
Submitted:
05 May 2024
Posted:
06 May 2024
You are already at the latest version
Abstract
Structural and functional asymmetries are traceable in every form of life and, at all cases, are a phenomenon of homology. Functionally speaking, the division of labour between the two halves of the brain is a basic characteristic of the nervous system that arose even before the appearance of vertebrates. The most prominent consequence of this specialization in humans and animals is handedness. Even if handedness is far more commonly associated with the presence of a nervous system, it is also observed in aneural organism such as plants. To date, little is known regarding the possible functional significance of handedness in plants, and many questions remain open (among these, whether it reflects a generalized behavioural asymmetry). Here, we propose a comparative perspective to the study of handedness, including plants, by taking advantage of the experimental models and paradigms already used to study laterality in humans and various animal species. By taking this approach, we aim to enrich our knowledge on the concept of handedness across natural kingdoms.
Keywords:
Handedness
; climbing plants
; lateralization
; asymmetry
; chirality
1. The Asymmetric Nature of the Universe
“Life as manifested to us is a function of the asymmetry of the universe
and of the consequences of this fact”
Pasteur, 1860
Nature is asymmetric at all levels. From plants to human beings, all organisms present a structure that is not identical to its mirrored image. Variation in the anatomical structure (i.e. position and orientation of organs) across and among species (e.g. plants and animals) is often described in relation to the primary organism’s axes [1]. Animals, for instance, exhibit frontally rather than laterally placed eyes and therefore divide their perceptual world according to coordinates such as right–left, top–bottom, and front–back. They present three whole-body axes as follows (Figure 1A, B): (i) the anterior–posterior (A–P) axis, which extends longitudinally from head to tail; (ii) the dorsal–ventral (D–V) axis in which ventral typically faces toward and dorsal away from a substrate; and (iii) the left–right (L–R) axis, which is defined in relation to a plane running along the anterior-posterior midline. In plants, the axes are related to the direction of growth of the organismal parts (e.g. stem, roots).
The main axis of plants’ growth is the apical–basal (A-B), which represents a single straight line from both the origin of the plant to the tip of the shoot or the roots (Figure 1C) [2,3]. Its direction (i.e. polarization) depends on various environmental cues (e.g. gravity; Hudson, 2000 [2,4]), and it remains active throughout plant development by defining the growth direction of both the stem and the roots [5,6]. The A–B axis is related only to the simplest growth forms of the plant. However, both the stem and the root system present their own relative elongation, which is characterized by a rhythmic repetition of leaves or secondary roots, respectively, at different distances. For instance, along the stem, leaves are added asymmetrically in the form of a single leaf primordium (i.e. groups of cells that will form into new leaves) to one side of the growing point as an investment around the stem. In this case, symmetry can also be observed in the leaves, which are characterized by several axes such as the proximal–distal (i.e. distance from the base to the apex; Figure 2A), the dorsal–ventral (i.e. distance from the adaxial to the abaxial side; Figure 2B), and the medial–lateral axis (i.e. distance from the midvein to the margin; Figure 2C). In plants, “proximal” and “distal” should be applied to organ parts that do not develop from an apical meristem (e.g. leaflets, petals; Figure 2A) or to organs with an apical meristem that branch from another site (e.g. branches or lateral roots). In adaxial–abaxial axis, the adaxial part represents the top of the leaf and abaxial the bottom of the leaf (Figure 2B). Then, the medial–lateral axis is used to describe laminar plant parts, such as many leaves and petals or some shoot axes (e.g. cactus paddles), that expand through the growth of marginal meristems (Figure 2C) [7].
Furthermore, symmetry can be observed in several plant organs (e.g. leaves, roots, shoots). It can be (i) radial (i.e. similar parts are arranged in a balanced way around the centre of the plant’s body; Figure 3A), such as the development of petals in flowers, roots, and shoots; (ii) dorsiventral (i.e. the front and back parts of the plant’s organs; Figure 3B) such as leaves; (iii) bilateral (i.e. the left and right parts of the plant’s body are symmetrical; Figure 3C), as in the flowers of pea plants; and (iv) helical/spiral (i.e. there is a central vertical axis around which the different plant’s organs, such as the stem, twist, either towards or away; Figure 3D), as in the arrangement of leaves along the stem (i.e. phyllotaxy) [8].
Asymmetries represent a way to adapt to the predominance of various environmental conditions. In animals such as human beings, functional asymmetries can be observed in brain asymmetries, such as the localization of the language centre in one hemisphere or the position of the organs in the human body (i.e. the heart is on the left, the liver is on the right side, and the small and large intestines coil in a chiral manner). On the other hand, in plants, the arrangement of leaves as a helix/spiral along the stem is necessary to balance the mass of the leaves along the stem and to provide an optimal structure for maximizing light capture, efficient gas exchange, and protection from excessive damage by insects, wind, or the sun [8,9,10].
The most relevant behavioural manifestation of lateralization is handedness (i.e. the consistent use of one effector rather than the other in performing certain tasks). In some cases, the meaning of handedness is clear as for hand dominance in human beings. In other cases, when bodies are characterized by irregular shapes (e.g. plants), the designation of handedness is more critical. At this point, the reader may wonder how handedness can be used to study lateralization in brainless organisms such as plants. Plants do not have a nervous system and do not develop along the same axes as animals do. However, as animals and plants co-evolved, they heavily interacted, and it is not surprising that several features of both groups of organisms are subjected to selection pressures such as symmetry vs. asymmetry. This opens up the possibility of a comparative taxonomic and evolutionary study of handedness focused on animals and plants. Naturally, we cannot equate brain asymmetries with those exhibited by plants. However, in terms of directional movement patterns, some factors (i.e. right-handed prevalence) can be compared across species. In the following paragraphs, we introduce the phenomenon of handedness in animals and plants. We then examine the empirical studies on handedness in separate animal species. This will lead us to propose a comparative perspective to the study of handedness in plants by taking advantage of the experimental models and paradigms already used to study laterality in various animal species. Finally, to determine the basis for valid cross-species comparisons, we highlight those factors that, from our perspective, should be considered in future research.
2. Handedness: One Term, Multiple Meanings
The concept of handedness (laterality or lateralization; hereafter, we shall use both terms interchangeably) includes the idea that an organism uses one effector (i.e. hand) more frequently than the other and the related idea that performance is more skilled or efficient with the preferred effector [1]. Handedness is commonly associated with the different specializations of function that are related to the left and right hemispheres [11]. Back in the day, there was the idea that directional asymmetry was unique to Homo sapiens given that right-handedness was linked to the emergence of language [12]. As commonly known, this applies to a majority of humans exhibiting a left-hemispheric dominance for the control of speech [13]. Nowadays, we are aware that lateralization is a characteristic that defines a wide range of species and that there was no discontinuity in its evolution.
Handedness is by far the most studied behavioural asymmetry in humans; in over 50 years of research, it has been tackled with consideration for genetic [14,15], behavioural [16], and environmental aspects [17]. Handedness can be distinguished into two forms: left- and right-handedness. Almost 90% of humans are right-handed, and this percentage holds consistently across cultures [14,18]. Handedness varies in strength (i.e. individuals who may be weakly or strongly lateralized) and direction (i.e. left or right) within and across species. Handedness is often task dependent (e.g. writing, reach-to-grasp, reach-to-eat), which makes it difficult to identify the factors driving the expression of handedness [19,20,21]. Furthermore, handedness can be observed at the population level (i.e. when most individuals within the same population show the same bias for the right or the left) or at the individual level (i.e. a consistent preference for a single individual irrespective of the population). In this context, it is not surprising that the first attempts to measure lateralization in animals used handedness as a parameter, as it is the most relevant behavioural manifestation of lateralization.
However, the extreme preference documented in our species for the right hand is apparently unmatched in other animal species. A recent review in non-human primates indicates, indeed, that population-level handedness is rare aside from our species [22]. Nevertheless, evidence of hand or limb population-level asymmetries has been documented in a host of vertebrates, including fish, rodents, birds, and anurans [23], as well as some species of prosimians [24]. Moreover, evidence of the predominance of right-handedness in captive chimpanzees for specific complex tasks (e.g. bimanual feeding) has suggested a continuity between humans and our closest relative [19]. An extensive literature in animal species [23] has indicated how strongly brain asymmetries affect everyday behaviour in terms of handedness, regardless of the presence of hands or limbs (e.g. fish and/or reptiles). In fish, behavioural asymmetries include a preferential ventral fin use for blue gourami, Trichogaster trichopterus [25], and lateralized pectoral spine stridulation in catfish, Ictalurus punctatus [26]. Snakes exhibit a lateralized use of a hemipenis according to different temperatures [27], whereas Roth [28], while observing coiling posture in cottonmouth snakes (used for defensive and offensive strikes directed at predators and prey), found a preference for counter-clockwise coiling. The above evidence suggests that the study of handedness has already been freed from the presence of limbs. Also, it allows researchers to extend the concept of handedness to organisms with no specialized functions such as plants.
3. Handedness in Plants
Many types of plants exhibit a helical structure with a preferred direction (e.g. honeysuckle grows as a left-handed helix). As previously reported, asymmetries represent a way to adapt to the prevailing environmental conditions at any time. In plants, a clear example of adaptability is represented by the helical growth (i.e. circumnutation) of both the stem and related extensions (e.g. tendrils, leaves) [29,30]. This movement involves many different behavioural patterns such as (i) the arrangement of the leaves or petals, (ii) the twisting of the flat plant’s organs to increase their rigidity, (iii) exploratory movements of the stem or tendrils (i.e. filamentary organs sensitive to contact and used exclusively for climbing), and (iv) the coiling of tendrils upon the potential support in the environment. For instance, climbing plants adopt the helical pattern of movement to find a potential support (e.g. wooden pole, tree trunk) in the environment and, once perceived, to direct their organs such as tendrils (i.e. modified leaves that are used by a climbing plant to climb a potential support) toward it for climbing [29,30,31]. Further, the direction of the tendrils’ helical growth determines the side of the coiling movement upon the support, which could be clockwise (i.e. on the left side), counter-clockwise (i.e. on the right side) or mixed (i.e. both directions; Figure 4A) [31,32,33,34,35,36,37]. This direction could be fixed within species or flexible between organs of the same plants. However, the majority of plants show a preference for twisting in the right or left direction, exhibiting a sort of “handedness” (i.e. the property exhibited by two forms of structures that are mirror images of each other) [37].
Handedness in plants is arbitrarily determined by the perspective of the viewer. This leads to discrepancies in the definition of right- and left-handed plants. In the past, some botanists such as Hugo von Mohl defined right-handed plants as those that climb a support “with the sun” or “like the clock hands” [34,36]. Whereas other botanists such as Charles Darwin claimed that when the circle made by the finger is clockwise, the spiral of the plant is right-handed [30]. On the contrary, when the circle is counter-clockwise, it is left-handed [34,36]. Kihara [36] adopted the terminology that described “clockwise” as “right-handed” and counter-clockwise as “left-handed.” Hashimoto [39] suggested defining it as “left-handed” clockwise helical growth. Indeed, clockwise arrangements of petals and leaves are correlated with left-handed helical epidermal cell files, and anti-clockwise arrangements are correlated with right-handed epidermal helix. To avoid misunderstandings, here we use the Hashimoto [39] definition: namely, everything that turns clockwise and moves away from the observer is considered left-handed (Figure 4A). However, counter-clockwise movement is right-handed (Figure 4A).
Examples of handedness in plants include (i), first, the spiral arrangement of leaves along the stem (i.e. phyllotaxy). For instance, the coconut tree (Cocos nucifera) shows a clockwise (i.e. left-handed) or counter-clockwise direction (i.e. right-handed) of phyllotaxy depending on the hemispheric locations. Indeed, right-handed forms are predominantly present in the Northern Hemisphere, with left-handed forms being present in the Southern Hemisphere [38,40,41]. (ii) Second is the helical growth of different organs of the plants (e.g. the stem, leaves, petals). The turn of the shoot of Arabidopsis (Arabidopsis thaliana L.) and the Ipomea purpurea (Ipomea purpurea Roth) plants is usually clockwise [42], whereas shoots of beans (Phaseolus vulgaris L.) and Fallopia baldschuanica (Polygonum baldschuanicum Regel) display a counter-clockwise direction [43]. (iii) Third is the coiling of the tendrils or stem in climbing plants. Evidence reported that 92% of climbers show a right-handed coil-direction preference independently of the hemispheric locations and latitude [32]. However, some plants such as pea plants (Pisum sativum L. cv, Alaska) show a mixed pattern of coiling direction (i.e. clockwise and/or counter-clockwise direction). Jaffe [35] observed that the prevalence of the tendril’s rotation in pea plants was predominantly clockwise (i.e. 53%) compared to counter-clockwise (i.e. 47%). Recent studies have confirmed these findings, showing that the direction of the circumnutation in pea plants during the approach-to-clasp movement towards a potential support (i.e. a wooden pole or another plant) can be either clockwise or counter-clockwise and that it can change within the same plant [44,45,46].
Given that the spiral-growth direction is fixed in given species, it has advanced that handedness in plants is genetically determined [47]. At the molecular and cellular level, the direction of the helical movement is linked with the arrangement of the arrays of the microtubules (i.e. hollow tubes made of alpha and beta tubulin that are a part of the cell’s cytoskeleton) in the cellular cortex and the related coiling of the cellulose microfibrils around the cell. Further, a range of microtubule-associated proteins (MAPs) are involved in helical growth development such as the SPIRAL1 (i.e. SKU6), SPIRAL2 (i.e. TORTIFOLIA1) and the gene GCP2. The right or left direction depends on the structural properties of the microtubule arrays. For instance, right-handed organ twisting is always associated with left-handed microtubule arrays and vice versa. In sum, both genetic and cellular investigation have demonstrated a strong link between microtubule arrangement and the potential twisting of cells and tissues [37].
4. Toward a Comparative Study of Handedness in Animals and Plants
An increasing number of studies have suggested that plant and animal behaviours show strong similarities (e.g. decision-making, learning) and that a neural architecture is not ubiquitously necessary to support certain abilities [44,45,46,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63]. However, how can we trace a territory in which handedness for animals and plants can coexist and, more importantly, be revealed?
Even though handedness in primates is task dependent, the situation is far more complex in plants. As reported previously, a critical point is represented by the fact that handedness in plants is determined by the observer’s perspective; therefore, it is quite difficult to unequivocally define plants as right- or left-handed [34,36,39]. Another critical aspect is how and when to measure handedness. Indeed, the spiral arrangement, the helical growth, and the coiling of tendrils or the stem are all used as indicators of handedness, and to our knowledge there is no agreement on what the most reliable measure may be or on the correlation between the various measurement. This led us to a further consideration: which phases of plant growth are most indicative to collect information on handedness? Given that all of the indicators are useful, what moment is the most critical to consider?
Having said that, we found evidence of what can be partially described as handedness at the population level in plants. Ninety-two percent of climbing plants appear to be right-handed considering the coiling direction [32], whereas this percentage drops at almost the chance level (53%) for some other plants [35]. It is rather unclear to what extent these two outcomes are comparable and to what extent they reflect an actual asymmetry at the population level. Do the authors estimate handedness at similar phases? Do the authors share the same criteria to assess handedness? Is it possible to hypothesize a scenario in which to investigate the variability of handedness in plants? The answer is, potentially yes but with great caution. Given the fact that handedness in plants can be measured in terms of direction (right vs. left) and, perhaps, in terms of extent (e.g. the timing in which the tendrils coil around the support), only systematic studies could clarify whether there is a direction of growth and climbing common to the majority of plants observed in the same conditions with a shared methodology. There is a second critical aspect that might be difficult to address as far as handedness in plants is concerned. In humans and other animal species, handedness is often measured through repeated observations. With plants, this is obviously difficult to achieve given that, on many occasions, the moment at which plants climb a support represents a non-repeatable event. A possible strategy to overcome this obstacle can be the study of plants that achieve their objective through a form of trial-and-error approach. For instance, Guerra and collaborators [45] investigated the growing movement of the apex and the tendrils from the germination of the seed until the clasping of the support. Results revealed that as the number of leaves progresses, the velocity and the time taken to perform a circumnutation increase. Also, this corresponds with a decrease in the number of circumnutation and “handedness” switches. In light of this, at the population level, even if there is a similar proportion of plants with clockwise, counter-clockwise, and mixed directions of circumnutation movement, single plants might exhibit a preference that could be revealed through the acquisition of repeated measures.
Furthermore, how can we measure the consistency of handedness at the individual level? In humans and other animal species, handedness is often task dependent. For example, in humans, manual preference is extremely strong if measured in a writing task but is greatly reduced as the task used to assess it varies. Ideally, handedness should be measured in different contexts and for as many functions as possible. It could be very informative to investigate handedness as a task-dependent function in plants. For instance, it might be of interest to assess how the handedness of climbers varies on the basis of various tasks (e.g. clasping a support) and/or the goal (e.g. competition or cooperation with other plants). In other words, investigating handedness at the individual level is a key factor in estimating the consistency of lateralization across various tasks.
However, as previously reported, the above approaches limit the categorization of the plant as right- or left-handed at the time they have completed their growth process. We are practically blind to the range of adjustments that precedes, for instance, the climbing of a support and that can enhance the analysis of lateralized behaviour in plants.
To uncover the nature of handedness in plants, we propose to characterize the movement of plants by means of the three-dimensional (3D) kinematical analysis. This methodology permits researchers to define the time course of changes in the position and orientation of the body or its part (e.g. effector) in terms of trajectories, velocities, and accelerations. In human beings and various animal species, this approach has been applied in studying the handedness of goal-directed motor programmes such as the reach-to-grasp movement at different stages of development. Evidence has shown that right-hand preference is already observed in the foetus during hand-to-face movements [64]. Both right- and left-handed foetuses were faster in reaching targets requiring greater precision (i.e. eye and mouth) with their dominant (vs. non-dominant) hand. Hand preference was also observed in young infants for grasping objects (i.e. toys or block), in which movements showed straighter and smoother paths of the dominant effector and a shorter movement time compared to the non-dominant one [65,66,67,68,69,70]. Similar findings have been observed in adults, whose movements performed with the dominant hand were generally faster, whereas grasping movements with the non-dominant hand presented a wider safety margin. In other words, movements performed with the non-dominant hand showed a longer movement time and more correction adjustments compared to movements with the dominant hand to possibly compensate for a higher endpoint variability (i.e. variability of finger positions upon the object). These findings have also been confirmed in non-human primates [71,72]. Evidence has reported that chimpanzees who used a precision grip to grasp small pieces of food were more likely to use their right hand and that this hand preference may reflect a property of the brain that is ancient and hardwired [71,72].
Recently, the kinematical approach has been extended to the study of goal-directed movement in plants. In specific, the approaching and clasping movements performed by different organs of a pea plant towards a potential support have been characterized [44,45,46,48,49,50,51,52,53,54,55,63,73]. These findings have demonstrated that pea plants are able to perceive an element in the environment and to modulate the kinematics of their movement in terms of velocity, acceleration, aperture of their tendrils, and smoothness on the basis of the feature of the to-be-grasped support [45,51,52,53,54,55], the task (e.g. decision-making) [46,63], and the context (e.g. competition or cooperation) [44,48]. The tracking and analysing of plants’ movement through time and space using dedicated in-house software [73] represent a potential tool for studying the distribution and functional significance of laterality in plants. Also, many of the kinematical features can be considered in a lateralized fashion. Figure 5 depicts the possible kinematical features to quantify specific properties of handedness in plants.
All of these measures might be applied to investigate laterality in plants with specific reference to climbing plants (e.g. peas and/or beans). Climbers represent an ideal model that, through their development, can somewhat represent the vegetal side of manual preference. For instance, pea plants are annual climbing plants that need to find a potential support in the environment in order to climb and to reach the light source, which is necessary for its survival. Their leaves are arranged asymmetrically along the stem, and each leaf has tendrils and filamentary organs that allow the plant to clasp an external support (Figure 6).
The morphological structure of pea plants will allow us to assess different aspects of handedness, such as whether (i) the clasping upon the support occurs predominantly in a clockwise or counter-clockwise direction; (ii) plants have a preference for climbing a support with leaves developed on the right or the left side of it and whether this preference could be observed in terms of a difference in kinematical patterning (e.g. velocity profiles, movement time, number of switch’s direction) and the smoothness of the movement (e.g. number of submovements, end-point variability). In this case, because plants do not have right and left sides, the R–L axis in plants should be determined by the observer's perspective (Figure 6); and (iii) the movement of right- or left-handed plants changes on the basis of the position of the support (e.g. on the left or right side with respect to the plant).
In sum, this approach will allow researchers to include plants in the “comparative” debate by capitalizing on paradigms and ideas already used to study laterality among organisms belonging to various species.
5. Conclusions
In the present review, we examined the question of laterality across kingdoms. What may emerge from our work is that, even though animals and plants are different organisms with different structures, they exhibit similarities in terms of directional movement patterns (i.e. right-handed prevalence), opening up the possibility of a comparative study of handedness across taxa. However, research on plants’ handedness is still in its early stages, and the existing literature is few and often controversial. Here, we suggest a comparative approach to the study of handedness in plants by adopting the experimental paradigms already used to study laterality in various animal species. The use of critical and specific methods would contribute to identifying, defining, and evaluating the functional aspects of handedness in plants as widely documented in various animal species. As knowledge about plant behaviour expands, the similarities between plant and animal behaviours are becoming increasingly evident. What may emerge from the study of handedness in plants and animals is the realization that they complement each other nicely and, if nothing else, once again demonstrate just how similar all free-living organisms are to one another.
Author Contributions
Conceptualization: M.D. S.G.; Writing - original draft: M.D. S.G.; Writing - review & editing: U.C. M.D. S.G. and B.B. All authors read and approved the final version of the manuscript.
Funding
This work has been funded by the European Union (ERC, ROOMors, Grant Number 101096728) to UC. However, the views and opinions expressed are those of the author(s) only and do not necessarily reflect those of the European Union or the European Research Council Executive Agency. Neither the European Union nor the granting authority can be held responsible. S.G. is supported by ERC grant (ERC, ROOMors, Grant Number 101096728) to UC. B.B. is supported by a MUR grant (no. 20227ZYLH9) to UC.
Data Availability Statement
Not applicable.
Acknowledgments
Figures 1–6 were created on BioRender.com.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hegstrom, R.A.; Kondepudi, D.K. The handedness of the universe. Sci. Am. 1990, 262, 108–115. [Google Scholar] [CrossRef]
- Hudson, A. Development of symmetry in plants. Annu. Rev. Plant Biol. 2000, 51, 349–370. [Google Scholar] [CrossRef]
- Jürgens, G. Axis formation in plant embryogenesis: cues and clues. Cell 1995, 81, 467–470. [Google Scholar] [CrossRef]
- Jeong, S.; Eilbert, E.; Bolbol, A.; Lukowitz, W. Going mainstream: How is the body axis of plants first initiated in the embryo? Dev. Biol. 2016, 419, 78–84. [Google Scholar] [CrossRef]
- Berger, F.; Brownlee, C. Establishment of the apical-basal axis in multicellular plant embryos. Biol. Cell. 1995, 84, 7–11. [Google Scholar] [CrossRef]
- Sachs, T. Cell polarity and tissue patterning in plants. Dev. 1991, 113, 83–93. [Google Scholar] [CrossRef]
- Dahdul, W.M.; Cui. H.; Mabee. P.M.; Mungall. C.J.; Osumi–Sutherland. D.; Walls. R.L.; Haendel, M.A. Nose to tail, roots to shoots: spatial descriptors for phenotypic diversity in the Biological Spatial Ontology. J. Biomed. Semant. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Hofmeister, W. AllgemeineMorphologie der Gewächse. Handbuch der physiologischen Botanik 1868, 1, 405–664. [Google Scholar]
- Braybrook, S.A.; Kuhlemeier, C. How a plant builds leaves. The Plant Cell. 2010, 22, 1006–1018. [Google Scholar] [CrossRef]
- Endress, P.K.; Doyle, J.A. Floral phyllotaxis in basal angiosperms: development and evolution. Curr. Opin. Plant Biol. 2007, 10, 52–57. [Google Scholar] [CrossRef]
- Warren, J.M. Handedness and laterality in humans and other animals. Physiol. Psychol. 1980, 8, 351–359. [Google Scholar] [CrossRef]
- Michel, G.F.; Babik, I.; Nelson, E.L.; Campbell, J.M.; Marcinowski, E.C. How the development of handedness could contribute to the development of language. Dev. Psychobiol. 2013, 55, 608–620. [Google Scholar] [CrossRef]
- Knecht, S.; Dräger, B.; Deppe, M.; Bobe, L.; Lohmann, H.; Flöel, A. Handedness and hemispheric language dominance in healthy humans. Brain 2000, 123, 2512–2518. [Google Scholar] [CrossRef]
- Annett, J. Motor Learning: A Review. In: Heuer, H. Kleinbeck, U. Schmidt, KH. (eds) Motor Behavior. Springer, Berlin, Heidelberg, 1985. [CrossRef]
- McManus, I.C. Handedness, language dominance and aphasia: a genetic model. Psychol. Med. Monogr. Suppl. 1985, 8, 3–40. [Google Scholar] [CrossRef]
- Rat-Fischer, L.; O'Regan, J.K.; Fagard, J. Handedness in infants' tool use. Dev. Psychobiol. 2013, 55, 860–868. [Google Scholar] [CrossRef]
- Domellöf, E.; Johansson, A.M.; Rönnqvist, L. Handedness in preterm born children: a systematic review and a meta–analysis. Neuropsychologia 2011, 49, 2299–2310. [Google Scholar] [CrossRef]
- Raymond, M.; Pontier, D. Is there geographical variation in human handedness? Laterality 2004, 9, 35–51. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Russell, J.L.; Lambeth, S.; Schapiro, S.J. Handedness and neuroanatomical asymmetries in captive chimpanzees: a summary of 15 years of research. Int. J. Primatol. 2007, 5, 146–181. [Google Scholar] [CrossRef]
- Crow, T. Directional asymmetry is the key to the origin of modern Homo sapiens (the Broca-Annett axiom): A reply to Rogers' review of The Speciation of Modern Homo Sapiens. Laterality: asymmetries of body, brain and cognition, 2004, 9, 233–242. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Cantalupo, C. Handedness in chimpanzees (Pan troglodytes) is associated with asymmetries of the primary motor cortex but not with homologous language areas. Behav. Neurosci. 2004, 118, 1176. [Google Scholar] [CrossRef]
- Caspar, K.R.; Pallasdies, F.; Mader, L.; Sartorelli, H.; Begall, S. The evolution and biological correlates of hand preferences in anthropoid primates. Elife 2022, 11, e77875. [Google Scholar] [CrossRef]
- Vallortigara, G.; Rogers, L. Survival with an asymmetrical brain: advantages and disadvantages of cerebral lateralization. BBS, 2005. [CrossRef]
- Watson, S.L.; Hanbury, D.B. Prosimian primates as models of laterality. Primatol. 2007, 5, 228–250. [Google Scholar] [CrossRef]
- Bisazza, A.; Sovrano, V.A.; Vallortigara, G. Consistency among different tasks of left–right asymmetries in lines of fish originally selected for opposite direction of lateralization in a detour task. Neuropsychologia 2001, 39, 1077–1085. [Google Scholar] [CrossRef]
- Fine, M.L.; Friel, J.P.; McElroy, D.; King, C.B.; Loesser, K.E.; Newton, S. Pectoral spine locking and sound production in the channel catfish Ictalurus punctatus. Copeia 1994, 777–790. [Google Scholar] [CrossRef]
- Shine, R.; Olsson, M.M.; LeMaster, M.P.; Moore, I.T.; Mason, R.T. Are snakes right–handed? Asymmetry in hemipenis size and usage in gartersnakes (Thamnophis sirtalis). Behav. Ecol. 2000, 11, 411–415. [Google Scholar] [CrossRef]
- Roth, E.D. ‘Handedness’ in snakes? Lateralization of coiling behaviour in a cottonmouth, Agkistrodon piscivorus leucostoma, population. Animal Behav. 2003, 66, 337–341. [Google Scholar] [CrossRef]
- Darwin, C.; Darwin, F. The power of movement in plants. John Murray, London. 1880. [CrossRef]
- Darwin, C. The movements and habits of climbing plants. John Murray, London. 1875.
- Gianoli, E. The behavioural ecology of climbing plants. AoB PLANTS 2015, 7, plv013. [Google Scholar] [CrossRef]
- Edwards, W.; Moles, A.T.; Franks, P. The global trend in plant twining direction. Glob. Ecol. Biogeogr. 2007, 16, 95–800. [Google Scholar] [CrossRef]
- Gardner, M. The ambidextrous universe. Penguin, Victoria, Australia. 1970.
- Jackson, D.B. A glossary of botanical terms. London. 4th ed. 1953. [Google Scholar]
- Jaffe, M.J. Physiological Studies on Pea Tendrils VIII. The Relationship of Circumnutation to Contact Coiling – With a Description of a Laboratory Intervalometer Using Integrated Digital Circuits. Physiol. Plant. 1972, 26, 73–80. [Google Scholar] [CrossRef]
- Kihara, H. Right–and left–handedness in plants–A review. ISO 690. 1973. [Google Scholar]
- Smyth, D.R. Helical growth in plant organs: mechanisms and significance. Dev. 2016, 143, 3272–3282. [Google Scholar] [CrossRef]
- Davis, T.A.; Davis, B. Association of coconut foliar spirality with latitude. Math. Model. 1987, 8, 730–733. [Google Scholar] [CrossRef]
- Hashimoto, T. Molecular genetic analysis of left–right handedness in plants. Philosophical Philos. Trans. R. Soc. 2002, 357, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Minorsky, P.V. Latitudinal differences in coconut foliar spiral direction: a re–evaluation and hypothesis. Ann. Bot. 1998, 82, 133–140. [Google Scholar] [CrossRef]
- Minorsky, P.V.; Bronstein, N.B. Natural experiments indicate that geomagnetic variations cause spatial and temporal variations in coconut palm asymmetry. Plant Physiol. 2006, 142, 40–44. [Google Scholar] [CrossRef]
- Schuster, J.; Engelmann, W. Circumnutations of Arabidopsis thaliana seedlings. Biol. Rhythm Res. 1997, 28, 422–440. [Google Scholar] [CrossRef]
- Millet, B.; Melin, D.; Badot, P.M. Circumnutation in Phaseolus vulgaris. I. Growth, osmotic potential and cell ultrastructure in the free-moving part of the shoot. Physiol. Plant. 1988, 72, 133–138. [Google Scholar] [CrossRef]
- Bonato, B.; Wang, Q. ; Guerra, S.; Simonetti, V.; Bulgheroni, M.; Quaggiotti, S.; Ruperti, B.; Castiello, U. ‘United we stand, divided we fall’: intertwining as evidence of joint actions in pea plants. AoB Plants 2024, 16, plad088. [Google Scholar] [CrossRef] [PubMed]
- Guerra, S. Bruno, G. Spoto, A. Panzeri, A. Wang, Q. Bonato, B. Simonetti, V. Castiello, U. Ascent and attachment in pea plants: a matter of iteration. Research square preprint. [CrossRef]
- Wang, Q.; Guerra, S.; Bonato, B.; Simonetti, V.; Bulgheroni, M.; Castiello, U. Decision–making underlying support searching in pea plants. Plants 2023, 12, 1597. [Google Scholar] [CrossRef] [PubMed]
- Silk, W.K.; Holbrook, N.M. The importance of frictional interactions in maintaining the stability of the twining habit. AJB. 2005, 92, 1820–1826. [Google Scholar] [CrossRef] [PubMed]
- Bonato, B.; Simonetti, V.; Bulgheroni, M.; Wang, Q.; Guerra, S.; Quaggiotti, S.; Ruperti, B.; Castiello, U. Evidence of motor intentions in plants: a kinematical study. J Comp Psycho. 2023. [Google Scholar] [CrossRef] [PubMed]
- Bonato, B.; Castiello, U.; Guerra, S.; Wang, Q. Motor cognition in plants: from thought to real experiments. Theor. Exp. Plant Physiol. 2024. [Google Scholar] [CrossRef]
- Castiello, U. (Re) claiming plants in comparative psychology. J. Comp. Psychol. 2021, 135, 127. [Google Scholar] [CrossRef]
- Ceccarini, F.; Guerra, S.; Peressotti, A.; Peressotti, F.; Bulgheroni, M.; Baccinelli, W.; Bonato, B.; Castiello, U. Speed–accuracy trade-off in plants. Psychon. Bull. Rev. 2020, 27, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Ceccarini, F.; Guerra, S.; Peressotti, A.; Peressotti, F.; Bulgheroni, M.; Baccinelli, W.; Bonato, B.; Castiello, U. On-line control of movement in plants. Biochem. Biophys. Res. Commun. 2020, 564, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Guerra, S.; Bonato, B.; Wang, Q.; Ceccarini, F.; Peressotti, A.; Peressotti, F.; Bacinelli, W.; Bulgheroni, M.; Castiello, U. The coding of object thickness in plants: when roots matter. J Comp Psycho. 2021, 135, 495–504. [Google Scholar] [CrossRef]
- Guerra, S.; Bonato, B.; Wang, Q.; Ceccarini, F.; Peressotti, A.; Peressotti, F.; Bacinelli, W.; Bulgheroni, M.; Castiello, U. Kinematic evidence of root–to–shoot signaling for the coding of support thickness in pea plants. Biology 2022, 11, 405. [Google Scholar] [CrossRef]
- Guerra, S.; Peressotti, A.; Peressotti, F.; Bulgheroni, M.; Bacinelli, W.; D’Amico, E.; Gómez, A.; Massaccesi, S.; Ceccarini, F.; Castiello, U. Flexible control of movement in plants. Sci Rep. 2019, 9, 16570. [Google Scholar] [CrossRef]
- Falik, O.; Reides, P.; Gersani, M.; Novoplansky, A. Root navigation by self inhibition. Plant, Cell Environ. 2005, 28, 562–569. [Google Scholar] [CrossRef]
- Gilroy, S.; Trewavas, T. Agency, teleonomy and signal transduction in plant systems. Biological J. Linn. Soc. 2023, 139, 514–529. [Google Scholar] [CrossRef]
- Novoplansky, A. Picking battles wisely: plant behaviour under competition. Plant, Cell Environ. 2009, 32, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Parise, A.G.; De Toledo, G.R.A.; de Carvalho Oliveira, T.F.; Souza, G.M.; Castiello, U, Gagliano, M. ; Marder, M. Do plants pay attention? A possible phenomenological–empirical approach. Prog. Biophys. Mol. Biol. 2022, 173, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Parise, A.G.; Marder, M. Extended plant cognition: a critical consideration of the concept. Theor. Exp. Plant Physiol. 2023. [Google Scholar] [CrossRef]
- Parise, A.G.; Reissig, G.N.; Basso, L.F.; Senko, L.G.S.; Oliveira, T.F.D.C.; De Toledo, G.R.A. Detection of different hosts from a distance alters the behaviour and bioelectrical activity of Cuscuta racemosa. Front. Plant Sci. 2021, 12, 594195. [Google Scholar] [CrossRef] [PubMed]
- Saito, K. A study on diameter-dependent support selection of the tendrils of Cayratia japonica. Sci. Rep. 2022, 12, 1–8. [Google Scholar] [CrossRef]
- Wang, Q.; Barbariol, T.; Susto, G.A.; Bonato, B.; Guerra, S.; Castiello, U. Classifying circumnutation in pea plants via supervised machine learning. Plants 2023, 12, 965. [Google Scholar] [CrossRef]
- Parma, V.; Brasselet, R.; Zoia, S.; Bulgheroni, M.; Castiello, U. The origin of human handedness and its role in pre-birth motor control. Sci. Rep. 2017, 7, 16804. [Google Scholar] [CrossRef]
- Cox, R.F.A.; Smitsman, W. Action planning in young children’s tool use. Developmental Science 2006, 9, 628–641. [Google Scholar] [CrossRef]
- Flindall, J. W.; Gonzalez, C. L. On the evolution of handedness: evidence for feeding biases. PLoS One 2013, 8, e78967. [Google Scholar] [CrossRef]
- Leconte, P.; Fagard, J. Which factors affect hand selection in children’s grasping in hemispace? Combined effects of task demand and motor dominance. Brain Cogn. 2006, 60, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Morange–Majoux, F.; Peze, A.; Bloch, H. Organisation of left and right hand movement in a prehension task: A longitudinal study from 20 to 32 weeks. Laterality: Asymmetries of Body, Brain Cogn. 2000, 5, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Rönnqvist, L.; Domellöf, E. Quantitative Assessment of Right and Left Reaching Movements in Infants: A Longitudinal Study from 6 to 36 Months. Dev. Psychobiol. 2006, 48, 444–459. [Google Scholar] [CrossRef] [PubMed]
- Sacrey, L.A.R.; Arnold, B.; Whishaw, I.Q.; Gonzalez, C. Precocious hand use preference in reach–to–eat behavior versus manual construction in 1– to 5–yearold children. Dev. Psychobiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Begliomini, C.; Nelini, C.; Caria, A.; Grodd, W.; Castiello, U. Cortical activations in humans grasp–related areas depend on hand used and handedness. PLoS ONE 2008, 3, e3388. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Cantalupo, C.; Wesley, M.J.; Hostetter, A.B.; Pilcher, D.L. Grip morphology and hand use in chimpanzees (Pan troglodytes): Evidence of a left hemisphere specialization in motor skill. J. Exp. Psychol. Gen. 2002, 131, 412. [Google Scholar] [CrossRef]
- Simonetti, V.; Bulgheroni, M.; Guerra, S.; Peressotti, A.; Peressotti, F.; Baccinelli, W.; Ceccarini, F.; Bonato, B.; Wang, Q.; Castiello, U. Can plants move like animals? A three–dimensional stereovision analysis of movement in plants. Animals 2021, 11, 1854. [Google Scholar] [CrossRef]
Figure 1.
Primary axes in animal and plants. In animals (A) and human beings (B), there are three main axes: the anterior–posterior (A–P) axis (from the head to the tail/feet), the dorsal–ventral (D-V) axis; the upper or back side of an organism), and the right–left (R–L) axis (defined in relation to a plane running along the anterior–posterior midline). In plants (C), the main axis is represented by the apical–basal (A–B) axis, which refers to the straight line from either the origin of the plant to the tip of the shoot or the roots.
Figure 1.
Primary axes in animal and plants. In animals (A) and human beings (B), there are three main axes: the anterior–posterior (A–P) axis (from the head to the tail/feet), the dorsal–ventral (D-V) axis; the upper or back side of an organism), and the right–left (R–L) axis (defined in relation to a plane running along the anterior–posterior midline). In plants (C), the main axis is represented by the apical–basal (A–B) axis, which refers to the straight line from either the origin of the plant to the tip of the shoot or the roots.
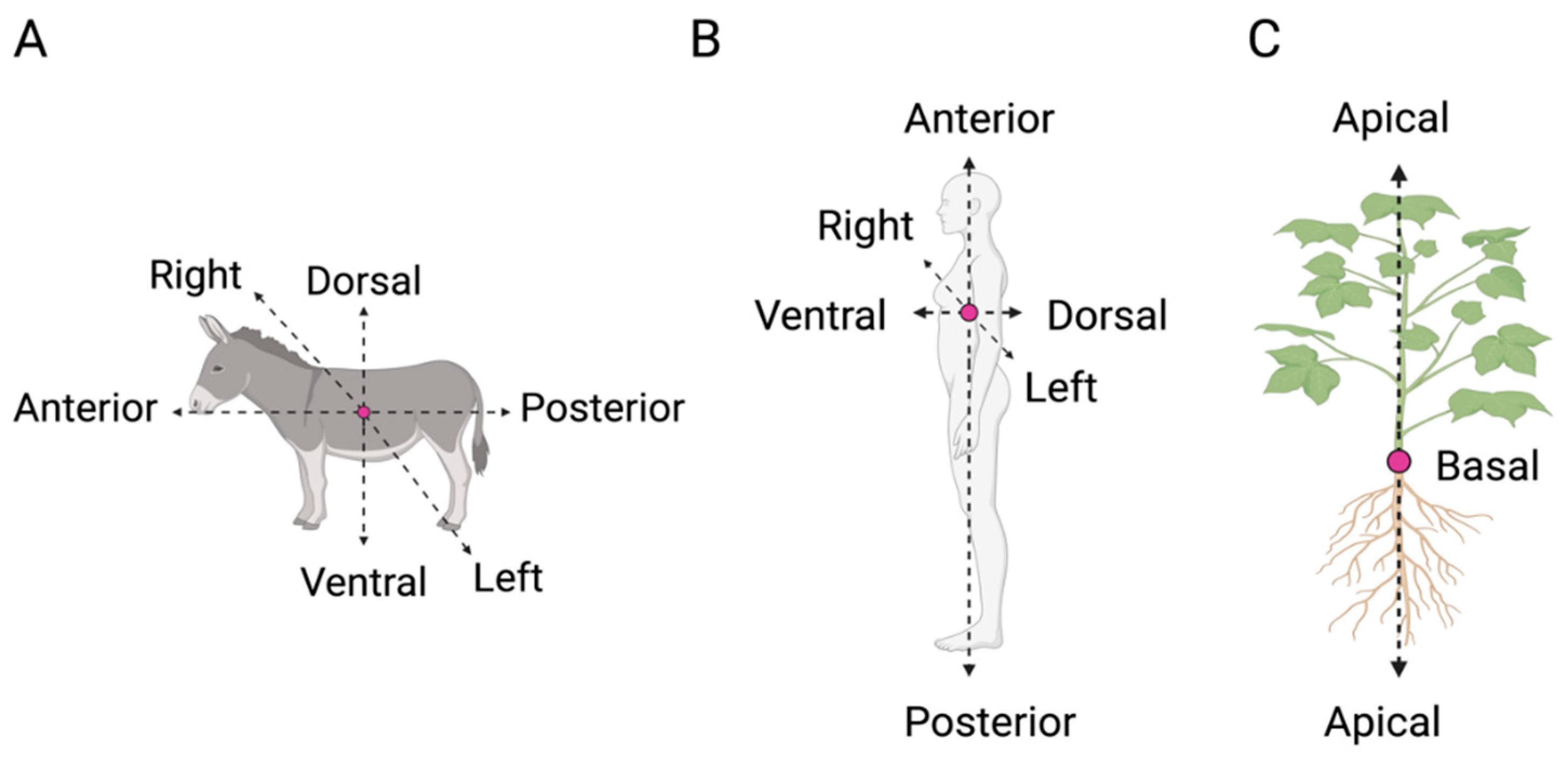
Figure 2.
Plants’ axes. Example of (A) proximal–distal, (B) dorsal–ventral, and (C) medial–lateral axes in the plants’ leaves.
Figure 2.
Plants’ axes. Example of (A) proximal–distal, (B) dorsal–ventral, and (C) medial–lateral axes in the plants’ leaves.
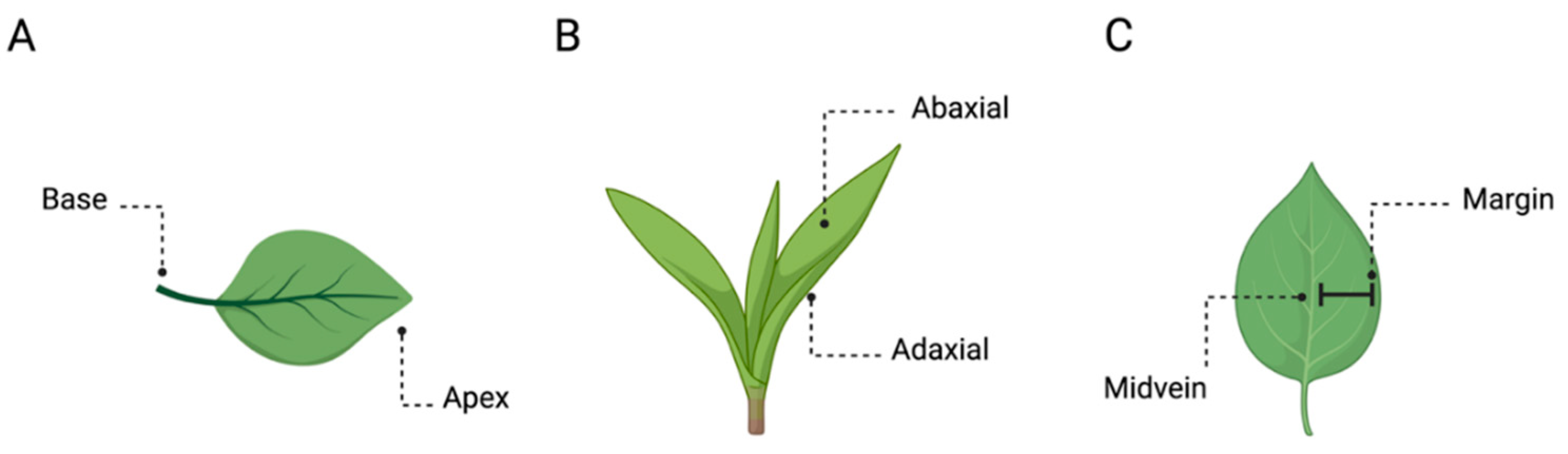
Figure 3.
Symmetry in plants. (A) Example of radial symmetry in flowers (e.g. wild roses); (B) dorsolateral symmetry (e.g. leaves); (C) bilateral symmetry (e.g. flower of a pea plant); and (D) helical/spiral symmetry (e.g. the arrangement of the leaves along the stem).
Figure 3.
Symmetry in plants. (A) Example of radial symmetry in flowers (e.g. wild roses); (B) dorsolateral symmetry (e.g. leaves); (C) bilateral symmetry (e.g. flower of a pea plant); and (D) helical/spiral symmetry (e.g. the arrangement of the leaves along the stem).
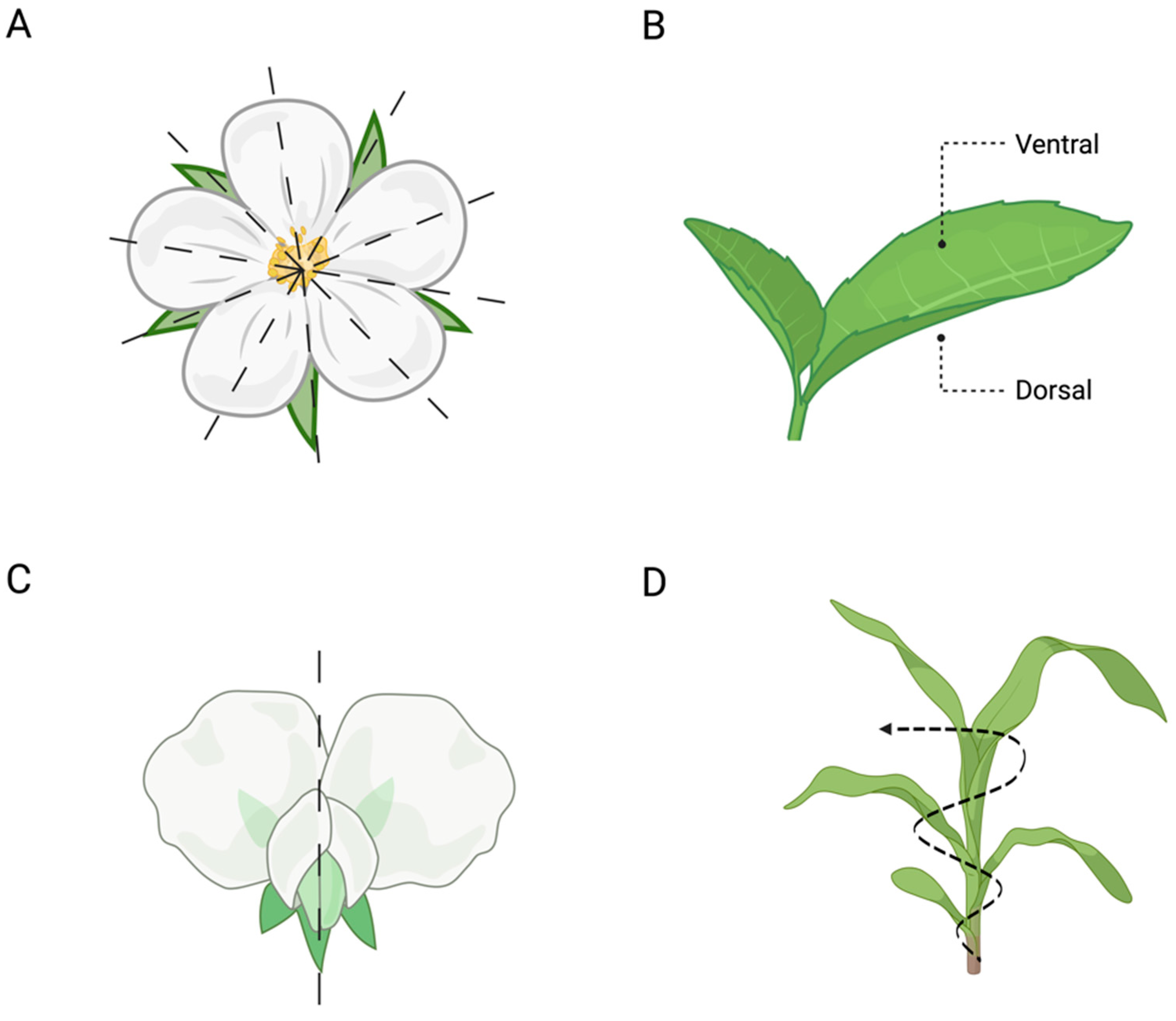
Figure 4.
(A) Graphical representation of the direction of circumnutating growing movement in plants, which could be: clockwise (i.e. left-handed), counter-clockwise (i.e. right-handed) or mixed. (B) Graphical representation of the direction of the coiling movement of the tendrils in pea plants.
Figure 4.
(A) Graphical representation of the direction of circumnutating growing movement in plants, which could be: clockwise (i.e. left-handed), counter-clockwise (i.e. right-handed) or mixed. (B) Graphical representation of the direction of the coiling movement of the tendrils in pea plants.
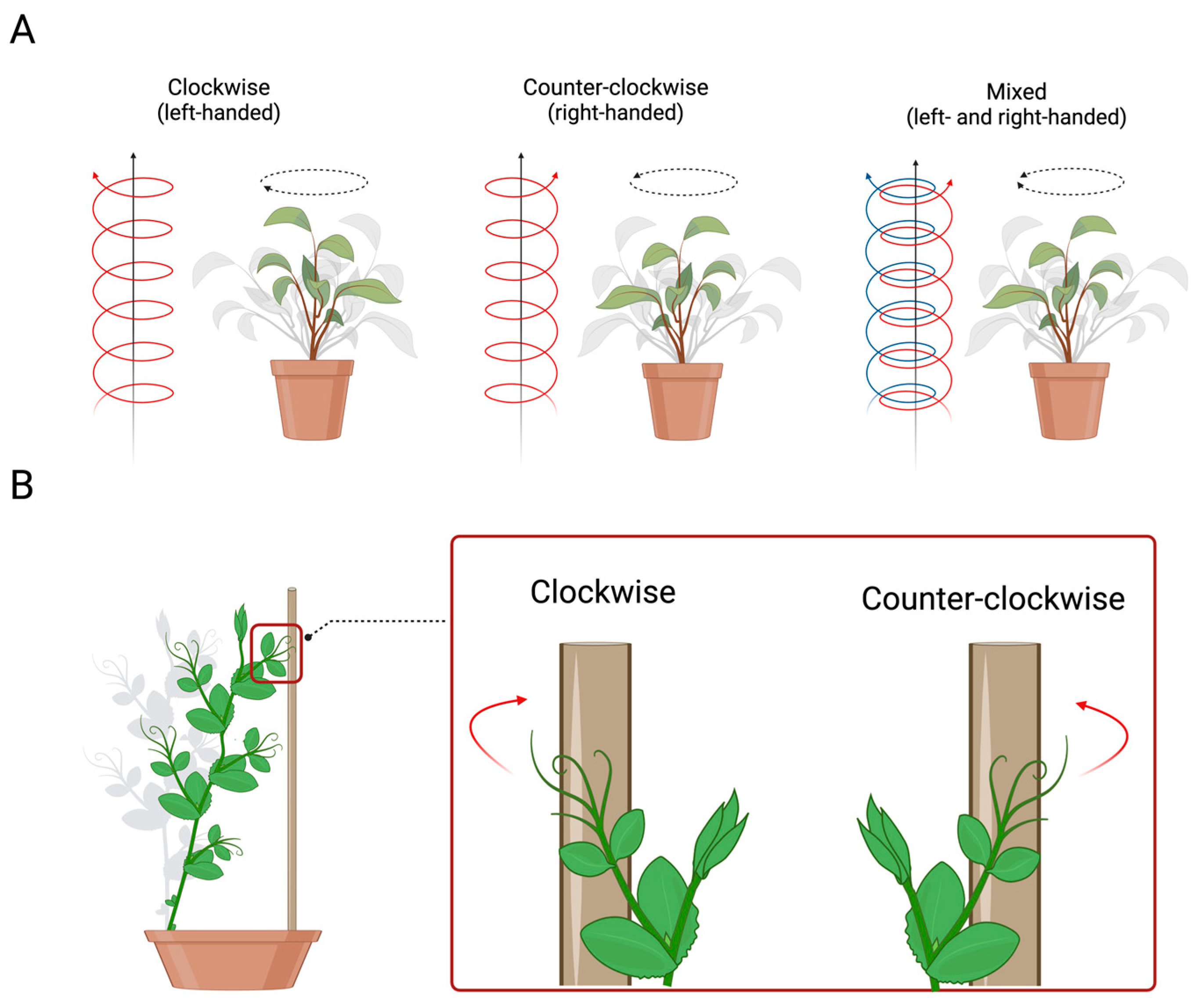
Figure 5.
Graphical representation of the kinematical measures to quantify the handedness in plants. (A) Graphical representation of the velocity profile of the tendrils of a pea plant during the approaching and clasping movement towards a potential support. Normalized movement time (%) refers to the time between the beginning and end of the movement. Acceleration/deceleration refers to the increasing and decreasing of the tendrils’ velocity before clasping the support. The peak of maximum velocity (%) refers to the maximum velocity reached by tendrils during the entire movement. Submovements refer to the corrective adjustments of the trajectories and velocity of the tendrils to execute a proper motor behaviour. (B) The path of the tendrils’ trajectories during the approaching and clasping of the support. (C) The end-point variability refers to the variability of the contact point of the tendrils upon the support. (D) Number of switch direction (i.e. clockwise or/and counter-clockwise) during circumnutation.
Figure 5.
Graphical representation of the kinematical measures to quantify the handedness in plants. (A) Graphical representation of the velocity profile of the tendrils of a pea plant during the approaching and clasping movement towards a potential support. Normalized movement time (%) refers to the time between the beginning and end of the movement. Acceleration/deceleration refers to the increasing and decreasing of the tendrils’ velocity before clasping the support. The peak of maximum velocity (%) refers to the maximum velocity reached by tendrils during the entire movement. Submovements refer to the corrective adjustments of the trajectories and velocity of the tendrils to execute a proper motor behaviour. (B) The path of the tendrils’ trajectories during the approaching and clasping of the support. (C) The end-point variability refers to the variability of the contact point of the tendrils upon the support. (D) Number of switch direction (i.e. clockwise or/and counter-clockwise) during circumnutation.
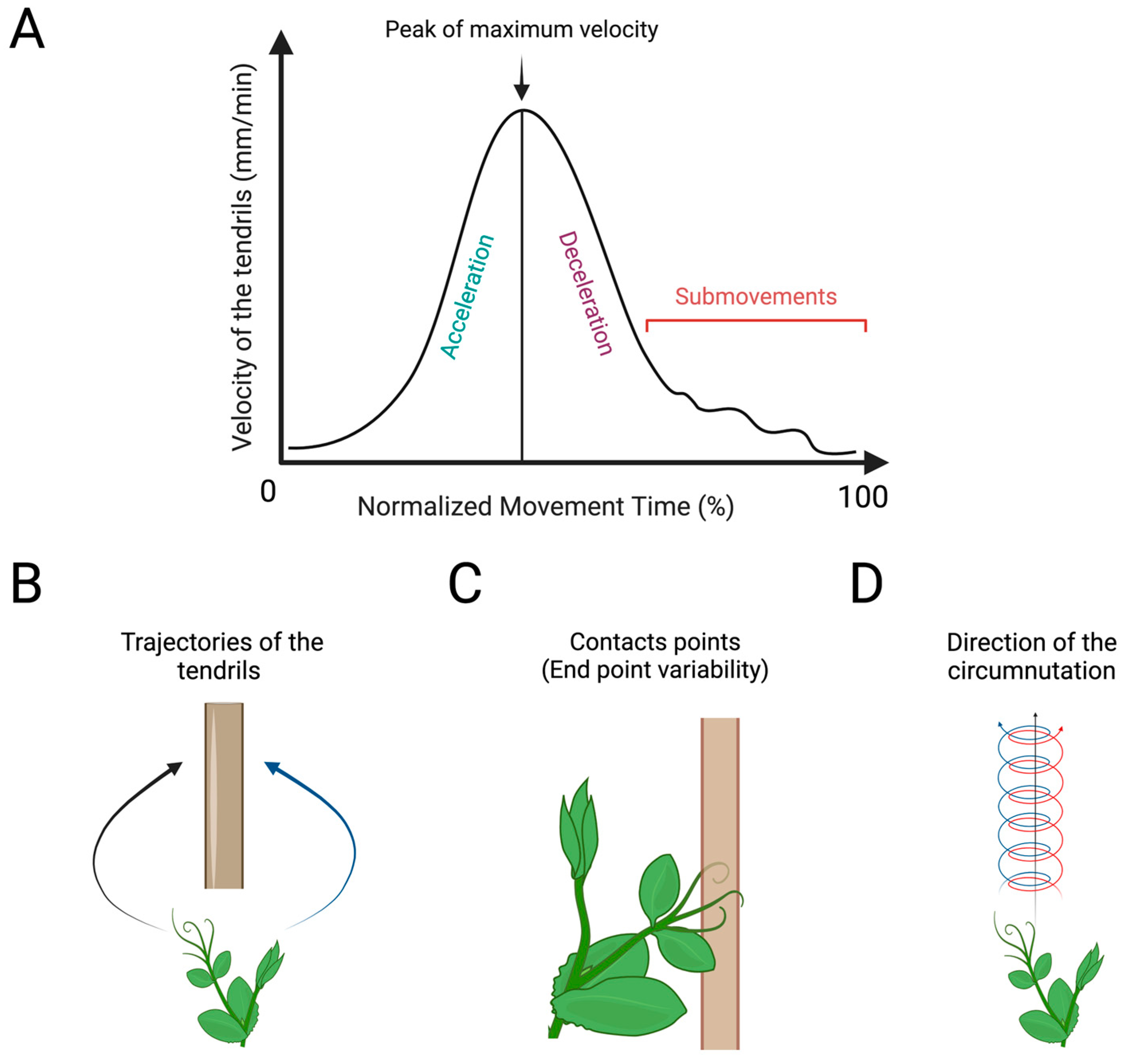
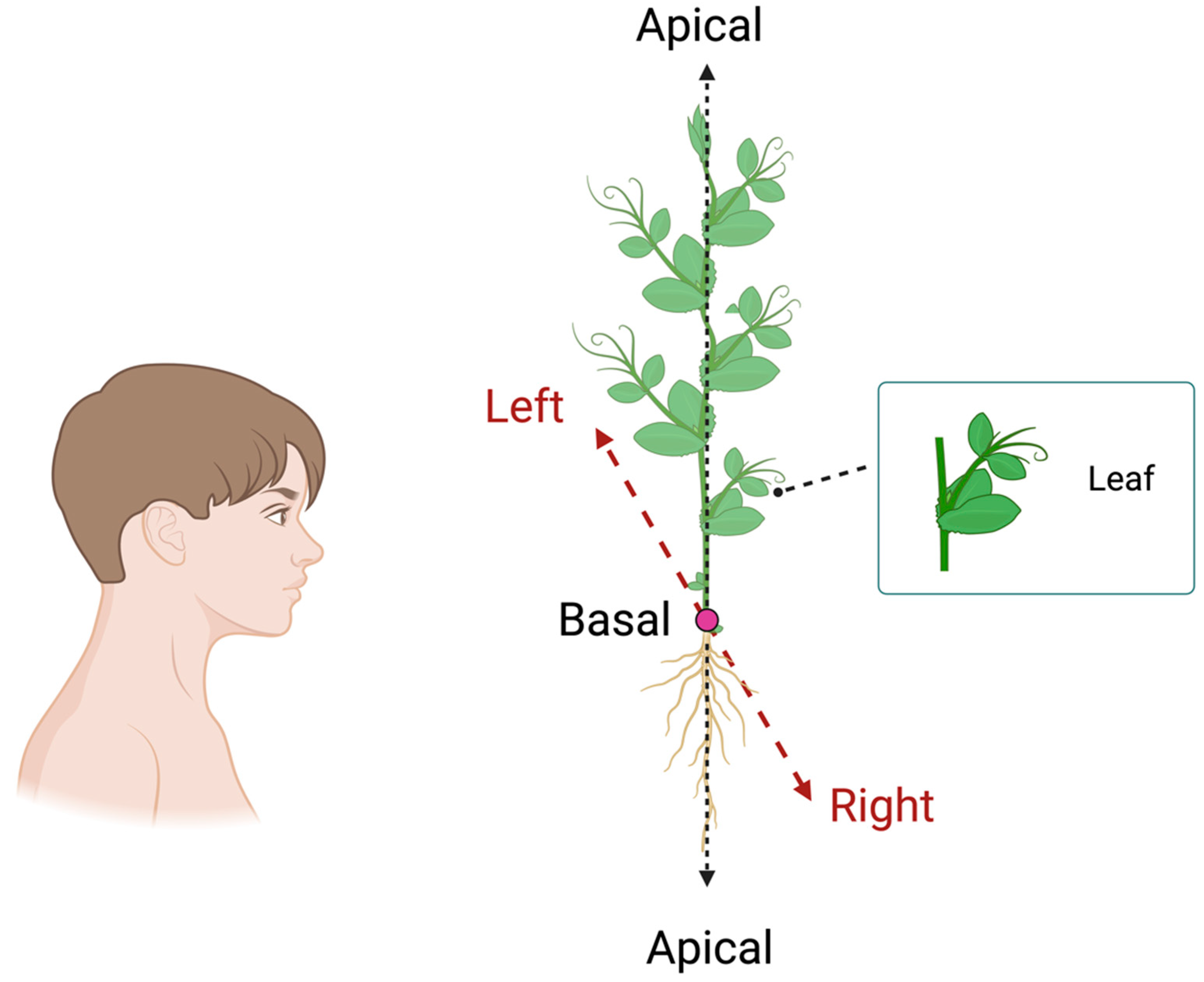
Figure 6.
Graphical representation of the morphological structure of the leaves of a pea plant. The black dashed lines refer to the apical–basal axis (A–B), whereas the red dashed lines refer to the right–left (R–L) axis, which is determined by the observer’s perspective.
Figure 6.
Graphical representation of the morphological structure of the leaves of a pea plant. The black dashed lines refer to the apical–basal axis (A–B), whereas the red dashed lines refer to the right–left (R–L) axis, which is determined by the observer’s perspective.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



