
Submitted:
30 April 2024
Posted:
01 May 2024
You are already at the latest version
Abstract
Mesenchymal stem cells due to their multipotent ability are considered one of the best candidates to be used in regenerative medicine. The most used source of those stem cells long the last decades was the bone marrow, despite its limited cell resources and the painful and very invasive surgical treatment. Therefore, the scientific community investigated many alternative sources for the harvesting of these stem cell subpopulations. Adipose tissue attracted the biggest interest thanks to special features, such as great tissue abundance, easy availability, and in the end the large amount of positive mesenchymal stem cells. Once it was verified that these stem cell subpopulations possess the same bone marrow differentiation potential, it became evident that they would be the ideal candidate to be used in regenerative and tissue engineering applications. Even the regenerative use of these cells long become a sensitive point because not all the acquired phenotypes resulted in the best source for plastic procedures. As documented in the literature, the ideal fillers for regenerative is represented by stem cell population. Therefore, it became crucial to investigate any kind of biological mechanism on the differentiative ability of those mesenchymal stem cells, especially their regulative factors. One of the most studied and important regulators are the miRNAs. Indeed, miRNA play a crucial role in the differentiation process of adipose stem cell cultures isolated from patients or obtained by commercial cell lines toward osteocytes, chondrocytes, and adipocytes differentiated populations.
Keywords:
mesenchymal stem cells (MSC)
; adipose tissue
; miRNA
; regenerative medicine
; adipose stem cells
1. Introduction
Adipose tissue is an endocrine organ, that acts as an energy triglycerides reservoir, and in case of necessity, it mobilizes energy by releasing fatty acids. This tissue is managed by endocrine, metabolic responses, and cellular composition through the potentially toxic buildup of lipids and their mobilization to limit biotoxicity due to lipid excess in peripheral organs [1,2]. Adipose tissue plays a crucial role in buffering chronic overnutrition by storing excess energy in the form of lipids. However, if this energy storage capacity is exceeded, it can result in pathological accumulation of lipids in other organs. There are three main types of adipose tissues, including subcutaneous and visceral white anabolic white (WAT), catabolic brown adipose tissue (BAT), and mixed adipose tissue. White adipose tissue mainly controls energy excesses by promoting hypertrophy or hyperplasia, which increases its energy-buffering ability. To achieve this, the tissue uses committed progenitors. On the other hand, brown adipose tissue is responsible for energy dissipation by producing heat and possesses multilocular fat cells with mitochondria, which promote the oxidation of lipids. WAT has been historically defined by anatomical location and the presence of parenchymal cells containing a single large lipid droplet. Researchers have debated the high adipocyte plasticity. [3].
It is crucial to conduct a deep study of all the cellular types that make up the adipose tissue, as well as the external factors that influence their development and adaptability. Adipose tissue is highly heterogeneous, with most cells exhibiting significant multipotency. The tissue primarily consists of mature adipocytes, in addition to a mixture of small mesenchymal stem cells (MSCs) and pre-adipocytes, endothelial cells, macrophages, and T regulatory cells. In particular, pre-adipocytes, which reached our attention, can proliferate and differentiate into mature adipocytes, providing the adipose tissue with a constant and very high functional adaptability. (Figure 1) [3,4]. The process of adipose tissue formation is well-regulated and occurs in different stages, with varying cell organization and differentiation pathways. Several studies have shown that different adipose tissue depots form at different times and possess unique molecular characteristics, indicating regional differences between them. Additionally, research into stem cell adipose biology has focused on understanding the molecular characteristics and differentiation potential of progenitor cells within this tissue [5,6,7,8,9]. Stem cells reside in specialized locations known as stem cell niches, where they remain dormant until adjacent cells or external signals from the microenvironment stimulate them to proliferate and differentiate. Adipogenesis is closely linked to other developmental processes, with angiogenesis being the most important among them. [10,11,12,13] Moreover, this process directly influences pre-adipocyte proliferation and differentiation, reducing in this way the vascularization during obesity, thus inducing apoptosis in dysfunctional adipocytes [14,15,16].
Furthermore, adipose tissue stromal components provide structural support and biochemical signals, maintaining tissue functionality. Extracellular matrix (ECM) components impact mesenchymal lineage fate, proliferation, and cell differentiation, while mesenchymal progenitors play an essential role in matrix remodeling. [17,18,19]. Adipocyte maturation upregulates collagen IV expression and several laminin complexes, while it reduces fibronectin expression during adipocyte differentiation [20,21,22]. Another regulation of adipogenesis regarding the so-called adipocytokines, this secreted class of molecules exerts a significant effect on every adjacent cell.
For these reasons, adipose tissue could be considered not only a simple energy storage tissue system but also an important endocrine and immune organ, even if it is not yet fully characterized.
As debated above, the adipocyte differentiation process mainly comprises two stages, the “determination phase” and then the “terminal differentiation phase”. [28,29,30,31,32]
During the beginning phase of adipogenesis, C/EBP-β and C/EBP-δ factors are identified as crucial transcriptional factors. These factors accumulate, causing adipose cells to re-enter the cell cycle and promoting the transition from active G1 to S phase. Specifically, when C/EBP-β is hyperphosphorylated and then activated by glycogen synthase kinase-3 B and mitogen-activated protein kinase (MAPK), it works together with C/EBP-δ to stimulate the expression of PPARγ and C/EBP-α. Hence, both C/EBP-β and C/EBP-δ play a vital role in regulating the transcription of genes that are critical for adipocyte differentiation. [33,34]. Therefore, both factors promote their expressions, but meanwhile, they can promote many other genes whose expression could induce specific adipocyte commitment or fate [35,36]
PPARγ represents an important nuclear receptor and it is considered a crucial transcription factor that drives brown or white adipocyte differentiation. Two PPARγ isoforms have been described: PPARγ1 which is constitutively expressed, and its expression is characteristic of fat tissues, and PPARγ2, regulated by the previous one, which regulates mainly adipocyte differentiation [37,38]. Adipocyte commitment results from the equilibrium between these factors, due to the dual face of these factors, which could be considered pro- and anti-adipogenicity transcription factors.
The Kruppel-like factors (KLFs), instead, of C2H2 a zinc-finger factor, regulates adipose tissue apoptosis, proliferation, and differentiation. It described a lot of different isoforms with different biological and molecular roles, such as KLF15 (which promotes GLUT-4 expression), KLF5 (induced by C/EBP-β and C/EBP-δ in the early adipocyte maturation), KLF9 (key pro-adipogenicity transcription factor through the middle adipogenesis stage), KLF2 and KLF7 (anti-adipogenicity factors, and in particular KLF2 represses the Pparg2 promoter) [39,40,41].
Many other groups studied other transcription factors, that could act specifically on the repression of adipogenesis, such as the GATA-binding family or forehead families Forkhead Box O1 and A2 (FOXO1 and FOXA2). So, we could conclude that adipogenesis resulted from positive or negative stimuli and regulation factors, many of which are hormones, cytokines, growth factors, and some pharmacological compounds. [42,43,44,45,46]. (Figure 2). To note, within adipose tissue heterogeneity, which is represented by a dynamic mixture of cellular and non-cellular elements, including progenitors, resident/recruited immune cells, fibroblasts, blood vessels, lymphatic vessels, peripheral nerves, and extracellular matrix (ECM), pre-adipocytes play a crucial role in adipose tissue plasticity. In this context, many studies demonstrated that miRNA expression in pre-adipocytes is altered during adipose tissue development and in obesity, therefore a better understanding of miRNAs' role in the proliferation and differentiation of adipocytes during fat cell development could provide new therapeutic strategies in regenerative medicine. The miRNAs are one of the most important biology and medicine scientific discoveries. miRNAs consist of a short class of RNA molecules, long about 20 nucleotides, endogenous, single-stranded, and non-coding RNA. They are involved in the negative post-transcription gene expression pairing of some specific mRNAs 3'UTR. The miRNA mediates RNA degradation or its translational repression [23]. Scientific evidence showed that miRNAs play a crucial role in the regulation of several biological behaviors, such as embryonic maturity, cell proliferation, metabolic homeostasis, and programmed cell death [24]. Nowadays, about 1,000 different genes coding for miRNAs have been recognized [25]. The interaction involves the formation of a double-stranded assembly between the miRNA '' seed and the mRNA “target” [26,27].
In recent literature, Michael et al. 2009 and Kim et al. 2010 have described the relationship between miRNAs and the regulation of early adipocyte differentiation. In particular, they studied how miR-27b overexpression correlates with adipogenesis, and both groups found that its abundance during human adipocyte differentiation decreases the PPARδ and C/EBPα induction. Therefore, the miR-27 gene family is potentially an important class of adipogenic-sis-negative regulators, useful as an anti-adipogenic factor. Then, another miRNA, the miR-519d, was studied by Martinelli et al. 2010 and is crucial in adipocyte development. This specific miRNA suppresses in a dose-dependently way the translation of PPARα protein, increasing lipid accumulation during preadipocyte differentiation. Moreover, Jordan et al. described the implication of miR-138 expression in the adipogenicity differentiation. In particular, miR-138 is downregulated during the adipogenicity differentiation of human adipose tissue-derived, while its overexpression in mesenchymal stem cells reduces lipid droplets accumulation [47]. In addition, Sun et al. 2009 highlighted the miR-31 role in mesenchymal stem cells (MSCs). When miR-31 was upregulated, adipogenic differentiation of MSCs was repressed, regulating C/EBPα expression. In addition, Tang et al. 2009 reported that during the differentiation of adipose-derived stem cells (ADSCs), the expression of miR-31 and miR-326 were significantly down-regulated. Furthermore, Esau et al. 2004 showed that miR-143 levels increased in differentiating adipocytes, and its ablation inhibits adipocyte differentiation, acting on the MAPK signaling pathway, even if in the terminal differentiation step [33]. Moreover, ERK5, which is involved in adipocyte differentiation, represents a target gene of miR-143, and Oskowitz et al. 2008 observed that relative overexpression promoted adipogenicity differentiation. Instead, Ling et al. 2011 studied the miR-375, which suppresses the phosphorylation levels of ERK1/2 and consequently promotes adipocyte differentiation [48]. Therefore, miRNA inhibitors toward MAPK could be used as a novel approach to reduce adipocyte differentiation and decrease lipid accumulation. In addition, the miR-143 affects negatively glucose homeostasis through the activation of the Akt pathway and specifically downregulates the oxysterol-binding- protein-related protein 8 (ORP8) [49]. Yi et al. 2011 described how the miR-143 could enhance adipogenesis with pleiotrophin (PTN) silencing, resulting in a negative adipogenesis differentiation through PTN/PI3K/AKT pathway. Despite the miR-375 modulates, by targeting PDK, the glucose-mediated stimulatory effect on insulin gene expression, inactivates the Akt pathway thanks to phosphatidyl-inositides generated by PI3K [50]. It described other miRNAs, including miR-210, miR-148a, miR-194, and miR-322, that induce adipogenesis, repressing Wingless-type MMTV integration site family members (Wnt) signaling. Other miRNAs, as miR-344, miR-27, and miR-181, inhibit adipogenesis secreting glycoproteins through their Frizzled (Fz) receptors and low-density lipoprotein receptor-related protein (LRP) co-receptors. As described in the literature, Wnt signaling blocks adipocyte differentiation inhibiting PPARγ and CEBPα expression [51,52]. Furthermore, the relationship between lipid metabolism and miRNA modulation has been highlighted in several papers. For instance, miR-210 overexpression induces hypertrophy and lipid droplet formation in the fat cell and its inhibition promotes the adipogenesis block [53]. miR-103 is reported to be upregulated during differentiation of human pre-adipocytes, and its overexpression in the presence of adipogenic stimuli increases triglyceride accumulation and adipogenic gene expression [54,55,56,57,58]. In mature adipocytes, long terminal differentiation, the upregulation or downregulation of specific miRNAs, such as miR-221, miR-125b, miR-34a, and miR-100, have different effects. In particular, miR-34a, positively upregulated during adipogenesis, is associated with BMI increasing [59]. Despite miR-448 suppressing adipocyte differentiation. Kruppel-like factor 5 (KLF5) contains a selective miR-448 binding site. Overexpression of miR-448 in pre-adipocytes suppresses KLF5, triglyceride accumulation, and adipogenic gene expression [60]. Moreover, miR-15a inhibition reduces preadipocyte size while promoting adipocyte proliferation. In preadipocytes, miR-15a has been shown to target DLK1 at mRNA and protein level [61]. Furthermore, miR-222 and miR-221 are decreased during adipogenesis but upregulated in obese adipocytes, contrary, miR-185 was upregulated in mature adipocytes while downregulated in obese men. In summary, there is a strong influence of the miRnoma on the potential differentiation and several miRNAs have been identified which can accelerate or inhibit adipocytes turnover.
2. Results
2.1. Different Culture Conditions Affect miRNA Expression
The miRNAs analysis comparison was performed between the two adipose stem subpopulations, ASphCs (primary adipose stromal cells grown as floating spheroids) and ADSCs (commercial adipose cells grown in adhesion), and their relative mature adipocyte populations ADA (ADSC-Derived Adipocytes) and SDA (ASphC-Derived Adipocytes). The lists of miRNAs and the cluster-grams obtained highlighted a wide negative transcriptional regulation when we compared ASphCs, ADSCs, and ADA with SDA (Figure 3 A) (Table 1). Furthermore, when we compared the two adipose stem populations it is possible to note a different miRNA pattern of expression with a score index of 85%. Thus, we could assume that the culture conditions, like non-adherence and fetal bovine serum (FBS) absence, are important for the expression of those miRNA expressions that are mutually expressed in ASphCs or ADSCs. Indeed, we demonstrated that ASphCs, which grow in the presence of a specific stem FBS-free medium, reside in a quiescent state and possess greater cellular plasticity than ADSCs [60]. Therefore, these multipotential differences could be due to these specific miRNA expressions that are involved in the key biological process.
More in detail, ASphCs overexpress the miR-126, which is a negative cell growth and angiogenesis regulator (silencing VEGF, FGF, and EGF-L). and the same cells showed the miR-146 family overexpression, which is composed of mir146a and mir146b. The first one is an important negative cell cycle regulator, and the second promotes adipogenesis commitment. In addition, ASphCs showed high expression levels of miR-199b that activate transcription factor HES1 by inhibiting leukemia inhibitory factor (LIF), together with a high level of miR-25 which is a p27 inhibitor control. ADSCs contrarily, have highlighted the overexpression of miR-143, a negative adipocyte commitment regulator, and miR-145 which inhibits SOX9 and ROCK genes. It also showed miR-494 overexpression that inhibits both the growth and angiogenesis potential of mesenchymal stem cells [49,50,51,52] (Figure 3 B) (Table 1).
2.2. MicroRNAs Regulate Adipocyte Differentiation
After completing the comparison of Mesenchymal stem cell cultures, we conducted a thorough investigation of the difference in miRNA regulation between their derivatives and mature adipocytes. (Figure 4) (Tab 2 and Tab 3).
Our analysis revealed a well-regulated miRNA process. Adipogenesis is a highly regulated process specifically in adipose stem cultures, where we found several miRNAs to be overexpressed, such as miR-31, which inhibits the C/EBP gene, and miR-27, which negatively represses the PPARγ gene. These two genes are crucial for adipogenesis, and their inhibition prevents adipocyte maturation. Furthermore, overexpression of miR-27 and high levels of miR-186, which activate the Wnt pathway, may explain the undifferentiated status of these stem cell populations. Additionally, the adipocyte lineage commitment of ASphCs is under the regulation control of miR-125b, which is overexpressed and responsible for MAPK silencing, as shown by data in the literature. ASphCs also show a high level of miR-100 and miR-199a3 expression, which inhibits the mTOR pathway and thus blocks cell cycle progression, leading to stemness maintenance at the expense of adipocyte commitment. Furthermore, miR-100, miR-31, miR-222, and Let7, which were commonly found to be overexpressed in ASphCs and ADSCs compared to the differentiated mature adipocytes derived from them, act as negative regulators of adipogenesis and cell cycle. This information is further illustrated in Figure 4 A-D and Table 2.
3. Discussion and Conclusions
All the data obtained from this vast miRNAs’ analysis on adipose stem cell subpopulations together with their differentiated progenies suggest to us that adipogenesis is carefully controlled by several factors and at many different levels, therefore to obtain a deed acknowledgment of these molecular mechanisms this descriptive analysis should be followed by a functional analysis. As described by many scientific works, adipose tissue is often used in regenerative medicine as an autologous filler for the correction of morpho-functional defects. Therefore, the adipocyte development control could be very helpful to exploit the autologous filler for regenerative purposes. In particular, we can speculate on a possible viable engineering therapy where an ectopic expression of miRNAs could be exploited to promote a specific commitment (i.e. we could get a pre-adipocyte population compared to a population of mature adipocytes, overexpressing mir27a or the mir31, limiting the harmful effect of adipocytes in lipofilling in cancer patients subjected to demolition surgery). Alternatively, we could try to overexpress mir100 or mir146a, succeeding in limiting the onset of breast cancer or succeeding in reverting a more aggressive phenotype of breast cancer to one that responds to anti-cancer therapies. Moreover, in the case of ectopic mir455, we could induce an adipose-type switch from WAT to BAT and thus minimize the damaging effects of obesity and harmful white adipose tissue paracrine secretion [55,56]. (Figure 5)
In conclusion, despite the broad possibility of understanding the role of microRNAs in the adipogenic pathway, an obvious limitation is the in-depth in vitro studies. A possible perspective could be based on the use of microRNAs identified as a possible strategy for the modulation of adipocyte-based processes. Furthermore, the delivery of such microRNAs through nanovesicles could deepen the study and mechanistic understanding of the biological and differentiation processes shown.
4. Materials and Methods
4.1. Adipose Tissue Samples and Cell Culture
The adipose tissues were extracted from a lipoaspirate tissue biopsy, in compliance with our department’s policy. Patients were treated in compliance with our department’s policy, following the patient’s written consent on adipose tissue harvest and its use for research purposes. The study was approved by the ethics committee Palermo-1 Polyclinic Paolo Giaccone of Palermo with report N°1/2016. The withdrawal of adipose tissue was performed under conscious sedation through local infiltration of 150 cm3 of Klein solution. A 10-gauge cannula connected to a 10-ml syringe with a luer-lock-type attack was used for the liposuction. Then 90–140 cm3 of subcutaneous adipose tissue was extracted from each patient. Lipoaspirates from healthy subjects were obtained using a liposuction procedure. These subjects had been previously screened to ensure the absence of chronic illness. The sample was centrifuged for 5 min at 700g. After centrifugation, the samples were stratified into three layers, the upper layer representing the oily component, the middle one consisting of a solid tissue layer, and the bottom layer composed of MSCs and blood elements. Lipoaspirates from donors were digested for 30 min at 37 °C, in the presence of collagenase (1.5 mg/ml; GIBCO) and hyaluronidase (20 mg/ml; Sigma). The digested samples were centrifuged and washed with PBS. The obtained pellet was suspended in serum-free stem cell-specific medium, supplemented with bFGF (10 ng/ml; Sigma) and EGF (20 ng/ml; Sigma), in ultra-low adhesion culture flasks (Corning) as described previously [61]. In these conditions, cells grew as floating spheroids, called AD-MSCs StemPro™ Human Adipose-Derived Stem Cells (ADSCs; ThermoFisher), cultured as the manufacturer recommended, were used as the MSC control.
4.2. Adipogenic Differentiation
Both ASphCs and ADSCs were trypsinized for 5 min at 37°C and cultured into adherent 24-well cell culture plates (50.000 cells/well). Cells were cultured in STEMPRO® Adipogenesis Differentiation Kit (Invitrogen) for up to 28 days. Cell viability, adhesion, and differentiation were assessed by daily observation using optical microscopy. We assess the mature adipocytes with AdipoRed TM Assay Reagent (Lonza). We referred to SDA: ASphC-Derived Adipocytes and to ADA: ADSC-Derived Adipocytes.
4.3. miRNA Gene Expression
We extracted the total RNAs by TRIzol® Reagent (Ambion) following the manufacturer's instructions. We evaluated miRNA expression by Megaplex pools protocol specific for a set of 384 microRNAs (pool A) as recommended by the manufacturer (Applied Biosystems). The relative quantification of microRNA expression was calculated using the equivalent Ct values where the original CT values are projected to 100% target efficiency. All the experiments were normalized using the global normalization method microRNA assays were performed for ASphCs, ADSCs, and their differentiated cells. At least two replicates were run for each sample. miRNAs with 3-fold changes were considered for analysis. [27]
Author Contributions
AG: Investigation, Data curation, Methodology, Writing–original draft, Writing–review and editing. SDF: Data curation, Methodology, Writing–original draft, Writing–review and editing. ALD Supervision, Writing–review and editing. GS: Conceptualization and Writing–review and editing.
Funding
This research was funded by “The National Biodiversity Future Center” grant number “CUP B83C22002930006”; by MUR grant number "PRIN P2022YAKJY"; and by European Union – NextGenerationEU initiative under the Italian Ministery of University and Research as a part of the PNRR – M4C2-l1.3 grant number “CUP B73C22001250006”. The APC was funded by the National Biodiversity Future Center.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by Ethics Committee 1 board, University of Palermo - Azienda Ospedaliera Universitaria “Paolo Giaccone” with report N°1/2016.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
No datasets have been created during the study. The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- R.H. Unger, Lipotoxic diseases, Annu. Rev. Med. 53 (2002) 319–336.
- Cinti S. The adipose organ: morphological perspectives of adipose tissues. Proc Nutr Soc. 2001 Aug;60(3):319-28. [CrossRef] [PubMed]
- Cinti S. The adipose organ. Prostaglandins Leukot Essent Fatty Acids. 2005 Jul;73(1):9-15. [CrossRef] [PubMed]
- Guzik TJ , Mangalat D, Korbut R. Adipocytokines - novel link between inflammation and vascular function , J Physiol Pharmacol. 2006 Dec;57(4):505- 28.
- R. Schofield, The relationship between the spleen colony-forming cell and the haemopoietic stem cell, Blood Cells 4 (1978) 7–25.
- V. Greco, S. Guo, Compartmentalized organization: a common and required feature of stem cell niches? Development 137 (2010) 1586–1594.
- K.A. Moore, I.R. Lemischka, Stem cells and their niches, Science 311 (2006) 1880–1885.
- L. Li, H. Clevers, Coexistence of quiescent and active adult stem cells in mammals, Science 327 (2010) 542–545.
- W. Tang, D. Zeve, J.M. Suh, D. Bosnakovski, M. Kyba, R.E. Hammer, M.D. Tallquist, J.M. Graff, White fat progenitor cells reside in the adipose vasculature, Science 322 (2008) 583–586.
- K. Iyama, Electronmicroscopical studies on the genesis of white adipocytes: differentiation of immature pericytes into adipocytes in transplanted preadipose tissue, Virchows Archiv. B, Cell pathology 31 (1979) 143–155.
- S. Cinti, A morphological study of the adipocyte precursor, J. Submicrosc. Cytol. 16 (1984) 243–251.
- H. Eto, H. Suga, D.Matsumoto, K. Inoue, N. Aoi, H. Kato, J. Araki, K. Yoshimura, Characterization of structure and cellular components of aspirated and excised adipose tissue, Plast. Reconstr. Surg. 124 (2009) 1087–1097.
- J.G. Granneman, P. Li, Y. Lu, J. Tilak, Seeing the trees in the forest: selective electroporation of adipocytes within adipose tissue, American Journal of Physiology.
- Y. Cao, Angiogenesis modulates adipogenesis and obesity, J. Clin. Invest. 117 (2007) 2362–2368.
- J. Han, J.-E. Lee, J. Jin, J.S. Lim, N. Oh, K. Kim, S.-I. Chang, M. Shibuya, H. Kim, G.Y. Koh, The spatiotemporal development of adipose tissue, Development 138 (2011) 5027–5037.
- K. Sun, I.W. Asterholm, C.M. Kusminski, A.C. Bueno, Z.V. Wang, J.W. Pollard, R.A. Brekken, P.E. Scherer, Dichotomous effects of VEGF-A on adipose tissue dysfunction, Proc. Natl. Acad. Sci. 109 (2012) 5874–5879.
- A.G. Cristancho, M.A. Lazar, Forming functional fat: a growing understanding of adipocyte differentiation, Nat. Rev. Mol. Cell Biol. 12 (2011) 722–734.
- Divoux, K. Clément, Architecture and the extracellular matrix: the stil unappreciated components of the adipose tissue, Obes. Rev. 12 (2011) e494– e503.
- E.M. Mariman, P. Wang, Adipocyte extracellular matrix composition, dynamics and role in obesity, Cell. Mol. Life Sci. 67 (2010) 1277–1292.
- Nakajima, S. Muroya, R.-i. Tanabe, K. Chikuni, Extracellular matrix development during differentiation into adipocytes with a unique increase in type V and VI collagen, Biol. Cell 94 (2002) 197–203.
- C. Pierleoni, F. Verdenelli, M. Castellucci, S. C., Fibronectins and basal lamina molecole expression in human subcutaneous white adipose tissue, Eur. J. Histochem. 42 (1998) 183–188.
- B.M. Spiegelman, C.A. Ginty, Fibronectin modulation of cell shape and lipogenic gene expression in 3T3-adipocytes, Cell 35 (1983) 657–666.
- Kalebm. Pauley And Edward K.L. Chan MicroRNA in autoimmunity and autoimmune diseases. J Autoimmun. 2009; 32(3-4): 189–194. [CrossRef]
- Stefani G, Slack FJ: Small Non-Codingrnas In Animal development.Natrevmol Cell Biol 2008, 9:219-230.
- Bartel, D.P. Micrornas: Genomics, Biogenesis, Mechanism, And Function. Cell, 116: 281-297, 2004.
- Du, T. & P.D. Zamore. 2005. Microprimer: The Biogenesis and Function Of Microrna. Development 132: 4645–4652.
- Filipowicz,W., S.N. Bhattacharyya&N. Sonenberg. 2008. Mechanisms of Post- Transcriptional Regulationby Micrornas: Are The Answers In Sight? Nat. Rev. Genet. 9: 102–114.
- Gesta S, Tseng YH, Kahn CR (2007) Developmental origin of fat:tracking obesity to its source. Cell 131: 242–56.
- Billon N, Monteiro MC, Dani C (2008) Developmental origin of adipocytes: new insights into a pending question. Biol Cell 100: 563–75.
- Considine RV, Nyce MR, Morales LM, Magosin SA, Sinha MK, Bauer TL, Rosato EL, Colberg J, Caro JF (1996) Paracrinestimulation of preadipocyte- enriched cell cultures by mature adipocytes. Am J Physiol 270: 895–99.
- Marques BG, Hausman DB, Martin RJ (1998) Association of fat cell size and paracrine growth factors in development of hyperplastic obesity. Am J Physiol 275: 1898–908.
- Kishida K, Kuriyama H, Funahashi T, Shimomura I, Kihara S,Ouchi N, Nishida M, Nishizawa H, Matsuda M, Takahashi M, Hotta K, Nakamura T, Yamashita S, Tochino Y, Matsuzawa Y(2000) Aquaporin Adipose, a Putative Glycerol Channel in Adipocytes. J Biol Chem 275: 20896–902.
- Tang QQ, Otto TC, Lane MD (2003b) CCAAT/enhancer-binding protein beta is required for mitotic clonal expansion during adipogenesis. Proc Natl Acad Sci USA 100: 850–5.
- Ramji DP, Foka P (2002) CCAAT/enhancer-binding proteins: structure, function and regulation. Biochem J 365:561–575.
- Wang GL, Shi X, Salisbury E et al (2006) Cyclin D3 maintains growth-inhibitory activity of C/ EBP a by stabilizing C/EBP a -cdk2 and C/EBP a -Brm complexes. Mol Cell Biol 26:2570–2582.
- Huang HJ, Donald JT (2007) Dynamic FoxO transcription factors. J Cell Sci 120:2479–2487.
- Kajimura S, Seale P, Tomaru T et al (2008) Regulation of the brown and white fat gene programs through a PRDM16/CtBP transcriptional complex. Genes Dev 22:1397–1409.
- Morrison RF, Farmer SR (1999) Role of PPARgamma in regulating a cascade expression of cyclindependent kinase inhibitors, p18(INK4c) and p21(Waf1/Cip1), during adipogenesis. J Biol Chem 274:17088–17097.
- Mori T, Sakaue H, Iguchi H et al (2005) Role of Kruppel-like factor 15 (KLF15) in transcriptional regulation of adipogenesis. J Biol Chem 280:12867–12875.
- Gray S, Feinberg MW, Hull S et al (2002) The Kruppel-like factor KLF15 regulates the insulinsensitive glucose transporter GLUT4. J Biol Chem 277:34322–34328.
- Wu J, Srinivasan SV, Neumann JC et al (2005) The KLF2 transcription factor does not affect the formation of preadipocytes but inhibits their differentiation into adipocytes. Biochemistry 44:11098–11105.
- Qian S, Li X, Zhang Y, et al. Characterization of adipocyte differentiation from human mesenchymal stem cells in bone marrow. BMC Dev Biol 2010; 10: 47.
- Esau C, Kang X, Peralta E, et al. MicroRNA-143 regulatesadipocyte differentiation. J Biol Chem 2004; 279: 52361-.
- Ortega FJ, Moreno-Navarrete JM, Pardo G, et al. MiRNA expression profile of human subcutaneous adipose and during adipocyte differentiation. PLoS ONE 2010; 5: e9022.
- Oskowitz A, Lu J, Penfornis P, et al. Human multipotent stromal cells from bone marrow and microRNA: Regulation of differentiation and leukemia inhibitory factor expression. Proc Natl Acad Sci USA 2008; 105: 18372.
- Yang Z, Bian C, Zhou H, et al. MicroRNA hsa-miR-138 Inhibits Adipogenic Differentiation of Human Adipose Tissue- Derived Mesenchymal .
- Yang Z, Bian C, Zhou H,Huang S,Wang S, Liao L, Zhao RC (2011) MicroRNA hsa-miR-138 inhibits adipogenic differentiation of human adipose tissue-derived mesenchymal stem cells through adenovirus EID-1. Stem Cells Dev 20: 259–67.
- Xu PZ, Vernooy SY, Guo M, Hay BA (2003) The drosophila MICRORNA Mir- 14 suppresses cell death and is required for normal fat metabolism. Curr Biol 13: 790–95.
- Jordan SD, Krüger M,Willmes DM, Redemann N,Wunderlich FT, Brönneke HS,Merkwirth C, Kashkar H, Olkkonen VM, Böttger T, Braun T, Seibler J, Brüning JC (2011) Obesity-induced overexpression of miRNA-143 inhibits insulin-stimulated AKT activation and impairs glucose metabolism. Nat Cell Biol 13: 434–46.
- Ouaamari AE, Baroukh N, Martens GA, Lebrun P, Pipeleers D, Van OE (2008) miR-375 targets 30-phosphoinositidedependent protein kinase-1 and regulates glucose-induced biological responses in pancreatic beta-cells. Diabetes : 2708–17.
- Ross SE, Hemati N, Longo KA, Bennett CN, Lucas PC, Erickson RL, MacDougald OA (2000) Inhibition of adipogenesis by WNT signaling. Science 289: 950–3.
- QinL,ChenY,NiuY,ChenW,WangQ,XiaoS,LiA,XieY,LiJ,ZhaoX,He ZY, Mo DL (2010) A deep investigation into the adipogenesis mechanism: profile of microRNAs regulating adipogenesis by modulating the canonical WNT/beta-catenin signaling pathway. BMC Genomics 11: 320.
- Xie H, Lim B, Lodish HF. MicroRNAs induced during adipogenesis that accelerate fat cell development are downregulated in obesity. Diabetes 2009; 58: 1050-7.
- Sun T, Fu M, Bookout AL, Kliewer SA, Mangelsdorf DJ. MicroRNA let-7 regulates 3T3-L1 adipogenesis. Mol Endocrinol 2009; 23: 925-31.
- Gerin I, Bommer GT, McCoin CS, et al. Roles for miRNA-378/378* in adipocyte gene expression and lipogenesis. Am J Physiol Endocrinol Metab 2010; 299: E198-206.
- Lund E, Güttinger S, Calado A, Dahlberg JE, Kutay U. Nuclear export of microRNA precursors. Science 2004; 303: 95-8.
- Nakanishi N, Nakagawa Y, Tokushige N, et al. The upregulation of microRNA- 335 is associated with lipid metabolism in liver and white adipose tissue of genetically obese mice. Biochem Biophys Res Commun 2009; 385: 492-6.
- Sun F, Wang J, Pan Q, et al. Characterization of function and regulation of miR- 24-1 and miR-31. Biochem Biophys Res Commun 2009; 380: 660.
- Esau C, Kang X, Peralta E, et al. MicroRNA-143 regulatesadipocyte differentiation. J Biol Chem 2004; 279: 52361-5.
- Ortega FJ, Moreno-Navarrete JM, Pardo G, et al. MiRNA expression profile of human subcutaneous adipose and during adipocyte differentiation. PLoS ONE 2010; 5: e9022.
- Andersen DC, Jensen CH, Schneider M, et al. MicroRNA-15a fine-tunes the level of Delta-like 1 homolog (DLK1) in proliferating 3T3-L1 preadipocytes. Exp Cell Res 2010; 316: 1681-91.
- Di Stefano AB, Leto Barone AA, Giammona A, et al (2016) Identifcation and expansion of adipose stem cells with enhanced bone regeneration properties. J Regen Med 5(1):1–11.
- Virzì F, Bianca P, Giammona A, Apuzzo T, Di Franco S, Mangiapane LR, Colorito ML, Catalano D, Scavo E, Nicotra A, Benfante A, Pistone G, Caputo V, Dieli F, Pirrello R, Stassi G. Combined platelet-rich plasma and lipofilling treatment provides great improvement in facial skin-induced lesion regeneration for scleroderma patients. Stem Cell Res Ther. 2017 Oct 23;8(1):236. [CrossRef] [PubMed] [PubMed Central]
Figure 1.
Mesenchymal stem cell commitment. The differentiation of mesenchymal stem cells into mature adipocytes involves both a preliminary lineage commitment and subsequent terminal differentiation.
Figure 1.
Mesenchymal stem cell commitment. The differentiation of mesenchymal stem cells into mature adipocytes involves both a preliminary lineage commitment and subsequent terminal differentiation.
Figure 2.
Adipogenesis maturation process. The process of how mesenchymal stem cells (MSCs) develop into mature fat cells by going through different stages of differentiation and maturation.
Figure 2.
Adipogenesis maturation process. The process of how mesenchymal stem cells (MSCs) develop into mature fat cells by going through different stages of differentiation and maturation.
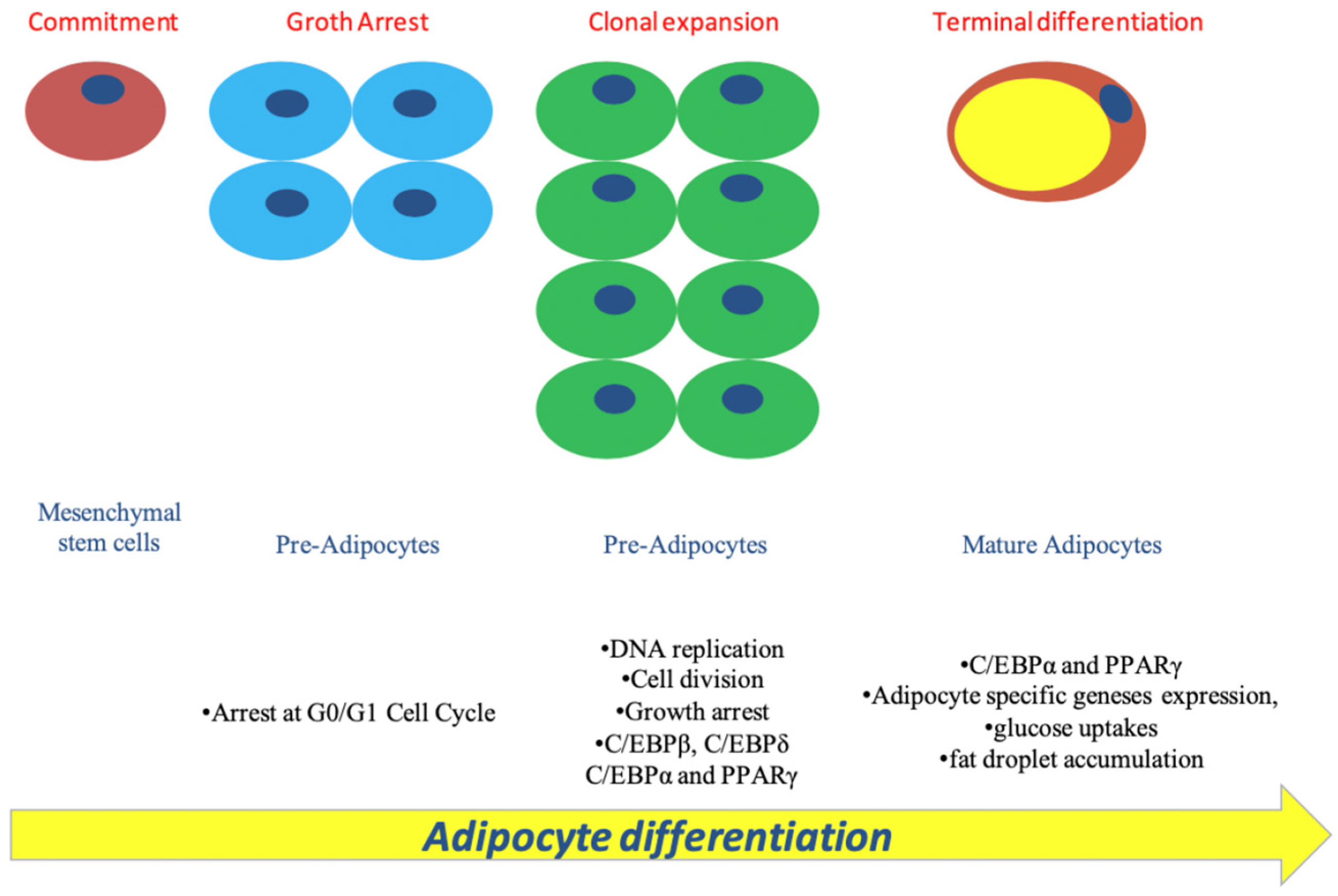
Figure 3.
The differences in terms of multi-difference efficiency could be due to miRNAs expression (A) Clustergrams of mesenchymal stem cell and their derived mature adipocytes ADA (ADSC-Derived Adipocytes) and SDA (ASphC-Derived Adipocytes), the score index is expressed using equivalent Cт values where the original Cт values are projected to 100% target efficiency. (B) Scatter plot of human miRNA expressed, and miRNAs expression reported as fold changes ADSCs compared with ASphCs.
Figure 3.
The differences in terms of multi-difference efficiency could be due to miRNAs expression (A) Clustergrams of mesenchymal stem cell and their derived mature adipocytes ADA (ADSC-Derived Adipocytes) and SDA (ASphC-Derived Adipocytes), the score index is expressed using equivalent Cт values where the original Cт values are projected to 100% target efficiency. (B) Scatter plot of human miRNA expressed, and miRNAs expression reported as fold changes ADSCs compared with ASphCs.
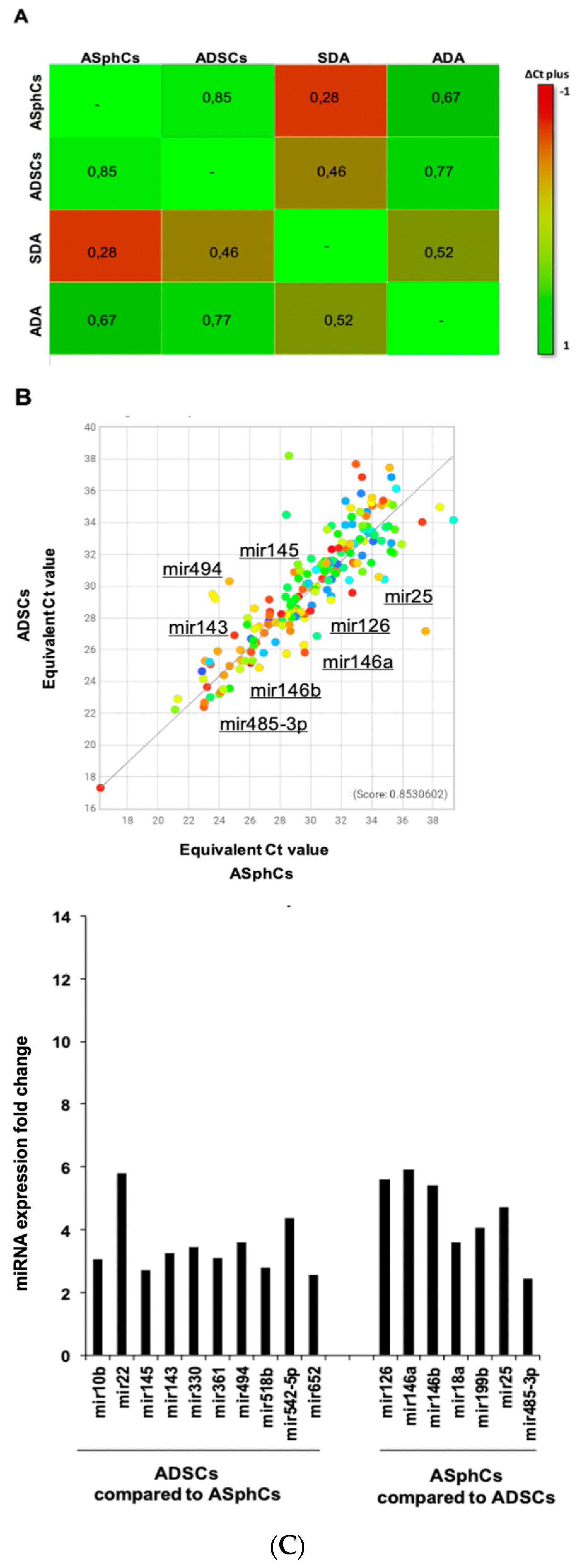
Figure 4.
The miRNAs acting on different pathways. (A) Scatter plot of human microRNA expressed by mesenchymal stem cell ASphCs and their derived mature adipocytes SDA (ASphC-Derived Adipocytes) and (C) relative miRNA expression reported as fold changes. (B) Scatter plots of human microRNA expressed by mesenchymal stem cell ADSC and their derived mature adipocytes ADA (ASphC-Derived Adipocytes) and (D) relative miRNA expression reported as fold changes (lower histogram panel). clustergrams of mesenchymal stem cell ASphCs and their derived mature adipocytes SDA (ASphC-Derived Adipocytes) genes related to adipogenesis genes.
Figure 4.
The miRNAs acting on different pathways. (A) Scatter plot of human microRNA expressed by mesenchymal stem cell ASphCs and their derived mature adipocytes SDA (ASphC-Derived Adipocytes) and (C) relative miRNA expression reported as fold changes. (B) Scatter plots of human microRNA expressed by mesenchymal stem cell ADSC and their derived mature adipocytes ADA (ASphC-Derived Adipocytes) and (D) relative miRNA expression reported as fold changes (lower histogram panel). clustergrams of mesenchymal stem cell ASphCs and their derived mature adipocytes SDA (ASphC-Derived Adipocytes) genes related to adipogenesis genes.
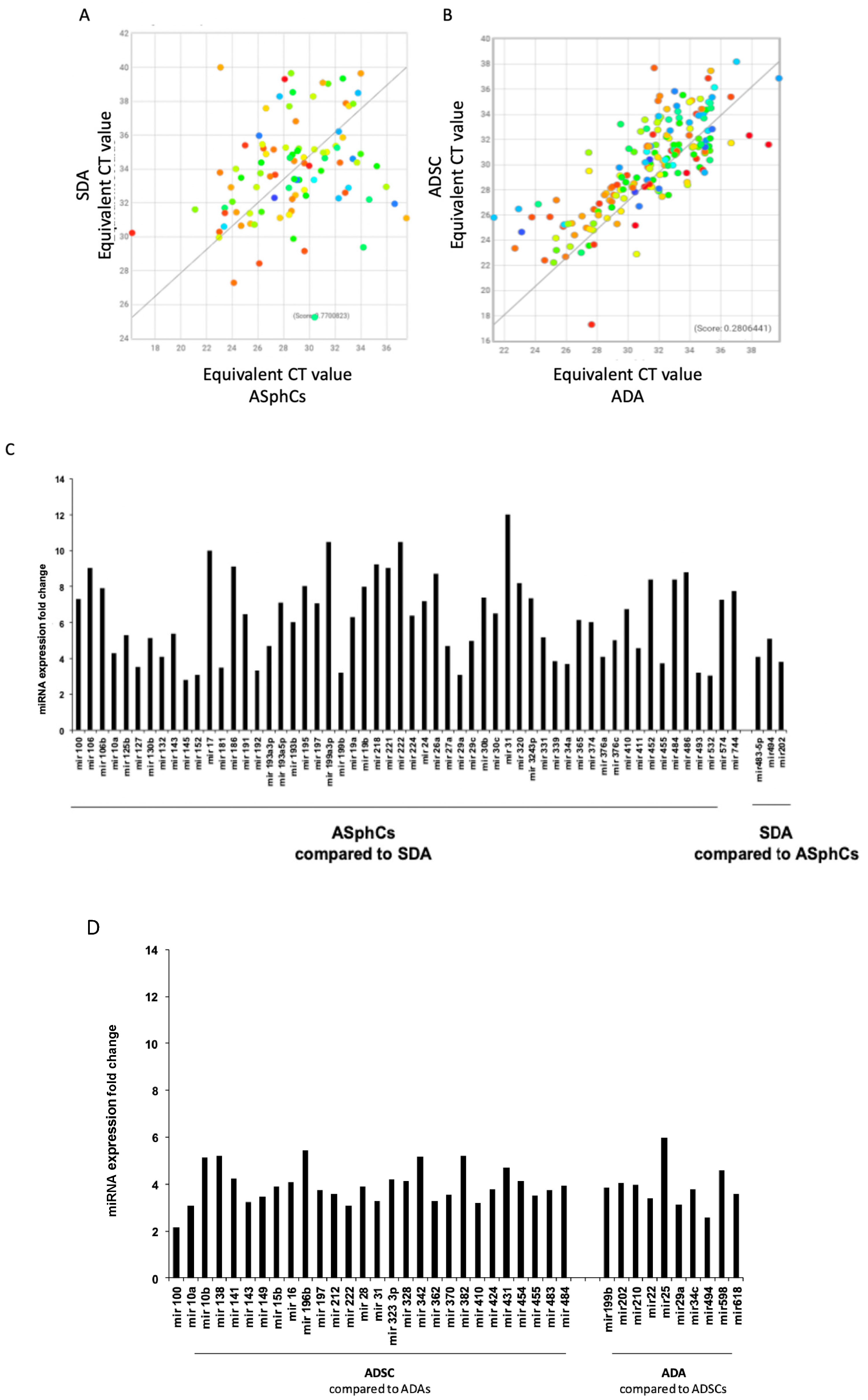
Figure 5.
miRNA families that regulate specific target genes involved in adipogenesis.
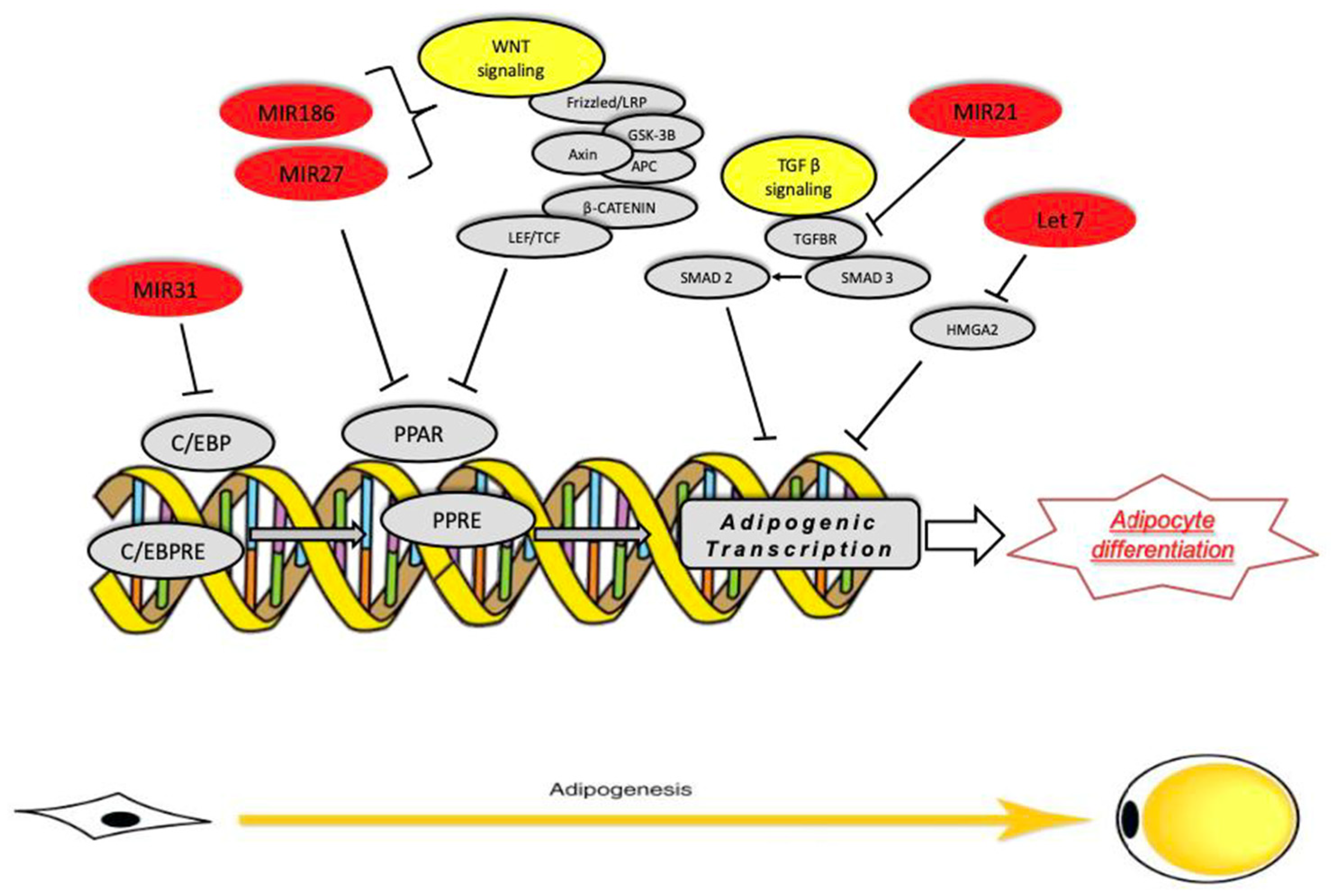

Table 1.
Mesenchymal stem cells miRNA list comparison.
| ASphCs vs ADSCs | |
|---|---|
| miRNA | Fold Change |
| miR-126 | 5,6 |
| miR-146a | 5,9 |
| miR-146b | 5,4 |
| miR-18a | 3,6 |
| miR-199b | 4,05 |
| miR-25 | 4,72 |
| miR-485-3p | 2,43 |
| ADSCs vs ASphCs | |
| miRNA | Fold Change |
| miR-10b | 3,05 |
| miR-22 | 5,8 |
| miR-145 | 2,71 |
| miR-143 | 3,24 |
| miR-330 | 3,45 |
| miR-361 | 3,1 |
| miR-494 | 3,6 |
| miR-518b | 2,79 |
| miR-542-5p | 4,36 |
| miR-652 | 2,55 |
Table 2.
miRNAs belonging to mesenchymal stem cells comparison with their derived adipocytes.
| ASphCs vs SDA | |||
|---|---|---|---|
| miRNA | Fold Change | ||
| miR-100 | 7,3 | ||
| miR-106 | 9,04 | ||
| miR-106b | 7,9 | ||
| miR-10a | 4,29 | ||
| miR-125b | 5,32 | ||
| miR-130b | 5,15 | ||
| miR-132 | 4,09 | ||
| miR-143 | 5,39 | ||
| miR-17 | 10 | ||
| miR-186 | 9,1 | ||
| miR-191 | 6,48 | ||
| miR-192 | 3,32 | ||
| miR-193A-3p | 4,7 | ||
| miR-193A-5p | 7,1 | ||
| miR-193b | 6,03 | ||
| miR-195 | 8,05 | ||
| miR-197 | 7,08 | ||
| miR-199A-3p | 10,5 | ||
| miR-19a | 6,3 | ||
| miR-19b | 8 | ||
| miR-21 | 2,6 | ||
| miR-218 | 9,25 | ||
| miR-221 | 9,03 | ||
| miR-222 | 10,5 | ||
| miR-224 | 6,4 | ||
| miR-24 | 7,2 | ||
| miR-26a | 8,7 | ||
| miR-27a | 4,7 | ||
| miR-29c | 5 | ||
| miR-30b | 7,4 | ||
| miR-30c | 6,5 | ||
| miR-31 | 12 | ||
| miR-320 | 8,2 | ||
| miR-324-3p | 7,36 | ||
| miR-331 | 5,2 | ||
| miR-365 | 6,16 | ||
| miR-374 | 6,02 | ||
| miR-376c | 5,04 | ||
| miR-410 | 6,75 | ||
| miR-411 | 4,6 | ||
| miR-452 | 8,4 | ||
| miR-455 | 3,75 | ||
| miR-484 | 8,39 | ||
| miR-486 | 8,8 | ||
| miR-574 | 7,27 | ||
| miR-744 | 7,75 | ||
| ADSCs vs ADA | |||
| miRNA | Fold Change | ||
| miR-100 | 2,17 | ||
| miR-10a | 3,1 | ||
| miR-10b | 5,11 | ||
| miR-138 | 5,19 | ||
| miR-141 | 4,23 | ||
| miR-143 | 3,22 | ||
| miR-141 | 3,48 | ||
| miR-149 | 3,89 | ||
| miR-15b | 4,09 | ||
| miR-196b | 5,45 | ||
| miR-197 | 3,74 | ||
| miR-212 | 3,58 | ||
| miR-222 | 3,08 | ||
| miR-28 | 3,9 | ||
| miR-31 | 3,29 | ||
| miR-32-3p | 4,21 | ||
| miR-328 | 4,13 | ||
| miR-342 | 5,17 | ||
| miR-362 | 3,26 | ||
| miR-370 | 3,53 | ||
| miR-382 | 5,22 | ||
| miR-410 | 3,21 | ||
| miR-424 | 3,76 | ||
| miR-431 | 4,69 | ||
| miR-454 | 4,13 | ||
| miR-455 | 3,52 | ||
| miR-483 | 3,74 | ||
| miR-484 | 3,94 | ||
| SDA vs ASphCs | |||
| miRNA | Fold Change | ||
| miR-483-5p | 4,1 | ||
| miR-494 | 5,11 | ||
| miR-202 | 3,82 | ||
| ADA vs ADSCs | |||
| miRNA | Fold Change | ||
| miR-199b | 3,87 | ||
| miR-202 | 4,05 | ||
| miR-210 | 3,99 | ||
| miR-22 | 3,39 | ||
| miR-25 | 5,96 | ||
| miR-29a | 3,14 | ||
| miR-34c | 3,79 | ||
| miR-494 | 2,57 | ||
| miR-59 | 4,6 | ||
| miR-618 | 3,59 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



