Antalya Bay, Turkey [37] | 36.737417 | 31.434361 | 26/11/2016 | 30 | Bottom trawl | 1 |
| 5 | Iskenderun, Turkey [37] | 36.654400 | 36.186183 | 21/05/2017 | 35 – 38 | Bottom trawl | 2 |
| 6 | Tripoli, Lebanon [53] | 34.410000 | 35.770000 | 15/11/2017 | N/A | Gillnet | 1 |
| 7 | Mersin Bay, Turkey [39] | 36.128833 | 33.520667 | 22/11/2017 | 95 | Bottom trawl | 1 |
| 8 | Antalya Bay, Turkey [39] | 36.061867 | 32.534233 | 09/01/2018 | 70 | Bottom trawl | 2 |
| 9 | Büyükeceli Coast (Mersin Bay)Turkey [38] | 36.123139 | 33.467944 | 05/10/2018 | 150 | Bottom trawl | 2 |
| 10 | Egypt [30] | El-Hamam - Sidi Kirayr. | 09/03/2019 | 40 – 60 | Bottom trawl | 1 | |
| 11 | Lattakia, Syria [54] | 35.518325 | 35.713492 | 29/07/2019 | 45 | Gillnet | 1 |
| 12 | Lattakia, Syria [55] | 35.243086 | 35.920000 | 24/09/2019 | 30 | Gillnet | 1 |
| 13 | Gialia, Cyprus [56] | 35.110000 | 32.490000 | 26/09/2019 | 55 | Gillnet | 1 |
| 14 | Banyas, Syria [55] | 35.518325 | 35.713492 | 29/09/2019 | 45 | Gillnet | 2 |
| 15 | Gökova Bay, Turkey [57] | 36.857889 | 27.896556 | 19/10/2019 | 15 – 20 | Longline | 1 |
| 16 | Güllük Bay, Turkey [57] | 36.857883 | 27.896561 | 17/11/2019 | 65 | Bottom trawl | 4 |
| 17 | Cyprus [58] | 34.747367 | 33.463400 | 14/07/2020 | 55 | Bottom trawl | 3 |
| 18 | Cyprus [58] | 34.964500 | 34.964500 | 15/07/2020 | 48 | Bottom trawl | 1 |
| 19 | Cyprus [58] | 34.759617 | 33.480650 | 16/07/2020 | 33 | Bottom trawl | 1 |
| 20 | Cyprus [58] | 34.924100 | 33.908050 | 24/07/2020 | 79 | Bottom trawl | 11 |
| 21 | Cyprus [58] | 35.081733 | 32.458700 | 24/07/2020 | 43 | Bottom trawl | 29 |
| 22 | Cyprus [58] | 34.635717 | 32.638517 | 27/03/2021 | 46 | Bottom trawl | 10 |
| 23 | Cyprus [58] | 34.661300 | 32.468650 | 27/03/2021 | 93 | Bottom trawl | 4 |
| 24 | Bandirma Bay, Turkey [53] | 40.416950 | 28.084000 | 11/06/2021 | 32 | Trammel net | 1 |
| 25 | Cyprus [58] | 34.693983 | 33.135567 | 04/08/2021 | 44 | Bottom trawl | 26 |
| 26 | Samos, Greece [53] | 37.706583 | 26.708783 | 07/11/2021 | 20 | Trammel net | 1 |
| 27 | Cyprus [58] | 35.060833 | 34.054383 | 08/08/2021 | 86 | Bottom trawl | 4 |
| 28 | Cyprus [58] | 34.750917 | 33.480933 | 08/08/2021 | 55 | Bottom trawl | 7 |
| 29 | Cyprus [58] | 34.750917 | 33.480933 | 08/08/2021 | 33 | Bottom trawl | 2 |
| 30 | Cyprus [58] | 34.692267 | 33.166750 | 08/08/2021 | 56 | Bottom trawl | 1 |
| 31 | Cyprus [58] | 34.699333 | 33.311817 | 13/09/2021 | 13 | Trammel net | 1 |
| 32 | Amorgos, Greece (current study) | 36.82760281 | 25.85879922 | 28/05/2023 | 24.5 - 30.6 | Trammel net | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



