Submitted:
01 May 2024
Posted:
02 May 2024
You are already at the latest version
Abstract
The development and evaluation of techniques that increase the efficiency of selection for planta plant resistance to pathogens is a challenge in all breeding programmes for disease resistance. Under field and laboratory conditions, hybrid populations (F1) of Coffea arabica derived from sources that carry the rust resistance gene SH1 and the CX.2385 line obtained from the Caturra variety × Timor Hybrid CIFC-1343 were evaluated. By means of survival curves, under controlled conditions, the probable times (P<0.05) for the development of symptoms associated with coffee rust were found to be highly variable. Similarly, phenotypic variation in resistance reactions was detected, and plants with incomplete resistance to rust, late development of the incubation period (PI), and no development of a latency period (PL) were identified. The results suggest the use of the detached leaf method as an alternative for the preselection of genotypes for resistance to coffee rust, which could reduce the number of plants used for field evaluations.
Keywords:
survival curves
; Coffea arabica genes
; incubation period
; latency period
; incomplete resistance
; coffee rust
1. Introduction
In C. arabica, the rust resistance genes SH1, SH2 and SH4 were identified through reactions induced to different pathotypes of Hemileia vastatrix [1], and it was proposed that the interactions between these resistance mechanisms and rust coffee plant, adjusted to the gene-for-gene theory, for every resistance gene in the plant there are avirulence genes in the pathogen [2]. In coffee, this relationship is often applied to resistance factors in the plant and virulence in the pathogen in the homozygous state, as long as the resistance comes from dominant genes and the virulence from recessive genes [3]. However, resistance genes in C. arabica and virulence genes in H. vastatrix are not completely dominant [4].
Currently, the protection conferred by the genes encoding C. arabica SH1, SH2, and SH4 is of little interest due to the rapidity with which variants of H. vastatrix can overcome it. However, plants harboring these genes may exhibit incomplete resistance when specific resistance is overcome by the pathogen [5,6]. In this way, incomplete resistance can become effective in the field for a longer time, providing protection to plants against pathogens that cause limiting diseases, compared to complete resistance, and its quantification can only be carried out through the study of races of the pathogen compatible with the host [7]. On the other hand, incomplete resistance is affected by various factors, including the environment [8], nutritional status [9,10,11], age, phenology, ontogenetic state of organs [8,12,13], environment [8], and production potential of varieties [14,15].
In C. arabica, incomplete resistance to H. vastatrix is affected by the age of the leaves [16], the productivity of the plant [17], the interaction between the genotype and the environment (GxA) [18,19,20,21,22], the physiological and nutritional status of the crop [9,10,11,23], and its genetic basis [24]. Studies on this last factor have shown that incomplete resistance in C. arabica is related to interactions between genes with greater effects [24] and genes with weaker effects that act in an additive manner [25]. The constitutive genetic background of the plant can have an effect, as can a homozygous state of the gene and its combination with other resistance mechanisms [24]. These mechanisms are probably similar to those of the genes with a greater effect, SH6 through SH9, identified in the Timor Hybrid (HdT) [3], and their control is oligogenic but is determined by a variable number of genes [25].
It has been estimated that the heritability of incomplete resistance is between 31% and 73% [4,25], and from segregating populations, varieties with resistance factors that are inherited separately or in different combinations can be obtained. Genetically diverse offspring are produced [26]. This has allowed the successful genetic improvement and development of coffee varieties with incomplete resistance to rust.
On the other hand, the development of varieties with durable genetic resistance to rust should be complemented with evaluations under natural conditions of the crop and under full pressure from the pathogen. The latter is the ideal condition for identifying and determining the response of the improved genotypes against the pathotypes of H. vastatrix that prevail in the natural conditions of C. arabica culture. In the case of Colombia, this evaluation process is recurrent and requires long periods of time and high economic resources, which can make up around 30% of the costs per cycle in the research processes in the early stages of the development of an improved coffee variety.
In the search for shorter times, fewer required resources, and more efficient selection processes for mechanisms in which there are no developed molecular markers, alternative methods for evaluating resistance to diseases, which require short evaluation times and replicate the infection reactions caused by the pathogen under field conditions used for different crops, which have been adjusted for coffee [5,27]. The use of such alternative methods makes it possible to measure the components of incomplete resistance [28]. In coffee, these components quantified under laboratory conditions present a high correlation (r = 0.83) with the severity of disease observed under natural conditions of the crop, which has allowed the selection of coffee genotypes with incomplete resistance to rust. [29]. However, this process involves the collection of continuous records, which are essential for determining the times at which one or more specific symptoms occur. This large volume of information is highly useful when investigating the behavior of plants against the attack of pathogens and disease development [30,31].
The information obtained in this type of research, when adjusted to statistical assumptions, is generally obtained from parametric statistics. When this does not occur, it is necessary to resort to other statistical approaches for analysis, including time to occurrence analysis. This method is also known as survival analysis or reliability analysis [32,33]. This type of analysis has been implemented to study the interactions between different species, including corn (Zea mays), spiroplasm (CSS), and corn and mycoplasma (MBSM) [34]; potato (Solanum tuberosum) and Clavibacter michiganensis [35]; cedar (Chamaecyparis lawsoniana) and Phytophthora lateralis [36]; soybean (Glycine max and G. soja) and Pythium aphanidermatum [37]; and chickpea (Cicer arietinum L.) and Phytophthora medicaginis [31].
Countless cases of plant‒pathogen interactions can be cited, but the times until the occurrence of events are rarely tracked [30,33,38], and coffee is no exception. Due to the importance in genetic improvement of developing techniques for the evaluation and selection of rust resistant genotypes and the ability to establish a balance between the time required for the analysis and the efficiency of the selection, the present investigation aimed to select genotypes resistant to coffee rust using the detached leaf inoculation method. This research was carried out in a hybrid population combining resistance mechanisms from the HdT CIFC-1343 and the SH1 gene present in wild genotypes of C. arabica from the Colombian Coffee Collection (CCC). The data were analyzed using nonparametric statistics, and the probability of occurrence of symptoms associated with the components of incomplete resistance was determined.
2. Materials and Methods
Twenty plants from three populations derived from complex crosses between Ethiopian introductions, carriers of the SH1 gene (CCC.32 and CCC.66), and varieties Caturra and Catuaí, which are susceptible to rust, were evaluated: Population 1: [[(Caturra × CCC.32) × (Caturra × CCC.66)] × CX.2385]; Population 2: [CX.2385 × [(Caturra × CCC.32) × (Caturra × CCC.66)]] and Population 3. [Catuaí × [(Caturra × CCC.66)] × CX.2385]. In previous works developed at the Centro de Investigação das Ferrugens do Cafeeiro (CIFC for its name in Portuguese), it was determined that these crosses segregate for the physiological resistance Groups C and EC (SH1.5, SH5), respectively. These populations were crossed with the CX.2385 line, a carrier of incomplete resistance to the disease, which was obtained from a cross between the Caturra variety and the HdT CIFC-1343. The resulting plants were established at the Naranjal Experimental Station of the National Coffee Research Center, Cenicafé, located in the municipality of Chinchiná, Department of Caldas, Colombia (04° 59′N, 75° 39′W, 1381 masl). This center has an average temperature of 21.4 °C, 2782 mm of annual precipitation, and 77.5% relative humidity.
First, the incidence of rust was quantified under field conditions using the scale of incidence in the field [39]. This scale takes the entire plant as the unit of observation. Between 2017 and 2019, two evaluations were carried out per year in the months with the highest incidence of the disease in Colombia (April and August). All the plants were evaluated for their incomplete resistance to rust using the detached leaf inoculation method [5,29]. Fully extended young leaves were taken from branches of the second third of the tree, located in the first two nodes, from the outside to the inside. The leaves were collected with the petiole, washed, and disinfected by immersion in a 3% hypochlorite solution under constant agitation for 30 to 40 seconds. Immediately afterward, they were washed with distilled water and then immersed in alcohol at a concentration of 70% for 1 minute. The alcohol was removed via rinses with distilled water, followed by rinses with sterile distilled water. Four leaves were deposited per genotype, arranged with the underside facing up, in transparent plastic boxes with a lid (37 cm wide by 26 cm high by 40 cm long) acting as a humid chamber. The leaves were then kept for 12 hours in complete darkness.
In parallel, the inoculum was obtained from the CX.2385 line and was prepared in solution with sterile distilled water, and 0.7 mg of urediniospores was added to each ml of water. The dispersion of the urediniospores and homogenization of the solution were carried out by shaking under ultrasound for 20 seconds. Finally, the solution was subjected to constant magnetic stirring, and with a micropipette, 8 drops of 5 µl were deposited on each leaf and then incubated in complete darkness for 48 hours. The inoculated leaves were subjected to alternating periods of illumination, 9 hours of light and 15 hours of darkness, for 60 days with LED light lamps set up 75 cm above the leaves. It was guaranteed that the interior humidity of each chamber stayed close to the saturation point, the temperature was between 22 and 24 °C, and from day 10 after inoculation (DAI) on, the progression of the disease was recorded (Figure 1) on the scale of increasing lesions for the evaluation of incomplete resistance [5].
The experiment was replicated four times in a completely randomized design. The data were analyzed by survival analysis, taking the leaf as an experimental unit and each inoculated site as an observation unit. Survival estimators until the appearance of each symptom were obtained by the Kaplan–Meier method [32] with 95% confidence intervals (P<0.05) using Equation (1):
where:
S (t) = P (T ≥ t)
- -
- S(t) = the survival function.
- -
- T = the most likely time at which symptoms develop in each genotype.
- -
- P(t) = the conditional probability function that describes the instantaneous risk for the symptom to develop at time t, from day 10 DAI to day 60 DAI.
3. Results
3.1. Incidence of Rust under Field Conditions
Six evaluations were made under field conditions, with maximum values of 3 and minimum values of 1 according to the incidence scale of Eskes & Toma-Braghini [39] (Table 1). Although grade 3 can be considered a degree of apparent susceptibility, it was classified as a resistance reaction due to the non-effect of the disease on production.
3.2. Probable Times of Symptom Onset under Controlled Conditions
The average DAI at the appearance of small, discoloured lesions (grade 1) and the percentage of inoculated sites that developed grade 1 lesions was evaluated. Variation between genotypes was seen (Figure 2, Table 2). The Caturra variety and the CX.2385 line exhibited the shortest incubation periods, with 14.34 and 14.94 DAI, respectively.
In all the plants evaluated, some tissue was affected by the fungus. The survival estimators (P<0.05) showed that the most likely times for the development of the fungus during the incubation period were not absolute but rather were highly variable (Figure 2). The incubation period was 14.34 DAI in the Caturra variety, 14.94 DAI in the CX.2385 line, and 22.50 DAI in the plant 128 (Population 3).
The symptom values recorded in the incubation period were variable, ranging from 32.8% to 100%. The lowest percentage (32.8%) was presented by plant 128, which did not develop more advanced symptoms. In the other plants, symptoms were detected in 87.5% to 100% of the lesions (Table 2). Lesions of genotypes 33, 34, 35, 51, and 53 and Caturra variety and CX.2385 line showed sporulation, with visible differences in density, indicating different levels of resistance for each plant (Table 2). In all cases, the disease developed gradually, with intense chlorotic zones, followed by the formation of uredosporic sori.
The times to reach the most advanced stages of the disease within the incubation period varied from plant to plant. The earliest expression of grade 2 was observed in the Caturra variety (18.2 DAI), the latest in plant 32 (36.4 DAI). CX.2385 line developed symptoms 4.3 days later than the Caturra variety and 13.9 days earlier than the most recent genotype (plant 32) (Figure 2).
The maximum grade reached by plant 65 was 2, which was visible in 93.8% of the lesions (Table 2) at 26 DAI. This time was 8 days later than that of the Caturra variety and 4.2 days later than that of CX.2385 line. In the other plants, symptoms arose between 19.0 and 23.0 DAI. Plants 35 and 51 developed symptoms at the same time (Figure 2).
In variety Caturra, grade 3 was observed 3.6 days after presenting grade 2. The progression of the disease was slower in plant 32, at 17.1 days after CX.2385 line and 22.2 days after the Caturra variety (Figure 2). Plants 31 and 127 presented equal times for the development of grade 3 symptoms. Grade 3 was present to different degrees, with values ranging from 6.3% to 100% of affectation (Table 2).
The shortest latency periods (grade 4) were recorded for the CX.2385 line (34.5 DAI) and the Caturra variety (36 DAI). This symptom developed in only 25% of the evaluated plants (33, 34, 35, 51, and 53) (Figure 2). The highest percentages of sporulating sites were observed in plants 51 and 53 (Table 2). Plant 34 was the latest to develop sporulating sites, 24 days later than the Caturra variety did and 25.5 days after the CX.2385 line did.
The stages of disease development greater than the latency period (grades 5, 6, and 7) were detected only in plants 33, 35, 51, and 53 and in the Caturra variety and CX.2385 line. Grade 6 was detected in CX.2385 line at 37.4 DAI, affecting 24% of the inoculated sites, and at 42.7 DAI in the Caturra variety, affecting 78.1%. With a difference of a few days, the same degree was recorded in plants 35, 51, and 53, which presented widely dissimilar infection values, ranging from 0.8% to 40.6% of affectation (Table 2).
Grade 7 was observed in the Caturra variety and CX.2385 line. Genotypes 35 and 51 developed the same symptoms at 48 and 44 DAI, respectively. The lesions appeared in variable percentages, between 5% and 30% (Table 2). The percentage of affectation of genotype 35 was 18.6% less than that of CX.2385 line and 71.8% less than that of the Caturra variety. The effect of plant 51 was 5.5% greater than that of CX.2385 line but 47.6% lower than that of the Caturra variety. The percentages of grade 7 lesions in the Caturra variety and CX.2385 line were widely dissimilar, with values greater than 75% in the Caturra variety and less than 25% in the CX.2385 line.
3.3. Probability of Survival to the Expression of Symptoms
Population 1, made up of plants 31, 32, 33, 34, and 35, presented heterogeneity in resistance reactions from day 10 to day 60 DAI (Figure 3). Under the exposure conditions, evaluation time, and inoculum used, the development of grade 4 fungi was not present in plants 31 and 32. In plants 31, 32, and 35, the development of the different symptoms did not occur quickly. In contrast, in plants 33 and 34, the symptoms were most common during the first 35 days of observation.
In population 2, there was also variation in the reactions of resistance to rust. Plants 52, 54, 55, 61, 62, 63, and 64 showed no development of a latency period (grade 4), similar to what we observed in plants 31 and 32 of population 1. Plants 51 and 53 had the highest incidence, which was similar to that of CX.2385 line. Plant 65, which did not develop to a degree higher than 2, stood out (Figure 4).
Population 3 presented resistance similar to that described for population 2 (Figure 5). Plants that did not develop grade 3 resistance symptoms were found. Plant 128 was of great interest and stood out among all the genotypes analyzed. This plant was characterized by late development of the infection period and a delayed symptom effect. Symptoms were occasionally observed at all the inoculated sites (Figure 5), reaching only grade 1, and no more advanced symptoms developed.
No plants of the Caturra variety or CX.2385 line survived to the development of the maximum degree of disease. In these genotypes, the disease developed and presented all its stages of development. The short durations (compared with those of the other genotypes) were the most likely, indicating a greater risk of disease development in the first days after contact between the inoculum and the host. The genotypes expressed all the symptoms of the disease in less than 60 days (Figure 6).
The controlled conditions in which the research was conducted guaranteed that all the evaluated genotypes presented an equal opportunity to be infected by the fungus. However, as the observation time increased, the probability that the disease did not develop on each genotype decreased. In the case of plants 65 and 128, a low probability of not developing during the infection period was observed (P<0.05), without symptoms developing beyond grade 2, an effect attributed to the genetic resistance that the plant presented the inoculum used. Likewise, the formation of chlorotic halos of variable size was observed, and in some cases swollen tissues were observed.
Regarding the controls, in the Caturra variety and CX.2385 line, the fungus developed spores, reaching the maximum degree according to the evaluation scale. The greatest effect was observed in the Caturra variety (Table 1).
4. Discussion
When the objective of a breeding program is to find resistant plants to limiting and potential diseases, it is necessary to evaluate large populations such that plants with favorable resistance alleles are likely to emerge. The size of the population will be a function of the number of loci that the breeders want to introgress into the same genotype, and the variation in the expression of the resistance components will be the result of the level of homozygosity of the resistance genes in the parents. In F1 plants, the absence of segregating patterns is expected when the resistance mechanisms are in a homozygous state. In contrast, the results obtained in this research suggest that rust resistance segregates within the evaluated populations, a behavior that has been reported in hybrid populations developed from sources of C. arabica [4]. In the plants evaluated in this work, an apparent segregation by resistance was observed, expressed by longer periods to develop a latency period, lower density of sporulating lesions, and, in some cases, early necrosis of the lesions, which is consistent with other reports [4].
In C. arabica, the percentages of urediniospores that germinate and infect tissues are similar in resistant and susceptible plants [40]. However, in genotypes considered resistant, the plant responses occur a few days after the fungus begins the infection process. These reactions in the plant against the attack of pathogens are modulated by the expression of proteins that regulate defense responses, blockages in the formation of the appressorium, mycelial growth and modifications of the cell wall, affecting the synthesis of lignin in the cells that surround stomatal tissue and sometimes cytoplasmic tissue [41,42,43,44].
All this leads to the development of chlorosis and swelling of the tissues and a reduction in the penetration of the fungus, making it difficult to establish, preventing colonization, and reducing the reproduction of the pathogen on the tissue [40,45]. These resistance reactions generally occur because the defense mechanisms that plants deploy to attack pathogens involve quantitative and polygenic inheritance [44].
In this study, no plants immune to the pathogen were identified, that is, all of them presented some degree of affectation. However, highly resistant plants with variable phenotypic responses, including the development of chlorotic zones of different intensities and swelling, were identified (plants 65 and 128). These symptoms are frequently observed in the Rubiaceae family, where tissue deformation occurs at the site of infection, accompanied by chlorotic regions, and these symptoms are considered reactions that occur in incomplete resistance responses of coffee to H. vastatrix [46]. In resistant genotypes of Coffea, during infection by H. vastatrix, the cells of the mesophyll develop hypertrophy as an effect of the activation of secondary metabolites, accumulation of phenolic components, and thickening of the cell wall [40]. These responses are the product of the activation of the metabolic pathway of shikimic acid, which is the same pathway that yields the biochemical compounds that lignin is made from. Lignin is responsible for the thickening of cell walls, the first barrier of plants in defense against the attack of phytopathogens.
These variations in resistance, to a large extent, are related to different factors, including environmental factors [9,10,11,16], genetic factors [8,24], and the product of interactions between them [18,19,20,21,22]. Therefore, to decrease the effects of interactions between genotype and the environment, this work was carried out under controlled conditions.
When any of the parents are not homozygous, segregating hybrid populations originate, in which case resistance and susceptibility are not absolute attributes of the genotype. In contrast, they are characterized by compatible or incompatible specific interactions between the host, its resistance mechanisms, the pathogen, and its virulence mechanisms [47]. In the case of resistance conferred by genes from C. arabica SH1, SH2, and SH4, its resistance capacity is regulated by its level of homozygosity and interaction with other resistance genes present in the plant [3,48].
The resistance reactions we observed show that the combination of the SH1 gene with the resistance genes derived from HdT can have a genetic control effect on the pathotype used, allowing the expression of various phenotypic resistance reactions and the apparent additive action of genes. Therefore, it is possible to phenotypically select resistant plants, facilitating the combination of resistance alleles in a genotype and providing an efficient barrier against the pathogen [48]. Additionally, in the HdT and its derived varieties, resistance is controlled by genes with independent segregation [49], allowing the genetic configurations of the progeny to be diverse in their reactions to disease [8].
In the three populations evaluated, 65% of the plants presented intermediate reactions in terms of intensity (grade 3), with a progressive increase in chlorotic tissue. This type of resistance reaction is most common in plants when they are affected by rust causing fungi [45]. In plants classified as susceptible, the resistance response is delayed, allowing the development, growth, and sporulation of fungus [40], consistent with what was observed within the plants of the evaluated populations. In this research, late phenotypic defense responses were observed, but these responses allow the pathogen to affect tissues in different proportions. In this sense, the level of resistance of the evaluated plants and the infectivity rates of the pathogen were decisive in each genotype to restrict the development of the fungus, and this plant‒pathogen interaction caused variations in each genotype’s risk per se of developing symptoms associated with the disease.
The survival curves allowed us to identify the probability and model the level of resistance of the plant to the development of H. vastatrix. What was observed was adjusted to what was expected in this type of experiment [31,32,35,36,37]. The phenotypic symptoms observed in plants 65 and 128 are often preceded by the cessation of growth and colonization of the pathogen [3,40], making this characteristic an indicator of its high genetic value and agronomic interest. We must bear in mind that when breeding and selection are carried out to seek genetic resistance against pathogens that limits crop production, yield reductions may occur [14,15]. Additionally, the relationship between genetic resistance to a pathogen and the production capacity of the plant will always be closely related to the effects of the environment and the genetic configuration of the varieties [8].
All plants presented incomplete resistance under field conditions, but plants 65 and 128 presented the lowest incidence of the disease under conditions of pressure from virulent races (Table 1). Therefore, strict selection was done based on the incidence values under controlled conditions and was complemented with the values obtained under field conditions. This last parameter (field evaluations) indisputably reflects the level of genetic resistance of the plant under conditions of high pressure from the inoculum to specific races.
In the selection process within any genetic improvement program for resistance to diseases, the selection parameters must always be as high as possible to reduce the future risk that the pathogen will pass through introgressed resistance mechanisms, which is a natural and inevitable process. Given this risk, the ideal strategy to mitigate the impacts of a possible loss of resistance is to make use of genetic diversity. Its uses and benefits are documented for different crops [50,51,52], coffee is no exception. Its genetic diversity has yielded a wide spectrum of benefits, including economic, environmental, and social benefits, as has been demonstrated with the varieties released by the National Federation of Coffee Growers and its research center Cenicafé in Colombia [53].
Although CX.2385 line exhibited a high disease incidence in the laboratory, with a grade of 7 on the scale of growing lesions and a grade of 6 under field conditions [5,12], the severity of these lesions under cultivation conditions does not seem to have negative effects on production, confirming its high incomplete resistance to rust. Contrasting findings were observed for the Caturra variety, which was highly susceptible to normal cultivation conditions in Colombia and whose production was harmed at any time of the year, at any altitude, and in the absence of chemical control for the disease. Therefore, from the identified and selected plants, we will continue to obtain segregating populations and to advance them to further generations to fix and select progenies for the various characteristics of agronomic value, which will benefit Colombian coffee growers.
In this research, the agronomic management of the plants and the research conditions, temperature, humidity, evaluation times, and light intensity were the same for all the genotypes. This ruled out environmental effects or effects attributed to the physiological state of the plant [9,16]; therefore, the variations in the resistance response observed are the expression of resistance characteristics of the genotype and are undoubtedly related to the genetic background of the parents and the combinations of resistance factors in the progeny.
5. Conclusions
This research identified hybrid plants (F1) that are highly resistant to coffee rust and that have become new sources of genetic resistance to the disease for the development of new coffee varieties in Colombia. New evidence is presented for the phenotypic response of the combination of resistance factors from C. arabica and HdT, which have different resistance spectra that are expressed when the genes are not combined or are acting in a separate way. Our results lead us to recommend the implementation of a plant preselection method under controlled conditions to identify the genetic potential for resistance to H. vastatrix early. In this way, fewer plants might be needed for field evaluation, which is crucial for the efficient use of economic resources in the selection and development of varieties.
Author Contributions
Conceptualization, JQC and CPFR, investigation, JQC, LFLM and writing — original draft preparation JQC, writing — review and editing, JQC, CPFR, and VMPV All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Coffee Research Center (Cenicafé) (Crossref Funder ID 100019597).
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank the university interns who were linked to the Genetic Improvement Program of the Coffee Research Center - Cenicafé during the research and who supported the assembly and registration of the information.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Noronha Wagner, M., & Bettencourt, A. J. (1967). Genetic study of the resistance of Coffea spp. to leaf rust - Identification and behavior of four factors conditioning disease reaction in Coffea arabica to twelve physiologic races of Hemileia vastatrix. Canadian Journal of Botany, 45(11), 2021-2031. [CrossRef]
- Flor, H. H. (1942). Inheritance of pathogenicity in Melampsora lini. Phytopathology, 32(8), 653-669.
- do Céu Silva, M., Rijo, L., Rodrigues Jr, C. J., & Vasconselos, M. I. (1992). Histological study of the heterozigosity effect of coffee resistance genes SH1 and SH4, SH5 towards Hemileia vastatrix. Broteria Genética, XIII(LXXX), 169-184.
- Eskes, A. B., & Carvalho, A. (1983). Variation for incomplete resistance to Hemileia vastatrix in Coffea arabica. Euphytica, 32, 625–637. [CrossRef]
- Leguizamón Caicedo, J. (1985). Contribution a la connaissnace de la resistance incomplete du cafier arabica (Coffea arabica L.) a la rouille orangee (Hemileia vastatrix Berk. et Br.). Montpellier: IRCC.
- Castillo Zapata, J., & Leguizamón Caicedo, J. (1992). Viruelencia de Hemileia vastatrix determinada por medio de plantas diferenciales de café en Colombia. Cenicafé, 43(4), 114-124.
- Rios, J. A., & Debona, D. (2018). Efeito epidemiológico da resistência de hospedeiro. En J. L. Dallagnol, Resistência genética de plantas a patógenos (pág. 437). Pelotas, RS - Brasil: UFPel.
- Ficke, A., Cowger, C., Bergstrom, G., & Brodal, G. (2018). Understanding Yield Loss and Pathogen Biology to Improve Disease Management: Septoria Nodorum Blotch - A Case Study in Wheat. Plant Diseases, 102(4), 696-707. [CrossRef]
- Toniutti, L., Breitler, J. C., Etienne, H., Campa, C., Doulbeau, S., Urban, et al., (2017). Influence of Environmental Conditions and Genetic Background of Arabica Coffee (C. arabica L) on Leaf Rust (Hemileia vastatrix) Pathogenesis. Frontiers in Plant Science, 8, 2025. [CrossRef]
- Pérez, C. P., Pozza, E. A., Pozza, A. A., de Freitas, A. S., Silva, M. G., & Gomes Guimarães, D. d. (2019). Impact of nitrogen and potassium on coffee rust. European Journal of Plant Pathology, 155, 219–229. [CrossRef]
- Ghufron Rosyady, M., Anom Wijaya, K., Wulanjari, D., & Wafa, A. (2020). Role of Mineral Elements to Induce the Resistance of Arabica Coffee Against Rust Disease at Lowland Area. E3S Web of Conferences, 142(03003), 03. [CrossRef]
- Eskes, A. B., & Toma-Braghini, M. (1982). The effect of leaf age on incomplete resistance of coffee to Hemileia vastatrix. Netherlands Journal of Plant Pathology, 88, 219–230. [CrossRef]
- Marla, S. R., Chu, K., Chintamanani, S., Multani, D. S., Klempien, A., De Leon., et al., (2018). Adult plant resistance in maize to northern leaf spot is a feature of partial loss-of-function alleles of Hm1. PLoS Pathogens, 14(10), e1007356. [CrossRef]
- Ning, Y., Liu, W., & Wang, G. L. (2017). Balancing Immunity and Yield in Crop Plants. Trends in Plant Science, 22(12), 1069-1079. [CrossRef]
- Li, W., Chern, M., Yin, J., Wang, J., & Chen, X. (2019). Recent advances in broad-spectrum resistance to the rice blast disease. Current Opinion in Plant Biology, 50, 114-120. [CrossRef]
- Countinho, T. A., Rijkenberg, F. J., & van Asch, M. (1994). The effect of leaf age on infection of Coffea genotypes by Hemileia vastatrix. Plant Pathology, 43(1), 97-103. [CrossRef]
- Alvarado Alvarado, G. (2011). El café y la roya: Estrategias de resistencia incompleta. Chinchiná: FNC - Cenicafé.
- DaMatta, F. M. (2004). Ecophysiological constraints on the production of shaded and unshaded coffee: a review. Field Crops Research, 86(2-3), 99-114. [CrossRef]
- Romero Guerrero, G., Herrera Pinilla, J. C., Ligarreto Moreno, G. A., & Alvarado Alvarado, G. (2008). Análisis genético de la resistencia incompleta a Hemileia vastatrix en progenies de Caturra x Híbrido de Timor. Cenicafé, 59(2), 103-119.
- Liebig, T., Ribeyre, F., Läderach, P., Poehling, H.-M., Asten, P. v., & Avelino, J. (2019). Interactive effects of altitude, microclimate and shading system on coffee leaf rust. Journal of Plant Interactions, 14(1), 407-415. [CrossRef]
- Belachew, K., Senbeta, G. A., Garedew, W., Barreto, R. W., & Del Ponte, E. M. (2020). Altitude is the main driver of coffee leaf rust epidemics: a large-scale survey in Ethiopia. Tropical Plant Pathology, 45, 511–521. [CrossRef]
- Merle, I., Pico, J., Granados, E., Boudrot, A., Tixier, P., Virginio Filho., et al., (2020). Unraveling the Complexity of Coffee Leaf Rust Behavior and Development in Different Coffea arabica Agroecosystems. Phytopathology, 110, 418-427. [CrossRef]
- Vasco, G. B., Pozza, E. A., Silva, M. G., Pozza, A. A., & Chaves, E. (2018). Interaction of K and B in the intensity of coffee rust in nutrient solution. Coffee Science, 13(2), 238-244. [CrossRef]
- Eskes, A. B., & Da Costa, W. M. (1983). Characterization of incomplete resistance to Hemileia vastatrix in the icatu coffee population. Euphytica, 32, 649–657. [CrossRef]
- Romero Guerrero, G., Alvarado Alvarado, G., Cortina Guerrero, H., Ligarreto Moreno, G., Galeano, N. F., & Herrera Pinilla, J. C. (2010). Partial resistance to leaf rust (Hemileia vastatrix) in coffee (Coffea arabica L.): genetic analysis and molecular characterization of putative candidate genes. Molecular Breeding, 25, 685–697. [CrossRef]
- Castillo Zapata, J., Moreno Ruíz, G., & López Duque, S. (1976). Uso de resistencia genética a Hemileia vastatrix Berk. y Br. existente en germoplasma de café en Colombia. Cenicafé, 27(1), 3-25.
- Karki, H. S., & Halterman, D. A. (2021). Phytophthora infestans (Late blight) Infection Assay in a Detached Leaf of Potato. 11(4), e3926. [CrossRef]
- Parlevliet, J. E. (1979). Components of Resistance that Reduce the Rate of Epidemic Development. Annual Review of Phytopathology (17), 203-222. [CrossRef]
- Ferreira de Mendonça, R., Cintra de Jesus Junior, W., Gava Ferrão, M., Bucker Moraes, W., Magno Busato, L., Gava Ferrão, R., et al., (2019). Genótipos de café conilon e sua reação à ferrugem alaranjada. Summa Phytopathologica, 45(3), 279-284. [CrossRef]
- Scherm, H., & Ojiambo, P. S. (2004). Applications of Survival Analysis in Botanical Epidemiology. Phytopathology, 94(9), 1022-1026. [CrossRef]
- Amalraj, A., Taylor, J., & Sutton, T. (2019). A hydroponics based high throughput screening system for Phytophthora root rot resistance in chickpea (Cicer arietinum L.). Plant Methods, 15, 82. [CrossRef]
- Kaplan, E. L., & Meier, P. (1958). Nonparametric Estimation from Incomplete Observation. Journal of the American Statistical Association, 53(282), 45-481. [CrossRef]
- Romano, A., & Stevanato, P. (2020). Germination Data Analysis by Time-to-Event Approaches. Plants, 9(5), 617. [CrossRef]
- Madden, L. V., & Nault, L. R. (1983). Differential Pathogenicity of Corn Stunting Mollicutes to Leafhopper Vectors in Dalbulus and Baldulus species. Phytopathology, 73, 1608-1614. [CrossRef]
- Westra G, A. A., Arneson, C. P., & Slack, S. A. (1994). Effect of Interaction of Inoculum Dose, Cultivar, and Geographic Location on the Development of Foliar Symptoms of Bacterial Ring of Potato. Ecology and Epidemiology, 84(4), 410-415.
- Jules, E. S., Kauffman, M. J., Ritts, W. D., & Carroll, A. L. (2002). Spread of an Invasive Pathogen over a Variable Landscape: A Nonnative Root Rot on Port Orford Cedar. Ecology, 83(11), 3167-3181. [CrossRef]
- Neher, D. A., Wilkinson, H. T., & Augspurger, C. K. (1992). Progression of damping-off epidemics in Glycine populations of even-age and mixed-age structure. Canadian Journal of Botany, 70(5), 1032-1038. [CrossRef]
- Stalpers, L. J., & Kaplan, E. L. (2018). Edward L. Kaplan and the Kaplan-Meier Survival Curve. Journal of the British Society for the History of Mathematics, 33(2), 109-135. [CrossRef]
- Eskes, A. B., & Toma Braghini, M. (1981). Assessment methods for resistance to coffee leaf rust (Hemileia vastatrix Berk. and Br.). FAO-Plant-Protection-Bulletin, 29(3/4), 56-66.
- do Céu Silva, M., Nicole, M., Guerra Guimaraes, L., & Rodrigues Jr., C. J. (2002). Hypersensitive cell death and post-haustorial defence responses arrest the orange rust (Hemileia vastatrix) growth in resistant coffee leaves. Physiological and Molecular Plant Pathology, 60(4), 169-183. [CrossRef]
- Andersen, E. J., Ali, S., Byamukama, E., Yen, Y., & Nepal, M. P. (2018). Disease Resistance Mechanisms in Plants. Genes, 9(1), 339. [CrossRef]
- Albert, M., Axtell, M. J., & Timko, M. P. (2021). Mechanisms of resistance and virulence in parasitic plant–host interactions. Plant Physiology, 185(4), 1282–1291. [CrossRef]
- do Céu Silva, M., Guerra-Guimarães, L., Diniz, I., Loureiro, A., Gil Azinheira, H., Carmo Pereira, A. P., et al., (2022). An Overview of the Mechanisms Involved in Coffee-Hemileia vastatrix Interactions: Plant and Pathogen Perspectives. Agronomy, 12(2), 326. [CrossRef]
- Simardeep, K., Kumar Samota, M., Choudhary, M., Choudhary, M., Pandey, A. K., Sharma, A., & Thakur, J. (2022). How do plants defend themselves against pathogens-Biochemical mechanisms and genetic interventions. Physiology and Molecular Biology of Plants, 28, 485–504. [CrossRef]
- Gill, U. S., Lee, S., & Mysore, K. S. (2015). Host Versus Nonhost Resistance: Distinct Wars with Similar Arsenals. Phytopathology, 105(5), 580-587. [CrossRef]
- Eskes, A. B. (1989). Resistance. En A. C. Kushalappa, & A. B. Eskes, Coffee Rust: Epidemiology, Resistance and Management (pág. 345). Boca Ratón, Florida: CRC Press. Inc.
- Zambolim, L., & Teixeira Caixeta, E. (2021). An Overview Of Physiological Specialization Of Coffee Leaf Rust – New Designation Of Pathotypes. International Journal of Current Research, 13(1), 15564 - 15575.
- Eskes, A. B. (1982). The effect of light intensity on incomplete resistance of coffee to Hemileia vastatrix. Netherlands Journal of Plant Pathology, 88, 191–202. [CrossRef]
- Capucho, A. S., Caixeta, E. T., Zambolim, E. M., & Zambolim, L. (2009). Herança da resistência do Híbrido de Timor UFV 443-03 à ferrugem-do-cafeeiro. Pesquisa Agropecuária Brasileira, 44(3), 276-282. [CrossRef]
- Parvatha Reddy, P. (2013). Variety Mixtures/Cultivar Mixtures/Multilines. En P. Parvatha Reddy, Recent advances in crop protection (págs. 201-221). India: Springer. [CrossRef]
- Chai, Y., Pardey, P. G., & Silverstein, K. T. (2022). Scientific selection: A century of increasing crop varietal diversity in US wheat. PNAS, 119(51), e2210773119. [CrossRef]
- Goulart Castro, D., Mendes de Moura, A., Botega Alves, N., Moretti Tomé, L., Silva Botelho, F., Rosário Neto, A., & Correa de Souza, D. (2022). Multiline aiming at phenotypic stability and rice blast resistance. Bioscience Journal, 38, e38100. [CrossRef]
- Flórez Ramos, C. P., Arias Suárez, J. C., Maldonado L, C. E., Cortina G, H. A., Moncada B, M. d., Quiroga Cardona, J., ... Duque O, H. (2018). Variedades Castillo® Zonales resistencia a la roya con mayor productividad. AVT489. Manizales, Caldas, Colombia.
Figure 1.
The scale of increasing lesions is used to measure incomplete resistance in detached leaves [5]. (0) Absence of visible injury. (1) Appearance of small, discoloured lesions. (2) Increased lesion surface and deeper discoloration. (3) Intensification of lesions and discoloration. (4) The appearance of the first spores. (5) Sporulation in less than 25% of the lesion surface. (6) Sporulation between 25% and 50% of the lesion surface. (7) Sporulation in more than 50% of the lesion surface.
Figure 1.
The scale of increasing lesions is used to measure incomplete resistance in detached leaves [5]. (0) Absence of visible injury. (1) Appearance of small, discoloured lesions. (2) Increased lesion surface and deeper discoloration. (3) Intensification of lesions and discoloration. (4) The appearance of the first spores. (5) Sporulation in less than 25% of the lesion surface. (6) Sporulation between 25% and 50% of the lesion surface. (7) Sporulation in more than 50% of the lesion surface.

Figure 2.
Probability of developing symptoms according to survival estimators (P<0.05).
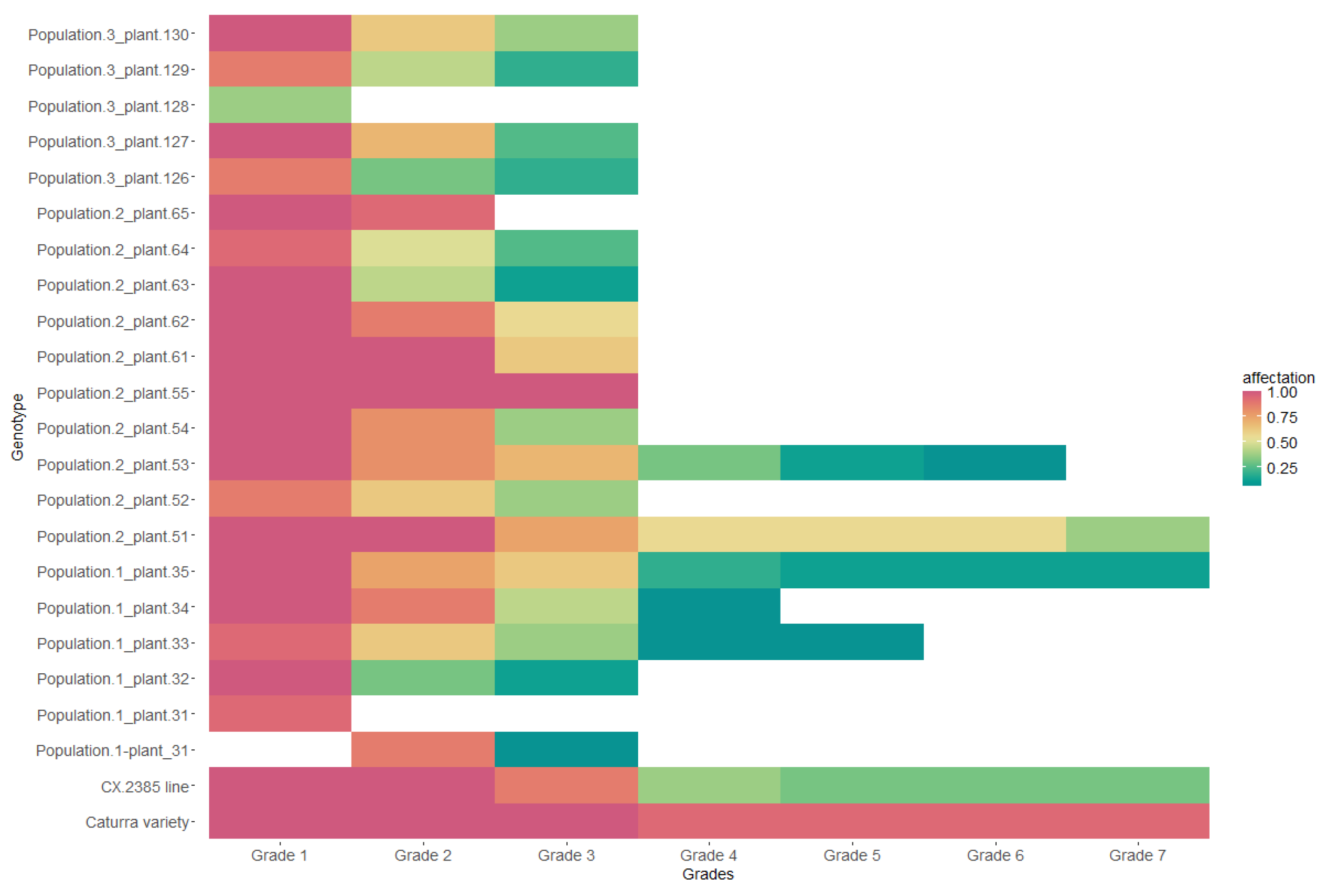
Figure 3.
Days vs. probability of survival to disease development in the genotypes of population 1 under laboratory conditions.
Figure 3.
Days vs. probability of survival to disease development in the genotypes of population 1 under laboratory conditions.
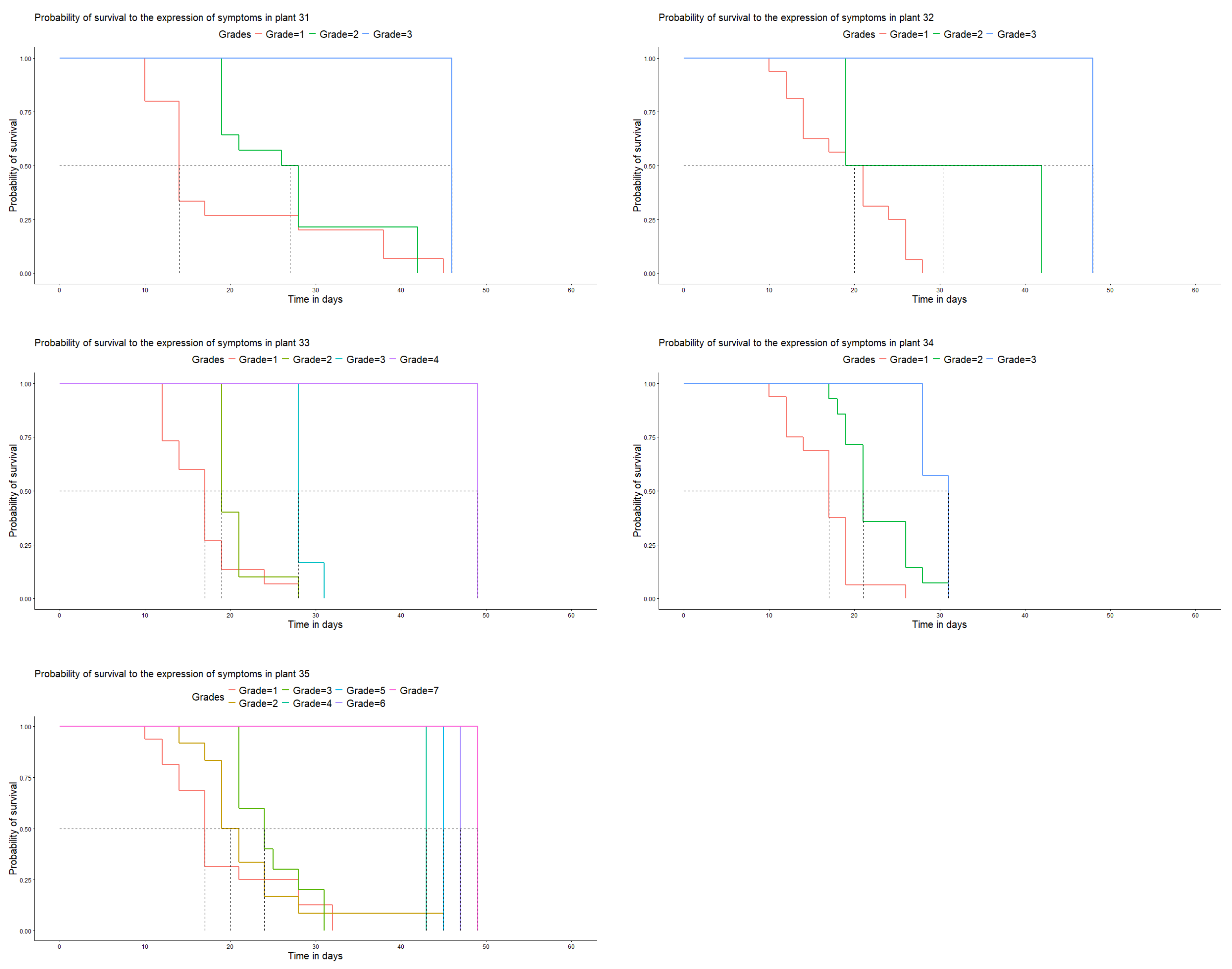
Figure 4.
Days vs. probability of survival to disease development in the genotypes of population 2 under laboratory conditions.
Figure 4.
Days vs. probability of survival to disease development in the genotypes of population 2 under laboratory conditions.
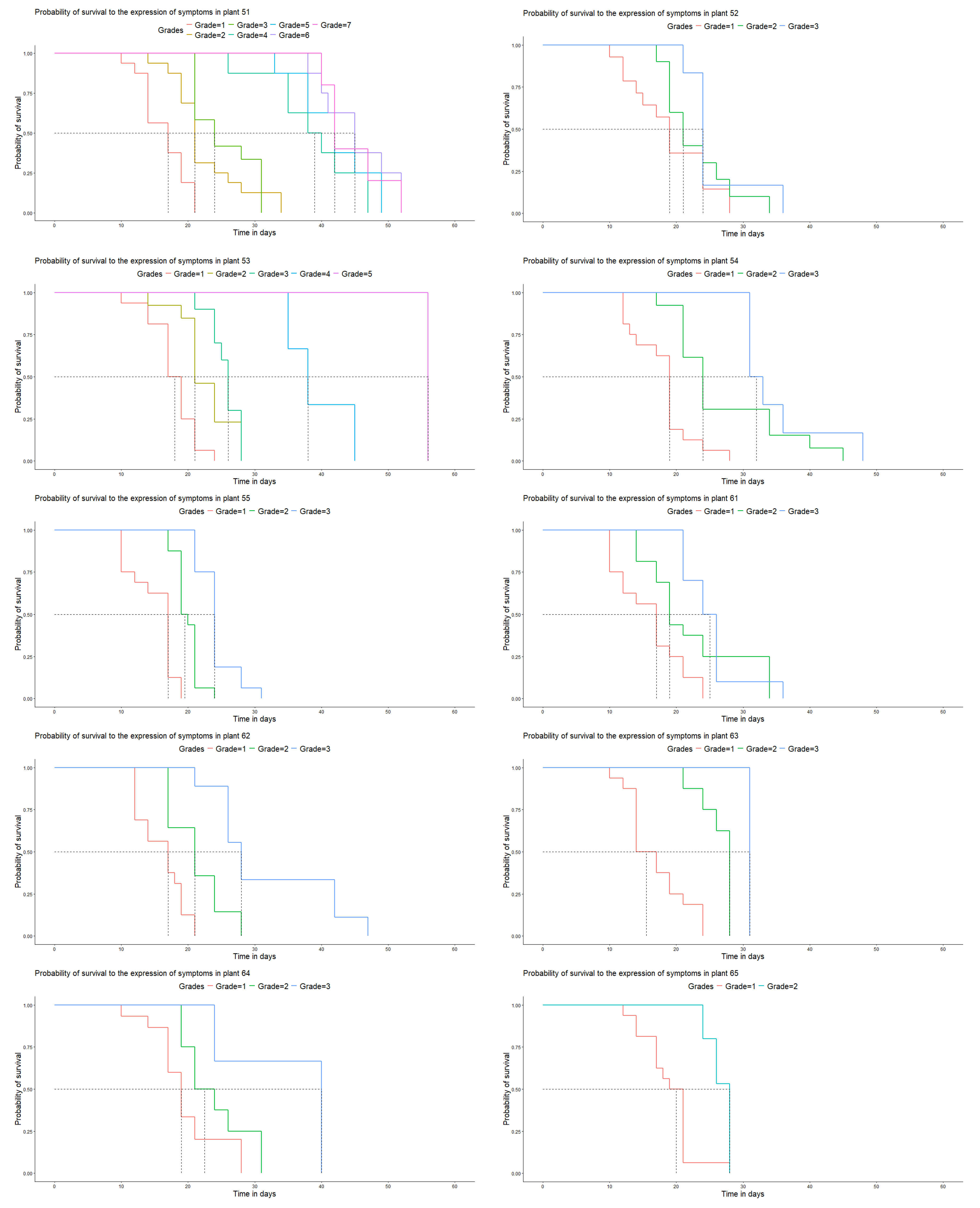
Figure 5.
Days vs. probability of survival to disease development in the genotypes of population 3 under laboratory conditions.
Figure 5.
Days vs. probability of survival to disease development in the genotypes of population 3 under laboratory conditions.
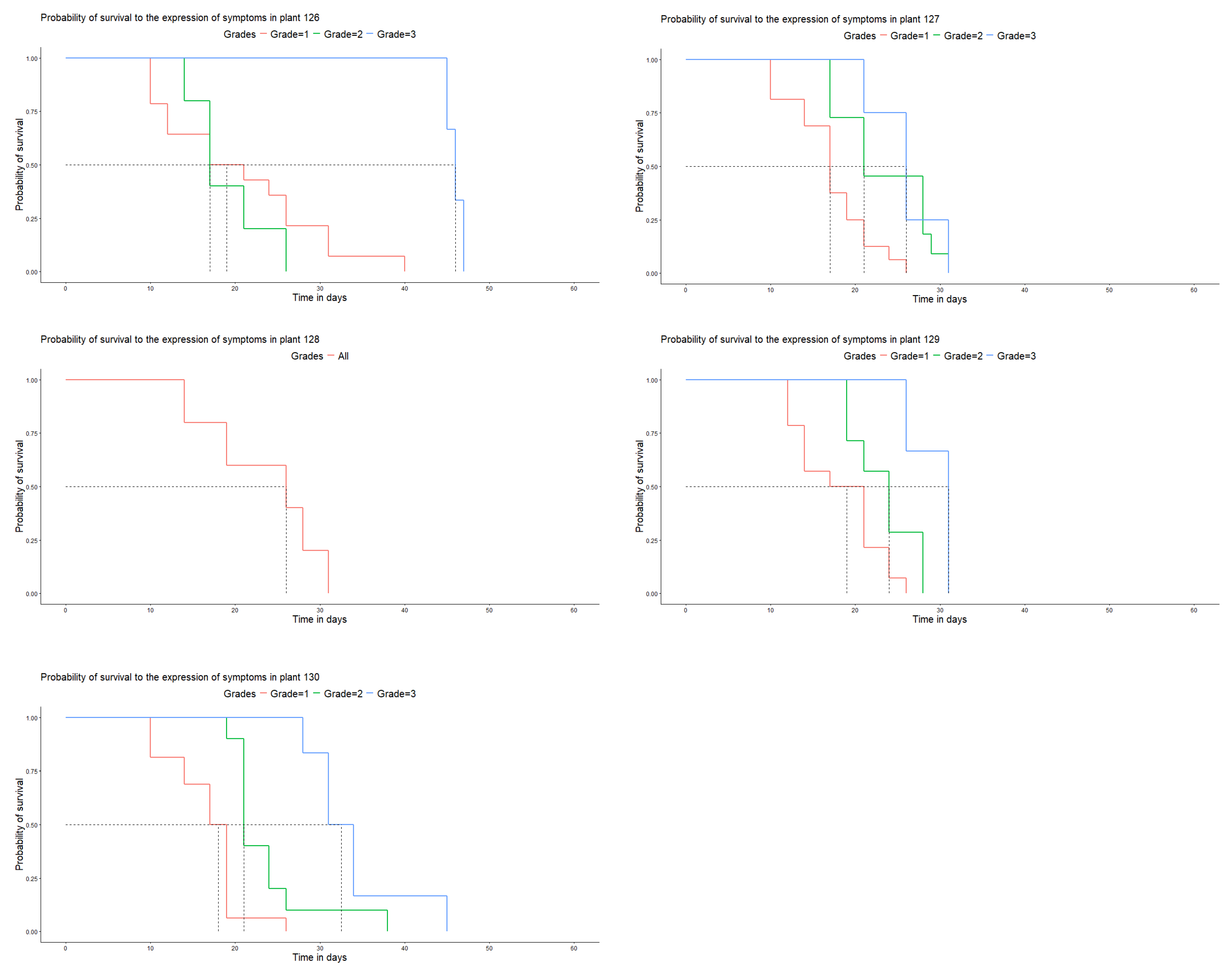
Figure 6.
Days vs. probability of survival to the development of the disease in the Caturra variety and CX.2385 line under laboratory conditions.
Figure 6.
Days vs. probability of survival to the development of the disease in the Caturra variety and CX.2385 line under laboratory conditions.
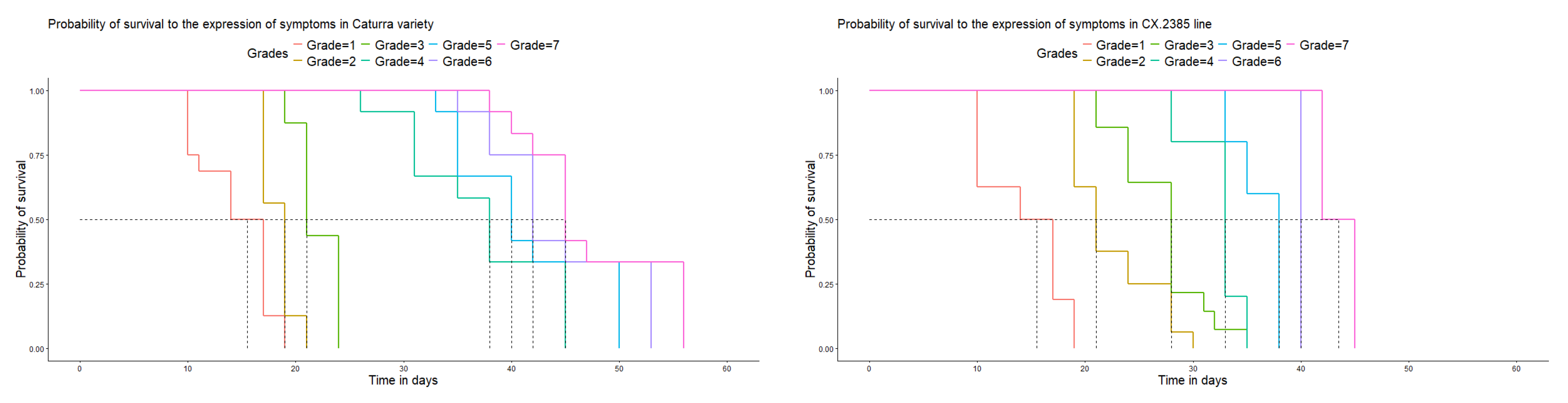

Table 1.
Rust incidence in field conditions of the populations evaluated using the Eskes & Toma-Braghini scale.
Table 1.
Rust incidence in field conditions of the populations evaluated using the Eskes & Toma-Braghini scale.
| Population | Rust incidence in field | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | Total Plants |
|
| 1.[(Caturra × CCC.32) × (Caturra × CCC.66)] × CX.2385 | 5 | 5 | |||||||||
| 2.CX.2385 × [(Caturra × CCC.32) × (Caturra × CCC.66)] | 1 | 9 | 10 | ||||||||
| 3.[Catuaí × (Caturra × CCC.66)] × CX.2385 | 1 | 4 | 5 | ||||||||
| Total Plants | 0 | 1 | 1 | 18 | 0 | 0 | 0 | 0 | 0 | 0 | 20 |
Table 2.
The observed symptoms and percentages of injuries were evaluated according to the scale of increasing injuries [5].
Table 2.
The observed symptoms and percentages of injuries were evaluated according to the scale of increasing injuries [5].
| Population | Plant | Percentages of involvement in the scale of increasing injuries | ||||||
|---|---|---|---|---|---|---|---|---|
| Grade 1 | Grade 2 | Grade 3 | Grade 4 | Grade 5 | Grade 6 | Grade 7 | ||
| 1 | 31 | 93.80 | 87.50 | 6.30 | ||||
| 32 | 100.00 | 27.30 | 9.40 | |||||
| 33 | 93.80 | 62.50 | 37.50 | 3.90 | 0.80 | |||
| 34 | 100.00 | 87.50 | 43.80 | 0.80 | ||||
| 35 | 100.00 | 75.00 | 62.50 | 6.30 | 5.50 | 5.50 | 5.50 | |
| 2 | 51 | 100.00 | 100.00 | 75.00 | 46.10 | 44.50 | 40.60 | 29.70 |
| 52 | 87,50 | 62.50 | 37.50 | |||||
| 53 | 100.00 | 81.30 | 63.30 | 18.00 | 6.30 | 0.80 | ||
| 54 | 100.00 | 81.30 | 37.50 | |||||
| 55 | 100.00 | 100.00 | 100.00 | |||||
| 61 | 100.00 | 100.00 | 62.50 | |||||
| 62 | 100.00 | 87.50 | 56.30 | |||||
| 63 | 100.00 | 43.80 | 12.50 | |||||
| 64 | 93.80 | 50.00 | 21.90 | |||||
| 65 | 100.00 | 93.80 | ||||||
| 3 | 126 | 87.50 | 31.30 | 18.80 | ||||
| 127 | 100.00 | 68.80 | 25.00 | |||||
| 128 | 32.80 | |||||||
| 129 | 87.50 | 43.80 | 18.80 | |||||
| 130 | 100.00 | 62.50 | 37.50 | |||||
| Caturra variety | 100.00 | 100.00 | 100.00 | 78.10 | 78.10 | 78.10 | 77.30 | |
| CX.2385 line | 100.00 | 100.00 | 87.50 | 26.60 | 25.80 | 24.20 | 24.20 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



