
Submitted:
28 April 2024
Posted:
29 April 2024
You are already at the latest version
Abstract
The study of biologically active substances from natural sources shows the prospect of obtaining a new type of antimicrobial agents, highly effective against pathogenic microorganisms and non-toxic to humans. Our study reveals for the first time that Cornu aspersum mucus has the potential to act as a source of antibacterial agents against bacterial pathogens such as Bacillus cereus 1085, Propionibacterium acnes 1897, Salmonella enterica 8691, Enterococcus faecalis 3915, and Enterococcus faecium 8754. Both mucus fractions with MW<20 kDa and MW>20 kDa showed antimicrobial activity and antioxidant properties that’s why they can be turned into an alternative agent to expensive synthetic antibacterial compounds. Using de novo sequencing 16 novel peptides with potential antibacterial activity were identified by in a fraction with MW<20 kDa. The results of electrophoretic and proteomic analysis of mucus fraction with MW >20 kDa presented proteins with potential antibacterial activity highly homologous to hemocyanins, mucus protein Aspernin, lectins, L-amino acid oxidase-like protein, and mucins (mucin-5AC, mucin-5B, mucin-2 and mucin-17). Thanks to the synergistic action of the bioactive components, the mucus fraction with MW > 20 kDa in low concentrations per 8–2 µg/mL showed promising antimicrobial activity against the tested bacteria, comparable to Vancomycin and without unwanted cytotoxic effects against non-target cells. Additionally, C. aspersum mucus fractions were found to have a positive effect on eukaryotic organisms by reducing the levels of intracellular oxidative damage and increasing the antioxidant capacity of the cell. These findings could serve as a basis for the development of a potent non-cytotoxic antibacterial agent to replace conventional antibiotics and avoid the development of antibiotic resistance.
Keywords:
Cornu aspersum mucus
; antimicrobial peptides
; de novo MS/MS sequencing
; proteomic analysis
; mucus proteins whit antimicrobial action
1. Introduction
The World Health Organization (WHO) has reported an alarming increase in the number of resistant bacterial strains to conventional antibiotics, which is a threat to public health safety [1]. Also, antibiotic resistance (AR) is a global problem with an important economic impact [2,3]. The causes of AMR are multifactorial, with the indiscriminate and prolonged use of antibiotics in both human and veterinary medicine and agriculture being paramount to the development and spread of drug-resistant microorganisms [4,5].
Bacteria are complex organisms and the effect of antibiotics on bacterial DNA is weak, but may provoke a new acquired resistance mechanism [6]. AMR is a public health problem, spreading through humans, animals (domestic and wild) and the environment (water and air) [4]. Antibiotic resistance is therefore a silent pandemic that requires global health solutions [7]. The problem of antibiotic resistance necessitates an urgent search for alternatives to conventional antibiotics with new modes of action and less susceptibility to bacterial resistance.
Antimicrobial peptides (AMPs) derived from natural sources have been shown to exhibit broad-spectrum antimicrobial activity with high specificity and low toxicity and are an excellent option to combat antibiotic resistance [8]. The mode of action of AMPs relates to osmotic lysis due to interaction with the bacterial membrane. Research also demonstrates that AMPs result in membrane damage, inhibit macromolecular synthesis, damage cellular organelle and DNA, inhibit enzymes and regulate host immunity [9]. The presence of a cation-rich moiety and hydrophobic amino acids is typical in AMPs, resulting in a cationic arrangement with amphiphilic (hydrophilic and hydrophobic) characteristics [10,11]. This enables specificity to prokaryotic membranes, as mammalian cells have a net-neutral charge due to the presence of zwitterionic phospholipids, for example, phosphatidylethanolamine, phosphatidylcholine or sphingomyelin [11,12].
AMPs play a crucial role against various infections in invertebrate species, which, unlike vertebrates, do not have an adaptive immune system and, therefore, rely solely on innate defence mechanisms [13]. Invertebrate organisms of the phylum Mollusca represent one of the largest reservoirs of pharmacologically active compounds due to their great diversity (surpassed only by arthropods) and their ability to adapt to almost all types of habitats [14,15]. Bioactive compounds from haemolymph and mucus from gastropods have been shown to be powerful bioactive combinations with potential applications in combating pathogenic bacteria [16,17,18,19,20,21,22,23,24].
In the last decade, several studies reported antibacterial and antioxidant properties, as well as healing potential in treating wounds on the mucus of some land snails [16,19,20,21,24,25,26,27]. The antimicrobial activity of land snail mucus is known to be related not only to the presence of AMPs but also to some antibacterial proteins and glycoproteins [29]. A number of studies have reported that some proteins and glycoproteins in the mucus of various land snails (Achatina fulica, H. aspersa, Cryptozona bistrialis, Lissachatina fulica and Hemiplecta differenta) are responsible for the antimicrobial properties of extracts from these snails [20,21,24,28,29,30,31,32].
The presented analyses in the current study are on new natural compounds with antimicrobial activity that upgrade our previous studies on the antibacterial activity of Cornu aspersum snail mucus. New information is provided on the antibacterial activity of two mucus fractions with MW<20 and MW>20 kDa from C. aspersum against 5 pathogenic bacteria (Bacillus cereus 1085, Propionibacterium acnes 1897, Salmonella enterica 8691, Enterococcus faecalis 3915, and Enterococcus faecium 8754) and the identified important mucus components associated with antimicrobial activity.
2. Results
2.1. Preparation of Snail Mucus Extract
The mucus was collected from snails C. aspersum, growing in Bulgarian eco-farms using patented technology, without disturbing the biological functions of snails [17,19]. The resulting crude mucus extract was homogenized and centrifuged to remove coarse impurities. The purified native mucus extract was separated into 2 major fractions by ultrafiltration under pressure with polyethersulfone membrane filters with pore sizes of 20 kDa (Microdyn Nadir™ from STERLITECH Corporation, Goleta, CA, USA).
2.2. Analysis and Characteristics of the Isolated Mucus Fraction with MW < 20 kDa
After purification by RP-HPLC on a Nucleosil C18 column, the molecular masses of the bioactive compounds in the fraction with MW < 20 kDa (Figure 1) were identified by mass spectrometry.
2.2.1. Molecular Mass Analysis and de Novo Sequencing of Peptides by Mass Spectrometry
A thorough characterization of a fraction with MW < 20 kDa is presented by Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry analyses (MADI-TOF-MS analysis) in the range to 3 kDa (Figure 2) and a range between 3 -20 kDa (Figure 3). The MS-spectrum presented in Figure 2 shows that the mucus fraction with MW < 3 kDa contained various peptides with different masses in the region between 900–3011 Da. Peptides determined as protonated molecule ions [M + H]+ at m/z 1376 Da, 1438 Da, 1738 Da, 1796 Da, 1909 Da, 1966 Da, and 2292 Da dominate the MS-spectrum (Figure 2).
Peptides and polypeptides with higher molecular weights detected in fraction with MW < 20 kDa are presented on MS-spectrum in Figure 3, as molecular protonated ions [M + H]+ at m/z 5573.742 Da, 6429.681 Da, 7337.199 Da, 11143.605 Da, 12464.216 Da, 15059.328 Da and 18005.169 Da as well as various other ions with lower intensity. The determined ions [M+H]+ at m/z 12.464 Da, 15059.328 Da and 18005.169 Da are in good agreement with similar proteins in snail mucus of C. aspersum [24,33], Helix pomatia [34] and A. fulica [35] reported previously.
The amino acid sequences (AAS) of low molecular peptides (with MW < 3 kDa) were identified by de novo sequencing experiments (MS/MS analysis) of the [M+H]+ ions. Following b- and y- fragmentation ions in MS/MS spectrum of peptide [M+H]+ at m/z 1966.11, the amino acid sequence LLLDNKGGGLVGGLLGGGGKGGG was identified (Figure 4).
Thus, the amino acid sequences of 16 new peptides with molecular masses below 3000 Da were identified and AAS of another 6 peptides were confirmed (Table 1). The determined physicochemical characteristics of the identified peptides, such as isoelectric points (pI), grand average of hydropathicity (GRAVY), and net charge by the ExPASy MW/pI tool program and ExPASy ProtParam tool are presented in Table 1. The analysis showed the presence of both cationic and anionic, as well as neutral amphipathic peptide structures.
The analysis showed the presence of cationic, anionic and neutral amphipathic peptide structures with generally hydrophobic surfaces, but 9 hydrophilic peptides were also identified (Nos. 1, 6–10, 12, and 15, Table 1). Based on the founded primary structures of peptides (Table 1) their antimicrobial activity was predicted using iAMPpred software (http://cabgrid.res.in:8080/amppred) (accessed on 6 March 2024), an extensive database [39]. The results showed that peptides Nos 3, 7, 8, 13, 17, 18, 20, 21–23, had the highest prognostic, antibacterial and antifungal activities, while peptides 8 and 17 had the highest prognostic antiviral activities.
The alignment of the AASs of peptides shown in Table 1 with database AMPs by CAMPSing software (http://www.campsign.bicnirrh.res.in/blast.php, accessed on 24 March 2024) revealed similarity with known antimicrobial peptides (presented in Supplementary Materials SI1). Peptides Nos 10, 13, 17, 18, 20, 21, 22, and 23 show high homology, with glycine-rich antimicrobial peptides—Procambarin, Holotricin, Microcin B, Acanthoscurrin and Ctenidin, with Gly/Leu rich antimicrobial peptide Leptoglycin, Glycine-rich protein GWK, while peptides Nos 7, 8, 12, and 15 revealed homologies with Defensin-like protein 196, Glycine-rich protein GWK, Crustin-like antimicrobial peptide, different forms of Gallinacin and Shepherin (presented in Supplementary Materials, SI1).
Presented evidence confirmed that identified mucus peptides belong to the AMP family. Furthermore, the obtained results can be considered as basic information in the study of bioactive peptides from the C. aspersum mucus extract and for their potential biomedical application.
2.2.2. Characterisation of Mucus Fraction with MW > 20 kDa by Electrophoretic and MALDI-MS Analyses
The electrophoretic profile determined by 12%-SDS-PAGE of mucus fraction from C. aspersum with MW > 20 kDa (at a concentration 1.4 mg/mL) clearly shows various protein bands with MWs major in the region between 20–200 kDa. The highest expression was observed for proteins with MW at 17.6 kDa, 26.71 kDa, 29–32 kDa, 39.115, between 48–60 kDa, 97.0 kDa and over 100 kDa (Figure 5A). Proteins with MW at 20.5 kDa, 23.6 kDa, 29.1 kDa, 35.93 kDa, 70.89 kDa, and 84.77 kDa are also present in the composition of the fraction with MW >20 kDa, although with a lower expression.
A more accurate analysis of molecular masses and protein intensities in the mucus fraction with MW 20 kDa was obtained by ImageQuantTM TL v8.2.0 software, after scanning of 12% SDS-PAGE (Figure 5B,C). Based on the performed analysis, 18 protein bands were identified, most of which are with MW between 23–200 kDa. The obtained results presented in Figure 5C are in good agreement with the results of MALDI-Tof-MS analysis in the 20–80 kDa region of fresh extract from C. aspersum mucus [27].
2.2.3. Identification of Proteins in Mucus Fraction with MW > 20 kDa from C. aspersum Snail
Proteins in the mucus fraction with MW>20 kDa were determined after a performed search in the database UniProt (https://www.uniprot.org, 03 March 2024) for proteins in molluscs and gastropods, as well as in published data for mucus proteins in snails with MW corresponding to electrophoretic analysis (Figure 5C). Based on the conducted search, it was found that the protein presented at protein band 17.565 kDa well corresponded to mucus protein [C. aspersum, QEG59314] detected at 17.5 kDa in [24]. Detected proteins in the range 20–31 kDa probably belong to families of glutathione peroxidase (GPx) and glutathione transferases (GST) because several studies have confirmed the antioxidant properties (glutathione transferase activity) of C. aspersum mucus [25,26,27].
Results from the database UniProt show that in this region are detected proteins: glutathione peroxidases from Biomphalaria glabrata with MW 21.252 kDa (A0A2C9L5T6), MW 22.812 kDa (A0A2C9JW73) and MW 24.101 kDa (A0A2C9JBG3), peroxiredoxin-5 from P. canaliculata (A0A2T7NZ99 MW 20.751 kDa) and glutathione S-transferases with MW 27.978 kDa (from P. canaliculata, A0A2T7PWN7); with MW 27.599 kDa (from B. glabrata, A0A9U8DVA0); with MW 27.774 kDa (from Haliotis rubra, XM_025244070.1), and probable glutathione S-transferase with MW 23.092 kDa (from Aplysia californica, XM_035968272.1). The proteins with MW between 30–40 kDa are in good agreement with the identified proteins in the study [24], which suggests some of them are lectins and are also related to the antimicrobial properties of mucus. Recently, Cerullo et al. reported about a new protein class termed Conserved Anterior Mollusk Proteins (CAMPs) detected in the mucus of C. aspersum with MW<40 kDa by proteomic analysis on SDS-PAGE [32].
Some of the proteins expressed in protein bands between 48–59 kDa probably correspond to functional units or polypeptides of N-glycosylated C. aspersum hemocyanin resulting from proteolytic processes. The presence of different forms of hemocyanin in the mucus of land snails has been reported in several studies in recent years [21,29,30].
The protein at 84–85 kDa probably corresponded to epiphragmin, identified in the adhesive mucus secration of land snails H. aspersa, H. pomatia, and Cernuella virgata [40,41,42,43].
The electrophoretic analysis also showed proteins with MW above 100 kDa, which most likely represent different types of mucins, proteoglycans and collagen, which were confirmed in recent studies on mucus proteins of land snails C. aspersum [32,41], A. fulica [29,30] and H. lucorum [29] through integrative “omics” approach.
2.2.4. Characterisation of Mucus Fraction with MW > 20 kDa by Tandem Mass Spectrometrical Analyses of 12% SDS-PAGE
In order to identify some of these hypothesised proteins, the protein bands from 12% SDS-PAGE were excised from the gel. After trypsin digestion, the extracted peptides were analysed tandem mass spectrometry and bioinformatics. The amino acid sequences (AASs) of extracted peptides from each protein band were determined by MS/MS analyses because the identification of proteins only by Mascot search of the experimental peptide masses as [M+H]+ did not lead to satisfactory results.
Limited proteomic information on gastropods in the NCBI database probably caused this unsuccessful protein identification. In Figure 7, the MALDI-MS/MS spectrum of peptide [M+H]+ at m/z 1670.93 extracted from the protein band at 48.75±1.5 kDa is presented.
The determined amino acid sequence HGSPIGVPYWDWTR, shows 100% identity (E=2e-09) with hemocyanin alpha-N from C. aspersum (AYO86684.1).
In this way, the rest of the proteins were also determined (Table 2). Most of the presented hits demonstrate identities above 60% and E-values between 1×10−13 and 2 (Table 2, Supplementary Materials:Supplementary SI Supplementary 2). It is known that the lower the E-value, the more “significant” the match is, suggesting a higher probability that the sequences share a common evolutionary origin.
The results shown in Table 2 confirm the presence of mucus protein [QEG59314, from C. aspersum]; NADH dehydrogenase subunit 6 [Albinaria caerulea]; glutathione S-transferase omega-1 [P. canaliculata]; H-type lectins included H. aspersa agglutinin, mucus protein with MW 39.115 kDa named [QEG59312 from C. aspersum] which probably is homology von Willebrand factor A domain-containing protein 3B; Functional unit βC-d of C. aspersum hemocyanin, L-amino-acid oxidase-like protein (as Achacin from L. fulica), FMRFamide-activated amiloride-sensitive sodium channel, zinc finger protein, elastin-like protein, several typs collagen (collagen alpha-1, collagen α-4, collagen alpha-6) and mucins (mucin-5AC, mucin-5B, mucin-2, and mucin-17-like proteins). Most of the detected proteins can be associated with antimicrobial and antioxidant properties of this protein mucus fraction.
2.3. Antimicrobial Effect of Two Fractions Isolated from C. aspersum Mucus
The antibacterial activity of the investigated peptides and polypeptides in both mucus fractions with MW < 20 kDa and MW > 20 kDa from the snail C. aspersum was determined against different bacterial strains by minimum inhibitory concentration (MIC) analysis. Two strains of gram-positive pathogenic bacterial isolates (Bacillus cereus 1085, Propionibacterium acnes 1897) and three strains of gram-negative ones (Salmonella enterica 8691, Enterococcus faecalis 3915, Enterococcus faecium 8754) were used as test microorganisms. The obtained results showed a different degree of effectiveness of the investigated substances.
The MIC values presented in Figure 8a,b show a higher antimicrobial potential of the fraction with MW > 20 kDa of C. aspersum mucus compared to the fraction with MW < 20 kDa, inhibiting at concentrations of 32–128 µg/mL more of 90% of the microbial growth of Gram+ pathogenic bacterial isolates Bacillus cereus 1085 and Propionibacterium acnes 1897. Furthermore, the effect against Gram+ microorganisms of the bioactive compounds in this fraction was observed even at low concentrations (2–4 µg/mL), having values higher than the IC50.
The antimicrobial activity of the mucus fraction with MW >20 kDa against Gram-negative bacteria - Salmonella enterica 8691, Enterococcus faecalis 3915, Enterococcus faecium 8754 is also well expressed. Even at concentrations of 8 µg/mL, more than 70% inhibition of bacterial growth was achieved (Figure 9a,b). Analysis of the peptide fraction with MW <20 kDa from C. aspersum mucus also showed the presence of a well-expressed antibacterial effect (Figure 9a). Still, the reported IC50 values were lower than those of the protein fraction with MW>20 kDa. They range between IC50–IC80 against Gram+ bacteria and have a maximum IC70 against Gram─ strains at the highest mucus concentrations tested. The weakest effect was reported for Salmonella enterica 8691. Interestingly, the antimicrobial activity of both investigated fractions of C. aspersum mucus is comparable to that of the antibiotic vancomycin, but only when higher concentrations are used. When peptide and protein mucus fractions are added to the culture medium in concentrations 8 μg/mL, their effect is greatly reduced.
2.4. Studying the in Vitro and in Vivo Effects of Snail Mucus on Eukaryotic Cell
The yeast Saccharomyces cerevisiae has been used as a model system for eukaryotic cell biology to assess the effect of both isolated bioactive fractions of C. aspersum mucus on eukaryotic cell metabolism. A time-kill kinetics test was performed, which revealed that the viability of mucus-treated S. cerevisiae cells was not significantly different from the not-treated ones (Figure 10).
Even at the 24h hour of cultivation, some growth stimulation effect has been observed, showing a 3–30% increase in the cellular biomass when snail mucus has been added to the nutrient media. Moreover, the observed improvement in the cell growth did not depend significantly on either the added snail mucus concentration or the bioactive fraction type—with MW<20 kD or with MW>20 kDa.
Next, the possible cytotoxic effect of the isolated bioactive snail mucus fractions on the eukaryotic cell has been studied. The S. cerevisiae cells were treated with concentrations of both fractions exceeding those used in characterizing their antibacterial activity. Then, the alternation in the levels of intracellularly generated ROS and carbonylated proteins was evaluated (Figure 11). It was revealed that in the cells treated with 256 μg/mL snail mucus either with MW < 20 kDa or with MW>20 kDa, ROS generation was decreased by 29% to 38%, respectively, compared to the untreated cells (Figure 11a). Simultaneously, the intracellular levels of oxidized proteins were not significantly different between treated and untreated cells, ranging between 18.37 ± 0.98 and 19.06 ± 0.87 μmol/mg of protein (Figure 11b).
To evaluate the possible mechanism involved in the growth-stimulating and cytoprotective effect of C. aspersum snail extracts on S. cerevisiae eukaryotic cells, intracellular GSH levels and TAC were evaluated (Figure 12). It was shown that exposure to both mucus fractions with MW<20 kD and MW>20 kD leads to an increase in the cytoplasmic GSH level of 35% and 25%, respectively (Figure 12a). The same tendency was observed when TAC was assessed. The exposure of S. cerevisiae cells to the action of snail BACs causes an increase in the total antioxidant capacity of the cell (Figure 12b).
3. Discussion
The biologically active substances with antibacterial action from natural sources provide the prospect of obtaining a new type of antimicrobial agents—with reduced or even absent toxicity towards people but highly effective towards inhibition of the pathogenic microorganisms [18]. Most Gastropods have high potential as sources of antimicrobial peptides and proteins. Several studies demonstrated the presence of highly effective peptides and proteins in the mucus of terrestrial snails [19,20,21,22,23,24,30] as well as freshwater [41] and marine snails [17,18,45]. The mucus secretions of snails and slugs play a pivotal role in the survival and interaction of the animals with the environment. It is crucial to continue identifying the natural antimicrobial potential of snail mucus because the rise of drug-resistant bacterial species continues to develop. To date, there are still few studies on the antimicrobial properties of the mucus of terrestrial molluscs and most of them have focused on the land snail, A. fulica [24].
The data obtained in the present study showed a strong antibacterial effect of the two fractions from the mucus C. aspersum against 2 Gram-positive pathogenic bacterial stains (B. cereus 1085, P. acnes 1897) and 3 strains of Gram-negative ones (S. enterica 8691, E. faecalis 3915, E. faecium 8754). Results of the in vitro study showed that high concentrations (128 μg/mL and 64 μg/mL) of both fractions with MW < 20 kDa and MW > 20 kDa exhibited antimicrobial activity against tested pathogenic bacterial strains comparable to antibiotic VCM.
The obtained data are in accordance with the results of other researchers, who prove the presence of specific oligo and polypeptides with antibacterial action in the mucus of land snails [19,20,21,23,24,37]. Our previous studies showed that the fraction with MW < 20 kDa shows strong antibacterial activity against P. aureofaciens AP9 in deep inoculation of the bacterium, while the fraction with MW between 10–30 kDa exhibits the highest antibacterial activity using surface inoculation of bacterial strain E. coli NBIMCC 8785 [19]. A protein fraction with Mw >20 kDa was also found to be most effective against the bacterial strain C. perfringens in deep anaerobic cultivation [19]. The observed differences in the antimicrobial activities against the tested pathogens are due to the difference in the active components of the two fractions.
The characterization of both fractions showed that their antimicrobial properties are due to different biocomponents, mainly antimicrobial peptides and polypeptides in the fraction with MW < 20 kDa and proteins with potential antibacterial properties in the fraction with MW > 20 kDa.
The peptide fraction with MW<20 kDa from C. aspersum mucus contains mostly various peptides < 3 kD with antimicrobial and antioxidant properties as some metabolites such as amino acids, allantoin, glycolic acid, γGSH and others [46]. In the current study, 16 newly antimicrobial peptides (shown in Table 1) were identified, as well as several peptides with higher molecular weight between 3–10 kDa and some proteins detected as [M+H]+ at m/z between 10–20 kDa. The identified peptides (Table 1) contain high levels of glycine, leucine, valine, and proline amino acid residues, as well as the presence of tryptophan, aspartic acid, phenylalanine, and arginine, which are associated with their antimicrobial activity and antioxidant properties. Amino acid sequence alignment comparison of C. aspersum mucus peptides by (http://www.campsign.bicnirrh.res.in/blast.php) CAMPSing software revealed high identity (more than 70%) with known AMPs (SI1). The structural characteristics of the peptides with the highest prognostic antibacterial and antifungal activity (Nos 17, 18, 21, 22, 23, Table 1) show high levels of glycine, leucine, valine, and proline residues, indicating that they belong to a new class of antimicrobial peptides, rich in Gly and Gly/Leu, which demonstrate broad-spectrum antibacterial activity [47].
Previous studies have also shown the presence of peptides with similar amino acid sequences revealed homology with mostly Leptoglycin, Acanthoscurrin and Ctenidin [19,36,37,38]. In addition, most peptides with high predictive therapeutic values are cationic and/or neutral (with one or two positively charged residues that are neutralized with negatively charged ones). Most of the identified peptides (Table 1) are hydrophobic, which is a known prerequisite for their antimicrobial activity because hydrophobicity not only modulates the peptide-membrane interaction but also affects cellular selectivity and is positively correlated with cytotoxic activity [44,48]. AMPs are known to kill microbes through mechanisms that primarily involve interactions between the charged amino acid residues of the peptide and components of the bacterial membranes of the target cells. There are several mechanism models to explain the action of these peptides, including the toroidal pore model, the barrel model, and membrane interaction according to the Shai–Huang–Matsazuki carpet model [48,49,50,51]. These interactions can lead to a number of effects, including membrane permeabilization, depolarization, leakage or lysis, leading to cell death.
In the current study, in a fraction with MW <20 kDa, some peptides with higher molecular masses between 10–20 kDa were identified, which are in good agreement with similar proteins in snail mucus of C. aspersum [24,33], Helix pomatia [34] and A. fulica [35] and may also contribute to the antibacterial activity against the tested bacterial pathogens. For example, polypeptide determined at 12.464 kDa (Figure 2) is probably a lectin previously found in a study by Pietrzyk et al., in the mucus of C. aspersum [33]. A similar protein with biological activity against Streptococcus mutans and Aggregatibacter actinomycetemcomitans was also found in A. fulica mucus with 11.45 kDa with [52]. The protein detected by us at m/z 18.005 kDa in a fraction with MW < 20 kDa using MALDI-MS analysis (Figure 2) probably corresponds to the anti-Pseudomonas aeruginosa protein (MW 17.5 kDa) described in a study by Pitt et al. [24].
Several studies have shown that mucus from C. aspersum snail also demonstrates antimicrobial activity due to one or more proteins with molecular masses between 30 and 100 kDa [19,20,24,52]. Based on the results of the electrophoretic analysis and proteomic analysis on SDS-PAGE of a fraction with MW > 20 kDa, a high homology was found with some proteins with potential antibacterial activity. Surprisingly, a protein at 17.5 kDa was detected on SDS-PAGE analysis of the fraction with MW > 20 kDa. Alignment of AASs obtained by MALDI-TOF-MS/MS showed homology to mucus protein from C. aspersum snail (ID QEG59314.1), identified by Pitt et al., as a lectin with potential antimicrobial activity against strains of Ps. aeruginosa. Some lectins have pathogen-recognition receptor properties, which may contribute to their antimicrobial activity. Natural lectins have powerful antibacterial properties because they bind to carbohydrates on microbial surfaces [53].
The results of the proteomic analysis show that peptides extracted from the protein band at 26 kDa are homologous on H-type lectin from B. glabrata and agglutinin from H. aspersa (HAA) and H. pomatia (HPA), which also play a role in the defense of the snail against pathogens. H-type lectins represent a new group of lectins that have been identified in invertebrates. In snails, H-type lectins (eg, HPA, HAA) protect fertilized eggs from bacteria because they are secreted by snail protein glands as a component of the perivitelline fluid [54,55]. Lectins are involved in many biological processes, including cell recognition, viral and bacterial pathogenesis, and inflammation [54]. Some studies have shown that high molecular weight lectin isolated from the mucus of the giant African snail A. fulica induces agglutination of Gram+ and Gram─ bacteria, although it does not inhibit bacterial growth [56]. In addition, H-type lectins recognize glycans present in the cell wall of pathogenic bacteria, for example group C Streptococcus sp. Agglutinins are glycoproteins in nature and play a fundamental role in the innate immune responses in invertebrates [57].
AASs of several peptides extracted from the protein band at 39.115 kDa (Table 3) show high homology with antibacterial mucus protein with MW ~37.5 kDa (QEG59312.1, C. aspersum) named ‘Aspernin’, active against different strains of Ps. aeruginosa [24]. According Pitt at al., the protein ‘Aspernin’ shares homology with proteins in another species of snail—Biomphalaria glabrata, such as H-type lectin domain-containing protein [B. glabrata] [24]. Our results also showed that some peptides derived from this protein band matched domains of the von Willebrand factor A superfamily. The proteins with MW between 30 and 40 kDa found in the fraction with MW > 20 kDa are in full agreement with the recently detected proteins by Pitt et al. that suggest that these proteins are lectins and are also related to the antimicrobial properties of the mucus [24].
Another identified protein in mucus fraction with MW>20 kDa with antibacterial activity is hemocyanins. Our results (presented in Figure 5 and Table 2) revealed that the intensive band in the range of 48.75±1.5 kDa and 97.0±1 kDa is composed of proteins demonstrating high homology to hemocyanin functional units and subunits. The high protein expression of different forms of hemocyanin can explain observed antibacterial activity against tested bacterial pathogens (Figure 8b1,b2 and Figure 9b1,b2,b3). Previous studies have also demonstrated that molluscan hemocyanin possesses antibiotic activity [58,59,60], which supports our hypothesis. Interest in hemocyanin as a bioactive molecule against pathogenic microorganisms is relatively recent [58,61,62]. Information about the antibiofilm and antiviral activity of hemocyanin from land snails is still very scarce. [21]. Moreover, hemocyanin-derived polypeptides (detected at 40, 60 and 80 kDa) with possible antimicrobial properties were previously reported by Suárez et al. [21]. Furthermore, Dolashka et al., 2016 reported the antimicrobial activity of the βc-HaH structural subunit of H. aspersa (also named C. aspersum), which demonstrated strong selective antimicrobial activity against S. aureus, Streptococcus epidermidis and Escherichia coli, while Ps. aeruginosa and E. faecium developed unconditionally with his presence [16]. Therefore, the observed antibacterial activity of fractions with MW> 20 kDa against B. cereus 1085, P. acnes 1897, S. enterica 8691, E. faecalis 3915, and E. faecium 8754, cannot be fully explained only by the identification of hemocyanin.
In addition, on the protein band at 52.89±1.0 kDa, another protein was found homological with the C. aspersum mucus protein (QEG59312.1) and an unnamed protein from Candidula unifasciata snail (CAG5131631.1 and CAG5131621.1). This protein is likely related to the antimicrobial activity of fraction with MW > 20 kDa. The study by Pitt et al., [24] also mentioned a protein with MW at ~50 kDa with potential antimicrobial activity against Ps. aeruginosa. Furthermore, we found that the protein detected at the range 56.94–59.04 kDa shared homology with proteins with L-amino acid oxidase activity, such as Achacin, which is established as an antibacterial N-glycosylated protein in the mucus of snails L. fulica, A. fulica. Several studies have associated the broad spectrum of antimicrobial properties of the mucus of the giant African snail A. fulica against Gram+ bacteria (Bacillus subtilis and Staphylococcus aureus), and Gram─ strains (Escherichia coli and Ps. aeruginosa) mainly with the presence of Achacin.
In another study [21], it was presented that the activity against the biofilm-forming bacteria S. aureus was found to be mainly due to the action of proteins hemocyanin and ‘Achacin’ and their fragments resulting from proteolysis processes. Indeed, Achacin, in addition to inhibiting bacterial growth, also appears to attack bacterial plasma membranes [31]. It was found that the antibacterial activity of Achacin depends on the production of H2O2, which is obtained from the oxidative deamination reaction. These data indicate that Achacin can also attack pathogens during other growth phases by increasing the local H2O2 concentration to not harm neighbouring host cells [30,63].
Extracellular matrix proteins constitute 40–50% of the mucus proteins of C. aspersum [30]. Our results have proven the presence of homologous proteins on mucin-5AC, mucin-5B, mucin-2, and mucin 17 like proteins, as well as collagens. Mucin proteins, included in snail mucus, can play roles for several biological functions, including adhesion, lubrication, and microbial protection [64,65]. Mucins are among the largest macromolecules, characterized by protein core and branching glycan chain. The structural diversity of mucins allows for their extensive biological diversity and unique physical characteristics. A tandem repeat domain located in the centre of the protein backbone, rich in serine, threonine, and proline, serves as an anchor for glycosylation. Mucin glycans are predominantly O-linked, but minor amounts of N-linked glycans also can be present [64,66]. It appears that mucins, which are an important component of snail mucus, play an important role as the first line of defense against bacterial infections. However, how the glycoprotein mucin—which makes up the mucus—interacts with the bacteria is still not fully understood [67]. For example, the MUC2 mucin established in the mucus fraction, but also found in the human colon, is capable of aggregating Gram+ bacteria such as Bacillus subtilis and E. faecalis by binding to peptidoglycan present in the cell wall, thereby inhibiting epithelial cell penetration. Furthermore, both mucins MUC5AC and MUC5B have been shown to inhibit up to 85% Pseudomonas aeruginosa isolated from sputum of patients with chronic infection [67]. Antimicrobial potentials of mucus mucins from different species of giant African land snails on some typed culture pathogenic bacteria were studied by Okeniyi et al., 2022 [68]. It was found that snail mucus extract had antimicrobial effects on Gram+ and Gram─ bacteria, but mucus mucins seem to lose its antibacterial potential with time [68]. A. marginata’s mucus secretions had a stronger antibacterial activity against B. subtilis when compared to mucus from A. achatina and A. fulica [22].
Moreover, the performed study demonstrates that both isolated C. aspersum mucus fractions with MW < 20 kD and MW > 20 kDa are not cytotoxic to eukaryotic cells but are characterized by growth-promoting potential. Bearing in mind that microorganisms and various Mollusca frequently form symbiotic relationships that are crucial to the ecology and evolution of species [69], it could be suggested that mucus from C. aspersum is used from S. cerevisiae as a supplementary substrate for its growth. Adding both mucus fractions to the nutrient media further leads to a substantial reduction in ROS levels. ROS generated during cell growth are involved in the roughly 60 distinct pathways that lead to protein oxidation, which includes carbonylation causing unbalanced cellular proteome and malfunctions [70]. Obtained results suggested that C. aspersum mucus possesses ROS scavenging properties, which automatically reflects their ability to reduce the level of carbonylated proteins in the eukaryotic organisms while combating the pathogenic bacterial strains. Thus, the two isolated bioactive fractions from C. aspersum can not only be used effectively to treat antibiotic-resistant pathogens, but their administration also has additional beneficial effects related to the reduction of oxidative stress in the host organism’s cells. In fact, other authors confirmed the radical scavenging activity of snail mucus, showing that P. canaliculata and L. fulica mucus have pro-oxidant capacity [71]. Furthermore, the same authors observed that the L. fulica mucus exhibited higher antioxidant capacity similar to those we found for the C. aspersum mucus low and high molecular weight mucus fractions. According to Wang et al. (2010) [72], low molecular weight components found in snail mucus, including uronic acid, phenolic compounds, vitamins C and E, and uric acid, determine their antioxidant qualities.
The specific characteristics of the many mucus peptides, such as their hydrophobicity, molecular mass, and AAS, are also related to its antioxidant capacity [73]. Several studies in recent years have confirmed the antioxidant properties of a C. aspersum mucus, and a fraction with MW > 20 kDa included antioxidant superoxide dismutase and glutathione transferase activity [25,26,27]. Antioxidants are substances that can protect cells from damage caused by unstable molecules known as free radicals or reactive oxygen species.
The study of the chemical composition of C. aspesum mucus fractions further confirmed that its antibacterial and cytoprotective properties are probably due to the complex action of the specific compounds found in it. The presence of specific innate antimicrobial peptides, as well as of antibacterial proteins such as mucus protein QEG59314.1 and QEG59312.1, different forms of hemocyanin, which serve as a precursor of broad-spectrum antimicrobial agents and microbial agglutination, determines the high antibacterial activity of the isolated bioactive fractions [59]. The demonstrated strong overall antioxidant activity in treated eukaryotic cells could be attributed to the role of both glutathione S-transferase and NADH dehydrogenase-like proteins found in the snail mucus.
Our study reveals that C. aspersum mucus has the potential to act as a source of antibacterial agents against bacterial pathogens B. cereus 1085, P. acnes 1897, S. enterica 8691, E. faecalis 3915, E. faecium 8754, for the first time. Both mucus fractions, demonstrated antimicrobial activity and antioxidant properties, may become an alternative agent to expensive synthetic antibacterial compounds. Application of a fraction with MW > 20 kDa showed promising antimicrobial activity comparable to Vancomycin and without adverse side effects. However, further studies are needed to use mucus secretion in solving the problem of antibiotic resistance in bacterial pathogens.
4. Materials and Methods
4.1. Preparation of Mucus Extract
The mucus extract was prepared from crude mucus collected from snail C. aspersum, grown in Bulgarian eco-farms using patented technology, so the snails survived without disturbing their biological functions [19]. After several filtration steps (also an object of patented technology), the native mucus extract was obtained. The protein concentration in the native mucus extract was determined by Bradford assay [74]. After ultrafiltration using a membrane filter from 20 kDa, the crude mucus extract was separated into two fractions: a peptide fraction with MW below 20 kDa and a fraction containing compounds with MW above 20 kDa.
The peptide fraction with MW below 20 kDa was additionally separated into three fractions using Amicon® Ultra-15 centrifugal tube filters with 3 and 10 kDa membranes. Finally, the following samples were obtained: Sample 1— a fraction with compounds of MW <3 kDa; Sample 2—a fraction with compounds of MW 3–10 kDa; Sample 3 fraction with compounds of Mw 10–200 kDa
4.2. HPLC Purification of Peptides from the Fraction Below 20 kDa
Reverse-phase high-performance liquid chromatography (RP-HPLC) on ZURA®; HPLC system (KNAUER, Berlin, Germany) was used to purify mucus fraction with MW below 20 kDa after lyophilization on a Nucleosil C18 column equilibrated with 0.1% trifluoroacetic acid (TFA, v/v) (solution A). Elution was performed by a gradient of water–acetonitrile formed by solutions A (0.1% TFA/water) and solution B (100% acetonitrile in 0.1% TFA (v/v)) at a flow rate of 1.0 mL/min, for 70 min as follows: to 10 min., A 100%; 11- 20 min., A 90%; 21- 50 min., A 40%; 51–60 min., A 0%; 61–63 min., A 0%; and 64–70 min., A 100%. Ultraviolet absorption was monitored at 216 nm.
The eluted fractions were collected and dried by vacuum concentration Speed-110 vac. The fractions were reconstituted in Milli Q water containing 0.10% TFA (v/v).
4.3. Molecular Mass Analysis and De Novo Sequencing of Peptides by Mass Spectrometry
The isolated peptide fraction with MW <3 kDa (Sample 1) was lyophilized and analysed by MALDI-TOF-TOF mass spectrometry on an AutoflexTM III. High Performance MALDI-TOF& TOF/TOF System (Bruker Daltonics, Bremen, Germany), which uses a 200 Hz frequency-tripled Nd–YAG laser operating at a wavelength of 355 nm. Analysis was carried out after mixing 2.0 μL of the sample with 2.0 μL of matrix solution (7 mg/mL of α-cyano-4-hydroxycinnamic acid (CHCA) in 50% ACN containing 0.1% TFA), but only 1.0 μL of the mixture was spotted on a stainless steel 192-well target plate. The samples were allowed to dry at room temperature before being analyzed. A total of 3500 shots were acquired in the MS mode, and a collision energy of 4200 was applied. The mixture of angiotensin I, Glu-1-fibrinopeptide B, ACTH (1–17), and ACTH was used for calibration of the mass spectrometer.
The MS/MS spectra were carried out in reflector mode with external calibration using fragments of Glu-fibrino-peptide B. Amino acid sequences of peptides were identified by precursor ion fragmentation using MALDI-MS/MS analysis.
4.4. Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
The eluted intravenous immunoglobulins were analyzed by 1D-sodium dodecyl sulphate-polyacrylamide gel electrophoresis (1D-SDS-PAGE) using 5% stacking gel and 12% resolving gel, according to the Laemmli method with modifications [75]. The samples were dissolved in Laemmli sample buffer containing 10 mM DTT as reducing agent. Staining Coomassie Brilliant Blue G-250 buffer was used for the visualization of proteins on the gel as well as a protein standard marker—mixture of proteins with molecular weight from 6.5 kDa to 200 kDa (SigmaMarkerTM, Sigma-Aldrich, Saint Louis, MO, USA).
4.5. Image Analysis of 10% SDS-PAGE by ImageQuant™ TL v8.2.0 Software
The obtained polyacrylamide gel (PAG) was captured on an Image Scanner III (GE Healthcare) and the image was opened within the ‘1D gel analysis’ utility of the Image Quant TL v8.2 software (GE Healthcare Bio-Sciences AB, Uppsala Sweden), which is highly automated and easy-to-use image analysis software. The background was performed through the “image rectangle” setting to compensate for the intensity of the image background. All bands were identified manually, including those in the standard protein marker, with a pen tool, as the bandwidth was fixed at 54 pixels.
The molecular weight analysis of each band was performed using the protein data standard SigmaMarkerTM (Sigma-Aldrich, Saint Louis, MO, USA) in the range of 6500–200,000 Da. Automatically, horizontal bands were drawn to the individual bands of the MW marker and calculated with the cubic curve spline.
Based on the pre-calculated amount of bands at the marker, the amount of bands tested is determined [76].
4.6. Tryptic In-Gel Digestion and Peptide Extraction
Protease digestion was run according to the work of Rosenfeld et al. with a slight modification [18]. The target protein bands excised from the SDS-PAGE gels were washed twice with a 150 µL mixture of 50% acetonitrile (ACN) and 200 mM NH4HCO3 each for 20 min at 30 °C to decolorize, as described previously in [77]. The digestion of proteins in gel was carried out with porcine trypsin (Promega, Madison, WI, USA). After drying the decolourized gels in the speed vac concentrator, a volume of 10 μL digestion buffer (50 mM ammonium bicarbonate, pH 7.8, containing modified trypsin) was added to them, and the Eppendorf tubes were kept on ice for 45 min to allow the gel pieces to be completely soaked with the protease solution.
Digestion was performed overnight at 37 °C, the supernatants were recovered, and the resulting peptides were extracted twice with 35 μL of 60% ACN/0.1% HCOOH. The extracts were pooled and dried in the speed vac concentrator.
4.7. Antibacterial Activity Assay
The antibacterial activity of the two bioactive fractions from C. aspersum was tested against 2 Gram-positive and 3 Gram-negative pathogenic bacteria listed below. It has been determined according to the procedure for the Minimum Inhibitory Concentrations (MICs) using the broth microdilution method according to Clinical Laboratory and Standards (CLSI) guidelines [78]. Briefly, the bacterial suspension cultured to the logarithmic phase was diluted to 0.5 McFarland Standard (approximately 1.5×108 CFU/mL) and then diluted 150 times to 1×106 CFU/mL using nutrient media. A 50 μL volume of undiluted and serial twofold dilutions of BACs with the Mueller Hinton broth (and Brain Heart Infusion broth for P. acnes) was dispensed in 96-well microtiter plates. Subsequently, an equal volume of adjusted inoculum (1×106 CFU/mL) was added to each well of the Microtiter plates up to a final volume of 100 μL. The nutrient media with bacterial culture without bioactive fractions was used as a negative control, and in the positive control, bioactive fractions were replaced by the glycopeptide antibiotic vancomycin (VCM)—(stock solution 40 mg/mL). The MIC value is accepted as the lowest concentration of snail BACs at which bacterial growth is completely inhibited.
| Bacteria strain | Causative agents |
| Bacillus cereus NBIMCC 1085 | Foodborne disease |
| Propionibacterium acnes DSM 1897 | Skin condition of acne, chronic endophthalmitis, corneal ulcers, sarcoidosis, etc. |
| Salmonella enterica NBIMCC 8691 | Salmonellosis |
| Enterococcus faecalis NBIMCC 3915 | Endocarditis, sepsis, urinary tract infections (UTIs), meningitis, etc. |
| Enterococcus faecium NBIMMCC 8754 | Bloodstream infections, urinary tract infections (UTIs), and wound infections associated with catheters or surgery are the most common infections. |
4.8. In vivo and In Vitro Effect on Eukaryotic Cell
The effect of both isolated fractions from the mucus of C. aspersum on the eukaryotic cell was evaluated using Saccharomyces cerevisiae as a model organism.
4.8.1. Cell Treatment and Viability Assay
Saccharomyces cerevisiae NBIMCC 584 were obtained from the National Bank for Industrial Microorganisms and Cell Cultures (Sofia, Bulgaria). The cells were cultured in a standard YPD medium containing 2% glucose, 1% yeast extract, and 1% peptone (pH 6.5) at 30 oC. A time-kill kinetics test has been performed, adding various concentrations of mucus fractions with MW<20 kD and MW>20 kD to the growth media (0, 64, 128, 256 μg/mL). Yeast growth was quantified after incubation for 0, 3, 6, 12 and 24 h. Absorbance was measured at 600 nm using a microplate reader.
For in vitro experiments, 1g S. cerevisiae biomass was treated with both mucus fractions at a final concentration of 256 μg/mL for 1 h at 25 oC, washed twice with distilled water and subjected to mechanical disruption according to the procedure described by [79]. Obtained cell-free extracts were further used for the biochemical analysis.
4.8.2. Measurement of the Mucus Cytotoxic Effect
The cytotoxic effects of the two mucus extracts were evaluated by measuring the alternations in the intracellular levels of ROS and carbonylated proteins. The generation of ROS in the S. cerevisiae cells was examined using the nitroblue tetrazolium test (NBT) method described by Kostova et al. (2008) [80]. The levels of oxidized proteins were assessed following the procedure of Mesquita et al. (2014) [81].
4.8.3. Determination of Cytoprotective Effect
5. Conclusions
In summary, our study reveals for the first time that C. aspersum mucus has the potential to act as a source of antibacterial agents against the bacterial pathogens—Bacillus cereus 1085, Propionibacterium acnes 1897, Salmonella enterica 8691, Enterococcus faecalis 3915 and Enterococcus faecium 8754. Both mucus fractions with MW <20 and MW>20 kDa, applied in the highest concentration of 128 μg/mL, demonstrated antimicrobial activity against tested pathogens that were comparable to Vancomycin. Also, the mucus fraction with MW>20 kDa showed promising activity even in concentrations between 8–2μg/mL. Furthermore, both mucus fractions are non-cytotoxic to eukaryotic cells and are characterized by growth-promoting potential and antioxidant properties. The study showed 16 novel peptides with MW between 1000–2900 Da identified by de novo sequencing in a mucus fraction with MW <20 kDa, with potential antibacterial activity. Based on the results of electrophoretic and proteome analysis of mucus fraction with MW >20 kDa, some proteins have been identified that demonstrate high homology to proteins with antibacterial potential, such as mucus protein called Aspernin, H-type lectins, hemocyanin, L-amino acid oxidase-like protein as well as mucins (mucin-5AC, mucin-5B, mucin-2 and mucin-17-like proteins) with MW > 100 kDa. Additionally, homology to glutathione S-transferase and NADH dehydrogenase was found.
These results present C. aspersum mucilage as a potent non-cytotoxic antibacterial agent—an alternative to conventional antibiotics while avoiding the development of antibiotic resistance.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Information 1 (SI1): An alignment of amino acid sequences of some identified peptides from C. aspersum mucus with known antimicrobial peptides with data base AMPs by CAMPSing, Matrix: BLOSUM 80 (http://www.campsign.bicnirrh.res.in/blast.php) and with proteins by BLAST (https://blast.ncbi.nlm.nih.gov); Supplementary Information 2 (SI2): An alignment of amino acid sequences of some identified peptides from protein bands of mucus fraction with MW>20 kDa with known proteins in Gastropoda (taxid:6448), land snail land snails (taxid:6527), and Mollusca (taxid:6447) with database non-redundant protein sequences (nr) and UniProtKB/Swiss-Prot, sequences algorithm blastp (protein-protein BLAST), filtered to match records with percent identity between 50% and 100% (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Funding
This research was carried out with the support of the project KP-06 PN61-8/2022 funded by the Bulgarian National Science Fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article and supplementary material.
Acknowledgments
Project BG05M2OP001-1.002-0019: ‘Clean Technologies for Sustainable Environment—Waters, Waste, Energy for a Circular Economy’.
Conflicts of Interest
The authors declare no conflicts of interest. The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analysis, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Garvey, M. Antimicrobial Peptides Demonstrate Activity against Resistant Bacterial Pathogens. Infect. Dis. Rep. 2023, 15, 454–469. [Google Scholar] [CrossRef] [PubMed]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Antibiotic Resistance: Moving From Individual Health Norms to Social Norms in One Health and Global Health. Front Microbiol. 2020, 11, 1914. [Google Scholar] [CrossRef] [PubMed]
- Final Report DRUG-RESISTANT INFECTIONS A Threat to Our Economic Future, March 2017, Jonas and World Bank Group Team, 2017.
- Walsh, T.R.; Gales, A.C.; Laxminarayan, R.; Dodd, P.C. Antimicrobial Resistance: Addressing a Global Threat to Humanity. PLoS Med 2023, 20, e1004264. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Orlek, A.; Anjum, M.F.; Mather, A.E.; Stoesser, N.; Walker, A.S. Factors associated with plasmid antibiotic resistance gene carriage revealed using large-scale multivariable analysis. Sci Rep. 2023, 13, 2500. [Google Scholar] [CrossRef] [PubMed]
- Akram, F.; Imtiaz, M.; ul Haq, I. Emergent crisis of antibiotic resistance: A silent pandemic threat to 21st century. Microb. Pathog. 2023, 174, 105923. [Google Scholar] [CrossRef] [PubMed]
- Aisenbrey, C.; Bechinger, B. Molecular packing of amphipathic peptides on the surface of lipid membranes. Langmuir 2014, 30, 10374–83. [Google Scholar] [CrossRef]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [PubMed]
- Garvey, M. Antimicrobial Peptides Demonstrate Activity against Resistant Bacterial Pathogens. Infect. Dis. Rep. 2023, 15, 454–469. [Google Scholar] [CrossRef] [PubMed]
- Pinilla, G.; Coronado, Y.T.; Chaves, G.; Muñoz, L.; Navarrete, J.; Salazar, L.M.; Taborda, C.P.; Muñoz, J.E. In Vitro Antifungal Activity of LL-37 Analogue Peptides against Candida spp. J. Fungi 2022, 7, 1173. [Google Scholar] [CrossRef]
- Tan, L.T.H.; Chan, K.G.; Pusparajah, P.; Lee, W.L.; Chuah, L.H.; Khan, T.M.; Goh, B.H. Targeting Membrane Lipid a Potential Cancer Cure? Front. Pharmacol. 2017, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Qiu, L.; Zhou, Z.; Song, L. Research progress on the mollusc immunity in China. Dev. Comp. Immunol. 2013, 39, 2–10. [Google Scholar] [CrossRef]
- Li, H.; Parisi, M.G.; Parrinello, N.; Cammarata, M.; Roch, P. Molluscan antimicrobial peptides, a review from activity-based evidences to computer-assisted sequences. Invertebr. Surviv. J. 2011, 8, 85–97. [Google Scholar]
- Smith, V.J.; Desbois, A.P.; Dyrynda, E.A. Conventional and unconventional antimicrobials from fish, marine invertebrates and micro-algae. Mar. Drugs 2010, 8, 1213–1262. [Google Scholar] [CrossRef] [PubMed]
- Dolashka, P.; Dolashki, A.; Velkova, L.; Stevanovic, S.; Molin, L.; Traldi, P.; Velikova, R.; Voelter, W. Bioactive compounds isolated from garden Snails. J. BioSci. Biotechnol. 2015, 147–155. [Google Scholar]
- Dolashka, P.; Dolashki, A.; Van Beeumen, J.; Floetenmeyer, M.; Velkova, L.; Stevanovic, S.; Voelter, W. Antimicrobial Activity of Molluscan Hemocyanins from Helix and Rapana Snails. Curr Pharm Biotechnol. 2016, 17, 263–70. [Google Scholar] [CrossRef]
- Kirilova, M.; Topalova, Y.; Velkova, L.; Dolashki, A.; Kaynarov, D.; Daskalova, E.; Zheleva, N. Antibacterial Action of Protein Fraction Isolated from Rapana venosa Hemolymph against Escherichia coli NBIMCC 8785. Pharmaceuticals 2024, 17, 68. [Google Scholar] [CrossRef]
- Dolashki, A. , Velkova L., Daskalova E., Zheleva N., Topalova Y., Atanasov V., Voelter W., Dolashka P. Antimicrobial Activities of Different Fractions from Mucus of the Garden Snail Cornu aspersum. Biomedicines 2020, 8, 315. [Google Scholar] [CrossRef]
- Pitt, S.; Graham, M.A.; Dedi, C.G.; Taylor-Harris, P.M.; Gunn, A. Antimicrobial properties of mucus from the brown garden snail Helix aspersa. Br. J. Biomed. Sci. 2015, 72, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Suárez, L.; Pereira, A.; Hidalgo, W.; Uribe, N. Antibacterial, Antibiofilm and Anti-Virulence Activity of Biactive Fractions from Mucus Secretion of Giant African Snail Achatina fulica against Staphylococcus aureus Strains. Antibiotics 2021, 10, 1548. [Google Scholar] [CrossRef]
- Etim, L.B.; Aleruchi, C.; Obande, G.A. Antibacterial Properties of Snail Mucus on Bacteria Isolated from Patients with Wound Infection. Br. Microbiol. Res. J. 2016, 11, 1–9. [Google Scholar] [CrossRef]
- Santana, W.; Melo, C.; Cardoso, J.; Pereira-Filho, R.; Rabelo, A.; Reis, F.; Albuquerque, R.L.C. Assessment of antimicrobial activity and healing potential of mucous secretion of Achatina fulica. Int. J. Morphol. 2012, 30, 365–373. [Google Scholar] [CrossRef]
- Pitt, S.J.; Hawthorne, J.A.; Garcia-Maya, M.; Alexandrovich, A.; Symonds, R.C.; Gunn, A. Identification and characterisation of anti-Pseudomonas aeruginosa proteins in mucus of the brown garden snail, Cornu aspersum. Br. J. Biomed. Sci. 2019, 76, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Brieva, A.; Philips, N.; Tejedor, R.; Guerrero, A.; Pivel, J.P.; Alonso-Lebrero, J.L.; Gonzalez, S. Molecular basis for the regenerative properties of a secretion of the mollusk Cryptomphalus aspersa. Ski. Pharmacol Physiol 2008, 21, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Kostadinova, N.; Voynikov, Y.; Dolashki, A.; Krumova, E.; Abrashev, R.; Kowalewski, D.; Stevanovic, S.; Velkova, L.; Velikova, R.; Dolashka, P. Antioxidative screening of fractions from the mucus of garden snail Cornu aspersum. Bul. Chem. Com. 2018, 50C, 176–183. [Google Scholar]
- Petrov, L.; Kachaunov, M.; Alexandrova, A.; Tsvetanova, E.; Georgieva, A.; Dolashki, A.; Velkova, L.; Dolashka, P. Snail Mucus Protective Effect on Ethanol-Induced Gastric Ulcers in Mice. Life 2022, 12, 1106. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Zhang, Z.; Li, G.; Sun, J.; Gu, T.; Ain, N.U.; Zhang, X.; Li, D. Extraction, structure, pharmacological activities and applications of polysaccharides and proteins isolated from snail mucus. Int J Biol Macromol. 2024, 258 Pt 1, 128878. [Google Scholar] [CrossRef]
- Deng, T.; Gao, D.; Song, X.; Zhou, Z.; Zhou, L.; Tao, M.; Jiang, Z.; Yang, L.; Luo, L.; Zhou, A.; Hu, L.; Qin, H.; Wu, M. A natural biological adhesive from snail mucus for wound repair. Nat Commun 2023, 14, 396. [Google Scholar] [CrossRef]
- Cilia, G.; Fratini, F. Antimicrobial properties of terrestrial snail and slug mucus. J Complement Integr Med. 2018, 15. [Google Scholar] [CrossRef]
- Ulagesan, S.; Kim, H.J. Antibacterial and Antifungal Activities of Proteins Extracted from Seven Different Snails. Appl. Sci. 2018, 8, 1362. [Google Scholar] [CrossRef]
- Cerullo, A.R.; McDermott, M.B.; Pepi, L.E.; et al. Comparative mucomic analysis of three functionally distinct Cornu aspersum Secretions. Nat Commun 2023, 14, 5361. [Google Scholar] [CrossRef] [PubMed]
- Pietrzyk, A.; Bujacz, A.; Mak, P.; Potempa, B.; Niedziela, T. Structural studies of Helix aspersa agglutinin complexed with GalNAc: a lectin that serves as a diagnostic tool. Int J Biol Macromol. 2015, 81, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Greistorfer, S.; Klepal, W.; Cyran, N.; Gugumuck, A.; Rudoll, L.; Suppan, J.; von Byern, J. Snail mucus − glandular origin and composition in Helix pomatia. Zoology 2017, 122, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Herluinus Mafranenda, D.N.H.; Kriswandini, I.L.; Arijani, R.E. Antimicrobial proteins of Snail mucus (Achatina fulica) against Streptococcus mutans and Aggregatibacter actinomycetemcomitans. Dent. J. 2014, 47, 31–36. [Google Scholar] [CrossRef]
- Dolashki, A.; Nissimova, A.; Daskalova, E.; Velkova, L.; Topalova, Y.; Hristova, P.; Traldi, P.; Voelter, W.; Dolashka, P. Structure and antibacterial activity of isolated peptides from the mucus of garden snail Cornu aspersum. Bulg. Chem. Commun. 2018, 50C, 195–200. [Google Scholar]
- Topalova, Y.; Belouhova, M.; Velkova, L.; Dolashki, A.; Zheleva, N.; Daskalova, E.; Kaynarov, D.; Voelter, W.; Dolashka, P. Effect and Mechanisms of Antibacterial Peptide Fraction from Mucus of C. aspersum against Escherichia coli NBIMCC 8785. Biomedicines 2022, 10, 672. [Google Scholar] [CrossRef] [PubMed]
- Velkova, L.; Nissimova, A.; Dolashki, A.; Daskalova, E.; Dolashka, P.; Topalova, Y. Glycine-rich peptides from C. aspersum snail with antibacterial activity. Bulg. Chem. Commun. 2018, 50, 169–175. [Google Scholar]
- Meher, P.; Sahu, T.; Saini, V.; Rao, A.R. Predicting antimicrobial peptides with improved accuracy by incorporating the compositional, physico-chemical and structural features into Chou’s general PseAAC. Sci. Rep. 2017, 7, 42362. [Google Scholar] [CrossRef] [PubMed]
- Pawlicki, J.M.; Pease, L.B.; Pierce, C.M.; Startz, T.P.; Zhang, Y.; Smith, A.M. The effect of molluscan glue proteins on gel mechanics. J. Exp. Biol. 2004, 207, 1127–1135. [Google Scholar] [CrossRef]
- Tachapuripunya, V.; Roytrakul, S.; Chumnanpuen, P.; E-kobon, T. Unveiling Putative Functions of Mucus Proteins and Their Tryptic Peptides in Seven Gastropod Species Using Comparative Proteomics and Machine Learning-Based Bioinformatics Predictions. Molecules 2021, 26, 3475. [Google Scholar] [CrossRef]
- Li, D.; Graham, L.D. Epiphragmin, the major protein of epiphragm mucus from the vineyard snail, Cernuella virgata. Comp. Biochem. Physiol. B: Biochem. Mol. Biol. 2007, 148, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Ballard, K.R.; Klein, A.H.; Hayes, R.A.; Wang, T.; Cummins, S.F. The protein and volatile components of trail mucus in the Common Garden Snail, Cornu aspersum. PLoS ONE 2021, 16, e0251565. [Google Scholar] [CrossRef]
- González García, M.; Rodríguez, A.; Alba, A.; Vázquez, A.A.; Morales Vicente, F.E.; Pérez-Erviti, J.; Spellerberg, B.; Stenger, S.; Grieshober, M.; Conzelmann, C.; et al. New Antibacterial Peptides from the Freshwater Mollusk Pomacea poeyana (Pilsbry, 1927). Biomolecules 2020, 10, 1473. [Google Scholar] [CrossRef] [PubMed]
- Ebou, A.; Koua, D.; Addablah, A.; Kakou-Ngazoa, S.; Dutertre, S. Combined Proteotranscriptomic-Based Strategy to Discover Novel Antimicrobial Peptides from Cone Snails. Biomedicines 2021, 9, 344. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, N.G.; Simova, S.D.; Dangalov, M.; Velkova, L.; Atanasov, V.; Dolashki, A.; Dolashka, P. An 1H NMR- and MS-Based Study of Metabolites Profiling of Garden Snail Helix aspersa Mucus. Metabolites 2020, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Sousa, J.C.; Berto, R.F.; Gois, E.A.; Fontenele-Cardi, N.C.; Honório-Júnior, J.E.; Konno, K.; Richardson, M.; Rocha, M.F.; Camargo, A.A.; Pimenta, D.C.; Cardi, B.A.; Carvalho, K.M. Leptoglycin: A new Glycine/Leucine-rich antimicrobial peptide isolated from the skin secretion of the South American frog Leptodactylus pentadactylus (Leptodactylidae). Toxicon 2009, 54, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Control of cell selectivity of antimicrobial peptides. Biochim Biophys Acta. 2009, 1788, 1687–92. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Raheem, N.; Straus, S.K. Mechanisms of Action for Antimicrobial Peptides with Antibacterial and Antibiofilm Functions. Front. Microbiol. 2019, 10, 2866. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef]
- Zhong, J.; Wang, W.; Yang, X.; Yan, X.; Liu, R. A novel cysteine-rich antimicrobial peptide from the mucus of the snail of Achatina fulica. Peptides 2013, 39, 1–5. [Google Scholar] [CrossRef]
- Breitenbach Barroso Coelho, L.C.; Marcelino Dos Santos Silva, P.; Felix de Oliveira, W.; de Moura, M.C.; Viana Pontual, E.; Soares Gomes, F.; Guedes Paiva, P.M.; Napoleão, T.H.; Dos Santos Correia, M.T. Lectins as antimicrobial agents. J Appl Microbiol. 2018, 125, 1238–1252. [Google Scholar] [CrossRef]
- Pietrzyk-Brzezinska, A.J.; Bujacz, A. H-type lectins - Structural characteristics and their applications in diagnostics, analytics and drug delivery. Int J Biol Macromol. 2020, 152, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Prokop, O.; Uhlenbruck, G.; Köhler, W. A new source of antibody-like substances having anti-blood group specificity: a discussion on the specificity of Helix agglutinins. Vox Sang. 1968, 14, 321–33. [Google Scholar] [CrossRef]
- Ito, S.; Shimizu, M.; Nagatsuka, M.; Kitajima, S.; Honda, M.; Tsuchiya, T.; Kanzawa, N. High molecular weight lectin isolated from the mucus of the giant African snail Achatina fulica. Biosci Biotechnol Biochem. 2011, 75, 20–5. [Google Scholar] [CrossRef]
- Maji, S.; Datta, U.; Lal Hembram, M. A new sperm agglutinin factor from marine snail Telescopium telescopium: An evaluation with goat (Capra hircus) cauda epididymal spermatozoa. Iran. J. Reprod. Med. 2010, 8, 10–17. [Google Scholar]
- 58 Coates, C.J.; Nairn, J. Diverse immune functions of hemocyanins. Dev. Comp. Immunol. 2014, 45, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Coates, C.J.; Costa-Paiva, E.M. Multifunctional Roles of Hemocyanins. Subcell Biochem. 2020, 94, 233–250. [Google Scholar] [CrossRef] [PubMed]
- Jeyachandran, S.; Chellapandian, H.; Park, K.; Kwak, I.S. Exploring the Antimicrobial Potential and Biofilm Inhibitory Properties of Hemocyanin from Hemifusus pugilinus (Born, 1778). Int J Mol Sci. 2023, 24, 11494. [Google Scholar] [CrossRef]
- Yang, S.; Huang, H.; Wang, F.; Aweya, J.J.; Zheng, Z.; Zhang, Y. Prediction and characterization of a novel hemocyanin-derived antimicrobial peptide from shrimp Litopenaeus vannamei. Amino Acids. 2018, 50, 995–1005. [Google Scholar] [CrossRef]
- Zhuang, J.; Coates, C.J.; Zhu, H.; Zhu, P.; Wu, Z.; Xie, L. Identification of candidate antimicrobial peptides derived from abalone hemocyanin. Dev. Comp. Immunol. 2015, 49, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Ehara, T.; Kitajima, S.; Kanzawa, N.; Tamiya, T.; Tsuchiya, T. Antimicrobial action of achacin is mediated by L-amino acid oxidase activity. FEBS Lett. 2002, 531, 509–512. [Google Scholar] [CrossRef] [PubMed]
- McDermott, M.; Cerullo, A.R.; Parziale, J.; Achrak, E.; Sultana, S.; Ferd, J.; Samad, S.; Deng, W.; Braunschweig, A.B.; Holford, M. Advancing discovery of snail mucins function and application. Front Bioeng Biotech. 2021, 9, 734023. [Google Scholar] [CrossRef]
- Rashad, M.; Sampò, S.; Cataldi, A.; Zara, S. Biological activities of gastropods secretions: snail and slug slime. Nat Prod Bioprospect 2023, 13, 42. [Google Scholar] [CrossRef]
- Corfield, A.P. Mucins: A Biologically Relevant Glycan Barrier in Mucosal protection. Biochim. Biophys. Acta 2015, 1850, 236–252. [Google Scholar] [CrossRef]
- Bakshani, C.R.; Morales-Garcia, A.L.; Althaus, M.; Wilcox, M.D.; Pearson, J.P.; Bythell, J.C.; Burgess, J.G. Evolutionary conservation of the antimicrobial function of mucus: a first defence against infection. NPJ Biofilms Microbiome 2018, 4, 14. [Google Scholar] [CrossRef]
- Okeniyi, F.A.; Oghenochuko, O.M.; Olawoye, S.O.; Animashahun, R.A.; Adeyonu, A.G.; Akpor, O.B. Antimicrobial potentials of mucus mucin from different species of giant African land snails on some typed culture pathogenic bacteria. Asian J Agric & Biol. 2022, 4, 202107294. [Google Scholar] [CrossRef]
- Zhukova, N.V.; Eliseikina, M.G.; Balakirev, E.S.; Ayala, F.J. Multiple bacterial partners in symbiosis with the nudibranch mollusk Rostanga alisae. Sci Rep. 2022, 12, 169. [Google Scholar] [CrossRef] [PubMed]
- Madian, A.G.; Regnier, F.E. Proteomic identification of carbonylated proteins and their oxidation sites. J Proteome Res. 2010, 9, 3766–80. [Google Scholar] [CrossRef]
- Phrompanya, P.; Suriyaruean, N.; Nantarat, N.; Saenphet, S.; Tragoolpua, Y.; Saenphet, K. Biological properties of mucus from land snails (Lissachatina fulica) and freshwater snails (Pomacea canaliculata) and histochemical study of mucous cells in their foot. Peer J. 2023, 11, e15827. [Google Scholar] [CrossRef]
- Wang, W.; Yi, J.; Ke, S.; Halmela, M. Gastropod biological fluid, method of making and refining and use. 2010. United States Patent Application 2010023 3111.
- Wang, L.; Yang, J.; Wang, Y.; Zhang, J.; Gao, Y.; Yuan, J.; Su, A.; Ju, X. Study on antioxidant activity and amino acid analysis of rapeseed protein hydrolysates. Int. J. Food Prop. 2016, 19, 1899–1911. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nat. 1970, 227, 680–5. [Google Scholar] [CrossRef] [PubMed]
- Lincz, L.F.; Scorgie, F.E.; Garg, M.B.; Gilbert, J.; Sakoff, J.A. A simplified method to calculate telomere length from Southern blot images of terminal restriction fragment lengths. Biotechniques. 2020, 68, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, J.; Capdevielle, J.; Guillemot, J.C.; Ferrara, P. In-gel digestion of proteins for internal sequence analysis after one- or two-dimensional gel electrophoresis. Anal. Biochem. 1992, 203, 173–179. [Google Scholar] [CrossRef] [PubMed]
- CLSI M100 Performance Standards for Antimicrobial Susceptibility Testing; Approved Standard-34th edition; March 2024.
- Daskalova, A.; Petrova, V.; Velkova, L.; Kujumdzieva, A.; Tomova, A.; Voelter, W.; Dolashka, P. Investigation of protein expression of Saccharomyces cerevisiae cells in quiescent and proliferating state before and after toxic stress. Biotechnol. Biotechnol. Equip. 2021, 35, 366–376. [Google Scholar] [CrossRef]
- Kostova, I.; Traykova, M.; Rastogi, V.K. New lanthanide complexes with antioxidant activity. Medicinal Chemistry 2008, 4, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, C.S.; Oliveira, R.; Bento, F.; Geraldo, D.; Rodrigues, J.V.; Marcos, J.C. Simplified 2,4-dinitrophenylhydrazine spectrophotometric assay for quantification of carbonyls in oxidized proteins. Anal. Biochem. 2014, 485, 69–71. [Google Scholar] [CrossRef]
- Zhang, Y. Role of glutathione in the accumulation of anticarcinogenic isothiocyanates and their glutathione conjugates by murine hepatoma cells. Carcinogenesis 2000, 21, 1175–1182. [Google Scholar] [CrossRef]
- Kumaran, A.; Karunakaran, R.J. Anti-oxidant activity of polyphenols from Phyllanthus debilis Klein ex Willd. J. Nat. Remedies 2006, 6, 141–146. [Google Scholar]
Figure 1.
Purification of RP-HPLC of mucus fraction with MW<20 kDa on a Nucleosil C18 column (250 mm x 10 mm) by gradient of water–acetonitrile (with 0.1% TFA) for 70 min at a flow rate of 1.0 mL/min.
Figure 1.
Purification of RP-HPLC of mucus fraction with MW<20 kDa on a Nucleosil C18 column (250 mm x 10 mm) by gradient of water–acetonitrile (with 0.1% TFA) for 70 min at a flow rate of 1.0 mL/min.
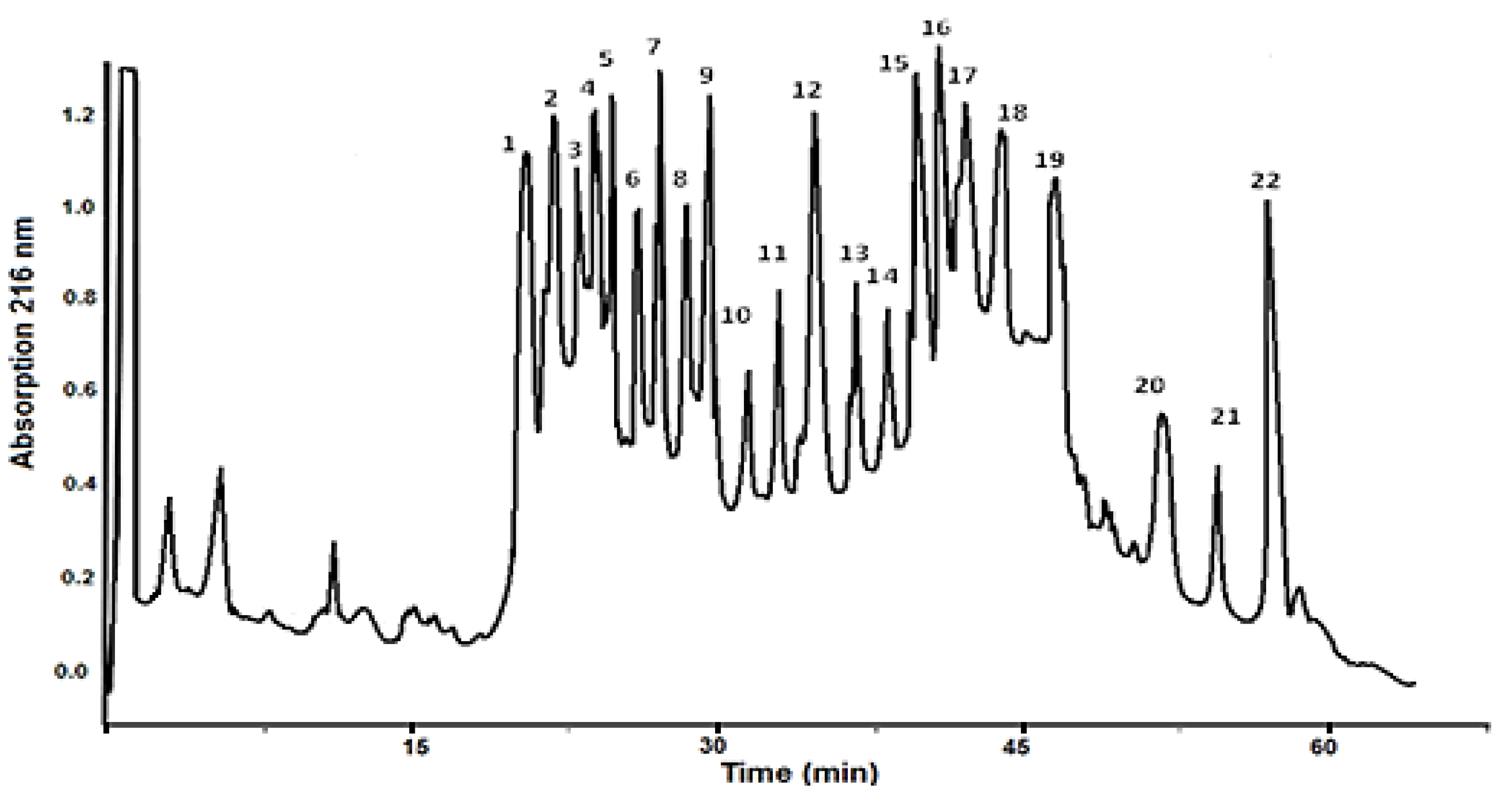
Figure 2.
MALDI-Tof-MS spectrum of a fraction with MW < 3 kDa. Standard peptide solution was used to calibrate the mass scale of the Autoflex™ III High-Performance MALDI-TOF and TOF/TOF Systems (Bruker Daltonics, Bremen, Germany).
Figure 2.
MALDI-Tof-MS spectrum of a fraction with MW < 3 kDa. Standard peptide solution was used to calibrate the mass scale of the Autoflex™ III High-Performance MALDI-TOF and TOF/TOF Systems (Bruker Daltonics, Bremen, Germany).
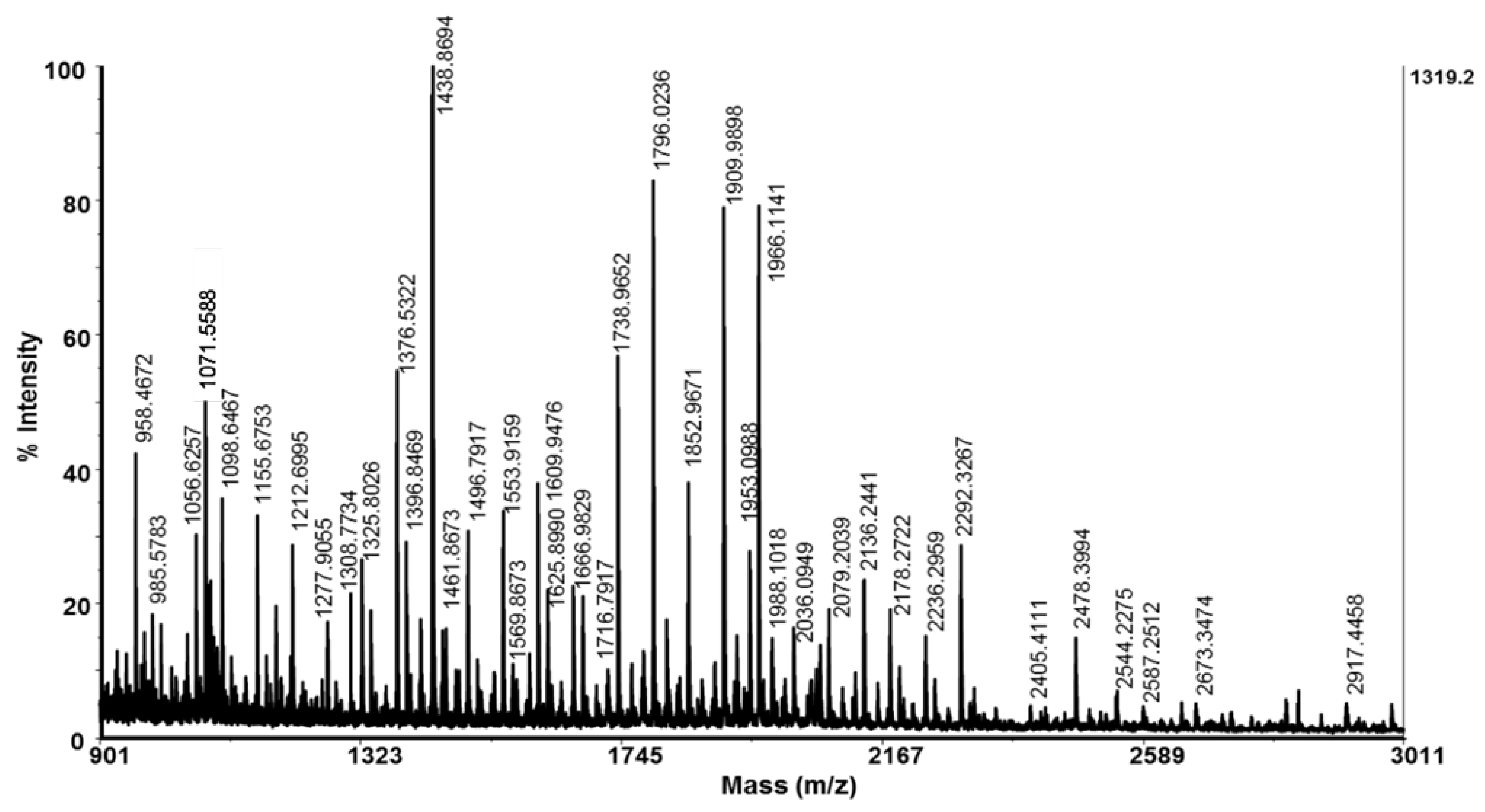
Figure 3.
MALDI-Tof-MS spectrum of fraction with MW<20 kDa recorded in range 3–20 kDa. Standard peptide solution was used to calibrate the mass scale of the Autoflex™ III High-Performance MALDI-TOF and TOF/TOF Systems (Bruker Daltonics, Bremen, Germany).
Figure 3.
MALDI-Tof-MS spectrum of fraction with MW<20 kDa recorded in range 3–20 kDa. Standard peptide solution was used to calibrate the mass scale of the Autoflex™ III High-Performance MALDI-TOF and TOF/TOF Systems (Bruker Daltonics, Bremen, Germany).
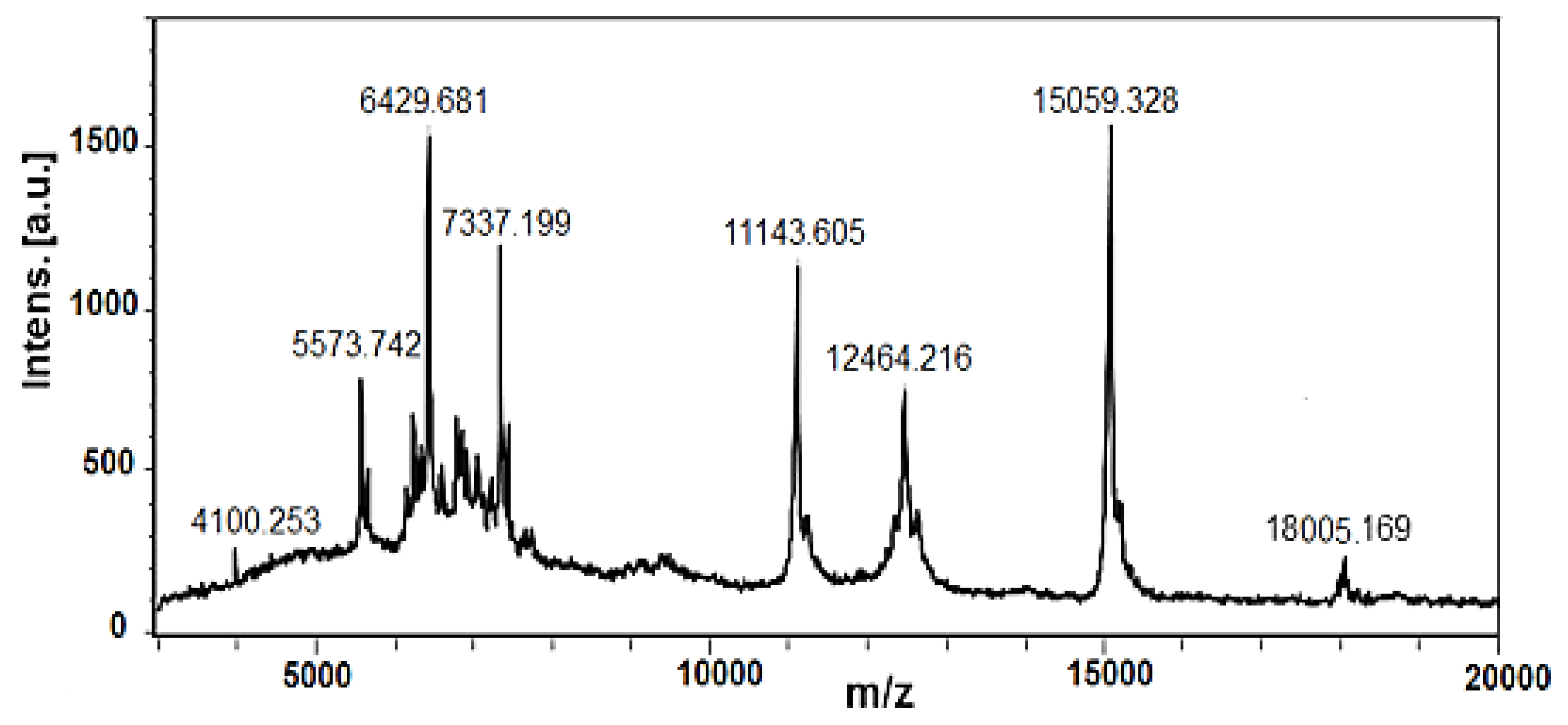
Figure 4.
MALDI-MS/MS spectrum of peptide [M+H]+ at m/z 1966.11Da and its amino acidic sequence LLLDNKGGGLVGGLLGGGGKGGG determined by de novo sequencing.
Figure 4.
MALDI-MS/MS spectrum of peptide [M+H]+ at m/z 1966.11Da and its amino acidic sequence LLLDNKGGGLVGGLLGGGGKGGG determined by de novo sequencing.
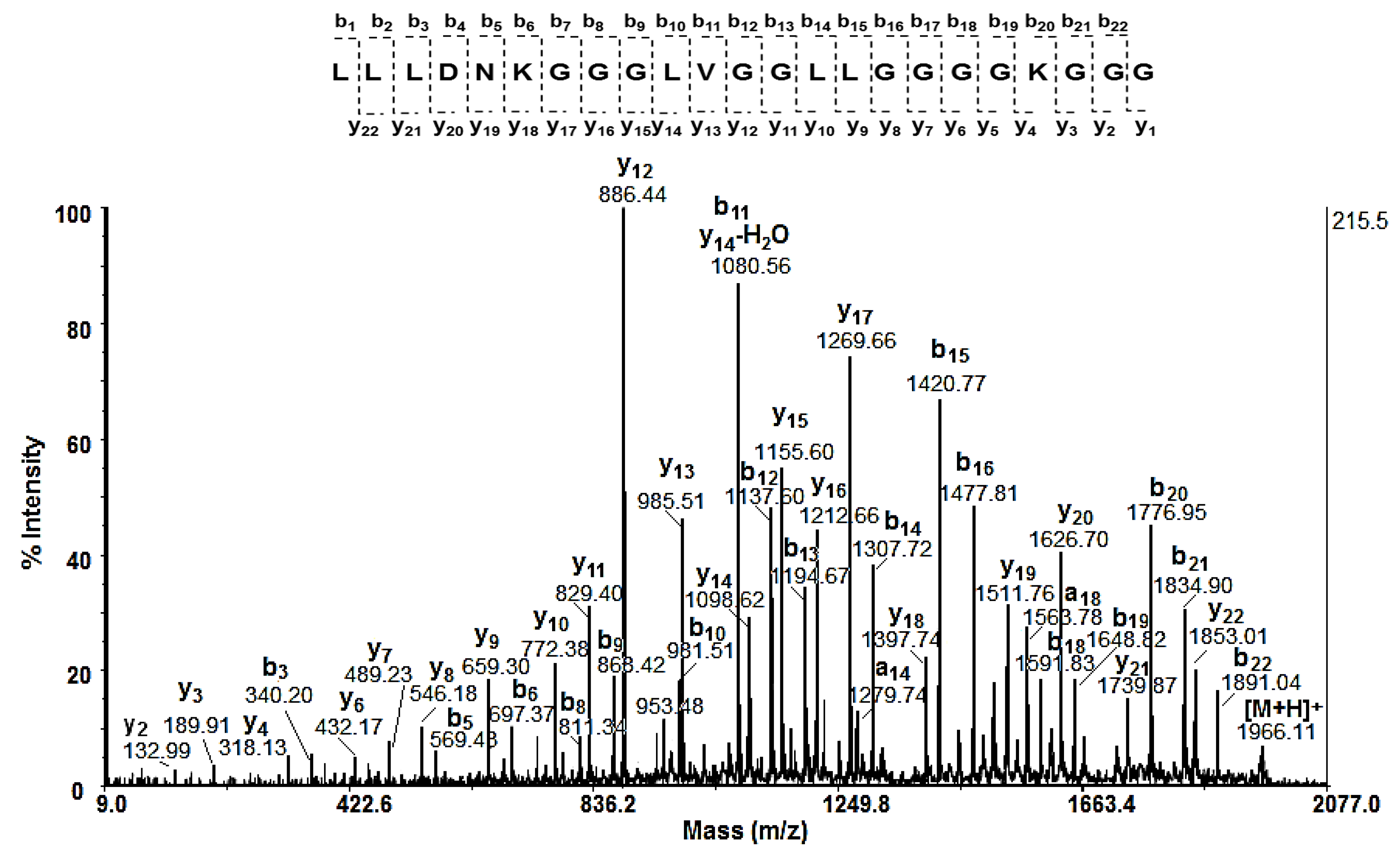
Figure 5.
Analys of molecular masses and protein intensities of 12% SDS-PAGE, scanned with a high resolution, using the ImageQuantTM TL v8.2.0 software. A) Electrophoretic pathway: 1) standard protein marker with MW between 200–6.5 kDa (SigmaMarkerTM, Sigma-Aldrich, Saint Louis, MO, USA); 2) mucus fraction from C. aspersum with MW>20 kDa; B) Electrophoretic profile of a standard protein molecular marker (electrophoretic Lane1) analyzed by ImageQuantTM TL; C) Analysis of the electrophoretic profile of mucus from C. aspersum with MW>20 kDa, using ImageQuantTM TL.
Figure 5.
Analys of molecular masses and protein intensities of 12% SDS-PAGE, scanned with a high resolution, using the ImageQuantTM TL v8.2.0 software. A) Electrophoretic pathway: 1) standard protein marker with MW between 200–6.5 kDa (SigmaMarkerTM, Sigma-Aldrich, Saint Louis, MO, USA); 2) mucus fraction from C. aspersum with MW>20 kDa; B) Electrophoretic profile of a standard protein molecular marker (electrophoretic Lane1) analyzed by ImageQuantTM TL; C) Analysis of the electrophoretic profile of mucus from C. aspersum with MW>20 kDa, using ImageQuantTM TL.
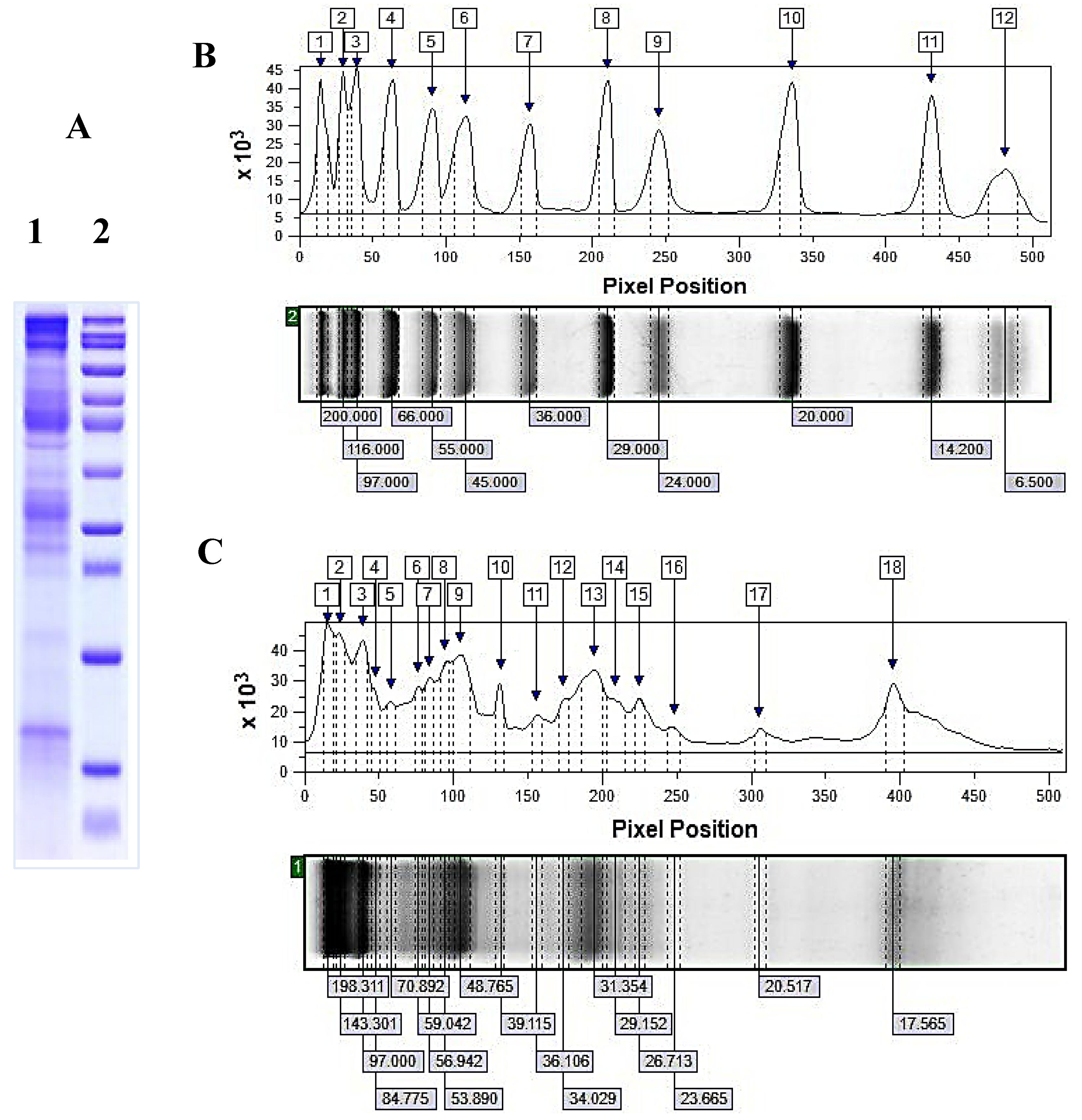
Figure 7.
MS/MS spectrum of peptide [M+H]+ at m/z 1670.93 from protein band at 48.75 kDa of fraction with MW >20 of C. aspersum mucus.
Figure 7.
MS/MS spectrum of peptide [M+H]+ at m/z 1670.93 from protein band at 48.75 kDa of fraction with MW >20 of C. aspersum mucus.
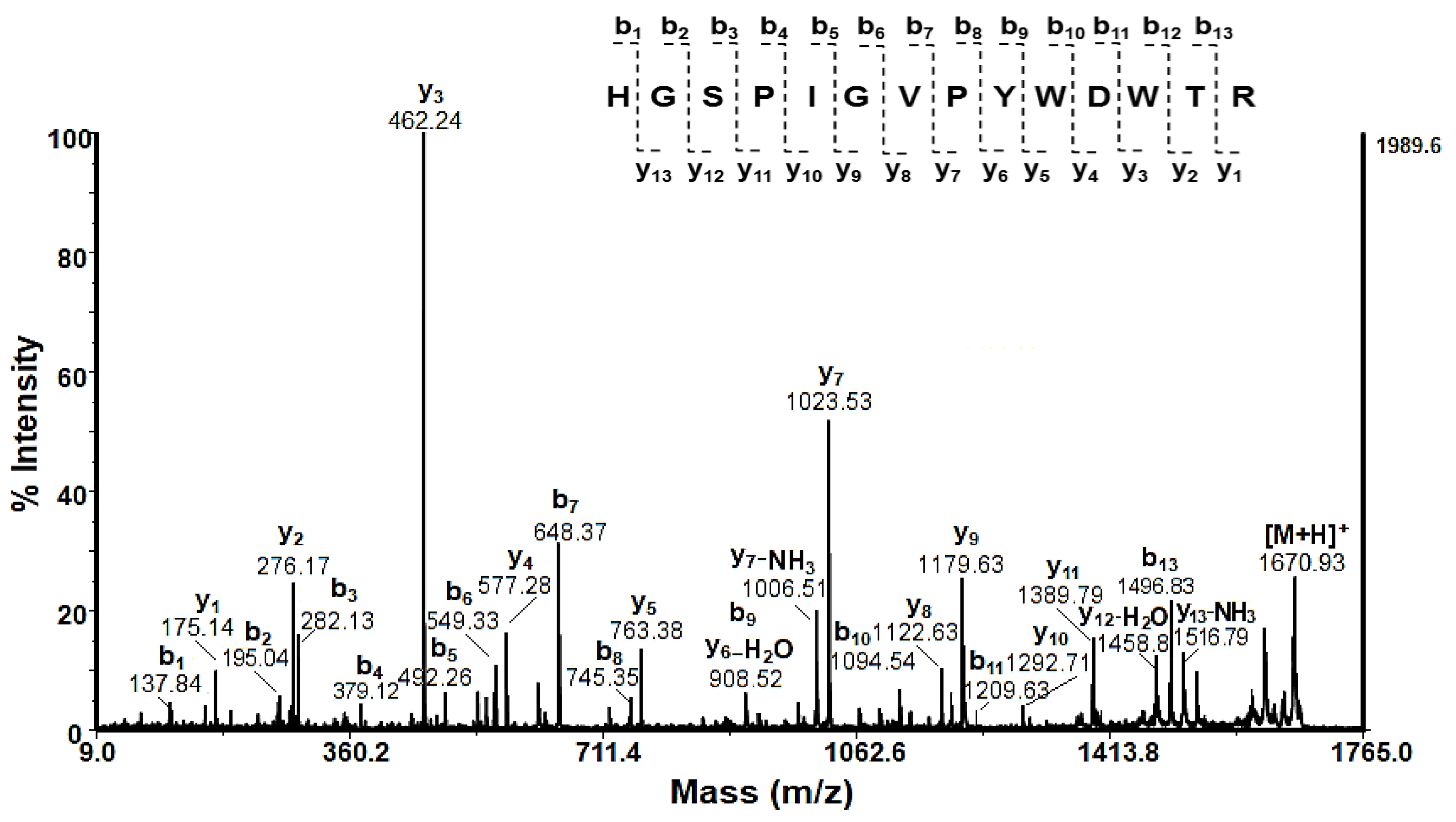
Figure 8.
Antibacterial activity of two mucus fractions from C. aspersum snail with MW < 20 kDa (a1, a2,) and MW > 20 kDa (b1, b2) against selected Gram-positive pathogenic strains. As a positive control the antibiotic Vancomycin (VCM) is used. The control (C) represents untreated cells. Treatments were carried out with concentrated (Conc) and diluted fractions of snail mucus.
Figure 8.
Antibacterial activity of two mucus fractions from C. aspersum snail with MW < 20 kDa (a1, a2,) and MW > 20 kDa (b1, b2) against selected Gram-positive pathogenic strains. As a positive control the antibiotic Vancomycin (VCM) is used. The control (C) represents untreated cells. Treatments were carried out with concentrated (Conc) and diluted fractions of snail mucus.
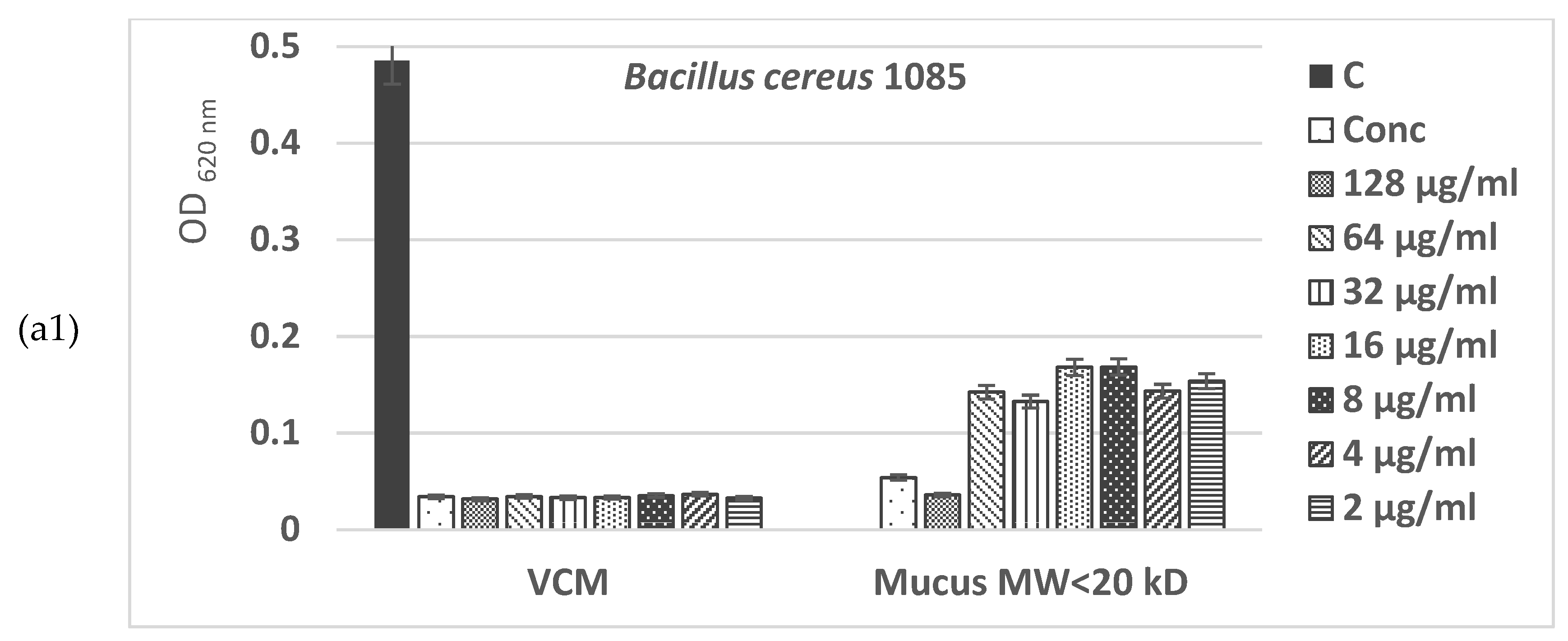
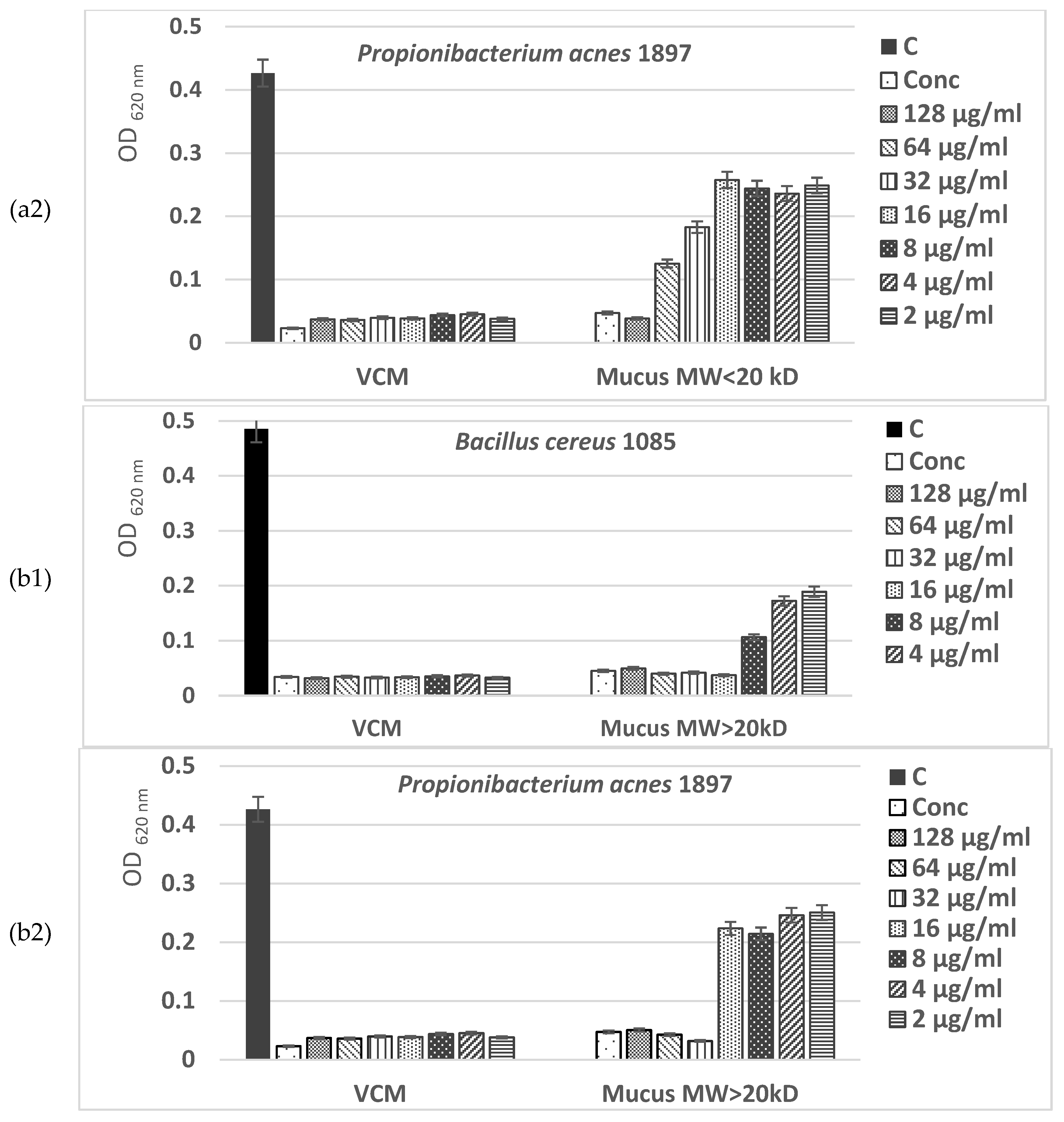
Figure 9.
Antibacterial activity of tow mucus fractions from C. aspersum snail with MW < 20 kDa (a) and MW > 20 kDa (b) against selected Gram (−) pathogenic strains. As a positive control the antibiotic Vancomycin (VCM) is used. The control (C) represents untreated cells. Treatments were carried out with concentrated (Conc) and diluted fractions of snail mucus.
Figure 9.
Antibacterial activity of tow mucus fractions from C. aspersum snail with MW < 20 kDa (a) and MW > 20 kDa (b) against selected Gram (−) pathogenic strains. As a positive control the antibiotic Vancomycin (VCM) is used. The control (C) represents untreated cells. Treatments were carried out with concentrated (Conc) and diluted fractions of snail mucus.
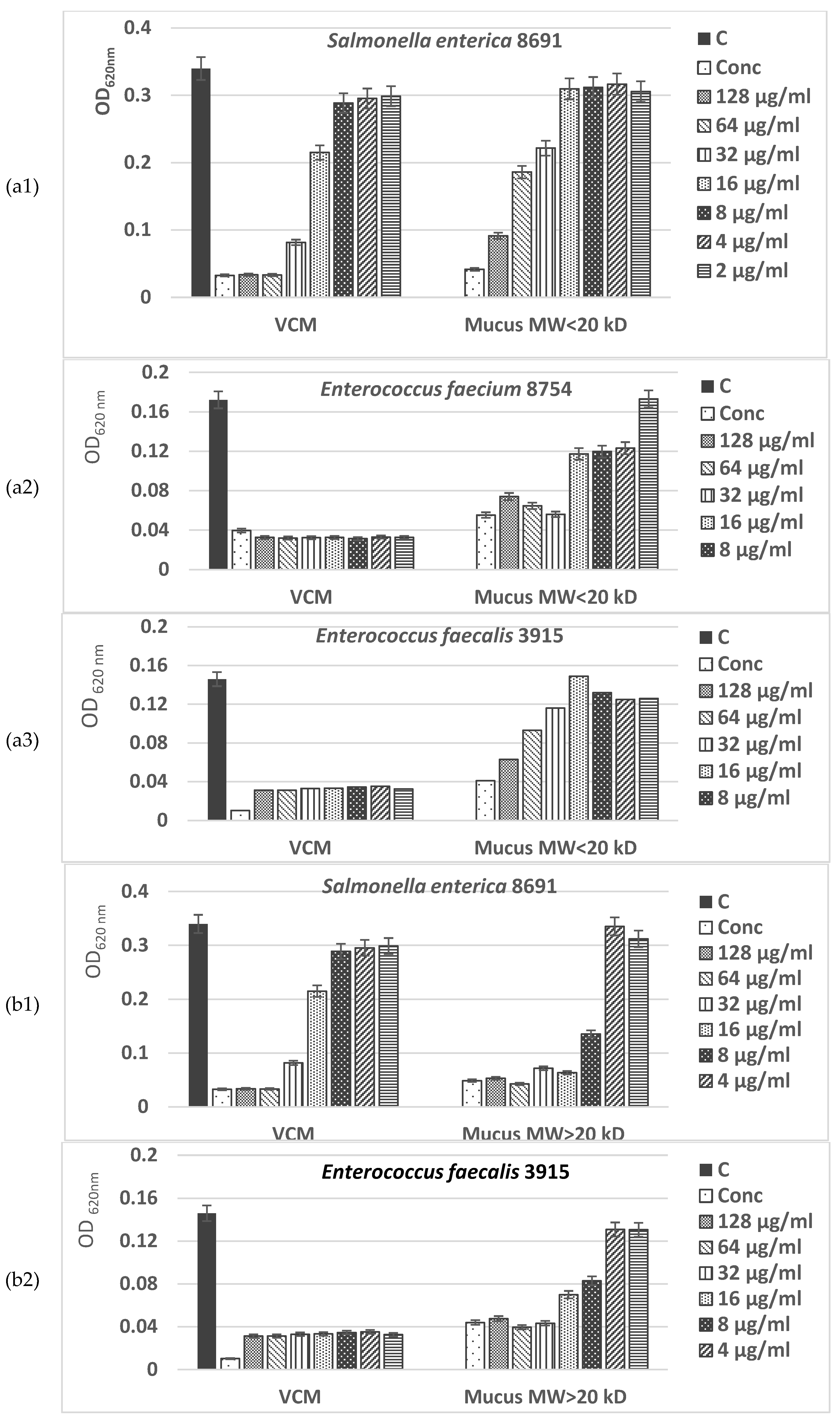
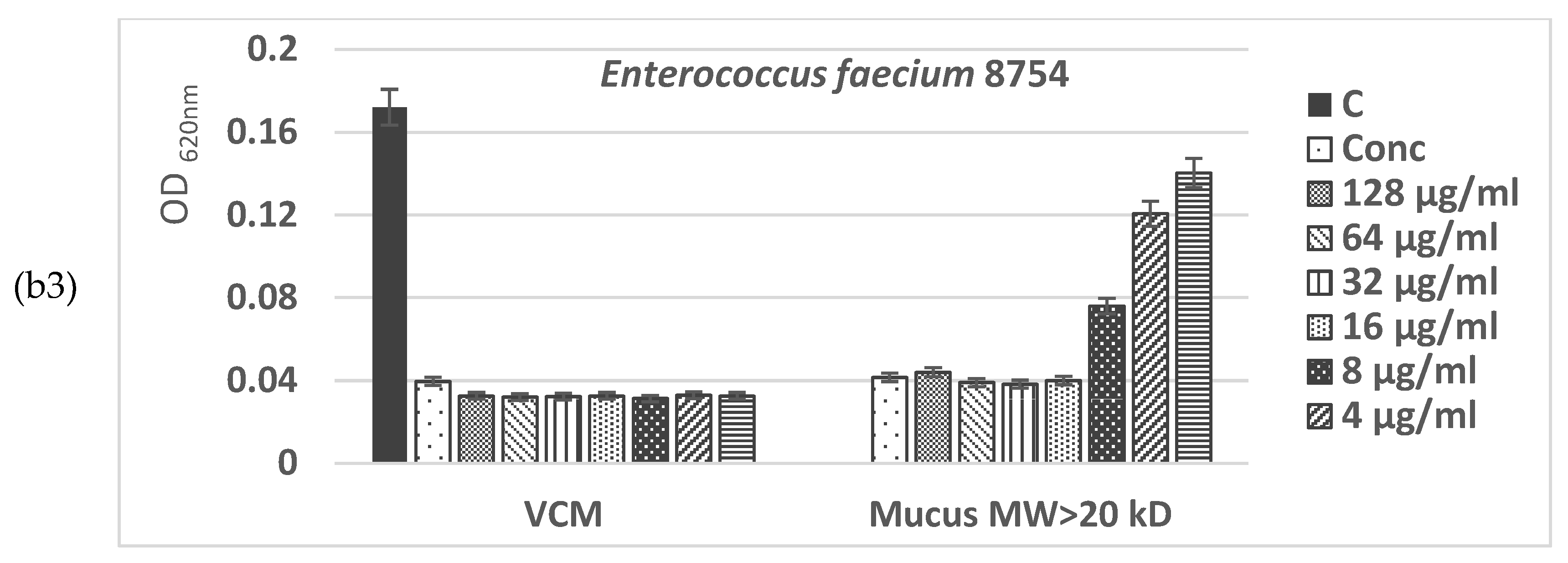
Figure 10.
Effect of C. aspersum mucus fraction with MW<20 kD (a) and MW>20 kD (b) on S. cerevisiae NBMCC 584 growth.
Figure 10.
Effect of C. aspersum mucus fraction with MW<20 kD (a) and MW>20 kD (b) on S. cerevisiae NBMCC 584 growth.
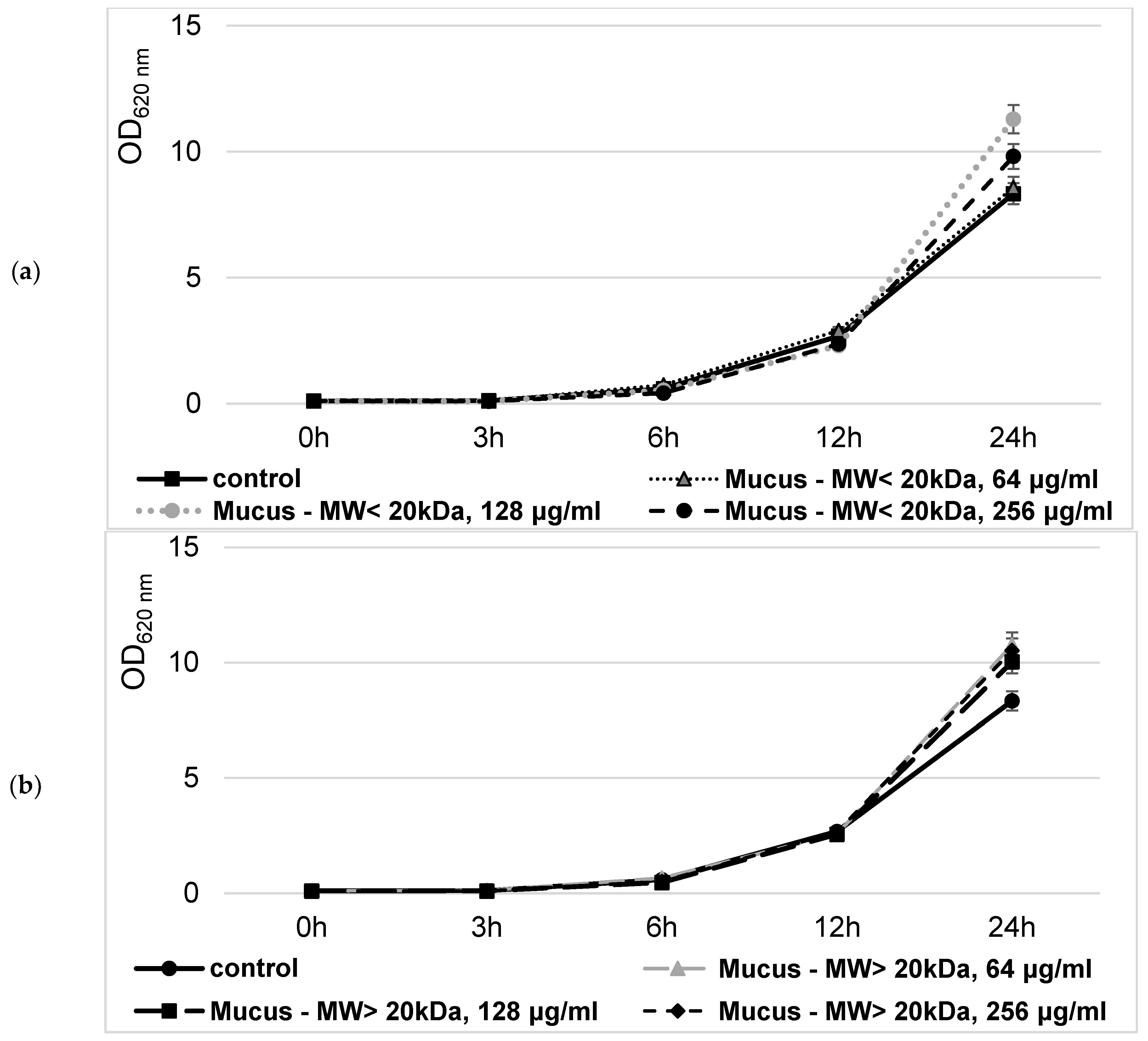
Figure 11.
Intracellular ROS (a) and carbonylated proteins’ (b) levels in S. cerevisiae NBMCC 584 cells grown in the presence of Cornu aspersum mucus fractions with MW<20 kD and MW>20 kDa.
Figure 11.
Intracellular ROS (a) and carbonylated proteins’ (b) levels in S. cerevisiae NBMCC 584 cells grown in the presence of Cornu aspersum mucus fractions with MW<20 kD and MW>20 kDa.
Figure 12.
Effect of C. aspersum mucus fractions with MW<20 kD and MW>20 kD on S. cerevisiae NBMCC 584 cellular reduced glutathione levels (GSH) (a) and total antioxidant capacity (TAC) (b).
Figure 12.
Effect of C. aspersum mucus fractions with MW<20 kD and MW>20 kD on S. cerevisiae NBMCC 584 cellular reduced glutathione levels (GSH) (a) and total antioxidant capacity (TAC) (b).

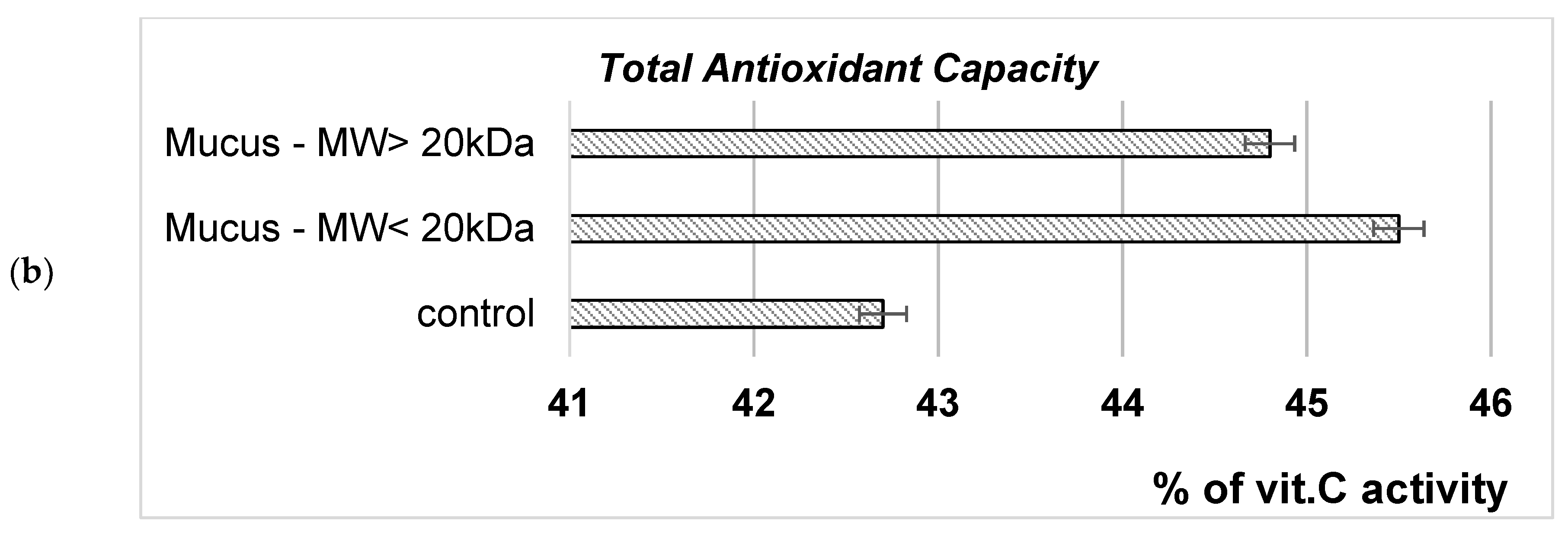

Table 1.
The primary structures of the peptides with MW below 3 kDa, from a garden snail C. aspersum, identified by de novo sequencing in MALDI-MS/MS.
Table 1.
The primary structures of the peptides with MW below 3 kDa, from a garden snail C. aspersum, identified by de novo sequencing in MALDI-MS/MS.
| No | Amino Acid Sequence of Peptides | [M+H]+ Da |
Calcul. monois. mass Da | pI | GRAVY | Net Charge | Predicted by iAMPpred software | ||
|---|---|---|---|---|---|---|---|---|---|
| Аntibacterial (%) | Аntiviral (%) | Аntifungal (%) | |||||||
| 1 | LLPFKEPDL | 1071.60 | 1070.60 | 4.37 | −0.600 | −2/+1 | 28 | 51 | 19 |
| 2 | ACGATLQLENCG | 1179.77 | 1178.51 | 4.00 | +0.350 | -1/0 | 29.3 | 35.7 | 43.7 |
| 3 | LNLGGNGANGLVGG | 1212.76 | 1211.63 | 5.52 | +0.321 | 0/0 | 74.0 | 43.1 | 74.3 |
| 4 | AGVGGAAGNPSTYVG | 1277.70 | 1276.60 | 5.57 | +0.260 | 0/0 | 25.1 | 7.4 | 11.3 |
| 5 | GGGMVKEDGSCLGV | 1308.77 | 1307.58 | 4.37 | +0.207 | −2/+1 | 40.4 | 31.4 | 33.7 |
| 6a | MLGGGVNSLRPPK | 1325.80 | 1324.73 | 11.0 | −0.262 | 0/+2 | 22.8 | 14.0 | 8.9 |
| 7 | CVGGAGGHGDSCAKGT | 1376.53 | 1375.56 | 6.73 | −0.106 | −1/+1 | 85.2 | 48.8 | 74.4 |
| 8 | GGGGYHTWGEGGKF | 1409.48 | 1408.62 | 6.75 | −0.964 | −1/+1 | 69.0 | 62.8 | 72.6 |
| 9 | MLNVAVNKGEVKH | 1438.86 | 1437.78 | 8.37 | −0.138 | −1/+2 | 56.4 | 38.0 | 19.7 |
| 10b | NLVGGSGGGGRGGANPLG | 1496.79 | 1495.75 | 9.75 | −0.217 | 0/+1 | 66.0 | 33.7 | 48.2 |
| 11 | GTMSPAGGEMGPVTAGVG | 1576.04 | 1574.71 | 4.00 | +0.250 | −1/0 | 13.1 | 24.8 | 8.3 |
| 12 | GTKGCGPGSCPPGDTVAGVG | 1716.79 | 1715.76 | 5.82 | −0.100 | −1/+1 | 23.8 | 20.2 | 25.8 |
| 13c | ACSLLLGGGGVGGGKGGGGHAG | 1738.96 | 1737.86 | 8.27 | +0.409 | 0/+1 | 82.6 | 49.5 | 67.0 |
| 14a | LLLDGFGGGLLVEHDPGS | 1796.00 | 1794.92 | 4.02 | +0.439 | -3/0 | 37 | 45 | 10 |
| 15d | MGGWGGLGGGHNGGWMPPK | 1852.96 | 1851.83 | 8.52 | −0.611 | 0/+1 | 69 | 56.0 | 57.0 |
| 16c | ACLTPVDHFFAGMPCGGGP | 1876.88 | 1875.81 | 5.08 | +0.542 | −1/0 | 32.4 | 42.6 | 20.1 |
| 17c | NGLFGGLGGGGHGGGGKGPGEGGG | 1909.98 | 1908.88 | 6.75 | −0.487 | −1/+1 | 89.5 | 67.2 | 79.5 |
| 18 | LLLDNKGGGLVGGLLGGGGKGGG | 1966.11 | 1965.10 | 8.59 | +0.322 | −1/+2 | 93 | 56 | 81 |
| 19 | GMVLLHCSPALDFHKTPAV | 2036.09 | 2035.04 | 6.91 | +0.616 | −1/+1 | 18 | 53 | 14 |
| 20 | LPFLLGVGGLLGGSVGGGGGGGGAPL | 2136.24 | 2135.17 | 5.52 | +1.023 | 0/0 | 66 | 33 | 36 |
| 21 | MVLDGKGGGGLLGGVLGGGKDAHLGG | 2292.32 | 2291.21 | 6.50 | +0.319 | −2/+2 | 84.3 | 59.0 | 71 |
| 22 | LLKDNGVGGLLGGGGAGGGGLVGGNLGGGAG | 2478.39 | 2477.30 | 5.84 | +0.439 | −1/+1 | 86.4 | 54 | 66 |
| 23 | KTSKLMVYLAGGGGGLLGGVGGGGGGAGGGGPGGL | 2843.76 | 2842.48 | 9.70 | +0.374 | 0/+2 | 76 | 47 | 67 |
Table 2.
AASs of peptides, determined after analysis of their MS/MS spectra. Proteins were identified after comparing AASs with a database of protein sequences by the Basic Local Alignment Search Tool (BLAST).
Table 2.
AASs of peptides, determined after analysis of their MS/MS spectra. Proteins were identified after comparing AASs with a database of protein sequences by the Basic Local Alignment Search Tool (BLAST).
| Band кDa |
AAS of peptide | Mass [M+H]+ | Protein name | UniProt ID | Identities |
|---|---|---|---|---|---|
| 17.5 | TAAFTEDTSVVTGR | 1454.63 | mucus protein [Cornu aspersum] | QEG59314.1 | 86%, E=8e-05 |
| FTAAFTEDTSVAEGR | 1601.74 | mucus protein [C. aspersum] | QEG59314.1 | 100%, E=2e-09 | |
| GNVAFTAAFTEDTSVAEGR | 1942.91 | mucus protein [C. aspersum] | QEG59314.1 | 100%, E=4e-13 | |
| GALNGNVAFTAAFTEDTSVAEGR | 2298.10 | mucus protein [C. aspersum] | QEG59314.1 | 100%,E=2e-11 | |
| 23.6 | GSCNWPMTILMLESLR | 1850.90 | NADH subunit 6 [Albinaria caerulea] | P48922 | 78%, E=0.005 |
| QYLQITWPSPR | 1388.7 | H-type lectin domain-cont. Protein B. glabrata | KAI8776186.1 | 100%,E=0.041 | |
| 26.7 | VPSDDPGR | 842.40 | Chain A, H. aspersa agglutinin[C. aspersum] | 4Q56_A | 100%,E=0.003 |
| DITFASPYCR | 1172.54 | Chain A, H. aspersa agglutinin C. aspersum | 4Q56_A | 90%,E=1e-04 | |
| NGGLVHPGPR | 1003.54 | uncharacterized protein [Pomacea canaliculata] | XP_025078819.1 | 89%, E=4.5 | |
| 31.35 | DWTLYVNTPLAPAR | 1616.84 | unnamed protein, partial [C. unifasciata] mucin 18b [Plakobranchus ocellatus] |
GFO09821.1 | 78%, E=0.33 75%, E=27 |
| FCPNAQRTR | 1092.54 | glutathione S-transferase omega-1-like [P. Canaliculata] [Elysia marginata] |
XP_025114871.1 GFR70608.1 |
89%, E=0.31 89%, E=0.31 |
|
| NPVGSVPVLELDGK | 1423.78 | glutathione S-transferase omega-1[P. Canaliculata] [B. Glabrata] containing protein [B.glabrata] |
XP_025099831.1 KAI8762368.1 |
86%, E=0.013 79%, E=0.02 |
|
| 33.0 | GPCFTPHTYTNWSWLR | 1965.91 | unnamed protein, Candidula unifasciata] “Bacterial protein of unknown function (HtrL_YibB) | CAG5121511.1 | 63%, E=4e-04 |
| DFLPPASLPDFAPSPPRVAER | 2279.18 | unnamed protein product, [Candidula unifasciata] | CAG5136685.1 | 53%, E=0.34 | |
| 39.1 | AGYLQITWPSPR | 1388.72 | mucus protein [C. aspersum] H-type lectin domain- |
QEG59312.1 KAI8776186.1 |
100%,E=1e-08 100% E=0.055 |
| ASVTGDLSNK | 991.51 | mucus protein [C. aspersum] | QEG59312.1 | 100%,E=8e-04 | |
| ELLGNVYRAAFTEDTSVAEGR | 2298.14 | mucus protein [C. aspersum] | QEG59314.1 | 77%, E=3e-08 | |
| VGSNGAR | 660.34 | mucus protein [C. aspersum] | QEG59312.1 | 100%E=0.004 | |
| ANVTFDLSEK | 1123.56 | von Willebrand factor A domain-containing protein 3B-like isoform X1 [B. glabrata] protein 3B isoform X2, [B. glabrata] |
XP_013069929.2 XP_013069930.2 |
100%, E=0.59 100%, E=0.59 |
|
| LFSTASGGR | 895.4632 | Collagen alpha-6(VI) chain, partial [Biomphalaria pfeifferi] | KAK0040452.1 | 89% E=1.9 | |
| LPLYEDPKLDVSSLR | 1744.83 | uncharacterized protein LOC101853718 [A. californica] | XP_012941625.1 | 83%, E=0.27 | |
| 48.7 | FDANPFFSGR | 1157.59 | Hemocyanin, βC chain unit D [Helix pomatia] | P12031.2 | 89%, E=9e-06 |
| HGSPIGVPYWDWTR | 1670.93 | Hemocyanin α N [C. aspersum] | AYO86684.1 | 100%, E=2e-09 | |
| LISEATYFNSR | 1300.71 | hemocyanin β [C. aspersum] βC chain unit D [H. pomatia] |
AYO86685.1, P12031.2 |
100%, E=2e-04 82%, E=6e-05 |
|
| FDPNPFFSGR | 1183.5 | Hemocyanin, βC chain unit D [H. pomatia] | P12031.2 | 100%,E=3e-07 | |
| EVFEQVEHALLAR | 1540.81 | Hemocyanin β [C. aspersum] βC chain unit D [H. pomatia] |
AYO86685.1 P12031.2 |
100%, E=0.006 88%, E=0.050 |
|
| QYLQITWSPPR | 1388.73 | fibrinogen-like protein A isoform [B. glabrata] | XP_055864456.1 | 78%, E=1.4 | |
| 52.8 | TDVTSALLGARCNEGGTNTHSPLR | 2470.21 | unnamed protein, partial [C. unifasciata] unnamed protein C. unifasciata |
CAG5131631.1 CAG5131621.1 |
70%, E=0.006 63%, E=0.26 |
| LTQHFNVGSNGAR | 1400.70 | mucus protein [C. aspersum] unnamed protein C. unifasciata unnamed protein partial [C. unifasciata] |
QEG59312.1 CAG5131621.1 CAG5131631.1 |
92%,E=0.001 77%, E=0.094 75%, E= 0.38 |
|
| MPDYDCGCCNGSNGSYGSGGGGGGGR | 2384.84 | unnamed protein, partial [C.unifasciata] unnamed protein product [C. unifasciata] |
CAG5126455.1 CAG5131834.1 |
80% , Е=0.23 63% Е=0.23 |
|
| 56.9 | DGMSAAPQAENAFALK | 1620.77 | Achacin; Flags: Precursor [L. Fulica] L-amino-acid oxidase (Escapin) A. Californica |
P35903.1 Q6IWZ0.1 |
71%, E=3e-04 63%, Е=0.95 |
| SGFPTTSKDR | 1095.54 | L-amino-acid oxidase (Escapin) [A. Californica | Q6IWZ0.1 | 100%, E=0.080 | |
| VGGGPSGVELDEFEARVELK | 2088.0 | Achacin ; Flags : Precursor [Lissachatina fulica] | P35903.1 | 41% E=0.40 | |
| AELKGDMQYTYADSEKVR | 2104.00 | fibrillin-3, partial [Biomphalaria pfeifferi] fibrillin-3, partial [B. Glabrata] |
KAK0048148.1 KAK6970092.1 |
48%, Е=0.22 48%, Е=0.22 |
|
| 70.8 | AKVQVIGVPDDRLLLR | 1792.08 | acyl-CoA synthetase family member 2, mito- chondrial-like, partial [Pomacea canaliculata] unnamed protein product, [C. unifasciata] |
XP_025086706.1 CAG5136482 |
91%, E=0.005 91%, E=0.005 |
| HGGGGGGFGGGGFGSR | 1320.65 | uncharacterized protein LOC124113905 [H. rufescens fibroin heavy chain isoform X1 [A. Californica] elastin-like [Ostrea edulis] Glycine, alanine asparagine -rich protein [Haliotis asinina] |
XP_046330350.2 XP_012943828.1 XP_056017499.1 P86732.1 |
100%,E=9e-04 81%, E=0.018 87%, E=0.038 65%, E=0.004 |
|
| 84.7 | YVLEDLSAADLELSR | 1693.86 | FMRFamide-activatedamiloride-sensitive sodium channel [C. aspersum] | Q25011.1 | 100%, E=0.14 |
| GSSVSLCVWDWAVLPR | 1774.8 | FMRFamide-activatedamiloride-sensitive sodium channel [C. aspersum] | Q25011.1 | 71%, E= 1.9 | |
| 97.0 | MVTSLYPGEDWAMLPR | 1865.89 | mucin-5AC-like, partial [Haliotis rufescens] | XP_048239711.1 | 56%, E= 0.15 |
| ETVSPGYSEGECTCASITQK | 2089.91 | mucin-5B-like isoform X4 [H. rubra] | XP_046552239.1 | 62%, E= 6.7 | |
| VFADFELHNIGASADVR | 1860.92 | hemocyanin β [C. aspersum] | AYO86685.1 | 100%, E= 2e-10 | |
| CQVSLPYWDWAVPLR | 1832.92 | hemocyanin, βC chain unit G [H. pomatia] | P56823.1 | 100%, E= 5e-04 | |
| LSGYDVSKVNEILK | 1564.86 | hemocyanin β [C. aspersum] | AYO86685.1 | 85%, E=8e-05 | |
| HLWLLHCFEHDLNGYEYDNLR | 2687.25 | hemocyanin β [C. aspersum] | AYO86685.1 | 87% E=8e-06 | |
| ANNARLTDASHDNPFSSYTLR | 2350.12 | hemocyanin β [C. aspersum] | AYO86685.1 | 94%, E=1e-09 | |
| 143.3 | VGRIESESYVVKKSK | 1708.96 | zinc finger protein 502-like [B. glabrata] 184 [B. glabrata] |
XP_055863608 KAI8795828.1 |
77%, E=0.12 77%, E=0.12 |
| VLTFYQSCDCVVDHCHR | 2024.88 | zinc finger protein 664-like [Patella vulgata] | XP_050401404.1 | 75%, E=1.5 | |
| GEPGSTGPPGLK | 1096.56 | collagen alpha-1(IV) chain-like, partial [P. canaliculata] | XP_025114158.1 | 100%, E=2e-04 | |
| NGPSGVELDEFEARVELK | 1988.99 | collagen alpha-1(XIII) chain – like isoform X7 [H. rubra] | XP_046573418.1 | 59%, E=2.5 | |
| FVDMDGMFSSAAGGRPEAR | 2000.89 | collagen α-4(VI) chain-like X2 [P. acuta] α-1(XII) chain-like [P. acuta] α-1(XII) chain-like isoform X6 [B. glabrata] |
XP_059154404.1 XP_059163536.1 XP_055860588.1 |
73%, E= 0.012 75%, E=0.26 68%, E=0.26 |
|
| RAYTDGMFSSAAGGRPEAR | 1999.94 | collagen α-4(VI) chain-like isoform [P. acuta] collagen α-4(VI) chain-like isoform [P. acuta] |
XP_059154404.1 XP_059154396.1 |
79%,E=0.031 79%, E=0.031 |
|
| CNEGGTNTHSPLR | 1385.62 | collagen α-6(VI) chain-like [Physella acuta] | XP_059164377.1 | 89%, E=6.1 | |
| 198.31 | NPGYLVSKVNEILK | 1573.89 | mucin-2-like [P. acuta] serine/arginine repetitive matrix protein 2 [B. glabrata] | XP_059140993.1 KAI8735946.1 |
71%, E=0.84; 71%, E=0.84 |
| DCRVRVGGPLFAPSPYLFER | 2279.1 | mucin-17-like [Haliotis rubra] | XP_046554219.1 | 52%,E=0.78 | |
| FNFLPLSADALESLR | 1692.90 | E3 ubiquitin-protein ligase HERC2-like isoform X2 [B. glabrata] | XP_055894969.1 | 83%, E= 0.50 | |
| ASSGSCDFSSSTAANR | 1547.64 | mucin-5AC-like isoform X2 [H. rufescens] mucin-5AC-like isoform X1 [H. rufescens] |
XP_048252836.1 XP_048252834.1 |
85%,E= 0.45 85%,E= 0.45 |
|
| GCAGCPQAGSK | 978.41 | fibrillin-1-like [B. glabrata] | XP_055888733.1 | 89%, Е=2.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



