
Submitted:
01 May 2024
Posted:
03 May 2024
You are already at the latest version
Abstract
The microbiome is capable of modulating the bioavailability of chemotherapy drugs mainly due to metabolizing these agents. Multiple cytostatic bacterial metabolites were recently identified that have cytostatic properties on cancer cells. In this study we addressed the question whether the cytostatic bacterial metabolites (cadaverine, indolepropionic acid, indoxylsulfate) can interfere with the cytostatic effects of the chemotherapy agents used in the management of breast cancer (doxorubicin, gemcitabine, irinotecan, methotrexate, rucaparib, 5-fluorouracil, paclitaxel). The chemotherapy drugs were applied in a wide concentration range to which a bacterial metabolite was added in a concentration within its serum reference range and the effects on cell proliferation were assessed. There was no interference between doxorubicin, gemcitabine, irinotecan, methotrexate, rucaparib and the bacterial metabolites. Nevertheless, cadaverine increased the Hill coefficient of the inhibitory curve of 5-fluorouracil and indolepropionic acid increased the IC50 value of 5-fluorouracil that are either disadvantageous effects or effects of unknown significance. Nevertheless, indolepropionic acid decreased the IC50 value of paclitaxel that is a potentially advantageous combination.
Keywords:
5-fluorouracil
; doxorubicin
; gemcitabine
; irinotecan
; methotrexate
; rucaparib
; paclitaxel
; cadaverine
; indolepropionic acid
; indoxylsulfate
; cell proliferation
; breast cancer
1. Introduction
Oncobiosis is the dysbiosis associated with neoplastic diseases. Oncobiosis associates with numerous cancers and affects multiple microbiome compartments [1,2,3]. There are three major pathways through which the oncobiome can supports tumor progression and metastasis formation: 1) direct colonization of the tumor tissue, 2) immune suppression, 3) the production of bacterial metabolites and toxins [3]. Although, these pathways are all active in breast cancer, metabolite production has key role [3].
Multiple bacterial metabolites were identified with cytostatic [3,4,5,6,7,8,9,10,11,12,13,14], pro-proliferative [15,16,17,18,19,20,21,22,23,24] or mixed [14] properties in breast cancer. These metabolites are chemically very diverse. Metabolites with cytostatic properties elicit pleiotropic effects involving the induction of an anti-Warburg type metabolic rearrangement and the induction of mild oxidative stress that block epithelial-mesenchymal transition, reduction of the proportions of cancer stem cells, culminating in cytostasis and reduced metastatic and recurrence potential [3].
There are numerous reports showing that the microbiome interferes with the metabolism and the effectiveness of the chemotherapy agents used in breast cancer management [25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40]. This raised a possibility for other indirect interactions, namely, cytostatic metabolites may add on or potentiate the effectiveness of chemotherapy agents. In this study we set out to investigate that possibility in a cell model of breast cancer.
2. Results
2.1. General Consideratons
For the studies we selected three well-characterized cytostatic bacterial metabolites, cadaverine (CAD), indolepropionic acid (IPA) and indoxylsulfate (IS) that were applied in concentrations corresponding to the top of the serum reference concentration of these metabolites as follows: CAD: 0.8 µM [41,42], IPA: 1 µM [43,44,45], IS: 4 µM [46]. We investigated doxorubicin (DOX), gemcitabine (GEM), irinotecan (IRI), methotrexate (MTX), rucaparib (RUCA), 5-fluorouracil (5FU) and paclitaxel (PAC) all applied in a serial dilution series as indicated on the corresponding figures (similar to [47] or [48]).
2.2. Bacterial Metabolites Do Not Interfere with Doxorubicin, Gemcitabine, Irinotecan, Methotrexate and Rucaparib Activity
We tested the effects of CAD, IS and IPA on the inhibitory properties of DOX, GEM, IRI, MTX and RUCA on cell proliferation. None of the metabolites impacted on the inhibitory activity of the chemotherapeutic agents, neither on the overall presentation of the inhibitory curves, nor on the kinetic readouts as the IC50 value or the Hill coefficient (Figure 1, Figure 2, Figure 3, Figure 4 and Figure 5).
2.3. Bacterial Metabolites Interfere with 5-Fluorouracil
The three bacterial metabolites were tested together with 5FU, an antimetabolite chemotherapeutic agent. CAD increased the Hill coefficient, but did not change the IC50 value (Figure 6). Unfortunately, IPA increased the IC50 value of 5FU but did not affect the Hill coefficient (Figure 6). IS did not impact on the kinetic values of 5FU (Figure 6).
2.4. Bacterial Metabolites Interfere with Paclitaxel
3. Discussion
Chemotherapy plays a pivotal role in the management of breast cancer. The chemotherapy regimens are built on anthracyclines, cyclophosphamides, taxanes, antimetabolites (5-fluorouracil, gemcitabine, capecitabine), navelbine [49], targeted therapeutic agents as trastuzumab, pertuzumab and trastuzumab-emtansine, lapatinib [50], endocrine therapy, including selective estrogen receptor modulators (SERMs), aromatase inhibitors and GNRH-analogs [50] and novel therapeutic agents as PARP inhibitors [51,52,53] or CDK4/6 (cyclin-dependent kinases) inhibitors [54]. In this study we assessed those inhibitors that can be utilized in cell-based model systems, as their action does not require the activation in the liver, interaction with the immune system (as for humanized antibodies) or systemic endocrine loops (e.g., SERMs).
In this study we investigated whether there is an interaction between cytostatic bacterial metabolites and the above-mentioned chemotherapy agents. Numerous bacterial metabolites were identified, the majority of which have cytostatic properties [3,4,5,6,7,8,9,10,11,12,13,14]. The production of these metabolites decline in breast cancer patients, nevertheless, administration of minute quantities of these metabolites reduce the mitotic rate and the metastatic potential of the primary tumor [4,5,6,7].
Multiple chemotherapy agents were shown to modulate the composition of the microbiome [55,56], bacterial metabolism of chemotherapy agents were also evidenced [32,34,35,36,37,38,39,40] and the efficiency of humanized antibodies [57] were also linked to compositional changes to the microbiome. These observations raised the possibility that bacterial metabolites may interfere with the cytostatic or cytotoxic effects of chemotherapy agents. Despite our negative observations on the interactions between bile acids and chemotherapy agents in pancreatic adenocarcinoma cells [58,59], in the current study we identified CAD and IPA that do interfere with PAC and 5FU. Neither metabolites displayed toxicity towards non-transformed cells in previous studies and can be applied in low concentrations [5,6].
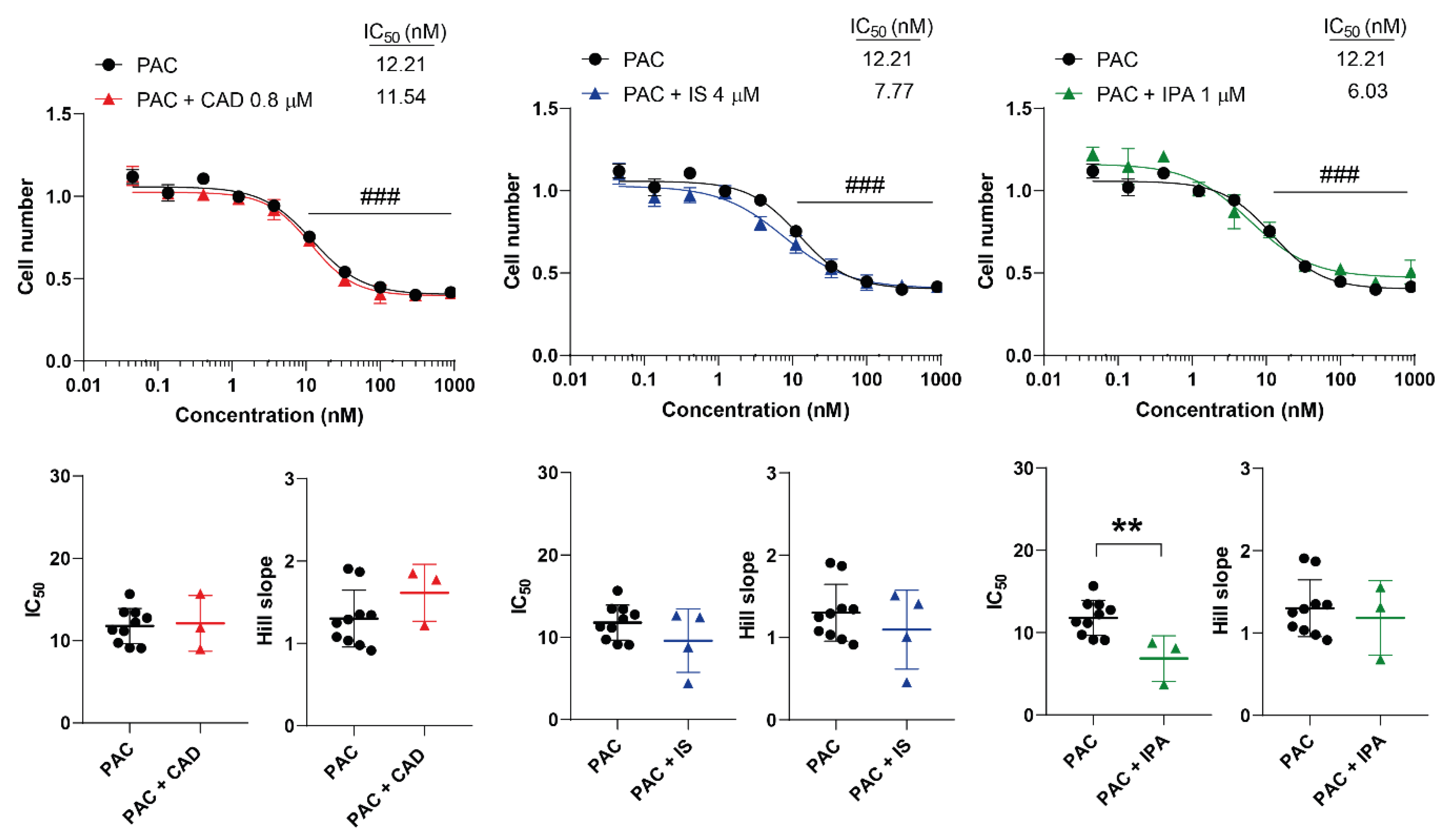
CAD increased the Hill coefficient of 5FU suggesting a more collaborative binding of the drug molecules [60] that has unknown pharmacological relevance. In contrast, IPA increased the IC50 value of 5FU suggesting a lower efficiency that has negative pharmacological and, likely, clinical consequences. While in the case of PAC the IC50 value was halved in the presence of IPA making this combination a potential. These findings also suggest that IPA may have adverse effects when PAC+5FU combinations are applied.
IPA is a bacterial metabolite that is the synthesized from tryptophan through deamination by tryptophanase (TnaA) [5,45]. A significant portion of tryptophan (4-6%) undergoes bacterial catabolism [61]. Multiple studies have shown that disturbances to indole/tryptophan metabolism correlates with survival in breast cancer ([6,7,62,63], reviewed in [6]). Our observations extend these by adding that higher IPA levels may support PAC responsiveness.
4. Materials and Methods
Chemicals
Bacterial metabolites (Cadaverine-CAD, cat # C8561; Indoxylsulfate-IS, cat # 13875; Indolepropionic acid-IPA, cat # 220027) were purchased from Sigma-Aldrich (St. Louis, MI, USA). All metabolites were dissolved in dimethyl-sulfoxide (DMSO) at a stock concentration of 100 mM. CAD was used at concentrations of 0.8 μM, IS at 4 μM and IPA at 1 μM, corresponding to normal human serum concentrations of these metabolites [41,42,43,44,45,46].
Chemotherapy drugs, Irinotecan (IRI, cat # I1406), 5-fluorouracil (5FU, cat # F6627), Methotrexate (MTX, cat # PHR1396), Rucaparib (RUCA, cat # PZ0036) and Gemcitabine (GEM, cat # G6423) were from Sigma-Aldrich. The drugs IRI, 5-FU, MTX and RUCA were dissolved in DMSO at a stock concentration of 100 mM; GEM was dissolved in water at a stock concentration of 100 mM. Liposomal Encapsuled Doxorubicin (DOX-NP, cat # 300112) was purchased from Avanti and a stock solution of 50 mM was prepared. Paclitaxel (PAC, cat # A0451335) was from Thermo Fisher Scientific (Waltham, MA, USA) and 50 mM stock solution was prepared in DMSO.
Chemotherapy compounds were used at different concentrations as indicated in the figures.
Cell Line
The 4T1 breast cancer cell line was obtained from the American Type Culture Collection. Cells were cultured in RPMI-1640 medium (Sigma-Aldrich, cat # R5886) containing 10% fetal bovine serum (FBS), 1% penicillin/streptomycin, 2 mM L-glutamine and 1% pyruvate at 37°C in a humidified incubator with 5% CO2. Cells were regularly checked for Mycoplasma contamination.
MTT Assay
4T1 cells were plated in 96-well plates (1.5 x 103 cell/well). On the next day cells were treated with chemotherapy agents alone or in combination with bacterial metabolites for 48 h. After treatments cell numbers was determined using an MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay. Briefly, cells were treated with MTT solution (0.5 mg/ml) and incubated at 37°C for 90 minutes. Then, culture medium was discarded and the formazan crystals were dissolved in DMSO. The absorbance was measured on a plate reader (Thermo Labsystems Multiskan MS, Walthman, MA, USA) at 540 nm. In the calculations, the absorbance values for the vehicle-treated cells were considered 1, and all treatment were expressed relative to 1.
Statistical Analysis
Each analyses were performed using GraphPad Prism 8 software. Experiments were repeated at least three times and results are presented as mean ± SEM values. Normal distribution of the values was tested using D’Agostino & Pearson normality test. Where appropriate, values were log-normalized or normalized using Box-Cox normalization method [64]. Nonlinear regression was performed using the GraphPad “[Inhibitor] vs. response - Variable slope (four parameters)” utility, from which IC50 and Hill slope values were obtained unless otherwise stated. Two-way analysis of variance test followed by Tukey’s honestly significant post hoc test were used for multiple comparisons.
5. Conclusions
The oncobiome was shown to modulate the efficacy or even limit the availability of chemotherapy agents. In this study we showed that, in contrast to previous negative findings in pancreatic adenocarcinoma models, IPA and CAD modulated the cytostatic activity of 5FU and PAC. Importantly, IPA decreased the IC50 values of PAC that is a beneficial interaction, as PAC concentrations can be decreased in combination with a low concentration non-toxic compound that may limit the side effects of PAC.
Supplementary Materials
No supplementary material is associated with the manuscript.
Author Contributions
Conceptualization, E.M. and P.B.; methodology, E.M., P.B., E.J. and S.S; software, S.S., P.N and T.I.B.; validation, E.M. and P.B.; formal analysis, E.M., P.B. and S.S.; investigation, S.S., T.I.B. P.N. and E.J..; resources, E.M. and P.B.; data curation, S.S.; writing—original draft preparation, S.S., E.M. and P.B; writing—review and editing, E.M and P.B.; visualization, E.M.; supervision, E.M. and P.B.; project administration, E.M and S.S.; funding acquisition, E.M and P.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the NKFIH (FK128387, K142141, TKP2021-EGA-19, TKP-EGA-20). Project no. TKP2021-EGA-19 has been implemented with the support provided from the National Research, Development and Innovation Fund of Hungary, financed under the TKP2021-EGA funding scheme. Grant from the Hungarian Academy of Sciences (POST-COVID2021-33). Supported by the University of Debrecen Program for Scientific Publication. This project has received funding from the HUN-REN Hungarian Research Network. The APC was funded by the University of Debrecen and the grants of the authors. The work was also supported by the ÚNKP-23 New National Excellence Program of the Ministry for Culture and Innovation from the Source of National research, Development, and Innovation Fund (ÚNKP-23-3-II-DE-151, ÚNKP-23-3-I-DE-184, ÚNKP-23-4-II-DE-172).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Primary data of the present manuscript can be found at https://figshare.com/s/6ecb8c6bd8b87284ae3c (DOI: 10.6084/m9.figshare.25678635).
Acknowledgments
The authors are grateful for the technical assistance of Ms. Kitti Barta.
Conflicts of Interest
Péter Bai is a CEO and shareholder of Holobiont Diagnostics LTD, a developer of cancer diagnostic tests. Other authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Sipos, A.; Ujlaki, G.; Mikó, E.; Maka, E.; Szabó, J.; Uray, K.; Krasznai, Z.; Bai, P. The role of the microbiome in ovarian cancer: mechanistic insights into oncobiosis and to bacterial metabolite signaling. Molecular medicine (Cambridge, Mass.) 2021, 27, 33. [CrossRef]
- Kiss, B.; Mikó, E.; Sebő, É.; Toth, J.; Ujlaki, G.; Szabó, J.; Uray, K.; Bai, P.; Árkosy, P. Oncobiosis and Microbial Metabolite Signaling in Pancreatic Adenocarcinoma. Cancers (Basel) 2020, 12, E1068. [CrossRef]
- Kovács, T.; Mikó, E.; Ujlaki, G.; Yousef, H.; Csontos, V.; Uray, K.; Bai, P. The involvement of oncobiosis and bacterial metabolite signaling in metastasis formation in breast cancer. Cancer and Metastasis Reviews 2021, 40, 1223-1249.
- Miko, E.; Vida, A.; Kovacs, T.; Ujlaki, G.; Trencsenyi, G.; Marton, J.; Sari, Z.; Kovacs, P.; Boratko, A.; Hujber, Z.; et al. Lithocholic acid, a bacterial metabolite reduces breast cancer cell proliferation and aggressiveness. Biochimica et Biophysica Acta - Bioenergetics 2018, 1859, 958-974.
- Kovács, T.; Mikó, E.; Vida, A.; Sebő, É.; Toth, J.; Csonka, T.; Boratkó, A.; Ujlaki, G.; Lente, G.; Kovács, P.; et al. Cadaverine, a metabolite of the microbiome, reduces breast cancer aggressiveness through trace amino acid receptors. Scientific Reports 2019, 9, 1300. [CrossRef]
- Sári, Z.; Mikó, E.; Kovács, T.; Jankó, L.; Csonka, T.; Sebő, E.; Toth, J.; Tóth, D.; Árkosy, P.; Boratkó, A.; et al. Indolepropionic acid, a metabolite of the microbiome, has cytostatic properties in breast cancer by activating AHR and PXR receptors and inducing oxidative stress. Cancers (Basel) 2020, 12, 2411. [CrossRef]
- Sári, Z.; Mikó, E.; Kovács, T.; Boratkó, A.; Ujlaki, G.; Jankó, L.; Kiss, B.; Uray, K.; Bai, P. Indoxylsulfate, a Metabolite of the Microbiome, Has Cytostatic Effects in Breast Cancer via Activation of AHR and PXR Receptors and Induction of Oxidative Stress Cancers (Basel) 2020, 12, 2915. [CrossRef]
- Wang, H.; Rong, X.; Zhao, G.; Zhou, Y.; Xiao, Y.; Ma, D.; Jin, X.; Wu, Y.; Yan, Y.; Yang, H.; et al. The microbial metabolite trimethylamine N-oxide promotes antitumor immunity in triple-negative breast cancer. Cell Metab 2022, 34, 581-594.e588. [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451-455.
- Rodrigues, M.F.; Carvalho, E.; Pezzuto, P.; Rumjanek, F.D.; Amoedo, N.D. Reciprocal modulation of histone deacetylase inhibitors sodium butyrate and trichostatin A on the energy metabolism of breast cancer cells. J Cell Biochem. 2015, 116, 797-808.
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain Fatty acids in health and disease. Adv Immunol 2014, 121, 91-119.
- Schulthess, J.; Pandey, S.; Capitani, M.; Rue-Albrecht, K.C.; Arnold, I.; Franchini, F.; Chomka, A.; Ilott, N.E.; Johnston, D.G.W.; Pires, E.; et al. The Short Chain Fatty Acid Butyrate Imprints an Antimicrobial Program in Macrophages. Immunity. 2019, 50, 432-445.
- Salimi, V.; Shahsavari, Z.; Safizadeh, B.; Hosseini, A.; Khademian, N.; Tavakoli-Yaraki, M. Sodium butyrate promotes apoptosis in breast cancer cells through reactive oxygen species (ROS) formation and mitochondrial impairment. Lipids Health Dis. 2017, 16, 208.
- Ujlaki, G.; Kovács, T.; Vida, A.; Kókai, E.; Rauch, B.; Schwarcz, S.; Mikó, E.; Janka, E.; Sipos, A.; Hegedűs, C.; et al. Identification of Bacterial Metabolites Modulating Breast Cancer Cell Proliferation and Epithelial-Mesenchymal Transition. Molecules (Basel, Switzerland) 2023, 28, 5898.
- Radde, B.N.; Ivanova, M.M.; Mai, H.X.; Salabei, J.K.; Hill, B.G.; Klinge, C.M. Bioenergetic differences between MCF-7 and T47D breast cancer cells and their regulation by oestradiol and tamoxifen. Biochem J. 2015, 465, 49-61.
- Radde, B.N.; Ivanova, M.M.; Mai, H.X.; Alizadeh-Rad, N.; Piell, K.; Van Hoose, P.; Cole, M.P.; Muluhngwi, P.; Kalbfleisch, T.S.; Rouchka, E.C.; et al. Nuclear respiratory factor-1 and bioenergetics in tamoxifen-resistant breast cancer cells. Exp Cell Res. 2016, 347, 222-231.
- Sotgia, F.; Lisanti, M.P. Mitochondrial mRNA transcripts predict overall survival, tumor recurrence and progression in serous ovarian cancer: Companion diagnostics for cancer therapy. Oncotarget. 2017, 8, 66925-66939.
- Gandhi, N.; Das, G.M. Metabolic Reprogramming in Breast Cancer and Its Therapeutic Implications. Cells. 2019, 8, 89. [CrossRef]
- Zacksenhaus, E.; Shrestha, M.; Liu, J.C.; Vorobieva, I.; Chung, P.E.D.; Ju, Y.; Nir, U.; Jiang, Z. Mitochondrial OXPHOS Induced by RB1 Deficiency in Breast Cancer: Implications for Anabolic Metabolism, Stemness, and Metastasis. Trends Cancer. 2017, 3, 768-779.
- Maximov, P.Y.; Abderrahman, B.; Curpan, R.F.; Hawsawi, Y.M.; Fan, P.; Jordan, V.C. A unifying biology of sex steroid-induced apoptosis in prostate and breast cancers. Endocr Relat Cancer. 2018, 25, R83-R113.
- Al-Howail, H.A.; Hakami, H.A.; Al-Otaibi, B.; Al-Mazrou, A.; Daghestani, M.H.; Al-Jammaz, I.; Al-Khalaf, H.H.; Aboussekhra, A. PAC down-regulates estrogen receptor alpha and suppresses epithelial-to-mesenchymal transition in breast cancer cells. BMC Cancer. 2016, 16, 540.
- Bouris, P.; Skandalis, S.S.; Piperigkou, Z.; Afratis, N.; Karamanou, K.; Aletras, A.J.; Moustakas, A.; Theocharis, A.D.; Karamanos, N.K. Estrogen receptor alpha mediates epithelial to mesenchymal transition, expression of specific matrix effectors and functional properties of breast cancer cells. Matrix Biol. 2015, 43, 42-60.
- Kulkoyluoglu-Cotul, E.; Arca, A.; Madak-Erdogan, Z. Crosstalk between Estrogen Signaling and Breast Cancer Metabolism. Trends Endocrinol Metab. 2019, 30, 25-38.
- Wu, Q.; Hatse, S.; Kenis, C.; Fernández-García, J.; Altea-Manzano, P.; Billen, J.; Planque, M.; Vandekeere, A.; Lambrechts, Y.; Richard, F.; et al. Serum methylmalonic acid concentrations at breast cancer diagnosis significantly correlate with clinical frailty. GeroScience 2024, 46, 1489-1498. [CrossRef]
- Cox, G.; Koteva, K.; Wright, G.D. An unusual class of anthracyclines potentiate Gram-positive antibiotics in intrinsically resistant Gram-negative bacteria. J Antimicrob Chemother. 2014, 69, 1844-1855.
- Westman, E.L.; Canova, M.J.; Radhi, I.J.; Koteva, K.; Kireeva, I.; Waglechner, N.; Wright, G.D. Bacterial inactivation of the anticancer drug doxorubicin. Chem Biol. 2012, 19, 1255-1264.
- Parajuli, P.; Pandey, R.P.; Nguyen, T.H.T.; Dhakal, D.; Sohng, J.K. Substrate Scope of O-Methyltransferase from Streptomyces peucetius for Biosynthesis of Diverse Natural Products Methoxides. Appl Biochem Biotechnol. 2018, 184, 1404-1420.
- Dhakal, D.; Lim, S.K.; Kim, D.H.; Kim, B.G.; Yamaguchi, T.; Sohng, J.K. Complete genome sequence of Streptomyces peucetius ATCC 27952, the producer of anticancer anthracyclines and diverse secondary metabolites. J Biotechnol. 2018, 267, 50-54.
- Zabala, D.; Brana, A.F.; Florez, A.B.; Salas, J.A.; Mendez, C. Engineering precursor metabolite pools for increasing production of antitumor mithramycins in Streptomyces argillaceus. Metab Eng. 2013, 20, 187-197.
- Dou, T.Y.; Luan, H.W.; Liu, X.B.; Li, S.Y.; Du, X.F.; Yang, L. Enzymatic hydrolysis of 7-xylosyltaxanes by an extracellular xylosidase from Cellulosimicrobium cellulans. Biotechnol Lett. 2015, 37, 1905-1910.
- Zhou, D.J.; Pan, J.; Yu, H.L.; Zheng, G.W.; Xu, J.H. Target-oriented discovery of a new esterase-producing strain Enterobacter sp. ECU1107 for whole cell-catalyzed production of (2S,3R)-3-phenylglycidate as a chiral synthon of Taxol. Appl Microbiol Biotechnol. 2013, 97, 6293-6300.
- Garcia-Gonzalez, A.P.; Ritter, A.D.; Shrestha, S.; Andersen, E.C.; Yilmaz, L.S.; Walhout, A.J.M. Bacterial Metabolism Affects the C. elegans Response to Cancer Chemotherapeutics. Cell. 2017, 169, 431-441.
- Scott, T.A.; Quintaneiro, L.M.; Norvaisas, P.; Lui, P.P.; Wilson, M.P.; Leung, K.Y.; Herrera-Dominguez, L.; Sudiwala, S.; Pessia, A.; Clayton, P.T.; et al. Host-Microbe Co-metabolism Dictates Cancer Drug Efficacy in C. elegans. Cell. 2017, 169, 442-456.
- Geller, L.T.; Straussman, R. Intratumoral bacteria may elicit chemoresistance by metabolizing anticancer agents. Mol Cell Oncol. 2018, 5, e1405139.
- Sandrini, M.P.; Shannon, O.; Clausen, A.R.; Bjorck, L.; Piskur, J. Deoxyribonucleoside kinases activate nucleoside antibiotics in severely pathogenic bacteria. Antimicrob Agents Chemother. 2007, 51, 2726-2732.
- Sandrini, M.P.; Clausen, A.R.; On, S.L.; Aarestrup, F.M.; Munch-Petersen, B.; Piskur, J. Nucleoside analogues are activated by bacterial deoxyribonucleoside kinases in a species-specific manner. J Antimicrob Chemother. 2007, 60, 510-520.
- Alexander, J.L.; Wilson, I.D.; Teare, J.; Marchesi, J.R.; Nicholson, J.K.; Kinross, J.M. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat Rev Gastroenterol Hepatol. 2017, 14, 356-365.
- Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; Shental, N.; Nejman, D.; Gavert, N.; Zwang, Y.; Cooper, Z.A.; Shee, K.; et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science. 2017, 357, 1156-1160.
- Vande Voorde, J.; Sabuncuoglu, S.; Noppen, S.; Hofer, A.; Ranjbarian, F.; Fieuws, S.; Balzarini, J.; Liekens, S. Nucleoside-catabolizing enzymes in mycoplasma-infected tumor cell cultures compromise the cytostatic activity of the anticancer drug gemcitabine. J Biol Chem. 2014, 289, 13054-13065. [CrossRef]
- Lehouritis, P.; Cummins, J.; Stanton, M.; Murphy, C.T.; McCarthy, F.O.; Reid, G.; Urbaniak, C.; Byrne, W.L.; Tangney, M. Local bacteria affect the efficacy of chemotherapeutic drugs. Sci Rep. 2015, 5:14554., . [CrossRef]
- Loser, C.; Folsch, U.R.; Paprotny, C.; Creutzfeldt, W. Polyamine concentrations in pancreatic tissue, serum, and urine of patients with pancreatic cancer. Pancreas. 1990, 5, 119-127.
- Loser, C.; Folsch, U.R.; Paprotny, C.; Creutzfeldt, W. Polyamines in colorectal cancer. Evaluation of polyamine concentrations in the colon tissue, serum, and urine of 50 patients with colorectal cancer. Cancer. 1990, 65, 958-966.
- Danaceau, J.P.; Anderson, G.M.; McMahon, W.M.; Crouch, D.J. A liquid chromatographic-tandem mass spectrometric method for the analysis of serotonin and related indoles in human whole blood. J Anal Toxicol. 2003, 27, 440-444.
- Rosas, H.D.; Doros, G.; Bhasin, S.; Thomas, B.; Gevorkian, S.; Malarick, K.; Matson, W.; Hersch, S.M. A systems-level “misunderstanding”: the plasma metabolome in Huntington’s disease. Ann Clin Transl Neurol. 2015, 2, 756-768.
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018, 9, 3294.
- Lin, C.-N.; Wu, I.W.; Huang, Y.-F.; Peng, S.-Y.; Huang, Y.-C.; Ning, H.-C. Measuring serum total and free indoxyl sulfate and p-cresyl sulfate in chronic kidney disease using UPLC-MS/MS. Journal of Food and Drug Analysis 2019, 27, 502-509. [CrossRef]
- Kacsir, I.; Sipos, A.; Kiss, T.; Major, E.; Bajusz, N.; Tóth, E.; Buglyo, P.; Somsak, L.; Kardos, G.; Bai, P.; et al. Half Sandwich-Type Osmium, Ruthenium, Iridium and Rhodium Complexes with Bidentate Glycosyl Heterocyclic Ligands Induce Cytostasis in Platinum-Resistant Ovarian Cancer Cells and Bacteriostasis in Gram-Positive Multiresistant Bacteria. Frontiers in Chemistry 2023, 11, 1086267.
- Kacsir, I.; Sipos, A.; Major, E.; Bajusz, N.; Bényei, A.; Buglyó, P.; Somsák, L.; Kardos, G.; Bai, P.; Bokor, É. Half-Sandwich Type Platinum-Group Metal Complexes of C-Glucosaminyl Azines: Synthesis and Antineoplastic and Antimicrobial Activities. Molecules (Basel, Switzerland) 2023, 28, 3058. [CrossRef]
- Senkus, E.; Kyriakides, S.; Ohno, S.; Penault-Llorca, F.; Poortmans, P.; Rutgers, E.; Zackrisson, S.; Cardoso, F. Primary breast cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann Oncol. 2015, 26, v8-30.
- Harbeck, N.; Gnant, M. Breast cancer. Lancet. 2017, 389, 1134-1150.
- Curtin, N.; Szabo, C. Therapeutic Applications of PARP Inhibitors: Anticancer Therapy and Beyond. Molecular Aspects of Medicine 2013, 6, 1043-1258.
- Bai, P. Biology of Poly(ADP-Ribose) Polymerases: The Factotums of Cell Maintenance. Molecular Cell 2015, 58, 947-958.
- Fong, P.C.; Boss, D.S.; Yap, T.A.; Tutt, A.; Wu, P.; Mergui-Roelvink, M.; Mortimer, P.; Swaisland, H.; Lau, A.; O’Connor, M.J.; et al. Inhibition of Poly(ADP-Ribose) Polymerase in Tumors from BRCA Mutation Carriers. N Engl J Med 2009, 361(2), 123-134.
- Kwapisz, D. Cyclin-dependent kinase 4/6 inhibitors in breast cancer: palbociclib, ribociclib, and abemaciclib. Breast Cancer Res Treat. 2017, 166, 41-54.
- Fan, Y.; Liang, L.; Tang, X.; Zhu, J.; Mu, L.; Wang, M.; Huang, X.; Gong, S.; Xu, J.; Liu, T.; et al. Changes in the gut microbiota structure and function in rats with doxorubicin-induced heart failure. Front Cell Infect Microbiol 2023, 13, 1135428. [CrossRef]
- Tian, Z.; Qiao, X.; Wang, Z.; Li, X.; Pan, Y.; Wei, X.; Lv, Z.; Li, P.; Du, Q.; Wei, W.; et al. Cisplatin and doxorubicin chemotherapy alters gut microbiota in a murine osteosarcoma model. Aging (Albany NY) 2024, 16, 1336-1351. [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 2015, 350, 1084-1089. [CrossRef]
- Schwarcz, S.; Kovács, P.; Kovács, T.; Ujlaki, G.; Nyerges, P.; Uray, K.; Bai, P.; Mikó, E. The pro- and antineoplastic effects of deoxycholic acid in pancreatic adenocarcinoma cell models. Mol Biol Rep 2023, 50, 5273-5282. [CrossRef]
- Schwarcz, S.; Kovács, P.; Nyerges, P.; Ujlaki, G.; Sipos, A.; Uray, K.; Bai, P.; Mikó, E. A bacterial metabolite, lithocholic acid has anti-carcinogenic effects in pancreatic adenocarcinoma. Research Square 2024.
- Gesztelyi, R.; Zsuga, J.; Kemeny-Beke, A.; Varga, B.; Juhasz, B.; Tosaki, A. The Hill equation and the origin of quantitative pharmacology. Archive for History of Exact Sciences 2012, 66, 427-438. [CrossRef]
- Yokoyama, M.T.; Carlson, J.R. Microbial metabolites of tryptophan in the intestinal tract with special reference to skatole. Am J Clin Nutr. 1979, 32, 173-178.
- Auslander, N.; Yizhak, K.; Weinstock, A.; Budhu, A.; Tang, W.; Wang, X.W.; Ambs, S.; Ruppin, E. A joint analysis of transcriptomic and metabolomic data uncovers enhanced enzyme-metabolite coupling in breast cancer. Sci Rep. 2016, 6, 29662.
- Tang, X.; Lin, C.C.; Spasojevic, I.; Iversen, E.S.; Chi, J.T.; Marks, J.R. A joint analysis of metabolomics and genetics of breast cancer. Breast Cancer Research 2014, 16, 415.
- Box, G.E.P.; Cox, D.R. An analysis of transformations. Journal of the Royal Statistical Society, B 1964, 26, 211-234.
Figure 1.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of doxorubicin. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with doxorubicin alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was determined using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ## and ### indicate p < 0.01 and p < 0.001, respectively, DOX-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, DOX– doxorubicin, IPA – indolepropionic acid, IS – indoxylsulfate.
Figure 1.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of doxorubicin. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with doxorubicin alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was determined using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ## and ### indicate p < 0.01 and p < 0.001, respectively, DOX-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, DOX– doxorubicin, IPA – indolepropionic acid, IS – indoxylsulfate.

Figure 2.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of gemcitabine. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with gemcitabine alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (three parameters)” utility) was performed on datasets to obtain IC50 values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values using the Shapiro-Wilk test. Dataset normality was achieved by the Box-Cox normalization method. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 values non-paired, two-sided t-test was applied. ## and ### indicate p < 0.01 and p < 0.001, respectively, GEM-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, GEM – gemcitabine, IPA – indolepropionic acid, IS – indoxylsulfate.
Figure 2.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of gemcitabine. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with gemcitabine alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (three parameters)” utility) was performed on datasets to obtain IC50 values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values using the Shapiro-Wilk test. Dataset normality was achieved by the Box-Cox normalization method. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 values non-paired, two-sided t-test was applied. ## and ### indicate p < 0.01 and p < 0.001, respectively, GEM-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, GEM – gemcitabine, IPA – indolepropionic acid, IS – indoxylsulfate.
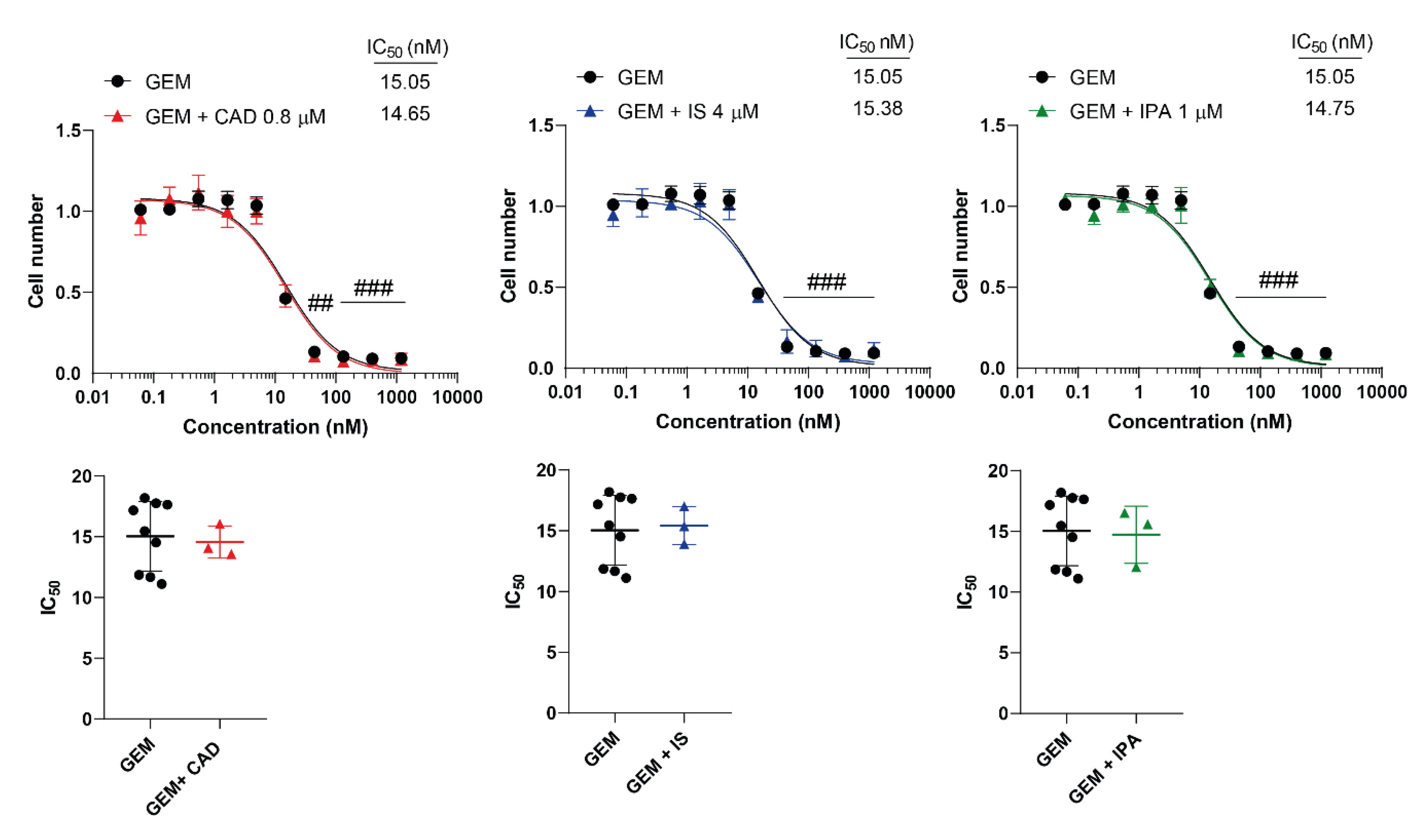
Figure 3.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of irinotecan. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with irinotecan alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ## and ### indicate p < 0.01 and p < 0.001, respectively, IRI-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IRI – irinotecan, IS – indoxylsulfate.
Figure 3.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of irinotecan. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with irinotecan alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ## and ### indicate p < 0.01 and p < 0.001, respectively, IRI-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IRI – irinotecan, IS – indoxylsulfate.
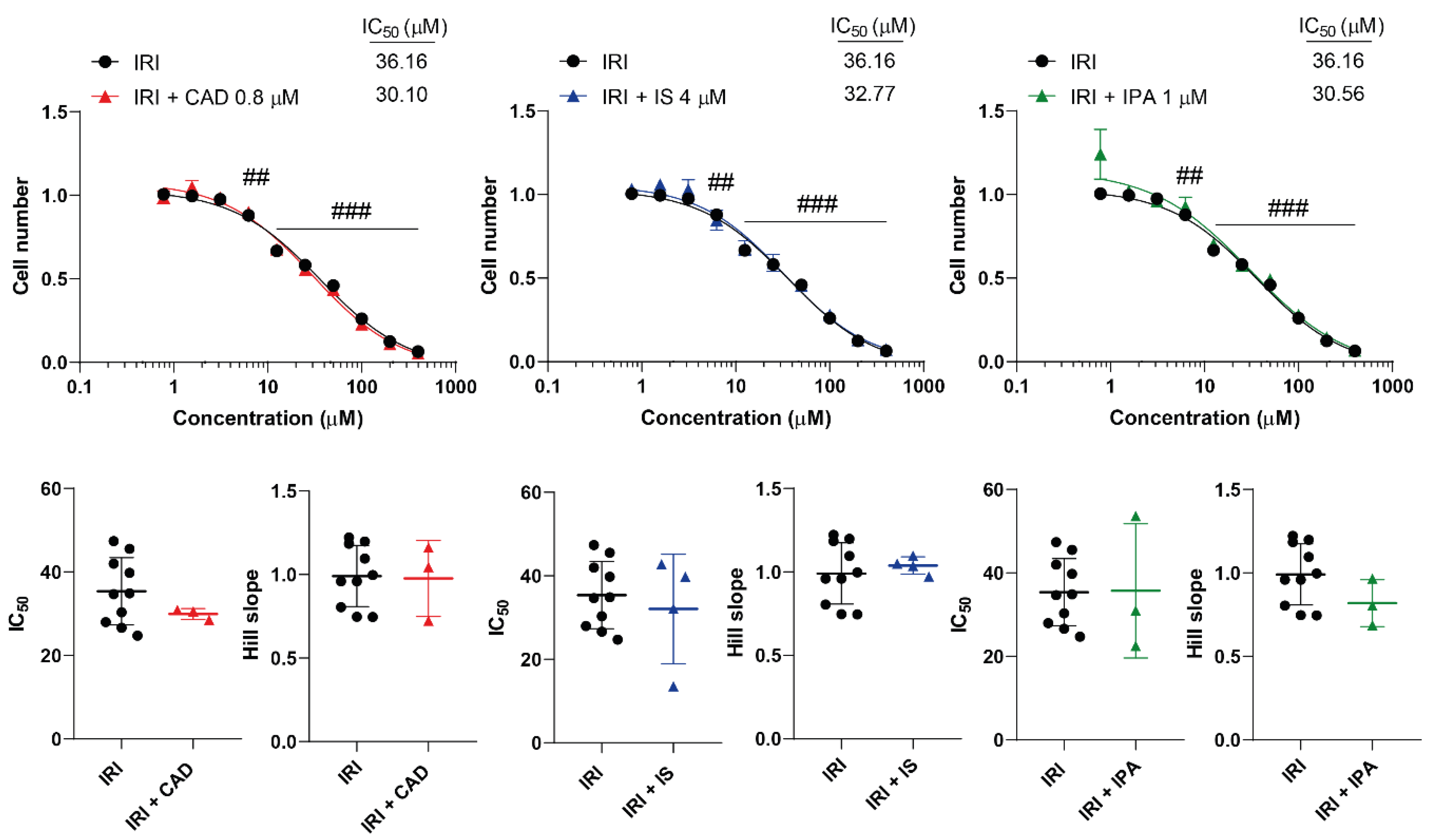
Figure 4.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of methotrexate. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with methotrexate alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ### indicate p < 0.001, respectively, MTX-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IS – indoxylsulfate, MTX – methotrexate.
Figure 4.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of methotrexate. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with methotrexate alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ### indicate p < 0.001, respectively, MTX-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IS – indoxylsulfate, MTX – methotrexate.
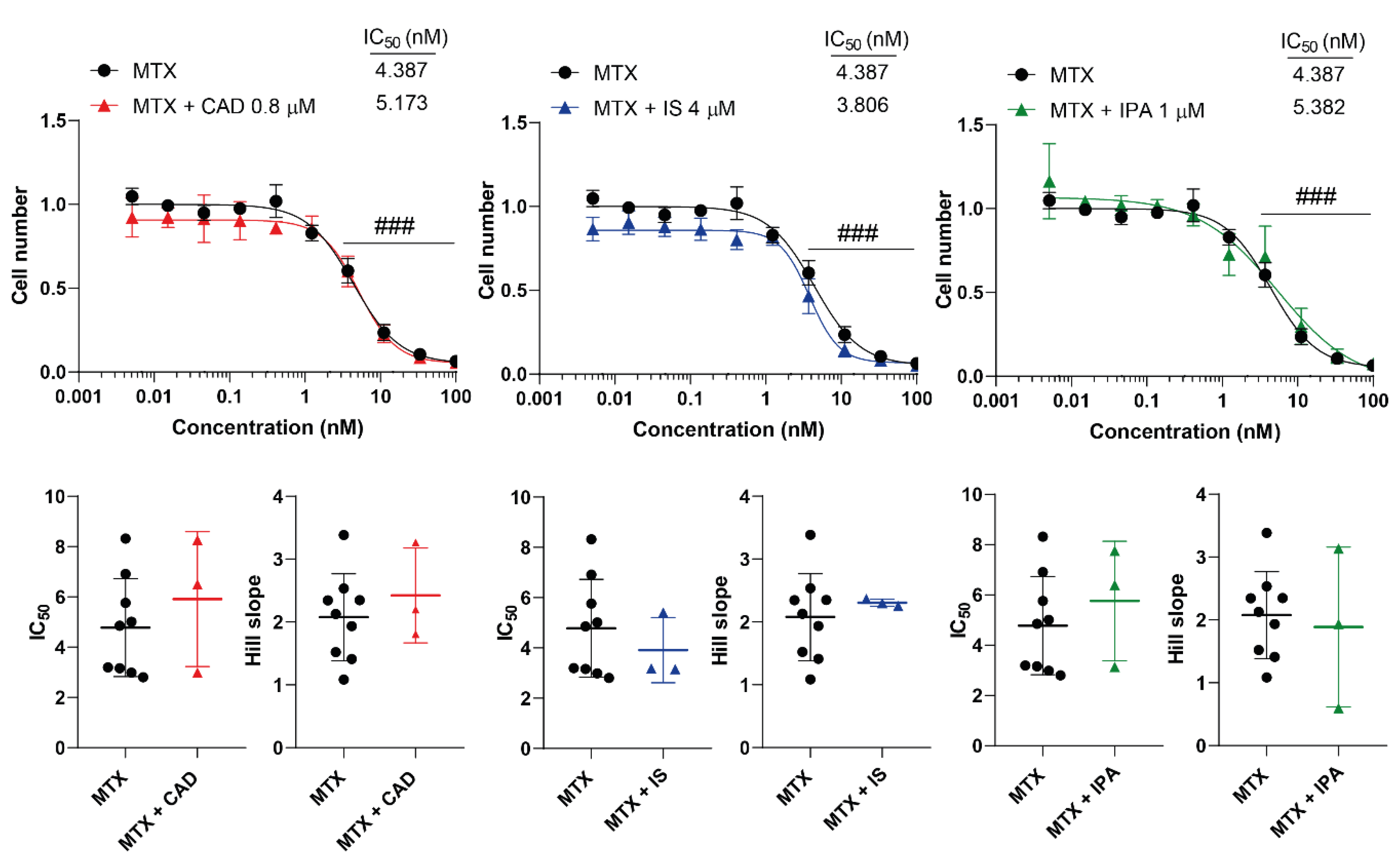
Figure 5.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of rucaparib. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with rucaparib alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ## and ### indicate p < 0.01 and p < 0.001, respectively, RUCA-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IS – indoxylsulfate, RUCA – rucaparib.
Figure 5.
Cytostatic bacterial metabolites do not interfere with the cytostatic effect of rucaparib. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with rucaparib alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ## and ### indicate p < 0.01 and p < 0.001, respectively, RUCA-treated vs. vehicle-treated cells. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IS – indoxylsulfate, RUCA – rucaparib.
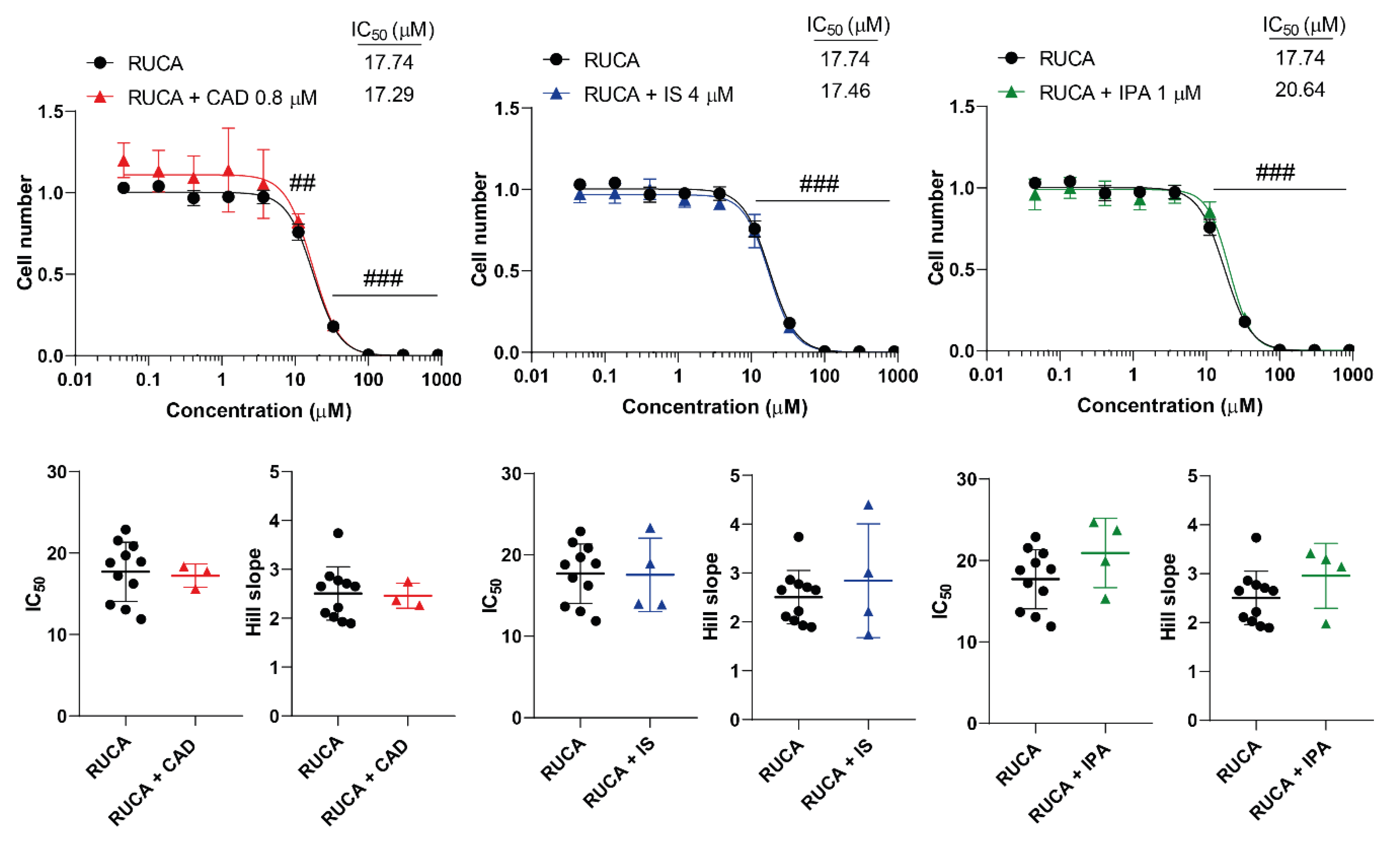
Figure 6.
Cadaverine and indolepropionic acid interfere with the cytostatic effects of 5-fluorouracil. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with 5-fluorouracil alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. To achieve normal distribution, datasets were log-normalized. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. #, ##, and ### indicate p < 0.05, p < 0.01, and p < 0.001, respectively, 5FU-treated vs. vehicle-treated cells. * represents significance at p < 0.05 between the indicated groups. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IS – indoxylsulfate, 5FU – 5-fluorouracil.
Figure 6.
Cadaverine and indolepropionic acid interfere with the cytostatic effects of 5-fluorouracil. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with 5-fluorouracil alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. To achieve normal distribution, datasets were log-normalized. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. #, ##, and ### indicate p < 0.05, p < 0.01, and p < 0.001, respectively, 5FU-treated vs. vehicle-treated cells. * represents significance at p < 0.05 between the indicated groups. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IS – indoxylsulfate, 5FU – 5-fluorouracil.
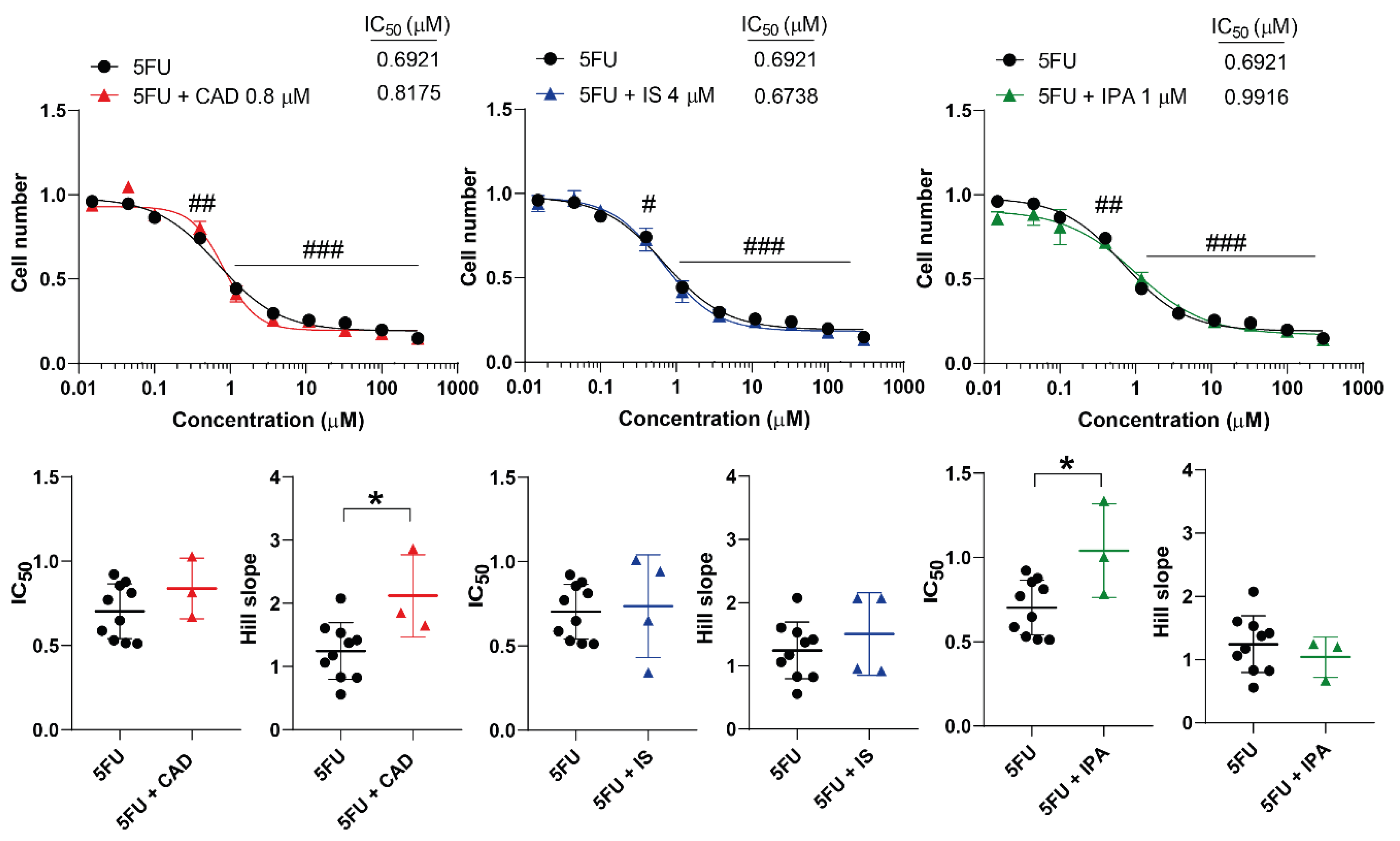
Figure 7.
Indolpropionic acid improves the cytostatic effect of paclitaxel. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with paclitaxel alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ### indicate p < 0.001, respectively, 5FU-treated vs. non-treated cells. ** represents significance at p < 0.01 between the indicated groups. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IS – indoxylsulfate, PAC - paclitaxel.
Figure 7.
Indolpropionic acid improves the cytostatic effect of paclitaxel. 4T1 cells were plated in 96-well plates (1500 cells/well). Cells were treated with paclitaxel alone or in combination with CAD (0.8 µM), IS (4 µM) or IPA (1 µM) for 48 hours, then cell numbers were determined by MTT assay. Data are presented as means ± SEM, from at least three biological replicates. Individual assays were measured in quadruplicate or in triplicate. Values were normalized to vehicle-treated cells (absorbance is equal to 1). Nonlinear regression (Graphpad “[Inhibitor] vs. response (four parameters)” utility) was performed on datasets to obtain IC50 and Hill slope values. Normality was determined for the inhibitory curves using the D’Agostino and Pearson normality test, while for the IC50 values or the Hill slope values using the Shapiro-Wilk test. Statistical difference between the inhibitory curves was performed using a two-way ANOVA test, and all data points were compared with each other (in Tukey post hoc tests). For the comparison of the IC50 and Hill slope values non-paired, two-sided t-test was applied. ### indicate p < 0.001, respectively, 5FU-treated vs. non-treated cells. ** represents significance at p < 0.01 between the indicated groups. Abbreviations: CAD – cadaverine, IPA – indolepropionic acid, IS – indoxylsulfate, PAC - paclitaxel.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



