
Submitted:
29 April 2024
Posted:
30 April 2024
You are already at the latest version
Abstract
The responses of inflammatory cytokines and brain-derived neurotrophic factor (BDNF) to a high-intensity interval training (HIIT) session were compared between 7 postmenopausal (57±2 y) and 8 premenopausal females (44±3 y) during the luteal phase of their menstrual cycle, when estrogen and progesterone are highest, and. HIIT consisted of 10 x 1 min running intervals (90% maximum heart rate) separated by 1 min recovery intervals. Blood samples were collected at baseline (fasted), pre-exercise (postprandial), and 0, 30 and 90 min post-HIIT and analyzed for interleukin (IL-6) and 10 (IL-10), tumour necrosis factor-alpha (TNF-α) and BDNF. IL-6 significantly increased from pre-exercise to 0 min post-HIIT in postmenopausal (+40%, p=0.01) but 30 min post-HIIT in premenopausal females (+60%, p=0.02). IL-6 remained elevated at 90 min post-HIIT in premenopausal (+104%, p=0.05) and to a higher degree in postmenopausal (+385%, p<0.001). IL-10 showed no response. TNF-α increased from pre- to 0 min post-HIIT (+10%, p=0.05), then decreased to below pre-exercise at 30 min (-10%, p=0.02) and 90 min (-5%, p=0.04) in both groups. BDNF increased immediate post-HIIT in premenopausal (+60%, p<0.001) but not postmenopausal females. The differences in IL-6 and BDNF response to HIIT between pre and postmenopausal females provide evidence of the role of female reproductive hormones in the regulation of these exercise-induced responses.
Keywords:
Menopause
; high-intensity interval training exercise
; interleukin 6
; interleukin 10
; tumour necrosis factor-alpha
; brain-derived neurotrophic factor
1. Introduction
Menopause is a pivotal time in female physiology characterized by the cessation of the menstrual cycle and the concomitant reductions in the female hormone concentrations [1]. One of the multiple physiological changes that females experience following menopause is the increase in circulating inflammatory cytokines, which have been typically attributed to the decrease in estrogen (estradiol in its most form in humans) concentrations [2]. Indeed, estrogen deficiency has been associated with increased expression and secretion of pro-inflammatory cytokines such as interleukins 1-beta (IL-1β) and tumour necrosis factor-alpha (TNF-α), as well as interleukin 6 (IL-6), which is considered both pro- and anti-inflammatory [3], suggesting an interaction between estrogen and these cytokines [4]. This is concerning as chronic inflammation, commonly shown by elevated resting concentrations of pro-inflammatory cytokines has been associated with hypertension, atherosclerosis, ischemic stroke, and heart disease [5,6,7]. In contrast, chronic exercise is associated with overall anti-inflammatory effects, which can mitigate chronic inflammatory conditions [8], which has been attributed to the transient spikes in cytokine concentrations following acute exercise bouts [9,10]. Specifically, acute exercise triggers changes in the circulating concentrations of IL-6, IL-1β and TNF-α, and anti-inflammatory cytokines such as interleukin 10 (IL-10), all of which typically peak immediately after exercise and return to baseline within 5 to 24 hours [11,12]. However, the extent and duration of these exercise-induced changes depend on various factors such as the exercise intensity, duration, and frequency [11,13], as well as on the individual’s training status [14], biological sex [15,16] and age [17,18]. Thus, it is also possible that the reductions in estrogen concentrations postmenopause, which has been previously shown to be associated with an increase in pro-inflammatory cytokines [3], could alter the magnitude and timing of these exercise-induced factors. However, studies in postmenopausal females examining the acute responses of these cytokines to exercise are lacking.
Moreover, menopause has been found to play a role in cognitive decline along with the progression of neurodegenerative diseases associated with low concentrations of brain-derived neurotrophic factor (BDNF) which is a crucial molecule involved in brain plasticity and cognition [19]. BDNF concentrations also increase post-exercise in an intensity-dependent manner [20,21,22,23] but most of these exercise studies are in male populations. According to a systematic review in older rodents, there are potential sex-based differences in the BDNF response to exercise, with healthy-aged or ovariectomized females exhibiting overall greater increases and cognitive improvements compared to healthy-aged males [24], suggesting female reproductive hormones may influence exercise-induced changes in BDNF. On the other hand, human studies examining the relationship between BDNF, and endogenous ovarian hormones have produced conflicting results due to large participant heterogeneity, and a lack of cycle phase standardization across studies [25,26]. Additionally, these studies only apply to resting conditions as there are no studies in postmenopausal females involving the response of BDNF to exercise. Since postmenopausal females have lower resting BDNF concentrations compared with premenopausal groups [26], the lack of information on how BDNF responds to acute exercise in these individuals is concerning. Examining the effects of various exercise modalities on BDNF in postmenopausal females is critical for prescribing exercise programs to manage menopause-induced chronic inflammation and cognitive decline.
The purpose of this secondary analysis was to compare the potential responses of IL-6, IL-10, TNF-α, and BDNF to a single high-intensity interval training (HIIT) exercise trial between postmenopausal females with low concentrations of reproductive hormones and premenopausal females during the luteal phase of their menstrual cycle when estrogen and progesterone are highest. Examining how exercise influences inflammatory cytokines and BDNF in postmenopausal females compared to premenopausal counterparts could provide valuable insights into the role of hormones in modulating exercise-induced responses and inform strategies for promoting health and wellness in women across the lifespan.
2. Materials and Methods
2.1. Participants
This study involves a secondary analysis of blood samples collected from a study examining the role of ovarian hormones in appetite regulation in pre versus postmenopausal females. Specifically, samples from 8 premenopausal females (44 ± 3 y, eumenorrheic, not currently taking oral contraceptives) and 7 postmenopausal females (57 ± 2 y, absence of menses for >12 months) recruited to participate in the original study were included in the present analysis. To be included in the study, participants were assessed as being healthy by the Get Active Questionnaire [27], were considered recreationally active (i.e., exercising for approximately 150 min of moderate to vigorous physical activity per week), were not taking any medications, and were free of injuries and other contraindications to exercise.
Premenopausal participants were included if they were non-hormonal contraceptive users and had no metabolic diseases such as diabetes. In addition, participants were excluded if they were pregnant, had been pregnant (for >3 months) within the past 3 years, or had plans to become pregnant during the time of study participation. They also had to have tracked their cycle for at least two months prior to participation. Participants were provided with a thermometer (HealthSmart® International) and instructed to monitor body temperature orally upon awakening each day following the onset of menstruation with a sustained increase of 0.3°C, indicative of ovulation. The temperature was reported daily to the researcher via text/email. Participants were also provided with ovulation kits (Easy@Home, Easy Healthcare Corporation, IL, USA), and asked to report the result via text/email to the researchers, to confirm surges in luteinizing hormone (LH). Testing during the luteal phase occurred 2-5 days following identification of ovulation. Post-menopausal females had no experience with hormonal replacement therapy (including bilateral oophorectomy in which both ovaries have been removed), had an absence of menses for >12 months, and an estradiol concentration of less than 108 pmol/L and a progesterone concentration of less than 4 nmol/L [28,29].
The study was conducted per the Declaration of Helsinki and received ethics approval from the Wilfrid Laurier University Research Ethics Board (REB #6900). Ethics approval for the secondary analysis was also received from the Brock University Research Ethics Board (REB #23-260). All participants agreed to participate in the study by signing informed consent. Experimental procedures took place in the Energy Metabolism Research Laboratory of Wilfrid Laurier University.
2.2. Familiarization
All participants completed a familiarization session during which they provided written informed consent and became acclimated with the laboratory. Body mass and height were measured using standard procedures (Health-o-meter Professional, Sunbeam Products Inc., Illinois, USA). Participants then completed a graded exercise test to exhaustion on a motorized treadmill (4Front, Woodway, WI, USA) to determine VO2max where oxygen consumption (VO2) and carbon dioxide production (VCO2) were measured continuously with an online gas collection system (MAX-II, AEI Technologies, PA, USA) and a silicon facemask (7400 series Vmask, Hans Rudolph Inc., KS, USA as previously described [30]. Participants began the test with a 5-min warm-up walking at between 4.8-8 km/h followed by fast walking or running at a self-selected pace between 6.4 and 11.2 km/h with incremental increases in grade (2%) applied every 2 min until volitional fatigue. Heart rate (HR) was continuously recorded using an integrated HR monitor (FT1, Polar Electro; QC, Canada). Following completion of the test and a 5 min walking cool-down period (4.8-8 km/h), a verification phase was completed to volitional fatigue where participants fast walked or ran to exhaustion at 105% of the speed and grade used to confirm V̇O2max [30]. Following the VO2max test and verification phase, the target intensity for the HIIT session (90% HRmax) was calculated using the ACSM running equation [31] using the speed and grade from the VO2max test and VO2 data. Participants completed two practice 1 min intervals (treadmill speed eliciting 90% HRmax) interspersed with 1 min rest to confirm the calculated exercise intensity was correct. Finally, at the end of the familiarization session, participants were asked if they had any food allergies that may prevent them from consuming the planned standardized breakfast during the HIIT session.
Premenopausal participants were provided with the ovulation kits before leaving the familiarization and asked to report the result via text/email to the researchers to determine the appropriate session timing. In addition, premenopausal participants were also asked to monitor and report body temperature upon awakening as increases of ~0.3°C indicated ovulation [28]. The premenopausal HIIT session was scheduled for ~7 d following a positive LH surge on the ovulation strips. The menstrual phase was confirmed to be within the luteal phase through a fasted blood sample of estradiol (E2) between 294-918 pmol/L and progesterone (P4) concentration >16 nmol/L [28].
2.3. Experimental Session
Participants arrived at the laboratory at 0800 h after a 12-hour overnight fast (i.e., only water after 2000 h). Moderate to vigorous activity and alcohol consumption were restricted for 24 h while caffeine consumption was restricted for 12 h before the session. A fasted blood draw (baseline sample) was taken at ~0815 h followed by a standardized breakfast (breakfast smoothie; 7 kcal/kg; 56% carbohydrate, 23% fat, and 21% protein; Gruppo, ON, Canada) at 0815-0830 h. During breakfast, participants were asked to fill out a short questionnaire about their typical sleep patterns, including how much sleep they received the night before coming into the lab. Following breakfast, participants were allowed 30 min for digestion and a subsequent blood sample was taken at ~0900 h (pre-HIIT sample). From 0900-0930 h, participants performed a HIIT exercise protocol which began with a 5-minute walking or jogging warmup at a self-selected pace between 4.8 and 8.0 km/h on the motorized treadmill (see above). Participants then started the HIIT protocol which consisted of 10 x 60 s bouts of running at a speed and grade that would elicit 90% of their maximum heart rate followed by 60 seconds of recovery. Heart rate was monitored throughout the session. Following the completion of the HIIT protocol, participants completed a 5-min walking cooldown at a self-selected pace. Upon completion of the cooldown, the first post-HIIT blood sample was taken at ~0930 h (0 min post-HIIT sample) and subsequent post-HIIT blood sampling occurred at ~1000 h (30 min post-HIIT sample), and 1100 h (90 min post-HIIT sample).
2.4. Blood Collection and Analysis
Approximately 3 mL of whole blood was collected into pre-chilled Vacutainer SST tubes (gold top) from each participant’s median cubital vein in the antecubital fossa while participants were lying in a supine position using a standard venipuncture technique during each session at each time point (baseline, pre-exercise, and 0 min, 30 min, and 90 min post-HIIT). Blood samples were inverted 8-10 times before sitting for 30 min at room temperature. The samples were then centrifuged at 3000g for 10 min at 4° before the serum was aliquoted into microcentrifuge tubes and stored at -80°C until analysis. In the present study, serum was used to measure circulating concentrations of IL-6, IL-10, TNF-α and BDNF.
Although this was a secondary analysis of samples from another study, the cytokines reported herein were analyzed for the first time for the present study using new commercially available immunoassay assay kits. Specifically, IL-6, IL-10 and TNF-α were measured with ELLA single assay kits (cat. # SPCKBPS-003028, ST01B-PS-000276 and ST01B-PS-002803, respectively; ProteinSimple, San Jose, CA). The inter-assay coefficients of variation were 9.9% for IL-6, 6.6% for IL-10 and 8.7% for TNF-α. The intra-assay coefficients of variation for IL-6, IL-10 and TNF-α were 4.6%, 6.69% and 2.54%, respectively. BDNF was analyzed using an ELISA kit (cat. #CYT306; EMD Millipore) with an inter-assay coefficient of variation of + 8.5 %, and an intra-assay coefficient of variation of + 3.7%.
2.5. Statistical Analysis
The data were assessed for normality using the Shapiro–Wilk test, and z-scores for skewness and kurtosis. There were a few missing values for each analyte due to low serum availability or non-detectable values, so the number of participants is different for each marker. A series of two-way repeated measures analysis of variance (RM-ANOVA) was used to examine the main effects of group (premenopausal versus postmenopausal) and time (5 points), as well as the time-by-trial interactions, for IL-6, IL-10, TNF-α, and BDNF. Statistical significance was set at p0.05. Effect sizes, including partial eta squared (η2) for ANOVA and Cohen’s d for post hoc comparisons, were calculated and interpreted using the Cohen criteria: 0.01 = small, 0.06 = moderate, 0.14 = large effect for partial η2, and 0.2 = small, 0.5 = medium, 0.8 = large effect for Cohen’s d [32,33]. The statistical analysis was performed using IBM SPSS Statistics 28 (SPSS Inc., Chicago, IL, USA).
3. Results
Participants in the premenopausal (44 ± 3 y) and postmenopausal (57 ± 2 y) groups had similar BMI (28 ± 5 kg/m2 and 28 ± 2 kg/m2, respectively), VO2max (29.2 ± 7.4 mL/kg/min and 30.2 ± 6.5 mL/kg/min, respectively), and reported similar hours of sleep both habitually (6.1 ± 0.6 h and 6.1 ± 1.5 h, respectively) and before the sessions (5.0 ± 2.0 h and 5.4 ± 2.4 h, respectively). As expected, the premenopausal females had significantly higher concentrations than the postmenopausal females of both estradiol (E2 = 137.3 ± 31.9 SD vs. 56.7 pmol/L, respectively; p < 0.001) and progesterone (P4 = 19.7 ± 4.8 vs. 0.60 ± 0.27 nmol/L). During HIIT, there were no significant differences between the premenopausal and postmenopausal females in either the average speeds (7.6 ± 1.6 km/h and 6.9 ± 0.5 km/h, respectively). Blood lactate concentrations were also not different (p > 0.05) between groups (Table 1). However, there was a time-by-group interaction (F = 7.51, p = 0.059, η2 = 0.16) as the increase of blood lactate from pre-exercise to 0 min post-HIIT was of a greater magnitude in postmenopausal compared to premenopausal females (+51% vs. +185%, respectively). Additionally, although in the premenopausal group, lactate concentrations returned to pre-exercise levels by 30 min post-HIIT and to the fasted levels by 90 min following HIIT, in the postmenopausal group, lactate concentrations remained higher than pre-exercise at 30 min (+46%) returning to pre-exercise 90 min later (Table 1).
IL-6 showed a significant group-by-time interaction (F = 10.9; p = 0.003; η2 = 0.52). Compared to pre-exercise, the IL-6 increase at 0 min post-HIIT was not significant (+18%, p = 0.189, d = 0.62) but it was significant at 30 min (+60%, p = 0.02, d = 1.02) and 90 min (+104%, p = 0.05, d = 2.19) post-HIIT in the premenopausal group. In contrast, IL-6 increased significantly from pre-exercise to 0 min post-HIIT (+40%, p = 0.01, d = 0.55) in the postmenopausal group and its increase from pre-exercise to 30 min (+80%, p < 0.001, d = 0.48) and 90 min post-HIIT (+385%, p < 0.001, d = 3.18) was of greater magnitude (2 to 3-fold) than in the premenopausal group (Figure 1).
For IL-10 (Figure 2), there was no group-by-time interaction (F = 0.81; p = 0.49; η2 = 0.0.8) and no main effect for time (F = 2.02; p = 0.11; η2 = 0.18) or group (F = 1.65; p = 0.23; η2 = 0.15).
There was no group-by-time interaction for TNF-α (F = 0.37; p = 0.73; η2 = 0.39). There was a main effect for time (F = 4.51; p = 0.018; η2 = 0.33), reflecting a 10% (p = 0.05, d = 0.38) increase in TNF-α from pre-exercise to 0 min post-exercise, which was followed by a decrease at 30 min (10%, p = 0.02, d = 0.26) and 90 min (5%, p = 0.04, d = 0.14) post-exercise to concentrations below pre-exercise, in both groups (Figure 3).
For BDNF, there was a significant group-by-time interaction (F = 7.998; p < 0.001; η2 = 0.25), reflecting an increase in BDNF at 0 min following HIIT in the premenopausal group (+69%, p < 0.001, d=1.56) but not in the postmenopausal group (-56%, p > 0.99, d=1.2 Figure 4). This was followed by +38% at 30 min (p = 0.94; d = 1.0) and -17% at 90 min (p > 0.99; d = 0.23) compared to pre-exercise in the premenopausal group and -31% at 30 min (p > 0.99; d = 0.52) and -3% 90 min (p > 0.99; d = 0.05) in postmenopausal group.
4. Discussion
This study provides valuable insights into the distinct acute response of exercise-induced factors following a single bout of HIIT in postmenopausal compared to premenopausal females. Specifically, postmenopausal participants exhibited an immediate increase in IL-6 concentrations following HIIT, whereas premenopausal females experienced a delayed spike in IL-6 30 min following HIIT. These effects were of large effect size and suggest that the time course of the increase in IL-6 following HIIT was different between groups. In contrast, no significant exercise-induced changes were observed in the IL-10 in either group while TNF-α concentrations slightly increased immediately after HIIT and then decreased to below pre-exercise concentrations 90 min post-HIIT in both groups. This suggests that the exercise-induced IL-10 and TNF-α response in females may be less influenced by ovarian hormonal status compared to IL-6. Moreover, BDNF concentrations increased after HIIT only in the premenopausal group, with no apparent response observed in the postmenopausal group. This difference in the BDNF response to HIIT, which was of large effect size, further underscores the potential influence of female reproductive hormones on BDNF regulation in response to exercise.
Greater concentrations of circulating inflammatory cytokines, leading to a higher risk of metabolic syndrome, are often seen in postmenopausal women due to the increase in visceral fat [34]. Considering the pre and postmenopausal females in this study had similar BMI, the similarity in their fasted and pre-exercise concentrations of circulating IL-6, IL-10 and TNFα was not surprising. However, the time course of the IL-6 response to HIIT was different between groups with concentrations being elevated immediately post-HIIT in the postmenopausal group the elevation was at 30 min post-HIIT in the premenopausal group. IL-6 remained elevated up to 90 min following HIIT in both groups, but this increase was of almost 3-fold greater magnitude in the postmenopausal group. We previously found IL-6 to be elevated compared to its pre-exercise concentrations 1 h after high-intensity running and cycling (39%, p<0.05) in young adult females [35], which supports the current finding of the IL-6 elevation 30 and 90 min following HIIT in both groups. However, we cannot explain mechanistically why the postmenopausal group had an earlier significant increase in IL-6 (i.e., immediately) following the HIIT protocol, or why 90 min later they had a much larger IL-6 increase compared to their premenopausal counterparts. The main suggestion is that the difference in the timing and magnitude of the IL-6 response is related to the low concentrations of female reproductive hormones, especially estrogen deficiency which has been previously associated with elevated concentrations of circulating inflammatory cytokines [2,36]. This speculation is based on the fact that the immediate exercise-induced response seen in postmenopausal females aligns with previous research in young adult males who also demonstrate exercise-induced increases in the circulating IL-6 immediately following different modes of a single bout of high-intensity interval exercise, although, in those studies, IL-6 returned to pre-exercise levels after 60 minutes [37,38]. In another study, however, both IL-6 and TNF-α increased immediately following an acute bout of high-intensity intermittent walking or continuous moderate-intensity walking in young adult males and remained elevated until at least 4 h post-exercise [39]. Thus, the different time course and magnitude of the IL-6 response to the current running protocol between pre and postmenopausal women suggests a direct relationship between IL-6 and female reproductive hormones.
Furthermore, the exercise-induced increase in IL-6 is typically directly related to exercise intensity, duration, muscle mass recruited, and muscle glycogen used [40]. While there were no differences between the pre and postmenopausal females in the HIIT speeds, the exercise-induced relative increase in blood lactate concentration at 0 min post-HIIT was of a greater magnitude in the postmenopausal females compared to the premenopausal females, and remained higher than pre-exercise until 30 min post-HIIT and higher than fasted levels 90 min following HIIT. These data suggest a higher lactate production and slower lactate clearance in the postmenopausal group. This is consistent with the aging-related decrease in oxidative muscle fibres, and increased dependency on carbohydrate energy production, which alters lactate production and kinetics during exercise and subsequently IL-6 secretion from muscle [40]. This lactate effect on the IL-6 release from skeletal muscle was shown in young adult males during high-intensity interval cycling and was confirmed using electrically stimulated muscle cell cultures [41]. Thus, further research is required to understand why the IL-6 increase in the postmenopausal group may appear immediately post-exercise (similar as in men) and continue to increase for 90 min post-exercise at a higher magnitude than what was observed in the premenopausal group.
The increase in TNF-α immediately following the HIIT session is in line with our previous work where TNF-α increased immediately following a high-intensity interval running session (10%, p < 0.05) in young adult females and males, but in that case, its concentrations at 1 h and 24 h post-exercise were no different than at pre-exercise, [35]. The present study also provides a novel finding that after this initial exercise-induced increase, TNF-α was significantly decreased at 30 and 90 min post-HIIT to concentrations slightly below pre-exercise. According to previous studies, exercise-induced increases in TNF-α may take as long as 24 to 48 h to return to pre-exercise levels following intense resistance training [42] and after a marathon race [43] in adult males 18-55 y. Other studies examining the response of TNF-α following exercise have also observed no change, which has been attributed to the suppression of TNF-α by the muscle-derived, anti-inflammatory IL-6 [44], which is consistent with the elevated IL-6 for up to 90 min after our HIIT protocol. Thus, the later decrease in TNF-α found herein is important as it could indicate a healthy balance of pro- versus anti-inflammatory status post-exercise. Indeed, post-exercise inflammation is a natural response of the immune system to tissue damage caused by physical exertion, but excess or prolonged inflammation can be harmful and slow down the recovery process. Therefore, it is important to balance the increase of pro-inflammatory cytokines with an increase in anti-inflammatory cytokines. IL-10 is such cytokine that plays a critical role in the anti-inflammatory properties, is mediated by IL-6, and is produced during exercise [45]. Indeed, in our previous study [35], IL-10 was elevated 5 min after running (20%, p < 0.05) and 1 h after both running and cycling (41% and 64%, respectively, p < 0.05) in both young adult females and males. In the present study, IL-10 did not change in response to HIIT, which somewhat contradicts the suggestion that IL-6 mediates the IL-10 response [45] since IL-6 did increase in both our groups. Because the previous study was performed on younger participants (18-25 y) it is possible that in the present study, the IL-10 response happened after the 90-min study period since IL-6 continued to increase further at 30 and 90 min following HIIT. Later blood draws, at 120- or 150-min post-exercise could potentially show whether IL-10 would increase in response to the substantial increase in IL-6 and would also help us understand the kinetics of the IL-6 response, i.e., how much longer it would take to return to baseline. In any case, corroborating with previous studies [35,42,45,46,47], both inflammatory cytokines (IL-6 and TNF-α) acutely increased following HIIT in our pre and postmenopausal females.
BDNF was originally characterized for its role in the brain for enhancing neuronal plasticity which is essential for learning and memory [19,48]. Additionally, BDNF is thought to play a role in central and peripheral molecular processes of energy metabolism and homeostasis [49]. Engaging in physical exercise has consistently demonstrated an elevation in the expression of BDNF in brain regions such as the hypothalamus, striatum, and various cortical areas [20,50,51]. In this study, we found BDNF to increase by 69% from pre-exercise to 0 min post-HIIT, and, although non-significantly, to remain 38% higher than pre-exercise at 30 min post-HIIT in premenopausal females but not in the premenopausal group. An increase in BDNF has also been reported after high-intensity exercise in groups of females and males aged 20–50 y [20,21,22,23,51,52]. However, exercise studies in postmenopausal females are lacking. Only one previous study examined the BDNF response to a single bout of 30-min aerobic exercise on the treadmill between pre and postmenopausal women and found that exercise did not lead to a significant change in BDNF at 0 and 30 h post-exercise in either group [53]. In their study, there was a large difference in average ages between the premenopausal (~28 y) and postmenopausal (~57 y) participants, making it difficult to conclude whether the difference in the BDNF response to HIIT was due to female reproductive hormones. On the other hand, resting BDNF concentrations have been reported to be lower in postmenopausal females compared to eumenorrheic premenopausal females and this difference has been attributed to their lower estradiol and progesterone as it was restored by a 6-month hormonal replacement treatment [54]. Herein we found no differences in resting BDNF concentrations probably because the age of our groups was not as far apart as in the study by Begliuomini et al. (2007)[26] as their premenopausal participants were in their early 20s. Indeed, in their study, BDNF was positively correlated with estradiol and progesterone and negatively correlated with menopausal age. Our study strengthens this claim on the role of female reproductive hormones on the BDNF regulation postmenopause by closing the age gap between the premenopausal and postmenopausal groups (44 ± 3 vs. 57 ± 2 y, respectively). However, other factors related to aging and neurodegenerative processes may still contribute to the observed differences. For example, in our previous study of young adult males (23.1 ± 3.0 y), who have very low estrogen and progesterone, we also found an acute exercise-induced increase following bouts of moderate and vigorous intensity, and high-intensity interval training [23]. Thus, further research is needed to elucidate the mechanisms underlying the differential BDNF response to exercise in postmenopausal females and better understand the role of reproductive hormones. Understanding these mechanisms could have implications for developing targeted interventions to promote brain health and cognitive function across the lifespan.
Acknowledging limitations is crucial for interpreting the findings of any study. Since this was a secondary analysis of available samples from another study, the minimum sample size of 12 participants (6 per group) was calculated to detect differences in appetite hormones using G*Power-2 with a large effect size of 0.467 [55,56,57]. We were still able to reach significance in IL-6, TNFα and BDNF with this sample which was comparable to that of a previous study examining post-exercise changes in BDNF [23]. However, we did have fewer participants than our previous study detecting changes in IL-10 [35]. Not accounting for changes in plasma volume during the exercise period is another potential limitation, as it could influence the observed changes in circulating markers. However, the plasma volume changes we have previously measured immediately after high-intensity interval exercise in females ranged between -3.82% (in running) to -6.68% (in cycling) (Kouvelioti et al. 2019). The substantial differences in the percent changes of IL-6 (ranging from 40% to 385%) and BDNF (60%) observed following HIIT indicate responses beyond the known effects of exercise-induced hemoconcentration, as evidenced by the modest changes in plasma volume previously reported (-3.82% % to -6.68%) post-HIIT [35]. Therefore, the study provides valuable insights despite these limitations.
In conclusion, the comparison of pre and postmenopausal females’ responses to a session of high-intensity interval exercise provides evidence of potential differences in inflammatory and neurotrophic responses based on hormonal status, emphasizing the importance of considering hormonal status in future exercise investigations and its impact on health outcomes in women across different stages of life. Moving forward, future studies with larger sample sizes and more comprehensive methods for measuring changes in plasma volume could further elucidate the observed differences and contribute to a better understanding of the relationship between age, hormonal status, exercise, and circulating markers related to inflammation and brain health.
Author Contributions
Conceptualization, P.K., R.E.K.M. and T.J.H.; methodology, T.J.H., P.K., R.E.K.M., J.A.L.T., S.F.M.; validation, X.X., Y.Y. and Z.Z.; formal analysis, A.G., A.M., P.K., and R.E.K.M.; investigation, A.G, A.M., M.I.R., J.A.L.T., D.P.D.B., S.F.M.; resources, P.K., R.E.K.M. and T.J.H.; writing—original draft preparation, A.G., M.I.R. and P.K.; writing—review and editing, T.J.H., R.E.K.M., J.T., and S.M.; visualization, P.K. and R.E.K.M.; supervision, P.K., T.J.H. and R.E.K.M.; project administration, P.K. and T.J.H.; funding acquisition, P.K., R.E.K.M. and T.J.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Sciences and Engineering Research Council of Canada (NSERC); grant numbers RGPIN-2023-03572 (P.K.), RGPIN-2016-06118 (T.J.H.), and RGPIN-2017-03904 (R.E.K.M).
Institutional Review Board Statement
The study was conducted per the Declaration of Helsinki and received ethics approval from the Wilfrid Laurier University Research Ethics Board (REB #6900). Ethics approval for the present secondary analysis was also received from the Brock University Research Ethics Board (REB #23-260). All participants agreed to participate in the study by signing informed consent. All experimental procedures took place in the Energy Metabolism Research Laboratory of Wilfrid Laurier University.
Informed Consent Statement
Informed consent was obtained from all participants involved in the study.
Data Availability Statement
The raw data supporting the findings of this study are available from the corresponding author PK upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nelson, H. D. (2008). Menopause. The Lancet, 371(9614), 760–770. [CrossRef]
- Malutan, A. M., Dan, M., Nicolae, C., & Carmen, M. (2014). Proinflammatory and anti-inflammatory cytokine changes related to menopause. Przegla̜d Menopauzalny = Menopause Review, 13(3), 162–168. [CrossRef]
- Pacifici, R., Brown, C., Puscheck, E., Friedrich, E., Slatopolsky, E., Maggio, D., McCracken, R., & Avioli, L. V. (1991). Effect of surgical menopause and estrogen replacement on cytokine release from human blood mononuclear cells. Proceedings of the National Academy of Sciences of the United States of America, 88(12), 5134. [CrossRef]
- Pfeilschifter, J., Köditz, R., Pfohl, M., & Schatz, H. (2002). Changes in Proinflammatory Cytokine Activity after Menopause. Endocrine Reviews, 23(1), 90–119. [CrossRef]
- Caiazzo, E., Sharma, M., Rezig, A. O., Morsy, M., Ialenti, A., Pellicori, P., Bruzzese, D., Maffia, P., & Guzik, T. J. (2023, July). Inflammatory Cytokines and Risk of Developing Hypertension: A Systematic Review and Meta-Analysis. 19th World Congress of Basic & Clinical Pharmacology (WCP2023), Glasgow, Scotland. [CrossRef]
- Su, J.-H., Luo, M.-Y., Liang, N.-, Gong, S.-X., Chen, W., Huang, W.-Q., Tian, Y., & Wang, A.-P. (2021). Interleukin-6: A Novel Target for Cardio-Cerebrovascular Diseases. Frontiers in Pharmacology, 12, 745061. [CrossRef]
- Feghali, C. A., & Wright, T. M. (1997). Cytokines in acute and chronic inflammation. Frontiers in Bioscience: A Journal and Virtual Library, 2, d12-26. [CrossRef]
- Docherty, S., Harley, R., McAuley, J. J., Crowe, L. A. N., Pedret, C., Kirwan, P. D., Siebert, S., & Millar, N. L. (2022). The effect of exercise on cytokines: Implications for musculoskeletal health: a narrative review. BMC Sports Science, Medicine and Rehabilitation, 14(1), 5. [CrossRef]
- Allen, J., Sun, Y., & Woods, J. (2015). Exercise and the Regulation of Inflammatory Responses. Progress in Molecular Biology and Translational Science, 135. [CrossRef]
- Gleeson, M., Bishop, N. C., Stensel, D. J., Lindley, M. R., Mastana, S. S., & Nimmo, M. A. (2011). The anti-inflammatory effects of exercise: Mechanisms and implications for the prevention and treatment of disease. Nature Reviews Immunology, 11(9), 607–615. [CrossRef]
- Azizbeigi, K., Azarbayjani, M. A., Atashak, S., & Stannard, S. (2015). Effect of Moderate and High Resistance Training Intensity on Indices of Inflammatory and Oxidative Stress. Research in Sports Medicine (Print), 23, 73–87. [CrossRef]
- Cerqueira, É., Marinho, D. A., Neiva, H. P., & Lourenço, O. (2020). Inflammatory Effects of High and Moderate Intensity Exercise—A Systematic Review. Frontiers in Physiology, 10, 1550. [CrossRef]
- Zwetsloot, K. A., John, C. S., Lawrence, M. M., Battista, R. A., & Shanely, R. A. (2014). High-intensity interval training induces a modest systemic inflammatory response in active, young men. Journal of Inflammation Research, 7, 9–17. [CrossRef]
- Kapilevich, L. V., Zakharova, A. N., Kabachkova, A. V., Kironenko, T. A., & Orlov, S. N. (2017). Dynamic and Static Exercises Differentially Affect Plasma Cytokine Content in Elite Endurance- and Strength-Trained Athletes and Untrained Volunteers. Frontiers in Physiology, 8, 35. [CrossRef]
- Kraemer, W. J., & Ratamess, N. A. (2005). Hormonal Responses and Adaptations to Resistance Exercise and Training. Sports Medicine, 35(4), 339–361. [CrossRef]
- Fragala, M. S., Kraemer, W. J., Denegar, C. R., Maresh, C. M., Mastro, A. M., & Volek, J. S. (2011). Neuroendocrine-Immune Interactions and Responses to Exercise. Sports Medicine, 41(8), 621–639. [CrossRef]
- Bogdanis, G. C. (2012). Effects of Physical Activity and Inactivity on Muscle Fatigue. Frontiers in Physiology, 3, 142. [CrossRef]
- Kany, S., Vollrath, J. T., & Relja, B. (2019). Cytokines in Inflammatory Disease. International Journal of Molecular Sciences, 20(23), 6008. [CrossRef]
- Pluchino, N., Russo, M., Santoro, A. N., Litta, P., Cela, V., & Genazzani, A. R. (2013). Steroid hormones and BDNF. Neuroscience, 239, 271–279. [CrossRef]
- Gyorkos, A., Baker, M. H., Miutz, L. N., Lown, D. A., Jones, M. A., & Houghton-Rahrig, L. D. (2019). Carbohydrate-restricted Diet and Exercise Increase Brain-derived Neurotrophic Factor and Cognitive Function: A Randomized Crossover Trial. Cureus, 11(9), e5604. [CrossRef]
- Marston, K. J., Newton, M. J., Brown, B. M., Rainey-Smith, S. R., Bird, S., Martins, R. N., & Peiffer, J. J. (2017). Intense resistance exercise increases peripheral brain-derived neurotrophic factor. Journal of Science and Medicine in Sport, 20(10), 899–903. [CrossRef]
- Rasmussen, P., Brassard, P., Adser, H., Pedersen, M. V., Leick, L., Hart, E., Secher, N. H., Pedersen, B. K., & Pilegaard, H. (2009). Evidence for a release of brain-derived neurotrophic factor from the brain during exercise. Experimental Physiology, 94(10), 1062–1069. [CrossRef]
- Reycraft, J. T., Islam, H., Townsend, L. K., Hayward, G. C., Hazell, T. J., & Macpherson, R. E. K. (2020). Exercise Intensity and Recovery on Circulating Brain-derived Neurotrophic Factor. Medicine and Science in Sports and Exercise, 52(5), 1210–1217. [CrossRef]
- Barha, C. K., Falck, R. S., Davis, J. C., Nagamatsu, L. S., & Liu-Ambrose, T. (2017). Sex differences in aerobic exercise efficacy to improve cognition: A systematic review and meta-analysis of studies in older rodents. Frontiers in Neuroendocrinology, 46, 86–105. [CrossRef]
- El-Sayes, J., Turco, C. V., Skelly, L. E., Nicolini, C., Fahnestock, M., Gibala, M. J., & Nelson, A. J. (2019). The Effects of Biological Sex and Ovarian Hormones on Exercise-Induced Neuroplasticity. Neuroscience, 410, 29–40. [CrossRef]
- Begliuomini, S., Casarosa, E., Pluchino, N., Lenzi, E., Centofanti, M., Freschi, L., Pieri, M., Genazzani, A. D., Luisi, S., & Genazzani, A. R. (2007). Influence of endogenous and exogenous sex hormones on plasma brain-derived neurotrophic factor. Human Reproduction (Oxford, England), 22(4), 995–1002. [CrossRef]
- Fowles, J. R., O’Brien, M. W., Wojcik, W. R., d’Entremont, L., & Shields, C. A. (2017). A pilot study: Validity and reliability of the CSEP-PATH PASB-Q and a new leisure time physical activity questionnaire to assess physical activity and sedentary behaviours. Applied Physiology, Nutrition, and Metabolism = Physiologie Appliquee, Nutrition Et Metabolisme, 42(6), 677–680. [CrossRef]
- Elliott-Sale, K. J., Minahan, C. L., de Jonge, X. A. K. J., Ackerman, K. E., Sipilä, S., Constantini, N. W., Lebrun, C. M., & Hackney, A. C. (2021). Methodological Considerations for Studies in Sport and Exercise Science with Women as Participants: A Working Guide for Standards of Practice for Research on Women. Sports Medicine (Auckland, N.Z.), 51(5), 843–861. [CrossRef]
- Hagobian, T. A., & Braun, B. (2010). Physical activity and hormonal regulation of appetite: Sex differences and weight control. Exercise and Sport Sciences Reviews, 38(1), 25–30. [CrossRef]
- McCarthy, S. F., Islam, H., & Hazell, T. J. (2020). The emerging role of lactate as a mediator of exercise-induced appetite suppression. American Journal of Physiology. Endocrinology and Metabolism, 319(4), E814–E819. [CrossRef]
- D.), S. G. (Ph, Dwyer, G. B., & Medicine, A. C. of S. (2007). ACSM’s Metabolic Calculations Handbook. Lippincott Williams & Wilkins.
- Cohen, J. (1988). Statistical Power Analysis for the Behavioral Sciences (2nd ed.). Routledge. [CrossRef]
- Cumming, G. (2013). Cohen’s d needs to be readily interpretable: Comment on Shieh (2013). Behavior Research Methods, 45. [CrossRef]
- Sinatora, R., Chagas, E., Mattera, F., Mellem, L., de Oliveira dos Santos, A., Pereira, L., Aranão, A., Landgraf Guiguer, E., Cressoni Araujo, A., Haber, J., Campos, L., & Barbalho, S. (2022). Relationship of Inflammatory Markers and Metabolic Syndrome in Postmenopausal Women. Metabolites, 12, 73. [CrossRef]
- Kouvelioti, R., Kurgan, N., Falk, B., Ward, W. E., Josse, A. R., & Klentrou, P. (2019). Cytokine and Sclerostin Response to High-Intensity Interval Running versus Cycling. Medicine and Science in Sports and Exercise, 51(12), 2458–2464. [CrossRef]
- Karim, R., Stanczyk, F. Z., Hodis, H. N., Cushman, M., Lobo, R. A., Hwang, J., & Mack, W. J. (2010). Associations between markers of inflammation and physiological and pharmacological levels of circulating sex hormones in postmenopausal women. Menopause (New York, N.Y.), 17(4), 785–790.
- Arent, S. M., Senso, M., Golem, D. L., & McKeever, K. H. (2010). The effects of theaflavin-enriched black tea extract on muscle soreness, oxidative stress, inflammation, and endocrine responses to acute anaerobic interval training: A randomized, double-blind, crossover study. Journal of the International Society of Sports Nutrition, 7(1), 11. [CrossRef]
- Mezil, Y. A., Allison, D., Kish, K., Ditor, D., Ward, W. E., Tsiani, E., & Klentrou, P. (2015). Response of Bone Turnover Markers and Cytokines to High-Intensity Low-Impact Exercise. Medicine and Science in Sports and Exercise, 47(7), 1495–1502. [CrossRef]
- Febbraio, M. A., & Pedersen, B. K. (2002). Muscle-derived interleukin-6: Mechanisms for activation and possible biological roles. FASEB Journal: Official Publication of the Federation of American Societies for Experimental Biology, 16(11), 1335–1347. [CrossRef]
- Hojman, P., Brolin, C., Nørgaard-Christensen, N., Dethlefsen, C., Lauenborg, B., Olsen, C. K., Åbom, M. M., Krag, T., Gehl, J., & Pedersen, B. K. (2019). IL-6 release from muscles during exercise is stimulated by lactate-dependent protease activity. American Journal of Physiology. Endocrinology and Metabolism, 316(5), E940–E947. [CrossRef]
- Zaldivar, F., Wang-Rodriguez, J., Nemet, D., Schwindt, C., Galassetti, P., Mills, P. J., Wilson, L. D., & Cooper, D. M. (2006). Constitutive pro- and anti-inflammatory cytokine and growth factor response to exercise in leukocytes. Journal of Applied Physiology (Bethesda, Md.: 1985), 100(4), 1124–1133. [CrossRef]
- de Sousa, C. A. Z., Sierra, A. P. R., Martínez Galán, B. S., Maciel, J. F. de S., Manoel, R., Barbeiro, H. V., de Souza, H. P., & Cury-Boaventura, M. F. (2021). Time Course and Role of Exercise-Induced Cytokines in Muscle Damage and Repair After a Marathon Race. Frontiers in Physiology, 12, 752144. [CrossRef]
- Petersen, A. M. W., & Pedersen, B. K. (2005). The anti-inflammatory effect of exercise. Journal of Applied Physiology (Bethesda, Md.: 1985), 98(4), 1154–1162. [CrossRef]
- Islam, H., Neudorf, H., Mui, A. L., & Little, J. P. (2021). Interpreting “anti-inflammatory” cytokine responses to exercise: Focus on interleukin-10. The Journal of Physiology, 599(23), 5163–5177. [CrossRef]
- Kasapis, C., & Thompson, P. D. (2005). The effects of physical activity on serum C-reactive protein and inflammatory markers: A systematic review. Journal of the American College of Cardiology, 45(10), 1563–1569. [CrossRef]
- Townsend, J. R., Hoffman, J. R., Fragala, M. S., Jajtner, A. R., Gonzalez, A. M., Wells, A. J., Mangine, G. T., Fukuda, D. H., & Stout, J. R. (2015). TNF-α and TNFR1 responses to recovery therapies following acute resistance exercise. Frontiers in Physiology, 6, 48. [CrossRef]
- Bathina, S., & Das, U. N. (2015). Brain-derived neurotrophic factor and its clinical implications. Archives of Medical Science : AMS, 11(6), 1164–1178. [CrossRef]
- Knaepen, K., Goekint, M., Heyman, E. M., & Meeusen, R. (2010). Neuroplasticity - exercise-induced response of peripheral brain-derived neurotrophic factor: A systematic review of experimental studies in human subjects. Sports Medicine (Auckland, N.Z.), 40(9), 765–801. [CrossRef]
- Sleiman, S. F., Henry, J., Al-Haddad, R., El Hayek, L., Abou Haidar, E., Stringer, T., Ulja, D., Karuppagounder, S. S., Holson, E. B., Ratan, R. R., Ninan, I., & Chao, M. V. (2016). Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body β-hydroxybutyrate. eLife, 5, e15092. [CrossRef]
- Vaynman, S., Ying, Z., & Gomez-Pinilla, F. (2004). Hippocampal BDNF mediates the efficacy of exercise on synaptic plasticity and cognition. The European Journal of Neuroscience, 20(10), 2580–2590. [CrossRef]
- Matthews, V. B., Aström, M.-B., Chan, M. H. S., Bruce, C. R., Krabbe, K. S., Prelovsek, O., Akerström, T., Yfanti, C., Broholm, C., Mortensen, O. H., Penkowa, M., Hojman, P., Zankari, A., Watt, M. J., Bruunsgaard, H., Pedersen, B. K., & Febbraio, M. A. (2009). Brain-derived neurotrophic factor is produced by skeletal muscle cells in response to contraction and enhances fat oxidation via activation of AMP-activated protein kinase. Diabetologia, 52(7), 1409–1418. [CrossRef]
- Mazo, C. E., Miranda, E. R., Shadiow, J., Vesia, M., & Haus, J. M. (2022). High Intensity Acute Aerobic Exercise Elicits Alterations in Circulating and Skeletal Muscle Tissue Expression of Neuroprotective Exerkines. Brain Plasticity (Amsterdam, Netherlands), 8(1), 5–18. [CrossRef]
- Wiet, R. (2018). The effects of acute aerobic exercise on BDNF levels and cognition in postmenopausal women [ClevelandStateUniversity]. https://etd.ohiolink.edu/acprod/odb_etd/etd/r/1501/10?clear=10&p10_accession_num=csu1527873335485353.
- Berchtold, N. C., Kesslak, J. P., Pike, C. J., Adlard, P. A., & Cotman, C. W. (2001). Estrogen and exercise interact to regulate brain-derived neurotrophic factor mRNA and protein expression in the hippocampus. The European Journal of Neuroscience, 14(12), 1992–2002. [CrossRef]
- Islam, H., Townsend, L. K., & Hazell, T. J. (2017). Modified sprint interval training protocols. Part I. Physiological responses. Applied Physiology, Nutrition, and Metabolism, 42(4), 339–346. [CrossRef]
- McCarthy, S. F., Ferguson, E. J., Jarosz, C., Kenno, K. A., & Hazell, T. J. (2023). Similar Postexercise Hypotension After MICT, HIIT, and SIT Exercises in Middle-Age Adults. Medicine and Science in Sports and Exercise, 55(1), 101–109. [CrossRef]
- Vanderheyden, L. W., McKie, G. L., Howe, G. J., & Hazell, T. J. (2020). Greater lactate accumulation following an acute bout of high-intensity exercise in males suppresses acylated ghrelin and appetite postexercise. Journal of Applied Physiology (Bethesda, Md.: 1985), 128(5), 1321–1328. [CrossRef]
Figure 1.
Serum concentrations (mean ± SD) of interleukin 6 (IL-6) before and after high-intensity interval training exercise in premenopausal (n=8) and postmenopausal (n=7) females. Significant differences are denoted as *p<0.05 from pre-exercise to post-HIIT for the premenopausal group; *p<0.05 and **p<0.001 from pre-exercise to post-HIIT for the postmenopausal group in the post hoc pairwise comparisons.
Figure 1.
Serum concentrations (mean ± SD) of interleukin 6 (IL-6) before and after high-intensity interval training exercise in premenopausal (n=8) and postmenopausal (n=7) females. Significant differences are denoted as *p<0.05 from pre-exercise to post-HIIT for the premenopausal group; *p<0.05 and **p<0.001 from pre-exercise to post-HIIT for the postmenopausal group in the post hoc pairwise comparisons.
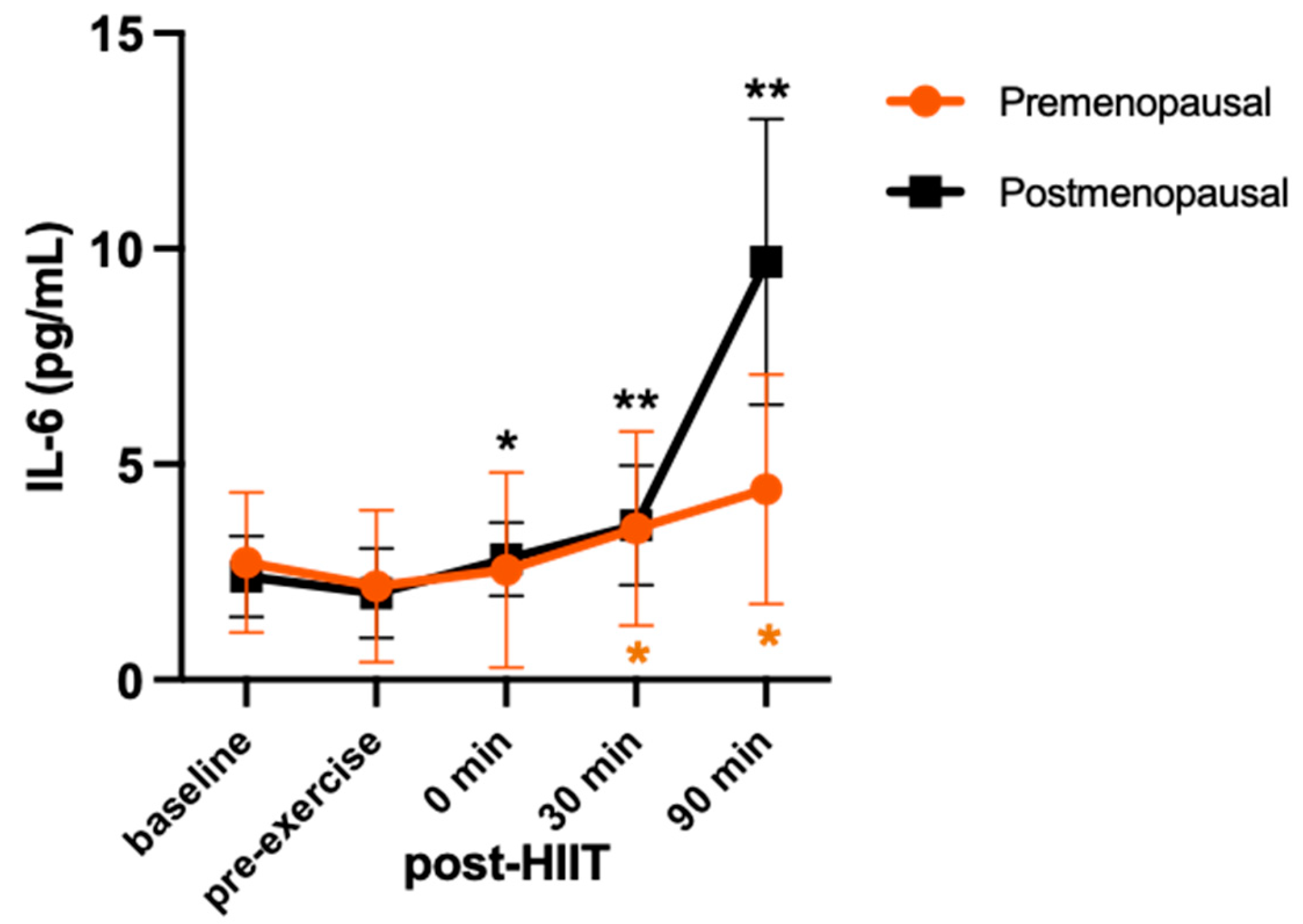
Figure 2.
Serum concentrations (mean ± SD) of interleukin 10 (IL-10) before and after high-intensity interval training exercise in premenopausal (n=8) and postmenopausal (n=7) females.
Figure 2.
Serum concentrations (mean ± SD) of interleukin 10 (IL-10) before and after high-intensity interval training exercise in premenopausal (n=8) and postmenopausal (n=7) females.
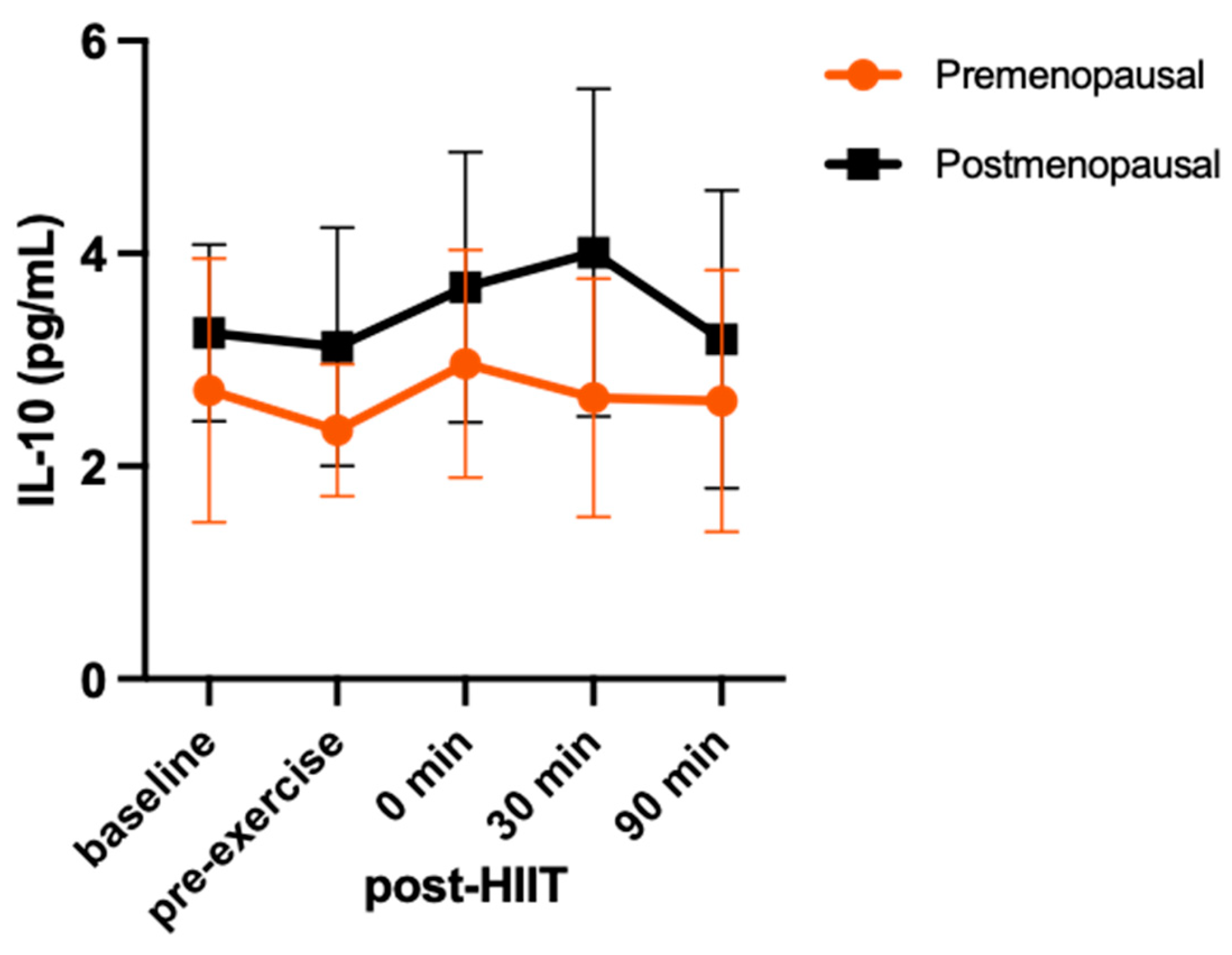
Figure 3.
Serum concentrations (mean ± SD) of tumor necrosis factor alpha (TNF-α) before and after high-intensity interval exercise in premenopausal (n=8) and postmenopausal (n=7) females. Significant differences from pre-exercise to post-HIIT are denoted as *p<0.05 for both groups combined in the post hoc pairwise comparisons.
Figure 3.
Serum concentrations (mean ± SD) of tumor necrosis factor alpha (TNF-α) before and after high-intensity interval exercise in premenopausal (n=8) and postmenopausal (n=7) females. Significant differences from pre-exercise to post-HIIT are denoted as *p<0.05 for both groups combined in the post hoc pairwise comparisons.
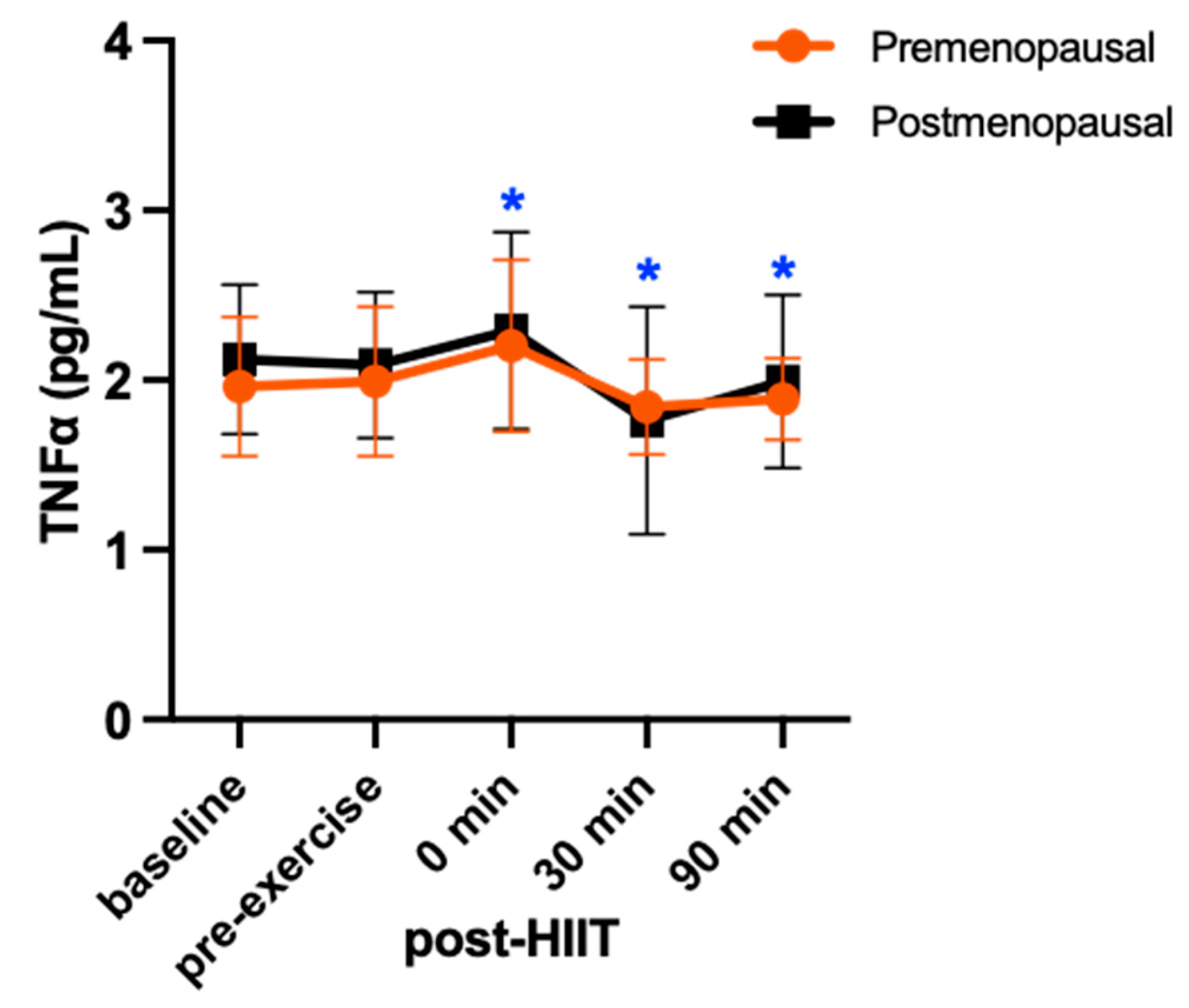
Figure 4.
Serum concentrations (mean ± SD) of brain-derived neurotrophic factor (BDNF) before and after high-intensity interval exercise in premenopausal (n=8) and postmenopausal (n=7) females. Significant differences are denoted as **p<0.001 from pre-exercise to post-HIIT for the premenopausal group in the post hoc pairwise comparisons.
Figure 4.
Serum concentrations (mean ± SD) of brain-derived neurotrophic factor (BDNF) before and after high-intensity interval exercise in premenopausal (n=8) and postmenopausal (n=7) females. Significant differences are denoted as **p<0.001 from pre-exercise to post-HIIT for the premenopausal group in the post hoc pairwise comparisons.
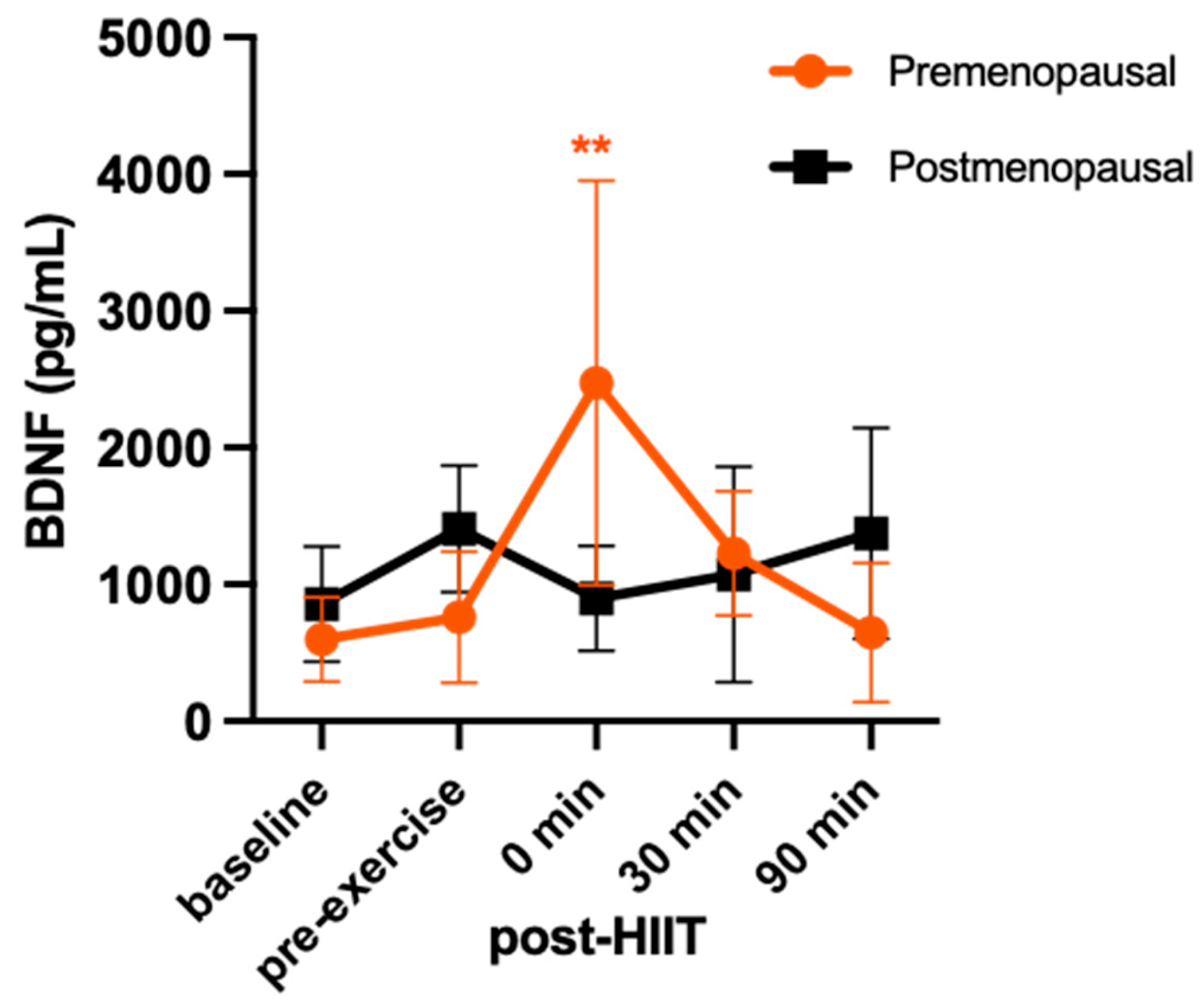

Table 1.
Blood lactate concentrations (mean ± SD) across time during the high-intensity interval training (HIIT) session in premenopausal (n = 8) and postmenopausal (n = 7) females.
Table 1.
Blood lactate concentrations (mean ± SD) across time during the high-intensity interval training (HIIT) session in premenopausal (n = 8) and postmenopausal (n = 7) females.
| Baseline (fasted) | pre-exercise (postprandial) |
0 min post-HIIT |
30 min post-HIIT |
30 min Post-HIIT |
|
|---|---|---|---|---|---|
| Blood Lactate (mmol/L) | |||||
| Premenopausal | 0.72 ± 0.52* | 1.19 ± 0.70# | 1.80 ± 1.02#* | 1.14 ± 0.60# | 0.77 ± 0.36* |
| Postmenopausal | 0.50 ± 0.18* | 0.80 ± 0.29# | 2.28 ± 0.94#* | 1.17 ± 0.35#* | 0.86 ± 0.17# |
Significant differences are denoted as #p 0.05 from baseline and *p 0.05 from pre-exercise.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



