
Submitted:
30 April 2024
Posted:
30 April 2024
You are already at the latest version
Abstract
Sugarcane mosaic virus (SCMV) and sugarcane streak mosaic virus (SCSMV) are the primary viruses responsible for mosaic diseases in sugarcane. In this study, the mosaic viruses SCMV and SCSMV with the involvement of sorghum mosaic virus (SrMV) were surveyed from eight commercial sugarcane cultivars grown in fields at four locations in East Java, Indonesia. Sugarcane leaves with mosaic symptoms were assessed for disease incidence and severity, and symptomatic leaves were collected for analysis. The incidence and severity of mosaic symptoms were the highest in the PS881 and NX04 cultivars grown in Kediri compared to the other cultivars. RT-PCR analysis showed that SCSMV was present in all sugarcane cultivars and locations. Interestingly, double mixed infections with SCMV were found in PS881, PS882, and Cening, whereas triple mixed infections with SrMV were found in the PS881 cultivar. Phylogenetic analysis grouped SCSMV, SCMV, and SrMV together with isolates from Thailand (KP987832.1), Florida, USA (MT725538.1), and Agentina (EU189037.1). The expression of ascorbate peroxidase gene (Apx) was increased in the virus-infected sugarcane cultivars and prominently elevated in the triple mixed infection of PS881 cultivars. However, the catalase gene (Cat) was slightly upregulated in the mixed infection group. The chlorophyll content was significantly reduced in the leaves of PS58, PS881, PS882, NX04, and HW Merah, in parallel with the expression of the photosystem I gene (PsaA). The expression of photosynthetic genes, such as ribulose-1,5 bisphosphate carboxylase small sub-unit (RbcS), phosphoenolpyruvate carboxylase (Pepc), sucrose-phosphate synthase (Sps1), and PsaA, was significantly reduced and almost depleted in the triple mixed infection of PS881, except for Pepc expression, which remained approximately half of that in the control healthy sugarcane. The decrease in photosynthetic activity was confirmed by the decrease in ribulose-1,5 bisphosphate carboxylase (Rubisco) and PEPC proteins, as detected by immunoblot analysis. The decrease in photosynthetic parameters indicated that the mosaic viral infection substantially reduced the growth and yield of sugarcane.
Keywords:
sugarcane
; mosaic virus infection
; mixed infections
; photosynthetic parameters
1. Introduction
Sugarcane (Saccharum spp) is an important industrial crop for sugar production, contributing nearly 80% of the world’s sugar production [1]. The plant also produces ethanol as a biofuel and other economically important products, such as animal feed, paper, organic fertilizer, and industrial enzymes [2]. Sugarcane production in Indonesia is mainly located on Java Island (56.3 %), Southern Sumatra (26.4%), and other spotted regions, such as South Sulawesi and West Nusa Tenggara [3]. Sugarcane is asexually propagated using stalks that are easily transmitted and infected by various plant viruses, such as sugarcane mosaic disease [4,5] and yellow leaf disease [6]. Mosaic is one of the most important sugarcane diseases, and infection leads to the manifestation of mosaic-like symptoms, which develop as irregular patterns of light and dark green or yellow patches or streaks [7,8]. Mixed infections with two or more viruses are frequent in plants and result in severe symptoms [9]. Mosaic diseases in sugarcane with mixed infections of sugarcane mosaic virus (SCMV), sugarcane streak mosaic virus (SCSMV), and sorghum mosaic virus (SrMV) have been reported in China [9,10]; SCMV and SrMV in the USA [11]; and SCMV and SCSMV in India [12].
Biotic stress caused by viral infection increases oxidative stress, leading to increased synthesis of reactive oxygen species (ROS). Mosaic virus infection significantly upregulates ROS-producing genes, such as NAD oxidase, malate dehydrogenase, and flavin-binding monooxygenase [13]. ROS production can potentially lead to damage of cellular components and programmed cell death [14,15]. However, ROS accumulation can be detoxified by non-enzymatic and enzymatic antioxidants, such as catalase (Cat), superoxide dismutase, ascorbate peroxidase (Apx), and glutathione reductase [16,17,18]. The antioxidant defense mechanim for scavenging ROS protects cells from oxygen radical [19].
SCMV infection stimulates malate synthesis by enhancing pyruvate orthophosphate dikinase (PPDK) activity, leading to ROS accumulation, which requires light illumination [20]. As a C4 plant, sugarcane shows higher rates of photosynthesis under high light intensities. Atmospheric CO2 is fixed by phosphoenolpyruvate carboxylase (PEPC) using the substrate phosphoenolpyruvate (PEP) to produce oxaloacetate and malate in the C4 cycle. Malate is then transported into the bundle sheath cell. Malate is then decarboxylated in the bundle sheath cells to produce CO2 molecules and refixed by ribulose-1,5-bisphosphate carboxylase (Rubisco), which forms sucrose and starch [21]. To maintain the C4 cycle, PEP is regenerated from pyruvate using PPDK and then used by PEPC to produce malate. It is well known that PPDK activity is regulated by light [22], including several carbon-assimilating enzymes, such as PEPC and sucrose phosphate synthase (SPS) [23]. However, the effect of mosaic viral infection on photosynthetic activity remains poorly understood.
Mosaic diseases can significantly reduce chlorophyll content, photosynthetic efficiency, yield, and quality of sugarcane [7,24,25]. The expression of light-harvesting proteins and primary carbon-assimilating enzymes is high in resistant cultivars [13] and non-infected sugarcane [25]. The expression of NADPH, Rubisco, and photosystem I (PSI) reaction center is upregulated in mosaic-resistant compared to that in susceptible sugarcane. Protein analysis has confirmed that the activities of phosphoenolpyruvate carboxylase (PEPC) and Rubisco are decreased in virus-infected sugarcane seedlings [24,25]. The alterations in the photosynthesis-related activities result in a slowdown in sugarcane growth [17], reducing sucrose content and sugarcane yield up to 75% [7]. In addition, mosaic virus infection reduces the chlorophyll content in cassava [26] and poplar [27], and perturbs pigment biosynthesis in tomato [28]. These results indicate that mosaic virus infection downregulates the expression of photosynthesis-related genes [27,29,30].
Mosaic symptoms in sugarcane are frequently associated with several different viral infections. Multiple or mixed viral infections have also been reported in sweet potatoes in Uganda [30] and cotton plants in China [10]. Mixed infections from two or more viral interactions are categorized as synergistic, antagonistic, or neutral interaction [9,31] that induce a decline in plant vigor and productivity [32]. The combined infection of viruses that act synergistically causes symptom severity and plant death or severely reduces yield [33], as reported in maize [34]. In the present study, the identification and distribution of the mosaic virus were surveyed in field-grown sugarcane located in four regions of East Java, Indonesia. Symptomatic sugarcane leaves were observed and collected for molecular detection and identification of three major RNA viruses, SCSMV, SCMV, and SrMV, via RT-PCR using specific primer pairs. The effect of viral infection on photosynthetic gene expression was determined using RT-PCR and immunoblot analysis. We found that mixed infection with mosaic viruses (SCMV, SCSMV, and SrMV) significantly reduced chlorophyll content and photosynthetic gene expression in sugarcane.
2. Materials and Methods
2.1. Survey Area and Leaf Sample Collection
Mosaic symptoms were observed in commercial field-grown sugarcane in four locations: the districts of Kediri, Lumajang, Jember, and Bondowoso in East Java Province. This province is the primary location for sugarcane production in Indonesia. The surveys were conducted from September to November 2022 in 3–4 months plant age in sugarcane plantation under the authority of PT. Perkebunan Nusantara X and PT. Perkebunan Nusantara XI. Sugarcane mosaic symptoms were assessed based on the disease incidence and severity. Disease incidence was determined as the percentage of sugarcane mosaic symptoms relative to the total observed sugarcane in the field. Disease severity was estimated using the scoring system of the young sugarcane leaf area showing symptoms at the position of the first and third leaves from the green top, according to previously described methods [5]. The severity percentage was calculated using the disease severity index (DSI) = [sum (class frequency × score of rating class)/(total number of plants × (maximal disease index)] x 100 [35]. A total of 62 sugarcane leaf samples with mosaic symptoms were disinfected with 75% alcohol and immersed in liquid nitrogen for further experiments.
2.2. Total RNA Extraction and Mosaic Virus Determination
Total RNA was extracted from approximately 0.5 g of sugarcane leaf using the RNAprep pure plant plus kit (Tiangen, China) for the mosaic virus detection [36]. The total RNA concentration was measured using a NanoVuePlus-UV Spectrophotometer (GE Healthcare, USA). Three major mosaics, SCSMV, SCMV, and SrMV, were identified via RT-PCR using specific primer pairs for the CP gene, as shown in Table 1. First-strand cDNA was synthesized from 1 µg total RNA using ReverTra AceTM Kit (Toyobo, Japan) and the PCR reaction was conducted using a Kit of KOD-Plus-Neo (Toyobo, Japan). The PCR mixture (50 µL) contained 5 µL of 10x buffer KOD-Plus-Neo, 5 µL of 2 mM dNTPs, 3 µL of 25 mM MgSO4, 1 µL of 10 µmol/L specific primer for corresponding the mosaic virus, and 1 µL DNA Polymerase KOD-Plus-Neo. PCR analysis was performed at 95 °C for 5 min, followed by 35 cycles at 95 °C for 30 s denaturation, 53 °C (SCSMV), 58 °C (SCMV), or 52 °C (SrMV) for 30 s annealing, 72 °C for 1 min extension, and followed by final extension at 72 °C for 5 min using T100TM Thermal Cycler (Bio-Rad, USA). The product of PCR amplified was run on 1% agarose gel, stained with ethidium bromide, and documented using GelDoc (Major Science, USA). The corresponding PCR products were excised and purified from agarose gel using the GenepHlowTM Gel/PCR Kit (Geneaid, Taiwan) and used for nucleotide sequence determination.
2.3. DNA Sequencing and Phylogenetic Analysis
The PCR products were delivered to a DNA sequencing provider (1st BASE, Singapore) for nucleotide sequencing. The obtained nucleotide sequences were aligned using the BioEdit Sequence Alignment Editor V 7.2.5, and confirmed to be specific viruses using BLAST/N online (https://blast.ncbi.nlm.nih.gov). The nucleotide sequences of SCSMV, SCMV, and SrMV were used to construct a phylogenetic tree using the neighbor-joining (NJ) method (1000 bootstrap replicates) by Molecular Evolutionary Genetics Analysis (MEGA) software version 10.2.6 [37].
2.4. Quantitative Real-Time PCR
Quantitative real-time PCR (qRT-PCR) was carried out using a CFX connectTM Real-TimeSystem (Bio-Rad, USA) to analyze expression levels of photosynthesis-related genes and ROS scavenging. The 25-µL reaction system containing 12.5 µL of 2x SYBR Green master mix (ThunderbirdTM SYBR qPCR Mix, Toyobo, Japan), 1 µL of 10 µmol/L of forward and reverse primers, 1.5 µL of 50 ng cDNA, and nuclease-free water to adjust the volume reaction. Gene-specific primer pairs are listed in Table 1. The qRT-PCR program was 95 °C for 30 s, followed by 40 cycles at 95 °C for 15 s, the temperature of primer-dependent annealing (Table 1) for 15 s, elongation at 72 °C for 15 s, and final extension at 72 °C for 60 s. Each sample was conducted in duplicate from three independent biological replicates and the expression was normalized using β-tubulin expression. The relative gene expression was calculated via Livak’s method using the formula 2-∆∆CT [38].
2.6. Protein Extraction and Immunoblotting
Frozen sugarcane leaves (1 g) were ground in liquid nitrogen and proteins were extracted in a buffer containing 50 mM 3-morpholino propane sulfonic acid (MOPS)-NaOH (pH 7.5), 1 mM ethylenediaminetetraacetic acid (EDTA), 2.5 mM dithiothreitol (DTT), and 10% of polyvinylpyrrolidone (PVP). The soluble protein fraction was obtained by centrifugation at 14,000xg at 4 °C for 10 min and the protein concentration was determined using a Bradford reagent (Bio-rad, USA).
The levels of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) and phosphoenolpyruvate carboxylase (PEPC) proteins were determined by immunoblotting with specific polyclonal antibodies against Rubisco and PEPC [39]. The soluble protein was separated using SDS-PAGE (12.5% acrylamide), 15 µg for PEPC, and 10 µg for Rubisco, subsequently transferred onto a polyvinylidene fluoride (PVDV) membrane (Thermo Scientific, USA) using a semi-dry trans-blotter (Bio-Rad, USA). The membranes were incubated overnight with polyclonal antibodies against Rubisco and PEPC. After washing with Tris-buffered saline, the membrane was incubated with the secondary antibody, goat anti-rabbit IgG alkaline phosphatase conjugate (Bio-Rad, USA), at 1:3000 for 90 min. Specific bands of Rubisco and PEPC were visualized by the enzymatic reaction of the alkaline phosphatase, 5-bromo-4-chloro 3-indolyl-phosphate, and nitro blue tetrazolium (NBT) (Wako, Japan).
2.7. Analysis of Total Chlorophyll of Leaves
Total chlorophyll content was determined using an optical method [40] with slight modifications. Symptomatic sugarcane leaves (0.1 g) were finely cut and incubated with 5 mL of a solution containing a mixture of acetone and ethanol (2:1 v/v) in the dark for 24 h. Each sample was analyzed in triplicate from three independent biological replicates. The absorbance of the solution was read using a spectrophotometer (UV-VIS double beam spectrophotometer (Hitachi, Japan) at 645 nm and 663 nm. A solution without the sample was used as the blank. The concentration of total chlorophyll was estimated using the following equation: mg total chlorophyll/g tissue: 20.2 x (absorbance of 645 nm) + 8.02 x (absorbance of 663 nm) × final volume of chlorophyll extract × fresh weight of leaf extract [41].
3. Results
3.1. Mosaic Symptom, Incidence, and Severity
Mosaic symptoms were observed in four sugarcane plantation locations, and 62 leaf samples were collected. Mosaic symptoms, presenting as blotchy or streaky patterns of yellow and green color with varying lengths, sizes, and irregularities, were recorded to determine the incidence percentage. The incidence of symptoms observed in Kediri was higher than that in the other locations (Figure 1A). The highest incidence was in NX04, followed by PS881 that reached almost 80 and 54%, respectively. However, the incidence of PS881 was slightly lower in Kediri than that in a previous study in the Jember area [5]. Furthermore, the percentage of severity was high, above 80% in all cultivars (Figure 1B), and a high percentage was found in PS58, PS881, and NX04. Although the mosaic patterns were similar in all of the sugarcane varieties, but the symptoms differed in intensity (Supplementary Figure S1).
To determine whether a mosaic infection of one or more viral types occurred, RT-PCR was conducted using specific primer pairs of CP gene for the detection of SCMV, SCSMV, and SrMV (Table 1). These three viruses are considered major mosaic viruses in sugarcane. The analysis showed that the corresponding CP-SCSMV DNA, with a molecular size of 335 kb, was present in all sugarcane leaf samples (Figure 2A). This result indicated that SCSMV was more widespread in all cultivars and regions used in this experiment than in previously reported results [4]. Furthermore, the SCMV with 735 kb in size was found mainly in PS881 and PS882, and only a few in the Cening cultivars, but the corresponding DNA band was not detected in NX04, NX01, HW Merah, PS58, and NXI-4T (Figure 2B). Interestingly, a specific primer for SrMV detection, which is a rare mosaic virus infection, was found in the PS881 cultivar, but not in the other cultivars (Figure 2C). These results indicate the presence of a mixed mosaic virus infection with double and triple infections, which has been reported for the first time in an Indonesian sugarcane plantation.
3.2. Phylogenetic Analysis of the Mosaic Virus Isolates
To confirm infection by the mosaic virus, the corresponding CP-DNA of SCSMV, SCMV, and SrMV were determined by their nucleotide sequences. The sequences were correctly verified from each mosaic virus using NCBI BLAST, and phylogenetic trees were constructed using MEGA software. The sequence identity analysis indicated that nucleotide sequence of the targeted sequences shares 92.11% (KP987832.1), 92.58% (MT725538.1) and 97.39% (EU189037.1) homology for SCSMV, SCMV, and SrMV with those of other the virus isolates available in the GenBank, respectively. Phylogenetic analysis grouped the SCSMV with isolates from Thailand (KP987832.1), and the SCMV isolate was grouped with isolates from Florida, USA (MT725538.1) and Boetzingen, Germany (X98168.1) (Figure 3A,B). The SrMV isolate was grouped together with isolate from Argentina (EU189037.1), USA (EF078962.1), and China (DQ227695.1), (Figure 3C).
3.3. Expression of Genes for Apx and Cat
Environmental stresses, including biotic stresses, produce ROS that regulate cellular metabolism and inhibit cell growth and development. Plants have developed antioxidant mechanisms to overcome the detrimental effects of ROS. Among the antioxidant system, the expressions of Apx and Cat have been studied in response to pathogen infection [37,42]. Detection of Apx expression showed increases in viral infection compared to the asymptomatic healthy controls of field-grown sugarcane (Figure 4A). The expression was substantially higher in PS881 and NX04 than in the other cultivars, and a slight increase was observed in NX4T and NX01 among the infected sugarcane cultivars. The change in Apx expression was associated with the ROS detoxification pathway in the SCMV-resistant sugarcane genotype [13]. However, whether Apx expression in sugarcane indicates the degree of resistance to viral infections is not well understood.
To confirm the response of antioxidant expression to mosaic virus infection, RNA was isolated from single, double, and triple mixed infections of PS881 leaves, and the expression of Apx together with Cat genes was determined. The presence of mixed infections in the leaves of the PS881 cultivar was confirmed by detection of the virus using RT-PCR (Supplementary Figure S2). The expression of Cat was slightly induced in single and double infections and increased approximately 1.4-fold in mixed-triple infection by the addition of SrMV compared to that in healthy leaves (control) (Figure 4B). However, the expression of Apx significantly increased in the mixed infection group, and the highest expression was found in the triple infection group, reaching almost 3.9-fold. These results indicate that Apx expression was mostly affected by mosaic infection, rather than Cat expression.
3.4. Expression of Photosynthesis Related-Genes in Response to the Mosaic Virus Infection
The mosaic virus infection disturbed the chloroplast structure, which developed leaf chlorosis and malfunctioning of chloroplasts, and downregulated photosystem efficiency [24,43]. To observe the effect of viral infections on chloroplast disruption, the chlorophyll content was determined in the infected leaves. As predicted, the chlorophyll content was significantly reduced in the leaves of PS58, PS881, PS882, NX04, and HW Merah compared to leaves that were not symptomatic (Figure 5A). Furthermore, infection only slightly reduced chlorophyll content in the Cening, NXI-4T, and NX01 cultivars. These results clearly indicate that the mosaic virus infection causes chloroplast disruption or damage, leading to disturbances in photosynthetic activity.
qRT-PCR was used to determine whether the expression of PsaA, located in the chloroplast genome, was affected by viral infection. The expression of PsaA was reduced in the infected leaves of all sugarcane cultivars compared to that in healthy leaves, although the expression varied among the infected cultivars (Figure 5B). Consistent with the chlorophyll content, the expression of PsaA was markedly reduced in the leaves of PS58, PS881, and NX04. However, PsaA expression was slightly reduced in the other cultivars compared to that in healthy plants.
The change in chlorophyll content suggests that viral infection has a significant impact on energy metabolism and photosynthetic efficiency. These changes may contribute to alterations in photosynthetic and carbon-assimilating gene expression in infected plants. Understanding plant responses will provide valuable insights for the development of effective disease management strategies. The expression of photosynthetic genes for RbcS (Rubisco small subunit), Pepc, and Sps1 was measured along with PsaA gene in the triple mixed infection of sugarcane leaves (Figure 5C). The expression of PsaA gradually decreased with the degree of viral infection in single, double, and triple PS881 leaves. The expression of RbcS and Sps1 genes was gradually reduced in single and double infections and was almost depleted in the triple virus infection, including the expression of PsaA. Furthermore, the expression of Pepc was reduced by approximately half in the single-, double-, and triple-infected leaves compared to that in healthy leaves.
To ensure the consistency of gene expression under mosaic virus infection, which reduces photosynthetic activity, the protein content of Rubisco and PEPC was detected using immunoblotting. Rubisco and PEPC are key enzymes for carbon assimilation in C4 plants, such as sugarcane. Consistent with gene expression, the levels of PEPC and both Rubisco-large subunit (RbcL)) and -small subunit (RbcS) proteins decreased according to the degree of single, double, and triple infection in the PS881 cultivars (Figure 5D). Because Rubisco protein is a conservative and slow turnover [44], the reduction of Rubisco protein was still at detectable levels and not depleted as the level of mRNA in the triple-infected leaves.
4. Discussion
In the present study, mosaic diseases were surveyed in four sugarcane plantations located in Lumajang, Jember, Bondowoso, and Kediri in East Java, Indonesia. Eight sugarcane cultivars were observed for mosaic symptoms and sampled for viral analysis. The mosaic diseases were found to have a higher incidence in Kediri than that in the other locations, although they had a similarly high percentage of severity above 80%. The higher incidence in this location might have been caused by differences in integrated disease management, such as heated water treatment prior to planting. Among the sugarcane cultivars, PS881 and NX04 showed a higher incidence and were categorized as susceptible cultivars [4,5]. RT-PCR analysis revealed that SCSMV was spread in all sugarcane cultivars, but SCMV and SrMV were only observed in a few cultivars. Nucleotide sequences of the amplified CP and phylogenetic analyses confirmed the presence of SCSMV, SCMV, and SrMV in sugarcane. Interestingly, PS882 and PS881 cells were infected with double SCSMV-SCMV and triple SCSMV-SCMV-SrMV infections, respectively. These results indicate the presence of a mixed infection with the mosaic virus in sugarcane, which has not been reported in Indonesia.
Mixed infections with two or more viruses occur naturally in plants [9,45]. Infection with SCSMV, SCMV, and SrMV in sugarcane results in the manifestation of mosaic-like symptoms, which involve the appearance of irregular light and dark green patterns on leaves [8]. Therefore, it is not surprising that these three viruses infect sugarcane with high similarity in mosaic symptoms and are difficult to differentiate. Interactions between two or more plant viruses in mixed infections can be categorized as synergistic or antagonistic. A synergistic interaction refers to a situation in which two or more viral infections result in severe symptom [31]. PS881 with triple infection and PS882 with double infection showed a higher percentage of severity (Figure 1B) and mosaic symptom intensity (Supplementary Figure 1) than the other cultivars with single infection. These results indicate that mixed mosaic virus infection with SCSMV, SCMV, and SrMV is a synergistic interaction.
Infections caused by mosaic viruses induce ROS accumulation, and plants have developed antioxidant scavenging and detoxification systems to overcome these problems. The Apx is one of the most important genes for scavenging ROS [37]. The expression of Apx gene increased in all infected sugarcane cultivars, and the increased expression was the highest in the mixed infection of PS881 leaves (Figure 4B). Increased Apx expression can regulate ROS accumulation, which may be considered a characteristic of plant resistance to viral infections. However, elevated Apx expression may be required to balance the oxidation and antioxidation cascades to maintain redox homeostasis.
Viral infection has a significant impact on energy metabolism, respiration, and the photosynthetic rate. Viral proliferation requires energy from infected plant cells, which leads to ROS accumulation [20]. ROS act as signaling molecules and play dual roles. Elevated ROS production can cause cellular damage through lipid peroxidation, protein oxidation, nucleic acid degradation, and the initiation of programmed cell death in infected cells. Viral infection modified the chloroplast structure [26] and caused a reduction in chlorophyll content (Figure 5A) as well as the expression of PsaA gene, which is located in the chloroplast. Chloroplasts play a central role in the generation of energy during photosynthesis. The disruption of chloroplasts causes a reduction in the photosynthetic energy for carbon assimilation. The significant reduction in PsaA expression in the mixed infection of PS881 plants was followed by a reduction in photosynthetic carbon-assimilating genes, such as RbcS, Pepc, and Sps1 as well as their protein content (Figure 5C,D). These results indicate that mosaic virus infection damages chlorophyll and subsequently reduces carbon assimilation activity. Thus, infection with mosaic viruses significantly reduces photosynthetic efficiency, yield, and quality of sugarcane.
The use of resistant sugarcane cultivars is considered the most effective approach for managing viral diseases. Pathogen-derived resistance and RNA interference technologies have been applied to engineered sugarcane cultivars that are resistant to sugarcane mosaic viruses [36,46]. The RNAi approach has been reported to effectively induce high resistance against mosaic viruses in sugarcane [47]. Mixed infection with SCSMV and SCMV, with synergistic interactions, exacerbates sugarcane yield. A strategy employing an RNAi approach can be implemented by assembling a hairpin element composed of CP gene sequences from SCSMV and SCMV in tandem and an intron sequence as a loop. Dual resistance to synergistically interacting viruses has been reported in transgenic orchids [48] and wheat [49]. Therefore, RNAi can be used to induce dual resistance against SCSMV and SCMV in sugarcane.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Variation of mosaic symptoms intensity in leaves of sugarcane variety infected by SCSMV, SCMV, and SrMV. (A) Single infection by SCSMV in Cening, HW Merah, NXI-4T, PS58, PS882, NX01, and NX04 cultivars, (B) Mixed double infection by SCSMV and SCMV in PS881 and PS882 cultivars, and (C) Mixed triple infection by SCSMV, SCMV, and SrMV in PS881 cultivar; Figure S2. RT-PCR amplification of CP gene of SCSMV (left), SCMV (middle), and SrMV (right) in PS881 leaves. M, DNA molecular size marker; S, D, T, and C represent single, double and triple, and control non-symptomatic leaves, respectively. .
Author Contributions
Conceptualization, B.S. and N.D.; methodology, I.R.N., R.H. and W.D.S.; formal analysis, I.R.N. and W.S.; investigation, I.R.N., W.S. and S.; software and statistic, I.R.N., S. and R.H.; validation, N.D., data curation, R.H. and W.D.S.; writing-original draft preparation, I.R.N., W.D.S. and B.S.; writing-review editing, B.S. and R.H.; supervision, B.S.; project administration, N.D. and W.D.S.; funding acquisition, B.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Indonesian Ministry of Education, Culture, Research and Technology, grant number 0267/E5/AK.04/2022.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Purnama Okviandari, Retnosari Apriasti and Almira Siam Veda for their helps and Editage (www.editage.com) for English language editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aono, A. H.; Pimenta, R. J. G.; Garcia, A. L. B.; Correr, F. H.; Hosaka, G. K.; Carrasco, M. M.; Cardoso-Silva, C. B.; Mancini, M. C.; Sforça, D. A.; Dos Santos, L. B.; Nagai, J. S.; Pinto, L. R.; Landell, M. G. D. A.; Carneiro, M. S.; Balsalobre, T. W.; Quiles, M. G.; Pereira, W. A.; Margarido, G. R. A.; De Souza, A. P. The Wild Sugarcane and Sorghum Kinomes: Insights Into Expansion, Diversification, and Expression Patterns. Front. Plant Sci. 2021, 12, 668623. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.-T.; Hoang, Q.-T.; Nguyen, T.-T.; Pham, T.-A.; Cao, A.-D.; Pham, H.-D.; Le, V.-H.; Vu, T.-T.; Pham, N.-H.; Nguyen, T.-C.; To, K.-A.; Nguyen, V.-H.; Phi, Q.-T.; Tran, V.-H.; Dang, T.-T.; Lai, Q.-D.; Lionnet, R.; Chu-Ky, S. Research and Development Prospects for Sugarcane Industry in Vietnam. Sugar Tech 2022, 24, 1330–1341. [Google Scholar] [CrossRef] [PubMed]
- Putra, R.P.; Ranomahera, M.R.R; Rizaludin, M.S.; Supriyanto, R.; Dewi, V.A.K. Short Communication: Investigating Environmental Impacts of Long-Term Monoculture of Sugarcane Farming in Indonesia through DPSIR Framework. Biodiversitas 2020, 21. [Google Scholar] [CrossRef]
- Putra, L. K.; Kristini, A.; Achadian, E. M.; Damayanti, T. A. Sugarcane Streak Mosaic Virus in Indonesia: Distribution, Characterisation, Yield Losses and Management Approaches. Sugar Tech 2014, 16, 392–399. [Google Scholar] [CrossRef]
- Addy, H.; Nurmalasari; Wahyudi, A.; Sholeh, A.; Anugrah, C.; Iriyanto, F.; Darmanto, W.; Sugiharto, B. Detection and Response of Sugarcane against the Infection of Sugarcane Mosaic Virus (SCMV) in Indonesia. Agronomy 2017, 7. [CrossRef]
- Bertasello, L. E. T.; Da Silva, M. F.; Pinto, L. R.; Nóbile, P. M.; Carmo-Sousa, M.; Dos Anjos, I. A.; Perecin, D.; Spotti Lopes, J. R.; Gonçalves, M. C. Yellow Leaf Disease Resistance and Melanaphis Sacchari Preference in Commercial Sugarcane Cultivars. Plants 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Wang, Z.; Xu, F.; Pan, Y.-B.; Grisham, M. P.; Xu, L. Sugarcane Mosaic Disease: Characteristics, Identification and Control. Microorganisms 2021, 9. [Google Scholar] [CrossRef]
- Krishna, G.V; Kumar, V.M.; Varma, P.K.; Bhavani, B.; Kumar, G.V. Identification of Resistance to Sugarcane Mosaic Virus, Sugarcane Streak Mosaic Virus, and Sugarcane Bacilliform Virus in New Elite Sugarcane Accessions in India. Front. Microbiol. 2023, 14, 1276932. [Google Scholar] [CrossRef]
- Xu, Y.; Ghanim, M.; Liu, Y. Editorial: Mixed Infections of Plant Viruses in Nature and the Impact on Agriculture. Front. Microbiol. 2022, 13, 922607. [Google Scholar] [CrossRef]
- He, E.-Q.; Bao, W.-Q.; Sun, S.-R.; Hu, C.-Y.; Chen, J.-S.; Bi, Z.-W.; Xie, Y.; Lu, J.-J.; Gao, S.-J. Incidence and Distribution of Four Viruses Causing Diverse Mosaic Diseases of Sugarcane in China. Agronomy 2022, 12. [Google Scholar] [CrossRef]
- Filloux, D.; Fernandez, E.; Comstock, J. C.; Mollov, D.; Roumagnac, P.; Rott, P. Viral Metagenomic-Based Screening of Sugarcane from Florida Reveals Occurrence of Six Sugarcane-Infecting Viruses and High Prevalence of Sugarcane Yellow Leaf Virus. Plant Disease 2018, 102, 2317–2323. [Google Scholar] [CrossRef]
- Viswanathan, R.; Balamuralikrishnan, M.; Karuppaiah, R. Characterization and Genetic Diversity of Sugarcane Streak Mosaic Virus Causing Mosaic in Sugarcane. Virus Genes 2008, 36, 553–564. [Google Scholar] [CrossRef]
- Akbar, S.; Wei, Y.; Yuan, Y.; Khan, M. T.; Qin, L.; Powell, C. A.; Chen, B.; Zhang, M. Gene Expression Profiling of Reactive Oxygen Species (ROS) and Antioxidant Defense System Following Sugarcane Mosaic Virus (SCMV) Infection. BMC Plant Biol 2020, 20. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Sun, Y.; Zhao, Y.; Zhang, J.; Luo, L.; Li, M.; Wang, J.; Yu, H.; Liu, G.; Yang, L.; Xiong, G.; Zhou, J.-M.; Zuo, J.; Wang, Y.; Li, J. Deficient Plastidic Fatty Acid Synthesis Triggers Cell Death by Modulating Mitochondrial Reactive Oxygen Species. Cell Res 2015, 25, 621–633. [Google Scholar] [CrossRef]
- Sawitri, W. D.; Slameto; Sugiharto, B.; Kim, K-M. Identification of Chinese Cabbage Sentrin as a Suppressor of Bax-Induced Cell Death in Yeast. J. Microbiol. J. Microbiol. Biotechnol. 2012, 22, 600–606. [CrossRef]
- Huseynova, I. M.; Mirzayeva, S. M.; Sultanova, N. F.; Aliyeva, D. R.; Mustafayev, N. Sh.; Aliyev, J. A. Virus-Induced Changes in Photosynthetic Parameters and Peroxidase Isoenzyme Contents in Tomato (Solanum Lycopersicum L.) Plants. Photosynt. 2018, 56, 841–850. [Google Scholar] [CrossRef]
- Zandi, P.; Schnug, E. Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Manimekalai, R.; Narayanan, J.; Ranjini, R.; Gokul, M.; Selvi, A.; Kumar, P.; Gomathi, R. Hydrogen Peroxide-Induced Oxidative Stress in Sugarcane and Response Expression Pattern of Stress-Responsive Genes Through Quantitative RT-PCR. Sugar Tech 2018, 20, 681–691. [Google Scholar] [CrossRef]
- Sahu, P. K.; Jayalakshmi, K.; Tilgam, J.; Gupta, A.; Nagaraju, Y.; Kumar, A.; Hamid, S.; Singh, H. V.; Minkina, T.; Rajput, V. D.; Rajawat, M. V. S. ROS Generated from Biotic Stress: Effects on Plants and Alleviation by Endophytic Microbes. Front. Plant Sci. 2022, 13, 1042936. [Google Scholar] [CrossRef]
- Jiang, T.; Du, K.; Xie, J.; Sun, G.; Wang, P.; Chen, X.; Cao, Z.; Wang, B.; Chao, Q.; Li, X.; Fan, Z.; Zhou, T. Activated Malate Circulation Contributes to the Manifestation of Light-Dependent Mosaic Symptoms. Cell Reports 2023, 42. [Google Scholar] [CrossRef]
- Ludwig, M. The Roles of Organic Acids in C4 Photosynthesis. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Chastain, C. J.; Chollet, R. Regulation of Pyruvate, Orthophosphate Dikinase by ADP-/Pi-Dependent Reversible Phosphorylation in C3 and C4 Plants. Plant Physiology and Biochemistry 2003, 41, (6–7). [Google Scholar] [CrossRef]
- Huber, S. C.; Huber, J. L. Role and Regulation of Sucrose-Phosphate Synthase In Higher Plants. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1996, 47, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Akbar, S.; Yao, W.; Yu, K.; Qin, L.; Ruan, M.; Powell, C. A.; Chen, B.; Zhang, M. Photosynthetic Characterization and Expression Profiles of Sugarcane Infected by Sugarcane Mosaic Virus (SCMV). Photosynth Res 2021, 150, (1–3). [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Deng, Q.; Chen, J.; Shen, W. Physiological and Molecular Mechanisms Governing the Effect of Virus-Free Chewing Cane Seedlings on Yield and Quality. Sci Rep 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Zanini, A. A.; Di Feo, L.; Luna, D. F.; Paccioretti, P.; Collavino, A.; Rodriguez, M. S. Cassava Common Mosaic Virus Causes Photosynthetic Alterations Associated with Changes in Chloroplast Ultrastructure and Carbohydrate Metabolism of Cassava Plants. , 2020. 25 April. [CrossRef]
- Wang, L.; Zhang, W.; Shen, W.; Li, M.; Fu, Y.; Li, Z.; Li, J.; Liu, H.; Su, X.; Zhang, B.; Zhao, J. Integrated Transcriptome and microRNA Sequencing Analyses Reveal Gene Responses in Poplar Leaves Infected by the Novel Pathogen Bean Common Mosaic Virus (BCMV). Front. Plant Sci. 2023, 14, 1163232. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, I. M.; Peter Van Esse, H.; Ballester, A.-R.; Hogewoning, S. W.; Parra, N. O.; Paeleman, A.; Lievens, B.; Bovy, A. G.; Thomma, B. P. H. J. Differential Tomato Transcriptomic Responses Induced by Pepino Mosaic Virus Isolates with Differential Aggressiveness. Plant Physiol. 2011, 156, 301–318. [Google Scholar] [CrossRef] [PubMed]
- Pesti, R.; Kontra, L.; Paul, K.; Vass, I.; Csorba, T.; Havelda, Z.; Várallyay, É. Differential Gene Expression and Physiological Changes during Acute or Persistent Plant Virus Interactions May Contribute to Viral Symptom Differences. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Tugume, A. K.; Mukasa, S. B.; Valkonen, J. P. T. Mixed Infections of Four Viruses, the Incidence and Phylogenetic Relationships of Sweet Potato Chlorotic Fleck Virus (Betaflexiviridae) Isolates in Wild Species and Sweetpotatoes in Uganda and Evidence of Distinct Isolates in East Africa. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Syller, J. Facilitative and Antagonistic Interactions between Plant Viruses in Mixed Infections. Molecular Plant Pathology 2012, 13, 204–216. [Google Scholar] [CrossRef]
- Moreno, A. B.; López-Moya, J. J. When Viruses Play Team Sports: Mixed Infections in Plants. Phytopathology® 2020, 110, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Redinbaugh, M. G.; Stewart, L. R. Maize Lethal Necrosis: An Emerging, Synergistic Viral Disease. Annu. Rev. Virol. 2018, 5, 301–322. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, C.; Wang, C.; Qian, Y.; Li, Z.; Hong, J.; Zhou, X. Further Characterization of Maize Chlorotic Mottle Virus and Its Synergistic Interaction with Sugarcane Mosaic Virus in Maize. Sci Rep 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Bock, C. H.; Barbedo, J. G. A.; Del Ponte, E. M.; Bohnenkamp, D.; Mahlein, A.-K. From Visual Estimates to Fully Automated Sensor-Based Measurements of Plant Disease Severity: Status and Challenges for Improving Accuracy. Phytopathol Res 2020, 2. [Google Scholar] [CrossRef]
- Apriasti, R.; Widyaningrum, S.; Hidayati, W. N.; Sawitri, W. D.; Darsono, N.; Hase, T.; Sugiharto, B. Full Sequence of the Coat Protein Gene Is Required for the Induction of Pathogen-Derived Resistance against Sugarcane Mosaic Virus in Transgenic Sugarcane. Mol Biol Rep 2018, 45, 2749–2758. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Huang, N.; Wang, L.; Ling, H.; Sun, T.; Ahmad, W.; Muhammad, K.; Guo, J.; Xu, L.; Gao, S.; Que, Y.; Su, Y. A Novel L-Ascorbate Peroxidase 6 Gene, ScAPX6, Plays an Important Role in the Regulation of Response to Biotic and Abiotic Stresses in Sugarcane. Front. Plant Sci. 2018, 8, 2262. [Google Scholar] [CrossRef] [PubMed]
- Livak, K. J.; Schmittgen, T. D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Anur, R. M.; Mufithah, N.; Sawitri, W. D.; Sakakibara, H.; Sugiharto, B. Overexpression of Sucrose Phosphate Synthase Enhanced Sucrose Content and Biomass Production in Transgenic Sugarcane. Plants 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Patricio, M.; Camas-Anzueto, J.; Sanchez-Alegría, A.; Aguilar-González, A.; Gutiérrez-Miceli, F.; Escobar-Gómez, E.; Voisin, Y.; Rios-Rojas, C.; Grajales-Coutiño, R. Optical Method for Estimating the Chlorophyll Contents in Plant Leaves. Sensors 2018, 18. [Google Scholar] [CrossRef]
- Fiorentini, M.; Zenobi, S.; Giorgini, E.; Basili, D.; Conti, C.; Pro, C.; Monaci, E.; Orsini, R. Nitrogen and Chlorophyll Status Determination in Durum Wheat as Influenced by Fertilization and Soil Management: Preliminary Results. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, Y.; Zou, W.; Pan, Y.-B.; Lin, P.; Xu, L.; Grisham, M. P.; Ding, Q.; Su, Y.; Que, Y. Genome-Wide Characterization of Sugarcane Catalase Gene Family Identifies a ScCAT1 Gene Associated Disease Resistance. International Journal of Biological Macromolecules 2023, 232, 123398. [Google Scholar] [CrossRef]
- Bwalya, J.; Alazem, M.; Kim, K. Photosynthesis-related Genes Induce Resistance against Soybean Mosaic Virus: Evidence for Involvement of the RNA Silencing Pathway. Molecular Plant Pathology 2022, 23, 543–560. [Google Scholar] [CrossRef] [PubMed]
- Studer, R. A.; Christin, P.-A.; Williams, M. A.; Orengo, C. A. Stability-Activity Tradeoffs Constrain the Adaptive Evolution of RubisCO. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 2223–2228. [Google Scholar] [CrossRef]
- Komínková, M.; Ben Mansour, K.; Komínek, P.; Brožová, J.; Střalková, R. Multiple Infections with Viruses of the Family Tymoviridae in Czech Grapevines. Viruses 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Widyaningrum, S.; Pujiasih, D. R.; Sholeha, W.; Harmoko, R.; Sugiharto, B. Induction of Resistance to Sugarcane Mosaic Virus by RNA Interference Targeting Coat Protein Gene Silencing in Transgenic Sugarcane. Mol Biol Rep 2021, 48, 3047–3054. [Google Scholar] [CrossRef] [PubMed]
- Hidayati, W. N.; Apriasti, R.; Addy, H. S.; Sugiharto, B. Distinguishing Resistances of Transgenic Sugarcane Generated from RNA Interference and Pathogen-derived Resistance Approaches to Combating Sugarcane Mosaic Virus. Indones. J. Biotechnol. 2021, 26. [Google Scholar] [CrossRef]
- Chen, T.-Y.; Pai, H.; Hou, L.-Y.; Lee, S.-C.; Lin, T.-T.; Chang, C.-H.; Hsu, F.-C.; Hsu, Y.-H.; Lin, N.-S. Dual Resistance of Transgenic Plants against Cymbidium Mosaic Virus and Odontoglossum Ringspot Virus. Sci Rep 2019, 9. [Google Scholar] [CrossRef]
- Tatineni, S.; Sato, S.; Nersesian, N.; Alexander, J.; Quach, T.; Graybosch, R. A.; Clemente, T. E. Transgenic Wheat Harboring an RNAi Element Confers Dual Resistance Against Synergistically Interacting Wheat Streak Mosaic Virus and Triticum Mosaic Virus. MPMI 2020, 33, 108–122. [Google Scholar] [CrossRef]
Figure 1.
Percentage of mosaic disease incidence (A) and severity (B) on leaves of sugarcane cultivars grown in field. The incidence and severity were observed in eight cultivars grown in the field of four locations that were Lumajang, Jember, Bondowoso, and Kediri in East Java, Indonesia, and the percentages were determined as described in Materials and Methods.
Figure 1.
Percentage of mosaic disease incidence (A) and severity (B) on leaves of sugarcane cultivars grown in field. The incidence and severity were observed in eight cultivars grown in the field of four locations that were Lumajang, Jember, Bondowoso, and Kediri in East Java, Indonesia, and the percentages were determined as described in Materials and Methods.
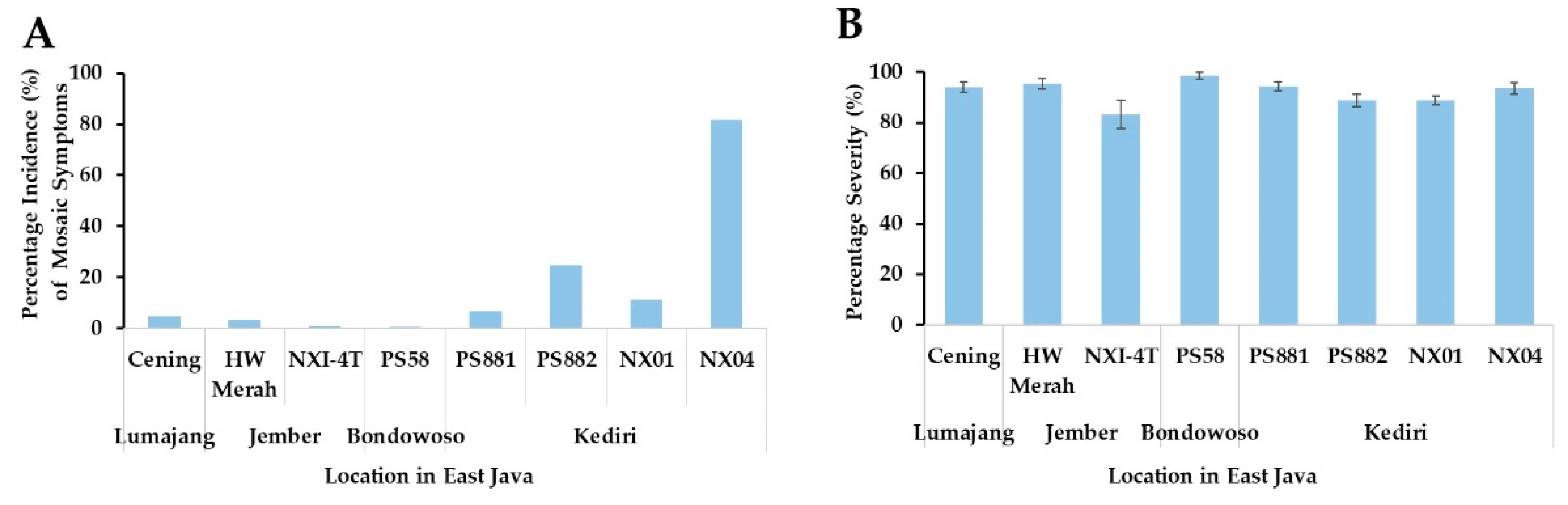
Figure 2.
RT-PCR amplification of the coat protein gene using specific primer for SCSMV (A), SCMV (B) and SrMV (C). RT-PCR was conducted using the RNA isolated from the infected leaves of eight sugarcane cultivars. The image is representing RT-PCR products from each sugarcane cultivars. M,1 kb DNA Ladder (Thermo Fisher Scientific, USA); C, control no symptomatic sugarcane; red asterisk indicating positive CP-SCMV DNA.
Figure 2.
RT-PCR amplification of the coat protein gene using specific primer for SCSMV (A), SCMV (B) and SrMV (C). RT-PCR was conducted using the RNA isolated from the infected leaves of eight sugarcane cultivars. The image is representing RT-PCR products from each sugarcane cultivars. M,1 kb DNA Ladder (Thermo Fisher Scientific, USA); C, control no symptomatic sugarcane; red asterisk indicating positive CP-SCMV DNA.
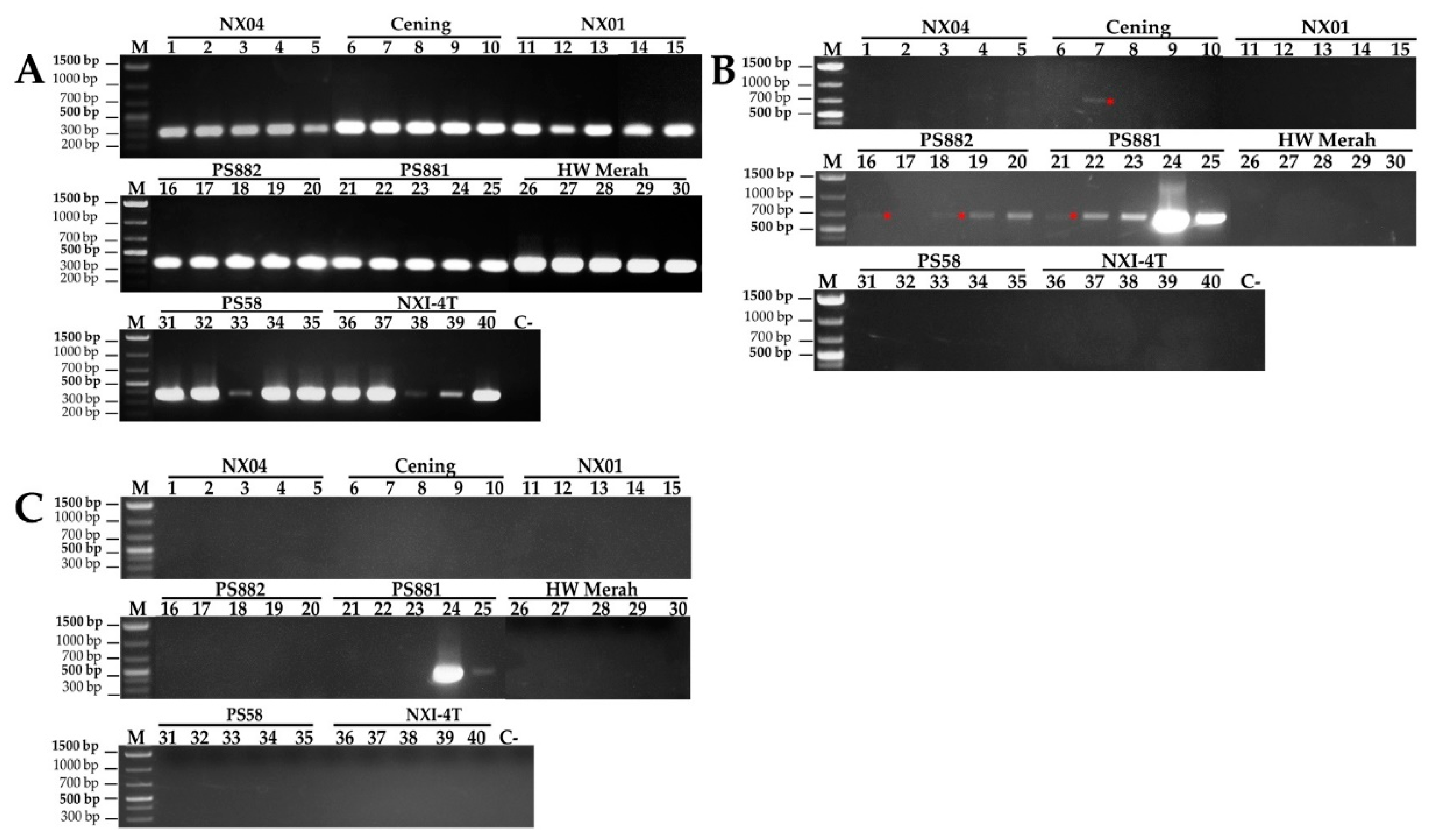
Figure 3.
Phylogenetic analysis of SCSMV (A) SCMV (B) and SrMV (C) based on the CP nucleotide sequences along with retrieved sequences from NCBI. Sequence cluster according to the geographical origin of the virus isolates, such as Thailand (KP987832.1) for SCSMV; USA (MT725538.1) and Germany (X98168.1) for SCMV; and Argentina (EU189037.1), USA (EF078962.1), China (DQ227695.1), and for SrMV.
Figure 3.
Phylogenetic analysis of SCSMV (A) SCMV (B) and SrMV (C) based on the CP nucleotide sequences along with retrieved sequences from NCBI. Sequence cluster according to the geographical origin of the virus isolates, such as Thailand (KP987832.1) for SCSMV; USA (MT725538.1) and Germany (X98168.1) for SCMV; and Argentina (EU189037.1), USA (EF078962.1), China (DQ227695.1), and for SrMV.

Figure 4.
Expression of Apx and Cat gene in response to the infection of mosaic virus in sugarcane leaves. Relative expression of Apx gene in a single infection (A), and Cat and Apx genes in mixed-triple infection in PS881 (B). Expression of β-tubulin was used as the internal control. The expression of Cat and Apx in control, single, double and triple infection were determined using PS881 sugarcane cultivars. Values are means ± SD of two biological replicates.
Figure 4.
Expression of Apx and Cat gene in response to the infection of mosaic virus in sugarcane leaves. Relative expression of Apx gene in a single infection (A), and Cat and Apx genes in mixed-triple infection in PS881 (B). Expression of β-tubulin was used as the internal control. The expression of Cat and Apx in control, single, double and triple infection were determined using PS881 sugarcane cultivars. Values are means ± SD of two biological replicates.
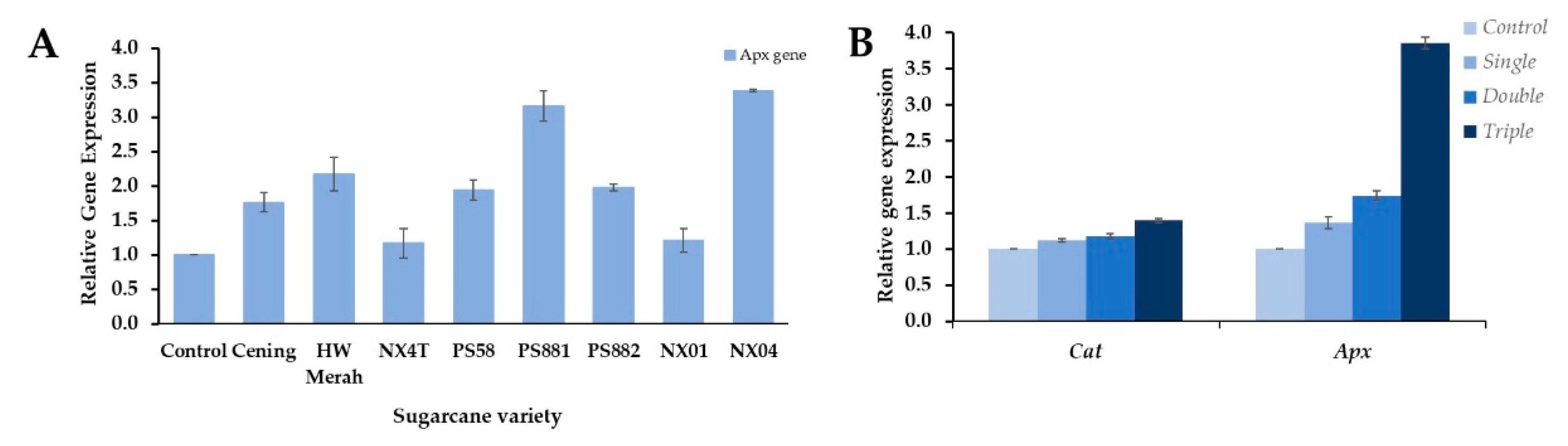
Figure 5.
Chlorophyll content and photosynthetic-related gene expression in response to the mosaic virus infection in sugarcane leaves. Total chlorophyll content (A) and PsaA expression (B) were determined in single infection sugarcane leaves. Expression of the genes PsaA, RbcS, Pepc, Sps1 (C), and levels of PEPC, and Rubisco proteins (D) were determined in control, single, double, and triple infection in PS881 leaves. Values are means ± SD of tree biological replicates. Inset, PEPC and Rubisco (large subunit-RbcL and small subunit-RbcS) proteins were detected by immunoblot analysis. C, S, D, and T represent control, single, double and triple infection, respectively. The intensities of protein bands were quantitively expressed using ImageJ free software (https://image.nih.gov/).
Figure 5.
Chlorophyll content and photosynthetic-related gene expression in response to the mosaic virus infection in sugarcane leaves. Total chlorophyll content (A) and PsaA expression (B) were determined in single infection sugarcane leaves. Expression of the genes PsaA, RbcS, Pepc, Sps1 (C), and levels of PEPC, and Rubisco proteins (D) were determined in control, single, double, and triple infection in PS881 leaves. Values are means ± SD of tree biological replicates. Inset, PEPC and Rubisco (large subunit-RbcL and small subunit-RbcS) proteins were detected by immunoblot analysis. C, S, D, and T represent control, single, double and triple infection, respectively. The intensities of protein bands were quantitively expressed using ImageJ free software (https://image.nih.gov/).
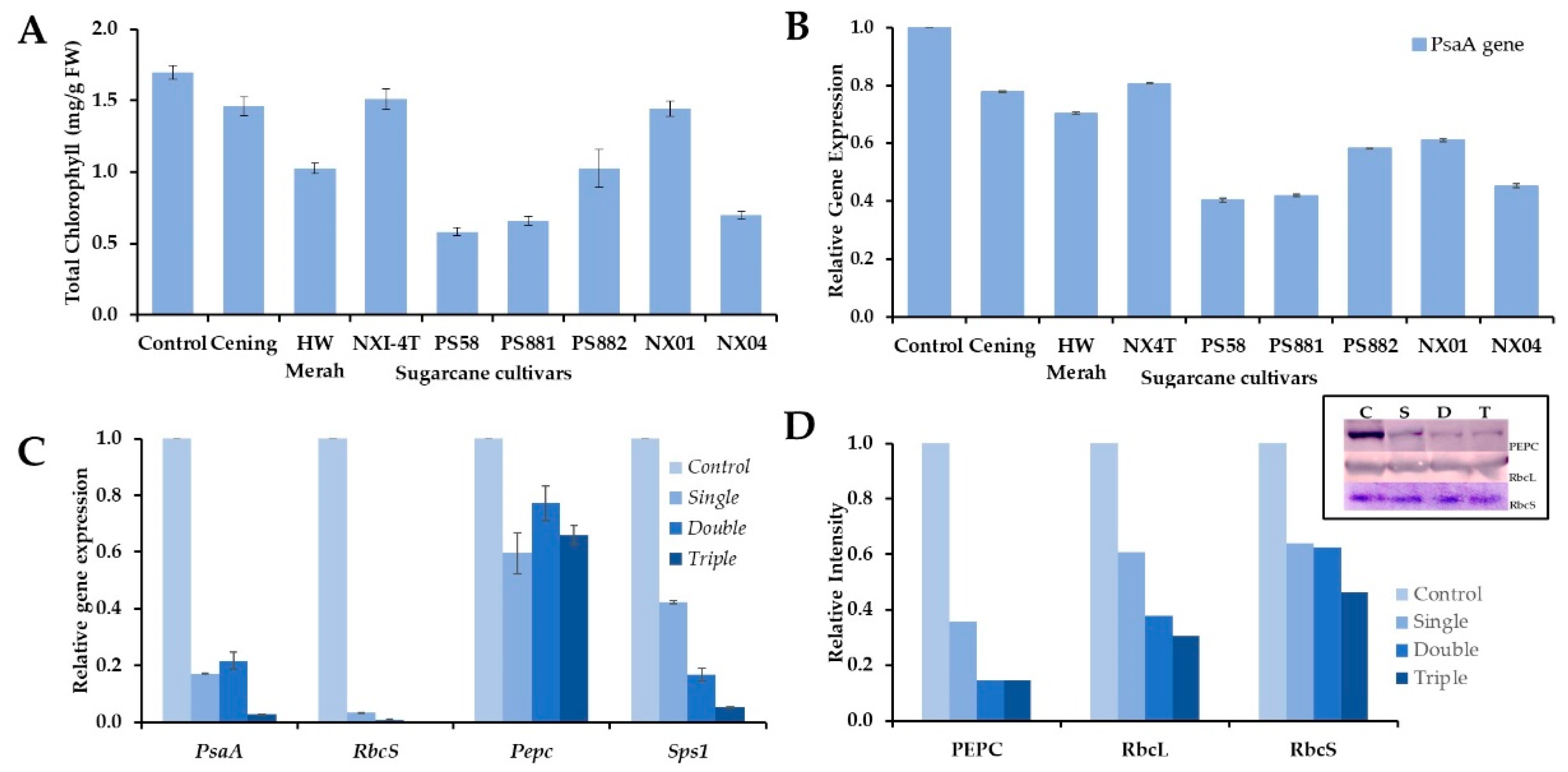

Table 1.
Description of primers used in this study.
| Primer | Sequence information (5’-3’) | Strategy | Amplicon length (bp) | Annealing temperature (°C) | Reference |
|---|---|---|---|---|---|
| CP-SCSMV-F | 5’-MTCTTCATCRGCCGCMTCRATAC-3’ | RT-PCR | 335 bp | 53 | This study |
| CP-SCSMV-R | 5’-AGAACTGAACCCACTTGTACGCC-3’ | ||||
| CP-SCMV-F | 5’-GACATATGGATGTAGATGCTGGTACGACA-3’ | RT-PCR | 735 bp | 58 | [36] |
| CP-SCMV-R | 5’-ATGGATCCTAGTGGTGCTGCTGCACTCCC-3’ | ||||
| CP-SrMV-F | 5’-KCAGATGCTGATGCGAAA-3’ | RT-PCR | 480 bp | 52 | This study |
| CP-SrMV-R | 5’-YGCTTCAGCTGCATCAC-3’ | ||||
| PEPC-F | 5’-TGGGTGGTGACCGTGATGG-3’ | RT-qPCR | 129 bp | 60 | This study |
| PEPC-R | 5’-GCAGCGCCACATAGAGAGC-3’ | ||||
| PsaA-F | 5’-AGGGGCTTATACCCTCAG-3’ | RT-qPCR | 121 bp | 52 | This study |
| PsaA-R | 5’-GGATTAGGTGCCTAACGGAC-3’ | ||||
| RbcS-F | 5’-CACTAGCTTCGCCAAAGT-3’ | RT-qPCR | 205 bp | 55 | This study |
| RbcS-R | 5’-AAGCCTTCCTTGCTGAAC-3’ | ||||
| SPS-F | 5’-GGGTCTCCATAGGACCATTA-3’ | RT-qPCR | 110 bp | 55 | This study |
| SPS-R | 5’-GGGGTGTTATTGTGTGAGTA-3’ | ||||
| CatF | 5’-GCTCAGTTCGACAGGGAACG-3’ | RT-qPCR | 208 bp | 55 | [18] |
| CatR | 5’-CACGTGGATCCCTCAAGGTC-3’ | ||||
| ApxF | 5’-GATTTGATTGCCGTGGCTGG-3’ | RT-qPCR | 134 bp | 55 | [37] |
| ApxR | 5’-TCTTCAGGAAGTTTGCCAGTTG-3’ | ||||
| Β-tubulin | 5’-CATCTTCGTGTGGAATCCT-3’ | RT-qPCR | 129 bp | 55 | This study |
| Β-tubulin | 5’-AACTCCAGAACAGACCGTA-3’ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



