
Submitted:
25 April 2024
Posted:
01 May 2024
You are already at the latest version
Abstract
The study evaluated the regenerative responses of the lacrimal functional unit (LFU) after Lacrimal Gland (LG) ablation. The LG of Wistar rats were submitted to G1) partial LG ablation, G2) partial ablation and transplantation of an allogeneic LG, or G3) total LG ablation, (n=7-10/group). The eye wipe test, slit-lamp image, tear flow, and histology were evaluated. RT-PCR analyzed inflammatory and proliferation mediators. The findings were compared to naïve controls after 1 and 2 months (M1 and M2). G3 presented increased corneal sensitivity, and the 03 groups have shown corneal neovascularization. Histology revealed changes in LG, and corneal inflammation. In the LG, there was an increase in MMP-9 mRNA of G1 and G2 at M1 and M2 and RUNX-1 at M1 and M2 in G1 and RUNX-3 mRNA at M1 in G1, and in G2 at M2. TNF-α mRNA rose in the corneas of G1 and G2 at M2. There was an increase in the IL-1β mRNA in the Trigeminal Ganglion of G1 at M1. Without changes in tear flow or evidence of LG regeneration, LG ablation or grafting are unreliable models for dry eye or LG repair in rats. The surgical manipulation extended inflammation to the LFU.
Keywords:
inflammation
; lacrimal functional unit
; regeneration
1. Introduction
It is controversial whether the main lacrimal gland (LG) is an indispensable organ for vision and ocular surface integrity [1,2,3]. However, it is clear that the lacrimal functional unit (LFU) responds together to challenges to one or more of its components and that evolutionally closer animals present similar responses to those challenges [2,3,4,5,6].
Damage to the main LG in humans and primates impacts other tissues of the LFU and induces dry eye disease (DED). However, it seems to be attenuated in the long term [1,7]. On the other hand, exorbital LG ablation in rodents has been used as animal models in short-term studies to evaluate the efficacy of novel therapies for dry eye disease [8,9].
The potential of mammalian LG auto or therapeutic-driven repair is the subject of many doubts and may be influenced by different mechanisms of damage and time-length of observation [10,11].
LG dysfunction can arise from different diseases or local injuries [4,12]. However, tissue ablation by surgery or radiotherapy may also elicit declines in LG support of the lacrimal functional unit (LFU) that deserves a deeper understanding [5,13,14,15].
There is evidence that autologous salivary gland or neonatal LG transplantation can restore sufficient LG structure and function to be of therapeutic value in dry eye models or patients [16,17,18,19]. However, it is unknown whether or not increases in LG secretion can be attributed to proliferative or repair events or it is just exudation related to inflammatory healing as a response to the surgical procedure, and whether this secretory activity persists for extended periods of time [8,20,21,22].
LG impact on the LFU and its repair capacity is specie specific and modulated by pro mitotic factors [6,10]. Removal, recession, extraction, excision, and ablation are similar terms used in medical science literature when referring to surgical extirpation of an organ. Here, we adopted the word ablation to describe the proposed partial or complete manipulation of the LG.
In humans, main LG agenesia, atrophy, or ablation induces persistent dry eye syndrome (DES) [1,23,24,25,26]. LG In monkeys, the complete ablation of the main LG led to a reduction in tears flow and ocular surface staining. The findings gradually reverted along 20 weeks, probably supported by the accessory glands [13]. In rats, total LG ablation is used to establish a dry eye syndrome model for searching for novel therapeutic options, pharmacological and surgical, based on the belief that it is an irreversible and reproducible model [8,27]. Recently, the extra orbital LG total ablation mice model was revealed to be more aggressive to the ocular surface than drug-induced cholinergic blockage combined with a dry environment [28]. Rabbit models are not investigated for the LG regeneration model as much as rats and mice; however, the larger organs offer an exciting model for surgical and grafting studies, where the evaporative and tear deficiency repercussions can be accessed [29].
In mice LG, injury-induced tissue inflammation is associated with apoptosis and autophagy [30]. Moreover, it changes protein expression and growth factors relevant to LG morphogenesis and regeneration. They include fibroblast growth factor-10 (FGF-10) and bone morphogenetic protein 7 (BMP-7) and signaling proteins such as nestin and Pax6 [30,31,32]. Their presence further substantiates the notion that following the damage. The regeneration would occur in three phases (i.e.,; inflammation, new tissue formation, and remodeling) where LG cells require molecular machinery and local stem cells to undergo proliferation and differentiation [19,33]. A recent review addressed the topic of LG regeneration, which involved several mediators, including BMP-7, and stem cells, mentioning various species but not in rat models [33].
Our hypothesis predicts that LG ablation is irreversible and leads to structural, molecular, and functional repercussions in the LFU. Still, partial LG ablation and grafting will reveal the proliferative agents and structural attempts to regenerate the LG.
We describe here different rat models of LG damage obtained by partial ablation (G1), allogeneic LG transplantation following partial ablation (G2), complete ablation (G3), and control (G4), where the changes induced in LG structure and function, and LFU health were compared. Furthermore, the possible signs of repair were evaluated in response to those interventions.
2. Results
2.1. LG partial Ablation (G1)
2.1.1. In Vivo Exams
After 1 and 2 months, the corneal sensitivity was not changed compared to the control group (Table 1). The slit-lamp exam revealed corneal neovascularization and punctate keratitis (Figure 1, upper and lower lane).
The results of a phenol red thread test mean wetting in mm indicate those tear secretion measurements of G1 were similar to control values along the observational period (Table 1).
Corneal epithelial cell impression cytology did not reveal metaplasia compared to controls at 1 or 2 months (P>0.05, Fisher test) (data not shown).
As expected, LG weight in G1 was significantly lower than controls (Table 1). The results indicate that LG weight after 1 or 2 months remained lower and did not recover the weight of the intact LG.
2.1.2. Ex Vivo Assays
To determine whether or not any of the invasive procedures induced changes in LG proliferation, PCNA expression levels were measured. PCNA expression levels remained similar in the LG of G1, G2, and control groups throughout the two months (data not shown).
LG histology comparing Control and G1 revealed that LG acinar and ductal structures were unchanged, except for mild cytoplasmic vacuolation and atypical nuclei suggestive of attempt mitosis or apoptosis (Figure 3). Corneal, conjunctival, and lid histology was not changed by partial LG ablation in G1 (Figure 4 for corneal histology in month 2).
Figure 2.
Representative histological images of LG of Controls (C), LG partial ablation (G1), and ablation followed by allogeneic LG transplant (G2). Samples were stained with H&E. Magnification 100x for the figures and 400x for inserts that show details of acinar cells structure (right lower corner). Black arrows indicate the border between graft (upper side) and host (lower side) LG.
Figure 2.
Representative histological images of LG of Controls (C), LG partial ablation (G1), and ablation followed by allogeneic LG transplant (G2). Samples were stained with H&E. Magnification 100x for the figures and 400x for inserts that show details of acinar cells structure (right lower corner). Black arrows indicate the border between graft (upper side) and host (lower side) LG.
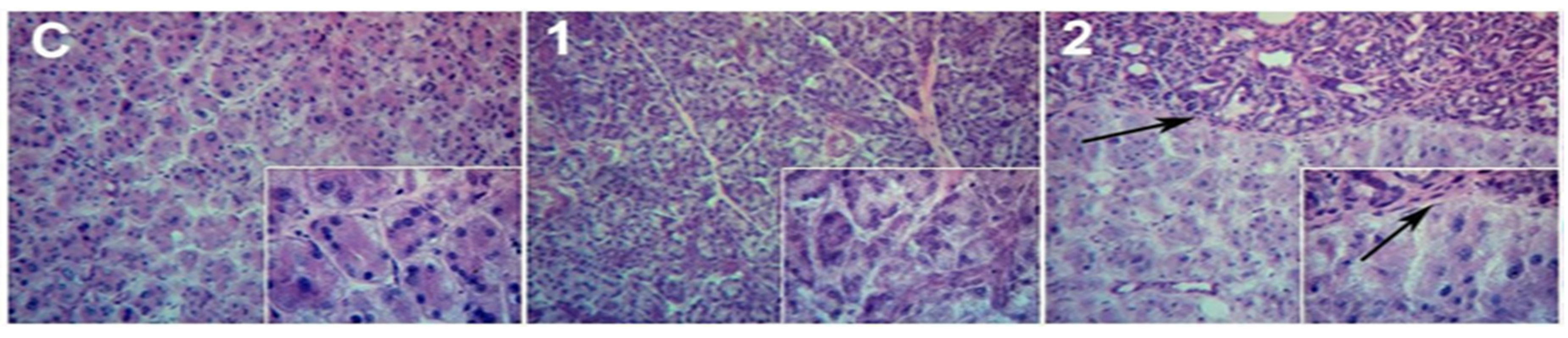
Figure 3.
Representative histological images of LG of Controls (C), LG partial ablation (G1), and ablation followed by allogeneic LG transplant (G2). Samples were stained with H&E. Magnification 100x for the figures and 400x for inserts that show details of acinar cells structure (right lower corner). Black arrows indicate the border between graft (upper side) and host (lower side) LG.
Figure 3.
Representative histological images of LG of Controls (C), LG partial ablation (G1), and ablation followed by allogeneic LG transplant (G2). Samples were stained with H&E. Magnification 100x for the figures and 400x for inserts that show details of acinar cells structure (right lower corner). Black arrows indicate the border between graft (upper side) and host (lower side) LG.
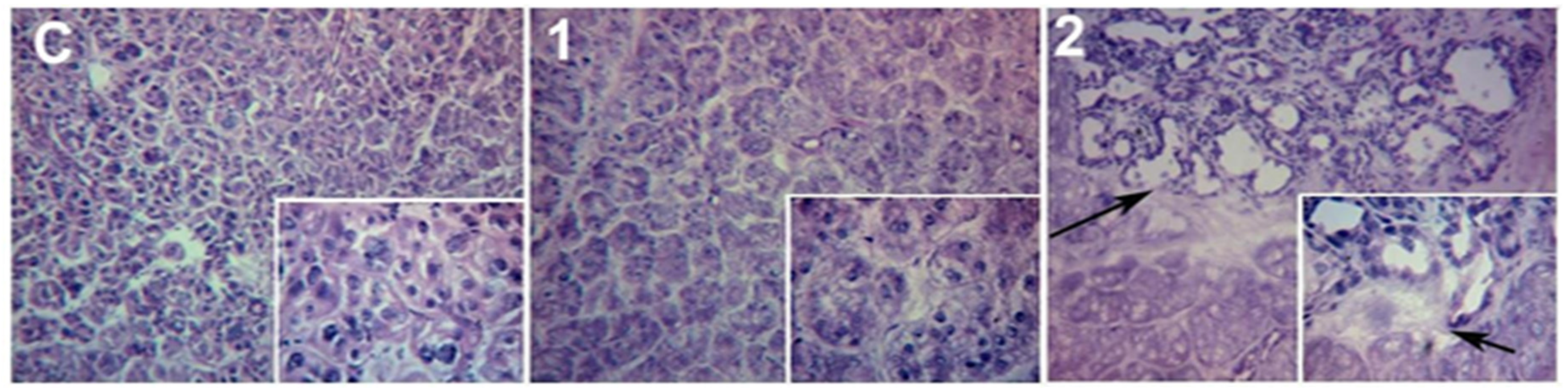
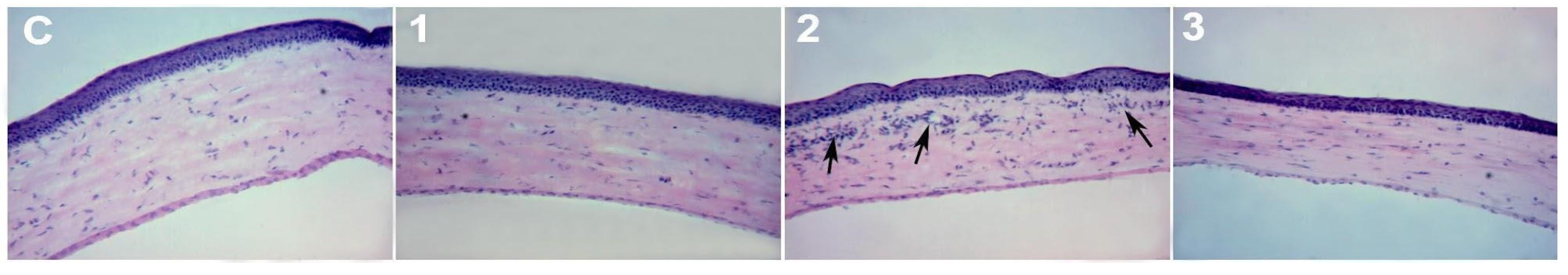
Figure 4.
Representative histological image of the cornea of Controls (C), LG partial ablation (G1), ablation followed by allogeneic LG transplant (G2), and total ablation (G3) at month 2. Samples were stained with H&E (100 x magnification). Black arrows indicate leukocytes and microvessels in the cornea stroma.
Figure 4.
Representative histological image of the cornea of Controls (C), LG partial ablation (G1), ablation followed by allogeneic LG transplant (G2), and total ablation (G3) at month 2. Samples were stained with H&E (100 x magnification). Black arrows indicate leukocytes and microvessels in the cornea stroma.
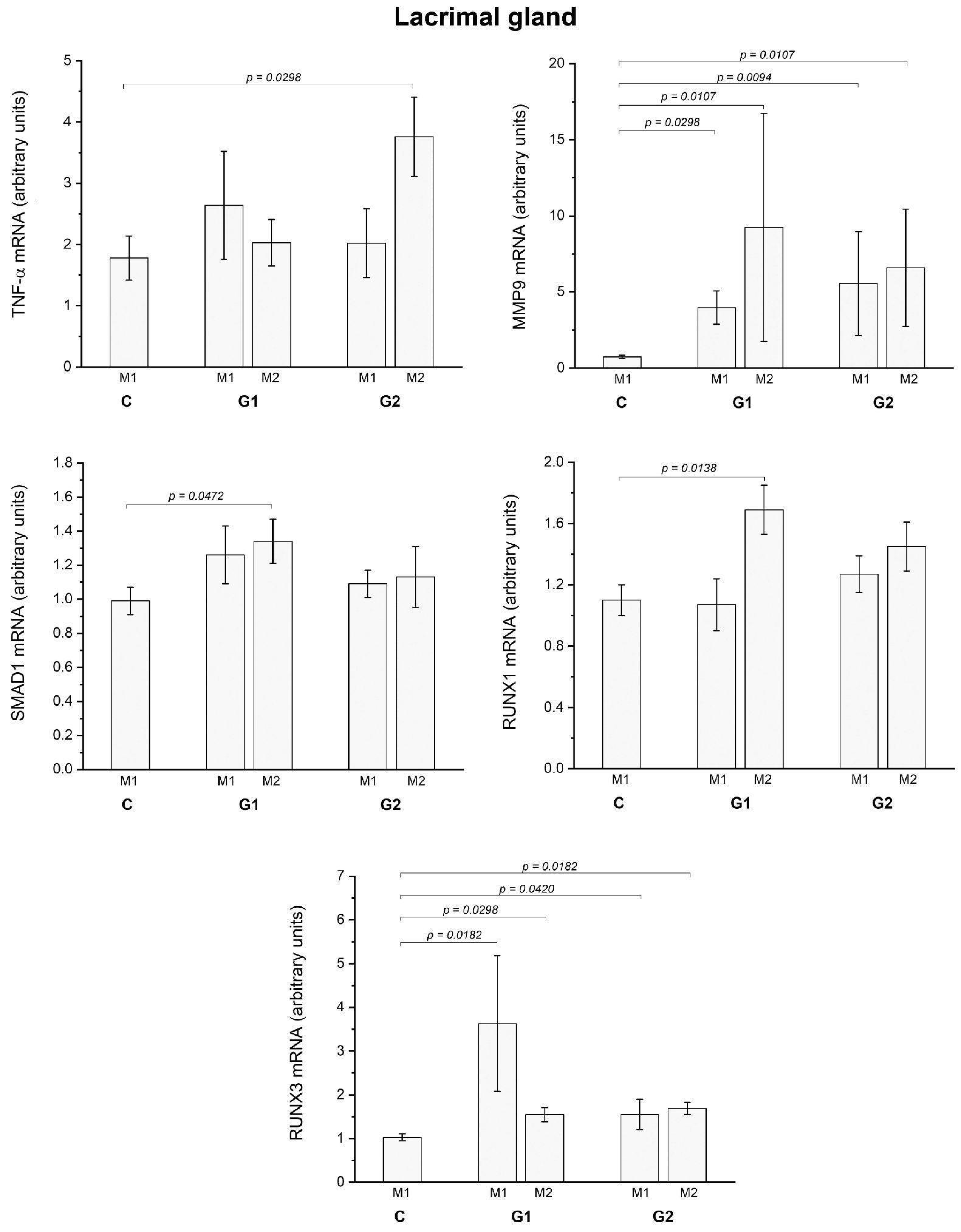
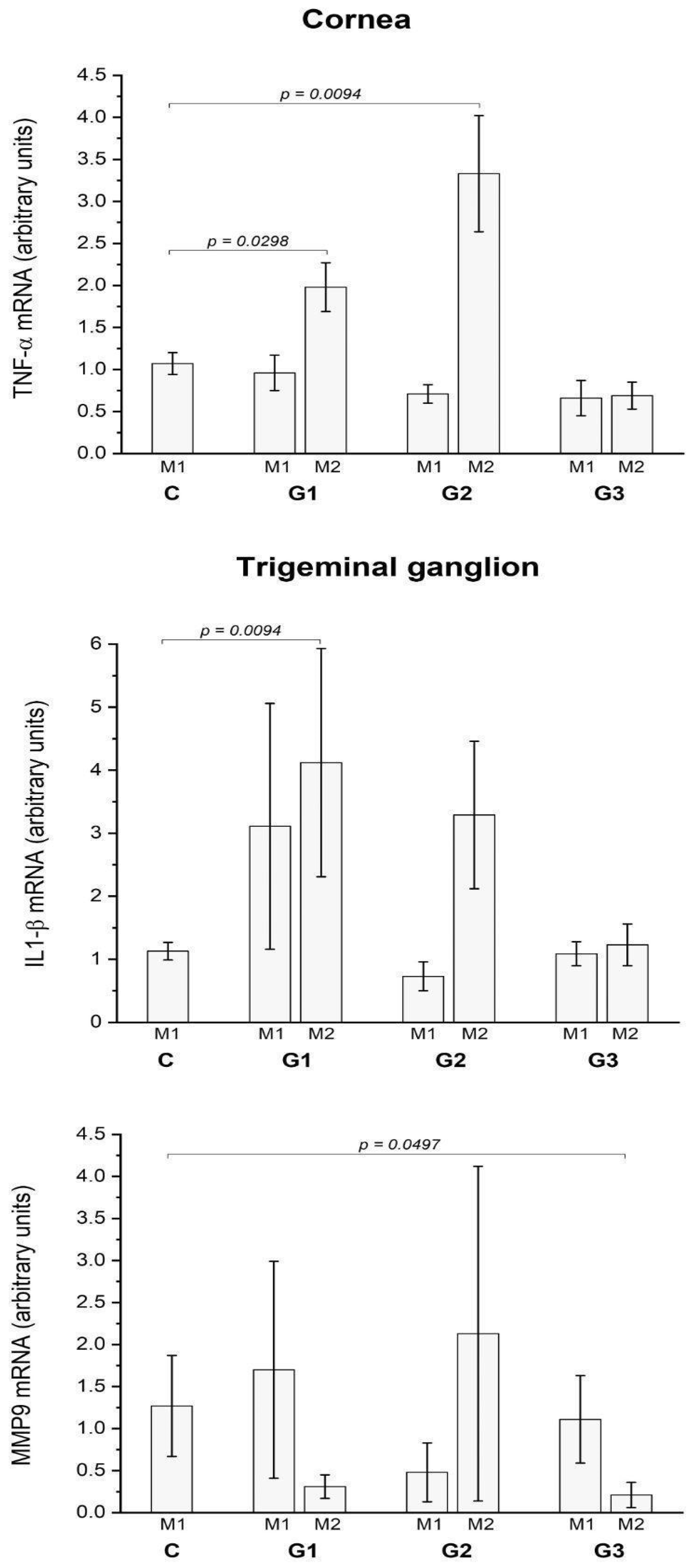
The RT PCR analysis of the mRNA expression of inflammatory mediators in the LG of G1 revealed that MMP-9 mRNA levels raised at months 1 and 2 (P=0.0242 and 0.0061, respectively) (Figure 5A).
The RT-PCR of the mRNA expression of proliferative mediators in LG of G1 revealed that RUNX-1 and RUNX-3 mRNA levels rose at month 1 (P=0.0138 and 0.0118, respectively) and RUNX-1 was also higher in G1 compared to controls at month 2 (P=0.0286) (Figure 5B,C).
Figure 5.
A: Comparative analysis of pro-inflammatory cytokines mRNA of Mmp9 by qPCR in the LG (expressed in arbitrary units, normalized by β-actin mRNA). The G1 (partial ablation) presented higher relative mRNA expression Mmp9 at month 1 (M1) (P=0.0242) and at month 2 (M2) (P=0.0061) compared to controls. The G2 (partial ablation + allogeneic grafting) presented higher relative mRNA expression of Mmp9 at M1 (P=0.0051) and at M2 (0.0061). 5. B: Comparative analysis of the mRNA of the pro mitotic peptide Runx1, in the LG by QRT-PCR (arbitrary units, normalized by β-actin mRNA). G1 showed higher expression of Runx1 at M1 and M2 (P=0.0138 and P=0.00118). 5.C: Comparative analysis of the mRNA of the pro mitotic peptide Runx3, in the LG by QRT-PCR (arbitrary units, normalized by β-actin mRNA) G1 showed higher expression of at M2 (P=0.0149). The statistical analysis applied the Mann-Whitney U test. The number of animals was CG M1: X rats, CG M2: x rats; G1 M1: x rats and G1 M2: x rats, G2 M1: x rats and G2 M2: x rats). The statistical analysis applied the Mann-Whitney U test.
Figure 5.
A: Comparative analysis of pro-inflammatory cytokines mRNA of Mmp9 by qPCR in the LG (expressed in arbitrary units, normalized by β-actin mRNA). The G1 (partial ablation) presented higher relative mRNA expression Mmp9 at month 1 (M1) (P=0.0242) and at month 2 (M2) (P=0.0061) compared to controls. The G2 (partial ablation + allogeneic grafting) presented higher relative mRNA expression of Mmp9 at M1 (P=0.0051) and at M2 (0.0061). 5. B: Comparative analysis of the mRNA of the pro mitotic peptide Runx1, in the LG by QRT-PCR (arbitrary units, normalized by β-actin mRNA). G1 showed higher expression of Runx1 at M1 and M2 (P=0.0138 and P=0.00118). 5.C: Comparative analysis of the mRNA of the pro mitotic peptide Runx3, in the LG by QRT-PCR (arbitrary units, normalized by β-actin mRNA) G1 showed higher expression of at M2 (P=0.0149). The statistical analysis applied the Mann-Whitney U test. The number of animals was CG M1: X rats, CG M2: x rats; G1 M1: x rats and G1 M2: x rats, G2 M1: x rats and G2 M2: x rats). The statistical analysis applied the Mann-Whitney U test.
The RT-PCR assay to compare the expression of inflammatory cytokines mRNA in the cornea and investigate the potential inflammatory repercussions of those interventions on the ocular surface, as indicated by the slit-lamp and histological analysis, revealed that G1 presented higher levels of TNF-α mRNA at month 2 (P=0.0298) (Figure 6A). The mRNA expression of MMP-9, IL-1β, and IL-6 did not significantly change in none of the groups and time points compared to the control group.
In addition, considering that LG intervention increased the corneal sensitivity in G3, we investigated the changes in pro-inflammatory cytokines mRNA expression in TG compared to the control group at 1 and 2 months after the intervention. The mRNA expression of IL-1β rose in TG of G1 at month 2 (P=0.0094) (Figure 6B). The other cytokines mRNA tested (IL-6 and TNF-α) presented no changes in TG compared to the control group at 1 or 2 months.
Figure 6.
A: Comparative analysis of pro-inflammatory cytokine mRNA Tnf by qPCR in the cornea (expressed in arbitrary units, normalized by β-actin mRNA). The G1 and G2 presented higher expression of Tnf at M2 (P=0.0257 and P=0.028, respectively). CG M2: x rats; G1 M1: x rats and G1 M2: x rats, G2 M1: x rats and G2 M2: x rats). The statistical analysis applied the Mann-Whitney U test. 6.B: Comparative analysis of pro-inflammatory cytokine mRNA Il1b by qPCR in the TG (expressed in arbitrary units, normalized by β-actin). G1 presented higher levels of Il1b at M1 (P=0.0286). CG M1: X rats, CG M2: x rats; G1 M1: x rats and G1 M2: x rats, G2 M1: x rats and G2 M2: x rats). The statistical analysis applied the Mann-Whitney U test. 6.C: Comparative analysis of pro-inflammatory cytokine mRNA MMP-9 by qPCR in the TG (expressed in arbitrary units, normalized by β-actin). G1 presented higher levels of Il1b at M1 (P=0.0286). CG M1: X rats, CG M2: x rats; G1 M1: x rats and G1 M2: x rats, G2 M1: x rats and G2 M2: x rats). The statistical analysis applied the Mann-Whitney U test.
Figure 6.
A: Comparative analysis of pro-inflammatory cytokine mRNA Tnf by qPCR in the cornea (expressed in arbitrary units, normalized by β-actin mRNA). The G1 and G2 presented higher expression of Tnf at M2 (P=0.0257 and P=0.028, respectively). CG M2: x rats; G1 M1: x rats and G1 M2: x rats, G2 M1: x rats and G2 M2: x rats). The statistical analysis applied the Mann-Whitney U test. 6.B: Comparative analysis of pro-inflammatory cytokine mRNA Il1b by qPCR in the TG (expressed in arbitrary units, normalized by β-actin). G1 presented higher levels of Il1b at M1 (P=0.0286). CG M1: X rats, CG M2: x rats; G1 M1: x rats and G1 M2: x rats, G2 M1: x rats and G2 M2: x rats). The statistical analysis applied the Mann-Whitney U test. 6.C: Comparative analysis of pro-inflammatory cytokine mRNA MMP-9 by qPCR in the TG (expressed in arbitrary units, normalized by β-actin). G1 presented higher levels of Il1b at M1 (P=0.0286). CG M1: X rats, CG M2: x rats; G1 M1: x rats and G1 M2: x rats, G2 M1: x rats and G2 M2: x rats). The statistical analysis applied the Mann-Whitney U test.
2.2. LG partial Ablation and Allogeneic Transplant (G2)
2.2.1. In Vivo Exams
Corneal sensitivity challenged by capsaicin and measured by the eye wipe test revealed a similar number of forepaw eye wipes comparing G2 and controls (Table 1). However, partial LG ablation followed by grafting showed pronounced corneal neovascularization in the slit lamp exam at month 1 (Figure 1, upper lane). Again, tear secretion measurements (Table 2) and corneal epithelial cell impression cytology results were similar to controls at 1 or 2 months (P>0.05, Fisher test) (data not shown).
2.2.2. Ex Vivo Assays
In G2, PCNA levels remained similar to control group LG throughout the two months (data not shown).
H&E stained LG from G2 at month one revealed that the transplanted tissue retained some of its attributes. However, the ductal structure dilated, leukocyte infiltration occurred, and acinar structure was lost in the grafted area. The host LG of G2 was similar to the control (Figure 2).
At month 2, the histology changes in G2 LG became more pronounced. The ducts were more dilated, acinar cells were absent, the parenchyma was replaced by fibrosis, and there was leukocyte infiltration. Only in the interlobular bundle the vascular and ducts were preserved (Figure 3).
Despite no striking corneal histology changes at month 1 (data not shown)., at month 2, inflammatory cell infiltration and micro-vessels appeared in the corneal stroma (Figure 4). These observations agree with the results of the in vivo slit lamp exam of the rat corneas at month 1 (Figure 1). Also, at month 2, conjunctival and lid histology were well preserved, as indicated by intact epithelial and associated structures (data not shown).
The RT PCR analysis of the mRNA expression of inflammatory mediators in the LG revealed that only MMP-9 mRNA levels raised at months 1 and 2 in G2 (P=0.0051 and 0.0061) (Figure 5A).
The RT-PCR of the mRNA expression of proliferative mediator RUNX-3 in LG of G2 was higher than in the control group at month 2 (P=0.0149) (Figure 5C).
The RT-PCR assay of inflammatory cytokines mRNA in the cornea revealed that G2 presented higher levels of TNF-α mRNA at month 2 (P=0.0094)(Figure 6A) in agreement with the inflammatory repercussions of those interventions on the ocular surface, as indicated by the slit-lamp and histological analysis.
No changes in pro-inflammatory cytokines mRNA expression in TG compared to the control group at 1 and 2 months after the intervention was observed in G2 (Figure 6B,C).
2.3. LG Total Ablation (G3)
2.3.1. In Vivo Exams
Eye wipe test, followed by capsaicin eye drops, revealed raised corneal sensitivity by a higher mean number of forepaw eye wipes in the G3 at month 2 (P=0.0438) (Table 1).
Slit-lamp examination revealed that corneas of G3 remained unchanged in appearance (Figure 1, upper lane) but presented mild punctate keratitis on slit lamp and cobalt filter exam with 2% sodium fluorescein dye staining (Figure 1, lower lane). Those observations were stable at month 2 but included mild peripheral corneal neovascularization (data not shown).
The phenol red thread test means wetting showed similar tear flow to controls (Table 1). Again, corneal epithelial cell impression cytology did not reveal any metaplasia among the groups compared to controls at 1 or 2 months (P>0.05, Fisher test) (data not shown).
LG weight was not registered since it was removed at the beginning of the experimental period, and the surgical site was empty at the end of 2 months.
2.3.2. Ex Vivo Assays
For the same reason indicated above, PCNA expression levels in LG and LG histology after the experimental period of 1 and 2 months were not investigated in G3. Moreover, no striking corneal histology changes were observed in G3 at month 2 (Figure 4), nor in the conjunctival and lid histology at month 2 (data not shown).
The RT PCR analysis of the mRNA expression of inflammatory or proliferative mediators in the LG of G3 was not investigated in G3 again because the total ablation of the LG did not preserve the minimum in vivo remaining sample for analysis.
The RT-PCR assay to compare the expression of inflammatory cytokines mRNA in the cornea confirmed the lack of inflammatory repercussions of total LG ablation on the ocular surface, as indicated by the similar levels of mRNA expression of MMP-9, IL-1β, TNF-α, and IL-6 compared to the control group corneas (Figure 6A).
Considering that LG total ablation increased the corneal sensitivity in G3, we investigated the changes in pro-inflammatory cytokines mRNA expression in TG compared to the control group at 1 and 2 months after the intervention. The only change was that MMP-9 mRNA expression reduced at month 2 (P=0.0497) (Figure 6C). The other cytokines mRNA tested (IL-1β, IL-6, and TNF-α) presented no changes in TG compared to the control group at 1 or 2 months. This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3. Discussion
This study was planned to respond to two major questions. First, whether exorbital LG ablation is a reliable animal model for dry eye. Second, whether spontaneous or graft-induced regeneration occurs after planned surgical manipulation. Moreover, we investigated the clinical, histological, and molecular journey of those procedures (i.e.,; LG partial ablation, partial ablation, and allogeneic grafting and total ablation, respectively G1, G2, and G3).
The most relevant findings are the changes in corneal sensitivity and neovascularization in response to the surgical manipulations of the LG. Inflammatory mediators mRNA have raise not just in the LG but also in the cornea and TG. Those findings demonstrated the extra glandular extension of those interventions. They are in accordance with previous descriptions of regional and systemic repercussions of LG losses in mice. A recent study observed the response to LG complete ablation in mice and reported changes on the contralateral side, including inflammation of the LG, keratitis, and reduction in conjunctival goblet cells [35]. Another study with LG complete ablation in mice induced an increase of IL-1β, TNF-α, and MMP-9 mRNA in the ipsilateral cornea [28]. Mice’s behavior changed after LG’s complete ablation, including a reduction in libido, anxiety, and increased corneal sensitivity, and those changes were different when comparing male and female sex [36].
Differently from data obtained from mice, the rat exorbital LG ablation induced only mild keratitis and unchanged tear flow at 01 and 2 months. These observations do not agree with previous descriptions of tear flow reduction in 50% in male Sprague-Dawley (SD) rats 8 weeks after exorbital LG ablation and caution against the use of the rat model of LG ablation in dry eye comparative therapeutic studies [27]. Reduction in tear flow is superior to 30% in rats after LG complete ablation was described when both the exorbital and intraorbital LGs are removed [37].
The regenerative capacity of the LG was not observed here, although conceivable since rodent LG cells grow in culture and present markers of identifiable local stem cells [33,38,39]. In our study, complete or partial ablation did not respond to tissue regeneration, PCNA levels in the LG were similar to controls (data not shown), and tissue weight did not increase over two months. Furthermore, the study showed that allogeneic LG grafting is not tenable as a therapeutic option for rat LG repair since this procedure was not tolerated based on increases in structural change and inflammation in the transplanted tissue, which is different in method and response to submandibular salivary gland grafting in SD male rats, the exorbital Lg removal created a model of dry eye and where the graft integrated and maintained its structure and function [8]. The overexpression of RUNX 1 and 3, and MMP9 mRNA in the partially ablated LG (G1) suggested an attempt to tissue repair, but the lack of response of other mediators, such as BMP7 and FGF, also involved in LG organogenesis and regeneration suggests that inductive actions are necessary [33,40,41]. This negative finding points to the need for the development of novel procedures to overcome LG graft failure, probably involving LG organoid transplantation.
In mice, IL-1α stimulates LG inflammation but also allows its repair; fewer restorative effect was observed after LG duct ligation [30,42,43]. It may indicate selective response to different methods of damage, distinct animal species response, and the time-length among the variables involved in the delicate process of LG regeneration.
The chosen model must mimic the clinical signs associated with a DED of interest. In this regard, models induced by hormone deprivation and neural damage, sensorial or autonomic, are useful to mimic clinical conditions, the mechanisms, and proposed therapeutic strategies [4,44].
In the future, promoting LG regeneration in response to severe tissue damage, whether induced by trauma, surgical removal, or radiotherapy, among other conditions, the most promising strategies will include LG organogenesis and bioengineering [19,39]. Before this can be undertaken in a broad sense, a better understanding is needed of the interactions among morphogenesis-related proteins and their complex protective and disruptive balance in the physiopathology of exocrine diseases as recently demonstrated by the role of BMP6 in the Sjögren’s syndrome [45].
The limitations of the present work are the absence of more time-points of evaluation, including the short-term outcome, and the absence of comparative analysis of rats of female sex under those conditions. Another limitation is associated with the rudimentary technique applied to test LG graft integration and its role as an inductor of regeneration. A simple appose of part of LG obtained from a sibling and closing the skin wound with cyanoacrylate glue does not meet the LG healing in the present work as previously described [14]. Bioengeenired LG grown in vitro and customized placed in the diseased eye appears to be the clue for each causative severe DED condition [46].
In conclusion, exorbital LG ablation did not induce dry eye or LG repair in rats in any of the ablation models used in this study. Nevertheless, the differences in inflammatory responses and tissue injury are type-dependent and not limited to the LG, but also extended to the TG and cornea. Such variability may have subtle effects on ocular surface health and should support decisions on future methodological decisions on the topic of the dry eye model and LG regeneration.
4. Materials and Methods
4.1. Experimental Design and Animal Procedures
Eight-week-old male Wistar rats (Rattus norvegicus) were obtained from the Animal Breeding Centre of the School of Medicine at Ribeirão Preto, University of São Paulo (FMRP-USP), Ribeirão Preto, SP, Brazil. Animals were given free access to standard rodent chow and water and randomly separated into 4 study groups.
All experimental procedures adhered to the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research and were approved by the committee on animal experimentation of the School of Medicine at Ribeirão Preto, University of São Paulo.
The surgical interventions and the in vivo comparative studies were performed after intraperitoneal anesthesia with a combination of ketamine (5 mg/100 g b.w.) (União Química Farmacêutica S.A, Embu-Guaçu, SP, Brazil) and xylazine (2 mg/100g b.w.) (Laboratorio Callier S.A., Barcelona, Spain), after ensuring the absence of corneal and caudal reflexes.
Exam data and samples were compared among of the four following experimental rat groups:
Naïve Control (C): the eyes and extra orbital lacrimal glands were kept intact. In vivo exam data were compared with the other groups and the eye globe and extra orbital LG were used as a control in laboratory assays.
Group 1 (G1): aseptic surgical removal of anterior half part of the right extra orbital LG, with a cut (5 mm in length) through the skin in a lateral area of the head 3 mm equidistant between the eye and ear, followed by two cyanoacrylate glue drops (Locite, Henkel Ltd.a, Diadema, SP, Brazil) to close the skin margins, and covered with a single, 5 mm application of antibiotic and anti-inflammatory ointment (Cylocort, União Química Farmacêutica Nacional S.A, Brasilia, DF, Brazil).
Group 2 (G2): The aseptic surgical removal procedure was the same as for group 1. Subsequently, an allogeneic transplant from a twin brother was performed on LG tissue, which had approximately the same size, and it was embedded in the extirpated site. The LG was removed from the anesthetized donor; then, its capsule was opened and excised; the tissue was cut with a surgical blade in the Petry dish and washed with saline. The LG transplantation was completed when the LG graft was juxtaposed in the same topographical site of the receptor animal, also anesthetized, after a similar piece of LG tissue was removed. After bleeding control, cyanoacrylate glue was applied to the borders of the skin to close the skin wound, followed by a 5 mm application of the same antibiotic and anti-inflammatory ointment (n=10 rats). The rationale was to investigate the possibility of the grafted tissue regarding histological and functional restoration.
Group 3 (G3): aseptic skin incision and exposure of right extra orbital LG, followed by complete LG ablation and homeostasis of the vascular beds. Afterward, the skin was glued and treated as mentioned above (n=10 rats). The skin wounds were undetectable two days later in all animals of the three groups (n=10 rats/group). The rationale was to investigate the impact of the absence of the LG on the OS and the possibility of the local germination of a novel LG and its functional restoration.
Animals were evaluated after 01 and 02 months of the procedure (n=05 rats /group per time point).
4.1.1. In Vivo Exam
At months 1 and 2 after surgical intervention, the body weight of the animals was registered.
4.1.2. Eye Wipe Test
At the end of the two experimental periods (1m and 2 m) for each group (i.e.,; C, G1, G2, and G3), the rats were submitted to the eye wipe test in response to capsaicin (CAP) to investigate the CO sensitivity. Following acclimation of the animals to Plexiglas transparent chambers for one hour, the right eyes of all models were treated with instillation of 20 μl of 10 μM CAP diluted in PBS at pH 7.2 and 25ºC (Sigma-Aldrich Brasil Ltd.a., COTIA, SP, BRAZIL).
The eye wipe behavior was recorded with a digital camera (DSC-W5, Sony, Japan) for 5 min after the instillation of CAP. Eye wipe movements with the forepaws (EWT) registered along of 3 min, starting 1 min after the CAP eye drop, were counted afterward from the digital movies recorded of each rat by a masked observer using an iMac computer (Apple Inc, Cupertino, CA, USA) and the means were compared among the groups.
Afterward, the rats were anesthetized with the same drugs and doses used for surgical interventions, and the eyes were examined with a slit lamp (Carl Zeiss, Germany). The corneal epithelial integrity was evaluated on a slit lamp and cobalt filter after 2% sodium fluorescein dye staining. The punctate keratitis was graded from 0 to 15, as previously described [34].
Tear flow was measured with the phenol red thread (PRT) for 30 seconds, in millimeters, and the values obtained were compared among the groups (Showa Yakuhin Kako Co; Ltd., Tokyo, Japan & Menicon USA Inc., Clovis, CA, USA).
4.1.3. Tissue Harvesting and Storage for Analysis
Extra orbital LG, eye globes with lids, and trigeminal ganglions (TG) were removed at the end of months 1 and 2 of the experimental period. The tissues were weighed, and the samples were processed for histology, qRT-PCR, and western blotting, as detailed below. The rats were euthanized with excess anesthesia (ketamine 15 mg/100 g of body weight and xylazine 6 mg/100 g body weight) and sodium Thiopental (1000 mg/kg) (Laboratório Cristália, São Paulo, SP, Brazil).
4.1.4. Impression Cytology
Cornea epithelial cells were harvested from the temporal area with 0.45 µmpore size filter paper for impression cytology (Millipore, Billerica, MA, USA), also at months 1 and 2.
Corneal epithelial cells adherent to the filter paper were fixed with a mixture of ethanol, 70% glacial acetic acid, and formalin, stained with periodic acid-Schiff (PAS) and hematoxylin. They were transferred to microscope slides. Squamous metaplasia of epithelial cells was staged in a masked fashion according to a four-stage classification scheme from 0 (normal morphology) to 3 (squamous metaplasia) based on the coloration and cytoplasmic appearance: nuclei size and presence of mucous secretion. The images were analyzed with an Olympus BX40 light microscope and photographed with a digital camera (Olympus Q-color 5) at 100 and 400 x magnification (Olympus Corporation, Tokyo, Japan).
4.1.5. Western Blotting
LG was homogenized in a buffer containing 50 mM Tris, pH 7.5, 500 mM NaCl, 0.1% Triton, and protease inhibitor cocktail set III (Calbiochem, San Diego, CA) with a polytron (Virsonic, Biopharma, Winchester, UK). PCNA expression levels in whole cell lysates were evaluated to assess cell proliferative activity.
After homogenization, the protein was measured by the biuret µdye test. Samples were treated with Laemmli buffer, and equal amounts of protein per sample (70 µg) were subjected to SDS-PAGE (10% Tris-acrylamide) in a Bio-Rad miniature lab gel apparatus (Miniprotean, Bio-Rad Laboratories, Richmond, CA, USA), in parallel with pre-stained protein standards and dithiothreitol (Bio-Rad, Hercules, CA, USA). Proteins were then electro-transferred from the gel to a 0.22 µm nitrocellulose membrane (Bio-Rad, Hercules, CA, USA) for 2 h at 120 V in a Bio-Rad miniature transfer apparatus (Miniprotean). After blocking, the membranes were incubated overnight using anti- PCNA or alpha-tubulin antibodies (Table 2), in a buffer containing 3% bovine serum albumin (BSA), and then washed three times with tris buffered saline/tween (TBS/T). Detection was performed using enhanced chemiluminescence (SuperSignal West Pico, Pierce, Rockford, IL) after 2-h incubation with a horseradish peroxidase-conjugated secondary antibody (1:10,000, Invitrogen, São Paulo, SP, Brazil). Band intensities were quantified by optical densitometry (Scion Image, Frederick, MD, USA), α-tubulin was used as an internal control.
4.1.6. Histology
The LG and cornea samples for histology were collected and frozen in OCT compound (Sakura Fine Tek Inc., Torrance, CA, USA). After tissue cutting (6 μm) and transferred to slides, the 10th to the 14th LG and corneal sections were submitted to hematoxylin/eosin staining (5 samples per animal, n=5/group). Digital photos were obtained (Nikon Eclipse E800, Nikon USA, Melville, NY), and the structure and area of the LG were compared in a masked manner.
4.1.7. Quantitative Real-Time PCR
The CO, LG, and TG tissues from the right side of the rats of the 4 groups were harvested and imbedded in RNA stabilization solution (RNAlater Solution, Ambion, Waltham, MA, USA) and stored at -80oC until proceeding to RNA extraction, quantification, quality evaluation, and Real-Time quantitative PCR (qPCR) analysis. The relative expression of the mRNA of proinflammatory cytokines Il1b, Il6, Tnf, Mmp9 was compared in LG, CO, and TG samples from the groups. In addition, the relative expression of the mRNA of the tissue repair elements in LG Bmp7, Runx1, Runx3, Fgf10, and Smad1 was compared in those groups using commercial primers and beta-actin mRNA as an internal control (Life Technologies, Carlsbad, CA, USA).
Total RNA samples were extracted from the tissues using PureLink MiniKit (Ambionby Life Technologies, USA), and the kit DNA-Free DNase Treatment & Removal (Ambionby Life Technologies, USA) to remove genomic DNA contamination, according to the manufacturer’s instructions and RNA was quantified with a spectrophotometer NanoDrop 2000c (Thermo Scientific, Wilmington, DE, USA).
Samples containing 500ng of total RNA of cornea tissue, 1000ng of total RNA of LG tissue, and 350ng of TG were used to synthesize the cDNA with the QuantiTect Reverse Transcription Kit (Qiagen, Germantown, MD, USA) in the ProFlex PCR System (Applied Biosystems, Carlsbad, CA, USA).
qPCR was performed using ViiA7 Real-time PCR System (Applied Biosystems, Carlsbad, CA, USA). The following hydrolysis probes were used in this study: Rn.PT 5838028824 (Il1b), Rn.PT 5813840513 (Il6), Rn.PT 5811142874 (Tnf), Rn.PT 587383134 (Mmp9), Rn.PT 5810180444 (Bmp7), Rn.PT 5810814634 (Fgf10), Rn.PT 589220704.g (β-actin)(all these from IDT); Rn00565555_m1 (Smad1), Rn00569082_m1 (Runx1), Rn00590466_m1 (Runx3)(Applied Biosystems, Carlsbad, CA, USA). Each amplification reaction was duplicated with 5.5 uL of QuantiNova Probe PCR Kit (Qiagen, Germantown, MD, USA), 0.5 μL of hydrolysis probe, and 4.5 μL of 1:4 dilution of the cDNA in a total volume of 10 μL. The cycles for real-time PCR were as follows: one cycle of 95°C for 2 minutes, 50 cycles of 5 seconds at 95°C, and 19 seconds at 60°C.
The relative quantification was determined by the Thermo Fisher Cloud Software, RQ version 3.7 (Life Technologies Corporation, Carlsbad, CA, USA).
4.2. Statistical Analysis
Data are reported as mean ± SEM. The normal distribution of the continuous data was investigated by the Shapiro-Wilk test. Since the data deviated from a normal distribution, the comparisons between the groups 1-3 and control, and between baseline versus 01 and 02 months of observation, were made using the Mann-Whitney U test for continuous data. The Fisher exact test was applied for categorical data, comparing each group with the corresponding control group (GraphPad 5.0 software; Prism, San Diego, CA). Densitometry values of western blotting were normalized to alpha-tubulin and reported as a ratio to validate loading equivalence. The RT-PCR values of each probe were compared as a ratio of β-actin for each tissue, condition, and time-point. The level of significance was set at p <0.05.
5. Conclusions
Without changes in tear flow or evidence of LG regeneration, LG ablation or grafting are unreliable models for dry eye or LG repair in rats. The surgical manipulation extended inflammation to the cornea and TG.
6. Patents
This section is not mandatory but may be added if there are patents resulting from the work reported in this manuscript.
Author Contributions
Conceptualization, EMR, EBMF and AAMS; methodology, AABM, AMSS, LECMS, MZF; formal analysis, EMR, AABM, DMG and MZF; investigation, EMR, AMSS and EBMF; resources, EMR; data curation, AABM, MZF and DMG; writing—original draft preparation, EMR; writing—review and editing, EMR; visualization, EMR and DMG; supervision, EMR; project administration, EMR; funding acquisition, EMR. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by following Brazilian institutions: The São Paulo Research Foundation (FAPESP)(FAPESP) 2015/20580-7, 2014/22451-7 and 2014/23211-0; The Brazilian National Council for Scientific and Technological Development (CNPq) Research Productivity Scholarship: 1C 302355/2017-0, 01/03/2018 to 28/02/2022; “Fundação de Apoio ao Ensino, Pesquisa e Assistência do Hospital das Clinicas” of Ribeirao Preto Medical School, São Paulo University (FAEPA), Research Core of Ocular Physiopathology and Therapeutics from University of São Paulo (NAP-FTO) 12.1.25431.01.7 and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, financial code 001.
Acknowledgments
The authors would like to thank Maria Helena Andrade from the Department of Ophthalmology, Otorhinolaryngology, and Head &Neck Surgery for assistance with this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Scherz, W.; Dohlman, C.H. Is the Lacrimal Gland Dispensable? Keratoconjunctivitis Sicca after Lacrimal Gland Removal. Arch. Ophthalmol. 1975, 93, 281–283. [Google Scholar] [CrossRef]
- Pflugfelder, S.C. Tear Dysfunction and the Cornea: LXVIII Edward Jackson Memorial Lecture. Am. J. Ophthalmol. 2011, 152, 900–909e1. [Google Scholar] [CrossRef]
- Stevenson, W.; Pugazhendhi, S.; Wang, M. Is the Main Lacrimal Gland Indispensable? Contributions of the Corneal and Conjunctival Epithelia. Surv. Ophthalmol. 2016, 61, 616–627. [Google Scholar] [CrossRef]
- Barros, J.F.F.; Sant’Ana, A.M.S.; Dias, L.C.; Murashima, A.d.A.B.; da Silva, L.E.C.M.; Fantucci, M.Z.; Rocha, E.M. Comparison of the Effects of Corneal and Lacrimal Gland Denervation on the Lacrimal Functional Unit of Rats. Arq. Bras. Oftalmol. 2022, 85, 59–67. [Google Scholar] [CrossRef]
- Stern, M.E.; Gao, J.; Siemasko, K.F.; Beuerman, R.W.; Pflugfelder, S.C. The Role of the Lacrimal Functional Unit in the Pathophysiology of Dry Eye. Exp. Eye Res. 2004, 78, 409–416. [Google Scholar] [CrossRef]
- Schechter, J.E.; Warren, D.W.; Mircheff, A.K. A Lacrimal Gland Is a Lacrimal Gland, but Rodent’s and Rabbit’s Are Not Human. Ocul. Surf. 2010, 8, 111–134. [Google Scholar] [CrossRef]
- Qin, Y.; Tan, X.; Zhang, Y.; Jie, Y.; Labbe, A.; Pan, Z. A New Nonhuman Primate Model of Severe Dry Eye. Cornea 2014, 33, 510–517. [Google Scholar] [CrossRef]
- Liu, W.-C.; Hsu, W.-M.; Lee, S.-M.; Lee, F.-L.; Yen, M.-Y.; Chou, C.-K.; Kao, S.-C. Transplantation of the Autologous Submandibular Gland to the Lacrimal Basin in Rats. Ophthalmic Res. 2004, 36, 195–199. [Google Scholar] [CrossRef]
- Odaka, A.; Toshida, H.; Ohta, T.; Tabuchi, N.; Koike, D.; Suto, C.; Murakami, A. Efficacy of Retinol Palmitate Eye Drops for Dry Eye in Rabbits with Lacrimal Gland Resection. Clin. Ophthalmol. 2012, 6, 1585–1593. [Google Scholar] [CrossRef]
- Zoukhri, D. Mechanisms Involved in Injury and Repair of the Murine Lacrimal Gland: Role of Programmed Cell Death and Mesenchymal Stem Cells. Ocul. Surf. 2010, 8, 60–69. [Google Scholar] [CrossRef]
- Tiwari, S.; Ali, M.J.; Vemuganti, G.K. Human Lacrimal Gland Regeneration: Perspectives and Review of Literature. Saudi J Ophthalmol 2014, 28, 12–18. [Google Scholar] [CrossRef]
- Bron, A.J.; de Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II Pathophysiology Report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef]
- Maitchouk, D.Y.; Beuerman, R.W.; Ohta, T.; Stern, M.; Varnell, R.J. Tear Production after Unilateral Removal of the Main Lacrimal Gland in Squirrel Monkeys. Arch. Ophthalmol. 2000, 118, 246–252. [Google Scholar] [CrossRef]
- Nemet, A.; Belkin, M.; Rosner, M. Transplantation of Newborn Lacrimal Gland Cells in a Rat Model of Reduced Tear Secretion. Isr. Med. Assoc. J. 2007, 9, 94–98. [Google Scholar]
- Rocha, E.M.; Cotrim, A.P.; Zheng, C.; Riveros, P.P.; Baum, B.J.; Chiorini, J.A. Recovery of Radiation-Induced Dry Eye and Corneal Damage by Pretreatment with Adenoviral Vector-Mediated Transfer of Erythropoietin to the Salivary Glands in Mice. Hum. Gene Ther. 2013, 24, 417–423. [Google Scholar] [CrossRef]
- Kumar, P.A.; Macleod, A.M.; O’Brien, B.M.; Hickey, M.J.; Knight, K.R. Microvascular Submandibular Gland Transfer for the Management of Xerophthalmia; an Experimental Study. Br. J. Plast. Surg. 1990, 43, 431–436. [Google Scholar] [CrossRef]
- MacLeod, A.M.; Robbins, S.P. Submandibular Gland Transfer in the Correction of Dry Eye. Aust. N. Z. J. Ophthalmol. 1992, 20, 99–103. [Google Scholar] [CrossRef]
- Geerling, G.; Borrelli, M. Adnexal Surgery for Severe Ocular Surface Disease. Seminars in Ophthalmology 2005, 20, 101–112. [Google Scholar] [CrossRef]
- Hirayama, M.; Ogawa, M.; Oshima, M.; Sekine, Y.; Ishida, K.; Yamashita, K.; Ikeda, K.; Shimmura, S.; Kawakita, T.; Tsubota, K.; et al. Functional Lacrimal Gland Regeneration by Transplantation of a Bioengineered Organ Germ. Nat. Commun. 2013, 4, 2497. [Google Scholar] [CrossRef]
- Nover, A. Experimental studies on regeneration and transplantation capacity of the lacrimal gland under local and general hormonal stimulation (thyroxin). II. Albrecht Von Graefes. Arch. Ophthalmol. 1954, 156, 98–118. [Google Scholar] [CrossRef]
- Geerling, G.; Sieg, P.; Bastian, G.O.; Laqua, H. Transplantation of the Autologous Submandibular Gland for Most Severe Cases of Keratoconjunctivitis Sicca. Ophthalmology 1998, 105, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Bagnoli, S. Morphological, cytochemical and body-weight aspects of regeneration in the submaxillary salivary gland of the rat. Arch. De Vecchi Anat. Patol. 1957, 26, 203–230. [Google Scholar] [PubMed]
- Mac Cord Medina, F.; Silvestre de Castro, R.; Leite, S.C.; Rocha, E.M.; de Melo Rocha, G. Management of Dry Eye Related to Systemic Diseases in Childhood and Longterm Follow-Up. Acta Ophthalmol. Scand. 2007, 85, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.B.; Shashi, V.; Kirse, D.J. Case Report: Aplasia of the Lacrimal and Major Salivary Glands (ALSG). Int. J. Pediatr. Otorhinolaryngol. 2009, 73, 899–901. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.F.; Akaishi, P.M.S.; de Molfetta, G.A.; Chodraui-Filho, S.; Cintra, M.; Toscano, A.; Silva, W.A., Jr.; Cruz, A.A.V. Lacrimal Gland Involvement in Blepharophimosis-Ptosis-Epicanthus Inversus Syndrome. Ophthalmology 2017, 124, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.P.; de Oliveira, F.R.; da Rocha, F.J.; Muglia, V.F.; Rocha, E.M. Lacrimal Gland Atrophy and Dry Eye Related to Isotretinoin, Androgen, and Prolactin: Differential Diagnosis for Sjögren’s Syndrome. Arq. Bras. Oftalmol. 2021, 84, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, T.; Murakami, T.; Fujita, H.; Nakamura, M.; Nakata, K. Improvement of Corneal Barrier Function by the P2Y(2) Agonist INS365 in a Rat Dry Eye Model. Invest. Ophthalmol. Vis. Sci. 2001, 42, 96–100. [Google Scholar]
- Stevenson, W.; Chen, Y.; Lee, S.-M.; Lee, H.S.; Hua, J.; Dohlman, T.; Shiang, T.; Dana, R. Extraorbital Lacrimal Gland Excision: A Reproducible Model of Severe Aqueous Tear-Deficient Dry Eye Disease. Cornea 2014, 33, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sharma, S.; Basu, S. Rabbit Models of Dry Eye Disease: Current Understanding and Unmet Needs for Translational Research. Exp. Eye Res. 2021, 206, 108538. [Google Scholar] [CrossRef]
- Zoukhri, D.; Fix, A.; Alroy, J.; Kublin, C.L. Mechanisms of Murine Lacrimal Gland Repair after Experimentally Induced Inflammation. Invest. Ophthalmol. Vis. Sci. 2008, 49, 4399–4406. [Google Scholar] [CrossRef]
- Makarenkova, H.P.; Ito, M.; Govindarajan, V.; Faber, S.C.; Sun, L.; McMahon, G.; Overbeek, P.A.; Lang, R.A. FGF10 Is an Inducer and Pax6 a Competence Factor for Lacrimal Gland Development. Development 2000, 127, 2563–2572. [Google Scholar] [CrossRef]
- Dean, C.; Ito, M.; Makarenkova, H.P.; Faber, S.C.; Lang, R.A. Bmp7 Regulates Branching Morphogenesis of the Lacrimal Gland by Promoting Mesenchymal Proliferation and Condensation. Development 2004, 131, 4155–4165. [Google Scholar] [CrossRef]
- Dietrich, J.; Schrader, S. Towards Lacrimal Gland Regeneration: Current Concepts and Experimental Approaches. Curr. Eye Res. 2020, 45, 230–240. [Google Scholar] [CrossRef]
- Wolffsohn, J.S.; Arita, R.; Chalmers, R.; Djalilian, A.; Dogru, M.; Dumbleton, K.; Gupta, P.K.; Karpecki, P.; Lazreg, S.; Pult, H.; et al. TFOS DEWS II Diagnostic Methodology Report. Ocul. Surf. 2017, 15, 539–574. [Google Scholar] [CrossRef]
- Kim, M.; Kim, S.Y.; Jeon, J.W.; Lee, H.K. Impact of Lacrimal Gland Extraction on the Contralateral Eye in an Animal Model for Dry Eye Disease. Korean J. Ophthalmol. 2022. [CrossRef]
- Mecum, N.E.; Cyr, D.; Malon, J.; Demers, D.; Cao, L.; Meng, I.D. Evaluation of Corneal Damage After Lacrimal Gland Excision in Male and Female Mice. Investigative Opthalmology & Visual Science 2019, 60, 3264. [Google Scholar]
- Kurose, M.; Meng, I.D. Dry Eye Modifies the Thermal and Menthol Responses in Rat Corneal Primary Afferent Cool Cells. J. Neurophysiol. 2013, 110, 495–504. [Google Scholar] [CrossRef]
- Malki, L.T.; Dias, A.C.; Jorge, A.G.; Módulo, C.M.; Rocha, E.M. Lacrimal Gland Primary Acinar Cell Culture: The Role of Insulin. Arq. Bras. Oftalmol. 2016, 79, 105–110. [Google Scholar] [CrossRef]
- Dietrich, J.; Roth, M.; König, S.; Geerling, G.; Mertsch, S.; Schrader, S. Analysis of Lacrimal Gland Derived Mesenchymal Stem Cell Secretome and Its Impact on Epithelial Cell Survival. Stem Cell Res. 2019, 38, 101477. [Google Scholar] [CrossRef]
- Hirayama, M.; Ko, S.B.H.; Kawakita, T.; Akiyama, T.; Goparaju, S.K.; Soma, A.; Nakatake, Y.; Sakota, M.; Chikazawa-Nohtomi, N.; Shimmura, S.; et al. Identification of Transcription Factors That Promote the Differentiation of Human Pluripotent Stem Cells into Lacrimal Gland Epithelium-like Cells. NPJ Aging Mech Dis 2017, 3, 1. [Google Scholar] [CrossRef]
- Hirayama, M.; Kawakita, T.; Tsubota, K.; Shimmura, S. Challenges and Strategies for Regenerating the Lacrimal Gland. Ocul. Surf. 2016, 14, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hirayama, M.; Kawakita, T.; Tsubota, K. A Ligation of the Lacrimal Excretory Duct in Mouse Induces Lacrimal Gland Inflammation with Proliferative Cells. Stem Cells Int. 2017, 2017, 4923426. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, J.; Schlegel, C.; Roth, M.; Witt, J.; Geerling, G.; Mertsch, S.; Schrader, S. Comparative Analysis on the Dynamic of Lacrimal Gland Damage and Regeneration after Interleukin-1α or Duct Ligation Induced Dry Eye Disease in Mice. Experimental Eye Research 2018, 172, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Jorge, A.G.; Módulo, C.M.; Dias, A.C.; Braz, A.M.; Filho, R.B.; Jordão, A.A., Jr.; de Paula, J.S.; Rocha, E.M. Aspirin Prevents Diabetic Oxidative Changes in Rat Lacrimal Gland Structure and Function. Endocrine 2009, 35, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.-Q.; Nakamura, H.; Tanaka, T.; Odani, T.; Perez, P.; Ji, Y.; French, B.N.; Pranzatelli, T.J.F.; Michael, D.G.; Yin, H.; et al. Lysosomal Exocytosis of HSP70 Stimulates Monocytic BMP6 Expression in Sjögren’s Syndrome. Journal of Clinical Investigation 2022, 132. [Google Scholar] [CrossRef]
- Massie, I.; Dietrich, J.; Roth, M.; Geerling, G.; Mertsch, S.; Schrader, S. Development of Causative Treatment Strategies for Lacrimal Gland Insufficiency by Tissue Engineering and Cell Therapy. Part 2: Reconstruction of Lacrimal Gland Tissue: What Has Been Achieved so Far and What Are the Remaining Challenges? Curr. Eye Res. 2016, 41, 1255–1265. [Google Scholar] [CrossRef]
Figure 1.
Upper lane: Slit lamp observation of the ocular surface of rats, submitted or not to LG intervention. The photos illustrate the outcome at a 1-month follow-up of eyes from controls (C), LG partial ablation (G1), ablation followed by allogeneic LG transplant (G2), and total LG ablation. Lower lane: Slit lamp observation of the ocular surface of rats, submitted or not to LG intervention, using a cobalt light filter and fluorescein dye. The photos illustrate the outcome at 1-month follow-up of eyes from controls (C′), LG partial ablation (G1′), ablation followed by allogeneic LG transplant (G2′), and total LG ablation (G3′).
Figure 1.
Upper lane: Slit lamp observation of the ocular surface of rats, submitted or not to LG intervention. The photos illustrate the outcome at a 1-month follow-up of eyes from controls (C), LG partial ablation (G1), ablation followed by allogeneic LG transplant (G2), and total LG ablation. Lower lane: Slit lamp observation of the ocular surface of rats, submitted or not to LG intervention, using a cobalt light filter and fluorescein dye. The photos illustrate the outcome at 1-month follow-up of eyes from controls (C′), LG partial ablation (G1′), ablation followed by allogeneic LG transplant (G2′), and total LG ablation (G3′).
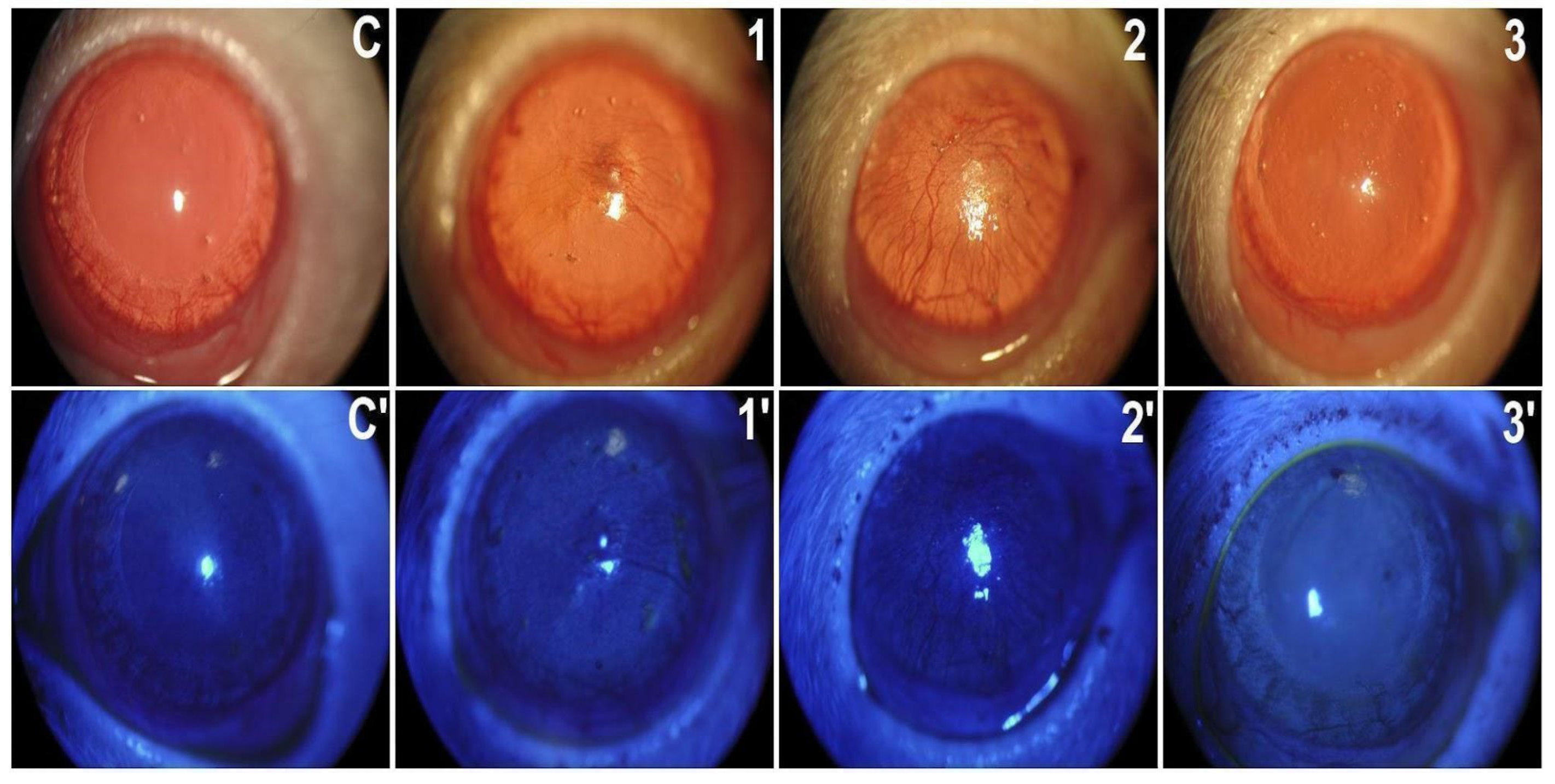

Table 1.
Behavioral and morphometric analysis of body weight, LG weight and tears flow (phenol red thread test) of the control (C) or the following intervention: LG partial ablation (G1), ablation followed by allogeneic LG transplant (G2), total ablation (G3). (mean ± SEM) (Mann-Whitney U test).
Table 1.
Behavioral and morphometric analysis of body weight, LG weight and tears flow (phenol red thread test) of the control (C) or the following intervention: LG partial ablation (G1), ablation followed by allogeneic LG transplant (G2), total ablation (G3). (mean ± SEM) (Mann-Whitney U test).
| Eye Wipe Test (Mean Wipes in 3min) | Phenol Red Thread Test (mm) | LG Weight (mg) | P versus Control | ||
|---|---|---|---|---|---|
| Control | 1m | 5.6 ± 1.4 | 8.1±2.0 | 132.6±15.4 | |
| 2m | 9.2±2.6 | 125.3±9.2 | |||
| G1 | 1m | 2.6 ± 1.2 | 6.8±1.7 | 50.3±18.3 | |
| 2m | 3.3 ± 1.3 | 10.4± 2.5 | 41.3±14.6# | # 0.0001 | |
| G2 | 1m | 14.4 ± 2.2 | 6.2±1.4 | 59.7±8.6* | * 0.0079 |
| 2m | 7.0 ± 1.1 | 5.8± 1.1 | 45.0±7.8# | # 0.0001 | |
| G3 | 1m | 4.7 ± 2.3 | 8.2±1.9 | N/A | |
| 2m | 15.3± 4.1 § | 5.2± 1.1 | N/A | § 0.0438 |
Table 2.
Antibodies used for Western blot analysis for comparison of apoptotic and proliferative response to interventions among experimental groups (G1 to G4) and control rats LG.
Table 2.
Antibodies used for Western blot analysis for comparison of apoptotic and proliferative response to interventions among experimental groups (G1 to G4) and control rats LG.
| Catalog Number | Isotype | |
|---|---|---|
| Molecular weight (kDa) | Concentration | |
| α-tubulin | Santa Cruz | |
| SC 8035 | Mouse | |
| Monoclonal | 55 | 200µg/mL |
| PCNA | Cell Signaling | |
| # 2586 | Mouse | |
| Monoclonal | 36 | 200 µg/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



