
Submitted:
29 April 2024
Posted:
01 May 2024
You are already at the latest version
Abstract
Hepatitis C virus (HCV) is an oncogenic virus that causes chronic liver disease in more than 80% of patients. During the last decade, efficient direct acting antivirals were introduced into clinical practice. However, clearance of the virus does not reduce risk of end-stage liver diseases to the level observed in patients that have never been infected. So, investigation of HCV pathogenesis is still warranted. Virus-induced changes in cell metabolism contribute to the development of HCV-associated liver pathologies. Here we studied the impact of the virus on the metabolism of polyamines and proline as well as on the urea cycle, which plays a crucial role in liver function. It was found that HCV strongly suppresses the expression of arginase, a key enzyme of the urea cycle, leading to accumulation of arginine and upregulates proline oxidase with a concomitant decrease in proline concentrations. Addition of exogenous proline moderately suppressed viral replication. HCV upregulated transcription but suppressed protein levels of polyamine-metabolizing enzymes. This resulted in a decrease in polyamine content in infected cells. Finally, compounds targeting polyamine metabolism demonstrated pronounced antiviral activity pointing to spermine and spermidine as compounds affecting HCV replication. These data expand our understanding of HCV’s imprint on cell metabolism.
Keywords:
hepatitis C virus
; polyamines
; urea cycle
; proline metabolism
; antiviral agents
1. Introduction
Hepatitis С virus (HCV) is responsible for 9% of chronic liver disease cases worldwide [1]. Currently the World Health Organization estimates 58 million people to be chronically infected with this virus [2]. Chronic hepatitis C (CHC) is often accompanied by persistent liver inflammation and gradual development of fibrosis [3]. As result, one in 3-10 patients will develop liver cirrhosis within a 20-30 year period [2,3,4]. Patients with liver cirrhosis have substantial risks of hepatic decompensation and hepatocellular carcinoma. HCV infection is the cause of death of almost 400 000 patients with end-stage liver disease each year [1]. Combinations of direct acting antivirals (DAAs) can clear the infection in almost every patient, though some CHC carriers with genotype 3 of the virus or with decompensated cirrhosis may not achieve sustained virological responses (SVR) [5,6]. While the high price of anti-HCV drugs still limits access to therapy in some parts of the world, insufficient screening allows between 68-94% of chronic hepatitis C cases to remain undiagnosed [7]. Moreover, SVR upon DAA treatment does not reduce the risk of liver cirrhosis and cancer to the level in the uninfected population [8]. Therefore, more insight into HCV pathogenesis is needed in order to develop drugs that may suppress or prevent liver dysfunction. One of the directions in this area is investigation of virus-induced changes in metabolism of infected cells.
HCV infection drives fibrogenesis and hepatocarcinogenesis by multiple mechanisms. These include chronic inflammation [9], production of profibrotic cytokines (i.e. TGFβ1) [10], interference of viral proteins with multiple signalling pathways in infected hepatocytes and concomitant inhibition of apoptosis and increased cell survival [11], overproduction of reactive oxygen species (ROS), dysregulation of antioxidant defence systems [12,13], and alteration of metabolic pathways [14]. HCV was reported to perturb multiple metabolic pathways including glycolysis, glutaminolysis, and amino acid metabolism. Specifically, this virus increases rates of glycolysis and lactate production [15,16,17] and enhances consumption of glutamine and its conversion into α-ketoglutarate to drive tricarbonic acid (TCA) cycle in a tumour-like fashion [18]. HCV also increases levels of nucleos(t)ides, some amino acids and fatty acids and decreases intracellular concentration of AcCoA [15,17,19]. Replication of the virus also relies on metabolites such as sphingomyelin that is critical for formation of double membrane vesicles where the virus replicase is localized [20]. However, the status of many other important metabolic pathways in HCV-infected hepatocytes remains unknown.
Biogenic polyamines are low molecular weight compounds that carry several positively charged amino groups and thus can interact electrostatically with negatively charged nucleic acids and proteins. Spermine and spermidine are found in all types of eukaryotic cells at high concentrations, with their levels generally correlating with cell proliferation rates [21]. Polyamines are synthesized from ornithine, a metabolite of the urea cycle that is also linked to proline and glutamine/glutamate metabolic pathways (Scheme 1) [22,23]. The rate-limiting step of polyamine biosynthesis is the conversion of ornithine into diamine putrescine (1,4-diaminobutane) catalysed by ornithine decarboxylase (ODC). Putrescine is then converted into spermidine and spermine by spermidine and spermine synthases (SRM, SMS), respectively with decarboxylated S-adenosyl methionine (dcSAM) being provided by S-adenosylmethionine decarboxylase (AMD). Polyamine degradation is achieved by two alternative pathways. The first one is a two-step process of acetylation by spermidine/spermine-N1-acetyl transferase (SSAT) with oxidation of acetylated spermine and spermidine by acetylpolyamine oxidase (PAOX) into spermidine and putrescine, respectively. An alternative pathway is a direct oxidation of spermine into spermidine by spermine oxidase (SMOX). Cells maintain levels of spermine and spermidine by multi-level control of expression of ODC, SSAT, SMOX, and, to some extent, AMD. Increased polyamine concentrations are a hallmark of hyperproliferative disorders such as cancer and autoimmune diseases [24]. At the same time, increased expression of SMOX that produces hydrogen peroxide and toxic acrolein is a feature of stroke, acute pancreatitis, or is the key factor of carcinogenesis during Helicobacter pylori or Bacteroides fragilis infections [25,26]. Thus, targeting polyamine-metabolizing enzymes with pharmacologic inhibitors/activators is considered as a promising strategy for treatment of cancer. An example is the considerable progress that was achieved in treatment of neuroblastoma by addition of ODC inhibitor difluoromethylornithine (DFMO) to the standard combination of drugs [27,28].
However, much less is known about the interplay between polyamines with viral infections [22]. Previously we reported that transient expression of HCV core and NS5A proteins affects expression of the key polyamine-metabolizing enzymes via enhanced ROS production [29]. Moreover, a stable Huh7 cell line harbouring a full-length HCV replicon exhibited suppressed expression of ODC and SSAT, up-regulated expression of SMOX, and diminished levels of spermine and spermidine. However, all this was shown in a non-infectious system, and warrants investigation in a model that ensures all steps of the viral life cycle. So far, nothing is known on whether polyamines are important for HCV replication, although other viruses have been shown to rely on spermine and spermidine at various stages of their life cycles [30]. The primary goal of this project was to investigate if hepatitis C virus interferes with metabolism of biogenic polyamines and related metabolites, and if compounds that target polyamine-metabolizing enzymes affect HCV replication.
2. Materials and Methods
2.1. Reagents
DFMO was a kind gift of Prof. P. Woster (Medical University of South Carolina, Charleston, SC, USA), N,N’-bis(2,3-butadienyl)-1,4-butanediamine (MDL72.527), deferiprone, ciclopirox and (S)-(+)-5-oxo-2 tetrahydrofuracarboxylic acid (THFA) were purchased from Sigma (St. Louis, MO, USA), L-canaline – from Caiman Chemical (Ann Arbor, MI, USA), while N1,N11-diethylnorspermine (DENSpm), N1-guanyl-1,7-diaminoheptane (GC7), N-(3-aminopropyl)cyclohexylamine (APCHA) and trans-4-methyl-cyclohecylamine (4MCHA) were supplied by Santa-Cruz Biotechnologies (Dallas, TX, USA). Sardomoside (SAM486A) was purchased from Cayman Chemicals (Ann Arbor, MI, USA). N1-Ethyl-N11-(cyclopropyl)-methyl-4,8-diazaundecane (CPENSpm) and N1-ethyl-N11-(cycloheptyl)-methyl-4,8-diazaundecane (CHENSpm) were kindly given by Dr. Tuomo Keinanen (University of Eastern Finland). 3-Aminooxy-1-aminopropane (APA) was synthesized as described in [31]. DMEM and DMEM-F12 media were from Life Technologies (Carlsbad, CA, USA). Plasmax medium was assembled as described earlier [32,33]. [14C]-labelled ornithine and AcCoA were from from GE Healthcare (Little Chalfont, UK). All other reagents were provided by Sigma. Oligonucleotides were synthesized by Evrogen J.S.C. (Moscow, Russia).
Huh7.5 cells and the plasmids pJFH1 and pSGR-JFH1 were kindly provided by Prof. C.M. Rice (The Rockefeller University, NY, USA), Prof. Takaji Wakita (National Institute of Infectious Diseases, Tokyo), and Apath L.L.C. (Brooklyn, NY, USA).
2.2. Cells and HCV Infection Systems
Huh7.5 cells were cultivated in a high glucose Dulbecco's Modified Eagle Medium (DMEM) with GlutaMax supplemented with 10% fetal bovine serum (Biosera, France) in a humid atmosphere at 37°C and split every three days. Alternatively, the cells were maintained in physiological Plasmax medium as described previously [32].
2.2.1. HCV Cell Culture (HCVcc) System
HCV viral stock was prepared using a plasmid pJFH1 according to a standard protocol [34]. Huh7.5 cells were seeded in DMEM-F12 medium on 6- or 12-well plates or 6 cm dishes 24 h before infection at a density of 4x105 cells/well of a 6-well plate. When the cells reached 70% confluency, they were infected at 0.1 MOI. Four hours post-infected the medium was changed with a fresh one, and the cells were kept for 3-10 days (as specified in figure legends). The cells were harvested by scrapping. During analysis of antiviral activity, compounds were added 4 h before infection and 4 h post-infection (upon change of virion-containing medium).
2.2.2. HCV Replicons
Subgenomic HCV replicon corresponding to JFH1 isolate was obtained by synthesis of the viral RNA via in vitro transcription from a plasmid pSGR-JFH1, and its delivery to Huh7.5 cells by electroporation as described in [35]. The cells were maintained in DMEM-F12 cells supplemented with G418 (500 ng/ml).
To assess role of polyamines in HCV replication, the cells harboring replicon were seeded in DMEM with standard of dialyzed serum (Gemini Bio, West Sacramento, CA, USA) supplemented with 1 mM aminoguanidine and, if required, a mixture of spermine and spermidine (100 µM each).
2.3. Quantification of Metabolites
2.3.1. Polyamines
The cells were grown on 6-well plates. After removal of culture cell medium, cells washed up twice by 1 mL PBS at 0ºC. Then 0.1 ml mQ was added per well and plate was subjected to three cycles of freezing (-196 ºC) and thawing at +4º C. After the final thaw, samples were transferred to 0.5 ml polyethylene tubes. Each well was washed up with additional 0.1 ml mQ, combined with the crude cell lysates and subjected to ultrasound treatment for 3 min at 0-4ºC. Debris was removed by centrifugation for 10 min at 14,000 g. Total protein content in supernatants was measured using Pierce™ BCA Protein Assay Kit (Thermo Scientific, USA) prior precipitation of protein by addition of 60% perchloric acid to the final 3% concentration with subsequent clarifying by centrifugation for 10 min at 14,000 g. The clarified lysates were lyophilised and kept at -80ºC prior to analysis.
Polyamines were quantified by high-pressure liquid chromatography (HPLC) with precolumn derivatization with dansyl chloride similarly to described in [36]. The analysis was performed by dissolving dry samples in 100 µl of water saturated at 20ºC Na2CO3.. A different ratio mixture of 1,6-diaminohexane and 1,7-diaminoheptane were used as an internal standard and hydrazine hydrate (2 µl) applied to quench the dansylation reaction.
The solution of the dansylated polyamines in toluene (after 2-step extraction with 200 µl of toluene) was vacuum dried, the residue was dissolved in 200 µl of methanol and applied on a reversed phase column (Cosmosil C18-MS-II, 250 x 4.6 mm, 5 µm, 100 Å). The column was eluted (1 mL/min) with the gradient: 0 min—0% B; 5 min—0% B; 60 min—100% B; 65 min—100%B, 70 min—0% B, 75 min—0% B. System A—30% acetonitrile, 69.5% H2O, 0.5% propionic acid. System B—79.5% acetonitrile, 20% tetrahydrofuran, 0.5% propionic acid. Column temperature 40 ºC, pressure 80–120 bar, fluorescent detection: λex 340 nm, λem 530 nm (detector RF-20A, Shimadzu Scientific Instrument, Columbia, MD, USA).
2.3.2. Arginine and Proline
Arginine and proline levels of the cell extracts were measured by a nontargeted metabolic profiling approach based on high-resolution mass spectrometry (HRMS) as previously described [37]. Prior HRMS analysis, the cells extracts were vacuum dried using a Savant SpeedVac Concentrator SPD101 (Thermo, Switzerland), and resuspended in 200 µL 1:1 acetonitrile:methanol (v/v) and transferred to total recovery LCMS vials. Data processing and statistical analysis were performed as in (cite again here publication above). Metabolic features were annotated based on their mass accuracy, isotopic pattern and MS/MS fragments if available.
2.4. Enzyme Activity Assays
2.5. Western Blotting
The cells were grown on 6-well plates or 6-cm dishes, harvested by scrapping, lysed in RIPA buffer, and protein concentrations in lysates were measured by BCA kit (Thermo Fischer Scientific). Ten micrograms of total protein were applied on 10-12% sodium dodecyl sulfate-polyacrylamide gel, and immunoblotting was further carried out as described previously [40]. Primary murine antibodies to β-actin (ab3280, 1:500), ASS (ab124465, 1:2000), and ODC1 (ab193338, 1:1000) as well as rabbit antibodies to ASL1 (ab97370, 1:2000) were from Abcam (Cambridge, UK), rabbit antibodies to arginase I (9819s, 1:1000) and SSAT (61586, 1:200) from Cell Signaling Technology (Danvers, MA, USA), to SMOX (hpa047117, 1:1000) from Sigma (St. Louis, MO, USA). Murine antibodies to OAT (sc-374243, 1:500), PRODH (sc-376401, 1:200) and PYCR (sc-243722, 1:200) were from Santa Cruz Biotechnology, Inc. (TX, USA). Secondary HRP-conjugated goat antibodies to mouse IgG (sc-2005, 1:3000) and goat antibodies to rabbit IgG (sc-2004, 1:3000) were from Santa Cruz Biotechnology. All antibodies were used in 1% nonfat dry milk (BioRad, Hercules, CA, USA) in PBS with 0.05% Tween 20 (PBS-T).
2.6. Immunofluorescence
Huh7.5 cells were seeded on 24-well plates, infected as described above, fixed with methanol-acetone mixture and stained with rabbit sera raised to HCV NS3 and secondary FITC-labelled antibodies to rabbit immunoglobulins (F0382, Sigma) as reported previously for [41].
2.7. Reverse Transcription and Real-Time PCR (RT-qPCR)
Real-time and quantitative PCR were carried out as described in [35] with minor modifications. Briefly, total RNA was purified using Extract RNA Reagent (Evrogen J.S.C., Moscow, Russia) and treated with a recombinant RNAse-free DNAse (Roche, Basel, Switzerland) using specifications from the manufacturers. cDNA synthesis was carried out with RevertAid reverse transcriptase (Thermo Scientific, Rockford, IL, USA) and a random hexamer primer according to standard protocols. PCR was performed using primers listed in a Table S1 in a LightCycler 96 System (Roche, Switzerland). A standard reaction mixture (10 μL) contained cDNA equivalent to 10 ng total RNA, 0.8 μM of the respective primers, and qPCRmix-HS SYBR (Evrogen). The amplification conditions were 55 °C for 5 min, 95°C for 10 min, followed by 40 cycles each at 95°C for 10 s and 57°C for 1 min (signal collection temperature). β-Glucoronidase (GUS) was used as a housekeeping gene. The results were analysed by ∆∆Ct approach.
2.8. Statistical Analysis
All data are presented as means ± standard error of means (S.E.M.). Statistical significance was determined by paired two-tailed t-test, Analysis of variance (ANOVA) with Tukey post-hoc test or Dunnett’s test using Graphpad Prism (7.0). A p-value <0.05 was considered as statistically significant, if not stated otherwise.
3. Results
3.1. Hepatitis C Virus Perturbs Metabolism of Biogenic Polyamines
Previously we showed that overexpression of several HCV enzymes altered expression of the key polyamine-metabolizing enzymes, whereas the long-lasting replication of its genome in human hepatoma Huh7 cells (i.e. replicon system) was accompanied by decreased expression of ODC and SSAT and reduced polyamine content [29]. In the present study we aimed to unveil the imprint of the virus on polyamine metabolism in the infectious cell culture model (HCVcc). Huh7.5 cells were infected with HCV at a multiplicity of infection (MOI) of 0.1, and mRNA levels of most polyamine-metabolizing enzymes and regulatory proteins were assessed 3-10 days post-infection. The results shown on Figure 1B,C, clearly show that HCV moderately upregulates transcription of both biosynthetic and catabolic enzymes, although for some of them analysis revealed only a trend towards statistical significance. As ornithine decarboxylase and spermidine/spermine-N1-acetyltransferase are regulated post-transcriptionally, levels of polyamine-metabolizing enzymes were also analysed by western blotting (ODC, SMOX) or by measuring intracellular activity in cell lysates (SSAT). HCV infection decreased levels of these enzymes, despite activation at the transcriptional level (Figure 1D,E). Quantification of polyamines revealed that HCV induces a time-dependent decrease of spermidine levels but an increase in spermine concentrations in a period up to 6 days (Figure 1F). This correlates with down-regulation of polyamine biosynthesis and spermine catabolism. Levels of putrescine were below the detection limit (0.05 nmol/mg total protein). However, over longer infection periods, spermine levels were also decreased (Figure 1F), likely due to exhaustion of their total levels. These data clearly demonstrate that HCV infection triggers a shift of the spermine-to-spermidine ratio during spread of the infection. However, at late stages of infection spermine accumulation normalized to a level slightly below that observed in uninfected cells.
3.2. Hepatitis C Virus Down-Regulates Expression of Arginase I
Since polyamines are synthesized from ornithine, a non-proteinogenic amino acid of the urea cycle [22], our next step was to study possible effects of the virus on expression of urea cycle enzymes. Expression of arginase (ARG) I and II, ornithine transcarbamylase 1 (OTC), argininosuccinate synthase (ASS), and argininosuccinate lyase 1 (ASL1) as well as of carbamoylphosphate synthase (CPS1) was evaluated by RT-qPCR and/or western blotting. This revealed that HCV down-regulates transcription of arginase 1 by several fold. Levels of extrahepatic arginase (ARGII) mRNA were increased at 6 dpi (Figure 2A) but absolute levels of ARGII remained below the levels of ARGI mRNA. In line with this, its expression was undetectable at the protein level by western blotting (not shown). Expression of other genes did not show any significant changes. Expression of OTC in this cell line was very low and even undetectable in some samples (Figure 2A), whereas other genes were expressed at much higher levels. Next, we assessed levels of arginine. Mass-spectrometry analysis indicated that HCV infection led to a time-dependent increase of the arginine pool in cells with suppressed ARGI expression.
To address the question if alteration of the urea cycle is a transient feature in HCV infection and occurs only shortly after virus entry, we assessed changes in expression of urea cycle enzymes in cells harbouring subgenomic HCV replicons. Indeed, HCV also decreased ARGI levels in this model of chronic infection (Figure 3D,E). In this model we also showed decreased expression of ASS and increased accumulation of ASL1. However, these latter changes were not observed in the HCVcc system.
Recently Tardito’s group reported that classical DMEM-F12 medium, that is often used in HCV research, has a profound imprint on cell metabolism [33]. Furthermore, triple-negative breast cancer cell lines in DMEM-F12 medium exhibit a reversed urea cycle, as arginine, present at a non-physiologically high concentrations in DMEM-F12, is converted not into ornithine via arginase but into argininosuccinate via ASL1. Therefore, we verified our data using a physiological plasma-resembling Plasmax medium that has normal levels of arginine. Indeed, a subgenomic HCV replicon was found to suppress expression of arginase I independently of the culture medium used (Figure 2F).
3.3. Hepatitis C Virus Up-Regulates Expression of Proline Dehydrogenase and Decreases Intracellular Level of Proline
Next, we evaluated the effect of HCV on the metabolism of proline, as its catabolism yields Δ1-pyrrolidine-5-carboxylate (P5C) that, in its turn, can be converted into ornithine by ornithine aminotransferase (OAT). The initial screening of transcription rates for genes involved in proline biosynthesis and catabolism identified proline dehydrogenase (PRODH, proline oxidase) as a gene whose expression is upregulated by the virus (Figure 3A). The levels of PRODH were elevated at the protein level as well (Figure 3B). The expression of pyrroline-5-carboxylate reductase 1 that catalyses the reverse reaction was unaltered (Figure 3A,B). Moderate changes were also observed for OAT at 3 dpi. The induction of the proline-catabolizing enzyme resulted in a decrease in intracellular proline concentration (Figure 3C). An even more pronounced induction of PRODH was observed in Huh7.5 cells harbouring the subgenomic virus replicon showing that the changes in proline catabolism are induced by the nonstructural viral proteins and suggesting that these changes also happen during long-lasting HCV infection (Figure 3D,E). Finally, as for the urea cycle, these changes occurred in both DMEM-F12 and Plasmax media (Figure 3F).
Since PRODH induction led to a decrease in intracellular proline concentration, our next step was to access the impact of these changes on the replication of the virus. Proline levels in culture media are usually within the 0.15-0.20 mM range [42]. We therefore elevated its concentration approximately by ten-fold, i.e. to 2.5 mM. However, this led only to a moderate (ap. 30%) reduction of replication levels in the subgenomic replicon model (Figure 3G), indicating that reduction of the proline pool in infected cells is dispensable for the virus. Similarly, a reduction in HCV replication levels was also achieved by treatment with the PRODH inhibitor (S)-(+)-5-oxo-2 tetrahydrofurancarboxylic acid (THFA) (Figure 3H).
3.4. HCV Does Not Affect Expression of the Genes that Channel Glutamate into Proline/Ornithine
Δ1-Pyrrolidine-5-carboxylate can be synthesized not only from proline but also from glutamate [43]. So, our next step was to assess the levels of ALDH18A1 that encodes Δ1-pyrrolidine-5-carboxylate synthase, an enzyme converting Glu into P5C, and ALDH4A1 that mediates the reverse reaction. However, RT-qPCR analysis showed no pronounced and statistically significant changes in transcription of their genes in response to HCV infection in HCVcc (Figure 3A) and subgenomic replicon (Figure 3D,F) systems.
3.5. Inhibition of Polyamine Biosynthesis or Catabolism Suppresses HCV Replication
Our final step was to assess importance of biogenic polyamines for HCV replication. It was performed by evaluating compounds that either inhibit or induce the key enzyme of polyamine metabolism as potential anti-HCV agents. These compounds included the inhibitors of ornithine decarboxylase (difluoromethylornithine, DFMO; 3-aminooxy-1-aminopropane, APA), polyamine oxidases (MDL72.527) or AMD (SAM486A), potent SSAT inducers N1,N11-diethylnorspermine (DENSpm) and N1-ethyl-N11-cyclopropylnorspermine (CPENS), inhibitors of eIF5a hypusination (a spermidine-dependent post-translational modification) GC7, deferiprone and ciclopirox. In addition, we evaluated N-(3-aminopropyl)cyclohexylamine (APCHA) and trans-4-methyl-cyclohecylamine (4MCHA) - the inhibitors of spermine and spermidine synthases. As additional control, we analysed another bis-alkylated spermine - N1-ethyl-N11-cycloheptylnorspermine (CHENS) that is a much weaker SSAT inducer [44]. Finally, we also tested L-canaline that inhibits ornithine aminotransferase, the enzyme that catalyses the conversion of P5C into ornithine. Noteworthy that the compounds were tested in non-toxic concentrations (Figure S1).
Initially these compounds were tested in Huh7.5 cells harboring the full-length HCV replicon (Figure 4A,E) with further evaluation in the HCVcc system (Figure 4C,D). Quantification of RNA levels revealed that the inducers of SSAT (Figure 4A), inhibitors of polyamine synthases and ornithine aminotransferase (Figure 4D,E) did not affect HCV replication. The hypusination pathway also seemed to be dispensable for HCV replication, as its classical selective inhibitor GC7 did not alter HCV RNA levels in either system, and other, less selective, inhibitors had opposite impact in HCVcc and replicon models (Figure 4D,E).
At the same time significant anti-HCV activity was displayed by DFMO, APA and SAM486A as well as MDL72.527, drugs that target polyamine biosynthesis and catabolism, respectively (Figure 4A,C,E). To analyse their mode of antiviral action, polyamine levels were quantified in treated naïve cells and the cells with HCV replicon. As can be seen from Table 1, treatment of naïve Huh7.5 cells with 20 µM DENSpm resulted in a 6.5-fold upregulation of SSAT transcription and a concomitant 2.8-fold decrease in spermine levels. Similar changes were observed in the replicon-harboring cells. The increase in SSAT expression was more pronounced in replicon-harboring cells compared to naïve cells due to lower gene expression in the untreated cells, but the total activity level became comparable to the level in the treated naïve cells. The treatment of the naïve Huh7.5 cells with 1 mM DFMO suppressed ODC activity just by two-fold with no changes in the levels of spermine or spermidine. Unfortunately, putrescine content in the hepatic cells was close to the limit of detection and quantification was not possible in all samples. Strikingly, in the cells with the HCV replicon, DFMO increased content of spermine and spermidine by 5-7 fold. Similar changes were seen for MDL72.527 that also exhibited antiviral activity. Since reduced HCV replication was observed in the samples with elevated polyamine content, the next step was to verify the influence of spermine and spermidine on HCV replication using other approaches. HCV RNA levels were evaluated in replicon-harboring cells maintained either in a standard DMEM supplemented with both polyamines in a combination with aminoguanidine, an inhibitor or their degradation by serum aminooxidases, or in a medium with a dialyzed serum (dDMEM), i.e. depleted of polar metabolites including polyamines. Indeed, in the presence of dialyzed serum HCV replication was elevated, whereas exogenous polyamines suppressed it (Figure 4B). Thus, high polyamine content in infected cells suppresses viral replication, and HCV reduces spermine and spermidine pools to alleviate their negative effect.
4. Discussion
Polyamines are ubiquitous compounds critical for cell growth and differentiation. Changes in their metabolism are associated with development of proliferative and metabolic disorders such as tumorigenesis and autoimmune pathologies (i.e. psoriasis) [45,46]. However, significance of spermine and spermidine for viral infections until recently was studied less extensively. Several viruses including Kaposi’s sarcoma-associated virus (KSAV) [47], Epstein-Barr virus (EBV) [48], Dengue virus [49], were shown to suppress expression of SSAT at transcriptional or translational levels. KSAV also induces expression of ODC, although only at latent stages of replication in 2D culture [50] or in 3D models [47]. A similar, although transient, increase in ODC was described in 3D organoids infected with KSAV [47]. As a result, at least some of these infections increase polyamine content in infected cells. In contrast, SARS-CoV-2 [51], porcine reproductive and respiratory syndrome virus (PRRSV) [52], and porcine endemic diarrhoea virus (PEDV) [53] upregulate expression of SSAT and thus decrease polyamine levels in infected cells. Exhaustion of spermine and spermidine levels due to lowered expression of ODC is also a feature of lytic KSAV [50] and Coxsackie virus B3 (CVB3) [54] infections. So, our data expand the list of viral pathogens that deregulate expression of polyamine-metabolising enzymes and decrease polyamine content.
Polyamines are important for replication of various viruses. In recent years the Mounce group demonstrated that polyamines ensure infectivity of virions either by direct incorporation into the virus particle [55] or by incorporation of cholesterol [56] thus conferring binding of virions to the cell surface and concomitant virus entry [55,57,58]. During polyamine exhaustion, non-infectious viral particles can be formed, as shown for Bunyaviruses [59]. So, it is not surprising that polyamine biosynthesis inhibitors such as DFMO or SAM486A exhibit activity towards members of the Filo- [60], Herpes- [50], Entero- [61], Picorna- [61], Alpha- [62], Corona- [57,61], Flavi- [61], Bunya- [61], Rhabdo- [61], and Hepadnaviridae [63] families. However, in all these cases their mechanism of antiviral action involved exhaustion of polyamine level in cells. In this study, to our great surprise inhibitory activity of the compounds was accompanied by increase in polyamine pool. The mechanism by which polyamines suppress HCV replication are not fully understood. On one hand, spermine and spermidine stimulate RNA polymerase activity of the HCV NS5B protein, however at the same time they inhibit RNA unwinding by the viral helicase [64].
Suppression of enteroviruses was earlier also shown for DENSpm [65]. Again, our study shows that these inhibitors also demonstrate anti-HCV activity and add APA to the list of antiviral agents. Moreover, we show that HCV replication can be inhibited by the inhibitor of polyamine oxidases MDL72.527 that was previously studied only for Ebola virus [66].
Polyamines also support replication of viruses by ensuring the hypusination of eIF5A. KSAV [47,50], Coxsackie virus B3 [67], vesicular stomatitis (VSV), influenza A, Zika and Chikungunya viruses [68] increase levels of hypusinated eIF5A. This factor enhances translation of the Ebola genome [69] and HIV transcription [70]. Inhibitors of deoxyhypusine synthase and deoxyhypusine hydrolase also exhibit a wide spectrum of antiviral activity [50,57,60,68,70]. Interestingly, neither of them had an anti-viral effect on HCV in our study.
One of most unexpected results of the study is the nonstandard response of Huh7.5 cells and especially the cells harbouring HCV replicon to the treatment with compounds that target polyamine-metabolizing enzymes such as DENSpm and especially DFMO. First, Huh7.5 cells demonstrated high resistance to DENSpm: this compound caused only moderate reduction of spermine and spermidine levels upon treatment, and the cells remained viable even upon treatment with 300 µM of the drug (Figure S1). Second, the cells also demonstrated resistance to DFMO which is an irreversible inhibitor of ODC: at subtoxic concentrations this drug decreased intracellular ODC activity by two-fold only. Third, a treatment of cells containing the HCV replicon led to an unprecedented elevation of the levels of both spermine and spermidine. It could be speculated that the effect can be due either to enhanced polyamine derived from serum import from culture medium or by some nonstandard feedback mechanism by which the cells counteract polyamine depletion. Such an effect was previously described by J. Janne’s group that raised DFMO-resistant human and murine leukaemia cell lines by maintaining cells in the presence of subtoxic drug concentrations and then observed either multiplication of the ODC1 gene or its enhanced transcription [71]. In the presence of DFMO this cell line enhanced ODC expression to counteract inactivation of the enzyme by the drug. Although we have not quantified ODC1 copy numbers in Huh7.5 cells with the HCV replicon, it cannot be excluded that ODC1 multiplication occurs during selection of the cells possibly mediated via HCV-enhanced ROS production [29,66].
Here we show that HCV decreases expression of ARGI and thus causes accumulation of its substrate arginine in the infected Huh7.5 cells. So far, the data on expression of Arginase 1 in the context of HCV infection have been discrepant. Several groups did not find any changes in Arg1 expression in liver samples of chronic hepatitis C virus patients without signs of steatosis [72,73], while the others observed increased staining of the enzyme in biopsy sections [74]. However, Arg1 downregulation was also reported for Huh7.5 cells harbouring the full-length HCV replicon [75]. Decreased expression of Arg1 and concomitant accumulation of arginine could be the feature of hepatocarcinoma cells during non-alcoholic steatohepatitis and metabolic syndrome [76]. So, we cannot rule out that down-regulation of Arg1 during HCV expression can contribute to viral pathogenesis.
Cell metabolism is affected not only by the virus but by the culture medium. Almost all studies are performed in cell lines maintained in classical media (such as MEM, DMEM, RPMI, F12 etc) formulated in 1950s or early 1960s [42]. Their recipes were to ensure rapid growth of cell biomass and longer period between replacement of conditioned medium with a fresh one. However, several lines of evidence have appeared during last decade that metabolism of cells maintained in classical medium sometimes does not mimic the processes in vivo (summarised in [42]). One of the most pronounced differences is addiction of various tumour cell lines on glutaminolysis when grown in DMEM or other “old” medium with no noticeable effect of antitumor/antiproliferative activity of a glutaminase 1 inhibitors in vivo or in 3D cell cultures [77,78]. The other example is an inverted urea cycle in cells maintained in DMEM-F12 medium like the one used in our study: arginine is converted not into ornithine or citrulline but into argininosuccinate, albeit with different efficiency in different cell lines [33]. Therefore, several groups recently developed media (HPLM, Plasmax) that resemble human blood plasma and showed “normalization” of the urea cycle [79,80]. So, we used Plasmax medium to verify the changes in cell metabolism in HCV-infected cells. Indeed, the key event, i.e. the suppressed expression of arginase 1, was consistent in case of both media.
Proline biosynthesis and catabolism may be not only the independent processes that regulate steady-state proline concentrations but also form a proline cycle [81]. It is formed by conversion of the amino acid into Δ1-pyrrolidine-5-caboxylate (P5C) in mitochondria by PRODH, export of this metabolite into cytoplasm with subsequent reverse transformation into Pro by Δ1-pyrrolidine-5-carboxylate reductase 3 (P5CR3, P5CRL). This cycle both feeds oxidative phosphorylation in mitochondria by providing electrons from the PRODH-reduced flavin (FADH2) and supports nucleotide biosynthesis via oxidation of NADPH into NADP+ by P5CRL. So, enhanced metabolic flux of proline and P5C may be a mechanism of activation of other metabolic pathways. However, upregulation of PRODH in the HCV-infected cells is not accompanied by changes in expression of proline biosynthetic enzymes, and of P5CRL in particular, thus excluding importance of proline cycle in regulation of the pentose phosphate pathway during HCV infection. Of note, PRODH expression is positively regulated by AMPK, p53, and PPARγ, and negatively by cMyc transcription factors. Since HCV is known to activate cMyc [18,82] and suppress p53 [83] and AMPK signalling [79], they are unlikely to mediate upregulation of PRODH during the infection. This points to PPARγ as the likely regulator of PRODH expression in HCV-infected cells.
Polyamines are synthesized from a non-proteinogenic amino acid ornithine which in turn can be produced either from arginine in the urea cycle or from glutamic semialdehyde (GSA) by ornithine aminotransferase (OAT). The latter is formed either via a non-enzymatic isomerization of P5C (i.e. from proline catabolism) or from glutamate by Δ1-pyrrolidine-5-carboxylate synthase (encoded by the ALDH18A1 gene) (i.e. from glutaminolysis). Few papers suggest that neoplastic transformation may switch an origin of ornithine from arginine to GSA, although these data were obtained from non-liver cells [84]. Since HCV activates both glutaminolysis [18] and proline catabolism and suppresses conversion of arginine into ornithine (this study), it is tempting to speculate that this virus also reprograms routes of ornithine production. Although origins of ornithine were not accessed using flux analysis by mass-spectrometry using 13C-labelled ornithine and glutamate/proline, we used an inhibitor of OAT – L-canaline. However, this compound only slightly affected viability of HCV-infected cells (Figure S1) and did not affect HCV replication levels thus not supporting our assumption.
Arginine:glycine amidinotransferase (GATM, AGAT) is an enzyme that catalyses an alternative arginine-to-ornithine conversion pathway. Its second product, guanidinoacetate is a precursor for the biosynthesis of creatine and creatinine by GAMT. It is generally accepted that the first reaction takes place in kidney, whereas the second – in the liver. However, GATM mRNA levels in the liver in hepatocarcinoma Huh7.5 cells as well as in nontumor hepatocyte-like HepaRG cells are at least six-fold higher than those of arginase 1 [85]. So, we cannot exclude that in HCV-infected cells ornithine is synthesized via a GATM-mediated reaction.
Agmatinase is the least studied enzyme of polyamine biosynthesis in mammalian cells. In prokaryotic and plant and fungi cells agmatinase produces ornithine from agmatine generated from arginine by arginine decarboxylase (ADC). Since mammals were thought not to encode ADC in its genome, almost no attempts to identify arginase in mammalian cells were made. In 2002 this gene was identified in HepG2 cells and upregulation of its transcription in the context of HBV infection was reported [86]. Later, ADC was also discovered in mammalian brain cells [87]. However, biological significance of agmatinase in mammals remains obscure, since the its substrate agmatine can originate either from gut biota or from food, as ADC has very low expression levels in a majority of tissues. Nevertheless, we included this gene into our analysis and demonstrated absence of its regulation in HCV-infected cells.
5. Conclusions
To sum up, we have demonstrated that HCV perturbs several metabolic pathways including the urea cycle, polyamine and proline metabolism. Biosynthesis and degradation of spermine and spermidine are critical for the virus, as pharmacological inhibitors of the respective enzymes display pronounced antiviral activity. Replenishment of proline levels also suppressed HCV replication. However, further studies are needed to unveil how these changes impact the development of the pathology associated with chronic HCV infection.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: primers used in the study; Figure S1: Cell toxicity of the polyamine metabolism inhibitors.
Author Contributions
Conceptualization, A.K., S.K, B.B. and A.I.; methodology, C.B., A.K., S.K, B.B. and A.I.; formal analysis, B.B. and A.I.; investigation, N.Z., O.K., O.S., J.M., S.D., D.Y., V.V.-E., L.M., B.G., O.I., I.K., M.G., B.R. and A.I.; resources, C.B., A.K., S.K., B.B. and A.I.; data curation, N.Z., O.K., O.S., J.M., D.Y., L.M., B.G., O.I., I.K., C.B., B.R., B.B. and A.I., ; writing—original draft preparation, O.S., B.B. and A.I.; writing—review and editing, N.Z., O.K., O.S., J.M., S.D., D.Y., V.V.-E., L.M., B.G., O.I., I.K., M.G., C.B., B.R., A.K., S.K., B.B. and A.I.; visualization, A.I.; supervision, B.B. and A.I.; project administration, B.B. and A.I..; funding acquisition, S.K., B.B. and A.I. All authors have read and agreed to the published version of the manuscript.
Funding
Most part of the study was supported by the Russian Science Foundation (project 19-74-10086), while infectious experiments in BSL-3 unit were supported by The DevWeCan French Laboratories of Excellence Network (Labex, Grant ANR-10-LABX-61) and PHC Kolmogorov. Interaction between the authors was supported by the 111 Project, Grant No. B12003.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The authors are immensely grateful to Dr. Tuomo A. Keinanen (University of Eastern Finland) for technical help in polyamine quantification.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Moon, A.M.; Singal, A.G.; Tapper, E.B. Contemporary Epidemiology of Chronic Liver Disease and Cirrhosis. Clin Gastroenterol Hepatol 2020, 18, 2650–2666. [Google Scholar] [CrossRef]
- Hepatitis, C.; Fact sheet. Updated 24.06.2022. Available online: https://www.who.int/en/news-room/fact-sheets/detail/hepatitis-c (accessed on 03.02.2023).
- Westbrook, R.H.; Dusheiko, G. Natural history of hepatitis C. Journal of hepatology 2014, 61, S58–68. [Google Scholar] [CrossRef]
- Lingala, S.; Ghany, M.G. Natural History of Hepatitis C. Gastroenterology clinics of North America 2015, 44, 717–734. [Google Scholar] [CrossRef]
- Falade-Nwulia, O.; Suarez-Cuervo, C.; Nelson, D.R.; Fried, M.W.; Segal, J.B.; Sulkowski, M.S. Oral Direct-Acting Agent Therapy for Hepatitis C Virus Infection: A Systematic Review. Ann Intern Med 2017, 166, 637–648. [Google Scholar] [CrossRef]
- Manns, M.P.; Maasoumy, B. Breakthroughs in hepatitis C research: from discovery to cure. Nature reviews. Gastroenterology & hepatology 2022, 19, 533–550. [Google Scholar] [CrossRef]
- Cooke, G.S.; Andrieux-Meyer, I.; Applegate, T.L.; Atun, R.; Burry, J.R.; Cheinquer, H.; Dusheiko, G.; Feld, J.J.; Gore, C.; Griswold, M.G.; et al. Accelerating the elimination of viral hepatitis: a Lancet Gastroenterology & Hepatology Commission. The lancet. Gastroenterology & hepatology 2019, 4, 135–184. [Google Scholar] [CrossRef]
- Negro, F. Residual risk of liver disease after hepatitis C virus eradication. Journal of hepatology 2021, 74, 952–963. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Murata, M.; Yoshida, K.; Sekimoto, G.; Uemura, Y.; Sakaida, N.; Kaibori, M.; Kamiyama, Y.; Nishizawa, M.; Fujisawa, J.; et al. Chronic inflammation associated with hepatitis C virus infection perturbs hepatic transforming growth factor beta signaling, promoting cirrhosis and hepatocellular carcinoma. Hepatology 2007, 46, 48–57. [Google Scholar] [CrossRef]
- Fabregat, I.; Caballero-Diaz, D. Transforming Growth Factor-beta-Induced Cell Plasticity in Liver Fibrosis and Hepatocarcinogenesis. Front Oncol 2018, 8, 357. [Google Scholar] [CrossRef]
- Virzi, A.; Roca Suarez, A.A.; Baumert, T.F.; Lupberger, J. Oncogenic Signaling Induced by HCV Infection. Viruses 2018, 10. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Tyurina, D.A.; Ivanova, O.N.; Kochetkov, S.N.; Bartosch, B.; Isaguliants, M.G. Oxidative stress, a trigger of hepatitis C and B virus-induced liver carcinogenesis. Oncotarget 2017, 8, 3895–3932. [Google Scholar] [CrossRef]
- Moriya, K.; Nakagawa, K.; Santa, T.; Shintani, Y.; Fujie, H.; Miyoshi, H.; Tsutsumi, T.; Miyazawa, T.; Ishibashi, K.; Horie, T.; et al. Oxidative stress in the absence of inflammation in a mouse model for hepatitis C virus-associated hepatocarcinogenesis. Cancer Res 2001, 61, 4365–4370. [Google Scholar]
- Levy, P.; Bartosch, B. Metabolic reprogramming: a hallmark of viral oncogenesis. Oncogene 2016, 35, 4155–4164. [Google Scholar] [CrossRef]
- Lupberger, J.; Croonenborghs, T.; Roca Suarez, A.A.; Van Renne, N.; Juhling, F.; Oudot, M.A.; Virzi, A.; Bandiera, S.; Jamey, C.; Meszaros, G.; et al. Combined Analysis of Metabolomes, Proteomes, and Transcriptomes of Hepatitis C Virus-Infected Cells and Liver to Identify Pathways Associated With Disease Development. Gastroenterology 2019, 157, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Ramiere, C.; Rodriguez, J.; Enache, L.S.; Lotteau, V.; Andre, P.; Diaz, O. Activity of Hexokinase Is Increased by Its Interaction with Hepatitis C Virus Protein NS5A. J Virol 2014, 88, 3246–3254. [Google Scholar] [CrossRef]
- Diamond, D.L.; Syder, A.J.; Jacobs, J.M.; Sorensen, C.M.; Walters, K.A.; Proll, S.C.; McDermott, J.E.; Gritsenko, M.A.; Zhang, Q.; Zhao, R.; et al. Temporal proteome and lipidome profiles reveal hepatitis C virus-associated reprogramming of hepatocellular metabolism and bioenergetics. PLoS pathogens 2010, 6, e1000719. [Google Scholar] [CrossRef] [PubMed]
- Levy, P.L.; Duponchel, S.; Eischeid, H.; Molle, J.; Michelet, M.; Diserens, G.; Vermathen, M.; Vermathen, P.; Dufour, J.F.; Dienes, H.P.; et al. Hepatitis C virus infection triggers a tumor-like glutamine metabolism. Hepatology 2016. [Google Scholar] [CrossRef]
- Roe, B.; Kensicki, E.; Mohney, R.; Hall, W.W. Metabolomic profile of hepatitis C virus-infected hepatocytes. PLoS One 2011, 6, e23641. [Google Scholar] [CrossRef]
- Gewaid, H.; Aoyagi, H.; Arita, M.; Watashi, K.; Suzuki, R.; Sakai, S.; Kumagai, K.; Yamaji, T.; Fukasawa, M.; Kato, F.; et al. Sphingomyelin Is Essential for the Structure and Function of the Double-Membrane Vesicles in Hepatitis C Virus RNA Replication Factories. J Virol 2020, 94. [Google Scholar] [CrossRef]
- Pegg, A.E. Mammalian polyamine metabolism and function. IUBMB Life 2009, 61, 880–894. [Google Scholar] [CrossRef]
- Smirnova, O.A.; Bartosch, B.; Zakirova, N.F.; Kochetkov, S.N.; Ivanov, A.V. Polyamine Metabolism and Oxidative Protein Folding in the ER as ROS-Producing Systems Neglected in Virology. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Ramsey, I.S.; Delling, M.; Clapham, D.E. An introduction to TRP channels. Annual review of physiology 2006, 68, 619–647. [Google Scholar] [CrossRef] [PubMed]
- Casero, R.A., Jr.; Murray Stewart, T.; Pegg, A.E. Polyamine metabolism and cancer: treatments, challenges and opportunities. Nat Rev Cancer 2018, 18, 681–695. [Google Scholar] [CrossRef]
- Chaturvedi, R.; Asim, M.; Romero-Gallo, J.; Barry, D.P.; Hoge, S.; de Sablet, T.; Delgado, A.G.; Wroblewski, L.E.; Piazuelo, M.B.; Yan, F.; et al. Spermine oxidase mediates the gastric cancer risk associated with Helicobacter pylori CagA. Gastroenterology 2011, 141, 1696–1708. [Google Scholar] [CrossRef]
- Goodwin, A.C.; Destefano Shields, C.E.; Wu, S.; Huso, D.L.; Wu, X.; Murray-Stewart, T.R.; Hacker-Prietz, A.; Rabizadeh, S.; Woster, P.M.; Sears, C.L.; et al. Polyamine catabolism contributes to enterotoxigenic Bacteroides fragilis-induced colon tumorigenesis. Proceedings of the National Academy of Sciences of the United States of America 2011, 108, 15354–15359. [Google Scholar] [CrossRef]
- Evageliou, N.F.; Haber, M.; Vu, A.; Laetsch, T.W.; Murray, J.; Gamble, L.D.; Cheng, N.C.; Liu, K.; Reese, M.; Corrigan, K.A.; et al. Polyamine Antagonist Therapies Inhibit Neuroblastoma Initiation and Progression. Clin Cancer Res 2016, 22, 4391–4404. [Google Scholar] [CrossRef] [PubMed]
- Saulnier Sholler, G.L.; Gerner, E.W.; Bergendahl, G.; MacArthur, R.B.; VanderWerff, A.; Ashikaga, T.; Bond, J.P.; Ferguson, W.; Roberts, W.; Wada, R.K.; et al. A Phase I Trial of DFMO Targeting Polyamine Addiction in Patients with Relapsed/Refractory Neuroblastoma. PLoS One 2015, 10, e0127246. [Google Scholar] [CrossRef]
- Smirnova, O.A.; A. , K.T.; Ivanova, O.N.; Hyvonen, M.T.; Khomutov, A.R.; Kochetkov, S.N.; Bartosch, B.; Ivanov, A.V. Hepatitis C virus alters metabolism of biogenic polyamines by affecting expression of key enzymes of their metabolism. Biochem Biophys Res Commun 2017. [Google Scholar] [CrossRef]
- Cruz-Pulido, Y.E.; Mounce, B.C. Good cop, bad cop: Polyamines play both sides in host immunity and viral replication. Semin Cell Dev Biol 2023, 146, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Khomutov, A.R.; Vepsalainen, J.J.; Shvetsov, A.S.; Hyvonen, T.; Keinanen, T.A.; Pustobaev, V.N.; Eloranta, T.O.; Khomutov, R.M. Synthesis of hydroxylamine analogues of polyamines. Tetrahedron 1996, 52, 13751–13766. [Google Scholar] [CrossRef]
- Golikov, M.V.; Karpenko, I.L.; Lipatova, A.V.; Ivanova, O.N.; Fedyakina, I.T.; Larichev, V.F.; Zakirova, N.F.; Leonova, O.G.; Popenko, V.I.; Bartosch, B.; et al. Cultivation of Cells in a Physiological Plasmax Medium Increases Mitochondrial Respiratory Capacity and Reduces Replication Levels of RNA Viruses. Antioxidants (Basel) 2022, 11, 97. [Google Scholar] [CrossRef] [PubMed]
- Vande Voorde, J.; Ackermann, T.; Pfetzer, N.; Sumpton, D.; Mackay, G.; Kalna, G.; Nixon, C.; Blyth, K.; Gottlieb, E.; Tardito, S. Improving the metabolic fidelity of cancer models with a physiological cell culture medium. Sci Adv 2019, 5, eaau7314. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Date, T.; Murayama, A.; Morikawa, K.; Akazawa, D.; Wakita, T. Cell culture and infection system for hepatitis C virus. Nat Protoc 2006, 1, 2334–2339. [Google Scholar] [CrossRef]
- Kukhanova, M.K.; Tunitskaya, V.L.; Smirnova, O.A.; Khomich, O.A.; Zakirova, N.F.; Ivanova, O.N.; Ziganshin, R.; Bartosch, B.; Kochetkov, S.N.; Ivanov, A.V. Hepatitis C Virus RNA-Dependent RNA Polymerase Is Regulated by Cysteine S-Glutathionylation. Oxidative medicine and cellular longevity 2019, 2019, 3196140. [Google Scholar] [CrossRef] [PubMed]
- Hyvonen, M.T.; Keinanen, T.A.; Nuraeva, G.K.; Yanvarev, D.V.; Khomutov, M.; Khurs, E.N.; Kochetkov, S.N.; Vepsalainen, J.; Zhgun, A.A.; Khomutov, A.R. Hydroxylamine Analogue of Agmatine: Magic Bullet for Arginine Decarboxylase. Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Rindlisbacher, B.; Schmid, C.; Geiser, T.; Bovet, C.; Funke-Chambour, M. Serum metabolic profiling identified a distinct metabolic signature in patients with idiopathic pulmonary fibrosis - a potential biomarker role for LysoPC. Respir Res 2018, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.R. Calf liver nuclear N-acetyltransferases. Purification and properties of two enzymes with both spermidine acetyltransferase and histone acetyltransferase activities. J Biol Chem 1978, 253, 233–237. [Google Scholar] [CrossRef]
- Janne, J.; Williams-Ashman, H.G. On the purification of L-ornithine decarboxylase from rat prostate and effects of thiol compounds on the enzyme. J Biol Chem 1971, 246, 1725–1732. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Smirnova, O.A.; Ivanova, O.N.; Masalova, O.V.; Kochetkov, S.N.; Isaguliants, M.G. Hepatitis C virus proteins activate NRF2/ARE pathway by distinct ROS-dependent and independent mechanisms in HUH7 cells. PLoS One 2011, 6, e24957. [Google Scholar] [CrossRef]
- Tunitskaya, V.L.; Eliseeva, O.V.; Valuev-Elliston, V.T.; Tyurina, D.A.; Zakirova, N.F.; Khomich, O.A.; Kalis, M.; Latyshev, O.E.; Starodubova, E.S.; Ivanova, O.N.; et al. Prokaryotic Expression, Purification and Immunogenicity in Rabbits of the Small Antigen of Hepatitis Delta Virus. Int J Mol Sci 2016, 17. [Google Scholar] [CrossRef]
- Golikov, M.V.; Bartosch, B.; Smirnova, O.A.; Ivanova, O.N.; Ivanov, A.V. Plasma-Like Culture Medium for the Study of Viruses. mBio 2023, 14, e0203522. [Google Scholar] [CrossRef] [PubMed]
- Chalecka, M.; Kazberuk, A.; Palka, J.; Surazynski, A. P5C as an Interface of Proline Interconvertible Amino Acids and Its Role in Regulation of Cell Survival and Apoptosis. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Lawson, K.R.; Marek, S.; Linehan, J.A.; Woster, P.M.; Casero, R.A., Jr.; Payne, C.M.; Gerner, E.W. Detoxification of the polyamine analogue N1-ethyl-N11-[(cycloheptyl)methy]-4,8-diazaundecane (CHENSpm) by polyamine oxidase. Clin Cancer Res 2002, 8, 1241–1247. [Google Scholar] [PubMed]
- Lou, F.; Sun, Y.; Xu, Z.; Niu, L.; Wang, Z.; Deng, S.; Liu, Z.; Zhou, H.; Bai, J.; Yin, Q.; et al. Excessive Polyamine Generation in Keratinocytes Promotes Self-RNA Sensing by Dendritic Cells in Psoriasis. Immunity 2020, 53, 204–216. [Google Scholar] [CrossRef]
- Wagner, A.; Wang, C.; Fessler, J.; DeTomaso, D.; Avila-Pacheco, J.; Kaminski, J.; Zaghouani, S.; Christian, E.; Thakore, P.; Schellhaass, B.; et al. Metabolic modeling of single Th17 cells reveals regulators of autoimmunity. Cell 2021, 184, 4168–4185. [Google Scholar] [CrossRef] [PubMed]
- Choi, U.Y.; Lee, J.J.; Park, A.; Jung, K.L.; Lee, S.A.; Choi, Y.J.; Lee, H.R.; Lai, C.J.; Eoh, H.; Jung, J.U. Herpesvirus-induced spermidine synthesis and eIF5A hypusination for viral episomal maintenance. Cell Rep 2022, 40, 111234. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Gan, Y.J.; Davis, T.O.; Scott, R.S. Downregulation of the polyamine regulator spermidine/spermine N(1)-acetyltransferase by Epstein-Barr virus in a Burkitt's lymphoma cell line. Virus Res 2013, 177, 11–21. [Google Scholar] [CrossRef]
- Pozzi, B.; Bragado, L.; Mammi, P.; Torti, M.F.; Gaioli, N.; Gebhard, L.G.; Garcia Sola, M.E.; Vaz-Drago, R.; Iglesias, N.G.; Garcia, C.C.; et al. Dengue virus targets RBM10 deregulating host cell splicing and innate immune response. Nucleic Acids Res 2020, 48, 6824–6838. [Google Scholar] [CrossRef]
- Fiches, G.N.; Wu, Z.; Zhou, D.; Biswas, A.; Li, T.W.; Kong, W.; Jean, M.; Santoso, N.G.; Zhu, J. Polyamine biosynthesis and eIF5A hypusination are modulated by the DNA tumor virus KSHV and promote KSHV viral infection. PLoS pathogens 2022, 18, e1010503. [Google Scholar] [CrossRef]
- Gassen, N.C.; Papies, J.; Bajaj, T.; Emanuel, J.; Dethloff, F.; Chua, R.L.; Trimpert, J.; Heinemann, N.; Niemeyer, C.; Weege, F.; et al. SARS-CoV-2-mediated dysregulation of metabolism and autophagy uncovers host-targeting antivirals. Nat Commun 2021, 12, 3818. [Google Scholar] [CrossRef]
- Zhou, Y.; Hou, Z.; Fang, L.; Ke, Q.; Xiong, Y.; Fang, P.; Xiao, S. Polyamine regulation of porcine reproductive and respiratory syndrome virus infection depends on spermidine-spermine acetyltransferase 1. Vet Microbiol 2020, 250, 108839. [Google Scholar] [CrossRef]
- Xie, H.; Ai, Q.; Tong, T.; Liao, M.; Fan, H. PEDV infection affects the expression of polyamine-related genes inhibiting viral proliferation. Virus Res 2022, 312, 198708. [Google Scholar] [CrossRef]
- Mastrodomenico, V.; LoMascolo, N.J.; Firpo, M.R.; Villanueva Guzman, M.D.M.; Zaporowski, A.; Mounce, B.C. Persistent Coxsackievirus B3 Infection in Pancreatic Ductal Cells In Vitro Downregulates Cellular Polyamine Metabolism. mSphere 2023, 8, e0003623. [Google Scholar] [CrossRef] [PubMed]
- Mastrodomenico, V.; Esin, J.J.; Qazi, S.; Khomutov, M.A.; Ivanov, A.V.; Mukhopadhyay, S.; Mounce, B.C. Virion-Associated Polyamines Transmit with Bunyaviruses to Maintain Infectivity and Promote Entry. ACS infectious diseases 2020, 6, 2490–2501. [Google Scholar] [CrossRef] [PubMed]
- Mastrodomenico, V.; LoMascolo, N.J.; Cruz-Pulido, Y.E.; Cunha, C.R.; Mounce, B.C. Polyamine-Linked Cholesterol Incorporation in Rift Valley Fever Virus Particles Promotes Infectivity. ACS infectious diseases 2022, 8, 1439–1448. [Google Scholar] [CrossRef]
- Firpo, M.R.; Mastrodomenico, V.; Hawkins, G.M.; Prot, M.; Levillayer, L.; Gallagher, T.; Simon-Loriere, E.; Mounce, B.C. Targeting Polyamines Inhibits Coronavirus Infection by Reducing Cellular Attachment and Entry. ACS infectious diseases 2021, 7, 1423–1432. [Google Scholar] [CrossRef]
- Kicmal, T.M.; Tate, P.M.; Dial, C.N.; Esin, J.J.; Mounce, B.C. Polyamine Depletion Abrogates Enterovirus Cellular Attachment. J Virol 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Mastrodomenico, V.; Esin, J.J.; Graham, M.L.; Tate, P.M.; Hawkins, G.M.; Sandler, Z.J.; Rademacher, D.J.; Kicmal, T.M.; Dial, C.N.; Mounce, B.C. Polyamine Depletion Inhibits Bunyavirus Infection via Generation of Noninfectious Interfering Virions. J Virol 2019, 93. [Google Scholar] [CrossRef]
- Olsen, M.E.; Filone, C.M.; Rozelle, D.; Mire, C.E.; Agans, K.N.; Hensley, L.; Connor, J.H. Polyamines and Hypusination Are Required for Ebolavirus Gene Expression and Replication. mBio 2016, 7. [Google Scholar] [CrossRef]
- Mounce, B.C.; Cesaro, T.; Moratorio, G.; Hooikaas, P.J.; Yakovleva, A.; Werneke, S.W.; Smith, E.C.; Poirier, E.Z.; Simon-Loriere, E.; Prot, M.; et al. Inhibition of Polyamine Biosynthesis Is a Broad-Spectrum Strategy against RNA Viruses. J Virol 2016, 90, 9683–9692. [Google Scholar] [CrossRef]
- Rojas-Luna, L.; Posadas-Modragon, A.; Avila-Trejo, A.M.; Alcantara-Farfan, V.; Rodriguez-Paez, L.I.; Santiago-Cruz, J.A.; Pastor-Alonso, M.O.; Aguilar-Faisal, J.L. Inhibition of chikungunya virus replication by N-omega-Chloroacetyl-L-Ornithine in C6/36, Vero cells and human fibroblast BJ. Antivir Ther 2023, 28, 13596535231155263. [Google Scholar] [CrossRef]
- Mao, B.; Wang, Z.; Pi, S.; Long, Q.; Chen, K.; Cui, J.; Huang, A.; Hu, Y. Difluoromethylornithine, a Decarboxylase 1 Inhibitor, Suppresses Hepatitis B Virus Replication by Reducing HBc Protein Levels. Front Cell Infect Microbiol 2020, 10, 158. [Google Scholar] [CrossRef]
- Korovina, A.N.; Tunitskaya, V.L.; Khomutov, M.A.; Simonian, A.R.; Khomutov, A.R.; Ivanov, A.I.; Kochetkov, S.N. Biogenic Polyamines Spermine and Spermidine Activate RNA Polymerase and Inhibit RNA Helicase of Hepatitis C Virus. Biochemistry (Moscow) 2012, 77, 1172–1180. [Google Scholar] [CrossRef]
- Hulsebosch, B.M.; Mounce, B.C. Polyamine Analog Diethylnorspermidine Restricts Coxsackievirus B3 and Is Overcome by 2A Protease Mutation In Vitro. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Smirnova, O.A.; Isaguliants, M.G.; Hyvonen, M.T.; Keinanen, T.A.; Tunitskaya, V.L.; Vepsalainen, J.; Alhonen, L.; Kochetkov, S.N.; Ivanov, A.V. Chemically induced oxidative stress increases polyamine levels by activating the transcription of ornithine decarboxylase and spermidine/spermine-N1-acetyltransferase in human hepatoma HUH7 cells. Biochimie 2012, 94, 1876–1883. [Google Scholar] [CrossRef]
- Firpo, M.R.; LoMascolo, N.J.; Petit, M.J.; Shah, P.S.; Mounce, B.C. Polyamines and eIF5A hypusination facilitate SREBP2 synthesis and cholesterol production leading to enhanced enterovirus attachment and infection. PLoS pathogens 2023, 19, e1011317. [Google Scholar] [CrossRef]
- Seoane, R.; Llamas-Gonzalez, Y.Y.; Vidal, S.; El Motiam, A.; Bouzaher, Y.H.; Fonseca, D.; Farras, R.; Garcia-Sastre, A.; Gonzalez-Santamaria, J.; Rivas, C. eIF5A is activated by virus infection or dsRNA and facilitates virus replication through modulation of interferon production. Front Cell Infect Microbiol 2022, 12, 960138. [Google Scholar] [CrossRef]
- Olsen, M.E.; Cressey, T.N.; Muhlberger, E.; Connor, J.H. Differential Mechanisms for the Involvement of Polyamines and Hypusinated eIF5A in Ebola Virus Gene Expression. J Virol 2018, 92. [Google Scholar] [CrossRef]
- Hoque, M.; Hanauske-Abel, H.M.; Palumbo, P.; Saxena, D.; D'Alliessi Gandolfi, D.; Park, M.H.; Pe'ery, T.; Mathews, M.B. Inhibition of HIV-1 gene expression by Ciclopirox and Deferiprone, drugs that prevent hypusination of eukaryotic initiation factor 5A. Retrovirology 2009, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Polvinen, K.; Sinervirta, R.; Alhonen, L.; Janne, J. Overproduction of ornithine decarboxylase confers an apparent growth advantage to mouse tumor cells. Biochem Biophys Res Commun 1988, 155, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Moreau, M.; Riviere, B.; Vegna, S.; Aoun, M.; Gard, C.; Ramos, J.; Assenat, E.; Hibner, U. Hepatitis C viral proteins perturb metabolic liver zonation. Journal of hepatology 2015, 62, 278–285. [Google Scholar] [CrossRef]
- Alisi, A.; Comparcola, D.; De Stefanis, C.; Nobili, V. Arginase 1: a potential marker of a common pattern of liver steatosis in HCV and NAFLD children. Journal of hepatology 2015, 62, 1207–1208. [Google Scholar] [CrossRef]
- Cao, W.; Sun, B.; Feitelson, M.A.; Wu, T.; Tur-Kaspa, R.; Fan, Q. Hepatitis C virus targets over-expression of arginase I in hepatocarcinogenesis. Int J Cancer 2009, 124, 2886–2892. [Google Scholar] [CrossRef] [PubMed]
- Gerresheim, G.K.; Bathke, J.; Michel, A.M.; Andreev, D.E.; Shalamova, L.A.; Rossbach, O.; Hu, P.; Glebe, D.; Fricke, M.; Marz, M.; et al. Cellular Gene Expression during Hepatitis C Virus Replication as Revealed by Ribosome Profiling. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Kakehashi, A.; Suzuki, S.; Ishii, N.; Okuno, T.; Kuwae, Y.; Fujioka, M.; Gi, M.; Stefanov, V.; Wanibuchi, H. Accumulation of 8-hydroxydeoxyguanosine, L-arginine and Glucose Metabolites by Liver Tumor Cells Are the Important Characteristic Features of Metabolic Syndrome and Non-Alcoholic Steatohepatitis-Associated Hepatocarcinogenesis. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Khadka, S.; Arthur, K.; Barekatain, Y.; Behr, E.; Washington, M.; Ackroyd, J.; Crowley, K.; Suriyamongkol, P.; Lin, Y.H.; Pham, C.D.; et al. Impaired anaplerosis is a major contributor to glycolysis inhibitor toxicity in glioma. Cancer & metabolism 2021, 9, 27. [Google Scholar] [CrossRef]
- Singleton, D.C.; Dechaume, A.L.; Murray, P.M.; Katt, W.P.; Baguley, B.C.; Leung, E.Y. Pyruvate anaplerosis is a mechanism of resistance to pharmacological glutaminase inhibition in triple-receptor negative breast cancer. Bmc Cancer 2020, 20, 470. [Google Scholar] [CrossRef]
- Mankouri, J.; Tedbury, P.R.; Gretton, S.; Hughes, M.E.; Griffin, S.D.; Dallas, M.L.; Green, K.A.; Hardie, D.G.; Peers, C.; Harris, M. Enhanced hepatitis C virus genome replication and lipid accumulation mediated by inhibition of AMP-activated protein kinase. Proceedings of the National Academy of Sciences of the United States of America 2010, 107, 11549–11554. [Google Scholar] [CrossRef]
- Cantor, J.R.; Abu-Remaileh, M.; Kanarek, N.; Freinkman, E.; Gao, X.; Louissaint, A., Jr.; Lewis, C.A.; Sabatini, D.M. Physiologic Medium Rewires Cellular Metabolism and Reveals Uric Acid as an Endogenous Inhibitor of UMP Synthase. Cell 2017, 169, 258–272. [Google Scholar] [CrossRef]
- Tanner, J.J.; Fendt, S.M.; Becker, D.F. The Proline Cycle As a Potential Cancer Therapy Target. Biochemistry-Us 2018, 57, 3433–3444. [Google Scholar] [CrossRef] [PubMed]
- Higgs, M.R.; Lerat, H.; Pawlotsky, J.M. Hepatitis C virus-induced activation of beta-catenin promotes c-Myc expression and a cascade of pro-carcinogenetic events. Oncogene 2013, 32, 4683–4693. [Google Scholar] [CrossRef]
- Nishimura, T.; Kohara, M.; Izumi, K.; Kasama, Y.; Hirata, Y.; Huang, Y.; Shuda, M.; Mukaidani, C.; Takano, T.; Tokunaga, Y.; et al. Hepatitis C virus impairs p53 via persistent overexpression of 3beta-hydroxysterol Delta24-reductase. J Biol Chem 2009, 284, 36442–36452. [Google Scholar] [CrossRef]
- Lee, M.S.; Dennis, C.; Naqvi, I.; Dailey, L.; Lorzadeh, A.; Ye, G.; Zaytouni, T.; Adler, A.; Hitchcock, D.S.; Lin, L.; et al. Ornithine aminotransferase supports polyamine synthesis in pancreatic cancer. Nature 2023, 616, 339–347. [Google Scholar] [CrossRef]
- Ivanova, O.N.; Snezhkina, A.V.; Krasnov, G.S.; Valuev-Elliston, V.T.; Khomich, O.A.; Khomutov, A.R.; Keinanen, T.A.; Alhonen, L.; Bartosch, B.; Kudryavtseva, A.V.; et al. Activation of Polyamine Catabolism by N(1),N(11)-Diethylnorspermine in Hepatic HepaRG Cells Induces Dedifferentiation and Mesenchymal-Like Phenotype. Cells 2018, 7. [Google Scholar] [CrossRef]
- Mistry, S.K.; Burwell, T.J.; Chambers, R.M.; Rudolph-Owen, L.; Spaltmann, F.; Cook, W.J.; Morris, S.M., Jr. Cloning of human agmatinase. An alternate path for polyamine synthesis induced in liver by hepatitis B virus. American journal of physiology. Gastrointestinal and liver physiology 2002, 282, G375–381. [Google Scholar] [CrossRef]
- Zhu, M.Y.; Iyo, A.; Piletz, J.E.; Regunathan, S. Expression of human arginine decarboxylase, the biosynthetic enzyme for agmatine. Biochim Biophys Acta 2004, 1670, 156–164. [Google Scholar] [CrossRef]
Scheme 1.
Scheme of metabolism of biogenic polyamines and adjacent pathways and their pharmacological inhibitors used in this study. Compounds that target metabolic enzymes are presented in red. P5C - Δ1-pyrrolidine-5-carboxylate.
Scheme 1.
Scheme of metabolism of biogenic polyamines and adjacent pathways and their pharmacological inhibitors used in this study. Compounds that target metabolic enzymes are presented in red. P5C - Δ1-pyrrolidine-5-carboxylate.
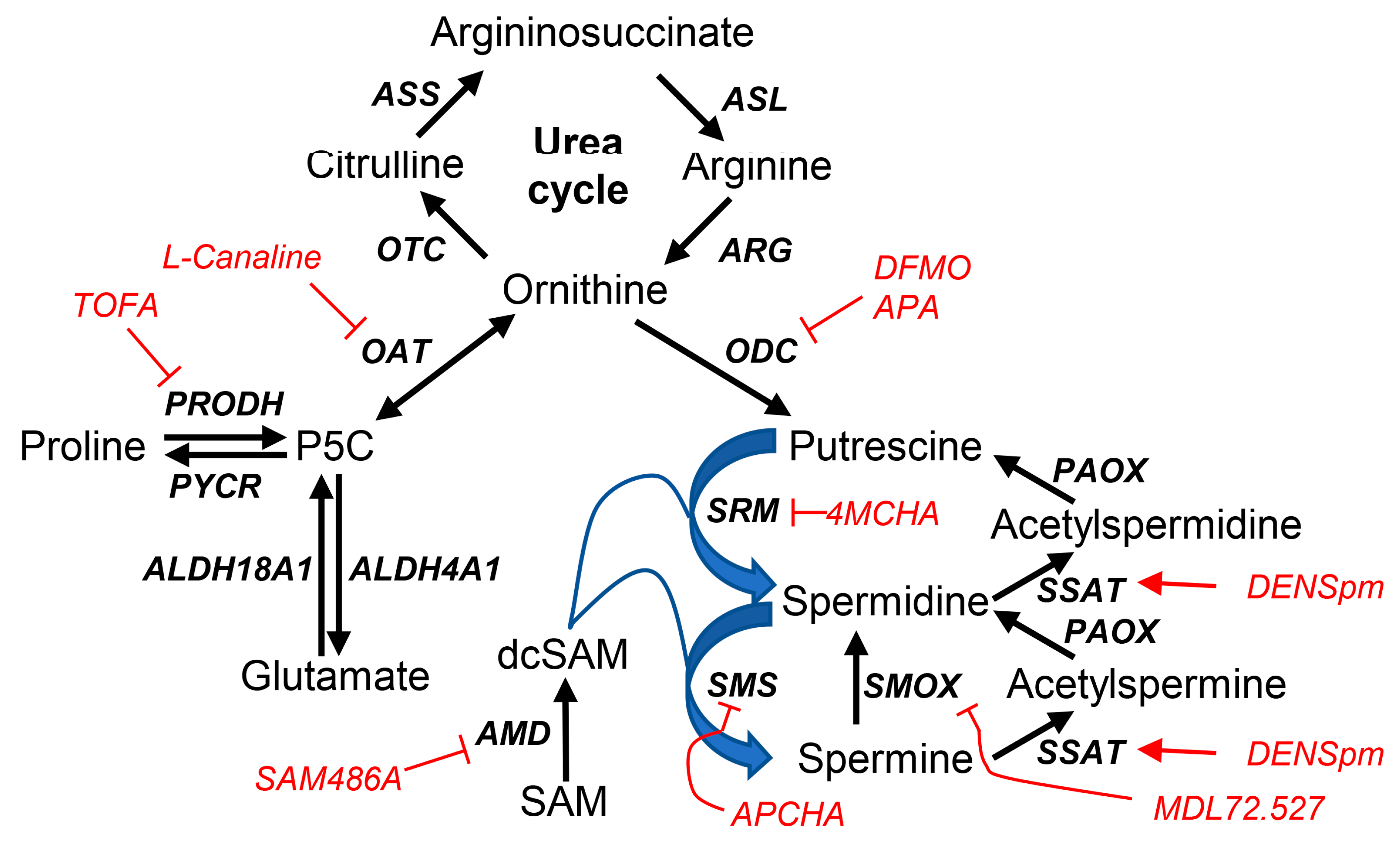
Figure 1.
HCV infection suppresses expression of polyamine-metabolizing enzymes at posttranscriptional level. (A-F) Huh7.5 cells were infected with HCV at MOI 0.1. (A) Infection spread was monitored by immunofluorescence staining using anti-NS3 rabbit sera and FITC-labelled antirabbit antibodies, the nuclei were counterstained with DAPI, and signals were visualized by confocal microscopy at 63x magnification. (B-D) Expression of polyamine-metabolizing proteins was assessed 3-10 days post-infection (dpi) by RT-qPCR (B-C) or western blotting (D). Intracellular SSAT activity was quantified by monitoring transfer of 14C-labelled acetyl group from [14C]-AcCoA to spermine molecule. (F) Polyamine levels were measured by HPLC. Graphs represent means ± SEM of four (B) or three (C,E,F) independent experiments. *p<0.05, **p<0.01, #p<0.1 compared to mock-infected cells by ANOVA with Dunnett’s post-hoc test (B,C,F) or paired t-test (E).
Figure 1.
HCV infection suppresses expression of polyamine-metabolizing enzymes at posttranscriptional level. (A-F) Huh7.5 cells were infected with HCV at MOI 0.1. (A) Infection spread was monitored by immunofluorescence staining using anti-NS3 rabbit sera and FITC-labelled antirabbit antibodies, the nuclei were counterstained with DAPI, and signals were visualized by confocal microscopy at 63x magnification. (B-D) Expression of polyamine-metabolizing proteins was assessed 3-10 days post-infection (dpi) by RT-qPCR (B-C) or western blotting (D). Intracellular SSAT activity was quantified by monitoring transfer of 14C-labelled acetyl group from [14C]-AcCoA to spermine molecule. (F) Polyamine levels were measured by HPLC. Graphs represent means ± SEM of four (B) or three (C,E,F) independent experiments. *p<0.05, **p<0.01, #p<0.1 compared to mock-infected cells by ANOVA with Dunnett’s post-hoc test (B,C,F) or paired t-test (E).
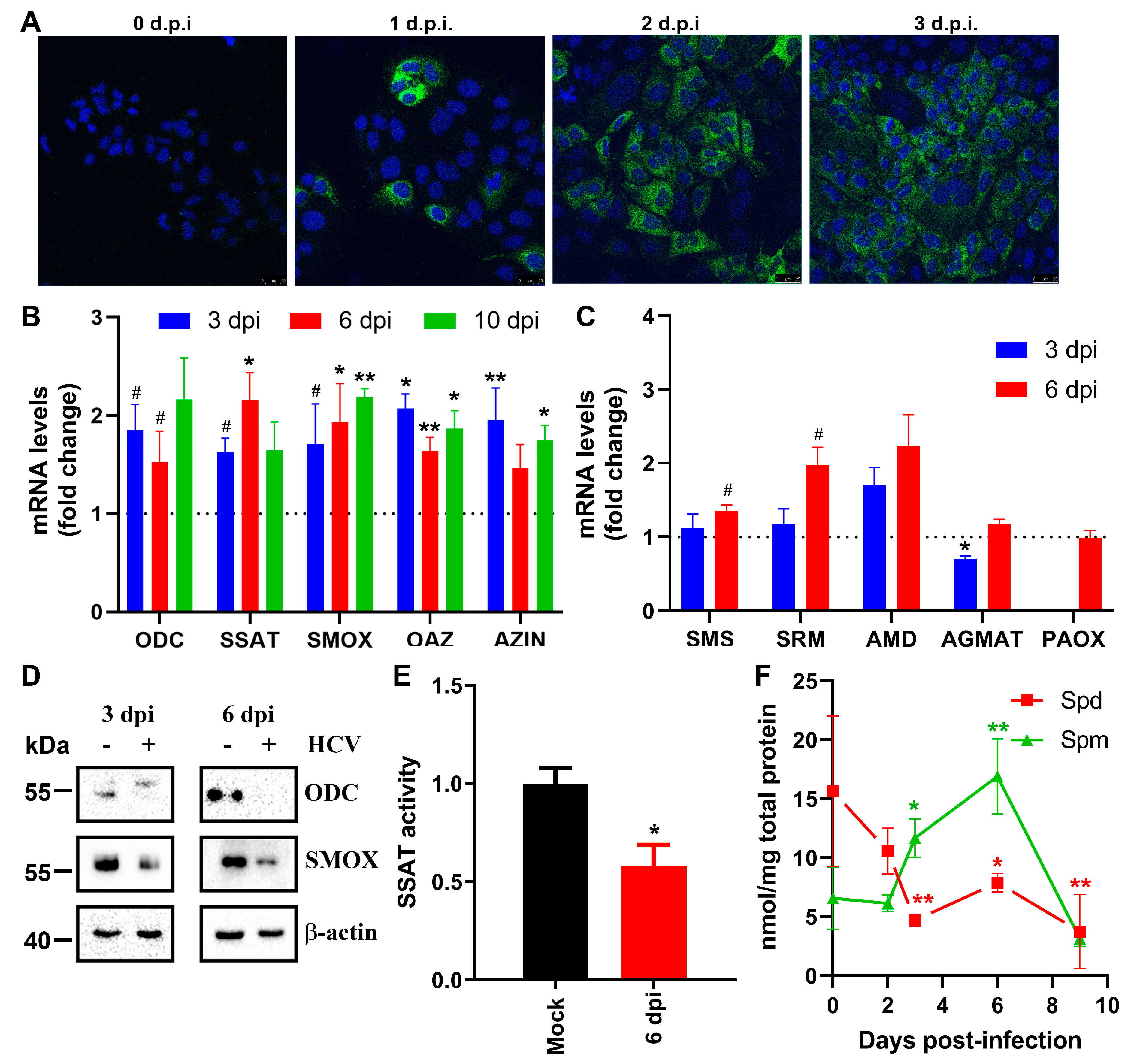
Figure 2.
HCV interferes with urea cycle by inhibiting expression of arginase 1. (A-C) Huh7.5 cells were infected with HCV at MOI 0.1 in DMEM-F12 media. (D-F) Alternatively, Huh7.5 cells harboring subgenomic HCV replicon or naïve Huh7.5 cells were maintained in DMEM-F12 (D,E) or Plasmax (F) media. Expression of urea cycle enzymes was assessed by RT-qPCR (A,D,F) or western blotting (B,E) with β-actin subpanels from 2B are the same as on Figure 1D as they are from the same experiment. © Relative intracellular arginine levels were quantified by LC-MS. Graphs represent means ± SEM of three independent experiments, with the exception of (A) RT-qPCR of cells 4 dpi (four independent experiments) and (F) cells in Plasmax medium (two experiments). *p<0.05, **p<0.01 compared to mock-infected cells or naïve Huh7.5 cells by ANOVA with Dunnett’s post-hoc test (A,C) or paired t-test (D,F). Note that actin blots on Panel B were from the same experiment as shown on Figure 1D.
Figure 2.
HCV interferes with urea cycle by inhibiting expression of arginase 1. (A-C) Huh7.5 cells were infected with HCV at MOI 0.1 in DMEM-F12 media. (D-F) Alternatively, Huh7.5 cells harboring subgenomic HCV replicon or naïve Huh7.5 cells were maintained in DMEM-F12 (D,E) or Plasmax (F) media. Expression of urea cycle enzymes was assessed by RT-qPCR (A,D,F) or western blotting (B,E) with β-actin subpanels from 2B are the same as on Figure 1D as they are from the same experiment. © Relative intracellular arginine levels were quantified by LC-MS. Graphs represent means ± SEM of three independent experiments, with the exception of (A) RT-qPCR of cells 4 dpi (four independent experiments) and (F) cells in Plasmax medium (two experiments). *p<0.05, **p<0.01 compared to mock-infected cells or naïve Huh7.5 cells by ANOVA with Dunnett’s post-hoc test (A,C) or paired t-test (D,F). Note that actin blots on Panel B were from the same experiment as shown on Figure 1D.
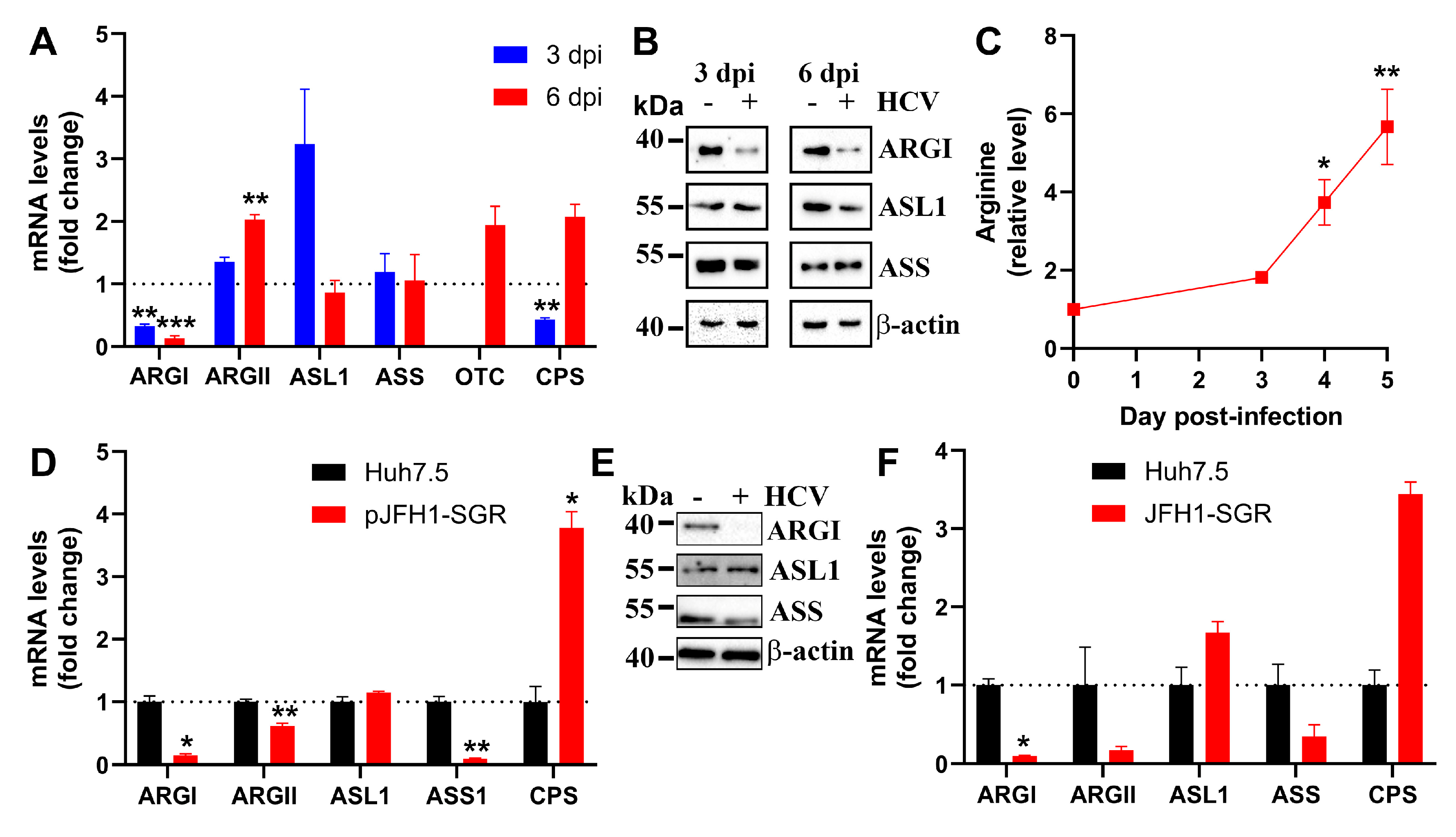
Figure 3.
HCV induces proline catabolism by up-regulating proline dehydrogenase. (A-C) Huh7.5 cells were infected with HCV at an MOI 0.1 in DMEM-F12 media. (D-G) Alternatively, Huh7.5 cells harboring subgenomic HCV replicon or naïve Huh7.5 cells were maintained in DMEM-F12 (D,E) or Plasmax (F) media or DMEM-F12 supplemented with 2.5 mM proline (G). Expression of proline- pyrrolidine-5-carboxylate-metabolizing enzymes was assessed by RT-qPCR (B,D,F) or western blotting (B,E). © Relative intracellular levels of proline were quantified by LC-MS. (H) Huh7.5 cells were treated with THFA 4 h prior to infection and 4 h post-infection with quantification of HCV RNA level 6 dpi. Graphs represent means ± SEM of three independent experiments, with the exception of (F) cells in Plasmax medium (two experiments). *p<0.05, **p<0.01 compared to mock-infected cells or naïve Huh7.5 cells by ANOVA with Dunnett’s post-hoc test (A) or paired t-test (D,F). The subpanel with β-actin on panel E are the same as on Figure 2E and subpanels from 2B (3 dpi) are the same as on 1D (3 dpi), as they are from the same experiments.
Figure 3.
HCV induces proline catabolism by up-regulating proline dehydrogenase. (A-C) Huh7.5 cells were infected with HCV at an MOI 0.1 in DMEM-F12 media. (D-G) Alternatively, Huh7.5 cells harboring subgenomic HCV replicon or naïve Huh7.5 cells were maintained in DMEM-F12 (D,E) or Plasmax (F) media or DMEM-F12 supplemented with 2.5 mM proline (G). Expression of proline- pyrrolidine-5-carboxylate-metabolizing enzymes was assessed by RT-qPCR (B,D,F) or western blotting (B,E). © Relative intracellular levels of proline were quantified by LC-MS. (H) Huh7.5 cells were treated with THFA 4 h prior to infection and 4 h post-infection with quantification of HCV RNA level 6 dpi. Graphs represent means ± SEM of three independent experiments, with the exception of (F) cells in Plasmax medium (two experiments). *p<0.05, **p<0.01 compared to mock-infected cells or naïve Huh7.5 cells by ANOVA with Dunnett’s post-hoc test (A) or paired t-test (D,F). The subpanel with β-actin on panel E are the same as on Figure 2E and subpanels from 2B (3 dpi) are the same as on 1D (3 dpi), as they are from the same experiments.
Figure 4.
Inhibitors of ornithine decarboxylase and polyamine oxidases inhibit HCV replication. (A) Huh7.5 cells harboring a subgenomic HCV replicon were treated with 20 µM DENSpm, CPENSpm, CHENSpm or MDL72.527 for 3 days. (B) Huh7.5 subgenomic replicon-harboing cells were cultivated in DMEM with standard (DMEM) or dialysed (dDMEM) FBS supplemented with 1 mM aminoguanidine alone or with a mixture of spermine and spermidine (100 µM each). (C,D) Huh7.5 cells were infected with HCV at 0.1 MOI and treated with DFMO, MDL72.527, DENSpm or with GC7 (10 and 30 µM), deferiprone (DFP, 200 and 600 µM), ciclopirox (25 and 75 µM), trans-4-methyl-cyclohecylamine (4MCHA), N-(3-aminopropyl)cyclohexylamine (APCHA) or L-canaline (30 and 100 µM) or SAM486A (1 and 3 µM) for 6 days. (E) Huh7.5 cells harboring subgenomic HCV replicon were treated similarly for 3 days. HCV RNA levels were quantified by RT-qPCR. Graphs represent means ± SEM of three independent experiments. **p<0.01 and ***p<0.001 by ANOVA with Dunnett’s post-hoc test (A) or Tukey’s test (B).
Figure 4.
Inhibitors of ornithine decarboxylase and polyamine oxidases inhibit HCV replication. (A) Huh7.5 cells harboring a subgenomic HCV replicon were treated with 20 µM DENSpm, CPENSpm, CHENSpm or MDL72.527 for 3 days. (B) Huh7.5 subgenomic replicon-harboing cells were cultivated in DMEM with standard (DMEM) or dialysed (dDMEM) FBS supplemented with 1 mM aminoguanidine alone or with a mixture of spermine and spermidine (100 µM each). (C,D) Huh7.5 cells were infected with HCV at 0.1 MOI and treated with DFMO, MDL72.527, DENSpm or with GC7 (10 and 30 µM), deferiprone (DFP, 200 and 600 µM), ciclopirox (25 and 75 µM), trans-4-methyl-cyclohecylamine (4MCHA), N-(3-aminopropyl)cyclohexylamine (APCHA) or L-canaline (30 and 100 µM) or SAM486A (1 and 3 µM) for 6 days. (E) Huh7.5 cells harboring subgenomic HCV replicon were treated similarly for 3 days. HCV RNA levels were quantified by RT-qPCR. Graphs represent means ± SEM of three independent experiments. **p<0.01 and ***p<0.001 by ANOVA with Dunnett’s post-hoc test (A) or Tukey’s test (B).
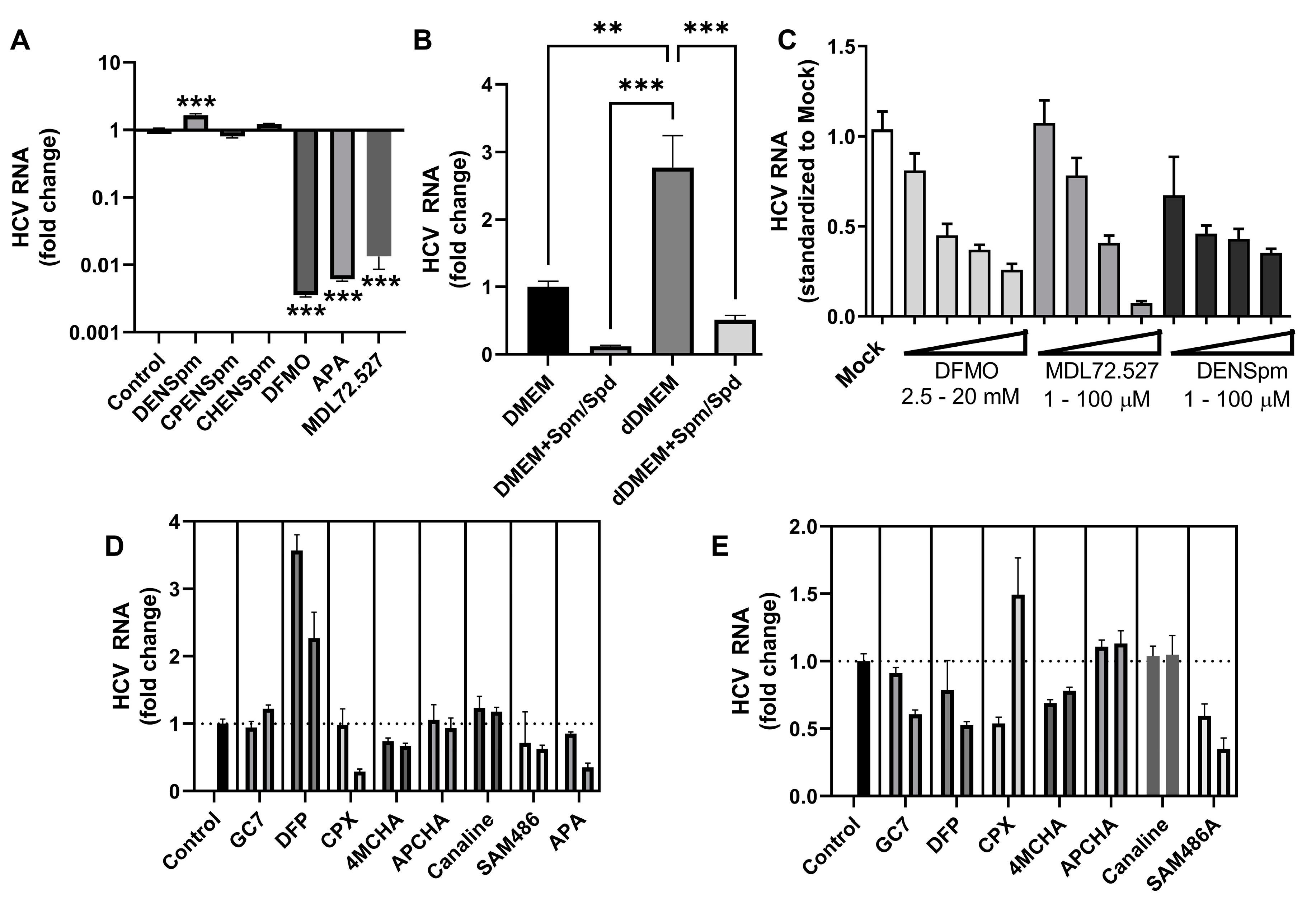

Table 1.
Influence of the compounds on polyamine metabolism.
| Cell line | Compound | Polyamine (nmol/mg DNA) |
Enzyme activity (nmol/h/mg total protein) |
||
|---|---|---|---|---|---|
| Spermidine | Spermine | ODC | SSAT | ||
| Naïve Huh7.5 | Control | 55,3 ± 10,8 | 49,6 ± 8,3 | 9,3 ± 1,2 | 33,1 ± 2,6 |
| DENSpm | 41,8 ± 9,5 | 17,9 ± 3,9 | 54,7 ± 3,2 | 217,5 ± 18,6 | |
| DFMO | 47,0 ± 10,1 | 37,2 ± 5,4 | 5,5 ± 0,6 | 2,7 ± 1,5 | |
| MDL72.527 | 63,5 ± 13,8 | 51,2 ± 13,7 | 4,4 ± 0,7 | 2,3 ± 0,2 | |
| HCV replicon- harboring Huh7.5 cells |
Control | 38,7 ± 6,1 | 27,9 ± 10,6 | 4,3 ± 1,1 | 8,6 ± 0,8 |
| DENSpm | 33,3 ± 8,3 | 14,1 ± 7,4 | 38,7 ± 2,1 | 190,3 ± 10,7 | |
| DFMO | 177,3 ± 42,2 | 140,1 ± 52,1 | 7,1 ± 1,5 | 1,2 ± 0,1 | |
| MDL72.527 | 227,6 ± 39,4 | 177,6 ± 27,8 | 17,4 ± 3,3 | 1,2 ± 0,2 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



