
Submitted:
02 May 2024
Posted:
06 May 2024
You are already at the latest version
Abstract
The chestnut tree (Castanea sativa Mill.) is gaining importance in the mountainous regions of southern Europe due to the high value of its fruits. It is essential to establish effective cultivation protocols, considering that this species is still relatively understudied. In this study, we present the outcomes of the initial establishment of a chestnut orchard conducted through a nutrient omission trial for four years. The treatments included a fertilization plan with nitrogen, phosphorus, potassium, and boron (NPKB), the control, and four other treatments corresponding to the omission of each nutrient (-NPKB, N-PKB, NP-KB, NPK-B). The -NPKB and NP-KB treatments showed significantly lower trunk circumferences and canopy volumes compared to the other treatments. The NPK-B treatment resulted in the lowest fruit production, with a total accumulated yield (2020-2022) of 0.56 kg tree–1, a value significantly lower than that of NPKB (1.12 kg tree–1) and N-PKB (1.19 kg tree–1). The assessment of nutrient concentrations in the leaves revealed plants with deficient levels of B and K in treatments that did not receive these nutrients. Conversely, N levels in the leaves in the -NPKB treatment fell within the sufficiency range (20 to 28 g kg–1). This suggests that the sufficiency range should be adjusted to higher values, given the treatment's effect on tree growth. It was also observed that the -NPKB treatment led to lower soil organic matter compared to the other treatments, likely due to reduced herbaceous vegetation development under the canopy, leading to decreased organic substrate deposition in the soil. The main findings of this study are that N and K were crucial elements for the optimal growth of chestnut trees, while B played a significant role in fruit production.
Keywords:
crop fertilization
; nutrient omission trial
; plant nutritional status
; sufficiency range
; soil organic matter
1. Introduction
Recent studies have highlighted the nutritional value of chestnuts, which is the main driver behind the increasing consumption [1]. The increase in market demand has also encouraged farmers to devote more attention to the cultivation of this species and led to an increase in the hectareage under chestnut (Castanea sativa Mill.) cultivation throughout the world. Over the last two decades (2002-2022), the global chestnut harvested area increased from 320,693 to 540,037 ha, while in Portugal, it rose from 30,614 to 49,940 ha [2].
In Portugal, farmers have also been intensifying cropping practices to increase crop productivity [3,4]. The use of fertilizers, for instance, has a high potential to improve crop yield, by replacing nutrients in the soil solution that are annually removed by the crops. The nutrients usually applied in almost all fertilization programs are the macronutrients nitrogen (N), phosphorus (P), and potassium (K), which are needed by plants in relatively large amounts, and soils are usually unable to make them available through natural processes at the rate at which they are removed by crops [5]. Additionally, in specific agro-environmental conditions, other essential nutrients can have high importance in the fertilization programs, even micronutrients. Boron (B) is probably the micronutrient that most frequently appears in deficiency in agricultural fields and that is regularly applied as a fertilizer [6,7,8].
Agricultural soils have a limited ability to supply N to crops, especially when they are poor in organic matter and have a low clay content [5,9]. Furthermore, N can be lost from the soil through various processes, including ammonia volatilization, nitrate leaching, and denitrification [5,10,11]. This makes N the nutrient to which crops most often respond to its application as a fertilizer [12,13,14] and the one associated with greater environmental damage. The common response of crops to the application of N and the environmental issues associated with the loss of this nutrient from the soil mean that the recommendation of this nutrient must be judicious and supported by field experimentation [15,16].
Along with N, the use of P in agriculture is one of the major concerns for crop fertilization in the 21st century. Although the loss of P from agricultural soils is not to be feared due to the lower solubility of the nutrient in the soil, the phosphate rocks from which phosphate fertilizers are made are running out, which raises serious problems for nutrient management in agriculture [17]. It is therefore important to know how to manage P fertilizers, applying them only when necessary and in the appropriate rates. While in certain world regions, crops tend to respond to the application of P [18,19], in others, this doesn't always occur. This is probably due to the higher natural availability of the element in the soil [20,21].
K is also a primary macronutrient with widespread use in agriculture. It is mainly used in NPK compound fertilizers [5]. The nutrient can enhance plant growth particularly under adverse environmental conditions, inducing tolerance to biotic and abiotic stresses [22,23]. Thus, studies reporting a positive effect of K application on crop growth and yield are not uncommon [24,25]. However, some soils have relatively high levels of K, which reduces crop response to the application of this nutrient. Khan et al. [26] reported the results of an extensive survey of more than 2,100 yield response trials, from which they concluded that plants rarely respond to K application. For this reason, despite the world's higher reserves of K compared to those of P [17], it is necessary to continue studying the response of plants to the application of the nutrient to rationalize its use.
B is a micronutrient of increased importance in dicot species, requiring higher amounts due to its functions in plasma membranes and cell walls compared to monocots [8,27]. On the other hand, in vast regions of the world, soils do not provide B to plants in adequate amounts, making it important to apply the nutrient as a fertilizer to maintain crop productivity [28,29,30]. In the north of Portugal, the importance of B in fertilization programs has been considered equivalent to that of macronutrient [30,31].
In the mountainous areas of Southern Europe, chestnut has gained increasing importance in recent decades. In some municipalities of Portugal, chestnut is currently the only catch crop, playing a decisive role in reducing the depopulation of rural areas. While some studies have already been carried out in mature orchards to try to understand how to fertilize these gigantic trees [32,33,34], few have been devoted to studying the response of young plants to fertilization. This early stage of the tree's life is also important as fertilizers can increase the rate of canopy growth and the speed at which the trees begin to bear fruit. Thus, this study reports the results of a nutrient omission trial conducted from planting grated plants to the fourth year of growth. It assessed the development of young trees, fruit production, nutritional status, and photosynthetic performance of plants. Additionally, the study examined the effect of continuous fertilization on soil fertility.
2. Materials and Methods
2.1. Experimental Conditions
The field experiment was carried out in Bragança, northeast of Portugal. The climate in the region is Csb type (temperate climate with dry and mild summer), registering an average annual temperature of 12.3 °C and total annual precipitation of 758.3 mm [35]. Meteorological records for the four years during which the field trial took place are presented in Figure 1.
The soil of the plot where the chestnut orchard was planted is a classified as Leptosol [36]. It originates from schist and has a sandy loam texture (soil separates are 623, 268 and 108 g kg–1 of sand, silt, and clay, respectively). The soil is acidic (pHH2O = 5.7) and poor in organic carbon (C) (0.92 g kg–1, Walkley-Black method). Extractable P and K levels (Egnér-Riehm method) were classified as low (21.5 mg kg–1 P2O5) and medium (73.4 mg kg–1 K2O), respectively, while extractable B (Azomethine-H) was found to be low (0.21 mg kg–1).
2.2. Experimental Design
The orchard was planted in the spring of 2018, with seedlings grafted the previous year, using the cultivar Judia as scion, and plants spaced 9 m × 9 m. The experiment was arranged as a nutrient omission trial. Five treatments were established, consisting of a basal fertilization plan with N, P, K and B, the four nutrients that are usually applied to dicot species in the region, and assigned in this study as the control treatment (NPKB). The other four treatments involved the removal of one of the mentioned nutrients from the basal fertilization plan, namely N (-NPKB), P (N-PKB), K (NP-KB) or B (NPK-B).
Each experimental unit consisted of four trees, replicated three times, resulting in a total of 12 trees per treatment. N, P, K and B were applied as ammonium nitrate (20.5% N, 50% NH4+, 50% NO3–), single superphosphate (18%, P2O5), potassium chloride (60% K2O) and borax (11% B). Increasing rates of nutrients were used over the years to meet the growing trees' increasing needs. In 2018, N, P and K were applied at a rate of 60 g per tree of N, P2O5, and K2O. Additionally, in the same year, B was applied at a rate of 1 g per tree. In subsequent years, the applied rates were based on those of the first year, with the amounts multiplied by a factor of 1.2. In the last year of the study, in 2022, the rates of nutrients applied per tree were 124 g of N, P2O5 and K2O and 2 g of B. Fertilizers were spread manually around the trees, initially in a circle with a 0.5 m radius, which was later extended to a 1 m radius following canopy growth. The fertilizers were incorporated into the soil with a cultivator.
2.3. Management of the Field Trial
During the summer following planting, the trees were watered to reduce the risk of mortality. Every two weeks, 10 L of water were applied per tree in circular furrows made around the trunk. In the following years, the trees were kept under rainfed conditions as is common in the region. Weed control was performed with two cultivator passes in the spring: one at the end of March, which also served to incorporate fertilizers into the soil, and another in mid-May. During the summer, from June to late August, two annual foliar sprays were applied to protect the young plants from fungal diseases. The commercial product Maestro® WG [25% (w/w) folpet and 50% (w/w) fosetyl-aluminum] was applied at a concentration of 300 g hL–1 and a rate of 3 kg ha–1 and Tocsin® WG [70% (w/w) thiophanate-methyl] applied at concentration and rate of 100 g hL–1 and 1 kg ha–1, respectively. At the end of June, light annual pruning was carried out, removing some basal branches, and ensuring a spherical shape of the tree canopy.
In October 2021 and 2022, trunk circumference and crown volume were measured. Trunk circumference was measured at a height of 40 cm, using a flexible tape measure adjustable to the trunk. The canopy volume (V) was calculated by measuring the average radius (r) at four horizontal positions around the trunk (North, South, East, and West) and halfway between the basal branches and the top of the canopy. The calculation followed a spherical canopy model, where V = 4/3 πr³.
From 2020, when the trees began to bear the first fruits, nut yield was also assessed. In the chestnut tree, the harvest is conducted by waiting for the fruits to fall to the ground, after which they are picked up and weighed. In this trial, we conducted three pickings during the natural fall period of the fruits to minimize the risk of them being taken by wild animals.
2.4. Soil and Leaf Sampling and Laboratory Analysis
Annually, at the end of July, leaves were collected for elemental chemical analysis. Mature leaves with the blade fully expanded were collected in the middle third of the branches of the year. The leaves were oven dried at 70 ºC and ground (1 mm mesh). In the elemental chemical analysis, N, P, K, calcium (Ca), magnesium (Mg), B, iron (Fe), copper (Cu), zinc (Zn) and manganese (Mn) were determined. Elemental analyzes of tissue samples were performed by Kjeldahl (N), colorimetry (B and P) and atomic absorption spectrophotometry (K, Ca, Mg, Fe, Mn, Cu, Zn) methods [37] after tissue samples have been digested with nitric acid in a microwave.
In November 2022, soil samples were collected at a depth of 0-0.20 m in the area close to the trunk where fertilizers were applied. Soil samples were oven-dried at 40 °C and sieved (2 mm mesh). They were analysed for pH (H2O and KCl) (soil: solution, 1:2.5), cation-exchange capacity (ammonium acetate, pH 7.0), organic C (wet digestion, Walkley-Black method) and extractable P and K (Egner-Riehm method, ammonium lactate extract). Soil B was extracted by hot-water and determined by the method of azomethine-H. These analytical procedures are described in detail in Van Reeuwijk [38]. The availability of other micronutrients (Cu, Fe, Zn, and Mn) in the soil was determined by atomic absorption spectrometry after extraction with diethylenetriaminepentaacetic acid (DTPA) buffered at pH 7.3, following the standard procedure of FAO [39]. Soil inorganic-N was determined in soil extracts prepared from 20 g of soil and 40 ml 2 M KCl. The suspension was shaken for 1 h and filtered through Watmann No. 42 filter paper. Nitrate and ammonium concentrations in the extracts were analysed in an UV–Vis spectrophotometer [40].
The soil samples were also used in a pot experiment to assess the bioavailability of nutrients in the soil. Pots of 3 kg of soil were sown with 30 ryegrass seeds and kept growing from December 4, 2022, to May 2023, during which two biomass cuts were performed. The pots were kept free of weeds and watered regularly. In addition to the evaluation of dry matter yield (DMY), the tissue samples were submitted to elemental chemical analysis, using the analytical methods mentioned above.
2.5. Data Analysis
The data was analysed for normality and homogeneity of variance using the Shapiro-Wilk and Levene’s test, respectively. The analysis of variance was performed as a one-way ANOVA, using the Statistical Package for the Social Sciences (SPSS) version 25 (IBM Corporation, New York, NY, USA). When significant differences were found, the means were separated by the Tukey HSD post hoc test (α = 0.05).
3. Results
3.1. Tree Growth and Fruit Yield
The measurements of trunk circumference resulted in the grouping of treatments into two categories, showing significant differences between them (see Figure 2). Trees in treatments without N (-NPKB) or K (NP-KB) exhibited thinner trunks compared to the others.
The canopy volume exhibited a similar response pattern to the trunk circumference, with treatments organized into the two response groups mentioned earlier and showing significant differences between them (Figure 3). However, in this case, the differences were much more pronounced.
Fruit production commenced in the third year after planting (2020). Figure 4 displays the outcomes of the initial harvests in 2020, 2021, and 2022. The treatments exhibited three response groups, where NPKB and N-PKB treatments resulted in the highest average accumulated fruit yields (1.12 and 1.19 kg tree–1, respectively). They were followed by -NPKB and NP-KB treatments (0.88 and 0.92 kg tree–1, respectively), and finally, the NPK-B treatment (0.56 kg tree–1). However, mean values within the first group did not significantly differ from those within the second, and those within the second did not differ significantly from those within the third. Significant differences were observed only between the treatments in the first group and the NPK-B treatment. The main difference between the latter result and that of trunk circumference and canopy volume is that the NPK-B treatment did not appear in the group with the highest values, appearing with the lowest average value among all treatments and with significant differences for the NPKB and N-PKB treatments.
3.2. Leaf Nutrient Concentration
Treatments significantly influenced leaf N concentration on all sampling dates (Figure 5). The treatment without N (-NPKB) consistently exhibited the lowest average values in all samplings. Generally, treatments that received N showed higher levels of the nutrient in leaves, often reaching or exceeding the upper limit of the sufficiency range. In contrast, the treatment without N resulted in the lowest values, although still above the lower limit of the sufficiency range.
Leaf P concentration varied significantly between treatments only in the first sampling date (Figure 5). Throughout all sampling dates, leaf P levels remained within the sufficiency range for all five treatments, including the one that did not receive P.
Leaf K concentrations varied significantly between treatments in all sampling dates (Figure 5). The treatment that did not receive K consistently displayed lower average values. When compared to the sufficiency range, average values were often found below the lower limit, particularly in the case of the NP-KB treatment.
Leaf B levels varied significantly between treatments across the four sampling dates (Figure 5). The treatment without B resulted in plants with very low levels of the nutrient in the leaves, falling below the lower limit of the sufficiency range. In treatments that received B, micronutrient levels in the leaves showed a decreasing trend between the first and third sampling dates. There was some evidence of B overfertilization in the early stages of growth when trees were very small. However, on subsequent dates, the values remained within the sufficiency range.
The leaf concentration of nutrients not included as treatments in the experimental design exhibited minimal variation with the effects of the treatments, and the results are provided in supplemental material (Figure S1). Leaf Ca concentration tended to be higher in the NP-KB treatment, with significant differences between treatments occurring in one of the four sampling dates. Leaf Mg levels did not vary significantly between treatments on any of the sampling dates. Leaf Fe levels showed little consistency, with average values interchanging over the sampling dates, and the same occurred with leaf Zn and Cu concentrations. Leaf Mn levels exhibited some consistency, being higher in the NPK-B treatment, with significant differences between treatments on two of the four sampling dates.
3.3. Soil Properties and Nutrient Bioavailability
Organic C varied significantly between treatments, with the -NPKB treatment showing lower values (Table 1). Soil pH, on the other hand, did not exhibit significant variations between treatments. Extractable P, K, and B demonstrated a clear trend of significantly lower values in the treatments that did not receive each respective nutrient. Metal micronutrients did not vary significantly with the treatments, except for Cu, which showed significantly higher values in the N-PKB treatments. Exchangeable bases and Al did not vary significantly with the treatments, except for K, which was part of the experimental design. Exchangeable acidity was significantly lower in the -NPKB treatment, while CEC did not exhibit significant variations with fertilizer treatments. Soil inorganic N levels, particularly N-NO3−, were lower in the -NPKB treatment compared to the others.
DMY differed significantly between treatments (Table 2). The highest values were found in the NPKB treatment, while the lowest were observed in the -NPKB treatment. The -NPKB treatment exhibited lower average tissue N concentrations, which were significantly different from those of the NPKB treatment. Tissue P levels did not vary significantly with fertilizer treatments, but tissue K and B levels were significantly lower in the treatments that did not receive these nutrients. Ca and Mg levels were the highest in the NP-KB treatment. Fe and Mn concentrations in plant tissue were the highest in the -NPKB treatment, whereas the levels of Zn and Cu did not vary significantly between treatments.
4. Discussion
4.1. The Lack of N and K Negatively Influenced Tree Growth, While the Absence of B Affected Fruit Yield
The growth of young trees, assessed by trunk thickening and canopy volume, was significantly affected by the availability of N and K in the soil. In treatments lacking either of these nutrients, the trees exhibited markedly less growth compared to those in other treatments. N is an essential element required by plants in substantial quantities. It plays a crucial role in various organic structures, notably proteins, nucleic acids, phytohormones, and chlorophylls. When N is deficient, plants clearly exhibit stunted growth [5,17]. In this region, where soils are typically shallow and low in organic matter and nutrients, significant differences in crop productivity often arise when comparing treatments that provide varying N rates [13,14,33].
K does not form part of organic structures, but it is equally required by plants in substantial quantities, playing vital roles in various plant functions. These include enzyme activation, regulation of osmotic fluid pressure with a significant impact on cell extension and stomatal aperture, and the transport of photoassimilates in plant vessels [5,17]. However, due to the high natural availability of K in some soils, the plant's response to K application is not as evident as its response to N application [16,41]. In this experiment, the potential significance of K application may have been influenced by its role in stomatal regulation, especially considering that the trial took place in rainfed conditions, and the growing season included a long, hot, and dry summer. K appears to have a particularly positive effect on crop production under adverse conditions, inducing increased tolerance to drought [22,23]. The nutrient's reduced mobility in the soil through diffusion and mass flow is also observed when soil water availability and plant transpiration decrease [5]. Therefore, lower K levels in the soil may have led to difficulties in nutrient uptake and negative effects on plant growth.
Fruit production was significantly impacted by the absence of B, with the average results of this treatment being the lowest and markedly inferior to the NPKB and N-PKB treatments. B, although it is a micronutrient and required in small quantities, plays crucial roles in plants associated with the biosynthesis and stability of biomembranes and cell walls [8,27]. The lack of B can substantially hinder blossom and fruit set. Plants experiencing B deficiency often exhibit poor flower production and pollen viability. Interestingly, this reproductive failure is frequently observed without deficiency symptoms in shoots, indicating that the B requirement for the reproductive process is greater than for shoot growth [27]. It is noteworthy that chestnut tree, being a dicot species, has higher B requirements compared to monocots [7,8,27]. Moreover, B is chronically deficient in the soils of this region, where a robust crop response to nutrient application, including in chestnut trees, has been consistently observed [21,42,43].
4.2. The Concentrations of N, K, and B in the Leaves Were Strongly Influenced by Their Applications, While P Had a Modest Effect
The treatment that did not receive N (-NPKB) showed lower N levels in the leaves compared to the other treatments. Nevertheless, the average values did not fall below the lower limit of the sufficiency range, which is commonly used in this crop and is set at 20 g kg–1 [21]. Therefore, at least in theory, the impact of not applying N should not be as negative for the plant as indicated by the results of trunk thickening and canopy size. However, the chestnut tree is a less-studied plant due to its economic importance being limited to only a few restricted areas globally [2]. It is plausible that the sufficiency range for different nutrients may not be adequately established, as it requires extensive studies on fertilizer application response. Unfortunately, in the case of chestnut tree, such studies are lacking. For instance, the Plant Analysis Handbook by Bryson et al. [9] provides sufficiency ranges for about 2000 cultivated species but does not include values for chestnut trees. The findings of this study suggest that the lower limit of the sufficiency range for N should be higher. With leaf N levels at 20 g kg–1, the plant may already be in deficiency, at least in a situation known as "hidden hunger," where the plant may appear relatively normal, but growth is reduced, and so is crop yield [9].
The N-PKB treatment did not significantly influence tree growth or fruit production and had very little impact on P concentration in the leaves. However, P is considered the second most limiting element for crop growth and yield in the world [19]. It is estimated that approximately 2/3 of the world's soils have limited P availability to plants, posing a significant constraint on agricultural productivity [18]. Furthermore, P levels in the soils of this region are often considered low when determined by the official method used in Portugal, the Egner-Riehm method [20,21]. However, in this region, there is a known lack of plant response to P application [32,44,45]. The use of this diagnostic method probably should be reconsidered because it seems that its results do not align with the nutrient's bioavailability in the soil, which determines plant responses in the field. The bioavailability of P in these soils has been shown to be higher than the results of the Egner-Riehm method suggest.
The K levels in the leaves were consistently low, often falling below the usual sufficiency range used for this crop [21]. The values for the NP-KB treatment were notably low, indicating a clear deficiency situation that resulted in lower tree productivity in this treatment. Strong constraints on potassium (K) mobility in the soil occur during the summer when soil moisture and plant transpiration decrease, making nutrient uptake challenging as mentioned above. Difficulties in K nutrition have been observed in rainfed crops, especially in plants where fruits develop during the summer [21,41]. Since fruit growth requires a significant amount of K, and fruits are a priority sink for the nutrient [17], the nutrient's translocation to the fruits in the summer leads to a decrease in K levels in the leaves. This reduction was accentuated in the treatment that did not receive K due to lower nutrient availability in the soil.
The B levels in the leaves were very low in the treatment that did not receive B, consistently falling below the lower limit of the sufficiency range reported by Arrobas et al. [21]. It's noteworthy that B uptake is hindered by long periods of drought, as B is generally absorbed through the water transpiration stream and not by active ion transport [5]. Dry conditions also delay the mineralization of B retained in organic matter [5], and this trial took place under rainfed conditions. Additionally, dicots have higher nutrient requirements than monocots, and the lack of B particularly affects the reproductive process [8,27]. Perhaps for these reasons, B deficiency is more widespread globally than that of any other micronutrient [6,7]. In this region, where soils have limited available B, the nutrient's importance in fertilization programs for dicots species is considered equivalent to that of N, despite being a micronutrient [30,31]. Thus, based on this study, the relationship between B deficiency and the plant's difficulty in fruit production seems evident, even though there was no recorded reduction in shoot growth.
The treatment receiving K resulted in lower Ca levels in the leaves compared to the NP-KB. Although dicotyledons have a higher root cation exchange capacity, facilitating the uptake of divalent cations [17], the increase in K in the soil solution and exchange complex, resulting from regular nutrient applications to the soil as fertilizer, may have reduced the opportunity for Ca uptake by roots. This phenomenon is commonly referred to in the literature as ionic antagonism [5]. The result may have been accentuated by the fact that the soil is acidic, and Ca levels in the soil are naturally low.
Possibly the most unexpected result is the consistent increase in Mn levels in the leaves in the plot that did not receive B. Interactions between nutrients are complex, particularly among micronutrients, and tend to be less understood [5]. However, most experimental results have shown that B promotes Mn accumulation in plants, suggesting a synergistic relationship between the two micronutrients [46,47]. In this study, the opposite seemed to occur. Perhaps the lower presence of fruits due to B deficiency reduced available sinks for nutrient translocation, and in the case of Mn, resulted in a higher concentration in the leaves. This could be because part of the Mn tends to accumulate in vacuoles, serving as a temporary storage pool for proper distribution to other plant parts [48].
4.3. The Variations in Nutrient Application Resulted in Differences in Soil Organic Matter and Nutrient Availability
The treatment that did not receive N stood out from the others by having lower soil organic matter content. Soil organic matter is influenced by environmental variables that determine the primary productivity of the agrosystem. It primarily results from the products of photosynthesis deposited in the soil by plant roots or aboveground parts, as well as the mycelia of mycorrhizal fungi associated with plants [5,49]. The absence of N application reduced the mineral N content in the soil and the development of herbaceous vegetation under the tree canopy, contributing to the decrease in soil organic matter. While this study did not assess herbaceous vegetation production under the canopy, the results of ryegrass, cultivated as an indicator of soil fertility, clearly showed that in the N-unfertilized treatment, the plant yielded much less dry matter than the treatments receiving N. In various studies, it has been observed that in orchards where fertilizer application is done under the canopy, a fertility pool quickly forms, including higher levels of organic matter, because of the stimulation in the growth of both the cultivated plant and the spontaneous herbaceous vegetation that develops under the canopy [45,50,51]. This is the same principle by which cover cropping strategies, compared to bare soil or herbicide use, increase soil organic matter [52,53,54].
The exchange acidity was significantly lower in the treatment that did not receive N compared to the other treatments, and the average pH values were higher in this treatment, although not significantly different from the others. The application of N-containing mineral fertilizers, especially those with urea or ammoniacal N components, is known to contribute to soil acidification through nitrification reactions. These reactions generate an excess of H+ ions during the bacterial conversion of NH4+ to NO3− [5,55]. In this study, the regular application of N, partly ammoniacal, may have contributed to soil acidification, consistent with findings in recent studies on N fertilization [50,56].
The non-application of K (NP-KB) or B (NPK-B) reduced the ryegrass DMY compared to the control treatment that received all nutrients (NPKB). It also significantly decreased the nutrient concentrations in the ryegrass tissues. The pots were irrigated, eliminating the argument of greater uptake difficulties under rainfed conditions, as mentioned for K and B in the field trial. Thus, it appears evident that, in this type of soil, the application of K and B is crucial for plant growth, as observed in previous studies [21,30,41,43]. It was also clear that in the treatment without K, plants showed higher levels of Ca and Mg in tissues, justifiable by the phenomenon of ionic antagonism in nutrient uptake between K and divalent cations [5,17].
The levels of Fe and Mn were higher in the pots of the -NPKB treatment. Plants in the -NPKB treatment produced much less biomass than the others, and the concentrations of Fe and Mn could be justified by the nutrient concentration phenomenon. In plants, concentration/dilution phenomena occur when there is a variation in DMY for a given amount of bioavailable nutrient in the medium [57]. However, in this case, irrigation of the pots may have had some influence. Irrigation causes wetting and drying cycles, with moments of poor soil aeration. The solubility of Fe and Mn in the soil depends heavily on the soil's redox potential [58,59]. The pots that did not receive N produced much less biomass, indicating lower transpiration during the trial. It is likely that the pots in the -NPKB treatment, with lower growth rates, lost water more slowly, maintaining more pronounced reduction conditions than in the other pots. This may have contributed to an increase in solubility and uptake of higher amounts of Fe and Mn in the -NPKB treatment.
5. Conclusions
The growth of the trees was negatively impacted by the lack of N and K application, resulting in trees with thinner trunks and smaller canopies. The absence of B seemed to affect the ability of young trees to initiate fruit production more than their vegetative growth. The non-application of P did not show significant effects on tree growth or fruit production, suggesting that, among the four nutrients, P may be the least relevant for the growth and fruit production process under the conditions of this study. The levels of K and B in the leaves were below the lower limits of their sufficiency ranges, clearly indicating deficiency situations for these nutrients. In the case of N, the nutrient levels in the leaves were well above the lower limit of the sufficiency range commonly used in this crop. Given the limited scientific literature on this topic, it appears evident that the limits of the N sufficiency range should be established at higher values. Furthermore, the non-application of N reduced the soil organic matter content. This result is likely due to the lower deposition of leaf litter, roots, and possibly mycorrhizal fungal structures, but mainly to the reduced development of herbaceous vegetation under the canopy, resulting in a lower deposition of organic substrate in the soil.
In summary, the results demonstrated the high importance of B for the reproductive process of the chestnut tree, a dicot species, and the unequivocal effects of N and K on tree growth. These three nutrients proved to be the most important for the initial establishment phase of chestnut trees under the conditions of this study.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
M.A.: funding acquisition, investigation, methodology, and writing—original draft preparation; S.R.: investigation; C.M.C.: methodology; writing—review and editing; M.Â.R.: conceptualisation, funding acquisition, project administration, data curation, writing—review and editing.
Funding
The authors are grateful to the Foundation for Science and Technology (FCT, Portugal) for financial support from national funds FCT/MCTES, to CIMO (UIDB/AGR/00690/2020), SusTEC (LA/P/0007/2020), and CITAB (UIDB/04033/2020)
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ramalhosa, E.; Pereira, E.L.; Silva, M.F.L. Valorização da castanha. [Fruit valorisation]. In Manual de boas Práticas do Castanheiro [Handbook of Good Management Practices on Chestnut Orchards]; Bento, A., Ribeiro, A.C., Eds.; Terras de Trás-os-Montes: Bragança, Portugal, 2020; pp. 235–244. (In Portuguese) [Google Scholar]
- FAOSTAT. 2024. Production: Crops and livestock products. Available online: https://www.fao.org/faostat/en/#data/QCL. (accessed on 26 February 2024).
- Almeida, A. Instalação da cultura [Crop planting]. In Manual de boas Práticas do Castanheiro [Handbook of Good Management Practices on Chestnut Orchards]; Bento, A., Ribeiro, A.C., Eds.; Terras de Trás-os-Montes: Bragança, Portugal, 2020; pp. 85–92. (In Portuguese) [Google Scholar]
- Patrício, M.A. Sistemas de condução e poda [Training and pruning systems]. In Manual de boas Práticas do Castanheiro [Handbook of Good Management Practices on Chestnut Orchards]; Bento, A., Ribeiro, A.C., Eds.; Terras de Trás-os-Montes: Bragança, Portugal, 2020; pp. 149–170. (In Portuguese) [Google Scholar]
- Weil, R.R.; Brady, N.C. The Nature and Properties of Soils, 15th ed; Global Edition: London, UK, 2017. [Google Scholar]
- Shorrocks, V.M. The Occurrence and Correction of Boron Deficiency. Plant Soil 1997, 193, 121–148. [Google Scholar] [CrossRef]
- Gupta, U.C. Boron. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 241–277. [Google Scholar]
- Wimmer, M.A.; Eichert, T. Review: Mechanisms for Boron Deficiency-Mediated Changes in Plant Water Relations. Plant Sci 2013, 203/204, 25–32. [Google Scholar] [CrossRef]
- Bryson, G.; Mills, H.A.; Sasseville, D.N.; Jones, J.B., Jr.; Barker, A.V. Plant Analysis Handobook III. A Guide to Sampling, Preparation, Analysis and Interpretation for Agronomic and Horticultural Crops; Micro-Macro Publishing, Inc.: Athens, GA, USA, 2014. [Google Scholar]
- Mulla, D.J.; Strock, J.S. Nitrogen transformation process in soils. In Nitrogen in Agricultural Systems, Agrono-My Monograph n.49; Schepers, J.S., Raun, W.R., Eds.; ASA, CSSA, SSSA: Madison, WI, USA, 2008; pp. 361–400. [Google Scholar] [CrossRef]
- Poikane, S.; Phillips, G.; Birk, S.; Free, G.; Kelly, M.G.; Willby, N.J. Deriving nutrient criteria to support ʽgoodʼ ecological status in European lakes: an empirically based approach to linking ecology and management. Sci. Total Environ. 2019, 650, 2074–2084. [Google Scholar] [CrossRef] [PubMed]
- Haberman, A.; Dag, A.; Shtern, N.; Zipori, I.; Erel, R.; et al. Significance of Proper Nitrogen Fertilization for Olive Productivity in Intensive Cultivation. Sci. Hortic. 2019, 246, 710–717. [Google Scholar] [CrossRef]
- Ferreira, I.Q.; Arrobas, M.; Moutinho-Pereira, J.M.; Correia, C.M.; Rodrigues, M.A. The Effect of Nitrogen Applications on the Growth of Young Olive Trees and Nitrogen Use Efficiency. Turk. J. Agric. For. 2020, 44, 278–289. [Google Scholar] [CrossRef]
- Lopes, J.I.; Gonçalves, A.; Brito, C.; Martins, S.; Pinto, L.; Moutinho-Pereira, J.; Raimundo, S.; Arrobas, M.; Rodrigues, M.A.; Correia, C.M. Inorganic Fertilization at High N Rate Increased Olive Yield of a Rainfed Orchard but Reduced Soil Organic Matter in Comparison to Three Organic Amendments. Agronomy 2021, 11, 2172. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Ferreira, I.Q.; Claro, A.M.; Arrobas, M. Fertiliser Recommendations for Olive Based Upon Nutrients Removed in Crop and Pruning. Sci. Hortic. 2012, 142, 205–211. [Google Scholar] [CrossRef]
- Arrobas, M.; Ferreira, I.Q.; Freitas, S.; Verdial, J.; Rodrigues, M.A. Guidelines for Fertilizer Use in Vineyards Based on Nutrient Content of Grapevine Parts. Sci. Horti 2014, 172, 191–198. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Moller, I.S.; White, P. Function of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Elsevier: London, UK, 2012; pp. 135–189. [Google Scholar] [CrossRef]
- Sepehr, E.; Rengel, Z.; Fateh, E.; Sadaghiani, M.R. Differential Capacity of Wheat Cultivars and White Lupin to Acquire Phosphorus from Rock Phosphate, Phytate and Soluble Phosphorus Sources. J. Plant Nutr. 2012, 35, 1180–1191. [Google Scholar] [CrossRef]
- Li, G.; Huang, G.; Li, H.; van Ittersum, M.K.; Leffelaar, P.A.; Zhang, F. Identifying Potential Strategies in the Key Sectors of China’s Food Chain to Implement Sustainable Phosphorus Management: A Review. Nutr. Cycl. Agroecosyst. 2016, 104, 341–359. [Google Scholar] [CrossRef]
- Ferreira, I.Q.; Rodrigues, M.A.; Moutinho-Pereira, J.M.; Correia, C.; Arrobas, M. Olive Tree Response to Applied Phosphorus in Field and Pot Experiments. Sci. Hortic. 2018, 234, 236–244. [Google Scholar] [CrossRef]
- Arrobas, M.; Afonso, S.; Rodrigues, M.A. Diagnosing the Nutritional Condition of Chestnut Groves by Soil and Leaf Analyses. Sci. Hortic. 2018, 228, 113–121. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. (2014) Regulation of Potassium Transport in Plants under Hostile Conditions: Implications for Abiotic and Biotic Stress Tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Senbayram, M.S.; Peiter, E. Potassium in Agriculture: Status and Perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef]
- Qiu, S.; Xie, J.; Zhao, S.; Xu, X.; Hou, Y.; Wang, X.; Zhou, W.; He, P.; Johnston, A.M.; Christie, P.; Jin, J. Long-Term Effects of Potassium Fertilization on Yield, Efficiency, and Soil Fertility Status in a Rain-Fed Maize System in Northeast China. Field Crops Res. 2014, 163, 1–9. [Google Scholar] [CrossRef]
- Zhao, S.; He, P.; Qiu, S.; Jia, L.; Liu, M.; Jin, J.; Johnston, A.M. Long-Term Effects of Potassium Fertilization and Straw Return on Soil Potassium Levels and Crop Yields in Northcentral China. Field Crops Res. 2014, 169, 116–122. [Google Scholar] [CrossRef]
- Khan, S.A.; Mulvaney, R.L.; Ellsworth, T.R. The Potassium Paradox: Implications for Soil Fertility, Crop Production and Human Health. Renew. Agric. Food Syst. 2013, 29(1), 3–27. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012; pp. 191–248. [Google Scholar] [CrossRef]
- Soyergin, S. Effects of Soil and Leaf Treatments to Eliminate Boron Deficiency in Olives. Commun. Soil Sci. Plant Anal. 2010, 41, 2004–2010. [Google Scholar] [CrossRef]
- Thapa, U.; Prasad, P.H.; Rai, R. Studies on Growth, Yield and Quality of Broccoli (Brassica oleracea L. var italica Plenck) as Influenced by Boron and Molybdenum. J. Plant Nutr. 2016, 39, 261–267. [Google Scholar] [CrossRef]
- Ferreira, I.Q.; Rodrigues, M.A.; Arrobas, M. Soil and Foliar Applied Boron in Olive: Tree Crop Growth and Yield, and Boron Remobilization within Plant Tissue. Span. J. Agric. Res. 2019, 17, e0901. [Google Scholar] [CrossRef]
- Arrobas, M.; Raimundo, S.; Conceição, N.; Moutinho-Pereira, J.; Correia, C.M.; Rodrigues, M.Â. On Sandy, Boron-Poor Soils, Liming Induced Severe Boron Deficiency and Drastically Reduced the Dry Matter Yield of Young Olive Trees. Plants 2023, 12, 4161. [Google Scholar] [CrossRef] [PubMed]
- Arrobas, M.; Afonso, S.; Ferreira, I.Q.; Moutinho-Pereira, J.M.; Correia, C.M.; Rodrigues, M.A. Liming and Application of Nitrogen, Phosphorus, Potassium and Boron on a Young Plantation of Chestnut. Turk. J. Agric. For. 2017, 41, 441–451. [Google Scholar] [CrossRef]
- Arrobas, M.; Belotto, L.B.; Marchetti, J.A.; Barroso, V.; Raimundo, S.; Cassol, L.C.; Correia, C.M.; Rodrigues, M.Â. Excessive Delay in Nutrient Release by Controlled-Release Fertilizers Can Reduce Chestnut Yield. Horticulturae 2022, 8, 1067. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Raimundo, S.; Pereira, A.; Arrobas, M. Large Chestnut Trees (Castanea sativa) Respond Poorly to Liming and Fertilizer Application. J. Soil Sci. Plant Nutr. 2020, 20, 1261–1270. [Google Scholar] [CrossRef]
- IPMA (Instituto Português do Mar e da Atmosfera); [Portuguese Institute of the Sea and the Atmosphere]. Normais Climatológicas [Climate Normals]. 2024. Available online: https://www.ipma.pt/pt/oclima/normais.clima/ (accessed on 15 April 2024). (In Portuguese).
- WRB. World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Temminghoff, E.E.; Houba, V.J. Plant Analysis Procedures, 2nd ed.; Kluwer Academic Publishers: London, UK, 2004. [Google Scholar] [CrossRef]
- Van Reeuwijk, L.P. Procedures for Soil Analysis, 6th ed.; Technical Paper 9; ISRIC (International Soil Reference Information Center); FAO (Food and Agriculture Organization of the United Nations): Wageningen, The Netherlands, 2002; ISBN 90-6672-044-1. [Google Scholar]
- FAO. Standard Operating Procedure for Soil Available Micronutrients (Cu, Fe, Mn, Zn) and Heavy Metals (Ni, Pb, Cd), DTPA Extraction Method; FAO: Rome, Italy, 2022; Available online: https://www.fao.org/3/cc0048en/cc0048en.pdf (accessed on 15 April 2024).
- Baird, R.B.; Eaton, A.D.; Rice, E.W. Nitrate by ultraviolet spectrophotometric method. In Standard Methods for the Examination of Water and Wastewater; American Public Health Association, American Water Works Association, Water Environment Federation: Washington, DC, USA, 2017. [Google Scholar]
- Ferreira, I.Q.; Arrobas, M.; Moutinho-Pereira, J.M.; Correia, C.; Rodrigues, M.A. Olive Response to Potassium Applications under Different Water Regimes and Cultivars. Nutr. Cycl. Agroecosyst. 2018, 112, 387–401. [Google Scholar] [CrossRef]
- Portela, E.A.C.; Ferreira-Cardoso, J.V.; Louzada, J.L. Boron Application on a Chestnut Orchard: Effect on Yield and Quality of Nuts. J. Plant Nutr. 2011, 34, 1245–1253. [Google Scholar] [CrossRef]
- Portela, E.; Ferreira-Cardoso, J.; Louzada, J.; Gomes-Laranjo, J. Assessment of Boron Application in Chestnuts: Nut Yield and Quality. J. Plant Nutr. 2015, 38, 973–987. [Google Scholar] [CrossRef]
- Arrobas, M.; Decker, J.V.; Feix, B.L.; Godoy, W.I.; Casali, C.A.; Correia, C.M.; Rodrigues, M.A. Biochar and Zeolites Did Not Improve Phosphorus Uptake or Crop Productivity in a Field Trial Performed in an Irrigated Intensive Farming System. Soil Use Manage. 2022, 38, 564–575. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Grade, V.; Barroso, V.; Pereira, A.; Cassol, L.C.; Arrobas, M. Chestnut Response to Organo-Mineral and Controlled-Release Fertilizers in Rainfed Growing Conditions. J. Soil Sci. Plant Nutr 2020, 20, 390–391. [Google Scholar] [CrossRef]
- Lopez-Lefebre, L.R.; Rivero, R.M.; Garcia, P.C.; Sánchez, E.; Ruiz, J.M.; Romero, L. Boron Effect on Mineral Nutrients of Tobacco. J. Plant Nutr. 2002, 25, 509–522. [Google Scholar] [CrossRef]
- Long, Y.; Peng, J. Interaction between Boron and Other Elements in Plants. Genes 2023, 14, 130. [Google Scholar] [CrossRef]
- Alejandro, S.; Höller, S.; Meier, B.; Peiter, E. Manganese in Plants: From Acquisition to Subcellular Allocation, Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Godbold, D.L.; Hoosbeek, M.R.; Lukac, M.; Cotrufo, M.F.; Janssens, I.A.; Ceulemans, R.; Polle, A.; Velthorst, E.J.; Scarascia Mugnozza, G.; Angelis, P.; et al. Mycorrhizal Hyphal Turnover as a Dominant Process for Carbon Input into Soil Organic Matter. Plant Soil 2006, 281, 15–24. [Google Scholar] [CrossRef]
- Silva, E.; Arrobas, M.; Gonçalves, A.; Martins, S.; Raimundo, S.; Pinto, L.; Brito, C.; Moutinho-Pereira, J.; Correia, C.M.; Rodrigues, M.A. A. Controlled-Release Fertilizer Improved Soil Fertility but not Olive Tree Performance. Nutr. Cycl. Agroecosyst. 2021, 120(1), 1–15. [Google Scholar] [CrossRef]
- Lopes, J.I.; Arrobas, M.; Raimundo, S.; Gonçalves, A.; Brito, C.; Martins, S.; Pinto, L.; Moutinho-Pereira, J.; Correia, C.M.; Rodrigues, M.A. Photosynthesis, Yield, Nutrient Availability and Soil Properties after Biochar, Zeolites or Mycorrhizal Inoculum Application to a Mature Rainfed Olive Orchard. Agriculture 2022, 12, 171. [Google Scholar] [CrossRef]
- Márquez-García, F.; Sánchez, E.J.G.; Castro-Garcia, S.; Ordóñez-Fernández, R. Improvement of Soil Carbon Sink by Cover Crops in Olive Orchards under Semiarid Conditions. Influence of the Type of Soil and Weed. Span. J. Agric. Res. 2013, 11, 335. [Google Scholar] [CrossRef]
- Torres, M.R.-R.; Ordóñez-Fernández, R.; Giráldez, J.V.; Márquez-García, J.; Laguna, A.; Carbonell-Bojollo, R. Efficiency of Four Different Seeded Plants and Native Vegetation as Cover Crops in the Control of Soil and Carbon Losses by Water Erosion in Olive Orchards. Land Degrad. Dev. 2018, 29, 2278–2290. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Arrobas, M. Cover cropping for increasing fruit production and farming sustainability. In Fruit Crops: Diagnosis and Management of Nutrient Constraints; Srivastava, A.K., Hu, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 279–295. [Google Scholar] [CrossRef]
- Neumann, G.; Römheld, V. Rhizosphere in relation to plant nutrition. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012; pp. 347–368. [Google Scholar] [CrossRef]
- Arrobas, M.; Silva, J.; Busato, M.R.; Ferreira, A.C.; Raimundo, S.; Pereira, A.; Finatto, T.; de Mello, N.A.; Correia, C.M.; Rodrigues, M.Â. Large Chestnut Trees Did Not Respond to Annual Fertiliser Applications, Requiring a Long-Term Approach to Establishing Effective Fertilisation Plans. Soil Syst. 2023, 7, 2. [Google Scholar] [CrossRef]
- Römheld, V. Diagnosis of deficiency and toxicity of nutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012; pp. 299–312. [Google Scholar] [CrossRef]
- George, E.; Horst, W.J.; Neumann, E. Adaptation of Plants to Adverse Chemical Soil Conditions. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 409–472. [Google Scholar] [CrossRef]
- Sparrow, L.A.; Uren, N.C. Manganese Oxidation and Reduction in Soils: Effects of Temperature, Water Potential, pH and Their Interactions. Soil Res. 2014, 52, 483–494. [Google Scholar] [CrossRef]
Figure 1.
Climatological normal, and average monthly temperature and accumulated precipitation from 2019 to 2022 recorded in the weather station of the Santa Apolónia farm in Bragança.
Figure 1.
Climatological normal, and average monthly temperature and accumulated precipitation from 2019 to 2022 recorded in the weather station of the Santa Apolónia farm in Bragança.
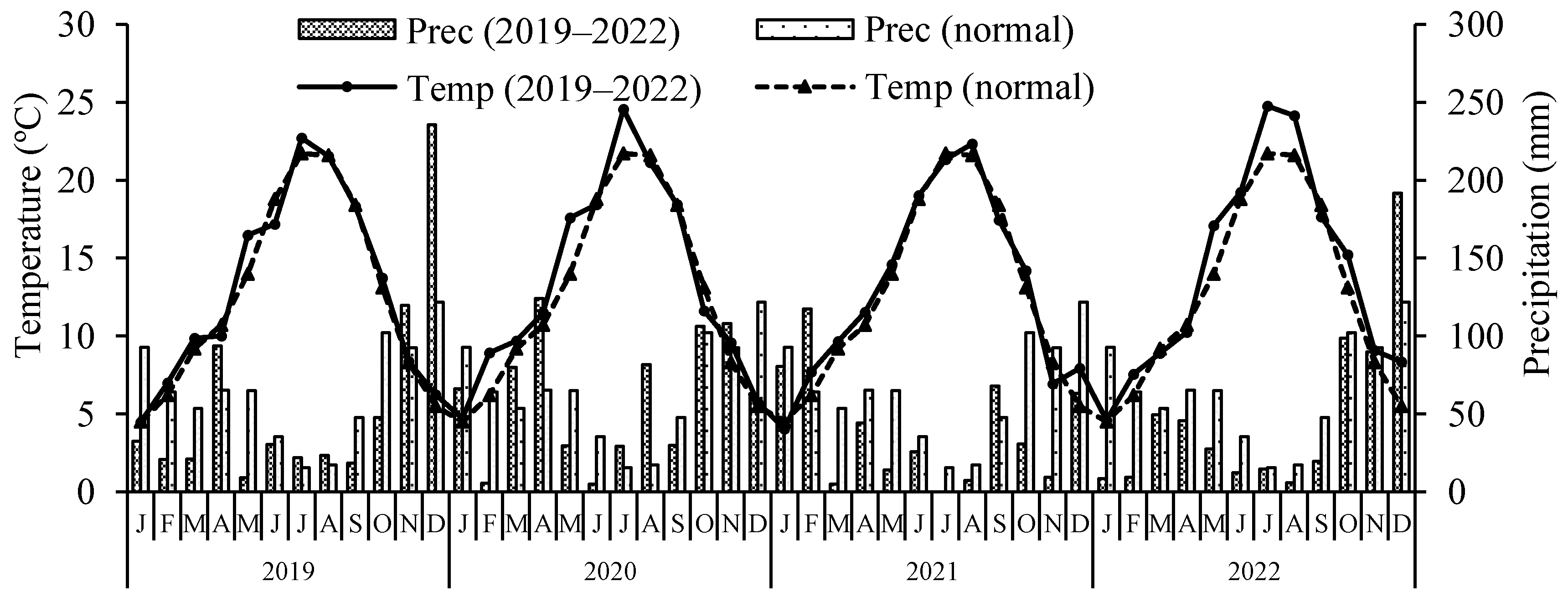
Figure 2.
Trunk circumference on two sampling dates (October 2021 and 2022) in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter do not differ significantly using the Tukey HSD test (α = 0.05). Vertical bars represent standard errors.
Figure 2.
Trunk circumference on two sampling dates (October 2021 and 2022) in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter do not differ significantly using the Tukey HSD test (α = 0.05). Vertical bars represent standard errors.
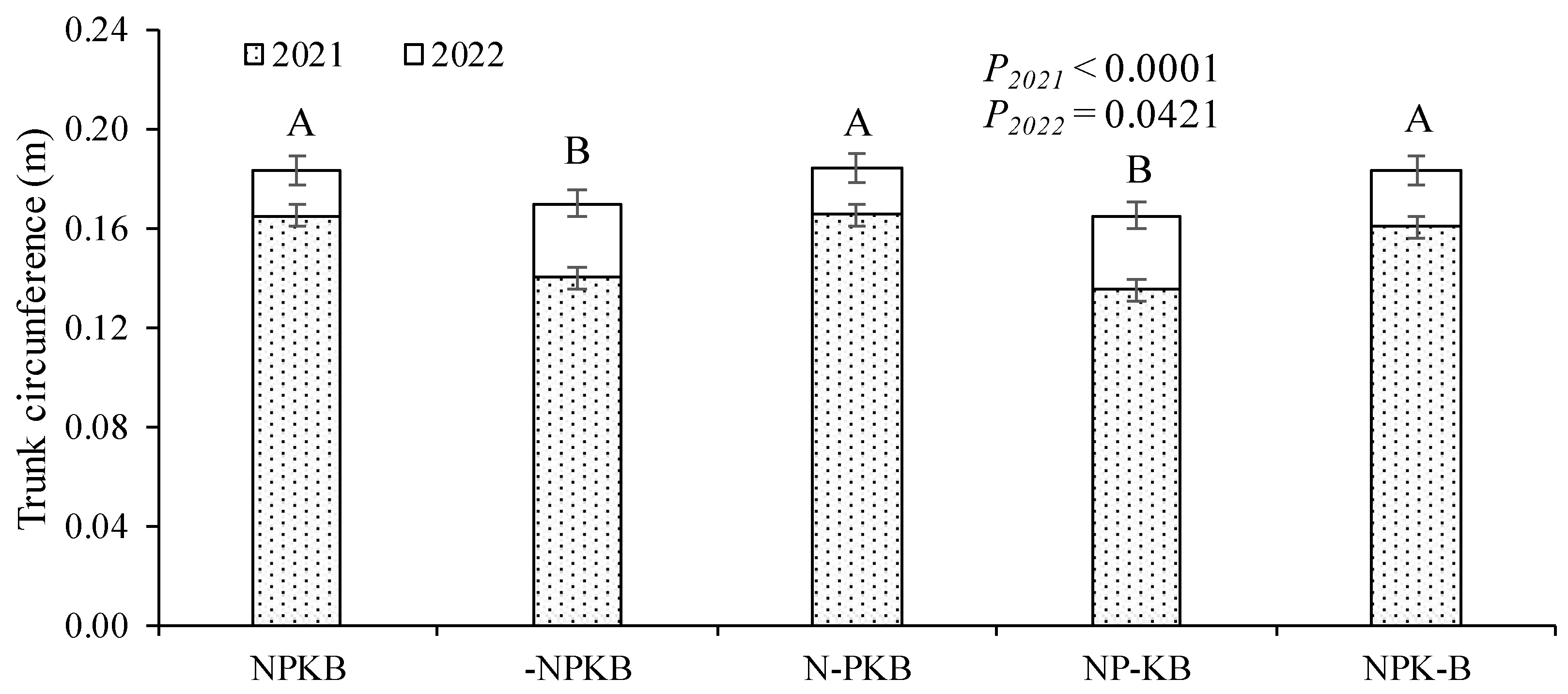
Figure 3.
Canopy volume on two sampling dates (October 2021 and 2022) in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter do not differ significantly using the Tukey HSD test (α = 0.05). Vertical bars represent standard errors.
Figure 3.
Canopy volume on two sampling dates (October 2021 and 2022) in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter do not differ significantly using the Tukey HSD test (α = 0.05). Vertical bars represent standard errors.
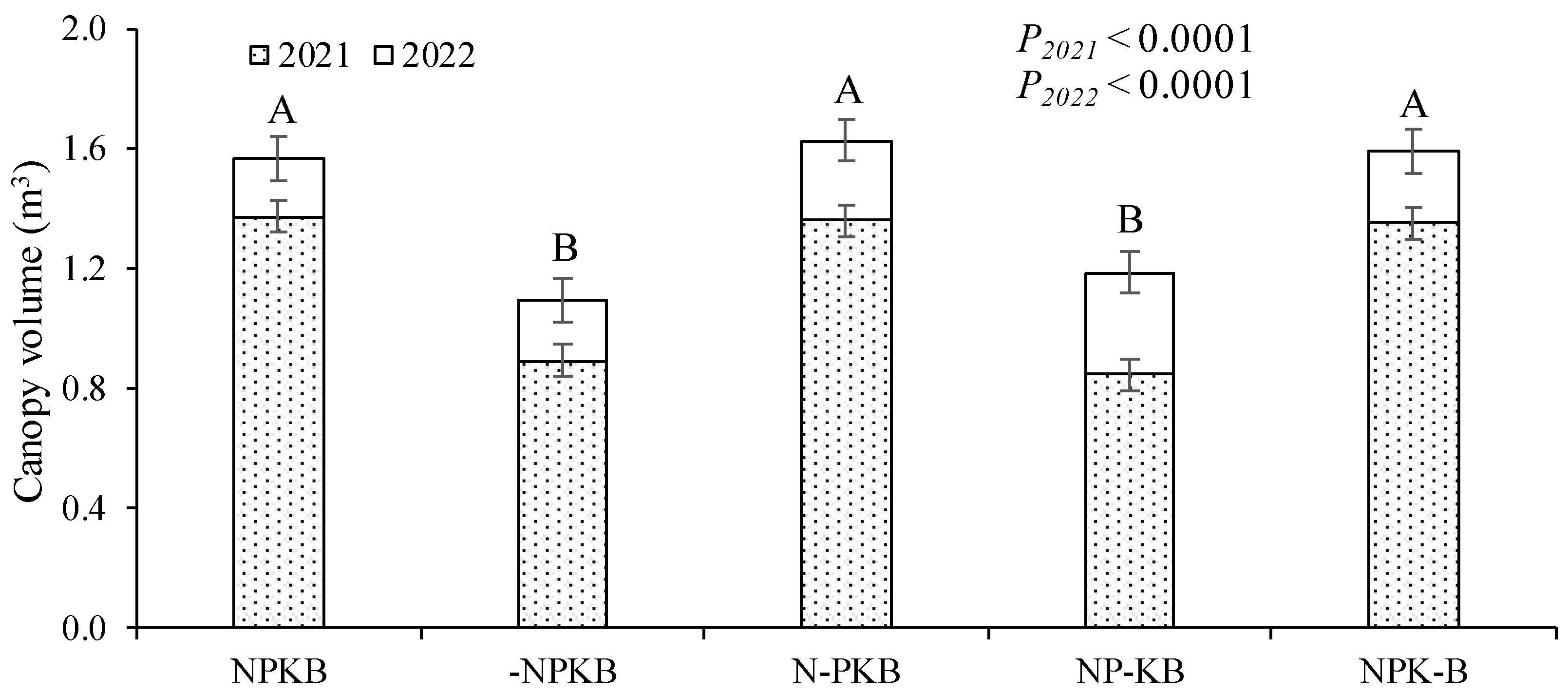
Figure 4.
Fruit yield in three successive harvests starting from the first year in which the trees produced fruit in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter (lower case for year and upper case for accumulated total) do not differ significantly using the Tukey HSD test (α = 0.05). Vertical bars represent standard errors.
Figure 4.
Fruit yield in three successive harvests starting from the first year in which the trees produced fruit in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter (lower case for year and upper case for accumulated total) do not differ significantly using the Tukey HSD test (α = 0.05). Vertical bars represent standard errors.
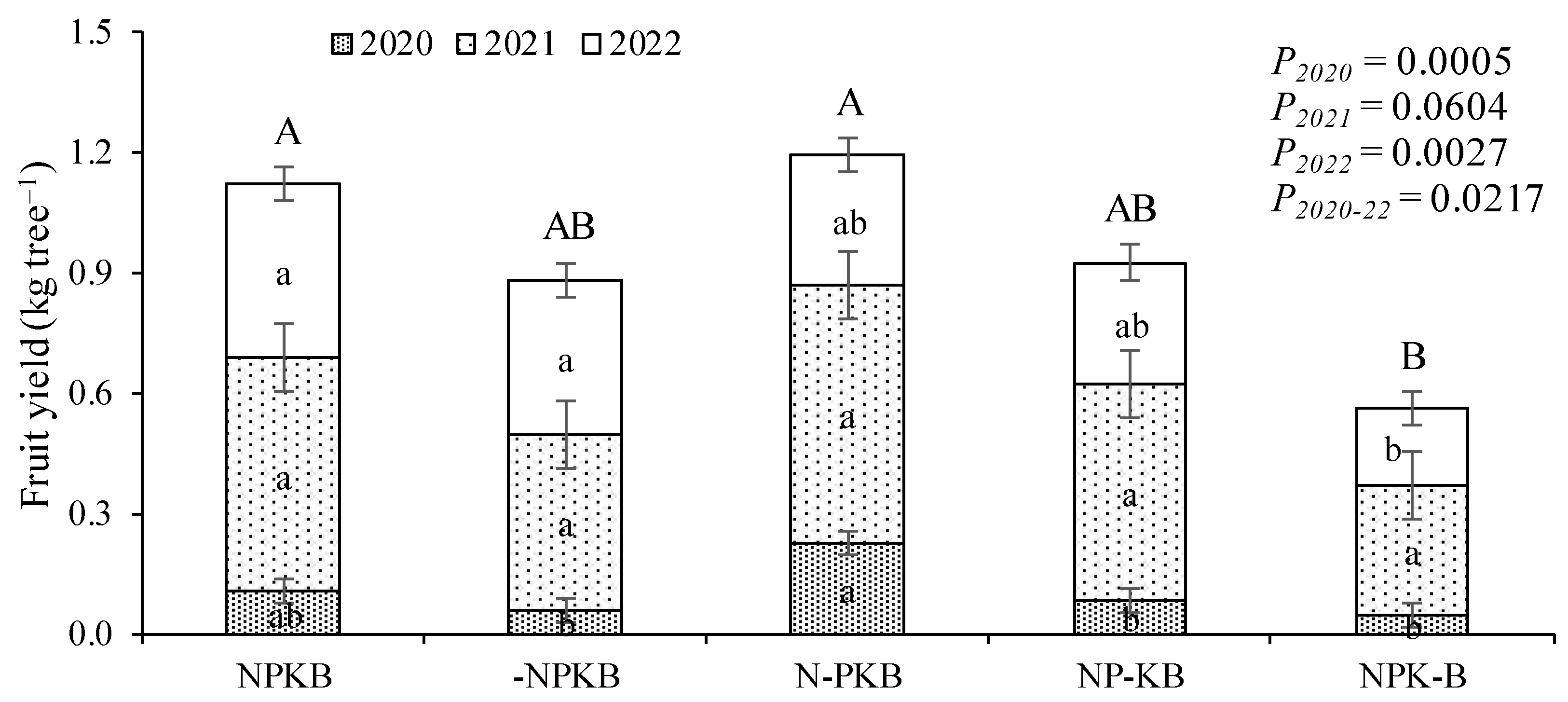
Figure 5.
Leaf concentration of the nutrients included in the experimental design, from samples taken in July (J) (2019-2022), in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). LLSR and HLSR are the lower and higher limit of the sufficiency range, respectively. The vertical bars are the standard error of the variance analysis. *, ** and *** significant differences at P < 0.05, P < 0.01 and P < 0.001; ns, not significant.
Figure 5.
Leaf concentration of the nutrients included in the experimental design, from samples taken in July (J) (2019-2022), in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). LLSR and HLSR are the lower and higher limit of the sufficiency range, respectively. The vertical bars are the standard error of the variance analysis. *, ** and *** significant differences at P < 0.05, P < 0.01 and P < 0.001; ns, not significant.
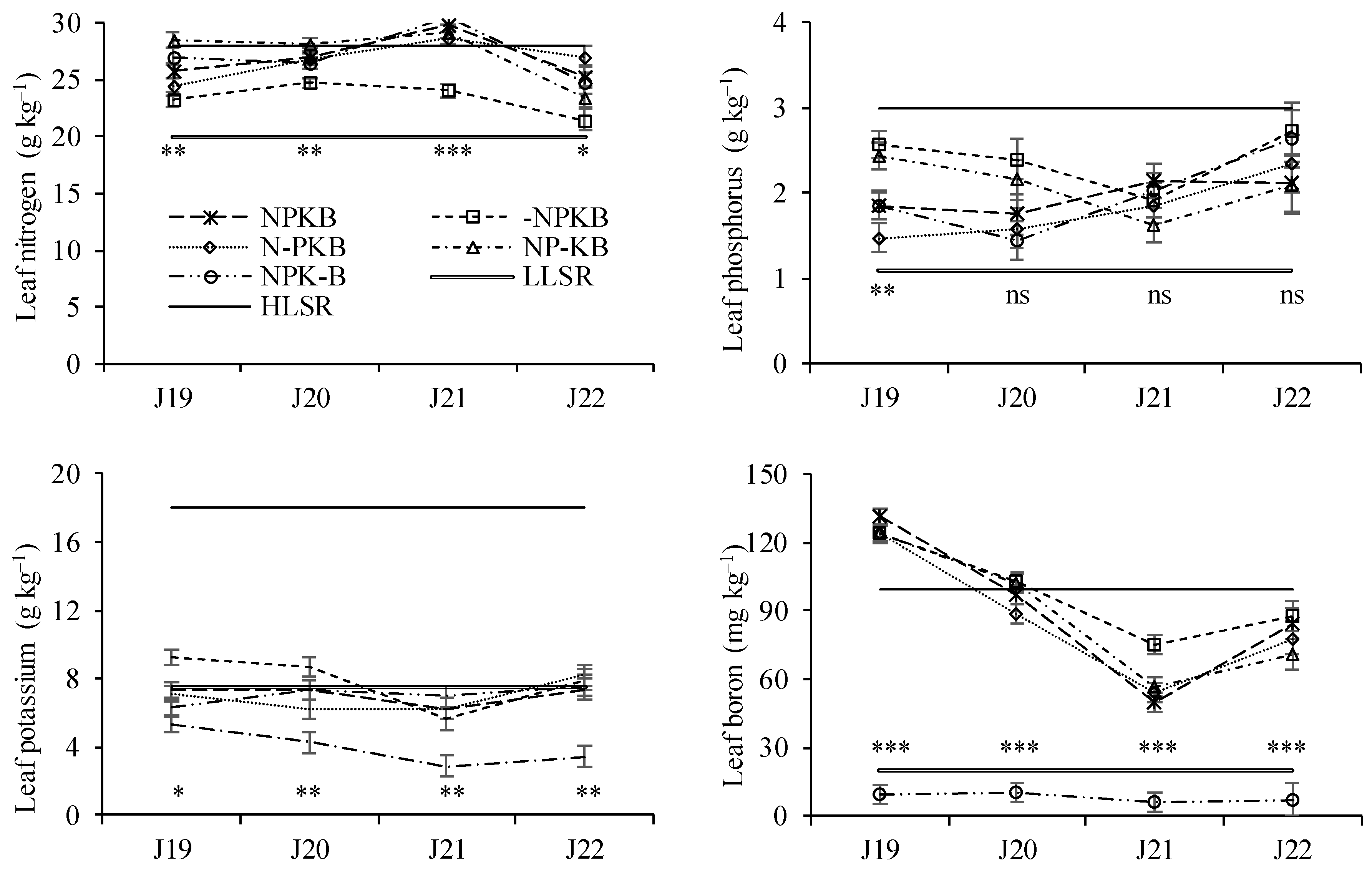

Table 1.
Soil properties from samples taken at 0-0.20 cm depth at the end of the experiment, in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter do not differ significantly using the Tukey HSD test (α = 0.05).
Table 1.
Soil properties from samples taken at 0-0.20 cm depth at the end of the experiment, in response to fertilizer treatments in a nutrient omission trial. The control received all four nutrients (nitrogen, phosphorus, potassium, and boron, NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter do not differ significantly using the Tukey HSD test (α = 0.05).
| Treatments | ||||||||
|---|---|---|---|---|---|---|---|---|
| Soil properties | NPKB | -NPKB | N-PKB | NP-KB | NPK-B | Prob. | SE | |
| 1OC (g kg−1) | 8.1 a | 5.3 b | 8.5 a | 9.0 a | 8.8 a | < 0.0001 | 0.34 | |
| 2pH (H2O) | 5.2 a | 5.5 a | 5.3 a | 5.2 a | 5.2 a | 0.2834 | 0.12 | |
| 2pH (KCl) | 4.0 a | 4.2 a | 4.1 a | 4.0 a | 4.1 a | 0.1472 | 0.07 | |
| Extractable macro and micronutrients (mg kg-1) | ||||||||
| 3Phosphorus (P2O5) | 212.5 a | 206.8 a | 41.7 c | 184.5 ab | 152.1 b | <0.0001 | 11.70 | |
| 3Potassium (K2O) | 313.0 a | 395.7 a | 386.0 a | 58.0 b | 370.0 a | <0.0001 | 26.06 | |
| 4Boron | 5.2 a | 5.7 a | 4.4 a | 4.3 a | 0.9 b | 0.0002 | 0.45 | |
| 5Iron | 26.2 a | 26.6 a | 33.9 a | 26.1 a | 25.2 a | 0.2084 | 2.66 | |
| 5Zinc | 0.4 a | 0.4 a | 0.3 a | 0.6 a | 0.3 a | 0.1244 | 0.07 | |
| 5Copper | 0.6 b | 0.4 b | 0.9 a | 0.5 b | 0.3 b | 0.0057 | 0.25 | |
| 5Manganese | 8.5 a | 6.7 a | 7.6 a | 6.4 a | 6.7 a | 0.2923 | 0.72 | |
| Exchangeable complex (cmolc kg−1) | ||||||||
| 6Calcium | 6.6 a | 5.4 a | 5.1 a | 4.9 a | 5.0 a | 0.4448 | 0.68 | |
| 6Magnesium | 1.8 a | 1.7 a | 1.5 a | 1.2 a | 1.5 a | 0.4679 | 0.21 | |
| 6Potassium | 0.9 ab | 1.2 a | 1.1 ab | 0.1 c | 0.9 b | < 0.0001 | 0.06 | |
| 6Sodium | 0.3 a | 0.3 a | 0.2 a | 0.2 a | 0.2 a | 0.1868 | 0.02 | |
| 7Aluminium | 1.3 a | 0.9 a | 1.2 a | 1.4 a | 1.3 a | 0.1172 | 0.12 | |
| 7Acidity | 3.5 b | 1.6 c | 3.1 b | 3.9 a | 4.0 a | <0.0001 | 0.08 | |
| 8CEC | 13.0 a | 10.2 a | 11.1 a | 10.4 a | 11.7 a | 0.1023 | 0.70 | |
| Inorganic nitrogen (mg kg-1) | ||||||||
| 9N-NH4+ | 48.1 a | 22.3 b | 60.8 a | 63.7 a | 44.3 ab | 0.0013 | 5.04 | |
| 10N-NO3− | 9.0 a | 1.2 b | 8.7 a | 7.6 a | 6.2 a | <0.0001 | 0.69 | |
1Organic carbon, wet digestion (Walkley–Black); 2Potentiometry; 3Ammonium lactate; 4Hot water, azomethine-H; 5Ammonium acetate and EDTA (ethylenediaminetetraacetic acid); 6Ammonium acetate; 7Potassium chloride; 8Cation Exchange Capacity; 9KCl extraction, colorimetry; 10UV/Vis spectrophotometry.
Table 2.
Dry mater yield (DMY) and tissue nutrient concentration in ryegrass grown on soil samples taken at the end of the field experiment beneath the plant canopy where the fertilizers were applied. Data from a nutrient omission trial in which the control received the four nutrients nitrogen, phosphorus, potassium, and boron (NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter do not differ significantly using the Tukey HSD test (α = 0.05).
Table 2.
Dry mater yield (DMY) and tissue nutrient concentration in ryegrass grown on soil samples taken at the end of the field experiment beneath the plant canopy where the fertilizers were applied. Data from a nutrient omission trial in which the control received the four nutrients nitrogen, phosphorus, potassium, and boron (NPKB), while others received only three out of the four, with the missing nutrient marked with (-). Means followed by the same letter do not differ significantly using the Tukey HSD test (α = 0.05).
| DMY | Nitrogen | Phosphorus | Potassium | Calcium | Magnesium | Boron | Iron | Manganese | Zinc | Copper | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| g pot–1 | g kg–1 | mg kg–1 | |||||||||
| NPKB | 6.1 a | 12.1 a | 2.8 a | 25.3 a | 5.2 bc | 2.6 bc | 124.2 a | 143.1 ab | 355.9 b | 17.3 a | 5.9 a |
| -NPKB | 2.5 c | 11.0 b | 3.0 a | 26.7 a | 5.4 bc | 2.7 abc | 129.2 a | 221.8 a | 492.4 a | 18.7 a | 6.0 a |
| N-PKB | 5.2 b | 11.1 ab | 2.4 a | 24.3 a | 3.9 c | 2.3 c | 119.7 a | 143.3 ab | 301.3 b | 16.3 b | 4.3 a |
| NP-KB | 4.5 b | 11.5 ab | 2.8 a | 8.4 b | 7.9 a | 3.1 a | 127.9 a | 110.6 b | 298.6 b | 18.9 a | 4.8 a |
| NPK-B | 4.7 b | 12.0 ab | 2.7 a | 25.3 a | 6.4 b | 2.9 ab | 11.0 b | 117.7 b | 344.3 b | 21.1 a | 5.4 a |
| Prob. | <0.0001 | 0.044 | 0.1183 | 0.0001 | <0.0001 | 0.0002 | <0.0001 | 0.0168 | <0.0001 | 0.0650 | 0.0981 |
| SE | 0.16 | 0.29 | 0.15 | 2.06 | 0.44 | 0.12 | 8.84 | 16.2 | 17.62 | 0.76 | 0.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



