
Submitted:
27 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
Obstructive sleep apnea (OSA) is a common sleep disorder that is associated with a wide variety of health conditions, including cardiovascular, cerebrovascular, metabolic, neoplastic, and neurocognitive manifestations. OSA as a chronic condition is mainly characterized by repeated upper airway obstructions during sleep that cause episodes of intermittent hypoxia (IH), resulting in tissue hypoxia-reoxygenation cycles. Decreased arterial oxygen pressure (PaO2) and hemoglobin saturation (SatO2) stimulate reflex responses to overcome the obstruction. The prevalence of OSA is significant worldwide, and an underrated problem when focusing on women during pregnancy. The physiological changes associated with pregnancy, especially during its latest stages, are related with a higher prevalence of OSA events in pregnant mothers, and associated with an increased risk of hypertension, pre-eclampsia and diabetes, among other deleterious consequences. Furthermore, OSA during pregnancy can interfere with normal fetal development and is associated with growth retardation, preterm birth, or low term weight. Carotid body overstimulation and hypoxia-reoxygenation episodes contribute to cardiovascular disease and oxidative stress, which can harm both mother and fetus and have long-lasting effects that can reach into adulthood. Because IH is the hallmark of OSA, this review examines the literature available about the impact of gestational intermittent hypoxia (GIH) on the respiratory system at maternal, fetal, and offspring levels. Offering the latest scientific data about OSA during pregnancy, we may help to tackle this condition with lifestyle changes and therapeutic approaches, that could influence the mothers, but also impact adult health problems, mostly unknown, inherited from these hypoxic episodes in the uterus.
Keywords:
gestational intermittent hypoxia
; intermittent hypoxia
; pregnancy
; obstructive sleep apnea
; respiratory system
; placenta
; fetus
; offspring
; oxidative stress
1. Introduction
Obstructive sleep apnea (OSA) is the most common sleep disorder worldwide [1]. It is associated with cardiovascular, cerebrovascular, metabolic, neoplastic and neurocognitive diseases [2,3]. OSA is characterized by repeated complete (apnea) or partial (hypoapnea) obstructions of the upper airway during sleep. With each obstruction, arterial oxygen pressure (PaO2) and hemoglobin saturation for oxygen (SatO2) decrease, making OSA a chronic condition of recurrent intermittent hypoxia (IH), the severity of which is proportional to changes in SatO2 [4]. The decrease in PaO2 activates the carotid body (CB), stimulating motor neurons of the inspiratory and dilator muscles of the upper airways and sleep-wake control areas of the central nervous system [5]. Muscle action and sudden arousals overcome the obstruction, PO2 is restored, and the cycle is repeated, resulting in IH, hypercapnia and sleep fragmentation [6]. CB stimulation activates respiratory reflexes, causing cardiovascular responses which minimize the deleterious effects of hypoxia. Repeated stimulation sensitizes CB chemoreceptors, increasing basal sympathetic tone, leading to hypertension and cardiovascular and metabolic pathology [7].
Epidemiological data on OSA indicate the magnitude of the social, health and economic problem it represents. Its prevalence is estimated to be 9-24% in men and 4-9% in women over 30 years of age, increasing with age [8]. In pregnant women, the prevalence is estimated to be around 8% in the first trimester and increases to nearly 20% in the third trimester of pregnancy [9,10].
Anatomical and physiological changes during pregnancy, or the presence of the disease prior to pregnancy, increase the prevalence of gestational OSA [11]. This pathology is associated with increased morbidity and mortality and can cause hypertension, pre-eclampsia and gestational diabetes mellitus [12]. Episodes of hypoxia-reoxygenation during pregnancy can affect fetal development and postnatal development, resulting in growth retardation, preterm delivery, low birth weight and even fetal death [13]. Adult changes, such as metabolic and epigenetic alterations in male offspring, have also been described in animal models where intermittent hypoxia exposure during pregnancy mimics the hypoxia-reoxygenation cycles of gestational OSA [14]. These episodes of hypoxia-reoxygenation increase the production of reactive oxygen species (ROS), causing tissue damage and an influx of inflammatory cells to the injured sites and into the fetus via the placenta [15].
The repeated episodes of IH that occur during sleep, and especially during pregnancy, can have significant respiratory effects on both the mother and the developing fetus. The following sections highlight the major respiratory changes that occur during pregnancy that may contribute to the development of gestational OSA and how it can affect the pregnant woman, the fetus, and the developing offspring, into adulthood.
2. Maternal Respiratory Changes Related to Pregnancy
2.1. Hormonal Changes
Pregnancy induces hormonal alterations, which are the main cause of changes in the respiratory pattern. Plasma levels of progesterone, which act as a trigger for the primary respiratory center, gradually increase during pregnancy, with one study showing a rise from 27.0 ± 1.5 ng/mL at week 13 to 146.0 ± 17.2 ng/mL at week 37, and then a decrease after delivery (3.5 ± 1.5 ng/mL) [16]. This remarkable increase is accompanied by a rise in the sensitivity of the respiratory center to CO2 [17]. In addition, progesterone acts as a bronchodilator lessening airway smooth muscle tone, and, by acting directly on the CB, increasing the peripheral ventilatory response to hypoxia.
Pregnancy also affects estrogen levels, which rise in parallel with progesterone levels Pregnancy causes an increase in the number and sensitivity of progesterone receptors in the areas of the central nervous system associated with respiration, mainly the hypothalamus and medulla oblongata [18]. Increases in prostaglandin levels during pregnancy have also been described to have a variety of effects, with PGF2α causing bronchial smooth muscle constriction and PGE1 and PGE2 causing bronchodilation [18].
2.2. Anatomical Changes
Pregnancy-induced uterine enlargement increases abdominal pressure, displacing the diaphragm upwards. As a result, pleural pressure increases, leading to small airways obstruction, which reduces functional residual capacity (FRC) (by 20-30%), expiratory reserve volume (ERV) (15-20%) and residual volume (RV) (by 20-25%). The decrease in the height of the chest is accompanied by an increase in other its dimensions, which maintains total lung capacity (TLC) [19]. Along with these changes in FRC and ERV, pregnancy is associated with an increase in the tidal volume (by 30-50%) and minute ventilation (by 20-50%), with only a small increase in respiratory frequency.
As the diaphragm moves upwards, the thorax also undergoes significant changes during pregnancy. There is an increase in the subcostal angle of the rib cage (from 68 to 103 degrees) due to relaxation of the lower rib cage ligaments, and an increase in the circumference of the lower chest wall (5 to 7 cm). These anatomical changes peak at week 37, and largely return to normal by week 24 after delivery (although the subcostal angle remains 20% wider than at baseline) [16]. This increase in chest wall size also helps to compensate for the upward displacement of the diaphragm mentioned above.
In addition, pregnant women have significant upper airway narrowing during the third trimester of pregnancy [20]. This upper airway narrowing may be related to the increased incidence of snoring and sleep-disordered breathing in pregnancy. Alongside this observation, women with pre-eclampsia have also been described to have narrower upper airways when in the supine position compared to pregnant and non-pregnant women without this condition [21]. Besides this narrowing, they have larger neck circumferences, which may be due to differences in fat deposition around the neck, or tissue oedema, which is present in most women with pre-eclampsia. All these factors would certainly promote or increase the severity of OSA.
Besides these effects, the mucosa of the upper airways is also affected during pregnancy, with an increase in mucopolysaccharide content and phagocytic activity, resulting in nasal congestion. This common nasal congestion is known as pregnancy rhinitis, and affects up to 65% of pregnant women, lasting 3 or more weeks [22] and resolving spontaneously 2 weeks after delivery. Although the etiology is unclear, some contributing factors may be smoking, nasal allergy or infections, with the potential of further complications. Nasal obstruction would exacerbate snoring and therefore sleep-disordered breathing.
The respiratory muscles play a key role in pushing during labour. It has been described in animal models that a long period of stretching causes muscle fibers to add sarcomeres in the longitudinal direction, thereby increasing tension [23]. This may well be what happens to the diaphragm and abdominal muscles during the long stretching period of pregnancy, to improve their efficiency in developing expulsive force during labor.
3. Effect of Intermittent Hypoxia in the Pregnant Women
The physiological changes associated with pregnancy may contribute to the development of new OSA in women with previously normal breathing during sleep or worsen an existing condition. The increased ventilatory drive in pregnant women is related to altered inspiratory pressures at the level of the upper airways, with a potential increase in collapsibility, while the characteristic respiratory alkalosis during pregnancy alters the ventilatory pattern, thus increasing the frequency of apneic/hypopneic events [24].
Among the aforementioned physiological changes associated with pregnancy, the decrease in FRC, alongside with increased oxygen consumption, makes pregnant women more susceptible to hypoxemia during obstructive respiratory events [25]. In a study following 12 pregnant women with preexisting mild to severe sleep-disordered breathing (either OSA or upper airway resistance syndrome) who had been treated with continuous positive airway pressure (CPAP) since early pregnancy, half had to increase the pressure of their CPAP device to prevent upper airway occlusion [26]. This readjustment clearly highlights the dynamic changes in upper airway physiology and the exacerbation of preexisting conditions in pregnant women.
The repeated episodes of intermittent hypoxia during pregnancy also have a few downstream effects, including activation of transcription factors (with changes in gene expression), production of ROS and inflammation, among others. One of the major transcription factors affected by hypoxemia is hypoxia-inducible factor-1 (HIF-1), and more specifically its HIF-1α subunit, which is activated in hypoxia and promotes the expression of genes with protective functions (angiogenesis or erythropoiesis) but also harmful inflammatory responses (see [27]). Therefore, elevated serum levels of HIF-1α have been reported in preeclamptic women [28), but also in men with severe OSA [29] whose levels returned to normal after CPAP therapy.
The continuous reoxygenations that follow intermittent hypoxia cycles promote the production of ROS exceeding antioxidant capacities. The accumulation of ROS entails oxidative stress and cellular damage (see [30]), while increasing the production of redox-sensitive transcription factors, such as the HIF-1 and the pro-inflammatory factor NF-κB, involved in the production of cytokines such as TNF-α and IL-1, related to a range of cardiovascular diseases.
In OSA patients and experimental OSA models, increased ROS signaling was found to cause several systemic and cellular responses [see 31], with reported improvements following CPAP therapy [32]. For pregnant women, the available data is scarcer. One study with pregnant women and controls found an association between sleep disruption and serum TNF-α levels [33]. They also found lower serum levels of the anti-inflammatory cytokine IL-4, which were related to longer sleep onset latency times.
More recently, [34] quantified plasma cytokine levels (TNF-α, IL-1β, IL-6, IL-8, and IL-10) during the third trimester of pregnancy in women with and without (less than 5 events/h) OSA, in a population homogeneous in age and body mass index. They reported a direct correlation between the obstructive apnea index and TNF-α levels, and between the apnea-hypopnea index (AHI) and IL-1β levels. These results led them to conclude that IH episodes in the pregnant population, especially during REM sleep, are associated with higher systemic inflammation than in healthy pregnant women without OSA. In addition, they also found an inverse correlation with newborn weight and age. Another study by the same group [35] found a higher sympathetic tone, as assessed by plasma normetanephrine levels, in women with gestational diabetes and OSA than in women with gestational diabetes alone. Additionally, they also found an association between lower nocturnal oxygenation and higher levels of IL-1β.
The cycles of hypoxia/reoxygenation also affect the vascular bed through morphological and functional modifications, leading ultimately to endothelium impairment (see [36]). For example, Yinon D et al [37] evaluated endothelial function using the reactive hyperaemia test. Briefly, they measured the ratio of pulse-wave amplitude between post and pre-occlusion of one arm as an endothelial function index (EFI). They found that pregnant women with pre-eclamptic toxemia, a pregnancy-specific disorder characterized by hypertension, proteinuria, and edema, had a higher respiratory disturbance index (RDI) and a lower EFI than controls. They speculated that the respiratory disturbances damaged the endothelium of blood vessels in women with pre-eclamptic toxemia.
One of the functions of the vascular endothelium is to release relaxing factors, such as NO, to induce vasodilation and prevent organ ischemia. Loss of this ability due to endothelial dysfunction leads to peripheral vasoconstriction and, potentially, hypertension (see [38]), among other deleterious effects. Injury to the endothelial wall, and therefore endothelial dysfunction, is considered to be a major contributor to the pathogenic mechanisms of cardiovascular diseases [39].
A retrospective analysis of 4326 pregnant women diagnosed with OSA in the United States found a direct association between sleep apnea and an increased likelihood of maternal-fetal complications [40]. Along with higher rates of pre-eclampsia and cesarean delivery, there was also a higher prevalence of pulmonary edema, although there was no significant association with an increased risk of mortality. Another study with pregnant women in Turkey [41] examined the relationship between OSA syndrome and pregnancy complications. They found a higher rate of maternal complications (preeclampsia, gestational diabetes, preterm labor, premature rupture of the membranes, and cesarean section) in patients at high risk of OSA syndrome, identified using the Stop Bang questionnaire, than in women at lower risk. Following this trend, a longitudinal study in Taiwan with 8346 pregnant women who reported adverse birth outcomes (premature delivery, abortion, and stillbirth) [42] found an association between the risk of these outcomes and OSA. Interestingly, they also found a dose-response effect, with women who suffered from OSA for a longer period being more likely to experience these adverse outcomes.
4. Effects of Intermittent Hypoxia in the Placenta
The placenta is an organ essential for feeding, breathing, protection and hormone production during fetal development [43]. During pregnancy, the placenta is exposed to a variety of environmental cues that can have an impact on the placental development, fetal growth, and the postnatal phenotype of the offspring. These factors include low oxygen levels, which can be caused by maternal anemia, umbilical cord occlusion, or poor placental vascularization, as well as high-altitude pregnancies or maternal OSA [44,45]. Studies which experimentally mimicked such conditions in animals, including mice, rats, and guinea pigs, have shown that placental and fetal weights are altered (see [46]). In general, these studies show that the specific effects on the placenta appear to depend on the type and the severity of the challenge, as well as its duration and timing relative to placentation. In addition, sexual dimorphism has been reported in the formation, function, and adaptation of the placenta [47], which, like the fetus, is genetically XX or XY.
Similarly to the postnatal role of the lung, the placenta is a “respiratory organ” where fetal blood gases are exchanged indirectly with the environment by a concentration gradient-dependent passive extraction of oxygen from maternal blood, like the process of diffusion and transfer of oxygen from alveolar gas. The human placenta consists of an extensively branched fetal villous tree that is bathed by maternal blood circulating in the intervillous space. The interface between fetal placental tissue and maternal blood is the syncytiotrophoblast, which acts as the endothelium for the intervillous space [48]. Oxygen transfer across the placental barrier is achieved as fetal blood circulates from the umbilical arteries to the capillaries within the placental villi and back to the umbilical vein, whereas maternal blood is filtered through a high-volume intervillous compartment, making the exchange process slower and less efficient than at the alveolar-capillary interface [49]. In addition, the placenta is a high consumer of oxygen, resulting in a PO2 in fetal blood leaving the placenta through the umbilical vein which is significantly lower than the PO2 in the maternal artery [49]. This is not a limitation under normal conditions of oxygenation but may compromise gas exchange during sustained hypoxia or with the intermittent hypoxia episodes that occur in OSA. Notably, there is evidence of hypoxia in the human placenta in OSA [50]. Fluctuations in oxygen delivery from the maternal circulation into the intervillous space following intermittent hypoxia during the process of gas exchange in the placenta result in hypoxia-reoxygenation events that can overwhelm local antioxidant defenses and lead to oxidative stress and pathological placental changes [51,52]. The effects of hypoxia-reoxygenation are well documented in other organs such as the heart, brain and intestine, where its detrimental effects are mainly due to its ability to generate high levels of ROS, oxidative stress and increased inflammation [53]. It is therefore biologically plausible that OSA, via one of these pathways, may adversely affect placental tissue perfusion and oxygenation, potentially predisposing to the development of placental-mediated adverse outcomes.
It has been described that the severity of maternal oxygen deprivation affects the structure and function of the placenta in a sex dependent manner. Maternally induced hypoxia of 12-13% during the last third of gestation in mice resulted in a selective expansion of the labyrinth vascular branching in placentas from female fetuses. In addition, mRNA expression of Glut1, Igf2 and Igf1r, genes important for placental growth, nutrient transport and glucocorticoid signaling, was reduced only in placentas from female fetuses [54,55]. Conversely, when maternal inspired oxygen is reduced to 10%, the labyrinth zone is reduced with a concomitant increase in the volume density of the junctional zone and activation of the invasive endovascular trophoblast cell lineage [56]. The hypoxia-induced changes in gross placental morphology have been associated with altered expression of genes and proteins involved in proliferation, apoptosis, oxidative stress, and cell lineage differentiation in which the hypoxia-inducible factors (HIF) participation appear to be essential [46,55,57].
Gestational IH impairs uterine artery function and causes placental oxidative stress and hypoxia [58]. GIH also has been reported to alter the placental structure in the rat. A morphological study [59] showed an increase in fetal capillary branching, expansion of maternal blood spaces, and an increased number of cells of the external trophectoderm in the placentas from GIH-exposed mothers. These changes precede the growth of the fetus and could lead to lower fetal weights.
Placental development during normal pregnancy occurs in a low-oxygen environment (~2-8% O2) and this "physiological hypoxia" appears to be critical for early placental development and angiogenesis [60,61]. Vascular endothelial growth factor (VEGF) and fibroblast growth factor-2 (FGF2) are two potent angiogenic factors that have been implicated in placental angiogenesis [62,63]. Hypoxia increased the expression of VEGF and its receptors VEGFR-1 and VEGFR-2 in endothelial cells [64]. In a different study hypoxia also increased the expression of FGF receptors and altered the distribution of FGF2 in different cellular compartments [65]. It has been reported that hypoxia may differentially modulate angiogenic factor-stimulated placental angiogenesis depending on the severity and duration of hypoxia exposure. Wang et al [66] showed in human placental artery endothelial (HPAE) cells that in vitro exposure to sustained hypoxia (3% O2) enhanced FGF2 and VEGF-stimulated HPAE cell proliferation but did so independently of the canonical MEK1/ERK1/2 and PI3K/AKT1 pathways, suggesting that hypoxia may induce a very complex signaling network in the placenta. Although limited research has explored whether intermittent hypoxia produces similar complex changes in placental signaling pattern, Weng et al [67] and Badran et al [58] found that in mice gestational IH also induced impairments in placental angiogenesis with an imbalance in production of pro-angiogenic (VEGF and PlGF) and anti-angiogenic factors (sFlt-1 and sENG), and in vascular remodeling.
Along the same lines, human trophoblasts proliferate in vitro under low O2 conditions, but differentiate at higher O2 levels, mimicking the developmental transition they undergo when invading the placental bed to establish the maternal-fetal circulation in vivo. Using both a human trophoblast cell line (HTR8/SVneo) and a primary culture of extravillous trophoblast cells (EVTs), Song et al [68] reported that, in contrast to sustained hypoxia, intermittent hypoxia blocks trophoblast proliferation and induces excessive trophoblast apoptosis by activating the ER stress signaling pathway. They found that intermittent hypoxia can also significantly impair the invasive and migratory ability of trophoblasts which they attributed in part to the reduced secretion of matrix metalloproteinase-2. These changes in trophoblast function may have deleterious implications for proper pregnancy development [69].
However, there is contrary evidence supporting the hypothesis that the influence of OSA on the development of adverse pregnancy outcomes may be mediated at sites other than the placenta. Findings in rats shows that HIF-1α mRNA and its target genes are not altered in placentas at E19 during gestational IH exposure despite a decrease in the arterial oxygen saturation in the mother to ~80–85% with each hypoxic episode [70] Also, gene expression profiling studies of placenta from women with obesity and OSA showed that there were no statistically significant differences in the placental transcriptome between OSA and no OSA women groups [71].
In this context, how intermittent hypoxemia is transmitted from the mother to the fetus in OSA deserves some comment. By simultaneously measuring PO2 in the maternal artery and umbilical vein, Almendros et al [55] showed in a sheep pregnancy model that the PO2 fluctuations resulting from experimental maternal obstructive apnea are significantly smaller in fetal blood than those registered in maternal blood, due to the damping effect of oxygen transfer across the placenta. This work raises an important question about the potential effects of intermittent hypoxia on the fetus during pregnancy, as the placenta may attenuate the severity of hypoxemic changes in the fetus. In addition to potential differences in the structure and physiology of the placenta in sheep and humans and, therefore, in the attenuation of the swings in PO2 which penetrate through the placenta in both species, the study by Almendros et al [72] rises several considerations when translating the findings to pregnant women. Since the severity of hypoxemic swings is attenuated in the fetus, it could be anticipated that hypoxia-reoxygenation, and thus the oxidative stress induced in the various fetal organs and tissues would be lower than that experienced by adult tissues [73]. It is also possible that the sensitivity and tolerance of fetal tissues to very short episodes of intermittent hypoxia, which have so far not been studied, are different from those in adults. Moreover, the impact of intermittent hypoxemia, even of low amplitude, could be different in the fetus and the adult, since fetal mechanisms to regulate the distribution of blood flow among organs are markedly different from those in adults [74,75,76]. These potential differences could influence the epigenetic changes observed in the offspring after GIH [77,78,79].
5. Fetal Respiratory Distress in Gestational Intermittent Hypoxia
As a result of maternal GIH, there is a reduction in oxygen saturation in the maternal blood which could compromise oxygen delivery from the placenta to the fetus, leading to fetal hypoxemia. However, the extent to which the placenta protects the fetus, and therefore the amount of oxygen that reaches the fetus is still unknown as mentioned above.
Although there is not a functional, fully developed gas exchange system in the lungs of the fetus, it doesn´t mean that they remain inactive. There are rhythmic contractions of the diaphragm and other respiratory muscles, known as fetal breathing movements (FBM), which play a key role in lung maturation. They are a distinctive adaptation to the intrauterine environment.
With completely unique characteristics that differ from neonatal and adult breathing, they also differ from maternal respiratory movements. In humans, FBM begin at around week-10 of gestation and increase in frequency with gestational age, occurring up to 35% of the time around week-30 of pregnancy [80,81]. They occur at a frequency of approximately 30-70 contractions/breaths per minute in each bout, followed by apneic periods that can last up to 2 hours near term [82]. Between 28 and 36 weeks, FBM are closely correlated with maternal REM states, with less frequent FBM occurring in non-REM states [83]. The frequency of FBM episodes is also correlated with other factors, such as glucose levels, with a higher incidence described in humans after maternal meals [84]. In terms of respiratory gases, high levels of PaCO2 (hypercapnia) increase the incidence of FBM, while low levels of PaO2 (hypoxia) inhibit FBM in studies in ewes [85] and humans [86].
Based on umbilical cord blood analysis, one study found that maternal snoring during pregnancy was associated with enhanced fetal erythropoiesis [87], suggesting that the fetus becomes hypoxic in utero, at least to some extent, during repeated bouts of intermittent hypoxia. These episodes of fetal hypoxemia, if prolonged or severe, could impair fetal development and increase the risk of adverse perinatal outcomes.
Due to the increased metabolic demands of the fetus during pregnancy, increases in O2 consumption (20%) and CO2 production (35%) have been described in pregnant individuals when compared with their respective controls. These changes do not correlate exactly with the changes in minute ventilation described in the previous section which were around 20 and 50%. This greater increase in minute ventilation translates into an increase in the alveolar (PAO2) and arterial partial pressures of O2 (PaO2) and a decrease in the corresponding partial pressures of CO2 (PACO2 and PaCO2). These increases in oxygen tension are important to facilitate oxygen transfer across the placenta. Thus, measurements of PaO2 during pregnancy found control values of PaO2 around 93 mmHg rising to 105-106 mmHg during the first trimester, and 101-106 mmHg near term [88]. Despite this increase in oxygen tension, the oxygen reserves of pregnant women are lower, making them, and consequently the developing fetus, more susceptible to hypoxic episodes during periods of apnea.
As for the PaCO2 levels, they have been shown to go from 37 mmHg in the non-pregnant state to 28-29 mmHg in the first trimester and 26-30 mmHg near term [88]. These low PaCO2 values lead to chronic respiratory alkalosis, with compensatory renal mechanisms acting to excrete bicarbonate. This causes stimulation of 2,3-diphosphoglycerate (2,3-DPG) synthesis. The resulting increase in 2,3-DPG levels reduces the affinity of maternal hemoglobin for O2, allowing more O2 to be delivered to the fetus, whose major hemoglobin, called fetal hemoglobin or HBf, has a higher affinity for O2 than the adult hemoglobin or HbA.
The transition from fetal to postnatal breathing includes, among other events, the absorption of lung fluid, gaseous expansion of the lungs, activation of the Hering-Breuer reflexes and the appearance of continuous respiration. Birth itself is a delicate process which has hindered more detailed research. Thus, many of the factors involved in the birth process are not yet fully understood. Asphyxia, umbilical cord occlusion or the rise in PaO2 with the first postnatal breath may be involved in the process [89,90], although neither the afferent input from the carotid sinus nerve [91] nor the interruption of umbilical circulation [92] seem to play a role in this process.
6. Offspring Respiratory Disorders following Gestational Intermittent Hypoxia
Although key components of the respiratory system such as phrenic motoneurons and diaphragm muscle begin to develop in utero, there is also an important postnatal maturation process. After birth there is a transition from fetal breathing movements to functional gas exchange. With a less efficient respiratory system, infants develop several counteractive mechanisms to compensate for the increased vulnerability of their respiratory system.
Looking at the evolution of some respiratory values in humans it is known that respiratory rates fall progressively with age, going from 40 breaths/min in neonates to around 12-18 breaths/min in adults, with tidal volume values (adjusted by weight) remaining virtually identical throughout life.
Besides the rapid breathing rate of the fetus, its minute ventilation is higher at any level of CO2 in neonates and infants compared to adults [93], even though chemoreceptor pCO2 sensitivity is almost mature at term. However, preterm infants showed an attenuated sensitivity (and slower response) to CO2 [94].
As for the peripheral pO2 receptors, they need a few weeks of readjustment [95] to gain sensitivity to the higher levels of oxygen of the environment compared to the uterus. This adaptation results in a depression of ventilatory responses to hypoxia that can last up to 6 months after birth in humans [96]. Immediately after birth, rats showed a weak ventilatory response to a hypoxic challenge [97], even though their ventilatory rate was relatively high, probably to compensate for their immature gas exchange machinery.
Regarding the airways, they are a component of the respiratory system that also shows differences between neonates and adults. Thus, the pediatric upper and lower airways are characterized by a higher resistance to flow, mainly due to a smaller diameter, higher collapsibility, and lower pharyngeal muscle tone [98]. Focusing on the upper airways, they have an elliptical shape in neonates. From the subglottic to the cricoid ring, the airways are narrower in the transverse dimension than in the anteroposterior dimension [99] whereas observations of airways in children over 1 year of age showed a circular shape. Smaller and softer, they are more prone to inspiratory collapse. The closure of newborn upper airways is behind central apneas. This closure may be due to either passive pharyngeal collapse or active laryngeal closure [100]. Nevertheless, the cartilaginous structure lengthens and stiffens with maturation during childhood.
Respiratory immaturity is associated with breathing instability, while immature breathing control manifests as central apneas. Both events have been linked, although controversially, to the sudden infant death syndrome [101]. There is an improvement with progressive maturation, with the frequency and duration of central apneas decreasing during the first 12 months of life, with values of 15 apneas longer than 3 seconds per hour reported at 12 months [102].
Newborns also have unique chest wall properties which disappear with time. Thus, newborns have up to 3 times higher compliance levels than adults to allow for greater expansion during breathing. The ribs also contribute to this end, as they are more flexible in the newborn, while they are positioned more horizontally than in the adults, moving only upwards, and not upwards and outwards, as in the adult rib cage. This difference is the main limitation to increase tidal volumes and the reason why infants under 12 months of age rely mainly on diaphragmatic breathing [103].
The values of the functional reserve capacity (FRC) are also subjected to significant changes, with values that increase rapidly after birth [104]. FRC is directly related to the number of alveoli; fewer alveoli in immature lungs provide less support and can lead to peripheral airways collapse. As the lungs grow, they undergo a process of alveolarization which also helps to increase the surface area available for gas exchange.
Exposure to adverse conditions in utero, such as those caused by GIH, may lead to epigenetic changes that affect the postnatal development of the respiratory control. Animal studies have shown that prenatal exposure to intermittent hypoxia can alter respiratory control mechanisms and increase the susceptibility to respiratory diseases later in life [105]. However, the effects of intermittent hypoxia protocols during pregnancy on the developing respiratory system are not yet fully characterized.
The first study of the effects of GIH on respiratory responses in neonatal rats found that normoxic ventilation was increased in GIH offspring compared to their respective controls (1- and 4-months of age), although hypoxic responses were similar [106]. It was postulated that this maladaptive response to postnatal hypoxia could contribute to events such as the sudden death syndrome. Nevertheless, this study had several limitations, one of which was that it didn´t distinguish between male and female pups.
A study of inflammatory status in neonatal GIH-treated rats recorded respiratory motor bursts in brainstem-spinal cord preparations [105]. In newborn rats (0-3 days old) burst frequency was unchanged in GIH animals, although the rhythm was more regular and the rise time tended to be faster, with no effects of gender on these variables in any of the groups. When testing the respiratory motor output elicited by an inflammatory challenge, males showed a significantly reduced burst frequency compared to normal conditions, with no differences between control and GIH animals. However, in preparations from female animals, only the control preparations showed a decrease in burst frequency with the inflammatory challenge, while the GIH group showed no change. Although GIH didn´t alter the expression of pro-inflammatory factors tested in the brainstem or spinal cord (IL-1β, TNFα and COX-2), it did affect the offspring response to a later postnatal inflammatory challenge, in a sex-dependent manner. The authors hypothesized that GIH reprograms the neuroinflammatory response pathways in the female animals so that there is less cytokine production during inflammatory challenges. Cytokines are linked to the sensitivity of the brainstem respiratory centers, which modulate the respiratory frequency in a complex way, ranging from hyperventilation and even respiratory stress syndrome (as in the worst cases of Covid-19, associated with cytokine storm) [107], to respiratory depression. If this were the case, female GIH offspring would have an impaired ability to trigger an immune response against infection, from birth, and perhaps even into adulthood, especially at the level of respiratory system pathologies.
At birth, the newborn comes into an oxygen-rich environment, and oxygenation through the lungs instead of the placenta raises tissue oxygenation within minutes. Reactive oxygen species (ROS) are normally neutralized by antioxidant defense systems. In preterm infants, these defenses are immature, making them more prone to oxidative tissue injury. In addition, preterm infants with inflammation also have an increased incidence of apnea and are at a greater risk for adverse neurodevelopmental outcomes and mortality rates (see [108]).
The frequent episodes of intermittent hypoxia associated with preterm birth have been linked to adverse respiratory outcomes, with a recent article [109] linking them to high levels of oxidative stress. Thus, more than 65% of preterm infants suffer at least one infection during their hospitalization [110]. By noninvasively measuring lipids, proteins, and DNA oxidative stress biomarkers in urine samples from 1 week and 1 month old premature babies (<31 weeks of gestation), the authors found a direct association between intermittent hypoxia and the various oxidative products quantified. In this sense, it could be possible that most of the effects of GIH on the offspring are indirectly caused by the maternal stress. Thus, OSA events in pregnant women were associated with higher levels of systemic inflammation, which was related to the number of obstructive events, especially during the REM phases of sleep [111].
It is known that gestational stress can induce permanent epigenetic changes in the offspring, as shown by an increase in the levels of some chemokines and altered expression of their receptors in a rat model [112]. In this context, sex-specific effects of stress during gestation and development of respiratory control have been described [113], with a higher rate of pathological apneas in newborn male rats, which was associated with a deficit of medulla-derived serotonin (5-HT). A recent study [114] has found that GIH impairs endothelial NO synthesis in rat offspring vessels, highlighting the fact that some detrimental effects of gestational hypoxia may persist well into adulthood.
Underscoring the importance of inflammation in the neonatal respiratory system, even low levels of inflammation in neonatal rats have been proved to impair respiratory control networks [115]. Using brainstem spinal cord preparations, these authors found differential sensitivities of the respiratory control areas to inflammation. Lower levels of inflammation were more disruptive, affecting amplitude and frequency, while more severe inflammation affected frequency only.
Neonatal inflammation also affected CB chemosensitivity, induced spontaneous apneas in vivo and caused significant O2 desaturation [116]. Levels of dopamine, an inhibitor of CB activity, were increased in the CB due to neonatal inflammation. Inflammatory responses in neonates and adults may involve similar mechanisms although it could be that neonatal signaling pathways are different (see [117]). Nevertheless, the paucity of scientific studies on GIH exposure and respiratory control makes it difficult to draw firm conclusions. Not many studies are available either on the role of the respiratory muscles, the final effectors in the respiratory control system. A study of the diaphragm function in rats exposed to GIH found it had no effect on the force-generating capacity of the diaphragm [118], in either male or female animals. Similar results were observed in the same study when the animals were subjected to intermittent hypoxia shortly after birth (postnatal days 22 and 42), suggesting a relative resilience of the diaphragm muscle to hypoxic stress during the perinatal period. However, when examining the sternohyoid, one of several pharyngeal dilator muscles, the same authors found evidence of weakness in the postnatally exposed animals [119]. This effect persisted into adulthood (16 weeks of age). They suggested that this mismatch between the diaphragm and sternohyoid muscles could contribute to a higher risk of airway collapse.
6. Conclusions
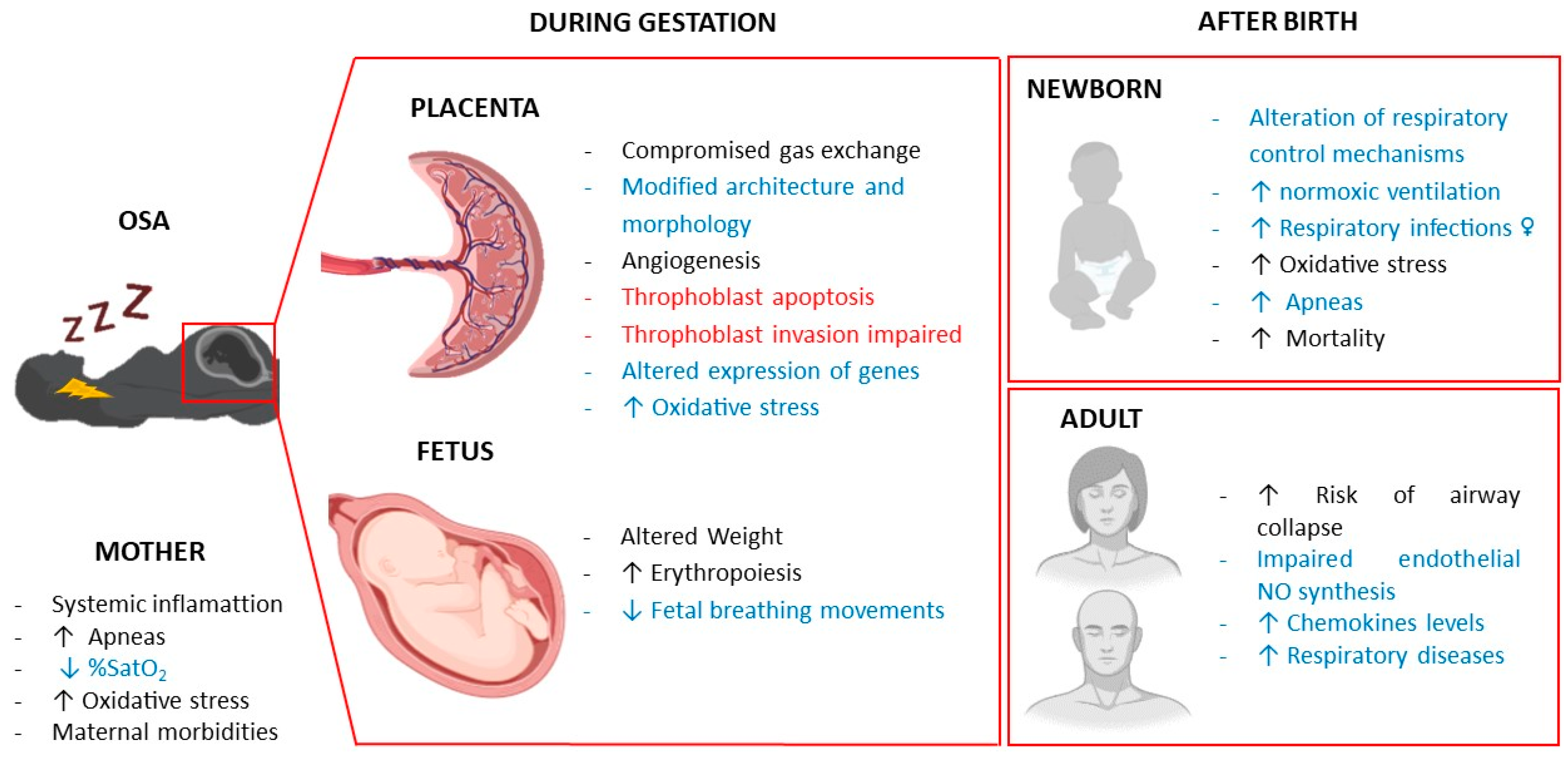
Overall, gestational intermittent hypoxia can have remarkable effects on both maternal and fetal health, with a concomitant risk of adverse pregnancy outcomes and also an increased risk of respiratory dysfunction in the offspring which may persist into adulthood (Figure 1).
Multidisciplinary care involving obstetricians, sleep specialists, and respiratory therapists should be the approach adopted by national health services for the comprehensive management of sleep apnea during pregnancy. Early detection of OSA during pregnancy should be an essential tool for timely intervention. Questionnaires and polysomnography should be first line tools to help clinicians identify women at higher risk for OSA and respiratory disturbances.
Once identified, CPAP therapy is usually the treatment of choice. It helps improve upper airway patency and maternal oxygenation. Other interventions include lifestyle changes, such as weight management, positional therapy, and avoidance of sedatives and alcohol.
Author Contributions
Conceptualization, J.P-L and A.R; writing original draft preparation, J.P-L and A.R.; writing—review and editing, P.I.A, E.O. and E.V-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Canever, J.B.; Zurman, G.; Vogel, F.; Sutil, D.V.; Diz, J.B.M.; Danielewicz, A.L.; Moreira, B.d.S.; Cimarosti, H.I.; de Avelar, N.C.P.; Canever, J.B.; et al. Worldwide prevalence of sleep problems in community-dwelling older adults: A systematic review and meta-analysis. Sleep Med. 2024, 119, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Heinzer, R.; Vat, S.; Marques-Vidal, P.; Marti-Soler, H.; Andries, D.; Tobback, N.; Mooser, V.; Preisig, M.; Malhotra, A.; Waeber, G.; et al. Prevalence of sleep-disordered breathing in the general population: the HypnoLaus study. Lancet Respir. Med. 2015, 3, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Mediano, O.; González Mangado, N.; Montserrat, J.M.; Alonso-Álvarez, M.L.; Almendros, I.; Alonso-Fernández, A.; Barbé, F.; Borsini, E.; Caballero-Eraso, C.; Cano-Pumarega, I.; et al. Spanish Sleep Network. International Consensus Document on Obstructive Sleep Apnea. Arch. Broconeumol. 2022, 58, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, B.; Dasti, S.; Park, Y.; Brown, T.; Davis, H.; Akhtar, B. Hour-to-Hour Variability of Oxygen Saturation in Sleep Apnea. Chest 1998, 113, 719–722. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.; Almaraz, L.; Obeso, A.; Rigual, R. Carotid body chemoreceptors: from natural stimuli to sensory discharges. Physiol. Rev. 1994, 74, 829–898. [Google Scholar] [CrossRef]
- Song, R.; Baker, T.L.; Watters, J.J.; Kumar, S. Obstructive Sleep Apnea-Associated Intermittent Hypoxia-Induced Immune Responses in Males, Pregnancies, and Offspring. Int. J. Mol. Sci. 2024, 25, 1852. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, N.R.; Peng, Y.; Nanduri, J. Carotid body hypersensitivity in intermittent hypoxia and obtructive sleep apnoea. J. Physiol. 2023, 601, 5481–5494. [Google Scholar] [CrossRef]
- Young, T. Rationale, Design, and Findings from the Wisconsin Sleep Cohort Study: Toward Understanding the Total Societal Burden of Sleep-Disordered Breathing. Sleep Med. Clin. 2009, 4, 37–46. [Google Scholar] [CrossRef]
- Pien, G.W.; I Pack, A.; Jackson, N.; Maislin, G.; A Macones, G.; Schwab, R.J. Risk factors for sleep-disordered breathing in pregnancy. Thorax 2013, 69, 371–377. [Google Scholar] [CrossRef]
- Zhu, B.; Bronas, U.G.; Carley, D.W.; Lee, K.; Steffen, A.; Kapella, M.C.; Izci-Balserak, B. Relationships between objective sleep parameters and inflammatory biomarkers in pregnancy. Ann. New York Acad. Sci. 2020, 1473, 62–73. [Google Scholar] [CrossRef]
- Louis, J.M.; Koch, M.A.; Reddy, U.M.; Silver, R.M.; Parker, C.B.; Facco, F.L.; Redline, S.; Nhan-Chang, C.-L.; Chung, J.H.; Pien, G.W.; et al. Predictors of sleep-disordered breathing in pregnancy. Am. J. Obstet. Gynecol. 2018, 218, 521–e1. [Google Scholar] [CrossRef]
- Eleftheriou, D.; Athanasiadou, K.I.; Sifnaios, E.; Vagiakis, E.; Katsaounou, P.; Psaltopoulou, T.; Paschou, S.A.; Trakada, G. Sleep disorders during pregnancy: an underestimated risk factor for gestational diabetes mellitus. Endocrine 2023, 83, 41–50. [Google Scholar] [CrossRef]
- Louis, J.M.; Mogos, M.F.; Salemi, J.L.; Redline, S.; Salihu, H.M. Obstructive Sleep Apnea and Severe Maternal-Infant Morbidity/Mortality in the United States, 1998-2009. Sleep 2014, 37, 843–849. [Google Scholar] [CrossRef]
- Khalyfa, A.; Cortese, R.; Qiao, Z.; Ye, H.; Bao, R.; Andrade, J.; Gozal, D. Late gestational intermittent hypoxia induces metabolic and epigenetic changes in male adult offspring mice. J. Physiol. 2017, 595, 2551–2568. [Google Scholar] [CrossRef]
- Dahlgren, J.; Samuelsson, A.-M.; Jansson, T.; Holmäng, A. Interleukin-6 in the Maternal Circulation Reaches the Rat Fetus in Mid-gestation. Pediatr. Res. 2006, 60, 147–151. [Google Scholar] [CrossRef]
- Contreras, G.; Gutiérrez, M.; Beroíza, T.; Fantín, A.; Oddó, H.; Villarroel, L.; Cruz, E.; Lisboa, C. Ventilatory Drive and Respiratory Muscle Function in Pregnancy. Am. Rev. Respir. Dis. 1991, 144, 837–841. [Google Scholar] [CrossRef]
- Lyons HA, Antonio R. The sensitivity of the respiratory center in pregnancy and after administration of progesterone. Trans Assoc Am Physicians 1959, 72, 173–180.
- Weinberger, S.E.; Weiss, S.T.; Cohen, W.R.; Weiss, J.W.; Johnson, T.S. Pregnancy and The Lung1,2. Am. Rev. Respir. Dis. 1980, 121, 559–581. [Google Scholar] [CrossRef]
- McAuliffe, F.; Kametas, N.; Costello, J.; Rafferty, G.F.; Greenough, A.; Nicolaides, K. Respiratory function in singleton and twin pregnancy. BJOG: Int. J. Obstet. Gynaecol. 2002, 109, 765–769. [Google Scholar] [CrossRef]
- Izci, B.; Vennelle, M.; Liston, W.A.; Dundas, K.C.; Calder, A.A.; Douglas, N.J. Sleep-disordered breathing and upper airway size in pregnancy and post-partum. Eur. Respir. J. 2006, 27, 321–327. [Google Scholar] [CrossRef]
- Izci, B.; Riha, R.L.; Martin, S.E.; Vennelle, M.; Liston, W.A.; Dundas, K.C.; Calder, A.A.; Douglas, N.J. The Upper Airway in Pregnancy and Pre-Eclampsia. Am. J. Respir. Crit. Care Med. 2003, 167, 137–140. [Google Scholar] [CrossRef]
- Bende, M.; Gredmark, T. Nasal Stuffiness During Pregnancy. Laryngoscope 1999, 109, 1108–1110. [Google Scholar] [CrossRef]
- Williams, P.; Goldspink, G. CHANGES IN SARCOMERE LENGTH AND PHYSIOLOGICAL PROPERTIES IN IMMOBILIZED MUSCLE. 1978, 127, 459–468.
- Johns, E.C.; Denison, F.C.; Reynolds, R.M. Sleep disordered breathing in pregnancy: A review of the pathophysiology of adverse pregnancy outcomes. Acta Physiol. 2020, 229, e13458. [Google Scholar] [CrossRef]
- Bourne, T.; Ogilvy, A.J.; Vickers, R.; Williamson, K. Nocturnal hypoxaemia in late pregnancy. Br. J. Anaesth. 1995, 75, 678–682. [Google Scholar] [CrossRef]
- Guilleminault, C.; Kreutzer, M.; Chang, J.L. Pregnancy, sleep disordered breathing and treatment with nasal continuous positive airway pressure. Sleep Med. 2004, 5, 43–51. [Google Scholar] [CrossRef]
- Garvey, J.F.; Taylor, C.T.; McNicholas, W.T. Cardiovascular disease in obstructive sleep apnoea syndrome: the role of intermittent hypoxia and inflammation. Eur. Respir. J. 2009, 33, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Rath, G.; Aggarwal, R.; Jawanjal, P.; Tripathi, R.; Batra, A. HIF-1 Alpha and Placental Growth Factor in Pregnancies Complicated With Preeclampsia: A Qualitative and Quantitative Analysis. J. Clin. Lab. Anal. 2014, 30, 75–83. [Google Scholar] [CrossRef]
- Lu, D.; Li, N.; Yao, X.; Zhou, L. Potential inflammatory markers in obstructive sleep apnea-hypopnea syndrome. Bosn. J. Basic Med. Sci. 2017, 17, 47–53. [Google Scholar] [CrossRef]
- Lavie, L. Oxidative stress inflammation and endothelial dysfunction in obstructive sleep apnea. Front. Biosci. 2012, E4, 1391–403. [Google Scholar] [CrossRef]
- Prabhakar, N.R.; Kumar, G.K.; Nanduri, J.; Semenza, G.L. ROS Signaling in Systemic and Cellular Responses to Chronic Intermittent Hypoxia. Antioxidants Redox Signal. 2007, 9, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; Mahmoudi, S.; Hattar, K.; Sibelius, U.; Olschewski, H.; Mayer, K.; Seeger, W.; Grimminger, F. Enhanced Release of Superoxide from Polymorphonuclear Neutrophils in Obstructive Sleep Apnea. Am. J. Respir. Crit. Care Med. 2000, 162, 566–570. [Google Scholar] [CrossRef]
- Okun, M.L.; Coussons-Read, M.E. Sleep disruption during pregnancy: How does it influence serum cytokines? J. Reprod. Immunol. 2007, 73, 158–165. [Google Scholar] [CrossRef]
- Alonso-Fernández, A.; Quetglas, C.R.; Mochales, A.H.; De Larrinaga, A. .R.; Barón, A.S.; Rodríguez, P.R.; Gil Gómez, A.V.; Martínez, C.P.; Marín, J.P.C.; Nicolau, M.B.; et al. Influence of Obstructive Sleep Apnea on Systemic Inflammation in Pregnancy. Front. Med. 2021, 8. [Google Scholar] [CrossRef]
- Serednytskyy, O.; Alonso-Fernández, A.; Ribot, C.; Herranz, A.; Álvarez, A.; Sánchez, A.; Rodríguez, P.; Gil, A.V.; Pía, C.; Cubero, J.P.; et al. Systemic inflammation and sympathetic activation in gestational diabetes mellitus with obstructive sleep apnea. BMC Pulm. Med. 2022, 22, 1–11. [Google Scholar] [CrossRef]
- Schulz, R. The vascular micromilieu in obstructive sleep apnoea. Eur. Respir. J. 2005, 25, 780–782. [Google Scholar] [CrossRef]
- Yinon, D.; Lowenstein, L.; Suraya, S.; Beloosesky, R.; Zmora, O.; Malhotra, A.; Pillar, G. Pre-eclampsia is associated with sleep-disordered breathing and endothelial dysfunction. Eur. Respir. J. 2006, 27, 328–333. [Google Scholar] [CrossRef]
- Dharmashankar, K.; Widlansky, M.E. Vascular Endothelial Function and Hypertension: Insights and Directions. Curr. Hypertens. Rep. 2010, 12, 448–455. [Google Scholar] [CrossRef]
- Hadi, H.A.R.; Carr, C.S.; Al Suwaidi, J. Endothelial Dysfunction: Cardiovascular Risk Factors, Therapy, and Outcome. Vasc. Health Risk Manag. 2005, 1, 183–198. [Google Scholar]
- Lui, B.; Burey, L.; Ma, X.; Kjaer, K.; Abramovitz, S.; White, R. Obstructive sleep apnea is associated with adverse maternal outcomes using a United States multistate database cohort, 2007–2014. Int. J. Obstet. Anesthesia 2020, 45, 74–82. [Google Scholar] [CrossRef]
- Sağ. ; Cakmak, B.; Üstünyurt, E. Obstructive sleep apnea syndrome is associated with maternal complications in pregnant women. Ginekol. Polska 2021, 92, 571–574. [Google Scholar] [CrossRef]
- Sun, J.; Lin, C.; Wu, F.; Chung, C.; Sun, C.; Chien, W. The association between obstructive sleep apnea and the risk of poor delivery events in women: A population-based nested case–control study. J. Nurs. Sch. 2021, 54, 31–37. [Google Scholar] [CrossRef]
- Maltepe, E.; Fisher, S.J. Placenta: The Forgotten Organ. Annu. Rev. Cell Dev. Biol. 2015, 31, 523–552. [Google Scholar] [CrossRef]
- Zamudio, S. The Placenta at High Altitude. High Alt. Med. Biol. 2003, 4, 171–191. [Google Scholar] [CrossRef]
- Hutter, D.; Kingdom, J.; Jaeggi, E. Causes and Mechanisms of Intrauterine Hypoxia and Its Impact on the Fetal Cardiovascular System: A Review. Int. J. Pediatr. 2010, 2010, 1–9. [Google Scholar] [CrossRef]
- Sferruzzi-Perri, A.N.; Camm, E.J. The Programming Power of the Placenta. Front. Physiol. 2016, 7, 33–33. [Google Scholar] [CrossRef]
- Kalisch-Smith, J.; Simmons, D.; Dickinson, H.; Moritz, K. Review: Sexual dimorphism in the formation, function and adaptation of the placenta. Placenta 2017, 54, 10–16. [Google Scholar] [CrossRef]
- Caruso, M.; Evangelista, M.; Parolini, O. Human Term Placental Cells: Phenotype, Properties and New Avenues in Regenerative Medicine. 2012, 1, 64–74.
- Goplerud, J.; Delivoriapapadopoulos, M. PHYSIOLOGY OF THE PLACENTA - GAS-EXCHANGE. 1985, 15, 270–278.
- Ravishankar, S.; Bourjeily, G.; Lambert-Messerlian, G.; He, M.; De Paepe, M.E.; Gündoğan, F. Evidence of Placental Hypoxia in Maternal Sleep Disordered Breathing. Pediatr. Dev. Pathol. 2015, 18, 380–386. [Google Scholar] [CrossRef]
- Al-Gubory, K.H.; Fowler, P.A.; Garrel, C. The roles of cellular reactive oxygen species, oxidative stress and antioxidants in pregnancy outcomes. Int. J. Biochem. Cell Biol. 2010, 42, 1634–1650. [Google Scholar] [CrossRef]
- Hung, T.-H.; Skepper, J.N.; Burton, G.J. In Vitro Ischemia-Reperfusion Injury in Term Human Placenta as a Model for Oxidative Stress in Pathological Pregnancies. Am. J. Pathol. 2001, 159, 1031–1043. [Google Scholar] [CrossRef]
- Lavie, L. Obstructive sleep apnoea syndrome – an oxidative stress disorder. Sleep Med. Rev. 2003, 7, 35–51. [Google Scholar] [CrossRef]
- Cuffe, J.S.M.; Walton, S.L.; Singh, R.R.; Spiers, J.G.; Bielefeldt-Ohmann, H.; Wilkinson, L.; Little, M.H.; Moritz, K.M. Mid- to late term hypoxia in the mouse alters placental morphology, glucocorticoid regulatory pathways and nutrient transporters in a sex-specific manner. J. Physiol. 2014, 592, 3127–3141. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.S.; Vaughan, O.R.; de Liger, E.F.; Fowden, A.L.; Sferruzzi-Perri, A.N. Placental phenotype and resource allocation to fetal growth are modified by the timing and degree of hypoxia during mouse pregnancy. J. Physiol. 2015, 594, 1341–1356. [Google Scholar] [CrossRef]
- Rosario, G.X.; Konno, T.; Soares, M.J. Maternal hypoxia activates endovascular trophoblast cell invasion. Dev. Biol. 2007, 314, 362–375. [Google Scholar] [CrossRef]
- Lum, D.H.; Kuwabara, P.E.; Zarkower, D.; Spence, A.M. Direct protein–protein interaction between the intracellular domain of TRA-2 and the transcription factor TRA-1A modulates feminizing activity in C. elegans. Genes Dev. 2000, 14, 3153–3165. [Google Scholar] [CrossRef]
- Badran, M.; Abuyassin, B.; Ayas, N.; Laher, I. Intermittent hypoxia impairs uterine artery function in pregnant mice. J. Physiol. 2019, 597, 2639–2650. [Google Scholar] [CrossRef] [PubMed]
- Arterial Chemoreceptors; Springer Nature: Dordrecht, GX, Netherlands, 2023; ISBN:.
- Genbacev, O.; Zhou, Y.; Ludlow, J.W.; Fisher, S.J. Regulation of Human Placental Development by Oxygen Tension. Science 1997, 277, 1669–1672. [Google Scholar] [CrossRef]
- Jauniaux, E.; Poston, L.; Burton, G.J. Placental-related diseases of pregnancy: involvement of oxidative stress and implications in human evolution. Hum. Reprod. Updat. 2006, 12, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, M.; Herr, F.; Münstedt, K.; Lang, U.; Liang, O.D. Angiogenesis and vasculogenesis in pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 110, S10–S18. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.-P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Ramakrishnan, S.; Anand, V.; Roy, S. Vascular Endothelial Growth Factor Signaling in Hypoxia and Inflammation. J. Neuroimmune Pharmacol. 2014, 9, 142–160. [Google Scholar] [CrossRef]
- Dailey, L.; Ambrosetti, D.; Mansukhani, A.; Basilico, C. Mechanisms underlying differential responses to FGF signaling. Cytokine Growth Factor Rev. 2005, 16, 233–247. [Google Scholar] [CrossRef]
- Wang, K.; Jiang, Y.-Z.; Chen, D.-B.; Zheng, J. Hypoxia Enhances FGF2- and VEGF-Stimulated Human Placental Artery Endothelial Cell Proliferation: Roles of MEK1/2/ERK1/2 and PI3K/AKT1 Pathways☆. Placenta 2009, 30, 1045–1051. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.; Huang, L.; Feng, H.; He, Q.; Lin, X.; Jiang, T.; Lin, J.; Wang, X.; Liu, Q. Gestational chronic intermittent hypoxia induces hypertension, proteinuria, and fetal growth restriction in mice. Sleep Breath. 2021, 26, 1661–1669. [Google Scholar] [CrossRef]
- Song, W.; Chang, W.-L.; Shan, D.; Gu, Y.; Gao, L.; Liang, S.; Guo, H.; Yu, J.; Liu, X. Intermittent Hypoxia Impairs Trophoblast Cell Viability by Triggering the Endoplasmic Reticulum Stress Pathway. Reprod. Sci. 2020, 27, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Giannubilo, S.R.; Cecati, M.; Marzioni, D.; Ciavattini, A. Circulating miRNAs and Preeclampsia: From Implantation to Epigenetics. Int. J. Mol. Sci. 2024, 25, 1418. [Google Scholar] [CrossRef]
- Vanderplow, A.M.; Kermath, B.A.; Bernhardt, C.R.; Gums, K.T.; Seablom, E.N.; Radcliff, A.B.; Ewald, A.C.; Jones, M.V.; Baker, T.L.; Watters, J.J.; et al. A feature of maternal sleep apnea during gestation causes autism-relevant neuronal and behavioral phenotypes in offspring. PLOS Biol. 2022, 20, e3001502. [Google Scholar] [CrossRef]
- Johns, E.C.; Halligan, D.L.; Tammsalu, T.; Hill, E.A.; Riha, R.L.; Denison, F.C.; Reynolds, R.M. Gene expression profiling of placentae from women with obesity and obstructive sleep apnoea. Placenta 2022, 121, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Almendros, I.; Martínez-Ros, P.; Farré, N.; Rubio-Zaragoza, M.; Torres, M.; Gutiérrez-Bautista. J.; Carrillo-Poveda, J.M.; Sopena-Juncosa, J.J.; Gozal, D.; Gonzalez-Bulnes, A.; et al. Placental oxygen transfer reduces hypoxia-reoxygenation swings in fetal blood in a sheep model of gestational sleep apnea. J. Appl. Physiol. 2019, 127, 745–752. [Google Scholar] [CrossRef]
- Almendros, I.; Farré, R.; Planas, A.M.; Torres, M.; Bonsignore, M.R.; Navajas, D.; Montserrat, J.M. Tissue Oxygenation in Brain, Muscle, and Fat in a Rat Model of Sleep Apnea: Differential Effect of Obstructive Apneas and Intermittent Hypoxia. Sleep 2011, 34, 1127–1133. [Google Scholar] [CrossRef]
- Cahill, L.S.; Zhou, Y.-Q.; Seed, M.; Macgowan, C.K.; Sled, J.G. Brain Sparing in Fetal Mice: BOLD MRI and Doppler Ultrasound Show Blood Redistribution During Hypoxia. J. Cereb. Blood Flow Metab. 2014, 34, 1082–1088. [Google Scholar] [CrossRef]
- Carter, AM. Placental gas exchange and the oxygen supply to the fetus. Compr Physiol 2015, 5, 1381–1403. [Google Scholar] [CrossRef] [PubMed]
- Giussani, D.A. The fetal brain sparing response to hypoxia: physiological mechanisms. J. Physiol. 2016, 594, 1215–1230. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, W.; Ciriello, J. Effect of maternal chronic intermittent hypoxia during gestation on offspring growth in the rat. Am. J. Obstet. Gynecol. 2013, 209, 564–e1. [Google Scholar] [CrossRef] [PubMed]
- Cortese, R.; Khalyfa, A.; Bao, R.; Andrade, J.; Gozal, D. Epigenomic profiling in visceral white adipose tissue of offspring of mice exposed to late gestational sleep fragmentation. Int. J. Obes. 2015, 39, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Cortese, R.; Gileles-Hillel, A.; Almendros, I.; Akbarpour, M.; Khalyfa, A.A.; Qiao, Z.; Garcia, T.; Andrade, J.; Gozal, D. Aorta macrophage inflammatory and epigenetic changes in a murine model of obstructive sleep apnea: Potential role of CD36. Sci. Rep. 2017, 7, 43648. [Google Scholar] [CrossRef] [PubMed]
- Koos, BJ. Breathing and sleep states in the fetus and at birth. In: Marcus CL, Carroll JL, Donnelly DF, Loughlin GM, editors. Sleep and breathing in children: developmental changes in breathing during sleep. 2nd ed. New York, NY: Informa Healthcare; 2008. p. 1–17.
- Koos BJ, Rajaee A. Fetal breathing movements and changes at birth. Adv Exp Med Biol. 2014, 814, 89–101. [CrossRef]
- Patrick, J.; Campbell, K.; Carmichael, L.; Natale, R.; Richardson, B. A definition of human fetal apnea and the distribution of fetal apneic intervals during the last ten weeks of pregnancy. Am. J. Obstet. Gynecol. 1980, 136, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Arduini, D.; Rizzo, G.; Giorlandino, C.; Valensise, H.; Dell'Acqua, S.; Romanini, C. The development of fetal behavioural states: A longitudinal study. Prenat. Diagn. 1986, 6, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Patrick, J.; Campbell, K.; Carmichael, L.; Natale, R.; Richardson, B. PATTERNS OF HUMAN-FETAL BREATHING DURING THE LAST 10 WEEKS OF PREGNANCY. 1980, 56, 24–30.
- Koos, B.J.; Matsuda, K.; Power, G.G. Fetal breathing and cardiovascular responses to graded methemoglobinemia in sheep. J. Appl. Physiol. 1990, 69, 136–140. [Google Scholar] [CrossRef]
- Platt, L.D.; Manning, F.A.; Lemay, M.; Sipos, L. Human fetal breathing: Relationship to fetal condition. Am. J. Obstet. Gynecol. 1978, 132, 514–518. [Google Scholar] [CrossRef]
- Tauman, R.; Many, A.; Deutsch, V.; Arvas, S.; Ascher-Landsberg, J.; Greenfeld, M.; Sivan, Y. Maternal snoring during pregnancy is associated with enhanced fetal erythropoiesis – a preliminary study. Sleep Med. 2011, 12, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Templeton A, Kelman GR. Maternal blood-gases, (PAo2-Pao2), physiological shunt and VD/VT in normal pregnancy. Br J Anaesth 1976, 48(10), 1001–4.
- Dawes, GS. Foetal and neonatal physiology; a comparative study of the changes at birth. Year Book Medical Publishers; 1968.
- Longo, L.D. The Rise of Fetal and Neonatal Physiology: Basic Science to Clinical Care, Perspectives in Physiology. Am Physiol Society 2013. [CrossRef]
- Herrington RT, Harned HS Jr, Ferreiro JI, Griffin CA 3rd. The role of the central nervous system in perinatal respiration: studies of chemoregulatory mechanisms in the term lamb. Pediatrics. 1971, 47(5), 857-64.
- Chou, P.J.; Ullrich, J.; Ackerman, B. Time of Onset of Effective Ventilation at Birth. Neonatology 1974, 24, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Cohen, G.; Katz-Salamon, M. Development of chemoreceptor responses in infants. Respir. Physiol. Neurobiol. 2005, 149, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Rigatto, H.; Verduzco, R.D.L.T.; Gates, D.B.; Dumont, F.S.; Kinkead, R.; Edwards, B.A.; Sands, S.A.; Skuza, E.M.; Brodecky, V.; Stockx, E.M.; et al. Effects of O2 on the ventilatory response to CO2 in preterm infants. J. Appl. Physiol. 1975, 39, 896–899. [Google Scholar] [CrossRef]
- Kumar P, Hanson MA. Re-setting of the hypoxic sensitivity of aortic chemoreceptors in the new-born lamb. J Dev Physiol. 1989, 11(4):199-206.
- Richardson, H.L.; Parslow, P.M.; Walker, A.M.; Harding, R.; Horne, R.S.C. Maturation of the initial ventilatory response to hypoxia in sleeping infants. J. Sleep Res. 2007, 16, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fehring, C.; Lowry, T.F.; Wong-Riley, M.T.T.; Stryker, C.; Camperchioli, D.W.; Mayer, C.A.; Alilain, W.J.; Martin, R.J.; MacFarlane, P.M.; et al. Postnatal development of metabolic rate during normoxia and acute hypoxia in rats: implication for a sensitive period. J. Appl. Physiol. 2009, 106, 1212–1222. [Google Scholar] [CrossRef] [PubMed]
- Trachsel, D.; Erb, T.O.; Hammer, J.; von Ungern-Sternberg, B.S. Developmental respiratory physiology. Pediatr. Anesthesia 2021, 32, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Wani, T.M.; Rafiq, M.; Akhter, N.; AlGhamdi, F.S.; Tobias, J.D. Upper airway in infants—a computed tomography-based analysis. Pediatr. Anesthesia 2017, 27, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Praud, J.-P.; Reix, P. Upper airways and neonatal respiration. Respir. Physiol. Neurobiol. 2005, 149, 131–141. [Google Scholar] [CrossRef]
- Ramanathan, R.; Corwin, M.J.; Hunt, C.E.; Lister, G.; Tinsley, L.R.; Baird, T.; Silvestri, J.M.; Crowell, D.H.; Hufford, D.; Martin, R.J.; et al. Cardiorespiratory Events Recorded on Home Monitors. 2001, 285, 2199–2207. [CrossRef]
- Daftary, A.S.; Jalou, H.E.; Shively, L.; Slaven, J.E.; Davis, S.D. Polysomnography Reference Values in Healthy Newborns. J. Clin. Sleep Med. 2019, 15, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Hershenson, M.B.; Colin, A.A.; Wohl, M.E.B.; Stark, A.R. Changes in the Contribution of the Rib Cage to Tidal Breathing during Infancy. Am. Rev. Respir. Dis. 1990, 141, 922–925. [Google Scholar] [CrossRef] [PubMed]
- Rosen, CL. Maturation of breathing during sleep. In: Marcus CL, Carroll JL, Donnelly DF, Loughlin GM, editors. Sleep and breathing in children: developmental changes in breathing during sleep. 2nd ed. New York, NY: Informa Healthcare; 2008. p. 117–30.
- Johnson, S.M.; Randhawa, K.S.; Epstein, J.J.; Gustafson, E.; Hocker, A.D.; Huxtable, A.G.; Baker, T.L.; Watters, J.J. Gestational intermittent hypoxia increases susceptibility to neuroinflammation and alters respiratory motor control in neonatal rats. Respir. Physiol. Neurobiol. 2018, 256, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Gozal, D.; Reeves, S.R.; Row, B.W.; Neville, J.J.; Guo, S.Z.; Lipton, A.J. Respiratory Effects of Gestational Intermittent Hypoxia in the Developing Rat. Am. J. Respir. Crit. Care Med. 2003, 167, 1540–1547. [Google Scholar] [CrossRef] [PubMed]
- Soy, M.; Keser, G.; Atagündüz, P.; Tabak, F.; Atagündüz, I.; Kayhan, S. Cytokine storm in COVID-19: pathogenesis and overview of anti-inflammatory agents used in treatment. Clin. Rheumatol. 2020, 39, 2085–2094. [Google Scholar] [CrossRef] [PubMed]
- Di Fiore, J.M.; Martin, R.J.; Gauda, E.B. Apnea of prematurity – Perfect storm. Respir. Physiol. Neurobiol. 2013, 189, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Raffay, T.M.; Di Fiore, J.M.; Chen, Z.; Sánchez-Illana. ; Vento, M.; Piñeiro-Ramos, J.D.; Kuligowski, J.; Martin, R.J.; Tatsuoka, C.; Minich, N.M.; et al. Hypoxemia events in preterm neonates are associated with urine oxidative biomarkers. Pediatr. Res. 2023, 94, 1444–1450. [Google Scholar] [CrossRef]
- Stoll, B.J.; Hansen, N.I.; Adams-Chapman, I.; Fanaroff, A.A.; Hintz, S.R.; Vohr, B.; Higgins, R.D.; National Institute of Child Health and Human Development Neonatal Research Network. Neurodevelopmental and Growth Impairment Among Extremely Low-Birth-Weight Infants with Neonatal Infection. JAMA 2004, 292, 2357–2365. [Google Scholar] [CrossRef]
- Alonso-Fernández, A.; Quetglas, C.R.; Mochales, A.H.; De Larrinaga, A. .R.; Barón, A.S.; Rodríguez, P.R.; Gil Gómez, A.V.; Martínez, C.P.; Marín, J.P.C.; Nicolau, M.B.; et al. Influence of Obstructive Sleep Apnea on Systemic Inflammation in Pregnancy. Front. Med. 2021, 8. [Google Scholar] [CrossRef]
- Ślusarczyk, J.; Trojan, E.; Głombik, K.; Budziszewska, B.; Kubera, M.; Lasoń, W.; Popiołek-Barczyk, K.; Mika, J.; Wędzony, K.; Basta-Kaim, A. Prenatal stress is a vulnerability factor for altered morphology and biological activity of microglia cells. Front. Cell. Neurosci. 2015, 9, 82–82. [Google Scholar] [CrossRef]
- Fournier, S.; Steele, S.; Julien, C.; Fournier, S.; Gulemetova, R.; Caravagna, C.; Soliz, J.; Bairam, A.; Kinkead, R. Gestational Stress Promotes Pathological Apneas and Sex-Specific Disruption of Respiratory Control Development in Newborn Rat. J. Neurosci. 2013, 33, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Lei, J.; Deng, F.; Zhao, C.; Xu, T.; Ji, B.; Fu, M.; Wang, X.; Sun, M.; Zhang, M.; et al. Gestational Hypoxia Impaired Endothelial Nitric Oxide Synthesis Via miR-155-5p/NADPH Oxidase/Reactive Oxygen Species Axis in Male Offspring Vessels. J. Am. Hear. Assoc. 2024, 13, e032079. [Google Scholar] [CrossRef] [PubMed]
- Morrison, N.R.; Johnson, S.M.; Hocker, A.D.; Kimyon, R.S.; Watters, J.J.; Huxtable, A.G. Time and dose-dependent impairment of neonatal respiratory motor activity after systemic inflammation. Respir. Physiol. Neurobiol. 2020, 272, 103314–103314. [Google Scholar] [CrossRef] [PubMed]
- Master, Z.R.; Porzionato, A.; Kesavan, K.; Mason, A.; Chavez-Valdez, R.; Shirahata, M.; Gauda, E.B. Lipopolysaccharide exposure during the early postnatal period adversely affects the structure and function of the developing rat carotid body. J. Appl. Physiol. 2016, 121, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Beyeler, S.A.; Hodges, M.R.; Huxtable, A.G. Impact of inflammation on developing respiratory control networks: rhythm generation, chemoreception and plasticity. Respir. Physiol. Neurobiol. 2019, 274, 103357–103357. [Google Scholar] [CrossRef] [PubMed]
- McDonald, F.B.; Dempsey, E.M.; O'Halloran, K.D. Effects of Gestational and Postnatal Exposure to Chronic Intermittent Hypoxia on Diaphragm Muscle Contractile Function in the Rat. Front. Physiol. 2016, 7. [Google Scholar] [CrossRef]
- McDonald, F.B.; Dempsey, E.M.; O'Halloran, K.D. Early Life Exposure to Chronic Intermittent Hypoxia Primes Increased Susceptibility to Hypoxia-Induced Weakness in Rat Sternohyoid Muscle during Adulthood. Front. Physiol. 2016, 7, 69. [Google Scholar] [CrossRef]
Figure 1.
Effects of gestational intermittent hypoxia on maternal, fetal and offspring respiratory system. In black, effects in humans; in red, in vitro effects; in blue, effects in animal models.
Figure 1.
Effects of gestational intermittent hypoxia on maternal, fetal and offspring respiratory system. In black, effects in humans; in red, in vitro effects; in blue, effects in animal models.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



