
Submitted:
30 April 2024
Posted:
30 April 2024
You are already at the latest version
Abstract
Resveratrol (RSV), a naturally occurring phytoalexin, is the most important stilbenoid synthesized by plants, as a defense mechanism, in response to microbial aggression, toxins or ultraviolet radiation. RSV came to the attention of researchers both as a potential chemopreventive agent and a possible explanation for the low incidence of cardiovascular disease (CVD) in French people with a high-fat diet. RSV is mainly administered as a food supplement, its properties being evaluated in vitro or in vivo, on various experimental models. RSV modulates signaling pathways that limit the spread of tumor cells, protects nerve cells from damage, is useful in the prevention of diabetes, and generally acts as an anti-aging natural compound. It was highlighted that RSV could ameliorate the consequences of an unhealthy lifestyle caused by an exaggerated caloric intake. This paper reviews the evidence supporting the beneficial effect of RSV for various pathological conditions, e.g., neoplastic diseases, neurodegeneration, metabolic syndrome, diabetes, obesity, CVDs, immune diseases, bacterial, viral, and fungal infections. The study also focused on the chromatographic analysis of trans-RSV (tRSV) in Romanian wine samples, providing a comprehensive overview of tRSV content across different types of wine.
Keywords:
resveratrol
; stilbenoids
; grapes
; wine
; biological activity
; applications
1. Introduction
Stilbenes are natural secondary metabolites also known as phytoalexins. Stilbenoids, respectively the hydroxylated derivatives of stilbenes, have in their structure two benzene nuclei connected by an ethylene bridge and belong to the category of polyphenolic compounds that are biosynthesized in the same way as flavonoids. Due to their role as phytoalexins, stilbenoids are very important in the defense of plants against pathogens. Up to now, more than 1000 stilbenoids have been isolated and identified, of which the best known are the natural components resveratrol (RSV – 3,5,4’-trihydroxy-stilbene), piceid, piceatannol, pterostilbene, astringin, viniferin, pallidol, hopeaphenol. Stilbenoids, primarily derived from grapes, are compounds of significant pharmacological interest because they offer numerous health benefits, particularly in preventing and potentially treating various chronic diseases related to aging, through anti-tumoral, antioxidant, anti-inflammatory, cardioprotective, neuroprotective, antidiabetic action [1,2,3].
The two natural isomers of RSV, cis-RSV (cRSV) and trans-RSV (tRSV), are biosynthesized almost entirely in grape skins; the highest concentrations of RSV were determined before the ripening of the grapes. tRSV has the highest biological activity, cRSV being occasionally identified only in some wines. It is believed that cRSV results from an enzymatic reaction during the wine fermentation process or by exposure of tRSV to ultraviolet (UV) radiation. tRSV is relatively stable and is preferred for chemical analyzes and syntheses [1].
RSV is more lipophilic than other phenolic compounds, potentially enhancing its cell permeability and absorption. However, its bioavailability is low due to fast metabolism, resulting in discrepancies between in vitro and in vivo results [2,3].
RSV came to the attention of researchers approximately five decades ago, being both a potential chemopreventive agent and a possible explanation for the so-called “French paradox”: the low incidence of cardiovascular disease (CVD) among the French, a people with a high-fat diet [1].
RSV is mainly administered as a food supplement, its properties being evaluated in vitro or in vivo, on various experimental models. RSV has potential beneficial effects in various diseases, e.g., diabetes, obesity, CVDs (hypertension), neoplastic diseases, Alzheimer’s disease (AD). In particular, RSV has been found to affect the expression of several genes, including genes encoding cytokines, caspases, matrix metalloproteinases (MMPs), adhesion molecules, growth factors. Moreover, RSV can modulate the activity of several signaling pathways, such as phosphatidylinositol 3’-kinase (PI3K)/Akt, Wnt, nuclear factor-kappa B (NF-κB), Notch [1,4,5].
RSV is being studied as a cancer prevention agent, effective both in vivo and in vitro, because it interferes with the cellular activities linked to the stages of cancer development: initiation, promotion, and progression. It exhibits antioxidant, anti-mutagenic, and anti-inflammatory properties. Furthermore, RSV inhibits cyclooxygenase (COX)-1 and COX-2 enzymes, which are involved in converting arachidonic acid into substances that promote inflammation, potentially stimulating tumor growth and weakening immune defenses. The two COXs can activate carcinogens to reactive forms that produce cytogenetic damage. In addition to COX and hydroperoxidase inhibition, RSV induces the differentiation of human leukemic promyelocytes [1,4].
RSV induces the expression of phase II enzymes of the metabolism of active principles, the activity of quinone reductase (involved in detoxification) and inhibits ribonucleotide reductase [an enzyme complex that catalyzes the reduction of ribonucleotides to deoxyribonucleotides necessary for deoxyribonucleic acid (DNA) synthesis], cytochrome P450 CYP1A1, DNA polymerase, COX, and nicotinamide adenine dinucleotide reduced form (NADH)–ubiquinone oxidoreductase. The influence of RSV on these enzyme systems explains its antioxidant, antimutagenic and anticarcinogenic potential [1,4].
RSV has a structure similar to that of diethylstilbestrol and therefore could exhibit a phytoestrogenic effect. Phytoestrogens have numerous physiological effects in humans, the moderate consumption of red wine at meals being useful for protecting the body against diseases and improving the aging process [6]. In vegetarians, but also in the case of Asians, people with a diet rich in phytoestrogens, there is a low incidence of breast, endometrial, ovarian, and prostate cancer (PC). Phytoestrogens are natural non-steroidal plant components, structurally and functionally similar to estrogens produced by the human body. Studies have shown that these compounds alleviate menopause symptoms and have anticancer properties, conferring protection against osteoporosis and hormone-dependent cancers (breast, prostate). In addition, phytoestrogens inhibit angiogenesis and cell cycle progression [7]. Phytoestrogens bind and activate estrogen receptors (ERs) but are less effective than estrogens of endogenous origin. RSV exerts its antiestrogenic activity, competing with 17β-estradiol for binding to ERs. By antagonizing the stimulatory effect of estradiol, RSV inhibits the growth of ER-positive human breast cells, acting both at the cellular level (cell growth) and at the molecular level (gene activation) [8,9].
Research highlighted the fact that moderate intake of wine can have beneficial effects on human health, attributed especially to phenolic derivatives. In addition to flavonoids, polyphenolic acids (e.g., hydroxybenzoic acids, hydroxycinnamic acids) and stilbenes are important compounds present in grapes and wine. The remarkable properties of wine are due to polyphenolic compounds, mainly through their antioxidant activity and ability to eliminate free radicals. In addition, these compounds [(+)-catechol, (–)-epicatechol, proanthocyanidins, quercetin, kaempferol, RSV, gallic acid, caffeic acid] have been reported to exhibit a multitude of pharmacological actions, including cardioprotective, antiatherogenic, anti-inflammatory, antitumor, antibacterial, antiviral. From a phytochemical point of view, grapes represent a natural combination of polyphenols with a synergistic effect in the prevention of diseases and maintaining the health of the body. Epidemiological studies and clinical trials have shown that a regular and moderate consumption of red wine (1–2 glasses/day) is associated with a low incidence of CVDs, diabetes, and some types of cancer. The bioavailability of polyphenolic compounds varies within quite wide limits, so the most abundant polyphenols in wine are not necessarily the ones that determine the highest levels of active metabolites in the target tissues. Since wine is a complex mixture, it is probable that various chemical components, primarily polyphenols and their metabolites, work together to positively impact human health [10,11,12].
This paper reviews the evidence supporting the beneficial effect of RSV for various pathological conditions, such as: neoplastic diseases, neurodegeneration, brain injury, metabolic syndrome, diabetes, obesity, non-alcoholic fatty liver disease (NAFLD), CVDs, immune disorders, bacterial, viral, and fungal infections. In addition, the study focused on the ultra-high-performance liquid chromatography–mass spectrometry (UHPLC–MS) analysis of tRSV in Romanian wine samples, both white and red varieties, providing a comprehensive overview of tRSV content across different types of wine.
2. Natural Occurrence of Resveratrol and Related Compounds
RSV has been identified in numerous natural resources of plant origin: e.g., grapevines, Vitis vinifera L. and V. labrusca L. (Vitaceae), the most important source; the roots of Polygonum cuspidatum Sieb., Japanese knotweed (Polygonaceae); the rhizomes and roots of Holoschoenus vulgaris Link. sin. Scirpus holoschoenus L., marsh cypress (Cyperaceae); the roots of Veratrum grandiflorum (Maxim. ex Baker) Loes., white sedge and V. formosanum O. Loes. (Liliaceae) – an endemic species in Taiwan; the leaves of Eucalyptus globulus Labill. and E. accedens W. Fitzg., eucalyptus (Myrtaceae); peanuts, Arachis hypogaea L. (Fabaceae) and peanut butter; the leaves of spruce, Picea abies L. (Pinaceae); the fruits of Vaccinium myrtillus L., blueberry, V. corymbosum L., American blueberry, and V. macrocarpon Ait., American cranberry (Ericaceae); cocoa beans, Theobroma cacao L. (Sterculiaceae) and chocolate; blackberry, Rubus fruticosus L. (Rosaceae); tomatoes, Lycopersicon esculentum Mill. (Solanaceae); the aerial part of Pterolobium hexapetallum (Roth) Santapau & Wagh (Fabaceae); the fruits and leaves of Cassia garrettiana Caraib. and C. quinquangulata Rich. (Fabaceae); the wood of Bauhinia racemosa Lam. (Fabaceae); sweet almonds, Prunus amygdalus Bartock sin. Prunus dulcis (Mill.) D. A. Webb (Rosaceae); different varieties of hops used mainly in the beer industry (Amarillo, Cascade, Nugget, Simcoe, Sterling, Tomahawk, Vanguard, Warrior, Wilhamette), Humulus lupulus L. sin. Lupulus amarus Gilib. (Cannabaceae); apples, raspberries, plums, pistachios, hazelnuts; and also, in the products derived from the processing of these raw materials (such as natural juices/syrups, various hydroalcoholic extracts, wines) [2,3,5,13,14,15].
Spermatophytes, such as grapevines, generate RSV, which is the most important stilbenoid synthesized by plants, as a defense mechanism, in response to microbial attack, toxins or UV radiation, as a naturally occurring phytoalexin. V. vinifera L., mainly in the form of wine, is a valuable dietary source of stilbenoids, such as tRSV, which has the most important anti-aging biological effects in in vitro and in vivo studies. Hence, the compounds found in highly resistant strains are essential for the development of resistant crops, natural spray reagents and as novel nutritional supplements or phytopharmaceuticals [3].
RSV is found in large quantities especially in the skin, but not in the pulp of the grapes. RSV content in wines obtained from different grape varieties is extremely variable. Typically, white wines (those produced from the White Burgunder, Riesling, Ortega, and Gewürztraminer varieties) contain about 10 times less RSV than red wines (from the Cabernet Mitos, Cabernet Cubin, Syrah, Spätburgunder/Pinot noir, Cabernet Sauvignon and Merlot). In red wine, tRSV amounts generally vary between 0.1 and 15 mg/L. RSV concentration is influenced by several factors, such as: grape variety, genotype, climate zone, soil type, exposure to light, pest control, agrotechnical works, maturity, post-harvest treatment, storage, preservation [2,13].
It was firstly reported the presence of RSV, as a bioactive compound, in eight Western Romanian propolis samples, along with kaempferol, quercetin, rosmarinic acid. The principal component analysis showed clustering of the propolis samples, according to the polyphenolic profile similarity [16].
The structure of phenanthrene derivatives, tri-O-methyl-RSV, methyl-tri-O-methyl-gallate, pterostilbene, RSV, methyl-gallate were determined by analytical and spectroscopic methods from the stems of P. hexapetallum [15]. Bioassay-directed purification of the active compounds from an ethanol extract of Psoralea corylifolia L., babchi (Fabaceae) led to the isolation of corylifolin and bakuchiol as DNA polymerase inhibitors and also, based on their structures, RSV was tested and demonstrated to be effective as a DNA polymerase inhibitor [17].
Viniferin is a derivative of RSV, which has several isomers: e.g., α-viniferin, β-viniferin, γ-viniferin, δ-viniferin, ε-viniferin, R-viniferin (vitisin A), R2-viniferin (vitisin B). δ-Viniferin is a dehydrodimer of RSV, respectively an isomer of ε-viniferin. It occurs in vitro by oxidative dimerization of RSV by plant peroxidases or fungal laccases. Also, δ-viniferin was recently identified in wines, in cell cultures and in grapevine leaves infected with Plasmopara viticola Berk. & M.A. Curtis (Peronosporaceae) or irradiated with UVC. δ-Viniferin, identified in higher concentration than ε-viniferin, is one of the most important phytoalexins derived from RSV. Viniferins have a series of important biological activities, such as: anti-inflammatory, anti-psoriatic, anti-diabetic, anti-parasitic, anti-cancer, anti-angiogenic, antioxidant, anti-melanogenic, neuroprotective, antiviral, antibacterial, antifungal, anti-diarrheal, anti-obesity [18,19].
Pallidol is a natural dimer of RSV, also identified in red wine, with antioxidant and antifungal properties. At low concentrations, pallidol has strong singlet oxygen inactivation effects, being ineffective in eliminating hydroxyl radicals or superoxide anions. Kinetic studies showed that the reaction of pallidol with singlet oxygen had a very high constant. Therefore, pallidol could be used as a pharmacological agent in singlet oxygen-mediated diseases, thus contributing to the beneficial health effects of red wine [20].
Hopeaphenol, identified for the first time in red wine, especially from the Merlot variety, has a stilbenoid tetramer structure. It was determined by reversed-phase high-performance liquid chromatography (RP–HPLC) analysis of wine samples from North Africa (Algeria), alongside the known stilbenoid components: tRSV, trans-piceid, trans-ε-viniferin, pallidol [21].
3. Anti-Tumoral Properties
Chemoprevention by non-toxic agents, such as RSV, has been demonstrated to provide protection against various cancer types. Nevertheless, the molecular mechanisms responsible for anti-tumorigenic or chemopreventive actions of this phytochemical continue to be mostly unidentified [22].
3.1. Leukemia and Lymphoma
RSV has been determined to inhibit carcinogenesis in murine models. Besides its chemopreventive action, it also induces apoptotic cell death in human leukemia cells, exhibiting specific involvement of the cluster of differentiation (CD)95-CD95L system in the anti-tumoral activity of RSV and emphasizing the chemotherapeutic ability of this natural compound [23]. It is a significant inhibitor of ribonucleotide reductase and DNA synthesis in mammalian cells, with possible utilizations as an antiproliferative and chemopreventive factor [24].
Studies demonstrate that RSV can inhibit every step of multistage carcinogenesis, by reducing viability and DNA synthesis ability of promyelocytic leukemia cells, inducing the apoptotic cell death, and decreasing the expression of anti-apoptotic B-cell lymphoma-2 (Bcl-2) [25]. In the concentration spectrum of 20 μM and higher, blocked the S-phase and caused apoptosis in the T-cell acute lymphoblastic leukemia (T-ALL), insufficient in functional p53 and p16 [26]. Other studies had the same results in human histiocytic lymphoma cells, indicating that RSV is an important cell cycle blocker, in addition to being a chemopreventive agent [27].
In the case of acute monocytic leukemia, researchers have shown that RSV inhibits the growth of THP-1 cells by inducing apoptosis, independent of the Fas/FasL signaling pathway and has no toxic effect on differentiated THP-1 cells [28]. Other data established RSV and its correspondent, piceatannol, are powerful generators of apoptosis in BJAB Burkitt-like lymphoma cells, with a median effective dose of 25 μM, autonomous of the CD95/Fas signaling cascade [29].
tRSV, ε-viniferin and vineatrol induced apoptosis in chronic leukemic B cells, by activating caspase-3, reducing the mitochondrial transmembrane potential, and inhibiting both the expressions of the Bcl-2 anti-apoptotic protein and the inducible nitric oxide (NO) synthase [30]. Some studies suggest the involvement of apoptosis signal-regulating kinase 1/c-Jun N-terminal kinase signaling in the regulation of FasL expression and following apoptosis induced by RSV in leukemia cells [31].
The inhibitory effect on hydrogen peroxide (H2O2)-induced apoptosis is attributed to the antioxidant property of RSV, which, in small concentrations, can inhibit death signaling in human leukemia cells via nicotinamide adenine dinucleotide phosphate reduced form (NADPH) oxidase-dependent increase of intracellular superoxide that stops mitochondrial H2O2 synthesis [32].
Researchers determined that RSV has the ability to sensitize cancer cells to X-irradiation. The study involved cervix carcinoma, chronic myeloid leukemia and multiple myeloma and demonstrated that the addition of RSV alone induced apoptosis and inhibited cell growth, while concomitant treatment with either RSV or X-irradiation caused a synergical reaction [33].
It has been discovered a plasma membrane receptor for RSV near the arginine–glycine–aspartate recognition site on integrin α(v)β3 that participates in the proapoptotic mechanism of RSV in tumor cells, along with stimulating the intranuclear COX-2 accumulation, correlated to the activation of p53 [34].
The possible anti-tumor actions of RSV against the natural killer (NK) cell malignancies, especially aggressive NK cell leukemias and lymphomas include strong G0/G1 cell cycle blockage, substantially restriction of cell proliferation and apoptosis. Furthermore, RSV caused synergistic activity on the apoptotic and antiproliferative effects of L-asparaginase against cancer cells [35].
RSV’s effectiveness varies among different human cancer cells, showing greater potential to induce cell death in leukemic cells than in solid tumor cells, by inhibiting Akt activation through Ras downregulation [36].
3.2. Glioma, Glioblastoma and Neuroblastoma
Neuroblastoma is an aggressive type of tumor, which requires a more efficient and less cytotoxic treatment. Studies indicate that the activity of RSV in neuroblastoma cells can be mediated by functionally activated p53 [37]. RSV intervenes in the proliferation, apoptosis, and cell cycle modification of neuroblastoma cells, inducing important cytotoxicity and increased apoptosis. S-phase accumulation is connected to the downregulation of p21 and up-regulation of cyclin E [38].
During an investigation in rat gliomas, RSV exerted important glioma cell cytotoxicity and apoptosis, induced antitumor effects on the spinal cord and intracerebral gliomas and inhibited angiogenesis in spinal cord gliomas [39]. RSV elevates cellular cytotoxicity and inhibits the proliferation of rat neuroblastoma cells, by activating mitochondria-mediated intrinsic caspase dependent pathway [40].
Human glioma cells were utilized to comprehend the molecular mechanisms of the anticancer effect of RSV. The phytocompound caused apoptosis of glioma cells, as evaluated by lactate dehydrogenase release and internucleosomal DNA fragmentation analysis. RSV activated caspase-3, which lead to cytochrome c release from mitochondria to the cytoplasm and activation of caspase-9 [41].
RSV has the ability to suppress the growth and proliferation of glioma and improve its apoptosis through upregulating the leucine repeat immunoglobulin-like protein 1 (LRIG1) gene expression, which stimulates antiglioma growth, suggesting that LRIG1 is a new biological objective in antiglioma cell proliferation and growth [42].
Researchers investigated autophagy induced by RSV in human glioma cells, which inhibited growth and induced cell apoptosis. Glioma cells firmly expressing green fluorescent protein (GFP) merged to LC3, engaged more GFP–LC3-labeled autophagosomes, and elevated the percentage of cells with GFP–LC3-labeled autophagosome. Also, RSV-induced autophagy was regulated by P38 and the extracellular signal-regulated kinase 1/2 (ERK1/2) pathway [43].
Glioblastoma multiforme (GBM) is the most destructive intracranial tumor that overexpresses the YKL-40 gene, related to the tumor stages of human primary astrocytoma and glioma cell proliferation. It was confirmed that RSV inhibits YKL-40 expression, by limiting the exertion of its supporter and decreasing messenger ribonucleic acid (mRNA) transcription and protein activity in vitro, involving the ERK1/2 pathway [44].
Invasion and metastasis of glioblastoma-initiating cells are accountable for the evolution and recurrence of GBM. In vitro and in vivo observations suggest that RSV inhibits the adhesion, invasion, and migration of glioblastoma-initiating cells. Furthermore, it suppresses the invasion of glioblastoma-initiating cells by the inhibition of PI3K/Akt/NF-κB signal transduction and the following suppression of MMP-2 expression [45].
Recent findings exhibited the effect of RSV for the amelioration of glioblastoma inflammatory response by reducing NOD-like receptor protein 3 (NLRP3) inflammasome activation through inhibition of the Janus tyrosine kinase 2/signal transducer and activator of the transcription 3 (JAK2/STAT3) pathway [46].
3.3. Meningioma
RSV may be effective against meningioma cells by curbing their growth and triggering apoptosis. It increases cleaved caspase-3 protein levels while reducing pro-caspase-3 protein and Bcl-2 mRNA levels. Additionally, the RSV mechanism includes the upregulation of miR-34a-3p, which targets the 3’-untranslated region (3’-UTR) of Bcl-2, decreasing its protein levels and thereby promoting apoptosis in these cells. Therefore, the RSV anti-proliferative and pro-apoptotic effects in meningioma cells are mediated through the regulation of miR-34a-3p and subsequent suppression of Bcl-2 [47].
3.4. Estrogen-Dependent Pituitary Tumor
Phytoestrogens have been widely studied, due to their capacity to reduce the incidence of various estrogen-dependent tumors. Some of them were proven to be active in pituitary tumor cells, by binding to the ERs, stimulating growth and causing an estrogenic response, such as prolactin secretion. Nevertheless, RSV does not attach to the ERs. Zearalenone, genistein and coumestrol have been demonstrated to induce prolactin secretion and to encourage growth, while RSV exhibited prolactin secretion, but without growth stimulation [48].
3.5. Breast Cancer
Breast cancer (BC) is one of the most prevalent tumors in females and is culpable for the principal cancer-related deaths after lung cancer. The anticancer mechanisms of RSV concerning BC involve the restriction of cell proliferation, decreasing of cell viability, invasion, and metastasis. However, its favorable therapeutic effects, it has limitations due to the reduced pharmacokinetics, stability, and poor water solubility, implying the help of nanodelivery systems to enhance its bioavailability [49,50]. RSV-modified mesoporous silica nanoparticles (NPs) inhibit BC progression more efficiently, in comparison to the phytochemical alone, via suppressing the NF-κB signaling pathway, indicating that they are a novel and more secure method for applying in the complex therapeutics of BC [51].
Some research involved RSV-loaded solid lipid NPs that were properly designed to improve BC cells. The NPs presented a higher capacity than free RSV in suppressing the proliferation of tumor cells and displayed much better inhibitory actions on the incursion and migration of BC cells [52,53]. Additional data, important in the BC applications, established that transferrin–cathepsin B cleavable peptide modified mesoporous silica NPs encapsulated RSV substantially reduced cell viability and enhanced cell apoptosis [54].
Research has shown that RSV activates the transcription of cytochrome P450 CYP1A1 and reduces the formation of carcinogen-induced preneoplastic lesions in mouse mammary tissues. It also prevents the growth of mouse skin tumors in two-stage models, inhibits the enzymatic actions of COX-1 and COX-2 in cell-free experiments, and blocks COX-2 mRNA along with the activation of protein kinase C (PKC) and AP-1-mediated gene expressions in mammary epithelial cells [55].
There has been investigated a possible mechanism by which RSV could control the cell cycle and apoptosis, exploring the human BC MCF-7 cell line. The results showed a dose-dependent anti-proliferative effect of RSV, that was correlated with a notable inhibition of cyclin D and cyclin-dependent kinase (CDK)4 proteins and stimulation of p53 and CDK inhibitor p21WAF1/CIP [56,57]. Apoptosis caused by RSV was determined to happen exclusively in BC cells exhibiting wild-type p53, however not in mutant p53-expressing cells [58].
The growth factor heregulin-beta1 (HRG-β1) promotes the tumorigenicity and metastasis of human BC cells. MMP-9 is a crucial enzyme in the degeneration of extracellular matrices and has an altered expression in BC infiltration and metastasis. RSV showed anticarcinogenic activities in both in vitro and in vivo explorations that imply it suppresses the MMP-9 expression in BC cells, connected with the downregulation of the mitogen-activated protein kinase (MAPK)/ERK signaling pathway [59].
Metastasis is an important cause of decease in patients with BC. In the evolution of the tumor, epithelial–mesenchymal transition (EMT) is essential to supporting the invasion and migration of cancer cells. RSV suppresses the migration of human BC cells by invalidating the transforming growth factor-beta1 (TGF-β1)-induced EMT and inhibits the lung metastasis in a xenograft-bearing mouse model [60]. The cytotoxic action of RSV on human BC cells may be correlated with its pro-oxidant effect, which inhibits casein kinase 2 activity, influencing cell viability and mitochondrial function [61].
3.6. Hepatocellular Carcinoma
Many studies are designed to examine the chemopreventive action of various doses of RSV against hepatocellular carcinoma (HCC). It was demonstrated that a 100 mg/kg dose of RSV can be a suitable treatment for HCC produced by diethylnitrosamine in rats [62]. Other evidence revealed that RSV suppresses tumor necrosis factor-alpha (TNF-α)-regulated MMP-9 expression and progression of human HCC cells, which correlates with the inhibition of the NF-κB signaling axis [63]. RSV reduces cell proliferation, produces reactive oxygen species (ROS), and generates DNA single-strand breaks. Due to its ability to regulate the expression of proteins implicated in the redox pathways and apoptosis, RSV induces hepatic cancer cell death by inhibiting the expression of antioxidant proteins [64].
It was demonstrated that several natural polyphenols stimulate the activity of telomerase. Recent studies show that silent information regulator 1 (SIRT1) and nuclear factor erythroid 2 (NF-E2)-related factor 2 (Nrf2) are implicated in the modulation of human telomerase reverse transcriptase (hTERT). RSV, together with kuromanin chloride and gallic acid, stimulate the hTERT gene expression in the hepatocellular tumor cells, probably by activation of the SIRT1/Nrf2 signaling cascade. Accordingly, through addressing hTERT or this axis, the contra-tumor action of this class of phytochemicals might be increased [65].
When treated with a mixture of bioactive substances, RSV and berberine, human HCC cells, Hep-G2 and Hep-3B, with various p53 status, showed distinct reactions in cell viability by p53-reliant apoptosis pathway activation. An efficient concentration of berberine, combined with RSV, could be decreased even to 50% in tumor therapy and substantially decreases the viability of wild type p53 Hep-G2 and null p53-mutant Hep-3B. Therefore, p53 status in hepatocellular tumor cell lines regulates the reactions to phytotherapy [66].
3.7. Pancreatic Carcinoma
Pancreatic tumor, one of the most frequent causes of cancer-connected mortality, has a weak outcome, brief survival rate and resistance to therapy. Studies have shown that RSV has chemopreventive and treatment action in pancreatic cancer: it reduces the propagation of pancreatic cancer cells; generates apoptosis and cell cycle arrest; prevents metastasis and infiltration of pancreatic cancer cells; obstructs the reproduction and viability of tumor stem cells; increases the chemoradiosensitization of cancer cells [67].
The examination of the mechanisms implicated in the in vitro antitumor activity of RSV revealed that it suppresses the proliferation of pancreatic cancer cells, which, after incubation with RSV, lead to cell apoptosis and cell cycle cessation. RSV generates the activation of caspases, modulates the activity of the contra-apoptotic proteins Bcl-xL, Bcl-2 and X-linked inhibitor of apoptosis protein (XIAP) and the proapoptotic protein Bax [68].
RSV performs dual functions in pancreatic tumors: as a cancer inhibitor by the up-regulation of Bax and as a carcinoma activator by the up-regulation of vascular endothelial growth factor-B (VEGF-B). However, the antitumor action of RSV is more powerful than the cancer stimulating one. As a result, the association of RSV with pharmacological inhibitor of VEGF-B can be a valuable clinical approach in the treatment of pancreatic cancer [69]. Moreover, polydopamine-based RSV–hyaluronidase nanomedicine inhibited pancreatic cancer cell invasive phenotype in hyaluronic acid enrichment tumor sphere model [70].
Hyperlipidemia acute pancreatitis is an important undercover threat concerning our health. Hence, researchers analyzed the preventive influence and the intrinsic mechanism of RSV in the hyperlipidemia acute pancreatitis in mice and discovered that the RSV pretreatment efficiently suppresses pancreatic pathological injury, inhibits inflammation and oxidative stress, by restraining the NF-κB signaling pathway and improving intestinal microbiota [71].
3.8. Esophageal Carcinoma
The purpose of several studies was to investigate the probability and possible mechanism of the inhibition of N-nitrosomethylbenzylamine-induced rat esophageal tumorigenesis by RSV. It substantially reduces the up-regulated expression of COX-1 and COX-2 and the enhanced levels of prostaglandin (PG)E2 synthesis and for that reason it can be a promising natural anti-carcinogenesis factor for the prevention and therapy of human esophageal cancer [72].
In vitro experiments were conducted to evaluate the RSV-induced apoptosis in esophageal cancer cells and its connection to the expressions of Bcl-2 and Bax. They concluded that RSV is capable of generating apoptosis in esophageal tumors by influencing the expressions of these two apoptosis-modulated genes: down-regulating Bcl-2 and up-regulating Bax [73].
3.9. Gastric Cancer
Gastric cancer (GC) is one of the most frequent tumors with serious malignancy. Numerous studies have determined the implication of Wnt, Nrf2, MAPK, and PI3K/Akt signaling pathways in this type of cancer. RSV involves several signaling pathways to generate apoptotic cell death along with obstructing the migration and metastasis of GC cells [74].
Some observations showed that RSV is capable of decreasing viability and generate apoptosis in GC cells by suppressing NF-κB activation [75]. Other studies established that RSV elevates superoxide dismutase (SOD) activity but reduces NF-κB transcriptional activity and heparanase enzymatic activity, which corresponds with the diminishing of invasion potential in GC cells [76].
It has also been reported that RSV inhibits the proliferation, migration, and invasion of human gastric tumor cells through the metastasis-associated lung adenocarcinoma transcript 1 (MALAT1)/micro-ribonucleic acid (miRNA)-383-5p/DNA damage-inducible transcript 4 pathway and generates apoptosis [77]. Overexpression of miRNA-155-5p, which generates oncogenesis in gastric tumors, is regulated by RSV, being one of the mechanisms that confirms the important antiproliferative activity of this phytochemical [78].
Chemotherapy toxicity is not principally caused by the medication itself, but is generated via cell-free chromatin particles, that are discharged from chemotherapy-induced decaying cells and are immediately internalized by healthy cells, at which point they produce double stranded DNA (dsDNA) breaks and stimulate inflammatory cytokines. It has been discovered that they can be inactivated by oxygen radicals that are induced after treatment with RSV and copper and consequently curtail chemotherapy toxicity [79].
3.10. Colorectal Carcinoma
Chemotherapy is universally used for treating colorectal tumors. Because of the high toxicity to normal cells and the emerging cancer resistance, it is preferable to use chemotherapy combined with natural bioactive agents that can assuage the side effects, as coadjuvants. It was confirmed that the co-administration of RSV along with chemotherapy has higher anti-proliferative effect, elevates chemotherapy-induced cytotoxicity, and synergistically has the ability to sensitize tumor cells to chemotherapy as a result of its oxidative, proapoptotic and anti-inflammatory action [80].
An RSV tetramer, vaticanol C, identified in the stem bark of Vatica rassak (Korth.) Blume, resak (Dipterocarpaceae), substantially inhibits cell proliferation via initiation of apoptosis, which was represented by nuclear alterations and DNA ladder development, in several distinct human colon cancer cell lines [81]. Another phytocompound, tRSV, has antiproliferative effect without cytotoxicity and generates apoptosis in human colorectal carcinoma, through a ROS-dependent apoptosis pathway [82].
Certain studies submitted that the anti-tumor activity of RSV is viable by activating Hippo/YAP signaling and moderately perturbing the YAP–TEAD interaction, a very useful information in colon cancer therapy [83]. Reports documented that 0.5 g or 1.0 g of RSV per os every day generate efficient concentrations in the human digestive tube to induce antiproliferative activity, showing great potential as a colorectal cancer chemopreventive agent [84]. The anti-proliferative effect of a standardized combination of compounds: tRSV, quercetin, vitamin E and selenium (Neoplasmoxan) was reported in mouse colorectal carcinoma, suggesting that this preparation may be a synergic treatment in addition to standard chemotherapy or radiotherapy [85].
When the comparative analysis of the anti-proliferative, proapoptotic and oxidative stress potential of RSV–zein NPs versus free RSV was performed against human colorectal carcinoma cells, the following observations were made: the NPs achieved cell cycle suppression promoted by enhanced cytotoxicity, response, and oxidative stress markers concentrations than RSV alone. This data suggests that the chemopreventive profile of RSV is expanded, as a result of the efficacious delivery system using histocompatible nano-dispersion [86].
3.11. Lung Cancer
The anti-tumor effect of RSV has been connected to transformations in the sphingolipid metabolism. The analysis performed on the enzymes accountable for the aggregation of two most relevant sphingolipids, apoptosis-promoting ceramide, and proliferation-stimulating sphingosine-1-phosphate (S1P), in human lung adenocarcinoma cells, revealed that RSV administration enhanced apoptosis-promoting ceramide and sphingosine and reduced sphingomyelin and S1P [87].
Studies show that RSV generated apoptosis in lung cancer cells by precisely targeting phosphoAkt (pAkt) and cellular FLICE-like inhibitory protein (c-FLIP) downregulation, as a result of proteasomal degradation contingent to epidermal growth factor receptor (EGFR) [88]. Other data determined that inhibits lung tumor cells proliferation by suppressing COX-2 expression [89]. New mechanisms of RSV tumor suppression have been discovered, implying that this phytochemical has the ability to reprogram M2-like tumor-associated macrophages and tumor-infiltrating CD8T effector cells [90].
Further investigations reported the implication of autophagy in RSV-induced apoptosis and its possible molecular mechanisms via Ca2+/adenosine-monophosphate activated-protein kinase (AMPK)–mammalian target of rapamycin (mTOR) signaling pathway in human lung adenocarcinoma cells [91]. In a non-small-cell lung cancer assay, RSV-induced autophagy, and apoptosis at levels higher than 55 μM were mediated by the nerve growth factor receptor–AMPK–mTOR signaling pathway [92].
There are numerous indications that cancer stem cells are a crucial conductor of carcinogenesis and tumor reoccurrence, involving cytokines and chemokines – the cellular and soluble constituents of the tumor microenvironment. The expression level of interleukin (IL)-6 is interconnected to the existence of lung cancer stem-like cells in human lung carcinoma. RSV has the ability to multi-target lung cancer stem-like cells and IL-6 in the tumor microenvironment, being an innovative approach in the therapy and prevention of lung cancer [93].
Regarding the lung cancer metastasis of melanoma, the cytotoxicity of RSV was assessed in immunocompetent mice. The in vitro development of melanoma cells was substantially inhibited by RSV. The average survival rate of mice was improved, and the lung tumor proliferation was suppressed through in vivo intraperitoneal injection of 40 mg/kg RSV. The lung tumors exhibited enhanced interferon-gamma (IFN-γ) and C-X-C motif chemokine ligand 10 (CXCL10) levels and reduced angiogenesis and infiltration [94].
3.12. Bladder Cancer
Relevant information about the influence of RSV on human bladder carcinoma cells disclose that its effects are dose-dependent and may encourage the creation of impending chemoprevention approaches. Elevated doses of RSV produce apoptosis of bladder tumor cells, while reduced dosage provides protection, by regulating Bcl-2 protein, during oxidative stress condition [95].
When the effects of RSV were evaluated in two human renal cell carcinoma cell lines, the findings indicated that it inhibits cell proliferation, migration, and infiltration, due to deactivation of the Akt and ERK1/2 signaling cascades, depending on concentration and time [96].
Recent studies demonstrated the antiproliferative action of RSV in bladder tumor cells, mediated by the ROS production, intervention in the cell cycle and suppression of cell migration, regardless of their tumor protein p53 (TP53) status. The enhanced ROS formation after RSV administration was concluded by decreased cell viability and proliferation in all bladder cancer cell lines [97].
3.13. Cervical Carcinoma
Cervical cancer is one of the most prevalent tumors in women, especially in developing countries. In order to find more affordable approaches in therapy, many studies included plant polyphenols, such as RSV or pterostilbene, which have presented antitumor and anti-human papillomavirus (HPV) effect opposed to cervical cancer cells. They reduce clonogenic survival, diminish cell migration, block cells at the S-phase and decrease the number of mitotic cells. The data confirmed a mechanism that concerns the downregulation of the HPV E6 oncoprotein, re-establishment of functional p53 protein and stimulation of apoptotic pathways, with pterostilbene being more effective compared to RSV [98].
RSV suppresses proliferation and generates apoptosis in human cervical cancer cells in a dose and time-relative process. The RSV-inflicted cell shrinkage and apoptosis is followed by the stimulation of caspase-3 and caspase-9, upregulation of the proapoptotic Bcl-2-associated X protein and downregulation of the contra-apoptotic genes Bcl-2 and Bcl-extra-large. Moreover, p53, an indispensable protein for cell survival and cell cycle continuity, presents RSV-induced increased expression levels in cervical carcinoma cells [99].
Currently, it was concluded that RSV causes the apoptosis and inhibition of the migration and invasion of cervical tumor cells by inhibiting the Hedgehog (Hh) signal pathway. This natural compound downregulates the expression levels of Hh signal pathway proteins (smoothened, zinc finger transcription factors and sonic Hh homolog). The Hh agonist purmorphamine inverts the effects of RSV [100].
3.14. Ovarian Cancer
Epithelial ovarian carcinoma constitutes the deadliest gynecological tumor and a considerable health problem, because of the serious mortality percentage. The potential antitumor action of RSV was appraised concurrently in in vitro and in vivo ovarian cancer. Even though it was efficient in inhibiting the in vitro cellular invasion of ovarian cancer cells, it was inactive in in vivo ovarian cancers in rats [101].
However, when the anti-cancer mechanism of RSV was evaluated in human ovarian cancer cells, it was validated that RSV generates ROS, which provokes autophagy and following apoptosis. RSV stimulates autophagy-related gene 5 (ATG5) expression and triggers LC3 cleavage. The RSV-induced apoptotic cell death was depreciated by pharmacological and also genetic suppression of autophagy, by silencing RNA (siRNA)-targeting of ATG5. Therefore, it was confirmed that autophagy and RSV-induced apoptosis are correlated in human ovarian cancer [102].
3.15. Osteosarcoma
Osteosarcoma is a high-degree bone neoplasm with serious invasive capacity. It was reported that RSV suppresses proliferation and glycolysis, generates apoptosis, and decreases the invasiveness of osteosarcoma cells in vitro. In addition to this, RSV downregulates the expression of interconnected Wnt/β-catenin signaling pathway target genes and enhances the level of E-cadherin. In an extended-time administration of RSV, the expression of connexin 43 (Cx43) was enhanced. Therefore, this data confirms that the anticancer influence of RSV on osteosarcoma cells is achieved via up-regulating Cx43 and E-cadherin and inhibiting the Wnt/β-catenin signaling pathway [103,104].
Various studies investigated the role of RSV on proliferation and apoptosis in osteosarcoma cell lines and associated it with the activation of the SIRT1/silent information regulator 2 (SIR2) family of nicotinamide dinucleotide oxidized form (NAD+)-dependent deacetylases, which influences the calorie restriction-regulated tumor inhibition. SIRT1 protein has an increased expression in cancer cells than in normal osteoblasts. RSV generates apoptosis in osteosarcoma cells conditional to the dosage but has insignificant impact on normal osteoblasts. Moreover, the proapoptotic action of RSV may be stimulated by dietary limitation induced by L-asparaginase [105].
Recent examinations certified that RSV prevents cell proliferation, migration, and invasion, initiating apoptotic cell termination in osteosarcoma cells. The mechanism might be the downregulation of NF-κB and Akt intracellular signaling transduction. Additionally, the association with pyrrolidine dithiocarbamate, an NF-κB inhibitor, determined combined inhibition of osteosarcoma evolution [106].
3.16. Epidermoid Carcinoma and Melanoma
Nonmelanoma skin cancer is the most frequent malignancy, caused by solar UV radiation, principally by its UVB segment (290–320 nm). RSV is a powerful antioxidant, known as “photochemopreventive agent” that can improve the damages provoked by UVB exposure to epidermis. In a study on hairless mice, the topical application of RSV suppressed the UVB-induced effects, such as: increase in bifold skin thickness, skin edema, induction of COX and ornithine decarboxylase (ODC) enzyme activities and protein expression of ODC, which are important factors in tumor development [107,108].
Regarding epidermoid carcinoma, the effect of RSV on transformed keratinocytes exhibiting mutant p53 was analyzed. It induced G1 cell cycle arrest, by significantly suppressing G1 cell cycle regulatory proteins, including cyclins A and D1, CDK6 and p53-independent induction of p21WAF1. RSV also down-regulated activating protein 1 DNA-binding and promoter activity, repressed MAPK/ERK1 or ERK1/2 signaling and inhibited c-Jun [109].
With reference to melanoma treatment, the aim of various surveys was to investigate the aptitude of RSV to complement chemotherapy. In vitro RSV substantially reduces melanoma cell viability, targeting only the cancer cells and exempting fibroblast cell lines. In comparison to the administration of temozolomide alone, the treatment of malignant cells with 50 μM RSV and temozolomide for 72 hours strongly augments cytotoxicity [110]. Western blot analysis confirmed that RSV generates the apoptosis of human melanoma cells by increasing the expression of Bcl-2-associated X protein and Bcl-2, presumably through the p53 pathway and stimulation of caspase-9 and caspase-3 [111].
In a study intended to evaluate the chemopreventive and antimutagenic action of RSV against skin neoplasm in vivo, croton oil-induced increase of ODC effects of dorsal epidermis cells in mouse and mouse ear edema model were used to highlight the anti-promotion activity of RSV. Also, 7,12-dimenthylbenz[a]anthracene (DMBA)/croton oil was utilized to cause mouse skin cancer. Administration of RSV reduced ODC activity in croton oil-induced dorsal epidermis. It also was demonstrated that RSV could inhibit mouse skin papilloma produced by DMBA/croton oil, which involves expansion of cancer occurrence duration, reducing the prevalence of papilloma and decreasing tumor number per mouse [112].
3.17. Oral Squamous Cell Carcinoma
Oral squamous cell carcinoma is the most typical tumor of the head and neck. Even though surgery, radiation, and chemotherapy are adequate therapeutic protocols, the medication tolerance and toxicity of oral cancer patients continue to be an issue. Various approaches on understanding the intrinsic mechanisms of RSV-induced apoptosis were conducted on this type of neoplasm. In the case of human tongue squamous cell carcinoma, the inhibitory effect of RSV was possible along the mitochondrial pathway and the suppression of cell invasion and migration by down-regulating the EMT-inducing transcription factors [113].
In vitro studies suggest that RSV influences the adhesion, migration, and infiltration of oral squamous cell carcinoma cells, therefore it has the ability to be a chemopreventive element for decreasing the invasion and metastasis [114]. Other data corroborated that RSV alone or in association with quercetin, in concentrations similar to red wines, are efficient inhibitors of oral squamous carcinoma cell evolution and proliferation [115].
In combination with copper, RSV can reduce tumor genome instability and inflammation, by inactivating the cell-free chromatin particles, with the help of oxygen radicals. Regarding the advanced squamous cell carcinoma of oral cavity, some results indicate that cell-free chromatin particles discharged into the tumor microenvironment from moribund malignant cells are universal generators for cancer manifestations and immune frontiers in remaining tumor cells. The capacity of this mixture to inactivate the chromatin particles, presents the possibility of an innovative and non-toxic therapeutic plan, which, instead of exterminating the cancer cells, provides treatment via inhibiting immune checkpoints and tumor hallmarks [116].
3.18. Nasopharyngeal Cancer
The underlying mechanism of RSV has been investigated in various tumors. Regarding human nasopharyngeal carcinoma cells, it inhibits cell proliferation by suppressing Ki67 and proliferating cell nuclear antigen (PCNA) proteins and induces apoptosis by altering its analogous proteins, cleaved caspase-3, and cleaved caspase-9. RSV therapy diminishes the enhanced expression of survivin that inactivates its reaction on cell growth and apoptosis. These results confirm that RSV can influence the cell proliferation and demise in nasopharyngeal carcinoma cells [117].
Administration of RSV suppresses cell viability and generates apoptosis of the human nasopharyngeal carcinoma cells. Moreover, it was determined that RSV stimulates caspase-3 and modifies the Bax/Bcl-2 apoptotic signaling, due to the up-regulation of AMPK activity and inhibition of p70S6K and S6 kinases, succeeding components of AMPK [118].
3.19. Prostate Cancer
PC is one of the most frequent kinds of tumors in men, related to the unrestrained growth of the prostate gland. The evolution of androgen-independent PC implicates fundamental ERK1/2 activation, promoted by the epidermal growth factor/transforming growth factor-alpha/EGF receptor pathway. It was ascertained that RSV inhibits the ERK1/2 activation, by altering PKC, selectively its isozyme PKCα, in neoplastic human prostate epithelial cells in vivo [119].
The antitumor effect of RSV in PC can be correlated to regulation of miRNAs, which are minuscule non-coding RNAs that antagonize gene expression. RSV down-regulates carcinogenic miRNAs and up-regulates cancer-inhibitor miRNAs in PC cells [120]. This mechanism may be elucidated by its capacity to inactivate (ERK1/2), Akt and reduce the concentrations of estrogen and insulin growth factor-1 receptor, suppressing the Akt/miR-21 signaling pathway [121].
New approaches support the benefit of RSV-loaded NPs for PC chemoprevention/chemotherapy, due to the lack of adverse reactions on normal cells. Polymeric NPs encapsulating RSV were developed and they generated substantially higher cytotoxicity to PC cells than free RSV, without negative cytotoxic repercussions on murine macrophages, even at 200 μM [122]. Similar results were obtained by the successful endeavor to expand systemic circulation and biological half-life of tRSV, utilizing solid lipid NPs, to raise its anti-tumor ability [123].
The function of RSV as a radiosensitizer by targeting cancer stem cells in radioresistant PC cells was analyzed, employing stem cell markers and EMT markers, such as vimentin and E-cadherin. Association of RSV administration with ionizing radiation, generated important decline in cell viability and stem cell markers genes and altered EMT markers [124].
Vasculogenic mimicry is specifically correlated to tumor progression and metastasis, being responsible for the poor prognosis of patients with PC. RSV decreases serum-induced invasion and vasculogenic mimicry formation in human PC cells. Blood-generated phosphorylation of erythropoietin-producing hepatocellular A2 and the expression of vascular endothelial (VE)-cadherin at the mRNA and protein positions are reduced by RSV. Blood-induced AKT signaling cascade, as well as metalloproteinase-2 and laminin subunit 5 gamma-2, are damaged by this phytochemical [125].
It is more and more acknowledged that the synergy of the glioma-associated oncogene axis and androgen receptor influences the evolution of PC. Recent in vitro and in vivo assays on xenograft models in mice confirmed that RSV suppresses the Hh signals and tumor necrosis factor receptor (TNFR)-associated factor 6 (TRAF6) levels, to ameliorate PC progression. The mechanism of action involved EMT progression, reducing the tumor size and expressions of vimentin, VEGF, and MMP-7, besides stimulating the expressions of E-cadherin and annexin 2 [126].
4. Neuroprotective Activity
4.1. Neurodegenerative Disorders
RSV, the polyphenol which can cross blood–brain barrier, has powerful neuroprotective effect on various neurodegenerative diseases by mitochondria modulation mechanism. Mitochondrial function alteration is the most frequent cause and pathological development in several neurodegenerative conditions, like amyotrophic lateral sclerosis, AD, Huntington’s disease, or Parkinson’s disease. The current accessible therapies for neurodegenerative disorders are only symptomatic treatments and have undeniable toxic reactions. In conclusion, there is a strong necessity for a safe, natural, and efficient therapy protocol [127,128,129].
AD is an escalating and neurodegenerative pathology of the cortex and hippocampus, which ultimately leads to cognitive deterioration. Despite its ambiguous etiology, the existence of β-amyloid peptides in the learning and memory areas is an indication of AD. It has been demonstrated that RSV has antioxidant, anti-inflammatory and neuroprotective effects and can reduce the toxicity and aggregation of β-amyloid peptides in the hippocampus, stimulate neurogenesis and avoid hippocampal impairment. Moreover, the antioxidant effect of RSV is very effective in neuronal differentiation, due to the activation of SIRT1. Because of the anti-inflammatory action, it inhibits M1 microglia activation, which plays a part in the induction of neurodegeneration and stimulates Th2 responses by enhancing anti-inflammatory cytokines and SIRT1 [130,131].
The mechanism and therapeutic action of RSV were explored in AD-like behavioral dysfunction caused by streptozotocin. The deteriorations in task-specific learning and memory provoked by the bilateral hippocampal microinjections of streptozotocin were improved by RSV. The mechanism is correlated to the modulation of brain-derived neurotrophic factor expression and synaptic plasticity-associated proteins in the hippocampus [132].
4.2. Chemical-Induced Neurotoxicity
The aqueous extract of RSV from methanol extracts of the roots of Vitis thunbergii var. sinuata (Regel) Rehder (Vitaceae) showed substantial neuroprotection contra glutamate-caused neurotoxicity in primary cultured rat cortical cells. By using fractionation processes, five RSV derivatives [vitisinols A, B, C, (+)-vitisin A and (+)-vitisin C] were isolated. Three of them presented the most important neuroprotective effect opposite to the neurotoxicity produced by glutamate, as recorded by a cell viability of relatively 75–85%, at concentrations varying from 0.1 μM to 10 μM [133].
4.3. Scopolamine-Induced Cognitive Impairment
Researchers evaluated the role of RSV on cognitive disorder caused by scopolamine, a muscarinic antagonist, in mice. Scopolamine induced considerable extension of transfer latency on elevated plus maze, decrease in step-down latency on a passive avoidance apparatus and also escape latency in Morris’ water maze test as well, demonstrating cognitive deterioration. Unfortunately, RSV administration did not annul the effects of scopolamine and, therefore, did not improve the loss in cognitive functions in mice [134].
4.4. Resveratrol against Brain Injury
Traumatic brain injury is the most frequent type of craniocerebral injury that generates neurological deterioration and cognitive impairment. RSV recovers cognitive activity in neurological dysfunctions and senescent prototypes as a result of its anti-inflammatory effect. Synapse creation is crucial for cognitive function and synaptophysin is one of the significant elements implicated in synapse induction. SIRT1 has a neuroprotective influence in several biological mechanisms, such as metabolism, inflammation, apoptosis, and autophagy. It was concluded that RSV enhances synaptophysin via activating the SIRT1/peroxisome proliferator-activated receptor-gamma coactivator-1 (PGC-1) cascade and promotes the post-traumatic brain injury cognitive activity [135].
The influence of RSV against brain injury caused by radiation in rats was assessed. The radiation group of rats exhibited an important reduction in catalase (CAT), SOD, glutathione peroxidase and glutathione reductase functions, including glutathione contents. The administration of RSV substantially improved these benchmarks in the brain tissues of the animals. This result is connected to the capacity of RSV to target the free radicals, stimulate the activity of the antioxidant enzymes, enhance glutathione content, and inhibit the inflammatory reactions and apoptosis markers [136].
5. Metabolic Disturbances
5.1. Metabolic Syndrome
Metabolic syndrome is an important health issue, correlated with enhanced risk factors for hepatic steatosis, which is the most frequent liver disorder nowadays. The protective actions of RSV over metabolic dysfunctions influenced by a high-fat high-fructose diet were analyzed. The following observations were made on metabolic syndrome rats: hepatic dysfunction, increased body weight, dyslipidemia, hepatic oxidative and inflammatory stress conditions, hepatic insulin resistance, stimulation of mRNA expression of sterol regulatory element binding protein 1c (SREBP-1c) and inhibition of mRNA expressions of peroxisome proliferator activated receptor-alpha (PPAR-α) and insulin receptor substrate-2 (IRS2). Treatment with RSV diminished liver steatosis, inflammatory state, and oxidative stress. Moreover, it ameliorated lipid profile, together with insulin reactivity and regressed modifications in hepatic mRNA expression levels of the investigated genes [137].
5.2. Antidiabetic Properties
Diabetic complications, such as cataract and nephropathy, have numerous and complex mechanisms. RSV was investigated for its inhibitory effect versus rat lens and kidney aldose reductase in vitro, together with its capacity to block the generation of advanced glycation end products. The phytoalexin considerably amended glycemic status and renal function in diabetic rats, with an important reduction in the production of advanced glycation end products in the kidneys. This evidence highlights the capability of RSV as a potential therapeutic factor opposed to long-term diabetic complications [138].
Quercetin and RSV have well-known favorable actions on diabetes and diabetic complication; nevertheless, the results of mixed treatment of these phytochemicals on diabetes are not completely elucidated. The combined treatment guarded the functions of hepatic glucose metabolic enzymes and structure of pancreatic β-cells from the diabetes, and it is remarkable that this cotreatment exhibited the most beneficial action on the diabetic rats [139]. Other assays proved that the nutritional polyphenols RSV and piceatannol reduce postprandial hyperglycemia and suggest that suppression of intestinal α-glucosidase function can be a possible mechanism associated to their antidiabetic effect [140].
5.3. Hyperlipidemia, Hepatic Steatosis, and Synaptic Impairment
In order to evaluate the impact of RSV on the lipid profile, dose-response meta-analysis of randomized controlled trials were conducted. The evidence showed that the intake of RSV may substantially reduce the total cholesterol, triglyceride, and low-density lipoprotein (LDL)-cholesterol levels, without modifying the level of high-density lipoprotein (HDL)-cholesterol. The decrease of LDL-cholesterol was more important in trials with an extended duration and in patients with type 2 diabetes mellitus, demonstrating that the dosage of the RSV treatment is a crucial element [141].
RSV produced via microorganism biotransformation exhibits better purity compared to the naturally obtained RSV, at a reduced expense than the chemically synthesized one. The beneficial actions of this novel product were examined in hamsters on a high-fat diet and substantially decreased liver enzymes, total cholesterol, triglycerides, serum glucose and liver weight. Moreover, HDL-cholesterol was also enhanced, while blood LDL-cholesterol was reduced. Triglycerides in liver tissue and fatty liver level were decreased and memory-associated proteins, phosphorylation of calmodulin-related protein kinase II and synaptophysin, were enhanced in the cerebrum of the hamsters. The microorganism-biotransformed RSV has the potential to defend patients from hyperlipidemia, hyperglycemia, hepatic steatosis, and synaptic alteration [142].
5.4. Anti-Obesity and Anti-Atherosclerotic Effects
Atherogenic or high lipid nutrition is recognized to generate CVDs and several studies explore the cardioprotective role of RSV. The treatment with RSV everted the anomalous blood lipid profile in a test involving atherogenic rats. Moreover, it improved the enzymatic and non-enzymatic antioxidants and stimulated the lipid metabolic proteins, demonstrating its beneficial role on lipid metabolism. Additionally, RSV repaired the tissue impairment and preserved the morphology of cardiac tissue. Therefore, RSV protects the cardiovascular system damaged by high lipid nutrition and is a favorable healthy lifestyle-adjuvant [143].
Red wines are considered a great nutritional provenience for tRSV, but also for its derivatives, such as ε-viniferin, an RSV dimer that has a better anti-adipogenesis role in metabolic disorders. Furthermore, ε-viniferin is more efficient than tRSV, due to its anti-obesity and anti-inflammatory actions in mice exposed to high-fat nutrition [144]. Other studies highlight that RSV, together with its dimers, ε-viniferin and δ-viniferin, are beneficial in restraining atherosclerosis via a related molecular mechanism, with distinct capability and competence [145].
Atherosclerosis is a pathological fundament of cardiovascular and cerebrovascular disorders, which involves signal transduction in its development. It was determined that RSV is very efficient in different stages of atherosclerosis evolution through modulating lipid metabolism, suppressing inflammatory reactions and ameliorating oxidative stress, via regulating the transforming growth factor/extracellular regulated protein kinases signaling axis [146].
5.5. Polycystic Ovary Syndrome
Polycystic ovary syndrome (PCOS) is the most frequent endocrinopathy concerning women of reproductive age, especially involving hyperandrogenism. Assays on individualized ovarian theca-interstitial cells indicated that RSV decreases androgen formation, substantially inhibiting ovarian and adrenal androgens by stimulating insulin sensitivity and reducing the insulin level [147]. Recent investigations assert that RSV presents valuable phytoestrogenic properties and induce hormonal regulation in rats with PCOS. RSV treatment generates enhanced levels of plasma adiponectin and estradiol, recuperation of normal ovarian morphology and elevated ovarian expression of nesfatin-1 and aromatase at the RNA and protein levels [148].
The nutraceutical preparation mixture consisting in RSV, alpha-lipoic acid, folic acid, vitamin D and vitamin B provided favorable results for alleviating the metabolic profile of women with PCOS. Following 12 weeks of therapy, body mass index, anthropometry and bioimpedance hallmarks were all decreased in the treated patients [149].
In a study participating 110 patients, the treatment with both RSV and myoinositol was substantially more efficient than metformin and pioglitazone in improving abnormal endocrine, metabolic markers and stress burden and has excellent potential in high-risk category of obese, oligo-anovulatory women with PCOS [150]. Another inquiry established that the protective actions of RSV on ovulation in rats with PCOS is correlated to the modulation of glycolysis-dependent mediators, such as pyruvate kinase isozyme M2, lactate dehydrogenase A and SIRT1 [151].
5.6. Anti-Aging Properties
RSV and caloric restriction are very potent treatment alternatives for anti-aging and were compared in vitro and in vivo, regarding their activities in longevity-related gene silencing information regulator, SIRT1. Interestingly, 10 μM RSV in vitro and a higher dose in vivo exhibited superior anti-aging effects and SIRT1-stimulation degrees, compared to caloric restriction, implying the promising potential of the phytocompound as a caloric restriction-mimetic [152].
RSV has a strong beneficial effect in lesion healing, regeneration and photoaging of the epidermis. Through the medium of complex mechanisms and pathways, it offers protection contra UVB radiation, which is the principal element in skin aging development. Moreover, it increases collagen synthesis via stimulating the ERs and alleviates wrinkles. In deteriorated tissues, it expedites epidermis regeneration and healing by triggering VEGF. Undeniably, RSV is a valuable polyphenol in cosmetology, dermatology, and plastic surgery, being utilized in anti-aging products or administered topically on scars and lesions. Future approaches may involve RSV in the pharmacotherapy of various dermatoses [153].
6. Hepatoprotective Effect
6.1. Liver Ischemia–Reperfusion
Ischemia–reperfusion (I–R) of the liver is a complex process that occurs during transplantation and surgery. The destructive repercussions of I–R derive from the intense generation of ROS, which can provoke instantaneous tissue deterioration and numerous disastrous cellular reactions, including apoptosis, organ collapse and inflammation. The formation of ROS in the I–R condition can impair the antioxidant system and produce liver depreciation. RSV has been demonstrated to possess antioxidant effects on liver lesion caused by I–R in mice. The results revealed that hepatic ischemia happens after liver transplantation and that the administration of RSV in advance of reperfusion improves graft survival as a consequence of suppression of protein response signaling pathways, particularly protein kinase R-like ER kinase (PERK) and inositol-requiring enzyme 1alpha (IRE1α) [154].
6.2. Non-Alcoholic Fatty Liver Disease
NAFLD is a progressive disorder concerning the liver that compels efficient therapies to prevent liver damage. A micronized formulation of tRSV, with higher absorption and enhanced antioxidant action, proved to be more efficient than simple RSV preparations in humans. It was successful in decreasing the liver adipose tissue, including reducing insulin resistance and hepatic enzymes, such as serum glutamate pyruvic transaminase and gamma-glutamyl transpeptidase [155].
NAFLD, a common cause of chronic liver disease worldwide, is linked to increased body weight and obesity. It leads to fat accumulation in the liver, resulting in insulin resistance, inflammation, oxidative stress, and altered lipid and glucose profiles. RSV may help manage NAFLD due to its anti-inflammatory, antioxidant properties, and effects similar to calorie restriction. In vitro and in vivo models show that RSV reduces liver fat and improves lipid and glucose metabolism through the activation of adenosine monophosphate (AMP)-activated protein kinase, SIRT1, and NF-κB signaling pathways [156].
Polydatin is a natural precursor and glycosylated type of RSV, with different bioactivity. The responses of polydatin and RSV were measured against energy homeostasis disproportion in mice with a high-fructose nutrition. Over-nourishment-activated ROS stimulate AMPK inhibition and metabolic dysfunction. It was firstly validated that polydatin treatment enhances the fecal concentrations of valeric and caproic acids, by altering gut microbiota; therefore, stimulating AMPK activation might be the intrinsic mechanism that polydatin is preferable to RSV in ameliorating the lipid dysmetabolism [157].
6.3. Anti-Hepatotoxic Activity
The liver is a crucial organ in the body and operates essential functions. RSV has favorable pharmacological effects in the therapy of several liver pathologies, such as fatty hepatitis, liver steatosis, liver carcinoma and liver fibrosis through its anti-inflammatory, anti-apoptotic, antioxidative and hepatoprotective actions. RSV reduces liver inflammation by inhibiting numerous proinflammatory cytokines. Additionally, it suppresses the NF-κB transcription factor, which initiates the inflammatory pathway and deactivates the PI3K/Akt/mTOR axis to inflict apoptosis. Furthermore, it decreases oxidative stress in hepatocytes by substantially diminishing malondialdehyde and NO and enhancing CAT and SOD. Because of its antioxidant, anti-inflammatory and anti-fibrotic effects, RSV curtails liver injury indicators. It is also a reliable natural antioxidant that improves the hepatotoxicity of noxious chemicals [158].
The hepatoprotective effect of RSV against ethanol-induced oxidative stress was examined in vivo. The fundamental mechanisms by which RSV exhibits its antioxidative activity on hepatic cells is explained through the generation of SOD activity and gene expression [159].
Recent investigations targeted the protective actions of RSV and avocado oil against paracetamol-generated hepatotoxicity in rats. The outcome illustrated that the total oxidant status was enlarged in the paracetamol group, while the total antioxidant status was enhanced in the RSV and RSV plus avocado oil groups. The histopathological analysis determined necrotic regions in the livers of the rats. The administration of both RSV and avocado oil in advance, inverted the oxidative stress parameters, DNA deterioration and necrosis caused by paracetamol [160].
7. Cardioprotective Properties
7.1. Coronary Heart Disease
Red wine is the most beneficial alcoholic beverage in reducing the incidence of coronary heart disease (CHD). There have been investigations of the mechanism of tRSV on human platelet aggregation and synthesis of eicosanoids from arachidonate by platelets. Numerous wine phenolic compounds and antioxidants were compared, tRSV and quercetin showing a dose-related reduction of platelet aggregation caused by thrombin and adenosine diphosphate (ADP). Moreover, tRSV suppressed the formation of thromboxane B2, hydroxyheptadecatrienoate and 12-hydroxyeicosatetraenoate, from arachidonate, conditioned by the dosage. This information emphasized the protective effect of red wine against atherosclerosis and CHD, due to tRSV [161].
7.2. Myocardial Ischemia–Reperfusion
The pre-administration of RSV has a benefic influence in mitochondrial activity and myocardial I–R damage, due to the suppression of stromal interaction molecule 2 (STIM2), through the medium of miRNA-20b-5p. This data has been recently verified in a study on 90 rats, involving assays to identify the expressions of Bcl-2, Bcl-2-associated X protein and cleaved-cysteine proteinase 3, together with the levels of mitochondrial membrane potential, ROS, and adenosine triphosphate (ATP) [162].
7.3. Resveratrol as Vascular Protective Natural Compound
Vascular metabolic dysfunction is exhibited in numerous recurrent disorders, such as atherosclerosis, hypertension, and diabetes mellitus. Current appraisals have determined that RSV has valuable influence on repairing metabolism in endothelial cells and vascular smooth muscle cells. The regulation of cell metabolism activity is connected to the suppression of glucose uptake, inhibition of glycolysis, prevention against damages caused by fatty acids through stimulation of fatty acid oxidation, reduction of lipogenesis by increasing lipolysis and elevation of glutamine uptake and synthesis [163]. Vascular smooth muscle cells senescence is a critical element that stimulates the progression of CVDs. RSV has the ability to prevent vascular senescence via inhibiting oxidative stress and mitochondrial alteration by activating E2F1/SOD2 pathway [164].
7.4. Resveratrol and Calcium Fructoborate Supplementation against Stable Angina Pectoris
The clinical and biological conditions of humans diagnosed with stable angina pectoris were investigated in a study that proposed to assess the results of per os treatment with calcium fructoborate (CaFB) and RSV. The following observations were highlighted: high-sensitivity C-reactive protein, an inflammation biomarker, was seriously reduced; the N-terminal prohormone of brain natriuretic peptide, a left ventricular function marker, was substantially decreased; the lipid markers were also influenced. The findings determined that the association of RSV with CaFB has favorable consequences in patients diagnosed with angina pectoris, improving their quality of life [165].
8. Immunostimulatory Activity
The obstructive effects of RSV on the release of mediators from bone marrow-derived mouse mast cells were verified in vitro. Regulated by immunoglobulin E (IgE), the discharge of histamine, TNF-α, leukotrienes, and PGD2 were seriously suppressed by RSV; in addition, A23187-modulated release of histamine and leukotrienes was also actively decreased. RSV did not exhibit cytotoxic effect against mast cells and can, therefore, be an important non-selective blocker of mediator release from activated mast cells [166].
The study of the influence of RSV alone and combined with cyclosporin A on the propagation of human peripheral blood T-lymphocytes, conversion into lymphoblasts, as well as IL-2 and INF-γ generation revealed that RSV can significantly inhibit the proliferation and transformation of lymphocytes and the synergic mixture of RSV and cyclosporine A can stimulate immune suppression [167].
9. Antioxidant Effect
RSV has been determined to defend LDLs against peroxidative degeneration. Being a tyrosine kinase suppressor, it also has the capacity to inhibit intracellular adhesion molecule 1 and vascular cell adhesion molecule 1 expressions and to stop the attachment of monocytes and granulocytes to endothelial cells. This mechanism is autonomous from its antioxidant action and provides a different interpretation [168].
Nutritional addition of RSV inhibits lipid peroxidation, the critical consequence of oxidative stress caused by chronic ethanol treatment. During a study on ethanol treated rats, RSV substantially counteracted the elevation in malondialdehyde, a marker of oxidative damage, concentrations in the liver, heart, brain, and testes, demonstrating its protective effect [169]. Numerous assays evaluated the inhibitory effect of RSV in oxidative stress. However, the association of RSV and vitamins C and E is more efficient in protecting the cells than any of these antioxidants alone [170].
10. Bone Protection
Nutritional supplementation of RSV has beneficial effects against hip fracture risk in elderly patients. This information was documented by research involving recently diagnosed patients of hip fracture. The investigations included the administration of RSV and piceid, in addition to RSV-rich foods, such as grapes, apples and nuts. The results were more conspicuous in female and non-obese subjects and highlighted that alimentation with RSV correlates with decreased risk of hip fracture [171].
11. Wound Healing Properties
The role of local and systemic injections of RSV on open cutaneous wound healing was compared in rats. Both of them elevated the histological scores for collagen deposition, chronic inflammation, and granulation. Neovascularization rates were substantially higher in the subcutaneous RSV treatment category than in the intraperitoneal one. Both systemic and local administrations importantly boosted wound healing and enhanced the tensile strength of the epidermis. Local subcutaneous injection of RSV emerged to be a better therapeutic approach, compared to the systemic treatment, due to the neovascularization to help wound healing [172]. On the other hand, topical administration of 5% RSV ointment was demonstrated to stimulate burn wound healing, by enhancing the progress of wound contraction, via collagen fiber synthesis, granulation tissue creation and epithelial regeneration in rats [173].
Diabetic injuries are problematic to heal because of constant inflammation and reduced angiogenesis. Platelet-derived extracellular vesicles are abundant in growth factors and cytokines that stimulate proliferation and angiogenesis. Nevertheless, individual medication therapy has reduced potency and release ability that can be improved by bioengineering, which combines active substances and materials to obtain synergic medication. A composite hydrogel was used as a wound cover for continuous RSV delivery from mesoporous silica NPs, combined with platelet-derived extracellular vesicles, and showed promising effects in modulating inflammation and angiogenesis of diabetic injuries and accelerating wound restoration [174,175].
12. Anti-Inflammatory Effect
Oxaliplatin is an average chemotherapy medication that causes neurotoxicity. To examine possible treatment variants for the neuropathic pain and inflammatory reaction induced by oxaliplatin, RSV was intrathecally injected into the spinal cord of rats. The findings revealed that RSV administration decreased COX-2 expression and obstructed ROS generation. The anti-inflammatory mechanism of RSV inhibited astrocytic activation and induced an antinociception effect that relieved neuropathic pain [176].
Assays of the significance of RSV in diabetic peripheral neuropathy emphasized the following results: the discomfort and temperature responses of diabetic mice were ameliorated; Nrf2 level was enhanced in the diabetic peripheral nerves and NF-κB axis suppression preserved nerves; RSV regulated the anti-inflammatory microenvironment of peripheral nerves through stimulating Nrf2 activation and the expression of phospho-p65 [177].
Due to its reduced bioavailability, the RSV NPs are a better solution and have higher neuroprotective role in a study involving rats with middle cerebral artery occlusion. Animals were tested for infarct volume, oxidative, inflammatory, and apoptotic markers. The mechanism of RSV involved the increased expressions of caspase-3 and caspase-9, and IL-1, IL-6 and TNF-α cytokines. Therefore, these findings show that suppression by RSV NPs has therapeutic effect in the ischemic stroke [178].
Recent explorations presented that the treatment with either monomeric tRSV or RSV dimer – gnetin C from Gnetum gnemon L., gnetum (Gnetaceae) seed extract – limits periodontitis, with higher decrease of bone loss being exhibited in the dimer group than the monomer batch and that these effects are connected to reduced oxidative stress and consequently minimization of local inflammation by downregulation of IL-1β, a proinflammatory cytokine [179]. Another finding suggests that RSV suppresses systemic local inflammatory markers and systemic endotoxin and implies that 500 mg per day of RSV is the optimal dosage for patients with periodontitis [180].
13. Antimicrobial Activity
RSV manifests antimicrobial action opposite to a remarkably extensive spectrum of bacterial and fungal species. It has the ability to modify bacterial expression of virulence nature, promoting decreased toxin formation, suppression of biofilm generation, limited motility and influencing the quorum sensing. In association with regular antibiotics, RSV stimulates the activity of aminoglycosides against Staphylococcus aureus, while it inhibits the noxious activity of fluoroquinolones against S. aureus and Escherichia coli. Moreover, topical administration of this nutraceutical has proven beneficial in acne lesions provoked by the Propionibacterium acnes. Recent data illustrates that the synergic combination of RSV with specific antibiotics may increase their antimicrobial potency, alleviating the crescent issue of antimicrobial resistance [181].
Candida albicans is the principal candidiasis-inducing fungal pathogen in humans and one of its most critical virulence elements is the capacity to synthesize biofilms. The antifungal effects of RSV alter the morphological transition of C. albicans following several hypha-inducing circumstances and reduce growth of the yeast-form and mycelia. Therefore, RSV has promising anti-Candida actions via suppressing existing or in the formation process C. albicans biofilms [182].
Topical bacterial infection produced by multidrug-resistant Pseudomonas aeruginosa is very difficult to treat, using polymyxins as the last alternative therapy, with unfavorable pharmacodynamics and pharmacokinetics, improbable to attain effective blood levels. Well-tolerated RSV had a synergic effect combined with polymyxin B therapy, when analyzed in vitro on antibacterial and anti-biofilm activities [183]. There was also established that Chilean red wines have antibacterial effect against Helicobacter pylori, due to the existence of RSV in their composition [184,185].
14. Antiviral Properties
Several studies described that RSV has a cooperative effect in association with the nucleoside analogues, such as zidovudine, zalcitabine and didanosine, stimulating their activity against human immunodeficiency virus type 1 (HIV-1). Individual RSV did not produce cell toxicity and inhibited viral replication, while in peripheral blood mononuclear cells infected with human T-cell lymphotropic virus-IIIB (HTLV-IIIB) isolate of HIV, it decreased the inhibitory concentration of the nucleoside analogues. Identical antiviral effect was documented when didanosine was merged with RSV in peripheral blood mononuclear cells infected with HIV-1. The inclusion of RSV determined a 10-time amplification of the antiviral action of didanosine in infected monocyte-derived macrophages. In a test on HTLV-IIIB-infected T-lymphocytes, the RSV–didanosine mixture, but not independently, inhibited the viral infection [186].
RSV has been demonstrated to have anti-herpes simplex virus (HSV) effect in vitro. To examine its activity in vivo, the abraded skin of mice was infected with HSV-1 and RSV cream was applied locally. The following findings were made: 25% RSV cream successfully inhibited lesion progression, while 12.5% RSV cream had the same effect only if applied one hour after infection; neither one was effective if applied twelve hours after infection. In addition, RSV cream, 10% docosanol cream and 5% acyclovir ointment were compared. RSV and acyclovir substantially obstructed the progression of HSV-1-caused epidermis lesions, while docosanol had no influence. The application of RSV provoked no dermal toxicity, such as erythema, scaling, crusting, lichenification or excoriation [187].
Regarding human cytomegalovirus replication, extremely high concentrations of RSV were necessary to generate cytotoxicity opposed to developing or stagnant human embryonic lung fibroblasts. RSV obstructed the virus-activated epidermal growth factor receptor and PI3K signal transduction as well as NF-κB and Sp1 transcription factor activation, immediately post-infection. The mechanisms of the cytomegalovirus, such as DNA replication, PI3K signaling and transcription factor activation, were all suppressed by RSV. However, the virucidal action was incompatible and also the antiviral effect of RSV was significantly reduced when administered four hours after infection. As a result, this phytocompound engaged amid attachment and entry, via arrest of epidermal growth factor receptor activation and its succeeding effectors [188].
Worldwide, rotavirus is the principal originator for viral gastroenteritis in children, with no efficient therapy. The anti-rotavirus impact of RSV was inspected in vitro, including production of virion descendants, viral polyprotein expression, genomic RNA synthesis, antigen clearance and alterations in proinflammatory cytokines/chemokines in infant mouse cells. The assays concluded that RSV strongly limited rotavirus replication by inhibiting RNA formation, protein expression, viroplasm plaque generation, progeny virion synthesis and cytopathy. RSV treatment ameliorated the gravity of diarrhea, reduced viral titers and alleviated correlated symptoms [189].
The high-occurrence hepatitis B virus infection requires successful therapeutics. A recent study prospected the influence of RSV on hepatitis B virus replication, utilizing in vitro and in vivo analysis, confirmed it suppresses cytotoxicity and virus replication. Through its mechanism of action via reducing miR-155 expression and stimulating autophagy, RSV proved to be a promising novel approach for the treatment of hepatitis B virus infection [190].
15. UHPLC–MS Analysis of trans-Resveratrol in Romanian Wines
15.1. Chemicals and Reagents
For the analysis of tRSV in wine samples through UHPLC–MS, the following chemicals and reagents were used: tRSV was purchased from Sigma-Aldrich (Taufkirchen, Germany) and utilized as a reference for identification and quantification processes. Acetonitrile, obtained from Merck (Darmstadt, Germany), served as one of the mobile phase components due to its excellent solvent properties and compatibility with MS. Formic acid, also sourced from Merck, was employed as a mobile phase modifier to enhance the ionization of tRSV during the MS detection phase. Ultrapure water was produced locally in our laboratory using the HALIOS 6 (Neptec, Montabaur, Germany) ultrapure water system and was utilized as the aqueous component of the mobile phase and for all preparations requiring solvent. All reagents were of analytical grade and were used without further purification to ensure the integrity and reproducibility of the results, ensuring optimal separation, identification, and quantification of tRSV in the studied wine samples.
15.2. Calibration Curve for Trans-Resveratrol Quantification
A stock solution of tRSV was prepared at a concentration of 1 mg/mL. From this stock solution, serial dilutions were made to create calibration standards at the following concentrations: 244.141, 122.070, 61.035, 30.518, and 15.259 ng/mL. These standards were prepared by diluting the stock solution with the initial mobile phase to achieve the precise concentrations needed. These diluted standards were then analyzed using the UHPLC–MS system under identical conditions to those of the wine samples. The resulting peak areas for tRSV at m/z 227 were plotted against their respective concentrations to construct the calibration curve. The linearity of the calibration curve was validated by the correlation coefficient (R2) obtained from the linear regression of the data. The curve was crucial for quantifying the concentration of tRSV in the wine samples by interpolating the sample peak areas against the calibration data. Measurements were made in triplicate to ensure the accuracy and reliability of the data. The limits of detection (LOD) and quantification (LOQ) for tRSV were determined based on the standard deviation (SD) of the response and the slope of the calibration curve, ensuring precise and reliable quantification [191,192,193].
15.3. Wine Samples
The study focused on the analysis of tRSV in four wine samples, all sourced from local Romanian vineyards to represent both white and red varieties, providing a comprehensive overview of tRSV content across different types of wine. The white wine samples included a Fetească Regală (W1), a traditional Romanian grape variety known for its aromatic profile and high acidity, and a Dry Muscat (W3), noted for its sweet floral aromas and dry finish. The red wine samples consisted of a Fetească Neagră (W2), an indigenous Romanian variety valued for its deep color, medium tannins, and berry flavors, and a Cabernet Sauvignon (W4), a globally recognized grape chosen for its robust structure and potential for aging. Each wine sample was stored and handled under controlled conditions to preserve its chemical integrity until analysis, aiming to explore the influence of grape type and wine style on the levels of tRSV and enhancing our understanding of how this phenolic compound varies within Romanian wines [194].
15.4. UHPLC–MS Analysis
The UHPLC–MS analysis of tRSV in wine samples was conducted using a Waters Arc System coupled with a Waters QDa detector. Initially, all samples were diluted five-fold and then filtered through 0.2 µm filters before being injected into the system, with an injection volume set at 5 µL. The mobile phase comprised two components: (A) was 0.1% formic acid in water, and (B) was 0.1% formic acid in acetonitrile. The flow rate was maintained at 0.8 mL/min. The gradient program was as follows: from 0 to 1.8 minutes, (B) increased from 2% to 9%; from 1.8 to 4 minutes, (B) held steady at 9%; from 4 to 10 minutes, (B) rose from 9% to 30%; from 10 to 15 minutes, (B) escalated from 30% to 90% and was maintained at 90% until 16 minutes; from 16 to 17 minutes, (B) rapidly decreased back to 2% for re-equilibration. The column used was a Waters CORTECS C18 (4.6×50 mm, 2.7 µm), with the temperature controlled at 28℃ and samples maintained at 10℃. A 15-minute equilibration period was allowed between each injection to ensure consistent performance. The QDa mass spectrometer operated in negative mode, with the capillary voltage set at 0.8 kV and the cone voltage at 15 V. Mass spectra were acquired in the range of m/z 100–400, with selected ion recording (SIR) at m/z 227 for the specific quantification of tRSV, providing a robust and sensitive method for analyzing the presence and concentration of this compound in the wine samples [191,192,193].
Our study included both white (W1 and W3) and red (W2 and W4) varieties, to evaluate the presence and concentration of tRSV. The calibration curve parameters, indicative of the method’s current performance, are detailed, providing insight into the potential for precise quantification. It is important to note that while this analysis offers valuable initial data, the method is undergoing further validation to refine its accuracy and reliability. The results herein serve as a foundation for subsequent validation efforts and offer an early look at tRSV levels in these wines, contributing to the broader understanding of its distribution in Romanian viniculture.
The calibration curve (Figure 1) parameters presented in Table 1 demonstrate a robust analytical method for the quantification of tRSV. The wide linearity range of 15.259 to 244.141 ng/mL indicates that the method is capable of accurately measuring tRSV across a range of concentrations that are relevant for wine samples. This range comfortably exceeds the expected concentration levels in typical wine samples, allowing for precise quantification even in wines with lower tRSV content.
R2 of 0.9979 reflects a high degree of linearity, suggesting that the response is directly proportional to the concentration over the range tested. This is critical for ensuring that the quantification is reliable, and that the method can be used confidently for routine analysis.
The equation of the calibration curve, Y = 181.7X - 197.6, provides the relationship between the instrument’s response and the concentration of tRSV. The positive slope (181.7) signifies a strong response to increasing concentrations, while the intercept (-197.6) is relatively small compared to the slope, indicating minimal background noise or interference.
LOD and LOQ are also indicators of the method’s sensitivity. With a LOD of 16.197 ng/mL and a LOQ of 49.081 ng/mL, the method is sensitive enough to detect and quantify even small amounts of tRSV. This is particularly important for scientific and regulatory purposes where it is necessary to detect all ranges of concentrations present in various wine samples.
The UHPLC chromatogram presents a distinct peak corresponding to the pure tRSV reference sample. The specificity of the peak is enhanced by the MS detection of the m/z 227 molecular ion, characteristic of tRSV. This suggests that the UHPLC–MS method provides high specificity, primarily due to the mass detection at m/z 227, ensuring that the observed peak can be confidently attributed to tRSV without interference from other substances. Such specificity is critical for accurate quantification in complex matrices like wine (Figure 2).
In addition to the distinct peak at the specific retention time (RT), the confirmation of tRSV in the samples was further substantiated by comparing the mass spectrum obtained between m/z 100–400. This range was meticulously examined to match the ionization pattern of the tRSV reference with the sample peaks. By ensuring that the spectra of the samples aligned with that of the reference tRSV, particularly the peak at m/z 227, the presence of tRSV could be confirmed with greater certainty. This comparative approach across the broad mass range enhances the reliability of the identification, ruling out the possibility of coincidental matches or false positives that might arise from other compounds eluting at similar RTs (Figure 3).
In Figure 4, we observe the elution profiles of four different wine samples labeled W1 through W4. Each chromatogram is overlaid to facilitate comparison, particularly focusing on the tRSV peak. For all samples, the tRSV peak was identified at the same RT, which is consistent with the reference peak, suggesting successful identification of tRSV in each wine. The peak intensities vary among the wine samples, which may reflect the different concentrations of tRSV in each type of wine. The RT’s consistency across all samples is a good indicator of the method’s reproducibility. Furthermore, the distinct peaks observed for tRSV in each sample, without interference from other peaks, confirm the specificity of the method, especially considering the mass detection confirmation at m/z 227. This comparative display allows for a visual assessment of the presence and relative quantity of tRSV across the different wine samples, providing an at-a-glance understanding of how tRSV concentrations may differ between white and red wines, as well as among different varieties. The clean baselines and sharp peak shapes are indicative of a well-optimized UHPLC method, essential for accurate quantification and comparison between samples (Figure 4).
Sample W1 (Fetească Regală variety) registers the highest concentration of tRSV among the white wines, suggesting that this varietal may naturally accumulate more of this compound, or that the vinification methods have been conducive to its retention. Moving to W3 (Dry Muscat variety), we can see that the tRSV content is significantly lower, which may reflect inherent varietal differences or distinct processing techniques affecting the final tRSV levels. When looking at the red wines, W2 (Fetească Neagră variety) contains the highest concentration of tRSV in the selection, aligning with the general observation that red wines often have higher levels of tRSV due to the extended contact with grape skins during fermentation, which typically enriches the wine with more phenolic compounds. W4 (Cabernet Sauvignon variety), however, has a notably lower level of tRSV, which might be influenced by a myriad of factors including viticultural conditions, grape harvesting parameters, or the specific methodologies employed during the winemaking process (Table 2).
The low SD associated with each measurement implies that the method used for quantification is both precise and reliable. This data provides valuable insights for winemaking strategies, could inform consumer choices regarding healthful components in wine, and may stimulate additional research into the beneficial properties of wine polyphenols.
16. Conclusions
Stilbenoids are natural compounds of the C6–C2–C6 type, consisting of two benzene nuclei linked together by an ethylene bridge. The double bond between the two benzene nuclei can have E (trans) or Z (cis) configuration. The trans configuration is the predominant form, much more pharmacologically active. For their role as phytoalexins, respectively hormones that trigger defense mechanisms, stilbenoids are synthesized by the plant kingdom in response to external stimuli, such as infection or UV radiation. More than 1000 stilbenoids have been isolated and identified, the best known of which are the natural components RSV, piceid, piceatannol, pterostilbene, astringin, rapontigenol, viniferin, pallidol, hopeaphenol. Grapes are the main source of natural stilbenoids. RSV is a potential chemopreventive agent, both in vivo and in vitro, considering its inhibitory effects on cellular processes associated with cancer induction and progression. The influence of RSV on numerous enzyme systems explains its antioxidant, antimutagenic and anticarcinogenic potential. In some bioassay systems, RSV has been shown to provide protection against several types of cancer. RSV has an antioxidant effect, but also numerous pharmacological actions useful for maintaining cardiovascular health and protection against aging, especially against diseases associated with aging, neurodegeneration, inflammatory, metabolic, or immune diseases and some types of cancer. RSV modulates signaling pathways that limit the spread of tumor cells, protects nerve cells from damage, is useful in the prevention of diabetes, and generally acts as an anti-aging agent that alleviates the symptoms of aging. It was highlighted that RSV could ameliorate the consequences of an unhealthy lifestyle caused by an exaggerated caloric intake. A moderate consumption of red wine, over a long period of time, protects the body against CHDs and could be the main factor responsible for the so-called “French paradox”. tRSV from the Romanian wine samples, both white and red varieties, can be identified and quantified by UHPLC–MS.
Author Contributions
Conceptualization, L.E.B., I.B. and G.D.M.; methodology, A.B., A.E.S., A.R., A.D. and M.V.C.; writing—original draft preparation, L.E.B., A.B. and G.D.M.; writing—review and editing, A.B., I.B. and G.D.M.; supervision, L.E.B., I.B. and C.B.; funding acquisition, I.B. and A.E.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The manuscript does not contain experiments on laboratory animals.
Informed Consent Statement
The manuscript does not contain clinical studies or patient data.
Data Availability Statement
Data described in the manuscript will be made publicly and freely available without restriction at: https://docs.google.com/document/d/1vVW1wFaxOrqwMIpVTSpMtgp8HrwQqx4r/edit?usp=sharing&ouid=106104952021876684289&rtpof=true&sd=true (accessed on 25 April 2024).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kaur, G.; Kaur, R.; Sodhi, G.K.; George, N.; Rath, S.K.; Walia, H.K.; Dwibedi, V.; Saxena, S. Stilbenes: A journey from folklore to pharmaceutical innovation. Arch. Microbiol. 2024, 206, 229. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.Y.; Gao, Y.; Shahidi, F. Stilbenoids: Chemistry, occurrence, bioavailability and health effects – a review. J. Food Bioact. 2021, 13, 20–31. [Google Scholar] [CrossRef]
- Pawlus, A.D.; Waffo-Téguo, P.; Shaver, J.; Mérillon, J.M. Stilbenoid chemistry from wine and the genus Vitis, a review. OENO One 2012, 46, 57–111. [Google Scholar] [CrossRef]
- Faisal, Z.; Mazhar, A.; Batool, S.A.; Akram, N.; Hassan, M.; Khan, M.U.; Afzaal, M.; Hassan, U.U.; Shah, Y.A.; Desta, D.T. Exploring the multimodal health-promoting properties of resveratrol: A comprehensive review. Food Sci. Nutr. 2024, 12, 2240–2258. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Bahroudi, Z.; Shoorei, H.; Hussen, B.M.; Talebi, S.F.; Baig, S.G.; Taheri, M.; Ayatollahi, S.A. Disease-associated regulation of gene expression by resveratrol: Special focus on the PI3K/AKT signaling pathway. Cancer Cell Int. 2022, 22, 298. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G. Nonalcoholic compounds of wine: The phytoestrogen resveratrol and moderate red wine consumption during menopause. Drugs Exp. Clin. Res. 1999, 25, 111–114. [Google Scholar] [PubMed]
- Huang, S.; Qi, B.; Yang, L.; Wang, X.; Huang, J.; Zhao, Y.; Hu, Y.; Xiao, W. Phytoestrogens, novel dietary supplements for breast cancer. Biomed. Pharmacother. 2023, 160, 114341. [Google Scholar] [CrossRef]
- Gehm, B.D.; McAndrews, J.M.; Chien, P.Y.; Jameson, J.L. Resveratrol, a polyphenolic compound found in grapes and wine, is an agonist for the estrogen receptor. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 14138–14143. [Google Scholar] [CrossRef] [PubMed]
- Alamolhodaei, N.S.; Tsatsakis, A.M.; Ramezani, M.; Hayes, A.W.; Karimi, G. Resveratrol as MDR reversion molecule in breast cancer: An overview. Food Chem. Toxicol. 2017, 103, 223–232. [Google Scholar] [CrossRef]
- Visioli, F.; Panaite, S.A.; Tomé-Carneiro, J. Wine’s phenolic compounds and health: A Pythagorean view. Molecules 2020, 25, 4105. [Google Scholar] [CrossRef]
- Rodríguez-Vaquero, M.J.; Vallejo, C.V.; Aredes-Fernández, P.A. Antibacterial, antioxidant and antihypertensive properties of polyphenols from Argentinean red wines varieties. Open J. Pharmacol. Pharmacother. 2020, 5, 1–6. [Google Scholar] [CrossRef]
- Magrone, T.; Magrone, M.; Russo, M.A.; Jirillo, E. Recent advances on the anti-inflammatory and antioxidant properties of red grape polyphenols: In vitro and in vivo studies. Antioxidants (Basel) 2019, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Frémont, L. Biological effects of resveratrol. Life Sci. 2000, 66, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.J.; Jyostna, D.; Krupadanam, G.L.D.; Srimannarayana, G. Phenanthrene and stilbenes from Pterolobium hexapetallum. Phytochemistry 1988, 27, 3625–3626. [Google Scholar] [CrossRef]
- Duca, A.; Sturza, A.; Moacă, E.A.; Negrea, M.; Lalescu, V.D.; Lungeanu, D.; Dehelean, C.A.; Muntean, D.M.; Alexa, E. Identification of resveratrol as bioactive compound of propolis from Western Romania and characterization of phenolic profile and antioxidant activity of ethanolic extracts. Molecules 2019, 24, 3368. [Google Scholar] [CrossRef]
- Sun, N.J.; Woo, S.H.; Cassady, J.M.; Snapka, R.M. DNA polymerase and topoisomerase II inhibitors from Psoralea corylifolia. J. Nat. Prod. 1998, 61, 362–366. [Google Scholar] [CrossRef]
- Fuloria, S.; Sekar, M.; Khattulanuar, F.S.; Gan, S.H.; Rani, N.N.I.M.; Ravi, S.; Subramaniyan, V.; Jeyabalan, S.; Begum, M.Y.; Chidambaram, K.; et al. Chemistry, biosynthesis and pharmacology of viniferin: Potential resveratrol-derived molecules for new drug discovery, development and therapy. Molecules 2022, 27, 5072. [Google Scholar] [CrossRef]
- Pezet, R.; Perret, C.; Jean-Denis, J.B.; Tabacchi, R.; Gindro, K.; Viret, O. Delta-viniferin, a resveratrol dehydrodimer: One of the major stilbenes synthesized by stressed grapevine leaves. J. Agric. Food Chem. 2003, 51, 5488–5492. [Google Scholar] [CrossRef]
- He, S.; Jiang, L.; Wu, B.; Pan, Y.; Sun, C. Pallidol, a resveratrol dimer from red wine, is a selective singlet oxygen quencher. Biochem. Biophys. Res. Commun. 2009, 379, 283–287. [Google Scholar] [CrossRef]
- Guebailia, H.A.; Chira, K.; Richard, T.; Mabrouk, T.; Furiga, A.; Vitrac, X.; Monti, J.P.; Delaunay, J.C.; Mérillon, J.M. Hopeaphenol: The first resveratrol tetramer in wines from North Africa. J. Agric. Food Chem. 2006, 54, 9559–9564. [Google Scholar] [CrossRef]
- Aziz, M.H.; Kumar, R.; Ahmad, N. Cancer chemoprevention by resveratrol: In vitro and in vivo studies and the underlying mechanisms (review). Int. J. Oncol. 2003, 23, 17–28. [Google Scholar] [CrossRef]
- Clément, M.V.; Hirpara, J.L.; Chawdhury, S.H.; Pervaiz, S. Chemopreventive agent resveratrol, a natural product derived from grapes, triggers CD95 signaling-dependent apoptosis in human tumor cells. Blood 1998, 92, 996–1002. [Google Scholar] [CrossRef]
- Fontecave, M.; Lepoivre, M.; Elleingand, E.; Gerez, C.; Guittet, O. Resveratrol, a remarkable inhibitor of ribonucleotide reductase. FEBS Lett. 1998, 421, 277–279. [Google Scholar] [CrossRef]
- Surh, Y.J.; Hurh, Y.J.; Kang, J.Y.; Lee, E.; Kong, G.; Lee, S.J. Resveratrol, an antioxidant present in red wine, induces apoptosis in human promyelocytic leukemia (HL-60) cells. Cancer Lett. 1999, 140, 1–10. [Google Scholar] [CrossRef]
- Bernhard, D.; Tinhofer, I.; Tonko, M.; Hübl, H.; Ausserlechner, M.J.; Greil, R.; Kofler, R.; Csordas, A. Resveratrol causes arrest in the S-phase prior to Fas-independent apoptosis in CEM-C7H2 acute leukemia cells. Cell Death Differ. 2000, 7, 834–842. [Google Scholar] [CrossRef]
- Park, J.W.; Choi, Y.J.; Jang, M.A.; Lee, Y.S.; Jun, D.Y.; Suh, S.I.; Baek, W.K.; Suh, M.H.; Jin, I.N.; Kwon, T.K. Chemopreventive agent resveratrol, a natural product derived from grapes, reversibly inhibits progression through S and G2 phases of the cell cycle in U937 cells. Cancer Lett. 2001, 163, 43–49. [Google Scholar] [CrossRef]
- Tsan, M.F.; White, J.E.; Maheshwari, J.G.; Bremner, T.A.; Sacco, J. Resveratrol induces Fas signalling-independent apoptosis in THP-1 human monocytic leukaemia cells. Br. J. Haematol. 2000, 109, 405–412. [Google Scholar] [CrossRef]
- Wieder, T.; Prokop, A.; Bagci, B.; Essmann, F.; Bernicke, D.; Schulze-Osthoff, K.; Dörken, B.; Schmalz, H.G.; Daniel, P.T.; Henze, G. Piceatannol, a hydroxylated analog of the chemopreventive agent resveratrol, is a potent inducer of apoptosis in the lymphoma cell line BJAB and in primary, leukemic lymphoblasts. Leukemia 2001, 15, 1735–1742. [Google Scholar] [CrossRef]
- Billard, C.; Izard, J.C.; Roman, V.; Kern, C.; Mathiot, C.; Mentz, F.; Kolb, J.P. Comparative antiproliferative and apoptotic effects of resveratrol, ε-viniferin and vine-shots derived polyphenols (vineatrols) on chronic B lymphocytic leukemia cells and normal human lymphocytes. Leuk. Lymphoma 2002, 43, 1991–2002. [Google Scholar] [CrossRef]
- Su, J.L.; Lin, M.T.; Hong, C.C.; Chang, C.C.; Shiah, S.G.; Wu, C.W.; Chen, S.T.; Chau, Y.P.; Kuo, M.L. Resveratrol induces FasL-related apoptosis through Cdc42 activation of ASK1/JNK-dependent signaling pathway in human leukemia HL-60 cells. Carcinogenesis 2005, 26, 1–10. [Google Scholar] [CrossRef]
- Ahmad, K.A.; Clement, M.V.; Hanif, I.M.; Pervaiz, S. Resveratrol inhibits drug-induced apoptosis in human leukemia cells by creating an intracellular milieu nonpermissive for death execution. Cancer Res. 2004, 64, 1452–1459. [Google Scholar] [CrossRef]
- Baatout, S.; Derradji, H.; Jacquet, P.; Ooms, D.; Michaux, A.; Mergeay, M. Enhanced radiation-induced apoptosis of cancer cell lines after treatment with resveratrol. Int. J. Mol. Med. 2004, 13, 895–902. [Google Scholar] [CrossRef]
- Lin, H.Y.; Tang, H.Y.; Davis, F.B.; Davis, P.J. Resveratrol and apoptosis. Ann. N. Y. Acad. Sci. 2011, 1215, 79–88. [Google Scholar] [CrossRef]
- Quoc Trung, L.; Espinoza, J.L.; Takami, A.; Nakao, S. Resveratrol induces cell cycle arrest and apoptosis in malignant NK cells via JAK2/STAT3 pathway inhibition. PLoS One 2013, 8, e55183. [Google Scholar] [CrossRef]
- Takashina, M.; Inoue, S.; Tomihara, K.; Tomita, K.; Hattori, K.; Zhao, Q.L.; Suzuki, T.; Noguchi, M.; Ohashi, W.; Hattori, Y. Different effect of resveratrol to induction of apoptosis depending on the type of human cancer cells. Int. J. Oncol. 2017, 50, 787–797. [Google Scholar] [CrossRef]
- Liontas, A.; Yeger, H. Curcumin and resveratrol induce apoptosis and nuclear translocation and activation of p53 in human neuroblastoma. Anticancer Res. 2004, 24, 987–998. [Google Scholar] [PubMed]
- Chen, Y.; Tseng, S.H.; Lai, H.S.; Chen, W.J. Resveratrol-induced cellular apoptosis and cell cycle arrest in neuroblastoma cells and antitumor effects on neuroblastoma in mice. Surgery 2004, 136, 57–66. [Google Scholar] [CrossRef]
- Tseng, S.H.; Lin, S.M.; Chen, J.C.; Su, Y.H.; Huang, H.Y.; Chen, C.K.; Lin, P.Y.; Chen, Y. Resveratrol suppresses the angiogenesis and tumor growth of gliomas in rats. Clin. Cancer Res. 2004, 10, 2190–2202. [Google Scholar] [CrossRef]
- Rahman, M.A.; Kim, N.H.; Kim, S.H.; Oh, S.M.; Huh, S.O. Antiproliferative and cytotoxic effects of resveratrol in mitochondria-mediated apoptosis in rat b103 neuroblastoma cells. Korean J. Physiol. Pharmacol. 2012, 16, 321–326. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, L.; Kuo, J.; Kuo, K.; Gautam, S.C.; Groc, L.; Rodriguez, A.I.; Koubi, D.; Hunter, T.J.; Corcoran, G.B.; et al. Resveratrol-induced apoptotic death in human U251 glioma cells. Mol. Cancer Ther. 2005, 4, 554–561. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Y.; Zhu, K.; Song, L.; Tao, M.; Huang, P.; Pan, Y. Resveratrol inhibits glioma cell growth via targeting LRIG1. J. BUON 2018, 23, 403–409. [Google Scholar]
- Yamamoto, M.; Suzuki, S.O.; Himeno, M. Resveratrol-induced autophagy in human U373 glioma cells. Oncol. Lett. 2010, 1, 489–493. [Google Scholar] [CrossRef]
- Zhang, W.; Murao, K.; Zhang, X.; Matsumoto, K.; Diah, S.; Okada, M.; Miyake, K.; Kawai, N.; Fei, Z.; Tamiya, T. Resveratrol represses YKL-40 expression in human glioma U87 cells. BMC Cancer 2010, 10, 593. [Google Scholar] [CrossRef]
- Jiao, Y.; Li, H.; Liu, Y.; Guo, A.; Xu, X.; Qu, X.; Wang, S.; Zhao, J.; Li, Y.; Cao, Y. Resveratrol inhibits the invasion of glioblastoma-initiating cells via down-regulation of the PI3K/Akt/NF-κB signaling pathway. Nutrients 2015, 7, 4383–4402. [Google Scholar] [CrossRef]
- Zhang, C.; Peng, Q.; Tang, Y.; Wang, C.; Wang, S.; Yu, D.; Hou, S.; Wang, Y.; Zhang, L.; Lin, N. Resveratrol ameliorates glioblastoma inflammatory response by reducing NLRP3 inflammasome activation through inhibition of the JAK2/STAT3 pathway. J. Cancer Res. Clin. Oncol. 2024, 150, 168. [Google Scholar] [CrossRef]
- Hu, S.A.; Wei, W.; Yuan, J.; Cheng, J. Resveratrol inhibits proliferation in HBL-52 meningioma cells. Onco Targets Ther. 2019, 12, 11579–11586. [Google Scholar] [CrossRef]
- Stahl, S.; Chun, T.Y.; Gray, W.G. Phytoestrogens act as estrogen agonists in an estrogen-responsive pituitary cell line. Toxicol. Appl. Pharmacol. 1998, 152, 41–48. [Google Scholar] [CrossRef]
- Behroozaghdam, M.; Dehghani, M.; Zabolian, A.; Kamali, D.; Javanshir, S.; Hasani Sadi, F.; Hashemi, M.; Tabari, T.; Rashidi, M.; Mirzaei, S.; et al. Resveratrol in breast cancer treatment: From cellular effects to molecular mechanisms of action. Cell. Mol. Life Sci. 2022, 79, 539. [Google Scholar] [CrossRef]
- Quarta, A.; Gaballo, A.; Pradhan, B.; Patra, S.; Jena, M.; Ragusa, A. Beneficial oxidative stress-related trans-resveratrol effects in the treatment and prevention of breast cancer. Appl. Sci. 2021, 11, 11041. [Google Scholar] [CrossRef]
- Gu, Y.; Fei, Z. Mesoporous silica nanoparticles loaded with resveratrol are used for targeted breast cancer therapy. J. Oncol. 2022, 2022, 8471331. [Google Scholar] [CrossRef]
- Khazaei, M.R.; Bozorgi, M.; Khazaei, M.; Aftabi, M.; Bozorgi, A. Resveratrol nanoformulation inhibits invasive breast cancer cell growth through autophagy induction: An in vitro study. Cell J. 2024, 26, 112–120. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, L.; Chen, T.; Guo, W.; Bao, X.; Wang, D.; Ren, B.; Wang, H.; Li, Y.; Wang, Y.; et al. Anticancer effects of resveratrol-loaded solid lipid nanoparticles on human breast cancer cells. Molecules 2017, 22, 1814. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Song, C.; Zhang, J.; Zhao, X. Targeted delivery and apoptosis induction activity of peptide-transferrin targeted mesoporous silica encapsulated resveratrol in MCF-7 cells. J. Pharm. Pharmacol. 2023, 75, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.P.L.; Pezzuto, J.M. Cancer chemopreventive activity of resveratrol. Ann. N. Y. Acad. Sci. 2002, 957, 210–229. [Google Scholar] [CrossRef] [PubMed]
- Kurzava Kendall, L.; Ma, Y.; Yang, T.; Lubecka, K.; Stefanska, B. Epigenetic effects of resveratrol on oncogenic signaling in breast cancer. Nutrients 2024, 16, 699. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.A.; Choi, B.T.; Lee, Y.T.; Park, D.I.; Rhee, S.H.; Park, K.Y.; Choi, Y.H. Resveratrol inhibits cell proliferation and induces apoptosis of human breast carcinoma MCF-7 cells. Oncol. Rep. 2004, 11, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Laux, M.T.; Aregullin, M.; Berry, J.P.; Flanders, J.A.; Rodriguez, E. Identification of a p53-dependent pathway in the induction of apoptosis of human breast cancer cells by the natural product, resveratrol. J. Altern. Complement. Med. 2004, 10, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.Y.; Chiang, E.P.I.; Sun, Y.C. Resveratrol inhibits heregulin-β1-mediated matrix metalloproteinase-9 expression and cell invasion in human breast cancer cells. J. Nutr. Biochem. 2008, 19, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, Q.M.; Lu, Y.Y.; Zhang, H.; Chen, Q.L.; Zhao, M.; Su, S.B. Resveratrol inhibits the migration and metastasis of MDA-MB-231 human breast cancer by reversing TGF-β1-induced epithelial–mesenchymal transition. Molecules 2019, 24, 1131. [Google Scholar] [CrossRef] [PubMed]
- da Costa, P.S.; Ramos, P.S.; Ferreira, C.; Silva, J.L.; El-Bacha, T.; Fialho, E. Pro-oxidant effect of resveratrol on human breast cancer MCF-7 cells is associated with CK2 inhibition. Nutr. Cancer 2022, 74, 2142–2151. [Google Scholar] [CrossRef] [PubMed]
- Karabekir, S.C.; Özgörgülü, A. Possible protective effects of resveratrol in hepatocellular carcinoma. Iran. J. Basic Med. Sci. 2020, 23, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Pan, C.; Zhao, S.; Wang, Z.; Zhang, H.; Wu, W. Resveratrol inhibits tumor necrosis factor-alpha-mediated matrix metalloproteinase-9 expression and invasion of human hepatocellular carcinoma cells. Biomed. Pharmacother. 2008, 62, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.Y.; Chong, S.A.; Nam, M.J. Resveratrol induces apoptosis in human SK-HEP-1 hepatic cancer cells. Cancer Genomics Proteomics 2009, 6, 263–268. [Google Scholar]
- Moghadam, D.; Zarei, R.; Vakili, S.; Ghojoghi, R.; Zarezade, V.; Veisi, A.; Sabaghan, M.; Azadbakht, O.; Behrouj, H. The effect of natural polyphenols Resveratrol, Gallic acid, and Kuromanin chloride on human telomerase reverse transcriptase (hTERT) expression in HepG2 hepatocellular carcinoma: Role of SIRT1/Nrf2 signaling pathway and oxidative stress. Mol. Biol. Rep. 2023, 50, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Skonieczna, M.; Adamiec-Organisciok, M.; Hudy, D.; Dziedzic, A.; Los, L.; Skladany, L.; Grgurevic, I.; Filipec-Kanizaj, T.; Jagodzinski, M.; Kukla, M.; et al. Hepatocellular cancer cell lines, Hep-3B and Hep-G2 display the pleiotropic response to resveratrol and berberine. Adv. Med. Sci. 2022, 67, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zong, L.; Chen, X.; Jiang, Z.; Nan, L.; Li, J.; Duan, W.; Lei, J.; Zhang, L.; Ma, J.; et al. Resveratrol in the treatment of pancreatic cancer. Ann. N. Y. Acad. Sci. 2015, 1348, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Sun, R.; Yu, Y.; Gou, S.; Zhao, G.; Wang, C. Antiproliferative effect of resveratrol in pancreatic cancer cells. Phytother. Res. 2010, 24, 1637–1644. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yang, L.; Tian, W.; Li, J.; Liu, J.; Zhu, M.; Zhang, Y.; Yang, Y.; Liu, F.; Zhang, Q.; et al. Resveratrol plays dual roles in pancreatic cancer cells. J. Cancer Res. Clin. Oncol. 2014, 140, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhang, L.; Yang, Y.; Wang, M.; Liu, T.; Ji, W.; Liu, Y.; Lv, H.; Zhao, Y.; Chen, X.; et al. Polydopamine-based resveratrol–hyaluronidase nanomedicine inhibited pancreatic cancer cell invasive phenotype in hyaluronic acid enrichment tumor sphere model. ACS Pharmacol. Transl. Sci. 2024, 7, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, G.; Chen, Y.; Mu, Z.; Zhou, H.; Zhang, L. Resveratrol pre-treatment alleviated caerulein-induced acute pancreatitis in high-fat diet-feeding mice via suppressing the NF-κB proinflammatory signaling and improving the gut microbiota. BMC Complement. Med. Ther. 2022, 22, 189. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.G.; Hong, T.; Shimada, Y.; Komoto, I.; Kawabe, A.; Ding, Y.; Kaganoi, J.; Hashimoto, Y.; Imamura, M. Suppression of N-nitrosomethylbenzylamine (NMBA)-induced esophageal tumorigenesis in F344 rats by resveratrol. Carcinogenesis 2002, 23, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.B.; Yan, Y.; Sun, Y.N.; Zhu, J.R. Resveratrol induces apoptosis in human esophageal carcinoma cells. World J. Gastroenterol. 2003, 9, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Ashrafizadeh, M.; Rafiei, H.; Mohammadinejad, R.; Farkhondeh, T.; Samarghandian, S. Anti-tumor activity of resveratrol against gastric cancer: A review of recent advances with an emphasis on molecular pathways. Cancer Cell Int. 2021, 21, 66. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, Y.; Zhu, B.; Liu, Q.; Yao, Q.; Zhao, G. Resveratrol induces apoptosis in SGC-7901 gastric cancer cells. Oncol. Lett. 2018, 16, 2949–2956. [Google Scholar] [CrossRef] [PubMed]
- Rojo, D.; Madrid, A.; Martín, S.S.; Párraga, M.; Silva Pinhal, M.A.; Villena, J.; Valenzuela-Valderrama, M. Resveratrol decreases the invasion potential of gastric cancer cells. Molecules 2022, 27, 3047. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Xia, L. Resveratrol inhibits the proliferation, invasion, and migration, and induces the apoptosis of human gastric cancer cells through the MALAT1/miR-383-5p/DDIT4 signaling pathway. J. Gastrointest. Oncol. 2022, 13, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Su, N.; Li, L.; Zhou, E.; Li, H.; Wu, S.; Cao, Z. Resveratrol downregulates miR-155-5p to block the malignant behavior of gastric cancer cells. Biomed Res. Int. 2022, 2022, 6968641. [Google Scholar] [CrossRef] [PubMed]
- Ostwal, V.; Ramaswamy, A.; Bhargava, P.; Srinivas, S.; Mandavkar, S.; Chaugule, D.; Peelay, Z.; Baheti, A.; Tandel, H.; Jadhav, V.K.; et al. A pro-oxidant combination of resveratrol and copper reduces chemotherapy-related non-haematological toxicities in advanced gastric cancer: Results of a prospective open label phase II single-arm study (RESCU III study). Med. Oncol. 2022, 40, 17. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Y.; Zhai, J.; Huan. X.K.; Xu, W.W.; Tian, J.; Farhood, B. A systematic review of the therapeutic potential of resveratrol during colorectal cancer chemotherapy. Mini Rev. Med. Chem. 2023, 23, 1137–1152. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Akao, Y.; Tanaka, T.; Iinuma, M.; Nozawa, Y. Vaticanol C, a novel resveratrol tetramer, inhibits cell growth through induction of apoptosis in colon cancer cell lines. Biol. Pharm. Bull. 2002, 25, 147–148. [Google Scholar] [CrossRef] [PubMed]
- Juan, M.E.; Wenzel, U.; Daniel, H.; Planas, J.M. Resveratrol induces apoptosis through ROS-dependent mitochondria pathway in HT-29 human colorectal carcinoma cells. J. Agric. Food Chem. 2008, 56, 4813–4818. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Luo, H.; Deng, Y.; Yao, X.; Zhang, J.; He, B. Resveratrol inhibits proliferation and induces apoptosis via the Hippo/YAP pathway in human colon cancer cells. Biochem. Biophys. Res. Commun. 2022, 636, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Brown, V.A.; Jones, D.J.L.; Britton, R.G.; Hemingway, D.; Miller, A.S.; West, K.P.; Booth, T.D.; Perloff, M.; Crowell, J.A.; et al. Clinical pharmacology of resveratrol and its metabolites in colorectal cancer patients. Cancer Res. 2010, 70, 7392–7399. [Google Scholar] [CrossRef] [PubMed]
- Czapla, J.; Drzyzga, A.; Matuszczak, S.; Pilny, E.; Cichoń, T.; Stojecki, K.; Smolarczyk, R. The complex composition of trans-resveratrol, quercetin, vitamin E and selenium inhibits the growth of colorectal carcinoma. Anticancer Res. 2022, 42, 4763–4772. [Google Scholar] [CrossRef] [PubMed]
- Khayat, M.T.; Zarka, M.A.; El-Telbany, D.F.A.; El-Halawany, A.M.; Kutbi, H.I.; Elkhatib, W.F.; Noreddin, A.M.; Khayyat, A.N.; El-Telbany, R.F.A.; Hammad, S.F.; et al. Intensification of resveratrol cytotoxicity, pro-apoptosis, oxidant potentials in human colorectal carcinoma HCT-116 cells using zein nanoparticles. Sci. Rep. 2022, 12, 15235. [Google Scholar] [CrossRef] [PubMed]
- Momchilova, A.; Pankov, R.; Staneva, G.; Pankov, S.; Krastev, P.; Vassileva, E.; Hazarosova, R.; Krastev, N.; Robev, B.; Nikolova, B.; et al. Resveratrol affects sphingolipid metabolism in A549 lung adenocarcinoma cells. Int. J. Mol. Sci. 2022, 23, 10870. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.; Iyer, A.K.V.; Yakisich, J.S.; Azad, N. Anti-tumorigenic effects of resveratrol in lung cancer cells through modulation of c-FLIP. Curr. Cancer Drug Targets 2017, 17, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, F.; Wang, F.; Li, J.; Lin, C.; Du, J. Resveratrol inhibits the proliferation of A549 cells by inhibiting the expression of COX-2. Onco Targets Ther. 2018, 11, 2981–2989. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, R.; Chang, Z.; Wang, X. Resveratrol activates CD8+ T cells through IL-18 bystander activation in lung adenocarcinoma. Front. Pharmacol. 2022, 13, 1031438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chiu, J.; Zhang, H.; Qi, T.; Tang, Q.; Ma, K.; Lu, H.; Li, G. Autophagic cell death induced by resveratrol depends on the Ca2+/AMPK/mTOR pathway in A549 cells. Biochem. Pharmacol. 2013, 86, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fan, Y.; Zhang, Y.; Liu, Y.; Yu, Y.; Ma, M. Resveratrol induces autophagy and apoptosis in non-small-cell lung cancer cells by activating the NGFR-AMPK-mTOR pathway. Nutrients 2022, 14, 2413. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Liang, C.; Wang, R.; Yi, K.; Zhou, X.; Li, X.; Chen, Y.; Miao, D.; Zhong, C.; Zhu, J. Resveratrol suppresses lung cancer by targeting cancer stem-like cells and regulating tumor microenvironment. J. Nutr. Biochem. 2022, 112, 109211. [Google Scholar] [CrossRef] [PubMed]
- Davoodvandi, A.; Darvish, M.; Borran, S.; Nejati, M.; Mazaheri, S.; Reza Tamtaji, O.; Hamblin, M.R.; Masoudian, N.; Mirzaei, H. The therapeutic potential of resveratrol in a mouse model of melanoma lung metastasis. Int. Immunopharmacol. 2020, 88, 106905. [Google Scholar] [CrossRef] [PubMed]
- Stocco, B.; Toledo, K.; Salvador, M.; Paulo, M.; Koyama, N.; Torqueti Toloi, M.R. Dose-dependent effect of resveratrol on bladder cancer cells: Chemoprevention and oxidative stress. Maturitas 2012, 72, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tang, H.; Zeng, X.; Ye, D.; Liu, J. Resveratrol inhibits proliferation, migration and invasion via Akt and ERK1/2 signaling pathways in renal cell carcinoma cells. Biomed. Pharmacother. 2018, 98, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Almeida, T.C.; Melo, A.S.; Lima, A.P.B.; Branquinho, R.T.; da Silva, G.N. Resveratrol induces the production of reactive oxygen species, interferes with the cell cycle, and inhibits the cell migration of bladder tumour cells with different TP53 status. Nat. Prod. Res. 2023, 37, 3838–3843. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, K.; AlSharif, D.; Mazza, C.; Syar, P.; Al Sharif, M.; Fata, J.E. Resveratrol and pterostilbene exhibit anticancer properties involving the downregulation of HPV oncoprotein E6 in cervical cancer cells. Nutrients 2018, 10, 243. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qiu, R.L.; Lin, Y.; Cai, Y.; Bian, Y.; Fan, Y.; Gao, X.J. Resveratrol suppresses human cervical carcinoma cell proliferation and elevates apoptosis via the mitochondrial and p53 signaling pathways. Oncol. Lett. 2018, 15, 9845–9851. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Liu, Z.; Zhou, X.; Peng, F.; Wang, Z.; Li, F.; Li, M. Resveratrol induces apoptosis, suppresses migration, and invasion of cervical cancer cells by inhibiting the Hedgehog signaling pathway. Biomed Res. Int. 2022, 2022, 8453011. [Google Scholar] [CrossRef] [PubMed]
- Stakleff, K.S.; Sloan, T.; Blanco, D.; Marcanthony, S.; Booth, T.D.; Bishayee, A. Resveratrol exerts differential effects in vitro and in vivo against ovarian cancer cells. Asian Pac. J. Cancer Prev. 2012, 13, 1333–1340. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Qin, Z.; Li, F.; Zhang, H.; Fang, Z.; Hao, E. Apoptotic cell death induced by resveratrol is partially mediated by the autophagy pathway in human ovarian cancer cells. PLoS One 2015, 10, e0129196. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Zheng, G.Z.; Xie, P.; Zhang, Q.H.; Lin, F.X.; Chang, B.; Hu, Q.X.; Du, S.X.; Li, X.D. Antitumor activity of resveratrol against human osteosarcoma cells: A key role of Cx43 and Wnt/β-catenin signaling pathway. Oncotarget 2017, 8, 111419–111432. [Google Scholar] [CrossRef] [PubMed]
- Viegas, S.; Marinheiro, D.; Bastos, V.; Daniel-da-Silva, A.L.; Vieira, R.; Oliveira, H.; Almeida, J.C.; Ferreira, B.J.M.L. Resveratrol-loaded polydimethylsiloxane–silica hybrid materials: Synthesis, characterization, and antitumoral activity. Polymers (Basel) 2024, 16, 879. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bäckesjö, C.M.; Haldosén, L.A.; Lindgren, U. Resveratrol inhibits proliferation and promotes apoptosis of osteosarcoma cells. Eur. J. Pharmacol. 2009, 609, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Wang, L.; Fu, S.; Jiang, B. Resveratrol is cytotoxic and acts synergistically with NF-κB inhibition in osteosarcoma MG-63 cells. Arch. Med. Sci. 2020, 17, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Afaq, F.; Adhami, V.M.; Ahmad, N. Prevention of short-term ultraviolet B radiation-mediated damages by resveratrol in SKH-1 hairless mice. Toxicol. Appl. Pharmacol. 2003, 186, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Synowiec-Wojtarowicz, A.; Krawczyk, A.; Kimsa-Dudek, M. The effect of resveratrol and static magnetic field interactions on the oxidation–reduction parameters of melanoma malignant cells. Appl. Sci. 2023, 13, 8042. [Google Scholar] [CrossRef]
- Kim, A.L.; Zhu, Y.; Zhu, H.; Han, L.; Kopelovich, L.; Bickers, D.R.; Athar, M. Resveratrol inhibits proliferation of human epidermoid carcinoma A431 cells by modulating MEK1 and AP-1 signalling pathways. Exp. Dermatol. 2006, 15, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Osmond, G.W.; Augustine, C.K.; Zipfel, P.A.; Padussis, J.; Tyler, D.S. Enhancing melanoma treatment with resveratrol. J. Surg. Res. 2012, 172, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Liu, B.; E, C.; Liu, J.; Zhang, Q.; Liu, J.; Chen, N.; Chen, R.; Zhu, R. Resveratrol inhibits the proliferation of human melanoma cells by inducing G1/S cell cycle arrest and apoptosis. Mol. Med. Rep. 2015, 11, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.D.; Cao, Y.; Wang, K.F.; Xu, S.F.; Han, R. [Chemopreventive effect of resveratrol to cancer]. Ai Zheng 2004, 23, 869–873. [Google Scholar] [PubMed]
- Kim, S.E.; Shin, S.H.; Lee, J.Y.; Kim, C.H.; Chung, I.K.; Kang, H.M.; Park, H.R.; Park, B.S.; Kim, I.R. Resveratrol induces mitochondrial apoptosis and inhibits epithelial–mesenchymal transition in oral squamous cell carcinoma cells. Nutr. Cancer 2018, 70, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Yang, G.; Xiang, W.; Pei-Jun, W.; Bin, Z. Effects of resveratrol on oral squamous cell carcinoma (OSCC) cells in vitro. J. Cancer Res. Clin. Oncol. 2014, 140, 371–374. [Google Scholar] [CrossRef] [PubMed]
- ElAttar, T.M.; Virji, A.S. Modulating effect of resveratrol and quercetin on oral cancer cell growth and proliferation. Anticancer Drugs 1999, 10, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Pilankar, A.; Singhavi, H.; Raghuram, G.V.; Siddiqui, S.; Khare, N.K.; Jadhav, V.; Tandel, H.; Pal, K.; Bhattacharjee, A.; Chaturvedi, P.; et al. A pro-oxidant combination of resveratrol and copper down-regulates hallmarks of cancer and immune checkpoints in patients with advanced oral cancer: Results of an exploratory study (RESCU 004). Front. Oncol. 2022, 12, 1000957. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Cheng, J.; Jiang, S.; Wen, J.; Jian, Y.; Wei, L.; Zhe, Z.; Fu-Qiang, J.; Peng, X. The antitumor effect of resveratrol on nasopharyngeal carcinoma cells. Front. Biosci. (Landmark Ed.) 2019, 24, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Zhao, L.; Qin, Y.; Zhang, M.; He, Y. Resveratrol inhibits proliferation and induces apoptosis of nasopharyngeal carcinoma cell line C666-1 through AMPK activation. Pharmazie 2015, 70, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.R.; O’Brian, C.A. Resveratrol antagonizes EGFR-dependent Erk1/2 activation in human androgen-independent prostate cancer cells with associated isozyme-selective PKC alpha inhibition. Invest. New Drugs 2004, 22, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Dhar, S.; Hicks, C.; Levenson, A.S. Resveratrol and prostate cancer: Promising role for microRNAs. Mol. Nutr. Food Res. 2011, 55, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Sheth, S.; Jajoo, S.; Kaur, T.; Mukherjea, D.; Sheehan, K.; Rybak, L.P.; Ramkumar, V. Resveratrol reduces prostate cancer growth and metastasis by inhibiting the Akt/MicroRNA-21 pathway. PLoS One 2012, 7, e51655. [Google Scholar] [CrossRef] [PubMed]
- Nassir, A.M.; Shahzad, N.; Ibrahim, I.A.A.; Ahmad, I.; Md, S.; Ain, M.R. Resveratrol-loaded PLGA nanoparticles mediated programmed cell death in prostate cancer cells. Saudi Pharm. J. 2018, 26, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.N.; Upadhyay, P.K.; Dewangan, H.K. Development, evaluation, pharmacokinetic and biodistribution estimation of resveratrol loaded solid lipid nanoparticles for prostate cancer targeting. J. Microencapsul. 2022, 39, 563–574. [Google Scholar] [CrossRef] [PubMed]
- El-Benhawy, S.A.; Morsi, M.I.; Fahmy, E.I.; Soula, M.A.; Khalil, F.A.Z.F.; Arab, A.R. Role of resveratrol as radiosensitizer by targeting cancer stem cells in radioresistant prostate cancer cells (PC-3). Asian Pac. J. Cancer Prev. 2021, 22, 3823–3837. [Google Scholar] [CrossRef] [PubMed]
- Han, D.S.; Lee, H.J.; Lee, E.O. Resveratrol suppresses serum-induced vasculogenic mimicry through impairing the EphA2/twist-VE-cadherin/AKT pathway in human prostate cancer PC-3 cells. Sci. Rep. 2022, 12, 20125. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Z.; Li, H.; Cao, J.; Nan, N.; Zhai, X.; Liu, Y.; Chong, T. Resveratrol inhibits TRAF6/PTCH/SMO signal and regulates prostate cancer progression. Cytotechnology 2022, 74, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Yadav, E.; Yadav, P.; Khan, M.M.U.; Singh, H.; Verma, A. Resveratrol: A potential therapeutic natural polyphenol for neurodegenerative diseases associated with mitochondrial dysfunction. Front. Pharmacol. 2022, 13, 922232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, H.; Fu, H. Protective effect of resveratrol combined with levodopa against oxidative damage in dopaminergic neurons. Cell Biochem. Biophys.2024, Apr 15. [CrossRef] [PubMed]
- Shamsher, E.; Khan, R.S.; Davis, B.M.; Dine, K.; Luong, V.; Cordeiro, M.F.; Shindler, K.S. Intranasal resveratrol nanoparticles enhance neuroprotection in a model of multiple sclerosis. Int. J. Mol. Sci. 2024, 25, 4047. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.A.Q.; Silva, J.P.B.; Romeiro, C.F.R.; Dos Santos, S.M.; Rodrigues, C.A.; Gonçalves, P.R.; Sakai, J.T.; Mendes, P.F.S.; Varela, E.L.P.; Monteiro, M.C. Neuroprotective mechanisms of resveratrol in Alzheimer’s disease: Role of SIRT1. Oxid. Med. Cell. Longev. 2018, 2018, 8152373. [Google Scholar] [CrossRef] [PubMed]
- Bartra, C.; Yuan, Y.; Vuraić, K.; Valdés-Quiroz, H.; Garcia-Baucells, P.; Slevin, M.; Pastorello, Y.; Suñol, C.; Sanfeliu, C. Resveratrol activates antioxidant protective mechanisms in cellular models of Alzheimer’s disease inflammation. Antioxidants (Basel) 2024, 13, 177. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.D.; Chen, X.X.; Yang, L.J.; Gao, X.R.; Xia, Q.R.; Qi, C.C.; Ge, J.F. Resveratrol ameliorates learning and memory impairments induced by bilateral hippocampal injection of streptozotocin in mice. Neurochem. Int. 2022, 159, 105385. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Yeo, M.A.; Kim, S.J.; Moon, H.I. Neuroprotective effects of resveratrol derivatives from the roots of Vitis thunbergii var. sinuata against glutamate-induced neurotoxicity in primary cultured rat cortical cells. Hum. Exp. Toxicol. 2011, 30, 1404–1408. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Gupta, L.K.; Mediratta, P.K.; Bhattacharya, S.K. Effect of resveratrol on scopolamine-induced cognitive impairment in mice. Pharmacol. Rep. 2012, 64, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhao, X.Y.; Meng, Q.P.; Teng, D.; Deng, K.; Lin, N. Resveratrol activates the SIRT1/PGC-1 pathway in mice to improve synaptic-related cognitive impairment after TBI. Brain Res. 2022, 1796, 148109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, X.B.; Zhao, J.C.; Gao, X.F.; Zhang, X.N.; Hou, K. Neuroprotective effect of resveratrol against radiation after surgically induced brain injury by reducing oxidative stress, inflammation, and apoptosis through NRf2/HO-1/NF-κB signaling pathway. J. Biochem. Mol. Toxicol. 2020, 34, e22600. [Google Scholar] [CrossRef] [PubMed]
- Reda, D.; Elshopakey, G.E.; Mahgoub, H.A.; Risha, E.F.; Khan, A.A.; Rajab, B.S.; El-Boshy, M.E.; Abdelhamid, F.M. Effects of resveratrol against induced metabolic syndrome in rats: Role of oxidative stress, inflammation, and insulin resistance. Evid. Based Complement. Alternat. Med. 2022, 2022, 3362005. [Google Scholar] [CrossRef] [PubMed]
- Ciddi, V.; Dodda, D. Therapeutic potential of resveratrol in diabetic complications: In vitro and in vivo studies. Pharmacol. Rep. 2014, 66, 799–803. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.K.; Kang, H.S. Anti-diabetic effect of cotreatment with quercetin and resveratrol in streptozotocin-induced diabetic rats. Biomol. Ther. (Seoul) 2018, 26, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.J.; Rimando, A.M.; Mizuno, C.S.; Mathews, S.T. α-Glucosidase inhibitory effect of resveratrol and piceatannol. J. Nutr. Biochem. 2017, 47, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Liao, W.; Xia, H.; Wang, S.; Sun, G. The effect of resveratrol on blood lipid profile: A dose-response meta-analysis of randomized controlled trials. Nutrients 2022, 14, 3755. [Google Scholar] [CrossRef] [PubMed]
- Su, C.H.; Wang, H.L.; Tsai, M.L.; Lin, Y.C.; Liao, J.M.; Yen, C.C.; Ting, H.C.; Yu, C.H. Protective effect of microorganism biotransformation-produced resveratrol on the high fat diet-induced hyperlipidemia, hepatic steatosis and synaptic impairment in hamsters. Int. J. Med. Sci. 2022, 19, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Liu, J.L.; Du, A.L. Cardioprotective effect of resveratrol on atherogenic diet-fed rats. Int. J. Clin. Exp. Pathol. 2014, 7, 7899–7906. [Google Scholar] [PubMed]
- Ohara, K.; Kusano, K.; Kitao, S.; Yanai, T.; Takata, R.; Kanauchi, O. ε-Viniferin, a resveratrol dimer, prevents diet-induced obesity in mice. Biochem. Biophys. Res. Commun. 2015, 468, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.W.; Nakamoto, Y.; Hisatome, T.; Yoshida, S.; Miyazaki, H. Resveratrol and its dimers ε-viniferin and δ-viniferin in red wine protect vascular endothelial cells by a similar mechanism with different potency and efficacy. Kaohsiung J. Med. Sci. 2020, 36, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Zhou, Y.; Xie, X. Resveratrol inhibiting TGF/ERK signaling pathway can improve atherosclerosis: Backgrounds, mechanisms and effects. Biomed. Pharmacother. 2022, 155, 113775. [Google Scholar] [CrossRef] [PubMed]
- Banaszewska, B.; Wrotyńska-Barczyńska, J.; Spaczynski, R.Z.; Pawelczyk, L.; Duleba, A.J. Effects of resveratrol on polycystic ovary syndrome: A double-blind, randomized, placebo-controlled trial. J. Clin. Endocrinol. Metab. 2016, 101, 4322–4328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhuang, L.; Gai, S.; Shan, Y.; Wang, S.; Li, F.; Chen, L.; Zhao, D.; Liu, X. Beneficial phytoestrogenic effects of resveratrol on polycystic ovary syndrome in rat model. Gynecol. Endocrinol. 2021, 37, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Malvasi, A.; Tinelli, A.; Lupica, G.; Vimercati, A.; Spyropoulou, K.; Dellino, M.; Mynbaev, O. Effects of a combination of resveratrol and alpha-lipoic acid on body weight and adipose composition in women with PCOS: A preliminary pilot study. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 6578–6582. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Shah, M.; Malik, M.O.; Ehtesham, E.; Habib, S.H.; Rauf, B. Treatment with combined resveratrol and myoinositol ameliorates endocrine, metabolic alterations and perceived stress response in women with PCOS: A double-blind randomized clinical trial. Endocrine 2023, 79, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Huo, P.; Li, M.; Le, J.; Zhu, C.; Yao, J.; Zhang, S. Resveratrol improves follicular development of PCOS rats via regulating glycolysis pathway and targeting SIRT1. Syst. Biol. Reprod. Med. 2023, 69, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, C.X.; Liu, Y.M.; Chen, K.L.; Chen, G. A comparative study of anti-aging properties and mechanism: Resveratrol and caloric restriction. Oncotarget 2017, 8, 65717–65729. [Google Scholar] [CrossRef] [PubMed]
- Leis, K.; Pisanko, K.; Jundziłł, A.; Mazur, E.; Mêcińska-Jundziłł, K.; Witmanowski, H. Resveratrol as a factor preventing skin aging and affecting its regeneration. Postepy Dermatol. Alergol. 2022, 39, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Totonchi, H.; Mokarram, P.; Karima, S.; Rezaei, R.; Dastghaib, S.; Koohpeyma, F.; Noori, S.; Azarpira, N. Resveratrol promotes liver cell survival in mice liver-induced ischemia–reperfusion through unfolded protein response: A possible approach in liver transplantation. BMC Pharmacol. Toxicol. 2022, 23, 74. [Google Scholar] [CrossRef] [PubMed]
- Theodotou, M.; Fokianos, K.; Moniatis, D.; Kadlenic, R.; Chrysikou, A.; Aristotelous, A.; Mouzouridou, A.; Diakides, J.; Stavrou, E. Effect of resveratrol on non-alcoholic fatty liver disease. Exp. Ther. Med. 2019, 18, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Tejada, S.; Capó, X.; Mascaró, C.M.; Monserrat-Mesquida, M.; Quetglas-Llabrés, M.M.; Pons, A.; Tur, J.A.; Sureda, A. Hepatoprotective effects of resveratrol in non-alcoholic fatty live disease. Curr. Pharm. Des. 2021, 27, 2558–2570. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Yang, L.; Zhong, W.; Hu, Y.; Tan, Y.; Ren, Z.; Ban, Q.; Yang, C.S.; Wang, Y.; Wang, Z. Polydatin, a glycoside of resveratrol, is better than resveratrol in alleviating non-alcoholic fatty liver disease in mice fed a high-fructose diet. Front. Nutr. 2022, 9, 857879. [Google Scholar] [CrossRef] [PubMed]
- Chupradit, S.; Bokov, D.; Zamanian, M.Y.; Heidari, M.; Hakimizadeh, E. Hepatoprotective and therapeutic effects of resveratrol: A focus on anti-inflammatory and antioxidative activities. Fundam. Clin. Pharmacol. 2022, 36, 468–485. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; Shaw, L.H.; Chang, P.J.; Tung, S.Y.; Chang, T.S.; Shen, C.H.; Hsieh, Y.Y.; Wei, K.L. Hepatoprotective effect of resveratrol against ethanol-induced oxidative stress through induction of superoxide dismutase in vivo and in vitro. Exp. Ther. Med. 2016, 11, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Gokkaya, E.O.; Yesilot, S.; Ozgocmen, M.; Aslankoc, R.; Aydin Acar, C. Protective effects of resveratrol and avocado oil against paracetamol-induced hepatotoxicity in rats. Drug Chem. Toxicol. 2022, 45, 2131–2139. [Google Scholar] [CrossRef] [PubMed]
- Pace-Asciak, C.R.; Hahn, S.; Diamandis, E.P.; Soleas, G.; Goldberg, D.M. The red wine phenolics trans-resveratrol and quercetin block human platelet aggregation and eicosanoid synthesis: Implications for protection against coronary heart disease. Clin. Chim. Acta 1995, 235, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Duan, Q.J.; Shen, J. [Resveratrol pretreatment improves mitochondrial function and alleviates myocardial ischemia–reperfusion injury by up-regulating mi R-20b-5p to inhibit STIM2]. Zhongguo Zhong Yao Za Zhi 2022, 47, 4987–4995. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Liu, C.; Zhang, Z.; Huang, K.; Wang, T.; Chen, S.; Li, Z. Progress in the preclinical and clinical study of resveratrol for vascular metabolic disease. Molecules 2022, 27, 7524. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yang, X.; Liao, Y.; Wang, C.; Wang, Y. Resveratrol alleviates Ang II-induced vascular smooth muscle cell senescence by upregulating E2F1/SOD2 axis. Toxicol. Res. (Camb.) 2022, 11, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Militaru, C.; Donoiu, I.; Craciun, A.; Scorei, I.D.; Bulearca, A.M.; Scorei, R.I. Oral resveratrol and calcium fructoborate supplementation in subjects with stable angina pectoris: Effects on lipid profiles, inflammation markers, and quality of life. Nutrition 2013, 29, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Baolin, L.; Inami, Y.; Tanaka, H.; Inagaki, N.; Iinuma, M.; Nagai, H. Resveratrol inhibits the release of mediators from bone marrow-derived mouse mast cells in vitro. Planta Med. 2004, 70, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wu, S.L.; Zhang, M.; Pan, C.E. [Effect of resveratrol alone and its combination with cyclosporin A on the immune function of human peripheral blood T lymphocytes]. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2003, 19, 549–551. [Google Scholar] [PubMed]
- Ferrero, M.E.; Bertelli, A.E.; Fulgenzi, A.; Pellegatta, F.; Corsi, M.M.; Bonfrate, M.; Ferrara, F.; De Caterina, R.; Giovannini, L.; Bertelli, A. Activity in vitro of resveratrol on granulocyte and monocyte adhesion to endothelium. Am. J. Clin. Nutr. 1998, 68, 1208–1214. [Google Scholar] [CrossRef] [PubMed]
- Kasdallah-Grissa, A.; Mornagui, B.; Aouani, E.; Hammami, M.; Gharbi, N.; Kamoun, A.; El-Fazaa, S. Protective effect of resveratrol on ethanol-induced lipid peroxidation in rats. Alcohol Alcohol. 2006, 41, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Chanvitayapongs, S.; Draczynska-Lusiak, B.; Sun, A.Y. Amelioration of oxidative stress by antioxidants and resveratrol in PC12 cells. Neuroreport 1997, 8, 1499–1502. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.M.; Li, S.Y.; Huang, Q.; Zeng, F.F.; Li, B.L.; Cao, W.T.; Chen, Y.M. Greater habitual resveratrol intakes were associated with lower risk of hip fracture – a 1:1 matched case-control study in Chinese elderly. Phytother. Res. 2023, 37, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Bilgic, T. Comparison of the effect of local and systemic injection of resveratrol on cutaneous wound healing in rats. Int. J. Low. Extrem. Wounds 2021, 20, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Yoksa, D.T.; Abba, Y.; Shamaki, B.U.; Satumari, N.A. Effects of resveratrol topical ointment on wound healing of full-thickness cutaneous burns in albino rats. J. Wound Care 2022, 31, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Dong, Y.; Xu, P.; Pan, Q.; Jia, K.; Jin, P.; Zhou, M.; Xu, Y.; Guo, R.; Cheng, B. A composite hydrogel containing resveratrol-laden nanoparticles and platelet-derived extracellular vesicles promotes wound healing in diabetic mice. Acta Biomater. 2022, 154, 212–230. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zheng, B.; Zheng, H.; Xu, G.; Jiang, H. Resveratrol promotes diabetic wound healing by inhibiting Notch pathway. J. Surg. Res. 2024, 297, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.B.; Wang, Y.J.; Wan, W.J.; Wu, J.; Wang, B.J.; Zhu, H.L.; Xie, M.; Liu, L. Resveratrol ameliorates oxaliplatin-induced neuropathic pain via anti-inflammatory effects in rats. Exp. Ther. Med. 2022, 24, 586. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yu, H.; Lin, Q.; Liu, X.; Cheng, Y.; Deng, B. Anti-inflammatory effect of resveratrol attenuates the severity of diabetic neuropathy by activating the Nrf2 pathway. Aging (Albany N.Y.) 2021, 13, 10659–10671. [Google Scholar] [CrossRef] [PubMed]
- Ashafaq, M.; Intakhab Alam, M.; Khan, A.; Islam, F.; Khuwaja, G.; Hussain, S.; Ali, R.; Alshahrani, S.; Antar Makeen, H.; Alhazmi, H.A.; et al. Nanoparticles of resveratrol attenuates oxidative stress and inflammation after ischemic stroke in rats. Int. Immunopharmacol. 2021, 94, 107494. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, E.; Tanaka, D.; Glogauer, M.; Tenenbaum, H.C.; Ikeda, Y. Healing effects of monomer and dimer resveratrol in a mouse periodontitis model. BMC Oral Health 2022, 22, 460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xu, S.; Xu, W.; Zhou, Y.; Luan, H.; Wang, D. Resveratrol decreases local inflammatory markers and systemic endotoxin in patients with aggressive periodontitis. Medicine (Baltimore) 2022, 101, e29393. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, M.; Ingmer, H. Antibacterial and antifungal properties of resveratrol. Int. J. Antimicrob. Agents 2019, 53, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Okamoto-Shibayama, K.; Yoshida, A.; Ishihara, K. Inhibitory effect of resveratrol on Candida albicans biofilm formation. Bull. Tokyo Dent. Coll. 2021, 62, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Liang, R.; Duan, J.; Song, S.; Pan, Y.; Liu, H.; Zhu, M.; Li, L. Synergistic antibacterial and anti-biofilm activities of resveratrol and polymyxin B against multidrug-resistant Pseudomonas aeruginosa. J. Antibiot. (Tokyo) 2022, 75, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Daroch, F.; Hoeneisen, M.; González, C.L.; Kawaguchi, F.; Salgado, F.; Solar, H.; García, A. In vitro antibacterial activity of Chilean red wines against Helicobacter pylori. Microbios 2001, 104, 79–85. [Google Scholar] [PubMed]
- Spósito, L.; Fonseca, D.; Gonçalves Carvalho, S.; Miguel Sábio, R.; Marena, G.D.; Maria Bauab, T.; Bagliotti Meneguin, A.; Parreira, P.; Martins, M.C.L.; Chorilli, M. Engineering resveratrol-loaded chitosan nanoparticles for potential use against Helicobacter pylori infection. Eur. J. Pharm. Biopharm. 2024, 199, 114280. [Google Scholar] [CrossRef] [PubMed]
- Heredia, A.; Davis, C.; Redfield, R. Synergistic inhibition of HIV-1 in activated and resting peripheral blood mononuclear cells, monocyte-derived macrophages, and selected drug-resistant isolates with nucleoside analogues combined with a natural product, resveratrol. J. Acquir. Immune Defic. Syndr. 2000, 25, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Docherty, J.J.; Smith, J.S.; Fu, M.M.; Stoner, T.; Booth, T. Effect of topically applied resveratrol on cutaneous herpes simplex virus infections in hairless mice. Antiviral Res. 2004, 61, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Evers, D.L.; Wang, X.; Huong, S.M.; Huang, D.Y.; Huang, E.S. 3,4’,5-Trihydroxy-trans-stilbene (resveratrol) inhibits human cytomegalovirus replication and virus-induced cellular signaling. Antiviral Res. 2004, 63, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liao, D.; Zhou, G.; Zhu, Z.; Cui, Y.; Pu, R. Antiviral activities of resveratrol against rotavirus in vitro and in vivo. Phytomedicine 2020, 77, 153230. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Li, J.; Lin, W.; Long, G. Effects of resveratrol on hepatitis B virus replication: In vitro and in vivo experiments. Intervirology 2022, 65, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Vlase, L.; Kiss, B.; Leucuta, S.E.; Gocan, S. A rapid method for determination of resveratrol in wines by HPLC-MS. J. Liq. Chromatogr. Relat. Technol. 2009, 32, 2105–2121. [Google Scholar] [CrossRef]
- Svilar, L.; Martin, J.C.; Defoort, C.; Paut, C.; Tourniaire, F.; Brochot, A. Quantification of trans-resveratrol and its metabolites in human plasma using ultra-high performance liquid chromatography tandem quadrupole–orbitrap mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2019, 1104, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yuan, H.; Liu, Y.; Wang, B.; Xu, X.; Xu, X.; Hussain, D.; Ma, L.; Chen, D. Current analytical strategies for the determination of resveratrol in foods. Food Chem. 2024, 431, 137182. [Google Scholar] [CrossRef] [PubMed]
- Crăciun, A.L.; Gutt, G. Optimization of experimental parameters in the solvent extraction of trans-resveratrol from pruning waste of Vitis vinifera, Fetească Neagră variety. Appl. Sci. 2023, 13, 823. [Google Scholar] [CrossRef]
Figure 1.
Calibration curve of trans-resveratrol displaying the detector response across a range of standard concentrations.
Figure 1.
Calibration curve of trans-resveratrol displaying the detector response across a range of standard concentrations.
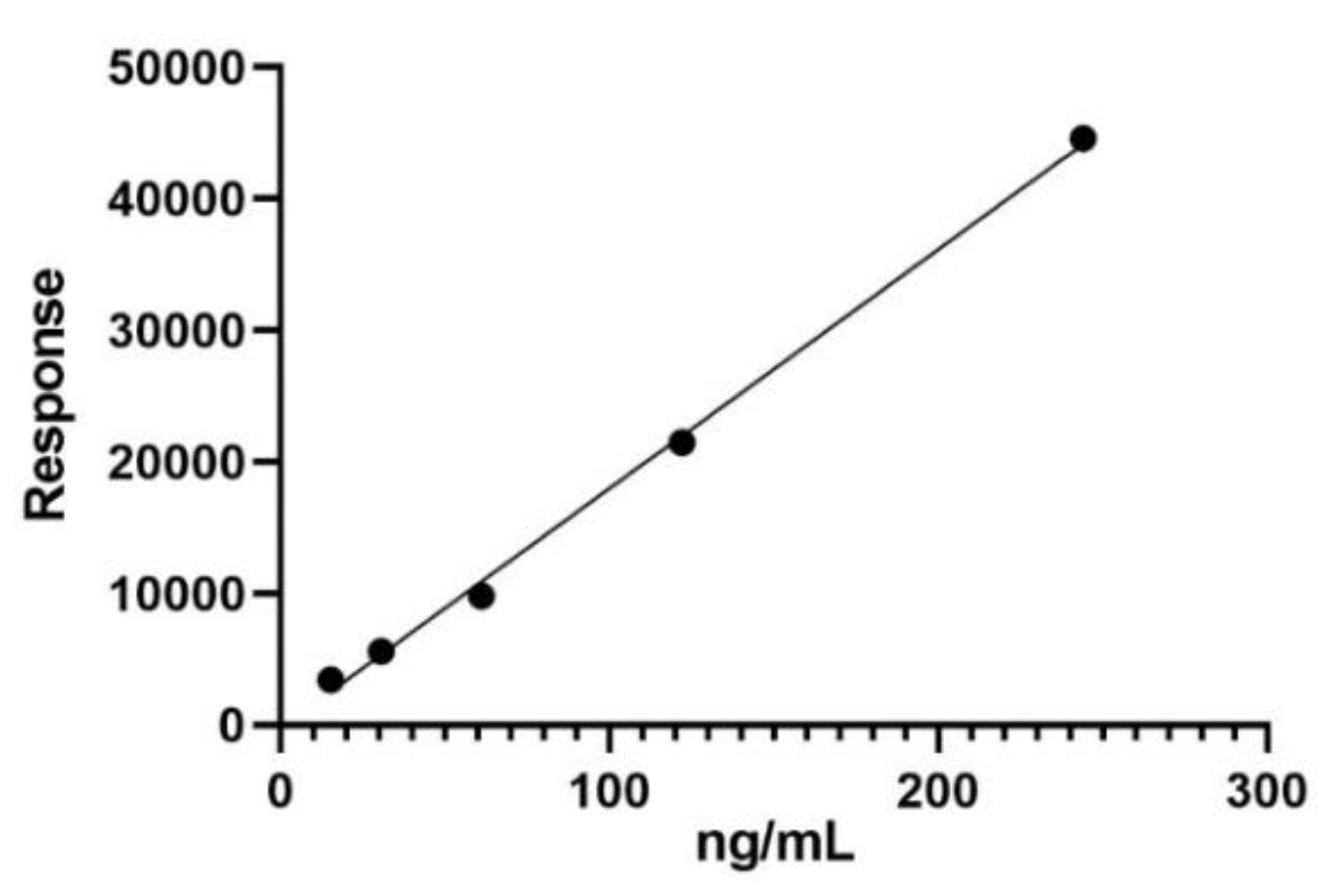
Figure 2.
Chromatogram of a trans-resveratrol standard showing a prominent peak at 9.67 minutes, indicative of its purity and concentration in the reference sample.
Figure 2.
Chromatogram of a trans-resveratrol standard showing a prominent peak at 9.67 minutes, indicative of its purity and concentration in the reference sample.
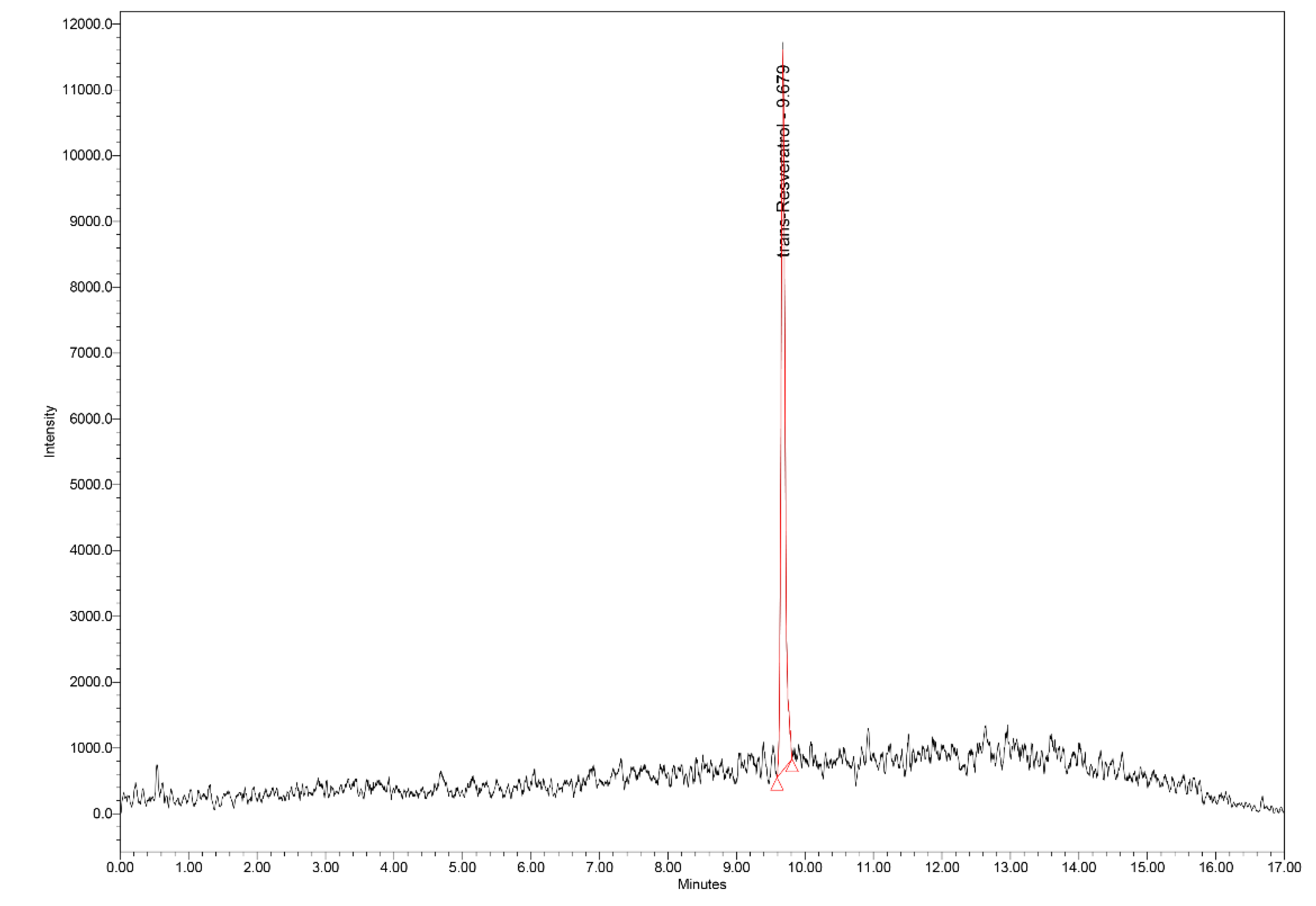
Figure 3.
Mass spectrum of trans-resveratrol illustrating the quantification of molecular ion peak at m/z 227, used for both confirmation and quantification of the analyte.
Figure 3.
Mass spectrum of trans-resveratrol illustrating the quantification of molecular ion peak at m/z 227, used for both confirmation and quantification of the analyte.
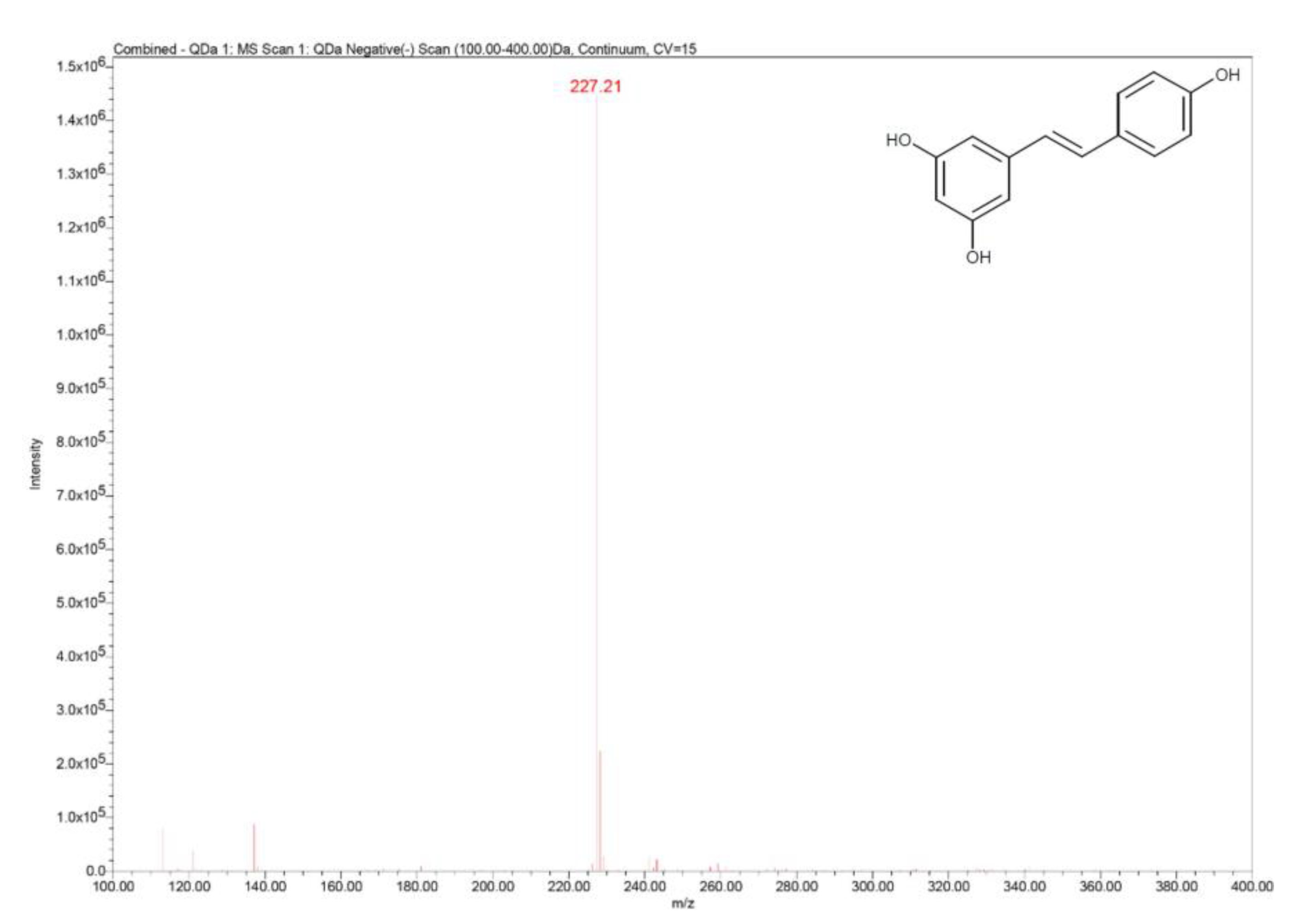
Figure 4.
Overlay of UHPLC chromatograms for Romanian wine samples, with the trans-resveratrol peak identified at the specific retention time across all samples. UHPLC: Ultra-high-performance liquid chromatography; W1: Fetească Regală; W2: Fetească Neagră; W3: Dry Muscat; W4: Cabernet Sauvignon.
Figure 4.
Overlay of UHPLC chromatograms for Romanian wine samples, with the trans-resveratrol peak identified at the specific retention time across all samples. UHPLC: Ultra-high-performance liquid chromatography; W1: Fetească Regală; W2: Fetească Neagră; W3: Dry Muscat; W4: Cabernet Sauvignon.
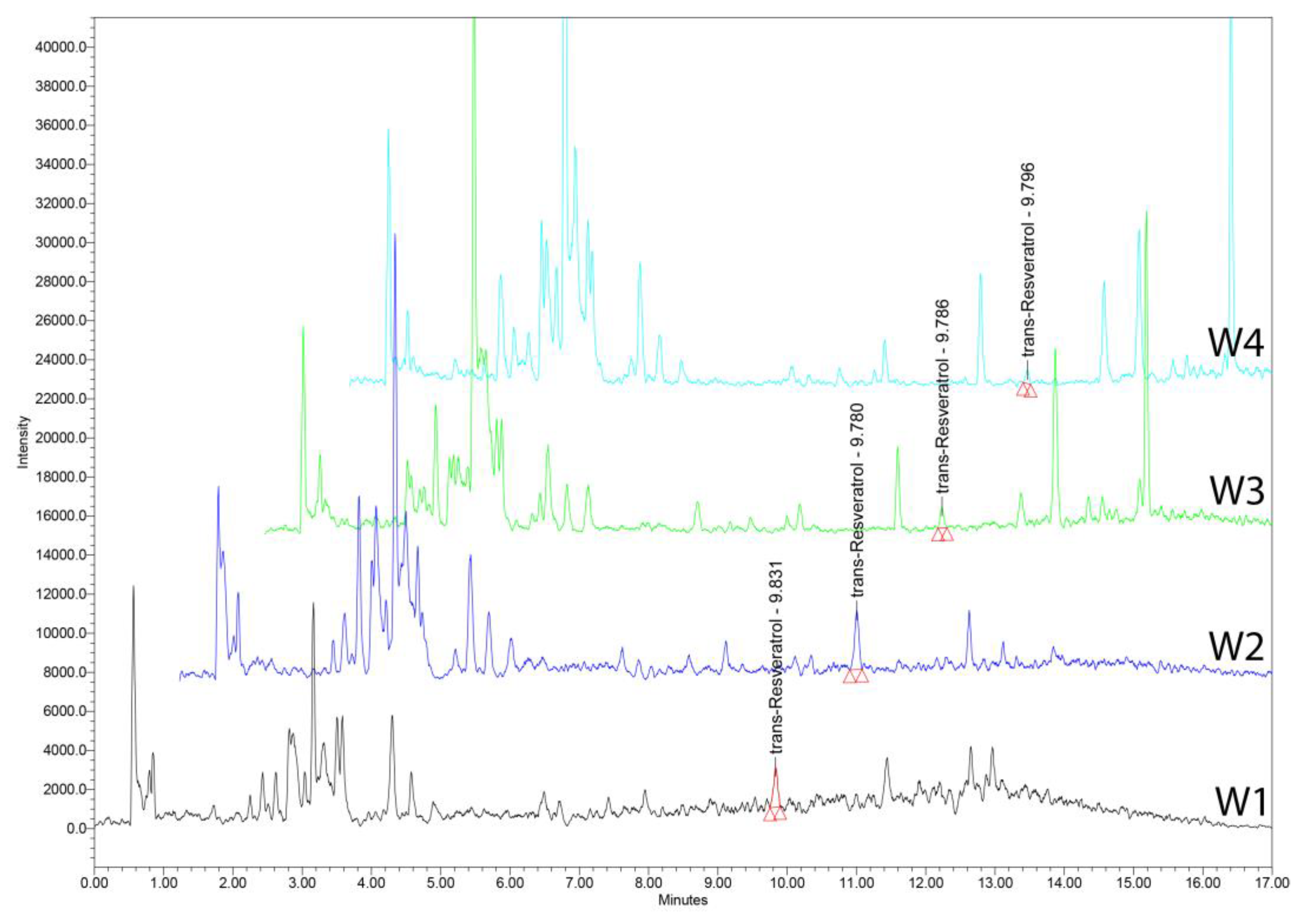

Table 1.
Summary of calibration curve parameters for trans-resveratrol quantification.
| Parameter | Values |
|---|---|
| Linearity range [ng/mL] | 15.259–244.141 |
| R2 | 0.9979 |
| Equation | Y = 181.7X - 197.6 |
| LOD [ng/mL] | 16.197 |
| LOQ [ng/mL] | 49.081 |
LOD: Limit of detection; LOQ: Limit of quantification; R2: Correlation coefficient.
Table 2.
trans-Resveratrol amounts detected in Romanian wine samples.
| Sample | trans-Resveratrol amount [ng/mL] |
|---|---|
| W1 | 243.291±2.51 |
| W2 | 372.042±2.75 |
| W3 | 108.487±1.98 |
| W4 | 69.166±1.05 |
White wine samples: W1: Fetească Regală; W3: Dry Muscat. Red wine samples: W2: Fetească Neagră; W4: Cabernet Sauvignon.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



