Submitted:
30 April 2024
Posted:
01 May 2024
You are already at the latest version
Abstract
Preeclampsia, a hypertensive disease of pregnancy of unknown etiology, is intensely studied as a model of cardiovascular (CVD) disease not only due to multiple shared pathologic elements, but also because changes that develop over decades in CVD disease appear and resolve within days in preeclampsia. Those affected by preeclampsia and their offspring experience increased lifetime risks of CVD. At the systemic level, preeclampsia is characterized by increased cellular, membrane, and blood levels of cholesterol, however, cholesterol dependent signaling such as canonical Wnt/βcatenin, Hedgehog, and endothelial nitric oxide synthase is downregulated indicating a cholesterol deficit with upregulation of cholesterol synthesis and efflux. Hypoxia related signaling in preeclampsia also appears paradoxical with increased Hypoxia Inducible Factors (HIF) in the placenta but measurably increased oxygen in maternal blood in placental villous spaces. This review addresses the molecular mechanisms by which excessive systemic cholesterol and deficient cholesterol dependent signaling may arise from the effects of dietary lipid variance causing the cellular hypoxia which characterizes preeclampsia.
Keywords:
preeclampsia
; hypoxia inducible factors
; cholesterol
; dietary lipids
; Hedgehog (Hh)
; Wnt/βcatenin
; membrane biophysics
; endothelial nitric oxide synthase
; dyslipidemia
; oxidative stress
1. Introduction
Lipids are intimately involved in preeclampsia pathology with increased risk for those with increased serum trans (TUFA)[1] or polyunsaturated fatty acids (PUFA)[2,3], reduced serum omega-3 PUFA[4], or dyslipidemia[5,6]; metabolomic studies consistently show sterol, phospholipid, and sphingolipid metabolism as major discriminatory pathways[7,8,9,10,11,12,13,14,15]. However, interventional studies of the effects of diet on preeclampsia are variable. For example, weight reduction[16] and reduction of dietary TUFA significantly reduce risk[17], yet adherence to a vegan diet increases risk[18]. Many studies show an increased risk of preeclampsia with increased dietary PUFA[19], but other populations show reduced risk with PUFA supplementation[20]. The role of cholesterol in the etiology of human preeclampsia is especially confusing with evidence of cholesterol excess systemically but deficit in cholesterol dependent signaling pathways. Standard medical advice is to reduce dietary cholesterol, yet some studies show those with higher cholesterol intakes have reduced preeclampsia risk[21].
An animal model of preeclampsia (Reduced Uterine Perfusion Pressure or RUPP) may be created by partially occluding vessels that supply the pregnant uterus. This ischemic model reproduces many of the clinical features of human preeclampsia including increased blood pressure and total peripheral resistance with impaired nitric oxide-dependent vasodilation, impaired renal glomerular filtration with proteinuria, reduced ejection fraction and cardiac output, immune dysfunction with increased prehypertensive cytokines, T cells, and B cells, as well as increased capillary permeability with cerebral edema and reduction in blood brain barrier integrity[22]. In laboratory animals, these clinical features may be traced directly to the effects of hypoxia related signaling and are mitigated by inhibiting HIF-2⍺[23]. Human preeclampsia is also characterized by increased HIF signaling[24]. Intriguingly, in humans with early onset preeclampsia (EOPE), measurements of oxygen concentrations in the villous space where maternal blood contacts the placental villous trophoblast and in the uterine vein leaving the maternal uterus do not show hypoxia but higher partial pressures of oxygen than in normal healthy pregnancies. However, in these pregnancies fetal oxygenation is reduced[25], indicating movement of oxygen across the placenta is impaired in those with EOPE. Because cholesterol presents a major impediment to diffusion of oxygen across membranes, elevated membrane cholesterol offers a mechanism for increased hypoxia related signaling from the placenta. However, the fundamental cause of increased membrane cholesterol and reduced movement of oxygen across the placenta is unknown.
Over the last century dietary lipid intake in the United States has markedly changed and environmental toxins such as perfluoroalkyl substances (PFAS), bisphenol A (BPA) and inhaled hydrocarbons have increased. This review first outlines the effects of dietary lipids and environmental membrane modifying agents on membrane biophysics, cholesterol sequestration, and oxygen diffusion, then integrates these findings with observational and interventional studies in those with preeclampsia. This integration provides a molecular mechanism for preeclampsia’s hypoxia and aberrant cholesterol signaling.
2. Changes in Dietary Lipids in the United States of America (US) over the Last Century
During the last century, the US population has undergone a widespread uncontrolled trial of the effects of dietary lipids on human health. From 1909 until 2011, the US Department of Agriculture (USDA) maintained data on foods consumed in the United States each year. Over this interval, the per capita daily consumption of saturated fats decreased from 28 pounds in 1912 to 13.4 pounds in 2011, while consumption of PUFA in vegetable oils increased from 11.3 to 64.5 pounds. The estimated per capita consumption of soybean oil, now the most used cooking oil in the US, increased over 1000-fold from 1909 to 1999.[26] Total fat intake increased from 41 to 79 pounds per capita per year, an increase almost totally due to an increase in PUFA. [27,28] In the last century, worldwide TUFA consumption rose from an optimal level of less than 1% of daily dietary fats to a high of 6.5% in some countries.[29,30] Although levels of TUFA consumption in packaged foods fell after 2013 when the USDA classified TUFA as a food additive[31] and the 2018 World Health Organization initiative limited dietary intake[32], TUFAs form at a rate of about 3.5 grams per 100 grams of polyunsaturated oil with each cooking episode.[33] Reuse of oil in deep fryers and the use of spent oil in livestock feeds means there is an ongoing yet unmeasured supply of TUFA in human diets. Humans evolved with an omega 6:3 ratio of about 1:1, while Western diets now average a ratio of 16:1 with levels rising to 50:1 in areas of urban India.[34] Archeologic studies estimate daily cholesterol intake of our hunter/gatherer genetic ancestors at 480 to 880mg per day,[35,36] yet average daily dietary cholesterol in the United States in 2014 was 293 mg per day.[37]
In summary, lipids in modern diets are characterized by 1) a marked increase in dietary fat almost wholly from polyunsaturated vegetable oils (increased unsaturation index), 2) a high omega 6:3 ratio, 3) increased trans fatty acids, 4) reduced dietary cholesterol and 5) reduced saturated fatty acids.
Because dietary fats are rapidly incorporated into tissues, these dietary variances may predictably change the biophysical and signaling properties of placental and maternal membranes in preeclampsia.
3. Environmental Lipids and Membrane Modifiers Affect Preeclampsia Risk
Although there is some disagreement[38], generally environmental studies demonstrate an increased risk of preeclampsia for those with chronic exposure to air pollution[39,40], traffic-related air pollution[41,42], indoor air pollution[43,44], and inhaled volatile organic compounds[45,46]. Exposure only to non-hydrocarbon particulate air pollution is not clearly associated increased preeclampsia risk[47,48]. An analysis of components of fine particulate air pollution shows specific components differentially increase preeclampsia risk with organic carbon components having greater adverse effect[49]. From these studies, it appears air pollution which contains pollutants such as benzene, toluene, xylene (volatile organic compounds) and oil, tar, gasoline, and diesel combustion components that may be absorbed through lung alveolar surface may be the source of increased preeclampsia risk. These compounds modify cell membrane fluidity and other biophysical and biochemical membrane properties[50] and are associated with clinical symptoms of increased susceptibility to vasospasm, increased blood pressure, and increased atherosclerotic lesions. They also dysregulate cellular signaling and gene expression[51].
4. Placental Hypoxia and Hypoxia Inducible Factors (HIF)
The term “hypoxia” is often used in discussing preeclampsia pathology partially due to the success of the RUPP animal model which uses ischemia to create preeclampsia-like pathology, but also due to the marked increase in HIFs in placentae of human preeclampsia [57]. Regulation of HIF is complex. With normal oxygen concentrations in placental and endothelial cytoplasm, HIFs are continuously produced but rapidly degraded through action of prolyl hydroxylase domains. However, at low oxygen tension, prolyl hydroxylation is suppressed causing stabilization of HIFs which translocate to the nucleus and stimulate transcription of over sixty hypoxia responsive genes regulating diverse pathways such as fibrosis[58], angiogenesis, erythropoiesis, and mitochondrial homeostasis[59]. Dysregulation of these pathways with elevation of HIFs is central to human preeclampsia pathophysiology[60]. Although HIFs may also be stimulated under normal oxygen conditions by other factors such as heat shock[61], increased mechanical stretch[62], reactive oxygen species[63], growth stimuli[64], thiamine deficiency[65], inflammation[66], and hyperglycemia[67,68], a significant part of preeclampsia’s elevated HIF likely arises from placental cellular hypoxia[69].
Low oxygen tension also stimulates changes in cholesterol homeostasis in human trophoblasts with increases in cholesterol synthesis and efflux to maternal serum, changes which occur in human preeclampsia[70,71]. Thus, cellular hypoxia may contribute both the to the increase in HIF signaling and changes in cholesterol homeostasis which characterize preeclampsia. However, the origin of placental and endothelial cell hypoxia remains a mystery.
Early animal models of preeclampsia, show dietary changes reduce placental uptake of oxygen. A 1965 study with Columbia-Sherman rats, investigates the effects of an experimental diet deficient in vitamin E to which 5% oxidized cod liver oil is added. With this diet, transport of sodium, potassium, and oxygen in the placenta is reduced[72]. Although HIF would not be discovered for almost thirty years[73], presumably reduction in oxygen uptake by the placenta would have resulted in placental cell hypoxia and elevated HIFs in these animals.
Early placental development in humans requires a hypoxic environment. In this environment, HIF-1⍺ and Transforming Growth Factor β3, which inhibits trophoblastic differentiation, are increased. In healthy pregnancies, toward the end of the first trimester spiral artery remodeling results in an increase in villous blood flow and a reduction in both HIF-1⍺ and in TGFβ3, stimulating normal placental development. However, in preeclampsia, the normal increase in villous blood flow does not occur, there is continued hypoxia with levels of HIF and TGFβ3 remaining high, and placental development is impaired[74].
These studies raise two fundamental questions about preeclampsia pathology: “What impedes movement of oxygen across the placenta?” and “What impairs placental development causing reduction in late first trimester villous blood flow?” We will first address the mechanism by which dietary and environmental elements may induce membrane changes which impede transplacental oxygen diffusion, then consider the role of these molecules in reducing cholesterol dependent angiogenic signaling and creating poor placentation.
5. Impedance to Transmembrane Oxygen Diffusion
Availability of oxygen in fetal tissues is dependent on three elements: oxygen saturation in maternal blood, exposure time of maternal RBCs to the villus trophoblast, and transmembrane oxygen diffusion time [75,76]. As mentioned above, in EOPE maternal blood in the villous space contains more, not less oxygen. Although hypertension and hemodynamic changes may alter the exposure time of RBCs at the trophoblast, hypoxia and reduced villous blood flow precede the onset of these changes by weeks with studies of placental exchange showing a decrease in oxygen conductance at the level of the villus trophoblast and fetal vasculature[77]. We will focus on the third element, that is, characteristics of the placental and fetal membranes which may reduce movement of oxygen from maternal red blood cells across the placenta to fetal blood.
Studies of in vitro artificial membranes and ex vivo natural membranes find increases in six membrane characteristics reduce trans membrane oxygen diffusion: unsaturation of membrane lipids, increased cytoskeletal elements, increased membrane stiffness, increased cholesterol, increased thickness, and increased protein content[78].
The role of membrane cholesterol in reducing trans membrane movement of oxygen is well studied in ophthalmology where cholesterol protects the highly oxidizable light sensing molecule rhodopsin and its surrounding PUFA from oxidative damage. Cholesterol’s effect may also be seen in the lens of the eye which is surrounded by a membrane with a very high cholesterol content that protects the lens from oxidative damage and prevents cataract formation. As the cholesterol concentration in plasma membranes rises above the normal of 10 to 20%, the membrane permeability coefficient (PM) for oxygen declines proportionally and movement of oxygen across membranes may be reduced to almost zero[79,80,81]. At concentrations over about 50%, membrane saturation occurs which allows cholesterol/cholesterol bilayer domains to form. These domains have a PM about one tenth that of surrounding membranes and present a significant barrier to oxygen transport[82]. Dietary and environmental lipids are largely responsible for determining the cholesterol content and biophysical properties of endothelial and placental membranes.
6. Effects of Molecular Components on Membrane Biophysics and Phase Separation
In the laboratory, a phospholipid bilayer forms spontaneously if phospholipids are mixed with cholesterol in an aqueous environment. The physical characteristics of this bilayer membrane are determined by the component lipids. A variety of head groups such as phosphatidylserine, phosphatidylethanolamine, phosphatidylcholine, sphingomyelin, and ceramide, each having one or two attached fatty acid tails creates an enormous diversity of lipids that compose membranes[83]. Each phospholipid or sphingolipid molecule differs in chemical and physical characteristics allowing properties such as fluidity or packing density, charge, phase separation, and permeability to be finely tuned for optimal membrane function. In general, longer fatty acids and fewer double bonds increase the membrane melting point, reduce membrane fluidity and permeability, and increase cholesterol affinity. Shorter fatty acids and more double bonds reduce packing density and cholesterol affinity resulting in a more fluid and permeable membrane. Oxidation of fatty acids and sterols generally reduces packing density causing a more fluid lipid bilayer membrane when compared to unoxidized analogs. Membrane composition also influences formation of cytoskeletal elements, stiffness, thickness, association of proteins, and formation of cholesterol crystals.
Even a small change in molecular structure may cause major differences in how a molecule behaves in a membrane. For example, elaidic and oleic acids have identical chemical formulae, each containing eighteen carbons with one double bond at position nine. Oleic acid’s double bond in a cis configuration gives this fatty acid’s shape a distinct kink which reduces packing density, thereby increasing membrane fluidity and permeability when compared to elaidic acid with a bond in the trans configuration. This simple change in the configuration of one double bond gives elaidic acid a melting point that is 31℃ higher than that of oleic acid. Elaidic acid also has a much higher cholesterol affinity requiring eight times as much cholesterol as oleic to be incorporated into a membrane[84]. Thus, substituting the trans fatty acid elaidic acid for oleic acid raises membrane cholesterol concentration, a finding common to CVD and preeclampsia[85]. Figure 1 and Figure 2 list some representative fatty acids and headgroup molecules which compose living cell membranes.
The high aversion of PUFA for cholesterol drives a process called phase separation in which membrane lipids with diverse melting points separate into liquid ordered and liquid disordered areas or domains. Molecules with higher melting points and high cholesterol affinity such as LCSFAs group together to form liquid ordered areas which correspond to rafts in living membranes with more fluid non-raft areas comprised of lower melting point lipids. Increased disparity of melting points in component lipids increases phase separation.
Figure 3.
A: Membrane Formation with Small Melting Point Disparity of Component Lipids. In vitro, on mixing a phospholipid and cholesterol in an aqueous environment, a phospholipid bilayer spontaneously forms. This process does not require DNA, proteins, or an external energy source.
Figure 3.
A: Membrane Formation with Small Melting Point Disparity of Component Lipids. In vitro, on mixing a phospholipid and cholesterol in an aqueous environment, a phospholipid bilayer spontaneously forms. This process does not require DNA, proteins, or an external energy source.
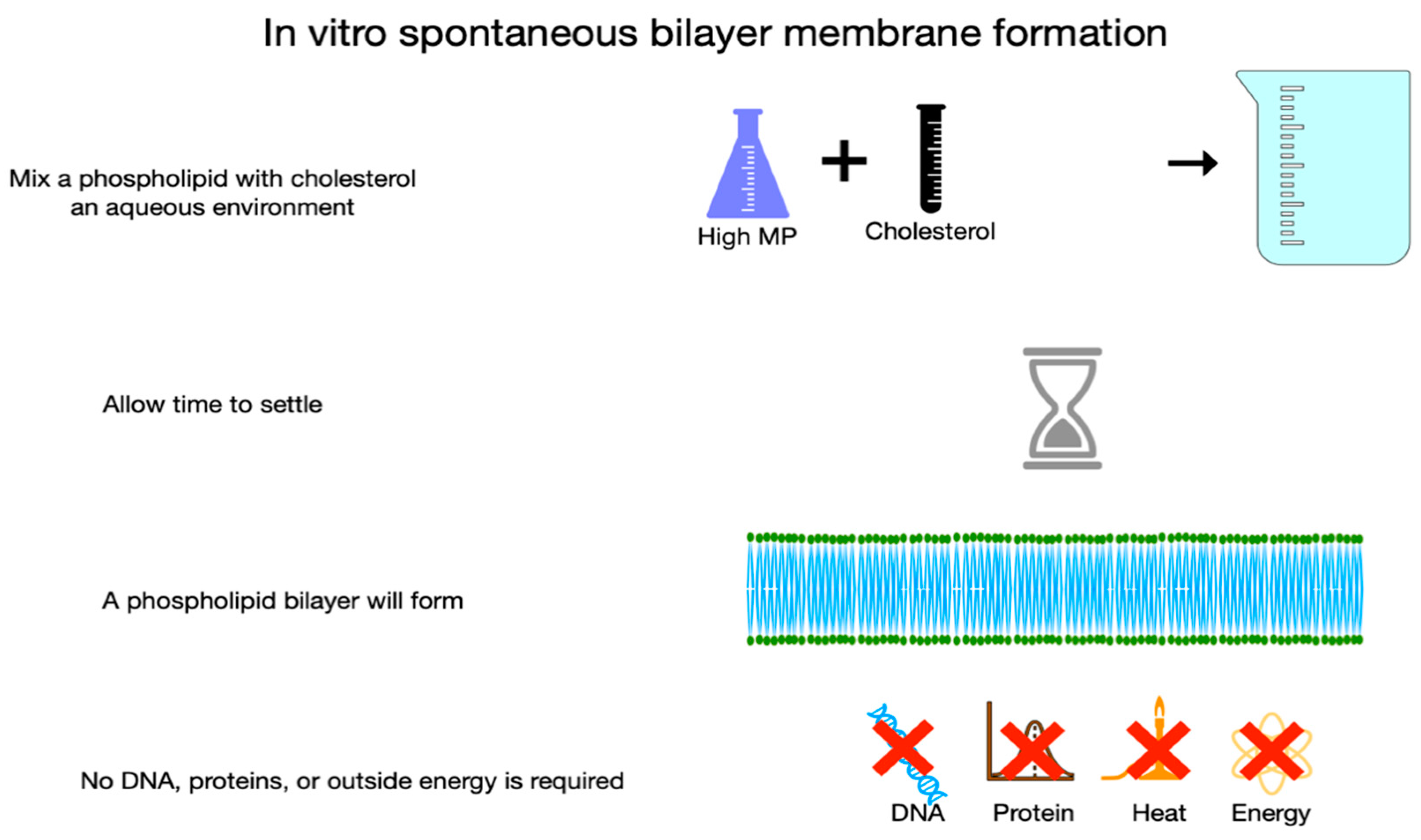
Figure B.
Membrane Formation with Wide Melting Point Disparity of Component Lipids. On adding a low melting point lipid to the mixture shown in Figure 3A, the lipids do not mix homogeneously allowing areas with greater and lesser order to form. PUFA’s aversion to cholesterol is the driving force in this process of phase separation in which areas of high melting point lipids gather with cholesterol away from low melting point PUFA. The thicker denser blue area in the middle of the bilayer membrane represents a liquid ordered area which corresponds to rafts in living cells. CVD is characterized by excess membrane rafts.
Figure B.
Membrane Formation with Wide Melting Point Disparity of Component Lipids. On adding a low melting point lipid to the mixture shown in Figure 3A, the lipids do not mix homogeneously allowing areas with greater and lesser order to form. PUFA’s aversion to cholesterol is the driving force in this process of phase separation in which areas of high melting point lipids gather with cholesterol away from low melting point PUFA. The thicker denser blue area in the middle of the bilayer membrane represents a liquid ordered area which corresponds to rafts in living cells. CVD is characterized by excess membrane rafts.
7. Dietary Challenges to Membrane Homeoviscosity
The importance of maintaining normal membrane viscosity to cell viability was first studied in single cell organisms and poikilotherms with work by Sinensky in the 1970s. This work defined a mechanism by which cells responded to cold or heat stress by metabolic adaptations of membrane composition to regain optimal membrane viscosity.[87] More recently, the Leventhal Lab investigated the effects of challenging membrane homeoviscosity in rats with a large dose of dietary PUFA. In these experiments, dietary lipids are rapidly incorporated into tissues with increased dietary PUFA resulting in excessively fluid membranes with high permeability, a similar situation to heat stress. To maintain optimal viscosity after incorporation of highly unsaturated fatty acids into a membrane, there is compensatory remodeling of the lipidome with upregulation of saturated fatty acids and cholesterol synthesis, resulting in recovery of normal membrane packing and permeability. If this compensatory response is blocked, the cell dies, indicating the critical necessity of maintaining normal membrane viscosity to cell viability[88]. These studies raise the possibility of major changes in dietary lipids challenging membrane homeoviscosity and causing lipid remodeling in humans.
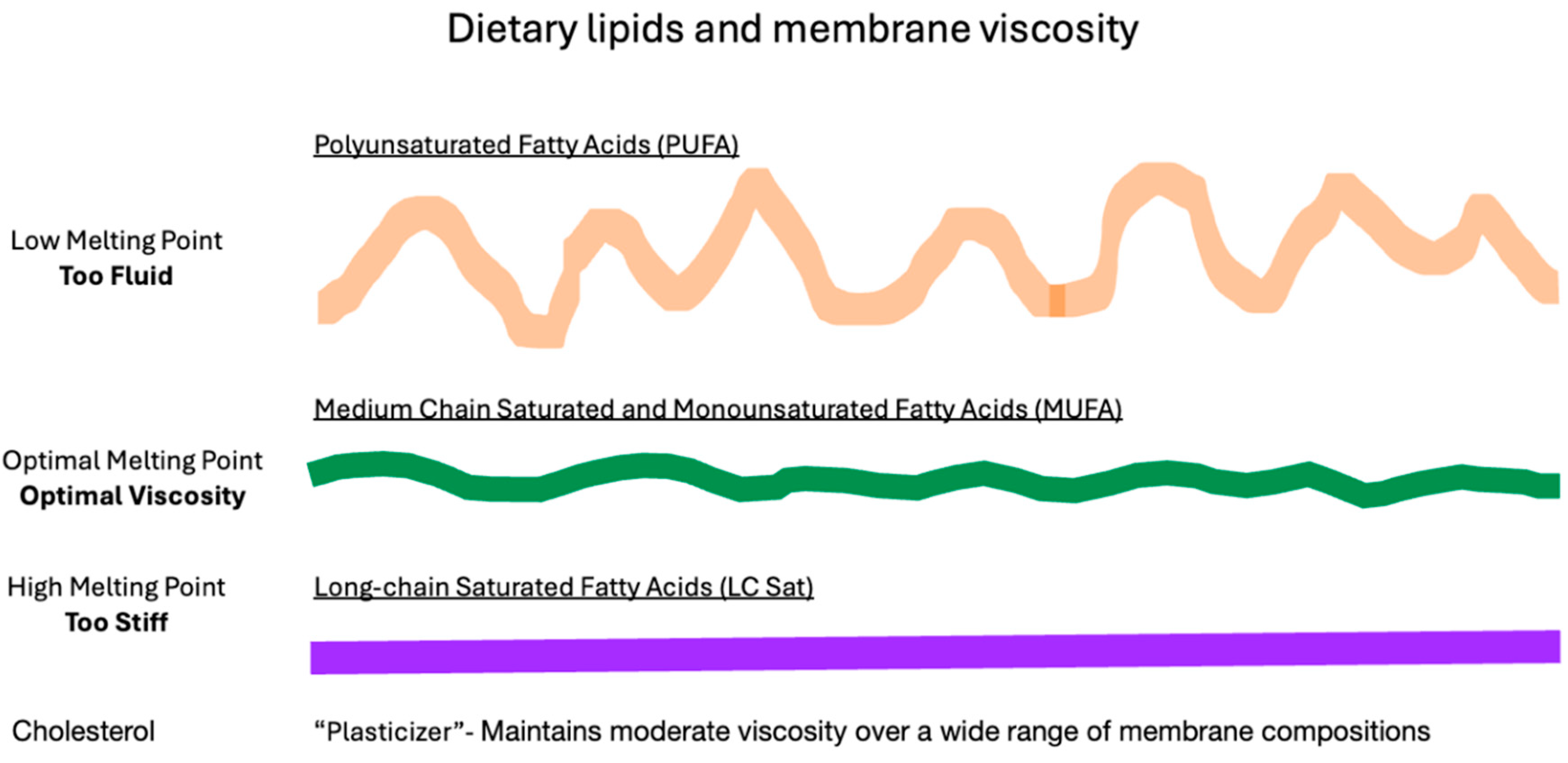
Figure 4.
Dietary Lipids and Membrane Viscosity. On challenging membrane homeostasis of laboratory animals with a high level of dietary low melting point PUFA, the fatty acids are rapidly incorporated into membranes causing the bilayer to become excessively fluid (represented by the wavey orange line in the top image). To compensate, increased synthesis and incorporation of cholesterol and high melting point LCSFA increases membrane rigidity (represented by the straight purple line in the bottom image), allowing membranes to regain normal viscosity (green moderately wavey line in the middle). Compensatory membrane remodeling is necessary for cell viability; if blocked, the cell dies.
Figure 4.
Dietary Lipids and Membrane Viscosity. On challenging membrane homeostasis of laboratory animals with a high level of dietary low melting point PUFA, the fatty acids are rapidly incorporated into membranes causing the bilayer to become excessively fluid (represented by the wavey orange line in the top image). To compensate, increased synthesis and incorporation of cholesterol and high melting point LCSFA increases membrane rigidity (represented by the straight purple line in the bottom image), allowing membranes to regain normal viscosity (green moderately wavey line in the middle). Compensatory membrane remodeling is necessary for cell viability; if blocked, the cell dies.
8. Excessively Fluid Lipid Bilayers Increase Membrane Stiffness
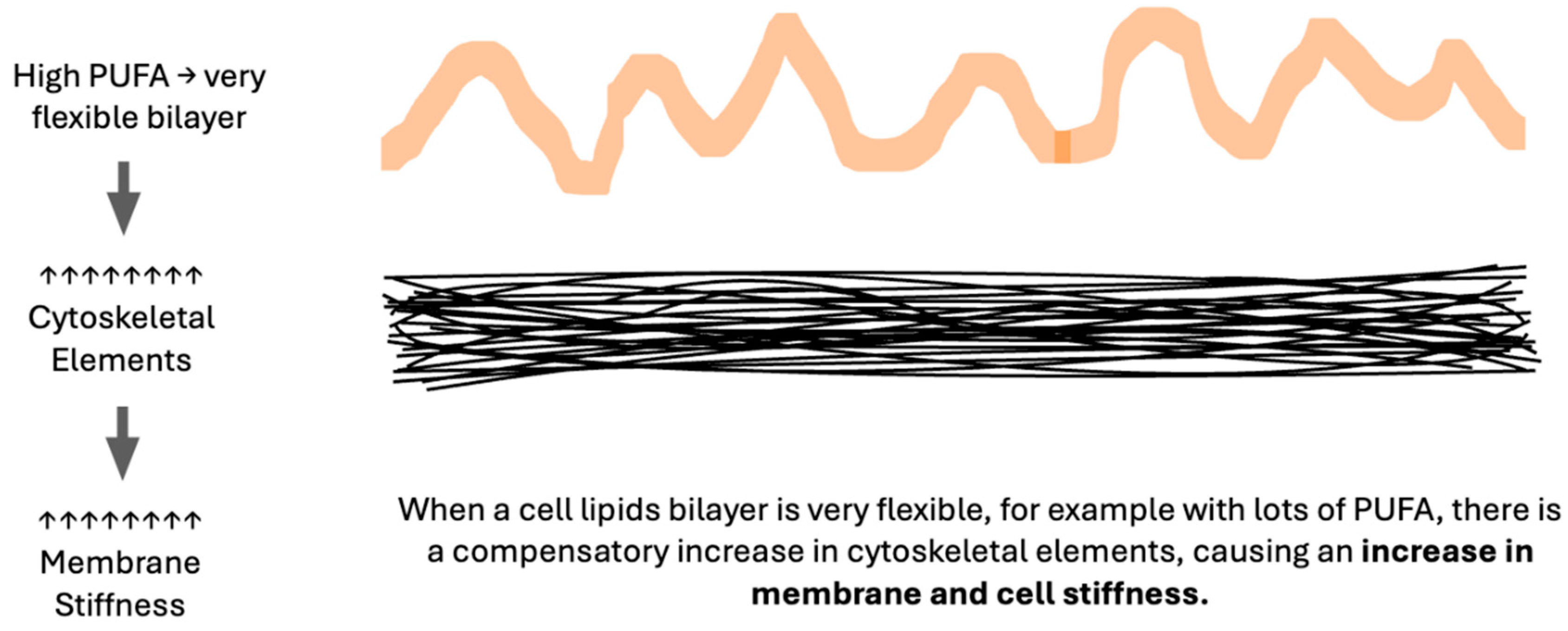
In considering the six factors which impede the movement of oxygen across membranes, that is, increased membrane unsaturation, cytoskeletal elements, stiffness, cholesterol, thickness, or proteins, an apparent inconsistency arises. An increase in unsaturation index and in membrane stiffness both reduce oxygen diffusion, yet increased unsaturation increases bilayer fluidity. A similar inconsistency is seen in preeclampsia literature with many lipidomic data sets indicating an increased unsaturation index in preeclampsia[89,90,91,92], yet measurably stiffer maternal RBC membranes[93,94,95] as is true in non-pregnant individuals with chronic hypertension[96]. The key to this paradox is the role of excessively fluid lipid bilayers in stimulating a compensatory increase in cytoskeletal elements, producing a stiffer cell as measured by RBC deformability or atomic force microscopy[97,98,99,100]. Thus, membrane stiffness is directly proportional to membrane lipid unsaturation index. (Figure 5) Actions of magnesium, a commonly used medication to reduce seizure risk in women with preeclampsia, include reduction of RBC stiffness[101] and disassembly of cytoskeletal elements[102].
Figure 5.
Excessively Fluid Bilayers Cause Compensatory Increases in the Cytoskeleton.
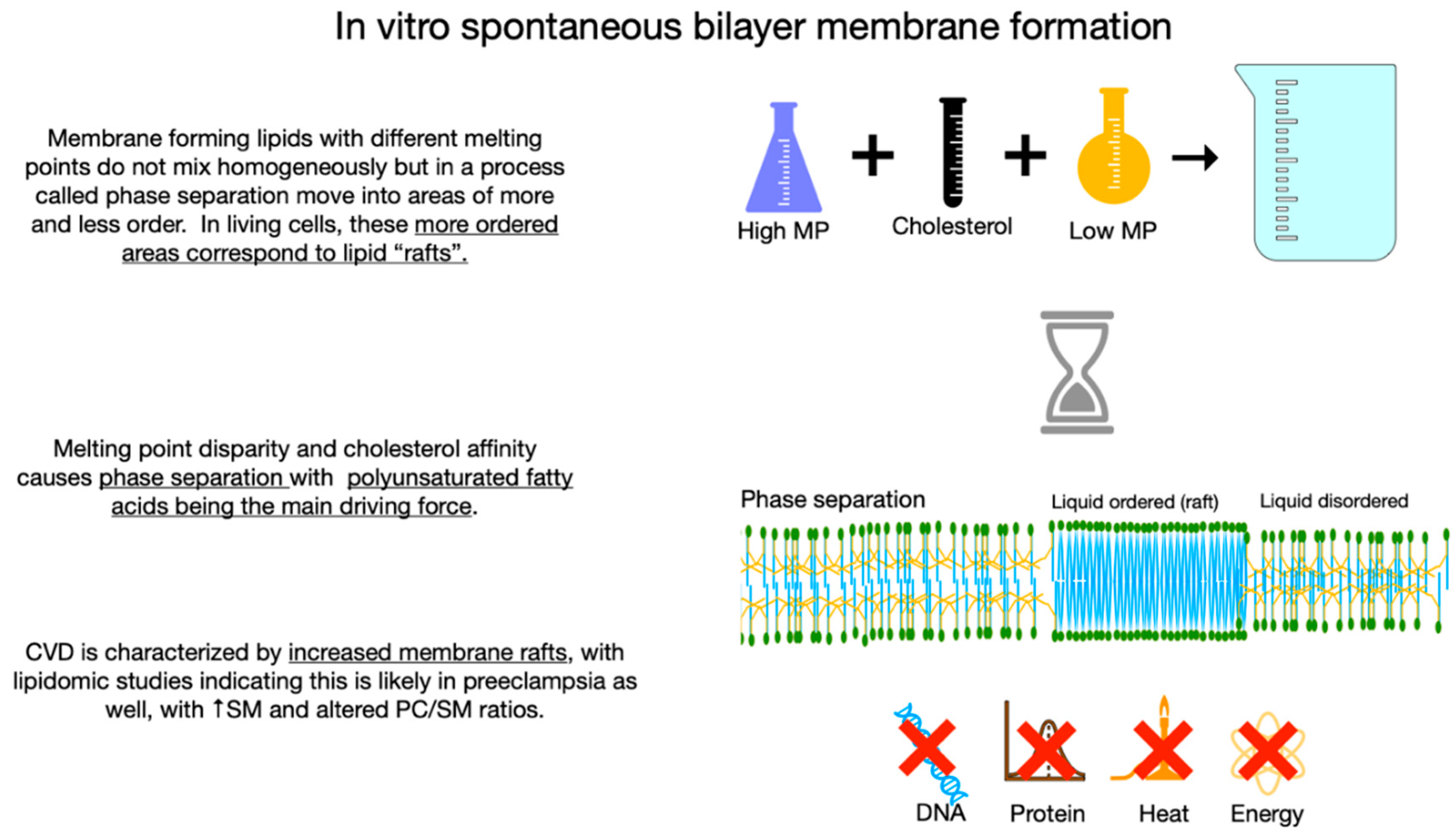
9. Melting Point Disparity, Membrane Rafts, and Cholesterol Content
In living cells, membrane rafts are defined as small, dynamic, sterol and sphingolipid enriched heterogeneous domains that compartmentalize cell membrane processes and serve as organizing centers for a complex network of signaling activity. Compared to more fluid areas of the cell membrane which are enriched in PUFA with very low cholesterol affinity, rafts are enriched in LCSFAs with a high cholesterol affinity, thus rafts represent areas of increased cholesterol[103]. Lipid rafts roughly correspond to liquid-ordered phases in model membrane systems. In these systems greater disparity in the melting point of membrane lipids favors increased phase separation and raft formation, meaning PUFAs with very low melting points are major driving forces for phase separation whilst monounsaturated fatty acids inhibit raft formation[104]. Animal studies show increased PUFA results in changes in membrane lateral organization causing larger rafts which function poorly as signaling platforms[105]. In human CVD, membrane rafts are increased in number and/or stability[106], with lipidomic evidence of a similar increase in preeclampsia[107].
Segregation of lipids into raft areas and non-raft areas may create microdomains with features which reduce trans membrane oxygen diffusion. The high unsaturation index, increased cytoskeletal elements, and stiffness of non-raft areas reduce oxygen diffusion. High cholesterol content, increased proteins, and high cholesterol affinity molecules such as LCSFA in raft areas also reduce oxygen diffusion. If the cholesterol content of raft areas is greater than 50%, cholesterol may precipitate as crystals or form cholesterol/cholesterol bilayer areas which reduce transmembrane oxygen diffusion to about 10% that of healthy membranes. In this way, high levels of dietary PUFA may serve as the driving force for cellular hypoxia in preeclampsia. (Figure 6A and Figure 6B).
Figure 6.
A: Oxygen Diffusion in Healthy Membranes. Healthy membranes present almost no barrier to oxygen diffusion. With primarily moderate melting point lipids, moderate cytoskeletal elements, less than 1% PUFA, small rafts, and minimal molecules with very high cholesterol affinity such as LCSFA and trans unsaturated fatty acids, the total cholesterol content of a healthy membrane is moderate. The cholesterol which is present is associated with molecules with low or moderate cholesterol affinity, so is accessible and available for signaling.
Figure 6.
A: Oxygen Diffusion in Healthy Membranes. Healthy membranes present almost no barrier to oxygen diffusion. With primarily moderate melting point lipids, moderate cytoskeletal elements, less than 1% PUFA, small rafts, and minimal molecules with very high cholesterol affinity such as LCSFA and trans unsaturated fatty acids, the total cholesterol content of a healthy membrane is moderate. The cholesterol which is present is associated with molecules with low or moderate cholesterol affinity, so is accessible and available for signaling.
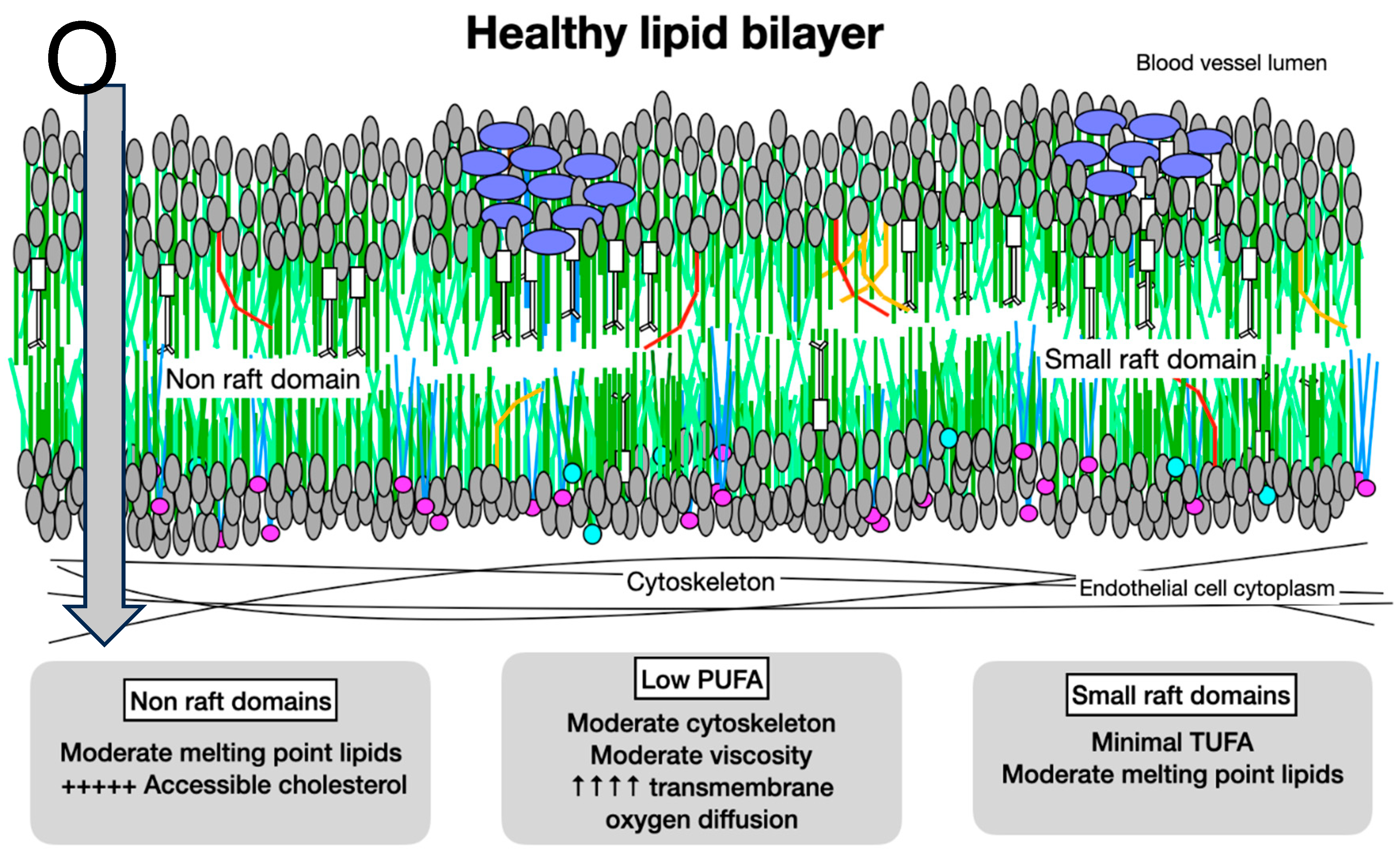
Figure 6.
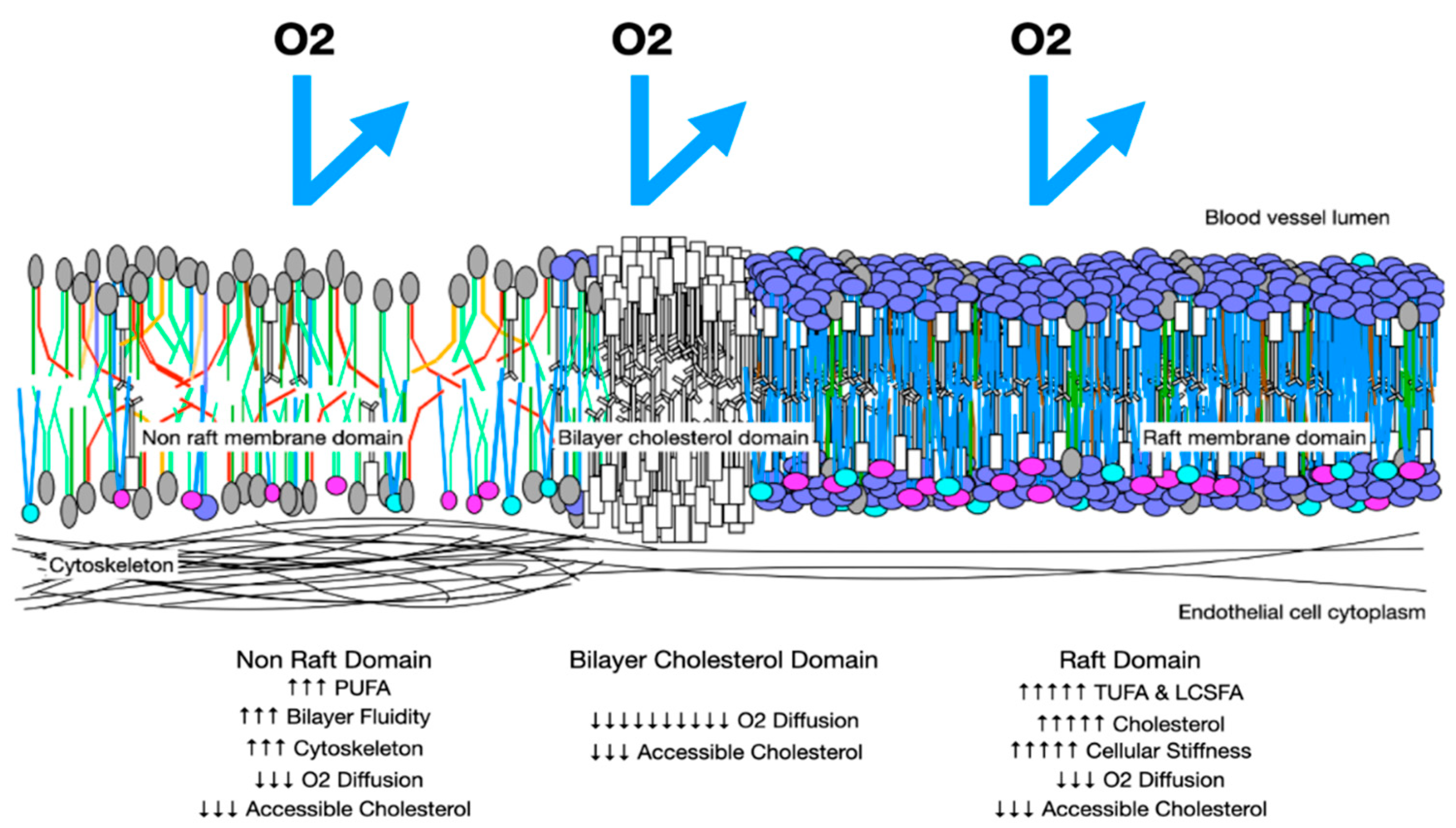
B: Oxygen Diffusion in Preeclampsia Membranes. In preeclampsia, membranes have more rafts with increased cholesterol and LCSFAs which increase membrane thickness and reduce trans-membrane oxygen diffusion. Areas with increased PUFA may have increased cytoskeletal elements and increased stiffness. These factors reduce trans-membrane oxygen diffusion. Cholesterol/cholesterol bilayer areas reduce oxygen DM to 10% that of a health membrane.
Figure 6.
B: Oxygen Diffusion in Preeclampsia Membranes. In preeclampsia, membranes have more rafts with increased cholesterol and LCSFAs which increase membrane thickness and reduce trans-membrane oxygen diffusion. Areas with increased PUFA may have increased cytoskeletal elements and increased stiffness. These factors reduce trans-membrane oxygen diffusion. Cholesterol/cholesterol bilayer areas reduce oxygen DM to 10% that of a health membrane.
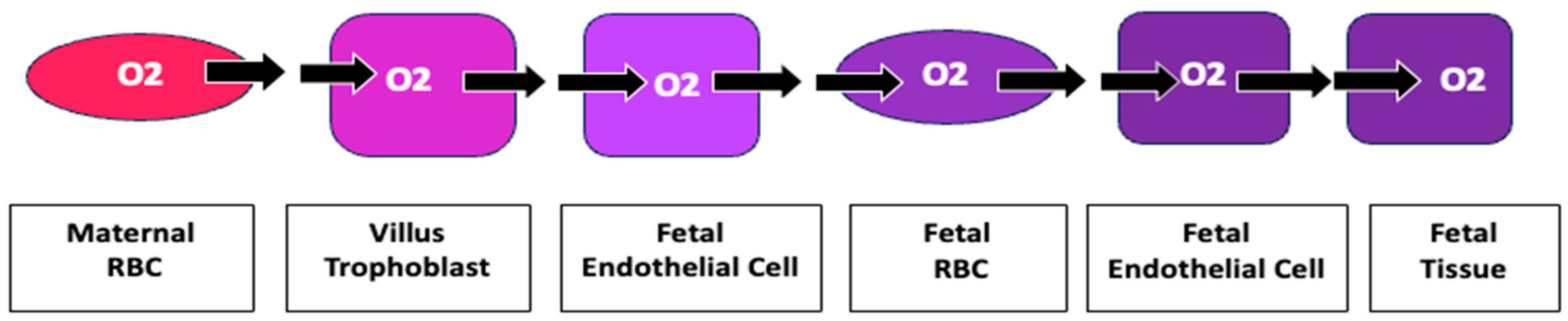
For an oxygen molecule to move from a maternal RBC to fetal tissue, it must pass through ten membranes. Increased impedance to diffusion of oxygen at each membrane provides an explanation for the measured increase in oxygen of maternal blood in the villous space as well as hypoxia of fetal blood (Figure 7).
Figure 7.
Oxygen’s Transplacental Path to Fetal Tissue Moving from a maternal RBC to fetal tissue, an oxygen molecule must pass through ten membranes: 1. maternal RBC, 2. Villous trophoblast maternal lumen side, 3. villous trophoblast stromal side, 4. fetal endothelial cell stromal side, 5. fetal endothelial cell lumen side, 6. fetal RBC at placental villus, 7. fetal RBC at fetal endothelial cell, 8. lumen side fetal endothelial cell, 9. tissue side fetal endothelial cell, 10. fetal tissue cell. Increased impedance at each membrane may become clinically relevant in creating the measured increased oxygenation of maternal villous blood and decreased fetal oxygenation.
Figure 7.
Oxygen’s Transplacental Path to Fetal Tissue Moving from a maternal RBC to fetal tissue, an oxygen molecule must pass through ten membranes: 1. maternal RBC, 2. Villous trophoblast maternal lumen side, 3. villous trophoblast stromal side, 4. fetal endothelial cell stromal side, 5. fetal endothelial cell lumen side, 6. fetal RBC at placental villus, 7. fetal RBC at fetal endothelial cell, 8. lumen side fetal endothelial cell, 9. tissue side fetal endothelial cell, 10. fetal tissue cell. Increased impedance at each membrane may become clinically relevant in creating the measured increased oxygenation of maternal villous blood and decreased fetal oxygenation.
10. Dyslipidemia: Increased Total Cholesterol, Reduced Cholesterol Dependent Signaling
The role of cholesterol in preeclampsia causation is enigmatic. At the systemic level, preeclampsia is associated with dyslipidemia and increased cholesterol in blood lipids, macrophages laden with esterified cholesterol, cholesterol crystals, increased oxysterols, and increased endothelial rafts with high cholesterol content; murine models of preeclampsia may be created by feeding cholesterol at supraphysiologic levels. Yet at the membrane signaling level, cholesterol deficiency is evidenced not only by reduced cholesterol dependent signaling processes such as Hh, Wnt/βcatenin, eNOS but also by increased vascular permeability, reduction in formation of primary cilia, and increased maternal cholesterol synthesis[108].
10. A. Accessible Membrane Cholesterol
The key to this paradox lies in distinguishing free or accessible membrane cholesterol which is available for signaling from sequestered or inaccessible cholesterol which is esterified, oxidized, crystalized, or complexed with other molecules rendering it unavailable for signaling or as an antioxidant[109]. In preeclampsia, seven mechanisms may increase sequestration and reduce maternal membrane cholesterol accessibility: 1) increased lipid raft formation, 2) cholesterol crystallization, 3) increased TUFA, 4) increased oxidative stress with cholesterol serving as an antioxidant by forming oxysterols to reduce free radical propagation, 5) increased cholesterol esters in which oxidized or damaged fatty acids are esterified to cholesterol for removal from cells, 6) formation of oxysterols 7) movement of cholesterol to the fetus and placenta[110,111].
By increasing raft formation, incorporation of PUFA and/or LCSFAs into tissues increases the maternal systemic and placental demands for cholesterol. As outlined in Section 5 and Section 6, raft formation is increased by high melting point disparity of membrane lipids caused by either excessive dietary low melting point lipids such as PUFA or high melting point lipids such as LCSFA. The extremely low cholesterol affinity of PUFA makes it a major driving force in raft formation. Increased PUFA also leads to increased oxidative stress as discussed in Section 10. Environmental molecules such as PFAS and PCB which change membrane viscosity may also drive this process and are associated with abnormal cholesterol homeostasis in humans[112,113].
With PUFA driving an increase in high cholesterol raft areas and incorporation of high cholesterol-affinity TUFA into rafts, localized cholesterol concentrations may rise dramatically causing precipitation of cholesterol crystals. Studies of gall stone formation link increased unsaturation in phosphatidylcholine to nucleation and growth of cholesterol crystals at lower cholesterol concentrations[114]. This mechanism may also account for increased cholesterol crystals found in lipid droplets typical of cardiovascular disease and nonalcoholic steatohepatitis[115]. Presumably, cholesterol precipitated in crystals is not available for membrane signaling [116]. By increasing lipid rafts, sequestration of cholesterol in cell membranes, promoting cholesterol crystallization, and increasing cholesterol oxidation and esterification, dietary PUFA, LCSFA and TUFA may cause the reduction in accessible cholesterol that is evident in preeclampsia.
Cholesterol is transferred from maternal circulation to the placenta, released from the placenta to the umbilical circulation and taken up by the fetus, with fetal cholesterol uptake independent of maternal cholesterol levels[117]. Early primate studies show around 42% of fetal serum cholesterol originates in the maternal circulation with a net cholesterol flux strongly from mother to fetus. Transplacental transfer of cholesterol from mother to fetus reduces maternal accessible membrane cholesterol and may partially explain the rapid resolution of preeclampsia following delivery.
10. B. Hedgehog Signaling in Preeclampsia
With normal placental development there is deep migration of placental cytotrophoblasts into maternal uterine spiral arteries with extensive endothelialization creating low resistance vascular sinuses and a broad interface between the separate maternal and fetal circulations. In contrast, in preeclampsia placental development is compromised with failure of cytotrophoblasts to transform from a proliferative epithelial subtype into an invasive endothelial subtype causing maternal vessels to remain narrow with reduced placental connection[118] causing placental hypoxia.
Control of placental development lies in a complex balance of angiogenic, antiangiogenic, and apoptotic factors with Vascular Endothelial Growth Factor (VEGF) as the nexus for a cascade of signaling[119]. Activation of the VEGF pathway results in cell division, migration, angiogenesis, and embryonic development. Positioned upstream of VEGF, Hedgehog (Hh) signaling is a key regulator of embryonic and placental development. Both VEGF and Hh signaling localize to projections from maternal endothelial cells called primary cilia which in preeclampsia are shorter, fewer in number[120], with diminished Hh signaling[121]. Placental Hh signaling which is downregulated in preeclampsia is highly correlated with fetal birth weight in murine models[122]. With Hh so intimately involved in all forms of animal growth and development, finding the mechanism controlling Hh signaling has been a top research goal for decades. Surprisingly, highly abundant cholesterol is the primary regulator of Hh signaling. However, it is not total cholesterol, but the availability of free or accessible membrane cholesterol which positively regulates Hh signaling [123,124]. Low accessible cholesterol may also explain the shortening, reduction in number and impaired function of maternal endothelial cell cilia[125].
10. C. Wnt/βcatenin Signaling
Acting on cell proliferation, migration, adhesion, and survival, Wnt/βcatenin signaling is also critical in mediating angiogenesis[126]. In preeclampsia, abnormalities in Wnt/βcatenin signaling lead to severe defects in placental development[127,128]. Wnt/βcatenin signaling may be divided into a canonical pathway which promotes proliferation and growth and a noncanonical pathway which promotes apoptosis, a critical factor in reduced placental development of preeclampsia[129]. In Wnt/βcatenin signaling, accessible cholesterol switches signaling from noncanonical to canonical pathways[130]. In this way a deficit of accessible cholesterol promotes trophoblast apoptosis while adequate accessible cholesterol promotes angiogenesis.
10. D. Endothelial Nitric Oxide Synthase Signaling
Synthesized from L-arginine in a reaction catalyzed by nitric oxide synthases, the soluble gas nitric oxide (NO), plays a role in regulating inflammation, oxidative stress, and vasodilation. Most studies show reduced NO activity in preeclampsia with diminished peripheral endothelial vasodilation and endothelial nitric oxide synthase (NOS3) activity[131] to which oxidative stress contributes[132]. Primarily located in caveolae[133], a type of membrane raft, NOS3 activity is ablated with application of methyl-β-cyclodextrin (MβCD) which selectively extracts cholesterol from the plasma membrane. After MβCD treatment, electron microscopy shows caveolae are no longer present, yet both caveolae and eNOS activity are restored with addition of cholesterol[134]. In murine models, statins may restore eNOS activity by reducing expression of caveolin-1, an NOS3 inhibiter[135]; in human monocytes statins redistribute cholesterol from non-caveolar areas of the cell membrane to caveolae[136]. Treatment of caveolae with oxidized LDL also ablates NOS3 activity, which can be restored by cholesterol treatment[137,138,139]. Studies of mice with absent NOS3 function show an exaggerated response to VEGF blocking factors with increased hypertension, hepatic dysfunction, thrombocytopenia, and renal dysfunction[140], all symptoms of human preeclampsia.
Each of these critical signaling pathways is downregulated in preeclampsia. A deficiency of accessible membrane cholesterol which may be induced by dietary lipid changes may cause this deficit.
10. E. Dyslipidemia and Accessible Cholesterol
In a reductionist view of lipoproteins, Low Density Lipoprotein (LDL) carries cholesterol from its site of synthesis in the liver to peripheral tissues where LDL receptors (LDLR) are produced by cells in response to deficient cholesterol. HDL cholesterol carries excess cholesterol from peripheral cells to the liver. Those with the highest levels of LDL cholesterol and the lowest levels of HDL cholesterol are at increased risk of preeclampsia and CVD. Cells with a cholesterol deficit are not able to produce an excess of cholesterol for export to HDL, therefore exhibit reduced cellular cholesterol efflux and increased cholesterol synthesis. Increased cholesterol synthesis and upregulation of LDLR are considered “pathologic” or “inappropriate” in CVD and preeclampsia because the total cholesterol content of peripheral cells including endothelial cells is elevated. However, because the regulation of cholesterol synthesis and LDLR formation are dependent on accessible membrane cholesterol rather than total cholesterol, upregulation of cholesterol and LDLR synthesis is appropriate to a situation with increased sequestered cholesterol but deficient free cholesterol, as occurs in diets with high levels of PUFA, TUFA, or LCSFAs.
Statins, medications prescribed for treatment of hypercholesterolemia, are marketed as inhibiters of hydroxymethylglutaryl-CoA reductase (HMG-CoAR) enzyme, a rate limiting step in cholesterol synthesis. Statins are effective in lowering total cholesterol, LDL, and triglyceride concentrations while increasing HDL concentrations. Pleiomorphic effects include immunomodulatory, antioxidant, anti-inflammatory, stabilization of plaque, and reduction of platelet aggregation.[141] Although blocking HMG-CoAR implies a reduction in cholesterol production, early cholesterol turnover studies with long term lovastatin therapy showed increased expression of HMG-CoAR as well as the LDL receptor and no significant change of total body exchangeable cholesterol[142]. In mice studies, statins robustly increase cholesterol synthesis primarily in the liver, stimulate fecal cholesterol elimination, increase expression of sterol regulatory element binding protein 2 (SREBP2) the transcription factor regulating cholesterol synthesis, and strongly induce gene expression of cholesterol synthesis enzymes with an increase in total body cholesterol synthesis[143]. A dietary study on the effect of dietary cholesterol found “The concentration of LDL-C (LDL cholesterol) increases in plasma when the daily intake of cholesterol was less than 900 mg, while the elevation of HDL-C, which has anti-atherogenic effects, occurred only in the range from 650 to 900 mg/day, and not when below 650 mg/day.”[144] That is, reduced dietary cholesterol increases dyslipidemia. Other studies show a high intake of eggs, one of the most concentrated dietary sources of cholesterol, is associated with beneficial serum lipid profiles[145]. Oxysterols which are formed by oxidation of cholesterol and are much more potent than cholesterol in down regulating cholesterol synthesis are reduced by statins[146]. Statins also increase serum monounsaturated fatty acids and reduce serum PUFA[147]. These studies raise the possibility the mode of action of statins is via increased accessible cholesterol by reduction of cholesterol sequestration in membrane rafts rather than by inhibition of cholesterol synthesis. By reducing serum PUFA, statins reduce raft formation and oxidative stress. With adequate accessible cholesterol available to esterify damaged (oxidized) fatty acids for removed from endothelial membranes and transport to the liver for disposal, membrane homeoviscosity may normalize.
11. Dietary Lipids and Oxidative Stress
Defined as an imbalance between oxidants and antioxidants, oxidative stress has long been recognized as a major element in preeclampsia pathology[148,149,150]. Oxidative stress occurs when free radicals, molecules containing an unpaired electron such as reactive oxygen and nitrogen species, interact with biological molecules causing damage. Some exogenous origins of free radicals are ultraviolet radiation, ionizing radiation, and heavy metals[151], with the primary endogenous source being the electron transport chain in mitochondria[152]. Because the surrounding lipid environment may markedly change a protein’s shape and function[153,154], variations in dietary lipids may alter mitochondrial lipids[155] causing conformational shifts in the structure of membrane proteins[156,157,158]. In this way, diet induced changes in lipids may distort electron transport proteins causing the escape of unpaired electrons and increased oxidative stress.
PUFA, being much more vulnerable to oxidation than are monounsaturated fatty acids and saturated fatty acids, markedly increases lipid, protein, and mitochondrial DNA oxidative damage[159]. Like the proliferation of nuclear fission in an atomic reactor, after absorbing a free radical PUFAs release more free radicals and propagate oxidation[160]. By absorbing a free radical without propagation, antioxidants such as glutathione peroxidase, superoxide dismutase, coenzyme Q10, selenium, vitamin C, Vitamin E, and cholesterol[161,162,163] slow the production of reactive species.
The biological consequences of reactive oxygen species include many pathways active in preeclampsia pathology including: 1) activation of redox-sensitive transcription factors such as NFKβ, COX-2, AP-1, and interleukin 1β which regulate gene transcription in response to cytokines, growth factors, stress, infections and also regulate inflammation, immunity, differentiation, proliferation, growth and survival, 2) increased TNF-⍺ an inflammatory cytokine, 3) activation of protein kinases such as MAPK, SAPK-JNK, and ASK1 which coordinate cell proliferation, differentiation, mobility, and survival, 4) increased levels of the soluble receptor for vascular endothelial growth factor (sFlt-1) which blocks the action of VEGF 5) increased VEGF, 6) loss of intracellular calcium homeostasis with increased unfolded protein response, endoplasmic reticulum and mitochondrial stress, and apoptosis, 7) lipid peroxidation induced change in membrane fluidity and function, 8) oxidation of proteins with both loss and gain of function and misfolding, and 9) nuclear and mitochondrial DNA damage[164,165,166].
The relationship between increased oxidative stress and preeclampsia has been noted for decades, yet the fundamental cause of the imbalance between antioxidants and free radicals remains unclear. Clinical trials using varied antioxidants to treat preeclampsia show no reduction in preeclampsia or preterm birth[167,168,169], leading a recent research team to ask, “Antioxidants in Pregnancy: Do We Really Need More Trials?”[170]. Studies of antioxidants in maternal blood are inconsistent with findings of both reduced and increased antioxidant levels[171,172]. It appears excess oxidative stress in preeclampsia arises from excess reactive oxygen and nitrogen species which may be generated from excess PUFA rather than primarily from a deficiency in antioxidants.
Because oxysterols produce a stronger inhibition of Hydroxymethylglutaryl-CoA reductase than does cholesterol, the presence of increased oxidative stress may reduce cholesterol synthesis and accessible cholesterol. Oxidized lipids also behave differently in cell membranes with oxysterols and oxidized PUFA producing a thinner and more fluid membrane which further reduces movement of oxygen across the membrane when compared to unoxidized molecules.
12. Lipids in Preeclampsia Screening and Epidemiology
Several factors are widely recognized to increase the risk of developing preeclampsia including previous preeclampsia, chronic hypertension, pregestational diabetes, multiple gestation, pre pregnancy BMI > 30, and antiphospholipid syndrome with lesser increased relative risk attributed to history of lupus erythematosus, stillbirth, pre pregnancy BMI > 25, nulliparity, prior placental abruption, assisted reproductive technology, chronic kidney disease, advanced maternal age and genetic susceptibility.[173] Although less generally noted, abnormal maternal serum lipids also convey increased risk for preeclampsia, with reduced maternal serum omega-3 fatty acids increasing relative risk 7.6 times, [174,175,176] elevated maternal red blood cell (RBC) TUFA increasing relative risk 3-7.4 times, [177,178] and high dietary intake of total PUFA increasing relative risk by 2.6 to 5 times. [179,180] Lipidomic studies reflect this pattern with placentae from affected women containing elevated levels of TUFA[181] and total PUFA.[182,183] Although there are exceptions[184], several studies show dietary TUFA intake is positively associated with preeclampsia incidence[185,186]. (Table 1)

Table 1.
Preeclampsia Risk Factors Compared to standard preeclampsia risk factors such as chronic hypertension, pregestational diabetes, multiple gestation, BMI>30, and antiphospholipid antibodies, lipid related factors such as elevated PUFA and TUFA and low omega 3 fatty acids confer substantial risk.
Table 1.
Preeclampsia Risk Factors Compared to standard preeclampsia risk factors such as chronic hypertension, pregestational diabetes, multiple gestation, BMI>30, and antiphospholipid antibodies, lipid related factors such as elevated PUFA and TUFA and low omega 3 fatty acids confer substantial risk.
| Condition | Relative risk |
|---|---|
| Previous pregnancy with preeclampsia | 8.4 |
| Chronic hypertension | 5.1 |
| Pregestational Diabetes | 3.7 |
| Multiple gestation | 2.9 |
| Pre-pregnancy BMI > 30 | 2.8 |
| Anti-phospholipid Antibodies | 2.8 |
| Reduced maternal serum omega-3 fatty acids | 7.6 |
| Increased maternal RBC TUFA | 3 to 7.4 |
| Increased dietary PUFA | 2.6 to 5 |
Membrane-forming lipids such as phospholipids, ceramides, and sphingomyelins have significantly different patterns in maternal and placental tissues in those with preeclampsia versus controls [187,188,189,190,191,192] . This marked lipid dysregulation supports their use in screening for preeclampsia[193,194]. A 2021 pilot study measuring the leptin/ceramide ratio outperformed the more established sFlt-1/PlGF ratio in predicting preeclampsia with superior detection and sensitivity[195]. Using maternal serum metabolic biomarkers including sphingolipids in mid trimester also performed better than sFlt-1/PlGF ratio in screening[196].
13. Diets and Preeclampsia
13. A. Diets Affect Preeclampsia Incidence
Because biologic membranes adapt their lipid composition in response to dietary lipids[197], dietary lipid variance may be the basis for the lipidomic changes of preeclampsia. The important role of diet in preeclampsia pathology is suggested by a 2023 meta-analysis of the effects of a high Dietary Inflammatory Index (DII) which indicates that each 1 unit increase in the DII score is associated with a 24% higher risk of preeclampsia[198]. A 2001 study from Norway showed a high dietary intake of energy, sucrose, and PUFA is associated with increased risk of preeclampsia[199]. A 2023 study of 106,808 pregnant US military personnel without preeclampsia risk factors on a standard military diet with 7% of the total caloric intake as PUFA and higher PUFA for those at high risk for CVD, shows a preeclampsia rate of 9.5 to 12% in all ethnic groups[200] compared to 3 to 8% rate in civilian populations. The Danish National Birth Cohort evaluated 66,738 pregnancies from 1996-2002 in which 0.03% were vegans (n=18). The rate of preeclampsia in omnivores was 2.6% with the rate in vegans 11.1%[201]. A meta-analysis of the effect of the Mediterranean Diet showed a reduction in preeclampsia risk with adherence to the Mediterranean Diet[202]. These studies emphasize the crucial role diet may play in preeclampsia pathology and the need to consider the role of dietary lipids in metabolic pathways, screening, and treatment.
13. B. Increased Dietary PUFA Increases Preeclampsia Incidence
Metabolic studies indicate a relationship between increased unsaturation index and preeclampsia. A 2005 study measuring urinary excretion of isoprostane, an indicator of oxidative damage to lipids, found increased dietary fat and PUFA correlate with increased urinary isoprostane excretion and a fivefold increase in preeclampsia risk[203]. A 2021 study from Indonesia with one of the world’s highest preeclampsia rates shows a positive correlation between maternal serum PUFA and preeclampsia incidence. “Severe preeclampsia showed the highest concentration of total PUFA and the lowest DHA percentage, with significantly higher Omega-6/Omega-3 ratio (p = 0.004) and lower omega-3 index (p < 0.002) compared to control.” [204] Fatty acid desaturases, which are the rate limiting steps in metabolism of PUFA, are higher in maternal RBCs of those with preeclampsia compared to controls[205].
13. C. Dietary Intervention Studies
Although prospective studies of dietary lipid manipulation in pregnancy are uncommon, available studies are encouraging. In a group of 800 subjects, a 2020 randomized controlled study reducing dietary trans fatty acids showed a reduction in TUFA between study groups which was accompanied by a significant reduction in preeclampsia risk (OR 0.56)[206]. A Norwegian study of probiotic milk intake in late pregnancy was significantly associated with lower preeclampsia risk (OR 0.80)[207]. A meta-analysis of nine studies using olive oil supplementation also showed reduced risk of preeclampsia[208]. Increased dietary fiber has consistently shown reduced preeclampsia risk[209] perhaps through the mechanism of improving gut dysbiosis and increasing production of short chain saturated fatty acids[210,211,212]. Supplementation of omega-3 fatty acids has had mixed results with initial studies showing no benefit, but a 2018 Cochran meta-analysis showing possible reduction of preeclampsia risk with a relative risk of 0.84 in general population with low quality evidence, but significant protection among women with high risk [213]. Weight management interventions in pregnancy also reduced preeclampsia risk with a meta-analysis of 88 studies showing a significant decrease in the risk of preeclampsia (RR 0.67)[214]. However, in a population with primarily a low-fat, high-carbohydrate diet which is likely deficient in choline, supplementation of PUFA reduced preeclampsia relative risk to 0.40.[215] Although the Effect of Simple, Targeted Diet in Pregnant Women With Metabolic Risk Factors on Pregnancy Outcomes (ESTEEM) trial treating women with metabolic risk factors with a Mediterranean-style diet showed no reduction in preeclampsia[216], a similar trial including enhanced extra virgin olive oil and pistachios in addition to the Mediterranean Diet did reduce composite maternal and fetal complications including preeclampsia although the target for olive oil intake was not met[217]. Overall, these studies suggest a combination strategy to reduce TUFA, PUFA, and the omega 6:3 ratio may be an effective strategy to reduce preeclampsia incidence.
13. D. Choline deficiency and Preeclampsia
Choline is an essential factor which may be synthesized in small quantities endogenously from phosphatidylethanolamine, although genetic polymorphisms interfere with synthesis. To prevent deficiency, it must be obtained exogenously[218] . With the best dietary sources of choline being animal-based foods, choline deficiency occurs more frequently in vegans. The National Health and Nutrition Examination Survey (NHANES) in the United States indicates fewer than 11% of pregnant people have adequate intake [219,220]. During pregnancy, choline requirements rise as choline is passed from maternal to fetal tissues with human placentae containing around 50 times more choline than the maternal blood[221]. Those with choline deficiency who are not pregnant exhibit symptoms like preeclampsia with liver changes such as hepato-steatosis with elevated transaminases[222,223], renal cortical necrosis with lipid accumulation and reduced reabsorption of water[224], as well as cardiovascular effects such as hypertension[225].
Choline is necessary for synthesis of phosphatidylcholine as well as sphingomyelin. In a choline deficient diet with a high carbohydrate intake, fewer phospholipid headgroups available to esterify to fatty acids may cause lengthening of fatty acid chains and an increased in LCSFAs. Rodent models confirm this analysis with lipidomics of choline deficient animals confirming major reductions in serum phospholipids and sphingolipids with a choline deficient diet which are restored after treatment with phosphatidylcholine[226].
13. E. Preeclampsia with PUFA Deficiency
Although many studies implicate excessive PUFA’s involvement in increased preeclampsia risk, dietary studies in populations with much different diets than the United States indicate potential involvement of LCSFA and/or deficient choline. For example, in low to moderate income populations with a baseline high carbohydrate, low protein, low PUFA diet, supplementation with PUFA shows a significant effect in reducing relative risk of preeclampsia[227]. In another population with a similar diet, use of a lipid based high PUFA nutritional supplement in underweight women diagnosed with preeclampsia found significant improvements in birthweight, gestational age at delivery, head circumference, and birth length[228]. To understand how these dietary interventions fit with the lipid raft model of preeclampsia outlined in this review, consideration must be given to the effects of excessive LCSFA with deficient PUFA and choline on membrane biophysics.
Returning to Figure 4 which illustrates homeoviscous adaptation with an excessively fluid lipid bilayer, the Leventhal experiments show the essential compensatory action of adding LCSFA and cholesterol to return the membrane to normal viscosity. In situations with choline and PUFA deficiency creating an excess of LCSFAs as may be found in high carbohydrate/low protein diets, membrane lipid bilayers may be excessively stiff. In this setting, the addition of PUFA in the form of essential fatty acids increases fluidity and restores membrane homeoviscosity. With a large membrane melting point disparity, lipid rafts and cholesterol sequestration are increased, creating preeclampsia without prominent elements of oxidative stress. From these studies, we may conclude the critical factor in the etiology of preeclampsia is perhaps the balance of low, moderate, and high melting point lipids and cholesterol rather than absolute values or deficiencies.
13. F. Dietary Cholesterol
Since the 1970s when the United States Department of Agriculture first published guidelines recommending limitation of dietary cholesterol to reduce cardiovascular disease until these guidelines were withdrawn in 2015, the role of dietary cholesterol in causation of CVD has been vigorously debated. More recent analyses not only confirm the absence of harm, but also show beneficial effects of dietary cholesterol challenges on human plasma lipoprotein subfractions[229]. In preeclampsia, one study shows a statistically significant lower dietary cholesterol intake in human subjects with preeclampsia (361.48 mg/day) versus controls (407.88 mg/day)[230]. A source of confusion in the role of cholesterol in dietary studies may arise from dietary questionnaires which frequently group dietary cholesterol, oxysterols, long-chain saturated fatty acids and trans fatty acids together under the category of “Western Style Diet” which is associated with an increase in preeclampsia incidence[231], thus, dietary cholesterol is condemned by association rather than causation.
14. Discussion
Consideration of the role of lipids in preeclampsia pathology present four major goals of lipid therapy:
- Reduce oxidative stress With very low melting-point PUFA as the driving force for oxidative stress, replacing dietary PUFA with medium-chain saturated fatty acids such as lauric acid (C12:0) in coconut oil, monounsaturated fatty acids such as oleic acid (C18:1) in olive oil, or short-chain saturated fatty acids such as butyric (C2-5:0) which are found in milk products or as products of gut bacteria should markedly reduce oxidative stress as these lipids are less vulnerable to oxidative damage. Avoiding dietary oxysterols which are found in dehydrated milk, eggs, and proteins also reduces oxidative stress[232]. These measures should lower blood pressure, restore normal angiogenic signaling, and reduce protein misfolding.
- Reduce lipid raft formation and hypoxia Reduction of dietary PUFA reduces disparity in membrane lipid melting points and reduces the need for endogenous LCSFA synthesis, thus reducing lipid raft formation and hypoxia. Reduction of membrane PUFA also reduces cholesterol crystallization, cholesterol/cholesterol bilayer domains which markedly impair the movement of oxygen across placental membranes. LCSFAs such as palmitoleic (C:16) and stearic acid (C18:0) found in beef, pork, lamb, and chicken may also stimulate excessive lipid raft formation and are elevated in some with preeclampsia[233,234], so intake should be low or moderate. supplementation of moderate melting point fatty acids to inhibit raft formation and of choline in those who are deficient should be beneficial[235].
- Increase accessible membrane cholesterol Increasing dietary cholesterol allows esterification and removal of damaged fatty acids as well as reducing oxidative stress. Reducing dietary TUFA by avoiding foods cooked at high temperature in oil and meats produced with feeds using spent deep fryer oils should reduce cholesterol sequestration in rafts and cholesterol esters. Supplementing dietary cholesterol provides accessible cholesterol to restore normal Wnt, Hh, and eNOS signaling, aids angiogenesis, improves vasodilation, and restores normal membrane rheology and permeability.
- Reduce the omega 6:3 ratio to less than 4:1 Monitoring the omega 6:3 ratio allows determination of cases in which an elevated 6:3 ratio (> 4:1) is clinically significant and whether supplementation has been effective. This measure should reduce inflammation and thrombosis.
Strategies to meet the goals of reducing oxidative stress, hypoxia, lipid raft formation, cholesterol sequestration, and optimizing the omega 6:3 ratio can be met over long periods of time through diets high in fruits and vegetables with reduced intake of total fats, PUFA, sugar, processed and high fat meats, deep fried foods, oxysterols, and excess calories[236]. Both the Mediterranean Diet and the New Nordic Diet have been effective in reducing preeclampsia. Ideally, months before conception, a person contemplating pregnancy would begin an optimal diet to reduce risk of preeclampsia. However, dietary change is difficult as evidenced by the high rate of obesity in the United States and the effects of cooking techniques have not yet been widely incorporated into dietary advice. Expansion of previous successful clinical trials to reduce preeclampsia which are aimed at reducing trans fatty acids, improving the omega 6:3 balance, and reducing excessive PUFA in those contemplating pregnancy provides a starting point to prevent preeclampsia.
Because dietary lipids are incorporated into endothelial cell membranes within hours of ingestion, there is reason for optimism that lipid rescue therapy for mothers hospitalized with preeclampsia may very rapidly affect a patient’s status within a clinically relevant time. With the volume of lipids in maternal endothelial cells, placenta, and transferred to the fetus increasing as gestation advances and because very premature deliveries carry higher fetal risks, perhaps the initial proof-of-concept protocol should target early preterm (< 30 weeks) gestations as lipid treatment is most likely to be successful in this situation.
15. Conclusions
This review explores the role dietary lipids may play in creating placental hypoxia and reducing accessible membrane cholesterol and cholesterol dependent signaling. By changing membrane viscosity and raft formation, dietary lipids, environmental factors such as inhaled hydrocarbons, and membrane viscosity modifiers such as pfas may reduce movement of oxygen across membranes, sequester cholesterol, and modify raft related signaling. Metabolomic, metabolic and interventional studies demonstrate close correlations between dietary lipid intake, environmental membrane modulators and preeclampsia risk. The encouraging results of interventional trials merits further investigation of the use of dietary lipids to prevent and treat preeclampsia.
References
- Williams, M.A.; King, I.B.; Sorensen, T.K.; Zingheim, R.W.; Troyer, B.L.; Zebelman, A.M.; Luthy, D.A. Risk of preeclampsia in relation to elaidic acid (trans fatty acid) in maternal erythrocytes. Gynecol Obstet Invest. 1998, 46, 84–7. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T.; Slott, M.; Solvoll, K.; Drevon, C.A.; Vollset, S.E.; Henriksen, T. High intake of energy, sucrose, and polyunsaturated fatty acids is associated with increased risk of preeclampsia. Am J Obstet Gynecol. 2001, 185, 451–8. [Google Scholar] [CrossRef] [PubMed]
- Irwinda, R.; Hiksas, R.; Siregar, A.A.; Saroyo, Y.B.; Wibowo, N. Long-chain polyunsaturated fatty acid (LC-PUFA) status in severe preeclampsia and preterm birth: a cross sectional study. Sci Rep. 2021, 11, 14701. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.A.; Zingheim, R.W.; King, I.B.; Zebelman, A.M. Omega-3 fatty acids in maternal erythrocytes and risk of preeclampsia. Epidemiology. 1995, 6, 232–7. [Google Scholar] [CrossRef] [PubMed]
- Wojcik-Baszko, D.; Charkiewicz, K.; Laudanski, P. Role of dyslipidemia in preeclampsia-A review of lipidomic analysis of blood, placenta, syncytiotrophoblast microvesicles and umbilical cord artery from women with preeclampsia. Prostaglandins Other Lipid Mediat. 2018, 139, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Poornima, I.G.; Indaram, M.; Ross, J.D.; Agarwala, A.; Wild, R.A. Hyperlipidemia and risk for preclampsia. J Clin Lipidol. 2022, 16, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Odenkirk, M.T.; Stratton, K.G.; Gritsenko, M.A.; Bramer, L.M.; Webb-Robertson, B.M.; Bloodsworth, K.J.; et al. Unveiling molecular signatures of preeclampsia and gestational diabetes mellitus with multi-omics and innovative cheminformatics visualization tools. Mol. Omics. 2020, 16, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Melland-Smith, M.; Ermini, L.; Chauvin, S.; Craig-Barnes, H.; Tagliafetto, A.; Todros, T.; et al. Disruption of sphingolipid metabolism augments ceramide-induced autophagy in preclampsia. Autophagy 2015, 11, 653–669. [CrossRef]
- Brown, S.H.; Eather, S.R.; Freeman, D.J.; Meyer, B.J.; Mitchell, T.W. A Lipidomic Analysis of Placenta in Preeclampsia: Evidence for Lipid Storage. PLoS One. 2016, 11, e0163972. [Google Scholar] [CrossRef]
- Del Gaudio, I.; Sasset, L.; Lorenzo, A.D.; and Wadsack, C. Sphingolipid signature of human feto- placental vasculature in preeclampsia. Int. J. Mol. Sci. 2020, 21, 1019. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Y.; Lv, Y.; Ding, H. Dissecting the Roles of Lipids in Preeclampsia. Metabolites. 2022, 12, 590. [Google Scholar] [CrossRef] [PubMed]
- Al, M.D.; van Houwelingen, A.C.; Badart-Smook, A.; Hasaart, T.H.; Roumen, F.J.; and Hornstra, G. The essential fatty acid status of mother and child in pregnancy-induced hypertension: A prospective longitudinal study. Am. J. Obstet. Gynecol. 1995, 172, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Young, S.; Esplin, M.S.; Peaden, B.; Tolley, H.D.; Porter, T.F.; et al. Detection and confirmation of serum lipid biomarkers for preeclampsia using direct infusion mass spectrometry. J. Lipid Res. 2016, 57, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Korkes HA, et al Lipidomic assessment of plasma and placenta of women with early-onset preeclampsia. PLoS One. 2014, 9, e110747.
- Baig, S.; Lim, J.Y.; Fernandis, A.Z.; Wenk, M.R.; Kale, A.; Su, L.L.; et al. Lipidomic analysis of human placental syncytiotrophoblast microvesicles in adverse pregnancy outcomes. Placenta 2013, 34, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Thangaratinam, S.; Rogozińska, E.; Jolly, K.; Glinkowski, S.; Duda, W.; Borowiack, E.; Roseboom, T.; Tomlinson, J. ; J; Kunz, R. ; Mol, B.W.; Coomarasamy, A.; Khan, K.S. Interventions to reduce or prevent obesity in pregnant women: a systematic review. Health Technol AssWalczakess. 2012, 16, iii–iv, 1. [Google Scholar] [CrossRef] [PubMed]
- Alamolhoda, S.H.; Simbar, M.; Mirmiran, P.; and Mirabi, P. Effect of low trans-fatty acid intakes on preeclampsia: A randomized controlled trial. J. Res. Med. Sci. 2020, 25, 112. [Google Scholar] [CrossRef] [PubMed]
- Hedegaard, S.; Nohr, E.A.; Olsen, S.F.; Halldorsson, T.I.; Renault, K.M. Adherence to different forms of plant-based diets and pregnancy outcomes in the Danish National Birth Cohort: A prospective observational study. Acta Obstet Gynecol Scand. 2024 Jan 24. Epub ahead of print. [CrossRef] [PubMed]
- Irwinda R, Hiksas R, Siregar AA, Saroyo YB, Wibowo N. polyunsaturated fatty acid (LC-PUFA) status in severe preeclampsia and preterm birth: a cross sectional study. Sci Rep. 2021, 11, 14701. [CrossRef]
- Kinshella MW, Omar S, Scherbinsky K, Vidler M, Magee LA, von Dadelszen P, Moore SE, Elango R, The Precise Conceptual Framework Working Group. Effects of Maternal Nutritional Supplements and Dietary Interventions on Placental Complications: An Umbrella Review, Meta-Analysis and Evidence Map. Nutrients. 2021, 13, 472. [CrossRef]
- [1] Li SN, Liu YH, Luo ZY, Cui YF, Cao Y, Fu WJ, Dou WF, Duan DD, Zhao XL, Chen YM, Lyu QJ, Chen QS, Zeng FF. The association between dietary fatty acid intake and the risk of developing preeclampsia: a matched case-control study. Sci Rep. 2021, 11, 4048. [CrossRef]
- Bakrania, B.A.; George, E.M.; Granger, J.P. Animal models of preeclampsia: investigating pathophysiology and therapeutic targets. Am J Obstet Gynecol. [CrossRef]
- Colson, A.; Depoix, C.L.; Lambert, I.; Leducq, C.; Bedin, M.; De Beukelaer, M.; Gatto, L.; Loriot, A.; Peers de Nieuwburgh, M.; Bouhna, K.; Baldin, P.; Hubinont, C.; Sonveaux, P.; Debiève, F. Specific HIF-2α (Hypoxia-Inducible Factor-2) Inhibitor PT2385 Mitigates Placental Dysfunction In Vitro and in a Rat Model of Preeclampsia (RUPP). Hypertension. 2023 May;80, 1011-1023. [CrossRef]
- Rajakumar, A.; Whitelock, K.A.; Weissfeld, L.A.; Daftary, A.R.; Markovic, N.; Conrad, K.P. Selective overexpression of the hypoxia-inducible transcription factor, HIF-2alpha, in placentas from women with preeclampsia. Biol Reprod. 2001, 64, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, B. Placental physioxia is based on spatial and temporal variations of placental oxygenation throughout pregnancy. J Reprod Immunol. 2023, 158, 103985. [Google Scholar] [CrossRef] [PubMed]
- Blasbalg, T.L.; Hibbeln, J.R.; Ramsden, C.E.; Majchrzak, S.F.; Rawlings, R.R. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr. 2011, 93, 950–62, Epub 2011 Mar 2. [Google Scholar] [CrossRef]
- Gerrior, S.; Bente, L.; and Hiza, H. (2004). Nutrient content of the U.S. Food supply, 1909-2000. (Home economics research report No. 56), Washington, DC: U.S. Department of Agriculture, Center for Nutrition Policy and Promotion.
- USDA/ERS U.S (2021). Department of agriculture, food availability (per capita) data system, Undated July 21.
- Craig-Schmidt, M.C. World-wide consumption of trans fatty acids. Atheroscler. Suppl. 2006, 7, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Wanders, A.J.; Zock, P.L.; and Brouwer, I.A. Trans fat intake and its dietary sources in general populations worldwide: A systematic review. Nutrients 2017, 9, 840. [Google Scholar] [CrossRef] [PubMed]
- 31. Federal Register, 1: Health and Human Services, Food and Drug Administration Revocation of Uses of Partially Hydrogenated Oils in Foods, Rules and Regulations, Action: Direct final rule: Vol. 88, 2023.
- World Health Organization Replace Trans Fat; an Action Pacakage to Eliminate Industrially produced Trans-Fatty Acids Executive Summary WHO/NMH/NHD/18.4 https://www.who.int/docs/default-source/documents/replace-transfats/replace-action-package.pdf.
- Bhardwaj, S.; Passi, S.J.; Misra, A.; Pant, K.K.; Anwar, K.; Pandey, R.; et al. Effect of heating/reheating of fats/oils, as used by Asian Indians, on trans fatty acid formation. Food Chem. 2016, 212, 663–670. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Cordain, L.; Miller, J.B.; Eaton, S.B.; Mann, N.; Holt, S.H.; and Speth, J.D. Plant–animal subsistence ratios and macronutrient energy estimations in worldwide hunter-gatherer diets. Am. J. Clin. Nutr. 2000, 71, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Cordain, L.; Eaton, S.B.; Miller, J.B.; Mann, N.; and Hill, K. The paradoxical nature of hunter-gatherer diets: Meat-based, yet non-atherogenic. Eur. J. Clin. Nutr. 2002, 56, S42–S52. [Google Scholar] [CrossRef]
- Xu, Z.; McClure, S.T.; and Appel, L.J. Dietary cholesterol intake and sources among U.S adults: Results from national health and nutrition examination surveys (NHANES), 2001⁻2014. Nutrients 2018, 10, 771. [Google Scholar] [CrossRef]
- Gogna, P.; Villeneuve, P.J.; Borghese, M.M.; King, W.D. An exposure-response meta-analysis of ambient PM2. 5 during pregnancy and preeclampsia. Environ Res. 2022, 210, 112934. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.; Stayner, L.; Slama, R.; Sørensen, M.; Figueras, F.; Nieuwenhuijsen, M.J.; Raaschou-Nielsen, O.; Dadvand, P. Ambient air pollution and pregnancy-induced hypertensive disorders: a systematic review and meta-analysis. Hypertension. 2014, 64, 494–500, Epub 2014 Jun 16. [Google Scholar] [CrossRef]
- van den Hooven EH, de Kluizenaar Y, Pierik FH, Hofman A, van Ratingen SW, Zandveld PY, Mackenbach JP, Steegers EA, Miedema HM, Jaddoe VW. Air pollution, blood pressure, and the risk of hypertensive complications during pregnancy: the generation R study. Hypertension. 2011, 57, 406–12. [CrossRef] [PubMed]
- National Toxicology Program. NTP monograph on the systematic review of traffic-related air pollution and hypertensive disorders of pregnancy. NTP Monogr. 2019 Dec;(7):NTP-MGRAPH-7. [CrossRef]
- Pereira, G.; Haggar, F.; Shand, A.W.; Bower, C.; Cook, A.; Nassar, N. Association between pre-eclampsia and locally derived traffic-related air pollution: a retrospective cohort study. J Epidemiol Community Health. 2013, 67, 147–52. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Yamamoto, S. Effect of indoor air pollution from biomass and solid fuel combustion on symptoms of preeclampsia/eclampsia in Indian women. Indoor Air. 2015, 25, 341–52, Epub 2014 Aug 11. [Google Scholar] [CrossRef]
- Robbins, T.; Kuhrt, K.; Vousden, N.; Seed, P.; Shennan, A. Household air pollution and incidence of eclampsia in eight low- and middle-income countries. Int J Gynaecol Obstet. 2023, 160, 449–450. [Google Scholar] [CrossRef] [PubMed]
- Nobles, C.J.; Williams, A.; Ouidir, M.; Sherman, S.; Mendola, P. Differential Effect of Ambient Air Pollution Exposure on Risk of Gestational Hypertension and Preeclampsia. Hypertension. 2019, 74, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Assibey-Mensah, V.; Glantz, J.C.; Hopke, P.K.; Jusko, T.A.; Thevenet-Morrison, K.; Chalupa, D.; Rich, D.Q. Wintertime Wood Smoke, Traffic Particle Pollution, and Preeclampsia. Hypertension. 2020, 75, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Dadvand, P.; Basagaña, X.; Figueras, F.; Amoly, E.; Tobias, A.; de Nazelle, A.; Querol, X.; Sunyer, J.; Nieuwenhuijsen, M.J. Saharan dust episodes and pregnancy. J Environ Monit. 2011, 13, 3222–8. [Google Scholar] [CrossRef]
- Bogan, M.; Al, B.; Kul, S.; Zengin, S.; Oktay, M.; Sabak, M.; Gümüşboğa, H.; Bayram, H. The effects of desert dust storms, air pollution, and temperature on morbidity due to spontaneous abortions and toxemia of pregnancy: 5-year analysis. Int J Biometeorol. 2021, 65, 1733–1739. [Google Scholar] [CrossRef]
- Michikawa, T.; Morokuma, S.; Yamazaki, S.; Takami, A.; Sugata, S.; Yoshino, A.; Takeda, Y.; Nakahara, K.; Saito, S.; Hoshi, J.; Kato, K.; Nitta, H.; Nishiwaki, Y. Exposure to chemical components of fine particulate matter and ozone, and placenta-mediated pregnancy complications in Tokyo: a register-based study. J Expo Sci Environ Epidemiol. 2022, 32, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Wrońska-Nofer, T.; Rosin, J.; Bartosz, G. Interaction of ethanol and xylene in their effects on erythrocytes and other haematological parameters in the rat. J Appl Toxicol. 1991, 11, 289–92. [Google Scholar] [CrossRef] [PubMed]
- Moghe, A.; Ghare, S.; Lamoreau, B.; Mohammad, M.; Barve, S.; McClain, C.; Joshi-Barve, S. Molecular mechanisms of acrolein toxicity: relevance to human disease. Toxicol Sci. 2015, 143, 242–55. [Google Scholar] [CrossRef] [PubMed]
- Shan, Q.; Li, H.; Chen, N.; Qu, F.; Guo, J. Understanding the Multiple Effects of PCBs on Lipid Metabolism. Diabetes Metab Syndr Obes. 2020, 13, 3691–3702. [Google Scholar] [CrossRef] [PubMed]
- Wikström, S.; Lindh, C.H.; Shu, H. Early pregnancy serum levels of perfluoroalkyl substances and risk of preeclampsia in Swedish women. Sci Rep 2019, 9, 9179. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Chen, Q.; Zhang, L.; Luo, K.; Chen, L.; Zhao, S.; Feng, L.; Zhang, J. Prenatal exposure to perfluoroalkyl and polyfluoroalkyl substances and the risk of hypertensive disorders of pregnancy. Environ Health. 2019, 18, 5. [Google Scholar] [CrossRef] [PubMed]
- Bommarito, P.A.; Ferguson, K.K.; Meeker, J.D.; McElrath, T.F.; Cantonwine, D.E. Maternal Levels of Perfluoroalkyl Substances (PFAS) during Early Pregnancy in Relation to Preeclampsia Subtypes and Biomarkers of Preeclampsia Risk. Environ Health Perspect. 2021, 129, 107004. [Google Scholar] [CrossRef]
- Eslami, B.; Malekafzali, H.; Rastkari, N.; Rashidi, B.H.; Djazayeri, A.; Naddafi, K. Association of serum concentrations of persistent organic pollutants (POPs) and risk of pre-eclampsia: a case-control study. J Environ Health Sci Eng. 2016, 14, 17. [Google Scholar] [CrossRef]
- Soleymanlou, N.; Jurisica, I.; Nevo, O.; Ietta, F.; Zhang, X.; Zamudio, S.; Post, M.; Caniggia, I. Molecular evidence of placental hypoxia in preeclampsia. J Clin Endocrinol Metab. 2005, 90, 4299–308. [Google Scholar] [CrossRef]
- Higgins, D.F.; Kimura, K.; Iwano, M.; Haase, V.H. Hypoxia-inducible factor signaling in the development of tissue fibrosis. Cell Cycle. 2008, 7, 1128–32. [Google Scholar] [CrossRef]
- Tirpe, A.A.; Gulei, D.; Ciortea, S.M.; Crivii, C.; Berindan-Neagoe, I. Hypoxia: Overview on Hypoxia-Mediated Mechanisms with a Focus on the Role of HIF Genes. Int J Mol Sci. 2019, 20, 6140. [Google Scholar] [CrossRef] [PubMed]
- Korkes, H.A.; De Oliveira, L.; Sass, N.; Salahuddin, S.; Karumanchi, S.A.; Rajakumar, A. Relationship between hypoxia and downstream pathogenic pathways in preeclampsia. Hypertens Pregnancy. 2017, 36, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Chang, C.; Hao, M.; Chen, M.; Woodley, D.T.; Schönthal, A.H.; Li, W. Heat shock protein-90alpha (Hsp90α) stabilizes hypoxia-inducible factor-1α (HIF-1α) in support of spermatogenesis and tumorigenesis. Cancer Gene Ther. 2021, 28, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.S.; Qiao, X.; Reslan, O.M.; Xia, Y.; Raffetto, J.D.; Paleolog, E.; Davies, A.H.; Khalil, R.A. Prolonged mechanical stretch is associated with upregulation of hypoxia-inducible factors and reduced contraction in rat inferior vena cava. J Vasc Surg. 2011, 53, 764–73. [Google Scholar] [CrossRef] [PubMed]
- Tafani, M.; Sansone, L.; Limana, F.; Arcangeli, T.; De Santis, E.; Polese, M.; Fini, M.; Russo, M.A. The Interplay of Reactive Oxygen Species, Hypoxia, Inflammation, and Sirtuins in Cancer Initiation and Progression. Oxid Med Cell Longev. 2016;2016:3907147. [CrossRef]
- Lum, J.J.; Bui, T.; Gruber, M.; Gordan, J.D.; DeBerardinis, R.J.; Covello, K.L.; Simon, M.C.; Thompson, C.B. The transcription factor HIF-1alpha plays a critical role in the growth factor-dependent regulation of both aerobic and anaerobic glycolysis. Genes Dev. 2007, 21, 1037–49. [Google Scholar] [CrossRef] [PubMed]
- Zera, K.; Zastre, J. Stabilization of the hypoxia-inducible transcription Factor-1 alpha (HIF-1α) in thiamine deficiency is mediated by pyruvate accumulation. Toxicol Appl Pharmacol. 2018, 355, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Watts, E.R.; Walmsley, S.R. Inflammation and Hypoxia: HIF and PHD Isoform Selectivity. Trends Mol Med. 2019, 25, 33–46. [Google Scholar] [CrossRef]
- Déry, M.A.; Michaud, M.D.; Richard, D.E. Hypoxia-inducible factor 1: regulation by hypoxic and non-hypoxic activators. Int J Biochem Cell Biol. 2005, 37, 535–40. [Google Scholar] [CrossRef]
- Schönenberger, M.J.; Kovacs, W.J. Hypoxia signaling pathways: modulators of oxygen-related organelles. Front Cell Dev Biol. 2015, 3, 42. [Google Scholar] [CrossRef]
- Liu, W.; Wang, S.J.; Lin, Q.D. Study on the expressions of PHD and HIF in placentas from normal pregnant women and patients with preeclampsia. Int J Biol Sci. 2014, 10, 278–84. [Google Scholar] [CrossRef]
- Fuenzalida, B.; Yañez, M.J.; Mueller, M.; Mistry, H.D.; Leiva, A.; Albrecht, C. Evidence for hypoxia-induced dysregulated cholesterol homeostasis in preeclampsia: Insights into the mechanisms from human placental cells and tissues. FASEB J. 2024, 38, e23431. [Google Scholar] [CrossRef] [PubMed]
- Coates, H.W.; Capell-Hattam, I.M.; Olzomer, E.M.; Du, X.; Farrell, R.; Yang, H.; Byrne, F.L.; Brown, A.J. Hypoxia truncates and constitutively activates the key cholesterol synthesis enzyme squalene monooxygenase. Elife. 2023, 12, e82843. [Google Scholar] [CrossRef] [PubMed]
- Diet induced experimental toxemia and cation transport. Nutr Rev. 1965, 23, 309–11. [CrossRef] [PubMed]
- Wang, G.L.; Semenza, G.L. General involvement of hypoxia-inducible factor 1 in transcriptional response to hypoxia. Proc Natl Acad Sci U S A. 1993, 90, 4304–8. [Google Scholar] [CrossRef] [PubMed]
- Caniggia, I.; Winter, J.; Lye, S.J.; Post, M. Oxygen and placental development during the first trimester: implications for the pathophysiology of pre-eclampsia. Placenta. 2000 Mar-Apr;21 Suppl A:S25-30. [CrossRef]
- Carter, A.M. Placental Gas Exchange and the Oxygen Supply to the Fetus. Compr Physiol. 2015, 5, 1381–403. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, E.S.; Brownbill, P.; Jones, N.W.; Abrahams, V.M.; Baker, P.N.; Sibley, C.P.; Crocker, I.P. Utero-placental haemodynamics in the pathogenesis of pre-eclampsia. Placenta. 2009, 30, 634–41. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, T.M.; Manwani, R.; Ohadike, C.; Wijesekara, J.; Baker, P.N. The placenta in pre-eclampsia and intrauterine growth restriction: studies on exchange surface areas, diffusion distances and villous membrane diffusive conductances. Placenta. 2007 Feb-Mar;28(2-3):233-8. [CrossRef]
- Subczynski, W.K.; Widomska, J.; Stein, N.; Swartz, H.M. Factors determining barrier properties to oxygen transport across model and cell plasma membranes based on EPR spin-label oximetry. Appl Magn Reson. 2021, 52, 1237–1260. [Google Scholar] [CrossRef] [PubMed]
- Plesnar, E.; Szczelina, R.; Subczynski, W.K.; Pasenkiewicz-Gierula, M. Is the cholesterol bilayer domain a barrier to oxygen transport into the eye lens? Biochim Biophys Acta Biomembr. 2018, 1860, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Albert, A.D.; Boesze-Battaglia, K. The role of cholesterol in rod outer segment membranes. Prog Lipid Res. 2005 Mar-May;44(2-3):99-124. [CrossRef]
- Zuniga-Hertz, J.P.; Patel, H.H. The Evolution of Cholesterol-Rich Membrane in Oxygen Adaption: The Respiratory System as a Model. Front Physiol. 2019, 10, 1340. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Widomska, J.; Stein, N.; Swartz, H.M. Factors determining barrier properties to oxygen transport across model and cell plasma membranes based on EPR spin-label oximetry. Appl Magn Reson. 2021, 52, 1237–1260. [Google Scholar] [CrossRef]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L.; Sullards, C.M.; Wang, E.; Murphy, R.C.; Barkley, R.M.; Leiker, T.J.; Raetz, C.R.; Guan, Z.; Laird, G.M.; Six, D.A.; Russell, D.W.; McDonald, J.G.; Subramaniam, S.; Fahy, E.; Dennis, E.A. Lipidomics reveals a remarkable diversity of lipids in human plasma. J Lipid Res. 2010, 51, 3299–305. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.L.; Mitchell, D.C.; and Litman, B.J. Trans fatty acid derived phospholipids show increased membrane cholesterol and reduced receptor activation as compared to their cis analogs. Biochemistry 2005, 44, 4458–4465. [Google Scholar] [CrossRef] [PubMed]
- Baumer, Y.; McCurdy, S.; Weatherby, T.M.; Mehta, N.N.; Halbherr, S.; Halbherr, P.; Yamazaki, N.; Boisvert, W.A. Hyperlipidemia-induced cholesterol crystal production by endothelial cells promotes atherogenesis. Nat Commun. 2017, 8, 1129. [Google Scholar] [CrossRef] [PubMed]
- Ekroos, K.; Ejsing, C.S.; Bahr, U.; Karas, M.; Simons, K.; Shevchenko, A. Charting molecular composition of phosphatidylcholines by fatty acid scanning and ion trap MS3 fragmentation. J Lipid Res. 2003, 44, 2181–92. [Google Scholar] [CrossRef] [PubMed]
- Sinensky, M. Homeoviscous adaptation--a homeostatic process that regulates the viscosity of membrane lipids in Escherichia coli. Proc Natl Acad Sci U S A. 1974, 71, 522–5. [Google Scholar] [CrossRef]
- Levental KR, Malmberg E, Symons JL, Fan YY, Chapkin RS, Ernst R, Levental I. Lipidomic and biophysical homeostasis of mammalian membranes counteracts dietary lipid perturbations to maintain cellular fitness. Nat Commun. 2020, 11, 1339.
- Irwinda, R.; Hiksas, R.; Siregar, A.A.; et al. Long-chain polyunsaturated fatty acid (LC-PUFA) status in severe preeclampsia and preterm birth: a cross sectional study. Sci Rep. [CrossRef]
- Brown, S.H.; Eather, S.R.; Freeman, D.J.; Meyer, B.J.; and Mitchell, T.W. A lipidomic analysis of placenta in preeclampsia: Evidence for lipid storage. PLoS One 2016, 11, e0163972. [Google Scholar] [CrossRef]
- Anand, S.; Young, S.; Esplin, M.S.; Peaden, B.; Tolley, H.D.; Porter, T.F.; Varner, M.W.; D'Alton, M.E.; Jackson, B.J.; Graves, S.W. Detection and confirmation of serum lipid biomarkers for preeclampsia using direct infusion mass spectrometry. J Lipid Res. 2016, 57, 687–96. [Google Scholar] [CrossRef]
- Clausen, T.; Slott, M.; Solvoll, K.; Drevon, C.A.; Vollset, S.E.; Henriksen, T. High intake of energy, sucrose, and polyunsaturated fatty acids is associated with increased risk of preeclampsia. Am J Obstet Gynecol. 2001, 185, 451–8. [Google Scholar] [CrossRef] [PubMed]
- Schauf, B.; Lang, U.; Stute, P.; Schneider, S.; Dietz, K.; Aydeniz, B.; Wallwiener, D. Reduced red blood cell deformability, an indicator for high fetal or maternal risk, is found in preeclampsia and IUGR. Hypertens Pregnancy. 2002;21, 147-60. [CrossRef]
- Csiszar, B.; Galos, G.; Funke, S.; Kevey, D.K.; Meggyes, M.; Szereday, L.; Kenyeres, P.; Toth, K.; Sandor, B. Peripartum Investigation of Red Blood Cell Properties in Women Diagnosed with Early-Onset Preeclampsia. Cells. 2021, 10, 2714. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, L. Blood rheology and pregnancy. Baillieres Clin Haematol. 1987, 1, 777–99. [Google Scholar] [CrossRef] [PubMed]
- Kazushi Tsuda, Ichiro Nishio, Membrane fluidity and hypertension:, American Journal of Hypertension, Volume 16, Issue 3, March 2003, Pages 259–261,doi.org/10.1016/S0895-7061(02)032570.
- Lühr Jennifer, J.; Amon, L.; Kräter, M.; Kubánková, M.; Sezgin, E.; et al. Maturation of monocyte derived DCs leads to increased cellular stiffness, higher membrane fluidity, and changed lipid composition. Front. Immunol. 2020, 11, 590121. [Google Scholar] [CrossRef] [PubMed]
- Efremov, Y.M.; Velay-Lizancos, M.; Weaver, C.J.; Athamneh, A.I.; Zavattieri, P.D.; Suter, D.M.; et al. Anisotropy vs isotropy in living cell indentation with AFM. Sci. Rep. 2019, 9, 5757. [Google Scholar] [CrossRef] [PubMed]
- Shentu, T.P.; Singh, D.K.; Oh, M.J.; Sun, S.; Sadaat, L.; Makino, A.; et al. The role of oxysterols in control of endothelial stiffness. J. Lipid Res. 2012, 53, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Byfield, F.J.; Aranda-Espinoza, H.; Romanenko, V.G.; Rothblat, G.H.; Levitan, I. Cholesterol depletion increases membrane stiffness of aortic endothelial cells. Biophys J. 2004, 87, 3336–43. [Google Scholar] [CrossRef] [PubMed]
- Schauf, B.; Becker, S.; Abele, H.; Klever, T.; Wallwiener, D.; Aydeniz, B. Effect of magnesium on red blood cell deformability in pregnancy. Hypertens Pregnancy. 2005;24, 17-27. [CrossRef]
- Prescott, A.R.; Comerford, J.G.; Magrath, R.; Lamb, N.J.; Warn, R.M. Effects of elevated intracellular magnesium on cytoskeletal integrity. J Cell Sci. 1988 Mar;89 ( Pt 3):321-9. [CrossRef]
- Simons, K.; and Toomre, D. Lipid rafts and signal transduction. Nat. Rev. Mol. Cell. Biol. 2000, 1, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Lorent, J.H.; Skinkle, A.D.; Levental, K.R.; Waxham, M.N.; Gorfe, A.A.; et al. Domain stability in biomimetic membranes driven by lipid polyunsaturation. J. Phys. Chem. B 2016, 120, 11930–11941. [Google Scholar] [CrossRef] [PubMed]
- Turk, H.F.; Chapkin, R.S. Membrane lipid raft organization is uniquely modified by n-3 polyunsaturated fatty acids. Prostaglandins Leukot Essent Fatty Acids. 2013, 88, 43–7. [Google Scholar] [CrossRef] [PubMed]
- Sviridov, D.; Mukhamedova, N.; Miller, Y.I. Lipid rafts as a therapeutic target. J Lipid Res. 2020, 61, 687–695. [Google Scholar] [CrossRef]
- Riquelme, G.; Vallejos, C.; de Gregorio, N.; Morales, B.; Godoy, V.; Berrios, M.; Bastías, N.; Rodríguez, C. Lipid rafts and cytoskeletal proteins in placental microvilli membranes from preeclamptic and IUGR pregnancies. J Membr Biol. 2011, 241, 127–40. [Google Scholar] [CrossRef]
- Lee, S.M.; Moon, J.Y.; Lim, B.Y.; Kim, S.M.; Park, C.W.; Kim, B.J.; Jun, J.K.; Norwitz, E.R.; Choi, M.H.; Park, J.S. Increased biosynthesis and accumulation of cholesterol in maternal plasma, but not amniotic fluid in pre-eclampsia. Sci Rep. 2019, 9, 1550. [Google Scholar] [CrossRef] [PubMed]
- Olsen, B.N.; Bielska, A.A.; Lee, T.; Daily, M.D.; Covey, D.F.; Schlesinger, P.H.; Baker, N.A.; Ory, D.S. The structural basis of cholesterol accessibility in membranes. Biophys J. 2013, 105, 1838–47. [Google Scholar] [CrossRef] [PubMed]
- Woollett, L.A. Review: Transport of maternal cholesterol to the fetal circulation. Placenta. 2011 Mar;32 Suppl 2(0 2):S218-21. [CrossRef]
- Horne, H.; Holme, A.M.; Roland, M.C.P.; Holm, M.B.; Haugen, G.; Henriksen, T.; Michelsen, T.M. Maternal-fetal cholesterol transfer in human term pregnancies. Placenta. 2019, 87, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhu, L.; Wang, M.; Sun, Q. Associations between Per- and Polyfluoroalkyl Substances Exposures and Blood Lipid Levels among Adults-A Meta-Analysis. Environ Health Perspect. 2023, 131, 56001. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.H.; Soh, S.X.H.; Wang, M.X.; Ong, J.; Seah, A.; Wong, Y.; Fang, Z.; Sim, S.; Lim, J.T. Perfluoroalkyl substances and lipid concentrations in the blood: A systematic review of epidemiological studies. Sci Total Environ. 2022, 850, 158036. [Google Scholar] [CrossRef] [PubMed]
- Tazuma S, Ochi H, Teramen K, Yamashita Y, Horikawa K, Miura H, Hirano N, Sasaki M, Aihara N, Hatsushika S, et al. Degree of fatty acyl chain unsaturation in biliary lecithin dictates cholesterol nucleation and crystal growth. Biochim Biophys Acta. 1994 Nov 17;1215(1-2):74-8. [CrossRef]
- Ioannou, G.N.; Subramanian, S.; Chait, A.; Haigh, W.G.; Yeh, M.M.; Farrell, G.C.; Lee, S.P.; Savard, C. Cholesterol crystallization within hepatocyte lipid droplets and its role in murine NASH. J Lipid Res. 2017, 58, 1067–1079. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Moon, J.Y.; Lim, B.Y.; Kim, S.M.; Park, C.W.; Kim, B.J.; Jun, J.K.; Norwitz, E.R.; Choi, M.H.; Park, J.S. Increased biosynthesis and accumulation of cholesterol in maternal plasma, but not amniotic fluid in pre-eclampsia. Sci Rep. 2019, 9, 1550. [Google Scholar] [CrossRef] [PubMed]
- Horne, H.; Holme, A.M.; Roland, M.C.P.; Holm, M.B.; Haugen, G.; Henriksen, T.; Michelsen, T.M. Maternal-fetal cholesterol transfer in human term pregnancies. Placenta. 2019, 87, 23–29. [Google Scholar] [CrossRef]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ Res. 2019, 124, 1094–1112, Erratum in: Circ Res. 2020, 126, e8. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, S.; Song, T.; Yin, Y.; Tan, C. Placental Angiogenesis in Mammals: A Review of the Regulatory Effects of Signaling Pathways and Functional Nutrients. Adv Nutr. 2021, 12, 2415–2434. [Google Scholar] [CrossRef]
- Wang, C.Y.; Tsai, P.Y.; Chen, T.Y.; Tsai, H.L.; Kuo, P.L.; and Su, M.T. Elevated miR-200a and miR-141 inhibit endocrine gland-derived vascular endothelial growth factor expression and ciliogenesis in preeclampsia. J. Physiol. 2019, 597, 3069–3083. [Google Scholar] [CrossRef] [PubMed]
- Lawson, N.D.; Vogel, A.M.; and Weinstein, B.M. Sonic hedgehog and vascular endothelial growth factor act upstream of the Notch pathway during arterial endothelial differentiation. Dev. Cell. 2002, 3, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Takai, H.; Kondoh, E.; Mogami, H.; Kawasaki, K.; Chigusa, Y.; Sato, M.; Kawamura, Y.; Murakami, R.; Matsumura, N.; Konishi, I.; Mandai, M. Placental Sonic Hedgehog Pathway Regulates Fetal Growth via the IGF Axis in Preeclampsia. J Clin Endocrinol Metab. 2019, 104, 4239–4252. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, A.; Rohatgi, R.; and Siebold, C. Cholesterol access in cellular membranes controls Hedgehog signaling. Nat. Chem. Biol. 2020, 16, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Kinnebrew, M.; Iverson, E.J.; Patel, B.B.; Pusapati, G.V.; Kong, J.H.; Johnson, K.A.; Luchetti, G.; Eckert, K.M.; McDonald, J.G.; Covey, D.F.; Siebold, C.; Radhakrishnan, A.; Rohatgi, R. Cholesterol accessibility at the ciliary membrane controls hedgehog signaling. Elife. 2019, 8, e50051. [Google Scholar] [CrossRef] [PubMed]
- Hart, N.R. A theoretical model of dietary lipid variance as the origin of primary ciliary dysfunction in preeclampsia. Front Mol Biosci. 2023, 10, 1173030. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, S.; Song, T.; Yin, Y.; Tan, C. Placental Angiogenesis in Mammals: A Review of the Regulatory Effects of Signaling Pathways and Functional Nutrients. Adv Nutr. 2021, 12, 2415–2434. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, Z.; Wang, X.; Zhang, L.; Shi, Y.; Wang, J.; Yan, H. Wnt/β-catenin signaling pathway in trophoblasts and abnormal activation in preeclampsia (Review). Mol Med Rep. 2017, 16, 1007–1013, Epub 2018 Mar 30. Erratum in: J Mol Histol. 2022, 53, 145-147. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Z.; Zeng, X.; Wang, J.; Zhang, L.; Song, W.; et al. Wnt/β-catenin signaling pathway in severe preeclampsia. J. Mol. Histol. 2022, 49, 317–327. [Google Scholar] [CrossRef] [PubMed]
- DiFederico, E.; Genbacev, O.; Fisher, S.J. Preeclampsia is associated with widespread apoptosis of placental cytotrophoblasts within the uterine wall. Am J Pathol. 1999, 155, 293–301. [Google Scholar] [CrossRef]
- Sheng, R.; Kim, H.; Lee, H.; Xin, Y.; Chen, Y.; Tian, W.; et al. Cholesterol selectively activates canonical Wnt signalling over non-canonical Wnt signalling. Nat. Commun. 2014, 5, 4393. [Google Scholar] [CrossRef] [PubMed]
- Osol, G.; Ko, N.L.; Mandalà, M. Altered Endothelial Nitric Oxide Signaling as a Paradigm for Maternal Vascular Maladaptation in Preeclampsia. Curr Hypertens Rep. 2017, 19, 82. [Google Scholar] [CrossRef] [PubMed]
- Guerby, P.; Tasta, O.; Swiader, A.; Pont, F.; Bujold, E.; Parant, O.; Vayssiere, C.; Salvayre, R.; Negre-Salvayre, A. Role of oxidative stress in the dysfunction of the placental endothelial nitric oxide synthase in preeclampsia. Redox Biol. 1018. [Google Scholar] [CrossRef]
- Goligorsky, M.S.; Li, H.; Brodsky, S.; Chen, J. Relationships between caveolae and eNOS: everything in proximity and the proximity of everything. Am J Physiol Renal Physiol. 2002, 283, F1–F10. [Google Scholar] [CrossRef]
- Rodal, S.K.; Skretting, G.; Garred, O.; Vilhardt, F.; van Deurs, B.; Sandvig, K. Extraction of cholesterol with methyl-beta-cyclodextrin perturbs formation of clathrin-coated endocytic vesicles. Mol Biol Cell. 1999, 10, 961–74. [Google Scholar] [CrossRef] [PubMed]
- Plenz, G.A.; Hofnagel, O.; Robenek, H. Differential modulation of caveolin-1 expression in cells of the vasculature by statins. Circulation. 2004, 109, e7–e8. [Google Scholar] [CrossRef]
- Salvary, T.; Gambert-Nicot, S.; Brindisi, M.C.; Meneveau, N.; Schiele, F.; Séronde, M.F.; Lorgis, L.; Zeller, M.; Cottin, Y.; Kantelip, J.P.; Gambert, P.; Davani, S. Pravastatin reverses the membrane cholesterol reorganization induced by myocardial infarction within lipid rafts in CD14(+)/CD16(-) circulating monocytes. Biochim Biophys Acta. 2012. Sep;1821, 1287-94. [CrossRef]
- Blair, A.; Shaul, P.W.; Yuhanna, I.S.; Conrad, P.A.; Smart, E.J. Oxidized low density lipoprotein displaces endothelial nitric-oxide synthase (eNOS) from plasmalemmal caveolae and impairs eNOS activation. J Biol Chem. 1999, 274, 32512–9. [Google Scholar] [CrossRef]
- Levitan, I.; Shentu, T.P. Impact of oxLDL on Cholesterol-Rich Membrane Rafts. J Lipids. 2011;2011:730209. [CrossRef]
- Gliozzi, M.; Scicchitano, M.; Bosco, F.; Musolino, V.; Carresi, C.; Scarano, F.; Maiuolo, J.; Nucera, S.; Maretta, A.; Paone, S.; Mollace, R.; Ruga, S.; Zito, M.C.; Macrì, R.; Oppedisano, F.; Palma, E.; Salvemini, D.; Muscoli, C.; Mollace, V. Modulation of Nitric Oxide Synthases by Oxidized LDLs: Role in Vascular Inflammation and Atherosclerosis Development. Int J Mol Sci. 2019, 20, 3294. [Google Scholar] [CrossRef]
- Palei, A.C.; Granger, J.P.; Spradley, F.T. Placental Ischemia Says "NO" to Proper NOS-Mediated Control of Vascular Tone and Blood Pressure in Preeclampsia. Int J Mol Sci. 2021, 22, 11261. [Google Scholar] [CrossRef]
- Sizar, O.; Khare, S.; Jamil, R.T.; Talati, R. Statin Medications. 2023 Feb 5. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 Jan–. [PubMed]
- Goldberg, I.J.; Holleran, S.; Ramakrishnan, R.; Adams, M.; Palmer, R.H.; Dell, R.B.; Goodman, D.S. Lack of effect of lovastatin therapy on the parameters of whole-body cholesterol metabolism. J Clin Invest. 1990, 86, 801–8. [Google Scholar] [CrossRef]
- Schonewille, M.; de Boer, J.F.; Mele, L.; Wolters, H.; Bloks, V.W.; Wolters, J.C.; Kuivenhoven, J.A.; Tietge, U.J.; Brufau, G.; Groen, A.K. Statins increase hepatic cholesterol synthesis and stimulate fecal cholesterol elimination in mice. J Lipid Res. 2016, 57, 1455–64. [Google Scholar] [CrossRef]
- Afonso, Milessa Silva, et al. “Molecular Pathways Underlying Cholesterol Homeostasis.” Nutrients 2018 Jun;10, 760. [CrossRef]
- Zhou, X.; Mott, M.M.; Yiannakou, I.; Bradlee, M.L.; Singer, M.R.; Moore, L.L. Eggs and a Fiber-Rich Diet Are Beneficially Associated with Lipid Levels in Framingham Offspring Study Adults. Curr Dev Nutr. 2024, 8, 102062. [Google Scholar] [CrossRef] [PubMed]
- Dias, Irundika H K et al. “Simvastatin reduces circulating oxysterol levels in men with hypercholesterolaemia.” Redox biology vol. 16 (2018): 139-145. [CrossRef]
- Wang, C.; Enssle, J.; Pietzner, A.; Schmöcker, C.; Weiland, L.; Ritter, O.; Jaensch, M.; Elbelt, U.; Pagonas, N.; Weylandt, K.H. Essential Polyunsaturated Fatty Acids in Blood from Patients with and without Catheter-Proven Coronary Artery Disease. Int J Mol Sci. 2022, 23, 766. [Google Scholar] [CrossRef]
- Stark, J.M. Pre-eclampsia and cytokine induced oxidative stress. Br J Obstet Gynaecol. 1993, 100, 105–9. [Google Scholar] [CrossRef] [PubMed]
- Goulopoulou, S.; Davidge, S.T. Molecular mechanisms of maternal vascular dysfunction in preeclampsia. Trends Mol Med. 2015, 21, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Grzeszczak, K.; Łanocha-Arendarczyk, N.; Malinowski, W.; Ziętek, P.; Kosik-Bogacka, D. Oxidative Stress in Pregnancy. Biomolecules. 2023, 13, 1768. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; Prakash Mishra, A.; Nigam, M.; El Rayess, Y.; Beyrouthy, M.E.; Polito, L.; Iriti, M.; Martins, N.; Martorell, M.; Docea, A.O.; Setzer, W.N.; Calina, D.; Cho, W.C.; Sharifi-Rad, J. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- McCord, J.M. The evolution of free radicals and oxidative stress. Am J Med. 2000, 108, 652–9. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, M.; Mileykovskaya, E.; Dowhan, W. Lipids in the assembly of membrane proteins and organization of protein supercomplexes: implications for lipid-linked disorders. Subcell Biochem. 2008;49:197-239. [CrossRef]
- Koshy, C.; Ziegler, C. Structural insights into functional lipid-protein interactions in secondary transporters. Biochim Biophys Acta. 2015, 1850, 476–87. [Google Scholar] [CrossRef]
- Mahler, C.A.; Snoke, D.B.; Cole, R.M.; Angelotti, A.; Sparagna, G.C.; Baskin, K.K.; Ni, A.; Belury, M.A. Consuming a Linoleate-Rich Diet Increases Concentrations of Tetralinoleoyl Cardiolipin in Mouse Liver and Alters Hepatic Mitochondrial Respiration. J Nutr. 2023 Dec 30:S0022-3166(23)72826-5. [CrossRef]
- Funai, K.; Summers, S.A.; Rutter, J. Reign in the membrane: How common lipids govern mitochondrial function. Curr Opin Cell Biol. 2020, 63, 162–173, Epub 2020 Feb 24. [Google Scholar] [CrossRef]
- Paradies, G.; Ruggiero, F.M.; Petrosillo, G.; Quagliariello, E. Peroxidative damage to cardiac mitochondria: cytochrome oxidase and cardiolipin alterations. FEBS Lett. 1998, 424, 155–8. [Google Scholar] [CrossRef]
- Bellanti, F.; Villani, R.; Tamborra, R.; Blonda, M.; Iannelli, G.; di Bello, G.; Facciorusso, A.; Poli, G.; Iuliano, L.; Avolio, C.; Vendemiale, G.; Serviddio, G. Synergistic interaction of fatty acids and oxysterols impairs mitochondrial function and limits liver adaptation during nafld progression. Redox Biol. 2018, 15, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, R.; Portero-Otin, M.; Sanz, A.; Requena, J.; Barja, G. Modification of the longevity-related degree of fatty acid unsaturation modulates oxidative damage to proteins and mitochondrial DNA in liver and brain. Exp Gerontol. 2004, 39, 725–33. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive oxygen species in living systems: Source, biochemistry, and role in human disease. Am. J. Med. 1991, 91, 14S–22S. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Noguchi, N.; Niki, E. Cholesterol is more readily oxidized than phospholipid linoleates in cell membranes to produce cholesterol hydroperoxides. Free Radic Biol Med. 2024, 211, 89–95. [Google Scholar] [CrossRef]
- Girao, H.; Mota, C.; and Pereira, P. Cholesterol may act as an antioxidant in lens membranes. Curr. Eye Res. 1999, 18, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.L. Another cholesterol hypothesis: cholesterol as antioxidant. Free Radic Biol Med. 1991;11, 47-61. [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract Res Clin Obstet Gynaecol. 2011, 25, 287–99. [Google Scholar] [CrossRef] [PubMed]
- Schieber M, Chandel NS. ROS function in redox signaling and oxidative stress. Curr Biol. 2014, 24, R453–R462. [CrossRef] [PubMed]
- Schäfer, G.; Cramer, T.; Suske, G.; Kemmner, W.; Wiedenmann, B.; Höcker, M. Oxidative stress regulates vascular endothelial growth factor-A gene transcription through Sp1- and Sp3-dependent activation of two proximal GC-rich promoter elements. J Biol Chem. 2003, 278, 8190–8. [Google Scholar] [CrossRef] [PubMed]
- Rumbold, A.; Duley, L.; Crowther, C.A.; Haslam, R.R. Antioxidants for preventing pre-eclampsia. Cochrane Database Syst Rev. 2008, 2008, CD004227. [Google Scholar] [CrossRef]
- Salles, A.M.; Galvao, T.F.; Silva, M.T.; Motta, L.C.; Pereira, M.G. Antioxidants for preventing preeclampsia: a systematic review. ScientificWorldJournal. 2012;2012:243476. Epub 2012 Apr 19. [CrossRef] [PubMed]
- Tenório, M.B.; Ferreira, R.C.; Moura, F.A.; Bueno, N.B.; Goulart, M.O.F.; Oliveira, A.C.M. Oral antioxidant therapy for prevention and treatment of preeclampsia: Meta-analysis of randomized controlled trials. Nutr Metab Cardiovasc Dis. 2018, 28, 865–876. [Google Scholar] [CrossRef]
- Di Fabrizio, C.; Giorgione, V.; Khalil, A.; Murdoch, C.E. Antioxidants in Pregnancy: Do We Really Need More Trials? Antioxidants (Basel). 2022, 11, 812. [Google Scholar] [CrossRef] [PubMed]
- Korkes HA, et al “Lipidomic assessment of plasma and placenta of women with early-onset preeclampsia. ” PLoS One. 2014, 9, e110747.
- Cohen, J.M.; Beddaoui, M.; Kramer, M.S.; Platt, R.W.; Basso, O.; Kahn, S.R. Maternal Antioxidant Levels in Pregnancy and Risk of Preeclampsia and Small for Gestational Age Birth: A Systematic Review and Meta=Analysis. PLoS One. 2015 Aug 6;(8):eO!#%!(@.
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ Res. 2019, 124, 1094–1112, Erratum in: Circ Res. 2020, 126, e8. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.A.; Zingheim, R.W.; King, I.B.; Zebelman, A.M. Omega-3 fatty acids in maternal erythrocytes and risk of preeclampsia. Epidemiology. 1995, 6, 232–7. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Sanchez, S.E.; Larrabure, G.; David, R.; Bralley, J.A.; Williams, M.A. Erythrocyte omega-3 and omega-6 polyunsaturated fatty acids and preeclampsia risk in Peruvian women. Arch Gynecol Obstet. 2006, 274, 97–103, Epub 2006 Mar 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zu, L.; Cai, W.; Cheng, Q.; Hua, T.; Peng, L.; Li, G.; Zhang, X. Metabolomics revealed decreased level of omega-3 PUFA-derived protective eicosanoids in pregnant women with pre-eclampsia. Clin Exp Pharmacol Physiol. 2019, 46, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.A.; King, I.B.; Sorensen, T.K.; Zingheim, R.W.; Troyer, B.L.; Zebelman, A.M.; Luthy, D.A. Risk of preeclampsia in relation to elaidic acid (trans fatty acid) in maternal erythrocytes. Gynecol Obstet Invest. 1998, 46, 84–7. [Google Scholar] [CrossRef] [PubMed]
- Mahomed, K.; Williams, M.A.; King, I.B.; Mudzamiri, S.; Woelk, G.B. Erythrocyte omega-3, omega-6 and trans fatty acids in relation to risk of preeclampsia among women delivering at Harare Maternity Hospital, Zimbabwe. Physiol Res. 2007;56, 37-50. [CrossRef]
- Clausen, T.; Slott, M.; Solvoll, K.; Drevon, C.A.; Vollset, S.E.; Henriksen, T. High intake of energy, sucrose, and polyunsaturated fatty acids is associated with increased risk of preeclampsia. Am J Obstet Gynecol. 2001, 185, 451–8. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Young, S.; Esplin, M.S.; Peaden, B.; Tolley, H.D.; Porter, T.F.; Varner, M.W.; D'Alton, M.E.; Jackson, B.J.; Graves, S.W. Detection and confirmation of serum lipid biomarkers for preeclampsia using direct infusion mass spectrometry. J Lipid Res. 2016, 57, 687–96. [Google Scholar] [CrossRef]
- Wada, Y.; Yoshida-Yamamoto, S.; Wada, Y.; Nakayama, M.; Mitsuda, N.; and Kitajima, H. Trans fatty acid accumulation in the human placenta. J. Mass Spectrom. 2017, 52, 139–143. [Google Scholar] [CrossRef]
- Brown, S.H.; Eather, S.R.; Freeman, D.J.; Meyer, B.J.; and Mitchell, T.W. A lipidomic analysis of placenta in preeclampsia: Evidence for lipid storage. PLoS One 2016, 11, e0163972. [Google Scholar] [CrossRef] [PubMed]
- Irwinda, R.; Hiksas, R.; Siregar, A.A.; et al. Long-chain polyunsaturated fatty acid (LC-PUFA) status in severe preeclampsia and preterm birth: a cross sectional study. Sci Rep, 2021; 11, 14701. [Google Scholar] [CrossRef]
- Chavarro, J.E.; Halldorsson, T.I.; Leth, T.; Bysted, A.; Olsen, S.F. A prospective study of trans fat intake and risk of preeclampsia in Denmark. Eur J Clin Nutr. 2011, 65, 944–51, Epub 2011 May 11. [Google Scholar] [CrossRef]
- Arvizu, M.; Minguez-Alarcon, L.; Wang, S.; Mitsunami, M.; Stuart, J.J.; Rich-Edwards, J.W.; Rosner, B.; Chavarro, J.E. Pre-pregnancy fat intake in relation to hypertensive disorders of pregnancy. Am J Clin Nutr. 2022, 116, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Alamolhoda, S.H.; Simbar, M.; Mirmiran, P.; Mirabi, P. Effect of low trans-fatty acid intakes on preeclampsia: A randomized controlled trial. J Res Med Sci. 2020, 25, 112. [Google Scholar] [CrossRef]
- Baig, S.; Lim, J.Y.; Fernandis, A.Z.; Wenk, M.R.; Kale, A.; Su, L.L.; Biswas, A.; Vasoo, S.; Shui, G.; Choolani, M. Lipidomic analysis of human placental syncytiotrophoblast microvesicles in adverse pregnancy outcomes. Placenta. 2013, 34, 436–42. [Google Scholar] [CrossRef] [PubMed]
- Bartho, L.A.; Keenan, E.; Walker, S.P.; MacDonald, T.M.; Nijagal, B.; Tong, S.; Kaitu'u-Lino, T.J. Plasma lipids are dysregulated preceding diagnosis of preeclampsia or delivery of a growth restricted infant. EBioMedicine. 2023, 94, 104704, Epub 2023 Jul 6. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- He, B.; Liu, Y.; Maurya, M.R.; Benny, P.; Lassiter, C.; Li, H.; Subramaniam, S.; Garmire, L.X. The maternal blood lipidome is indicative of the pathogenesis of severe preeclampsia. J Lipid Res. 2021;62:100118. Epub 2021 Sep 20. [CrossRef]
- Brown, S.H.; Eather, S.R.; Freeman, D.J.; Meyer, B.J.; Mitchell, T.W. A Lipidomic Analysis of Placenta in Preeclampsia: Evidence for Lipid Storage. PLoS One. 2016, 11, e0163972. [Google Scholar] [CrossRef] [PubMed]
- Lantzanaki, M.; Vavilis, T.; Harizopoulou, V.C.; Bili, H.; Goulis, D.G.; Vavilis, D. Ceramides during Pregnancy and Obstetrical Adverse Outcomes. Metabolites. 2023, 13, 1136. [Google Scholar] [CrossRef] [PubMed]
- Melland-Smith, M.; Ermini, L.; Chauvin, S.; Craig-Barnes, H.; Tagliaferro, A.; Todros, T.; Post, M.; Caniggia, I. Disruption of sphingolipid metabolism augments ceramide-induced autophagy in preeclampsia. Autophagy. 2015, 11, 653–69. [Google Scholar] [CrossRef]
- Dobierzewska, A.; Soman, S.; Illanes, S.E.; Morris, A.J. Plasma cross-gestational sphingolipidomic analyses reveal potential first trimester biomarkers of preeclampsia. PLoS One. 2017, 12, e0175118. [Google Scholar] [CrossRef]
- Patanapirunhakit, P.; Karlsson, H.; Mulder, M.; Ljunggren, S.; Graham, D.; Freeman, D. Sphingolipids in HDL - Potential markers for adaptation to pregnancy? Biochim Biophys Acta Mol Cell Biol Lipids. 2021, 1866, 158955. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Hao, S.; You, J.; Yao, X.; Li, Z.; Schilling, J.; Thyparambil, S.; Liao, W.L.; Zhou, X.; Mo, L.; Ladella, S.; Davies-Balch, S.R.; Zhao, H.; Fan, D.; Whitin, J.C.; Cohen, H.J.; McElhinney, D.B.; Wong, R.J.; Shaw, G.M.; Stevenson, D.K.; Sylvester, K.G.; Ling, X.B. Early-pregnancy prediction of risk for pre-eclampsia using maternal blood leptin/ceramide ratio: discovery and confirmation. BMJ Open. 2021, 11, e050963. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kang, Y.; Lee, E.M.; Jung, Y.M.; Hong, S.; Park, S.J.; Park, C.W.; Norwitz, E.R.; Lee, D.Y.; Park, J.S. Metabolomic biomarkers in midtrimester maternal plasma can accurately predict the development of preeclampsia. Sci Rep. 2020, 10, 16142. [Google Scholar] [CrossRef] [PubMed]
- Kummerow, F.A. Modification of cell membrane composition by dietary lipids and its implications for atherosclerosis. Ann N Y Acad Sci. 1983;414:29-43. [CrossRef] [PubMed]
- Hong, L.; Zhu, L.; Zhang, J.; Fu, Y.; Qi, X.; Zhao, M. Association of dietary inflammatory index with risk of gestational diabetes mellitus and preeclampsia: a systematic review and meta-analysis. Br J Nutr. 2024, 131, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T.; Slott, M.; Solvoll, K.; Drevon, C.A.; Vollset, S.E.; Henriksen, T. High intake of energy, sucrose, and polyunsaturated fatty acids is associated with increased risk of preeclampsia. Am J Obstet Gynecol. 2001, 185, 451–8. [Google Scholar] [CrossRef] [PubMed]
- Mu, T.S.; Duran-Stanton, A.; Stone, E.A.; Zarzabal, L.A.; Loewendorf, A. Rates of Preeclampsia and Post-preeclamptic Cardiovascular Disease Among US Military Servicewomen: A Retrospective Case-cohort Study. Mil Med. 2023 Aug 4:usad300. [CrossRef]
- Hedegaard, S.; Nohr, E.A.; Olsen, S.F.; Halldorsson, T.I.; Renault, K.M. Adherence to different forms of plant-based diets and pregnancy outcomes in the Danish National Birth Cohort: A prospective observational study. Acta Obstet Gynecol Scand. 2024 Jan 24. Epub ahead of print. PMID: 38263894. [CrossRef]
- Yang, J.; Song, Y.; Gaskins, A.J.; Li, L.J.; Huang, Z.; Eriksson, J.G.; Hu, F.B.; Chong, Y.S.; Zhang, C. Mediterranean diet and female reproductive health over lifespan: a systematic review and meta-analysis. Am J Obstet Gynecol. 2023, 229, 617–631, Epub 2023 Jul 26. [Google Scholar] [CrossRef] [PubMed]
- Scholl, T.O.; Leskiw, M.; Chen, X.; Sims, M.; Stein, T.P. Oxidative stress, diet, and the etiology of preeclampsia. Am J Clin Nutr. 2005, 81, 1390–6. [Google Scholar] [CrossRef] [PubMed]
- Irwinda R, Hiksas R, Siregar AA, Saroyo YB, Wibowo N. polyunsaturated fatty acid (LC-PUFA) status in severe preeclampsia and preterm birth: a cross sectional study. Sci Rep. 20Long-chain21 Jul 19;11, 14701. [CrossRef]
- Godhamgaonkar, A.A.; Wadhwani, N.S.; Randhir, K.N.; Selukar, S.S.; Dalvi, S.; Dangat, K.; Wagh, G.N.; Lalwani, S.; Chandhiok, N.; Kulkarni, B.; Fall, C.; Sachdev, H.P.S.; Gupte, S.; Joshi, S.R. Erythrocyte fatty acids and desaturase indices in early pregnancy are associated with risk of preeclampsia. Prostaglandins Leukot Essent Fatty Acids. 2023, 196, 102583. [Google Scholar] [CrossRef] [PubMed]
- Alamolhoda, S.H.; Simbar, M.; Mirmiran, P.; Mirabi, P. Effect of low trans-fatty acid intakes on preeclampsia: A randomized controlled trial. J Res Med Sci. 2020, 25, 112. [Google Scholar] [CrossRef]
- Nordqvist, M.; Jacobsson, B.; Brantsæter, A.L.; Myhre, R.; Nilsson, S.; Sengpiel, V. Timing of probiotic milk consumption during pregnancy and effects on the incidence of preeclampsia and preterm delivery: a prospective observational cohort study in Norway. BMJ Open. 2018, 8, e018021. [Google Scholar] [CrossRef]
- 6 Cortez-Ribeiro AC, Meireles M, Ferro-Lebres V, Almeida-de-Souza J. Olive oil consumption confers protective effects on maternal-fetal outcomes: A systematic review of the evidence. Nutr Res. 2023, 110, 87–95. [CrossRef] [PubMed]
- Frederick, I.O.; Williams, M.A.; Dashow, E.; Kestin, M.; Zhang, C.; Leisenring, W.M. Dietary fiber, potassium, magnesium and calcium in relation to the risk of preeclampsia. J Reprod Med. 2005, 50, 332–44. [Google Scholar] [PubMed]
- Luqman, A.; Hassan, A.; Ullah, M.; Naseem, S.; Ullah, M.; Zhang, L.; Din, A.U.; Ullah, K.; Ahmad, W.; Wang, G. Role of the intestinal microbiome and its therapeutic intervention in cardiovascular disorder. Front Immunol. 2024, 15, 1321395. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Wang, X.; Wang, J. Research progress on the correlation between gut microbiota and preeclampsia: microbiome changes, mechanisms and treatments. Front Cell Infect Microbiol. 2023, 13, 1256940. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Chen, Y.; Zhou, Q.; Wang, C.; Chen, L.; Di, W.; Zhang, Y. Short-chain fatty acids accompanying changes in the gut microbiome contribute to the development of hypertension in patients with preeclampsia. Clin Sci (Lond). 2020, 134, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Middleton, P.; Gomersall, J.C.; Gould, J.F.; Shepherd, E.; Olsen, S.F.; Makrides, M. Omega-3 fatty acid addition during pregnancy. Cochrane Database Syst Rev. 2018, 11, CD003402. [Google Scholar] [CrossRef] [PubMed]
- Thangaratinam, S.; Rogozińska, E.; Jolly, K.; Glinkowski, S.; Duda, W.; Borowiack, E.; Roseboom, T.; Tomlinson, J.; Walczak, J.; Kunz, R.; Mol, B.W.; Coomarasamy, A.; Khan, K.S. Interventions to reduce or prevent obesity in pregnant women: a systematic review. Health Technol Assess. 2012, 16, iii–iv, 1. [Google Scholar] [CrossRef] [PubMed]
- Kinshella MW, Omar S, Scherbinsky K, Vidler M, Magee LA, von Dadelszen P, Moore SE, Elango R, The Precise Conceptual Framework Working Group. Effects of Maternal Nutritional Supplements and Dietary Interventions on Placental Complications: An Umbrella Review, Meta-Analysis and Evidence Map. Nutrients. 2021, 13, 472. [CrossRef] [PubMed]
- H Al Wattar B, Dodds J, Placzek A, Beresford L, Spyreli E, Moore A, Gonzalez Carreras FJ, Austin F, Murugesu N, Roseboom TJ, Bes-Rastrollo M, Hitman GA, Hooper R, Khan KS, Thangaratinam S. ; ESTEEM study group. Mediterranean-style diet in pregnant women with metabolic risk factors (ESTEEM): A pragmatic multicentre randomised trial. PLoS Med. 2019, 16, e1002857. [CrossRef]
- Assaf-Balut C, García de la Torre N, Duran A, Fuentes M, Bordiú E, Del Valle L, Familiar C, Valerio J, Jiménez I, Herraiz MA, Izquierdo N, Torrejon MJ, Cuadrado MÁ, Ortega I, Illana FJ, Runkle I, de Miguel P, Moraga I, Montañez C, Barabash A, Cuesta M, Rubio MA, Calle-Pascual AL. A Mediterranean Diet with an Enhanced Consumption of Extra Virgin Olive Oil and Pistachios Improves Pregnancy Outcomes in Women Without Gestational Diabetes Mellitus: A Sub-Analysis of the St. Carlos Gestational Diabetes Mellitus Prevention Study. Ann Nutr Metab. 2019;74, 69-79. [CrossRef]
- Sherriff, J.L.; O'Sullivan, T.A.; Properzi, C.; Oddo, J.L.; Adams, L.A. Choline, Its Potential Role in Nonalcoholic Fatty Liver Disease, and the Case for Human and Bacterial Genes. Adv Nutr. 2016, 7, 5–13. [Google Scholar] [CrossRef]
- Sanders, L.M.; Zeisel, S.H. Choline: Dietary Requirements and Role in Brain Development. Nutr Today. 2007;42, 181-186. [CrossRef]
- Wallace TC, Fulgoni VL 3rd. Assessment of Total Choline Intakes in the United States. J Am Coll Nutr. 2016;35, 108-12. [CrossRef]
- Caudill, M.A. Pre- and postnatal health: evidence of increased choline needs. J Am Diet Assoc. 2010, 110, 1198–206. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.; Chen, L.; Deng, M.G.; Liang, Y.; Liu, F.; Nie, J.Q. Dietary choline intake and non-alcoholic fatty liver disease (NAFLD) in U. S. adults: National Health and Nutrition Examination Survey (NHANES) 2017-2018. Eur J Clin Nutr. 2023, 77, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Sherriff, J.L.; O'Sullivan, T.A.; Properzi, C.; Oddo, J.L.; Adams, L.A. Choline, Its Potential Role in Nonalcoholic Fatty Liver Disease, and the Case for Human and Bacterial Genes. Adv Nutr. 2016, 7, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.S.; Rossi, M.A.; Oliveira, J.S. Pathogenesis of the renal injury in choline deficiency: the role of catecholamines and acetylcholine. Br J Exp Pathol. 1979, 60, 613–9. [Google Scholar] [PubMed]
- Golzarand, M.; Bahadoran, Z.; Mirmiran, P.; Azizi, F. Dietary choline and betaine intake and risk of hypertension development: a 7.4-year follow-up. Food Funct. 2021, 12, 4072–4078. [Google Scholar] [CrossRef]
- Zhang, J.; Zang, X.; Lv, J.; Zhang, Y.; Lv, Z.; Yu, M. Changes in Lipidomics, Metabolomics, and the Gut Microbiota in CDAA-Induced NAFLD Mice after Polyene Phosphatidylcholine Treatment. Int J Mol Sci. 2023, 24, 1502. [Google Scholar] [CrossRef] [PubMed]
- Kinshella MW, Omar S, Scherbinsky K, Vidler M, Magee LA, von Dadelszen P, Moore SE, Elango R, The Precise Conceptual Framework Working Group. Effects of Maternal Nutritional Supplements and Dietary Interventions on Placental Complications: An Umbrella Review, Meta-Analysis and Evidence Map. Nutrients. 2021, 13, 472. [CrossRef] [PubMed]
- Mohammad, N.S.; Nazli, R.; Zafar, H.; Fatima, S. Effects of lipid based Multiple Micronutrients Supplement on the birth outcome of underweight pre-eclamptic women: A randomized clinical trial. Pak J Med Sci. 2022, 38, 219–226. [Google Scholar] [CrossRef]
- Fernandez, M.L.; Murillo, A.G. Is There a Correlation between Dietary and Blood Cholesterol? Evidence from Epidemiological Data and Clinical Interventions. Nutrients. 2022, 14, 2168. [Google Scholar] [CrossRef]
- Li, S.N.; Liu, Y.H.; Luo, Z.Y.; Cui, Y.F.; Cao, Y.; Fu, W.J.; Dou, W.F.; Duan, D.D.; Zhao, X.L.; Chen, Y.M.; Lyu, Q.J.; Chen, Q.S.; Zeng, F.F. The association between dietary fatty acid intake and the risk of developing preeclampsia: a matched case-control study. Sci Rep. 2021, 11, 4048. [Google Scholar] [CrossRef]
- Soliman, G.A. Dietary Cholesterol and the Lack of Evidence in Cardiovascular Disease. Nutrients. 2018, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Addis, P.B. Occurrence of lipid oxidation products in foods. Food Chem Toxicol. 1986 Oct-Nov;24(10-11):1021-30. [CrossRef]
- Herlambang, H.; Puspasari, A.; Maharani, C.; Enis, R.N.; Tarawifa, S.; Fitri, A.D.; Harahap, H.; Harahap, A.H.; Kusdiyah, E.; Syamsunarno, M.R.A.A. Comprehensive fatty acid fractionation profilling in preeclampsia: a case control study with multivariable analysis. BMC Pregnancy Childbirth. 2022, 22, 8. [Google Scholar] [CrossRef]
- Tan, B.; Ma, Y.; Zhang, L.; Li, N.; Zhang, J. The application of metabolomics analysis in the research of gestational diabetes mellitus and preeclampsia. J Obstet Gynaecol Res. 2020, 46, 1310–1318. [Google Scholar] [CrossRef] [PubMed]
- Sviridov, D.; Mukhamedova, N.; Miller, Y.I. Lipid rafts as a therapeutic target. J Lipid Res. 2020, 61, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Esquivel, M.K. Nutritional Status and Nutrients Related to Pre-Eclampsia Risk. Am J Lifestyle Med. 2022, 17, 41–45. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Representative Membrane Lipids. Note the very low melting points of the highly unsaturated omega-6 and omega-3 essential fatty acids as well as the very short chain saturated butyric acid (top of chart in red, orange, and yellow). Monounsaturated fatty acids such as oleic acid which is abundant in olive oil or medium chain saturated fatty acids such as lauric acid found in coconut oil have moderate melting points (green). Long chain saturated fatty acids (LCSFA) with no double bonds such as palmitic and stearic acids found in beef have high melting points and high cholesterol affinity (purple, blue and greys). Although cholesterol (white) has a very high melting point, due to its planar head and short tail, in a cell membrane it acts as a plasticizer maintaining normal viscosity over a wide range of membrane compositions. Cholesterol also reduces the membrane permeability to water. Although oleic acid and elaidic acid have the same chemical formula of eighteen carbons and one double bond at position nine, with a different bond structure, they have melting points over 30℃ apart. Elaidic acid also has a much higher cholesterol affinity. (Figure used with permission from Hart NR. A theoretical model of dietary lipid variance as the origin of primary ciliary dysfunction in preeclampsia. Front Mol Biosci. 2023 May 5;10:1173030. doi:10.3389/fmolb.2023.1173030.).
Figure 1.
Representative Membrane Lipids. Note the very low melting points of the highly unsaturated omega-6 and omega-3 essential fatty acids as well as the very short chain saturated butyric acid (top of chart in red, orange, and yellow). Monounsaturated fatty acids such as oleic acid which is abundant in olive oil or medium chain saturated fatty acids such as lauric acid found in coconut oil have moderate melting points (green). Long chain saturated fatty acids (LCSFA) with no double bonds such as palmitic and stearic acids found in beef have high melting points and high cholesterol affinity (purple, blue and greys). Although cholesterol (white) has a very high melting point, due to its planar head and short tail, in a cell membrane it acts as a plasticizer maintaining normal viscosity over a wide range of membrane compositions. Cholesterol also reduces the membrane permeability to water. Although oleic acid and elaidic acid have the same chemical formula of eighteen carbons and one double bond at position nine, with a different bond structure, they have melting points over 30℃ apart. Elaidic acid also has a much higher cholesterol affinity. (Figure used with permission from Hart NR. A theoretical model of dietary lipid variance as the origin of primary ciliary dysfunction in preeclampsia. Front Mol Biosci. 2023 May 5;10:1173030. doi:10.3389/fmolb.2023.1173030.).
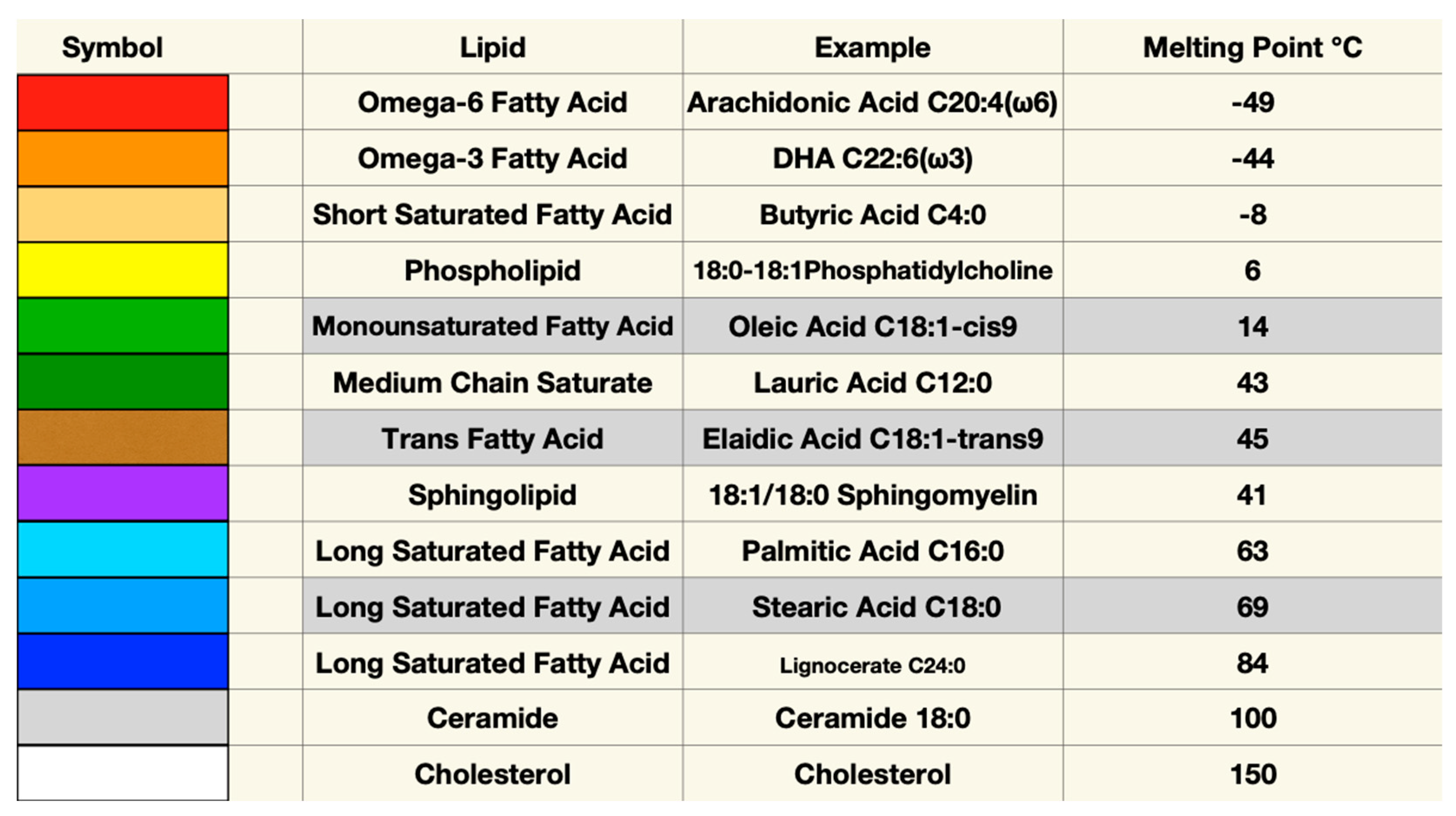
Figure 2.
Membrane Lipid Head Groups with Attached Fatty Acid Tails. Phosphatidylcholine is pictured as a representative glycerophospholipid (top right). Each phosphatidylcholine headgroup has two fatty acids attached with the most usual arrangement being a saturated fatty acid at position one and an unsaturated fatty acid at position two[86]. Sphingomyelin has a larger headgroup and generally has more LCSFA. Length and unsaturation of the component fatty acids gives phosphatidylcholines and sphingolipids different properties in the membrane. Cholesterol’s planar headgroup fits between the head groups of glycerol and sphingolipids, helps maintain normal viscosity over a wide range of membrane compositions, and reduces membrane permeability to water. Oxidized fatty acids reduce packing density, increase membrane fluidity, and increase permeability.
Figure 2.
Membrane Lipid Head Groups with Attached Fatty Acid Tails. Phosphatidylcholine is pictured as a representative glycerophospholipid (top right). Each phosphatidylcholine headgroup has two fatty acids attached with the most usual arrangement being a saturated fatty acid at position one and an unsaturated fatty acid at position two[86]. Sphingomyelin has a larger headgroup and generally has more LCSFA. Length and unsaturation of the component fatty acids gives phosphatidylcholines and sphingolipids different properties in the membrane. Cholesterol’s planar headgroup fits between the head groups of glycerol and sphingolipids, helps maintain normal viscosity over a wide range of membrane compositions, and reduces membrane permeability to water. Oxidized fatty acids reduce packing density, increase membrane fluidity, and increase permeability.
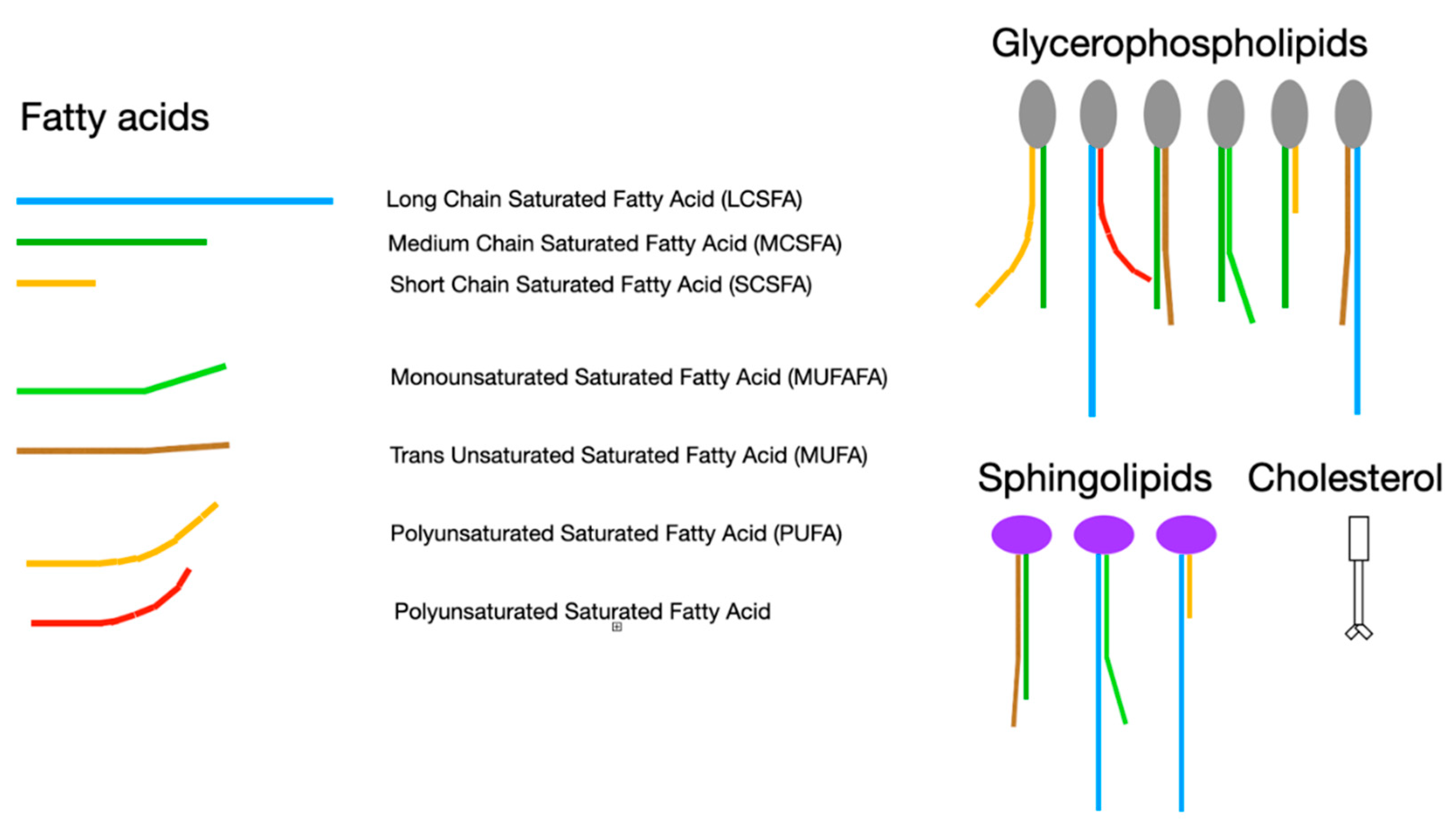
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



