
Submitted:
04 May 2024
Posted:
06 May 2024
You are already at the latest version
Abstract
This study utilized phytochemical screening to conduct qualitative analysis of plant extracts, aiming to identify various classes of secondary metabolites. Moreover, the antibacterial activity of different types of O. vulgare and S. triloba extracts was determined. To achieve the aim of the study, aqueous, ethanolic, and enzymatic extracts were prepared and screened for phytochemical capacity and antioxidant activities. Determination of the antibacterial activity included phenotypic screening of antibiotic susceptibility pattern of oral and food pathogenic bacterial strains, determination of Minimum inhibitory concentration and minimum bactericidal concentration - Microdilution broth test and in vitro valuation of antibacterial efficacies - anti-biofilm properties of the studied herbal extractions. Results: Our study evaluated the phytochemical composition and the antioxidant, antibacterial, and anti-biofilm properties of O. vulgare and S. triloba extracts. The analyzed samples contained bioactive compounds, such as phenolics and flavonoids, contributing to the observed strong antioxidant effect. Furthermore, they exhibited notable activity against oral biofilm formation and demonstrated significant antibacterial efficacy against dental caries microorganisms as well as food pathogens. Despite methodological variations, all extracts showed significant antioxidant capacity and promising antibacterial activity against various pathogens, including resistant strains, while also inhibiting biofilm formation. Although limited to two plant species and facing methodological constraints, the study lays a groundwork for future research, indicating the therapeutic potential of O. vulgare and S. triloba extracts. Further exploration is needed to report on underlying mechanisms and validate efficacy through clinical trials.
Keywords:
Origanum vulgare
; Salvia triloba extracts
; phytochemistry and biological capacity
; oral cariogenic and food-origin pathogenic bacteria
1. Introduction
Since ancient times, humans recognized the capacity of plants to generate chemical substances essential for the survival of all organisms inhabiting planet Earth [1]. Both the cause of Socrates' death and the development of active concentrations of salicylic acid (aspirin) involve these chemicals and they are among at least 100,000 chemical substances found in plants [2].
Phytochemistry divides plant metabolic products into two main categories: primary metabolites and secondary metabolites [3]. Although the conventional categorization of plant-derived organic compounds into primary metabolites, secondary metabolites, and hormones has been valuable over the years, recent studies have shed light on the complex interplay among different classes of plant secondary metabolites and plant metabolism [4]. Secondary metabolism accounts for 10% of a plant's total metabolic processes and involves biosynthetic pathways that produce molecules not directly involved in essential cellular functions, such as growth and photosynthesis [5]. These pathways play a variety of ecological roles including the establishment of symbiotic relationships, pollination, interspecies competition, and chemically mediated plant-plant interference (i.e., allelopathy). Contrary to earlier beliefs that deemed secondary metabolites merely as unnecessary by-products of primary metabolism, current understanding recognizes them as critical extensions of primary metabolism [6]. These compounds fulfill numerous ecophysiological functions, such as defending plants against pathogens and herbivores, helping them manage abiotic stress, and aiding in reproduction and seed dispersal through mechanisms like pollinator attraction and allelopathy [7].
Secondary metabolites exhibit a wide range of diversity and are categorized into several major groups based on their biosynthetic pathways [8]. Phenolic compounds constitute one such group, characterized by the presence of at least one aromatic ring, synthesized through the shikimic and/or malonic acid pathways [4]. Terpenes and steroids, another category, are produced via acetyl coenzyme A through the mevalonate pathway in the cytoplasm and the pyruvate-phosphoglycerinaldehyde pathway in plastids [8]. Nitrogenous secondary metabolites primarily derive from amino acids, comprising the third group. Lastly, phytohormones, also recognized as plant hormones, play crucial roles in regulating various organismal processes and cellular activities within plants. These hormones, belonging to different chemical groups, including indole derivatives, sesquiterpenes, alkenes, diterpenoid acids, aliphatic nitrogenous bases, phenolic organic acids, and terpenoid lactones, orchestrate diverse biological functions and biosynthesis within plants [9]. Understanding the concepts outlined above naturally prompts the inquiry into how chemicals synthesized by one organism might impact or be harnessed by another belonging to other classes. This line of thought gives rise to the idea of utilizing plants in their entirety or extracting specific chemicals from them to create formulations with antimicrobial properties, as well as antioxidant attributes [10].
Moreover, the exploration of the oral microbiota is an ever-evolving and captivating field of study [11,12]. Its importance transcends oral health alone, as numerous oral diseases, including dental caries and periodontitis, are linked to various non-communicable conditions such as diabetes, cardiovascular diseases, pneumonia, obesity, cancers, and premature birth [13,14]. Distributed within biofilms throughout the oral cavity, the oral microbiome forms a plethora of micro-ecosystems crucial for maintaining health equilibrium [15]. Any disruptions to this delicate balance can pave the way for the emergence of pathogens and subsequent disease [16]. Moreover, serving as the primary gateway to the gastrointestinal system, the oral cavity acts as the initial point of entry for xenobiotics into the body [17]. Imbalances in the diverse microecosystems within the oral cavity can lead to dysbiosis, highlighting the imperative of identifying the microbiome in a healthy state [18]. Therefore, there is an urgent need to investigate chemical substances derived from natural sources, as well as to design synthetic or semi-synthetic compounds that can act as regulators of biological processes such as inflammation and oxidative stress within the oral cavity. These compounds should also serve as effective antimicrobial agents against oral (especially cariogenic) pathogens.
Phytochemical research is expected to reveal undiscovered biomolecules from which pioneering remedies could develop [19,20,21]. This expectation becomes especially relevant as the threat of antibiotic resistance posed by pathogenic bacteria escalates significantly each year [22,23,24,25,26]. It is also true that the pharmaceutical industry faces serious economical, regulatory and scientifical difficulties to develop new classes of antibiotics [27]. Developing novel strategies to combat antibiotic resistance is now paramount [21]. Under these dire perspectives, plant extracts are recognized as a unique and valuable source of natural substances useful for drug discovery and advancement [20,28,29]. In the present research, we focus on the in vitro biological evaluation of the extracts of two plants: Origanum vulgare (O. vulgare) obtained from the mountainous regions of Epirus, Greece, and Salvia triloba (S. triloba), commonly known as Greek sage, originating from the island of Crete, Greece. The aim of this study is to conduct a screening involving the qualitative analysis of these plant extracts to identify their antioxidant, antibacterial, and anti-inflammatory actions.
2. Materials and Methods
The schematic representation of the experimental flow setup is depicted in Schema 1. An individual text field for explanations is provided below, primarily focusing on the methods or parameters requiring further clarification.
Scheme 1.
The schematic representation of the experimental flow setup.
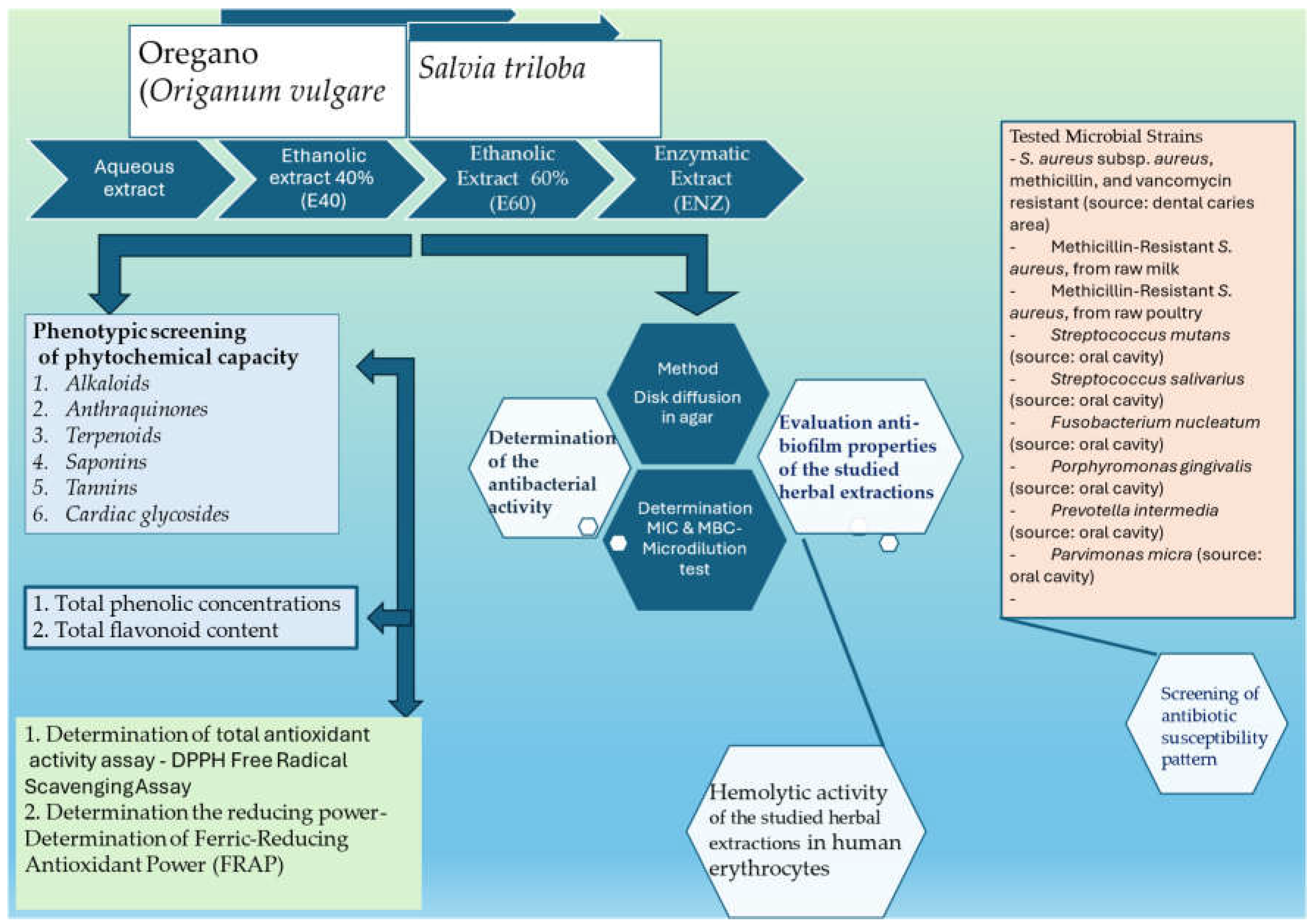
2.1. Plant Materials
The present study examined two plant species: (a) Wild oregano (Origanum vulgare) from the mountainous regions of Epirus, Greece; and (b) Greek sage (Salvia triloba) from Crete. Samples of these plants, gathered by locals, were air-dried at room temperature and then divided into distinct parts, including flowers, roots, leaves, bark, and stems. After complete drying, the samples were ground into a fine powder using a high-speed grinder, focusing on the leaves and flowers only. The powdered plant materials were stored at -18°C for further analysis.
2.1.1. Preparation Methods for Plant Extracts
The maceration method was employed for the preparation of the Aqueous, Ethanolic, and Enzymatic extracts [30].
2.1.2. Aqueous Extract (A)
Each plant powder was mixed with distilled water (DIW) at a ratio of 1:2 (w/v) and suspended in sterile flasks with continuous stirring at 30°C for 24 hours. Afterward, the mixture was filtered through sterilized Whatman No.1 filter paper, and the resulting liquid extracts were stored at -80°C for 24 hours. Concentration was achieved through evaporation under reduced pressure using a rotary evaporator (KNFRC 900, KNF Neuberger GmbH, Breisgau, Germany). Aqueous extracts underwent lyophilization in a freeze dryer for approximately 7 hours or overnight. Prior to use, these extracts were exposed to ultraviolet light overnight to eliminate potential microbial contaminants [1]. The letter 'A' was used as an abbreviation for convenience.
2.1.3. Ethanolic Extract (E)
To obtain ethanol extracts, we followed the procedure outlined in section 2.1.2, with the only difference being the use of an ethyl alcohol solution as the solvent instead of DIW. Two concentrations of 96% ethyl alcohol solution were used: 40% and 60%, both prepared from DIW [1]. The abbreviations used for the ethanolic extracts are E40 and E60, representing 40% and 60% concentrations, respectively.
2.1.4. Enzymatic Extract (ENZ)
One kilogram of pretreated fresh plant material (leaves and flowers) was immersed in a solution consisting of 2 kilograms of acidified DIW adjusted to pH=2 using concentrated hydrochloric acid and 1.0% pepsin (Merck KGaA, Darmstadt, Germany) [24]. After an incubation period at 37°C for 48 hours, hydrolysis was stopped by heating for 10 minutes. The resulting solution was divided into smaller 200-gram batches, manually compressed with a sterile pestle, and filtered through sterilized Whatman No.1 filter paper. Subsequently, the solvent from each batch was evaporated using a rotary evaporator (KNFRC 900, KNF Neuberger GmbH, Breisgau, Germany). The final step involved deep freezing at -80°C followed by lyophilization [1,31].
Keynotes for the pretreatment process: The plant material underwent several steps, including washing under running water, manual removal of unsuitable elements, and cutting into 1 mm pieces using a slicer to increase surface area. These pieces were then washed with phosphate-buffered saline (PBS) to eliminate intracellular vesicles released from damaged cells [1,32]. The abbreviation "ENZ" was used. Samples were prepared by weighing the crude extracts mentioned above and determining the volume of solvent [5% aqueous solution of dimethyl sulfoxide (DMSO)] needed to create a sample stock solution with a concentration of 10 mg/ml. Aqueous extracts were dissolved using sterile distilled water.
2.2. Screening the Phytochemical Capacity and Antioxidant Activities of Plant Extracts
2.2.1. Phytochemical Identification
To evaluate the phytochemical composition of the plant extracts, we conducted the following tests.
Clarification: For qualitative phytochemical screening, we prepared two types of aqueous extract solutions. The first was dissolved in sterile boiled water, designated as "A," while the second, an aqueous extract dissolved in a solvent composed of a 1:1 ratio of boiled water and 95% methanol, was labeled as "A*". All subsequent experiments were conducted in triplicate.
- -
-
The alkaloids in plant extract were identified, using (a) the Mayer’s reagent, (b) the Wagner’s reagent and (c) the Hager’s reagent (Picric acid test) as per the previous reports.
- -
-
In summary, fifty milligrams of the extract were dissolved in 5 mL of distilled water (DW) and then filtered. Two milliliters of the resulting extract solution were transferred to a test tube containing 10 mL of benzene. The mixture was vigorously shaken for 10 minutes and then filtered. Finally, 5 mL of a 10% ammonia solution was added to the test tube and shaken vigorously for an additional 30 seconds. The presence of free anthraquinones was indicated by the development of a pink, red, or violet color, evaluated based on a positivity rating.
- -
-
Detection of terpenoidsTo detect the presence of tri-terpenoids, the Salkowski test was conducted using chloroform and concentrated sulfuric acid (H2SO4) as reagents.
- -
-
Detection of saponins (Foam test) [32]Saponins were identified by the formation of persistent foam at 25°C. To achieve this, 500 milligrams of the extract were dissolved in 2 mL of distilled water. The resulting suspension was vigorously shaken for 15 minutes in a graduated cylinder. The presence of saponins was confirmed by the formation of a 2 cm layer of foam.
- -
-
Detection of tannins (Ferric chloride test - Braymer’s test) [33]To assess tannin content, a mixture was prepared from each plant extract. This involved dissolving 500 milligrams of the extract in 5 mL of ethanol, followed by sonication at 40 kHz for 5 minutes using a sonicator, and centrifugation at 190× g for 10 minutes. Subsequently, 1 mL of supernatant was collected. Then, 1 mL of 15% ferric chloride (FeCl3) was added, and the formation of a dark green or blue-black precipitate indicated the presence of tannin.
- -
-
milligrams of the extract were dissolved in distilled water (DW) and subsequently filtered. To 2 mL of the filtrate, 1.5 mL of glacial acetic acid, a single drop of 5% ferric chloride (FeCl3), and a drop of concentrated sulfuric acid (H2SO4) along the inner walls of the test tube were added. The transition to green-blue coloration at the upper layer and reddish-brown at the interface between the two layers confirmed the presence of cardiac glycosides.
2.2.2. Total Phenolic Concentrations [34]
The concentration of total phenolic compounds in the plant extracts was assessed using the Folin–Ciocalteu method, following the procedure outlined in our previous study. Folin–Ciocalteu reagent (120 µL) was mixed with 15 µL of each plant extract (final concentrations ranging from 4–40 µg/mL in methanol) in a 96-well plate. After 5 minutes, 120 µL of Na2CO3 solution (60 g/L) was added to each well and mixed. The plate was then incubated in darkness for 90 minutes before measurement. Absorbance was recorded at 725 nm using a microplate reader. The total phenolic content was determined from the standard curve of gallic acid in methanol, with a final concentration range of 2–40 µg/mL. Finally, the total concentration of phenolic compounds was expressed as gallic acid equivalents (GAE) in milligrams per gram of dry weight of the sample.
2.2.3. Estimation of Total Flavonoid Content
The total flavonoid content was determined using the aluminum chloride assay, with catechin serving as the standard for expressing total flavonoid content in milligrams of catechin equivalents (CE) per gram of dry weight of the sample (mg CE/g of DW) [34,35]. The total flavonoid content in the extracts was determined as follows: A volume of 0.50 mL of each plant extract was added to a 10 mL volumetric flask containing 4 mL of deionized water (DIW). To this flask, 0.3 mL of 5% NaNO2 was added. After 5 minutes of incubation, 0.3 mL of 10% AlCl3 was added. At the 6th minute, 2 mL of 1 M NaOH was added, and DIW was added to reach the mark. An orange-yellowish color developed. After 10 minutes of incubation, the absorbance was measured at 510 nm.
2.2.4. Total Antioxidant Activity Assay [35] (DPPH Free Radical Scavenging Assay)
The 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging assay was employed to assess the in vitro antioxidant activities of plant extracts, as detailed in our previous report.
2.2.5. The Reducing Power Assay
The evaluation of the reducing power of the tested plant extracts is based on the principle that molecules with the potential to undergo reduction react with potassium ferricyanide (Fe3+), generating potassium ferrocyanide (Fe2+). This resulting compound then reacts with ferric chloride, forming a ferric ferrous complex that exhibits an absorption peak at 700 nm. A greater absorbance of the reaction mixture signifies a higher reducing power [1,35].
Clarification: Each sample underwent triplicate testing for all the assays mentioned in Section 2.2.
2.3. Determination of the Antibacterial Activity
2.3.1. Tested Bacterial Strains
In this study, we examined nine (9) bacterial strains isolated from lesions of the oral cavity and food sources. These strains display diverse characteristics and have varying requirements for growth, nutrient substrates, and optimal incubation conditions.
The strains of the pathogenic bacteria that were tested as cell- targets, are mentioned in Table 1.
All the mentioned strains were identified and classified using standard laboratory procedures from the protocols followed by the Departments of Medicine and Dentistry at the National and Kapodistrian University of Athens. In our present study, we introduced bacteria with diverse metabolic growth processes and nutritional requirements. As a result, we employed a variety of nutrient media and incubation conditions, as detailed in our previous publication [1].
2.3.1.1. Phenotypic Screening of Antibiotic Susceptibility Pattern of Bacterial Strains
Antibiotic susceptibility tests were conducted for all tested bacteria using the Kirby-Bauer disc diffusion method, following the guidelines set forth by The National Committee for Clinical Laboratory Standards (later renamed The Clinical Laboratory Standard Institute-CLSI). According to relevant clinical guidelines, the following antibiotics were included in the analysis of antimicrobial profiles [36,37,38,39]: β-lactams [(amoxicillin/clavulanic acid; AMC, 20/10 µg), aminopenicillins (amoxicillin; A, 30 µg), especially for Staphylococcus aureus strains, markers for methicillin resistance (oxacillin, flucloxacillin)], Glycopeptide (Vancomycin; VA, 30 µg), second-generation cephalosporins (cefuroxime; CFX, 30μg), third-generation cephalosporins (cefotaxime; CFT 30μg), clindamycin (CLI, 2μg), Aminoglycosides (Gentamicin; GEN, 10μg), macrolides (erythromycin; ERY, 15μg), Tetracycline (TER, 30μg), fluoroquinolones (ciprofloxacin; CIP, 5μg), carbapenems (Imipenem; IMI, 10μg), and nitroimidazole (Metronidazole; MET, 5 µg).
Furthermore, utilizing the reference broth microdilution method, we determined the Minimum Inhibitory Concentration (MIC) values of the aforementioned antimicrobial agents against the tested bacterial strains, adhering to the guidelines of EUCAST and CLSI for each essential oil (EO) and reference antimicrobials [39,40,41,42,43].
2.3.2. 1st Assay for Evaluation of the Antibacterial Activity - Diffusion in Agar Test
We initiated the assessment of the antibacterial efficacy of the studied extracts using disc diffusion in agar assays [40]. Overnight cultures of each bacterial strain were prepared and plated onto agar petri dishes, with specific media selected based on the growth requirements of each strain. Dried Mueller–Hinton agar and Brain Heart Infusion agar were employed for Staphylococcus and Streptococcus strains, respectively, while Brucella Agar supplemented with 5% Sheep Blood, hemin, and menadione was utilized for obligate anaerobic isolates.
Sterile filter paper discs (6 mm diameter, Whatman No.1) saturated with herb extracts at concentrations ranging from 10% to 100% (v/v) were carefully placed on the surface of each plate using sterile forceps. Aqueous extracts were diluted with sterile distilled water, while discs loaded with sterile distilled water served as negative controls. After allowing the plates to incubate at room temperature for 2 hours to facilitate bacterial diffusion into the agar media, they were then placed in a 37°C incubator overnight for S. aureus strains, precisely 36 hours for Streptococcus strains, and at least 72 hours for obligate anaerobic strains. Anaerobic conditions were meticulously maintained throughout the incubation period for obligate anaerobic strains using anaerobic rectangular jars equipped with Anaerocult A gas packs. At the end of the designated incubation period, the zone of inhibition, commonly referred to as the "halo," was meticulously assessed, with each experiment meticulously conducted in triplicate.
2.3.3. 2nd Assay for Evaluation of the Antibacterial Activity - Determination Minimum Inhibitory Concentration and Minimum Bactericidal Concentration - Microdilution Broth Test
To determine the minimum inhibitory concentration (MIC) using the microdilution broth method in 96-well microplates [1,44,45], plant extracts were initially diluted to 400 mg/mL crude extraction in a 5% aqueous solution of dimethyl sulfoxide (DMSO), except for aqueous extracts, which were diluted in ultrapure water. Serial dilutions were then prepared using ultrapure water at concentrations ranging from 200 mg/mL to 0.0975 mg/mL. The bacterial inoculum was standardized to McFarland scale standard 1, and aerobic or facultative anaerobic strains were inoculated in double-strength Mueller-Hinton broth (MHB), while anaerobic bacteria were cultured in sBHI broth supplemented with 5 μg/mL hemin and 1 μg/mL menadione. Incubation conditions followed those detailed in the preceding sections. To validate the procedure, control wells containing only the liquid medium, the medium with inoculum, or chlorhexidine (CHX) 0.2% were included. Following incubation, the liquid media in each well were stained with an aqueous resazurin solution (Sigma-Aldrich, St Louis, MO, USA) at a concentration of 0.02%. The 96-well microplates were then re-incubated for an additional two hours. The MIC was determined as the lowest concentration where the resazurin staining remained blue, indicating inhibition. A color change to pink-purple or pink indicated resazurin reduction and bacterial growth. All experiments were conducted in triplicate. To ascertain the Minimum Bactericidal Concentration (MBC), a 20-μL aliquot from wells that exhibited no growth or from the well corresponding to the MIC reading was transferred onto plates containing Mueller-Hinton Agar or, for anaerobic bacterial strains, 5% defibrinated sheep blood Brucella agar enriched with 5 μg/mL hemin, 1 μg/mL menadione, and 2 g/L yeast. The plates were then incubated at 37°C for 24 hours, 36 hours, and 37 hours for S. aureus strains, facultative anaerobic Streptococcus strains, and obligate anaerobic isolates, respectively. Colony growth on the plates was confirmed at the end of this period. MBC was defined as the lowest extract concentration resulting in complete bacterial elimination.
2.4. In Vitro Evaluation of Antibacterial Efficacies - Anti-Biofilm Properties of the Studied Herbal Extractions
To assess the biofilm-forming ability of the bacterial isolates [46,47,48], we utilized a semi-quantitative approach employing collagen type I-coated 96-well flat-bottom microplates (Thermo Scientific™Nunc™, Waltham, MA, USA). Bacterial cultures were prepared in specific broths tailored to their growth requirements: Trypticase Soy Broth supplemented with 1% glucose (TSBG) for aerobes or facultative anaerobes, and Tryptic Soy Broth supplemented with yeast extract, L-cysteine hydrochloride, hemin, and menadione for obligate anaerobic isolates. Each well of the microplate received 100 μL of bacterial suspension (adjusted to a turbidity of 10^8 cfu/mL) and was incubated either aerobically at 37°C for 24-36 hours or anaerobically for a minimum of 48 hours. Following incubation, the wells were rinsed twice with Phosphate-Buffered Saline (PBS, pH 7.4) and fixed with absolute methanol. The fixed bacterial cells were then stained with 0.1% crystal violet dye for 30 minutes, washed to remove excess dye, and solubilized with 33% glacial acetic acid. Absorbance at 595 nm was measured to quantify the attached cells, with negative control wells containing only TSB. Each experiment was conducted in triplicate, and the average absorbance value was compared with the cut-off value (ODc), set as three standard deviations above the mean OD of the negative control.
Based on these findings, the isolates were categorized as non-biofilm producers (OD≤ODc), weak biofilm producers (ODc < OD≤2×ODc), moderate biofilm producers (2×ODc < OD≤4×ODc), or strong biofilm producers (4×ODc < OD).
To assess the anti-biofilm properties of each studied herbal extract, we followed the procedure outlined just above, with the exception that the bacterial suspension in each well was co-cultured with sub-MIC concentrations of the plant extracts as treatment. Wells without any plant extract served as the control samples. The anti-biofilm activity as a percentage (%) of inhibition was calculated using the following Equation (1) [49]:
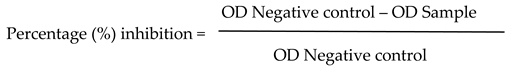
Based on the results, the tested herbal extracts were classified as: Excellent (++++) AB activity (> 95% inhibition); Very Good (+++) AB activity (> 95-80 % inhibition); Good (++) AB activity (> 80-50 % inhibition); Poor (+) AB activity (more than 0–50% inhibition); No (-) AB activity (0 % or less). AB meaning: Anti-biofilm
2.5. Hemolytic Potential of Tested Plant Extracts in Human Erythrocytes
Blood samples (AB type) were obtained from the local hospital, sourced from blood bags with EDTA, slated for disposal due to incomplete blood collection. The blood was centrifuged at 1500 rpm (15 minutes at 25°C) to separate erythrocytes from plasma and then washed thrice with 10 mL of PBS (pH 7.4). The erythrocytes were suspended in PBS, and subsequent steps involved multiple washes with 0.2 M PBS (pH 7.4) and centrifugation at 300 rpm for 10 minutes, followed by resuspension in saline solution (0.9% NaCl). After thorough washing, the erythrocytes were diluted 1:100 in PBS pH 7 to yield a 1% erythrocyte suspension.
To assess the hemolytic activity of the extracts, various concentrations ranging from 12.50 to 1000 μg/mL were mixed with NaCl solution (0.85%), and a 2% suspension of human erythrocytes was added. The mixture was then incubated at 37°C for 60 minutes. Absorbance was measured over time, and hemolysis rates were calculated as a percentage of total hemolysis after one hour.
A negative control comprising only erythrocytes (500 mL of erythrocyte suspension and 1500 mL of PBS buffer, without extract) represented 0% hemolysis, while a positive control included 1% Triton X-100 (hemolyzing agent), indicating 100% hemolysis [50]. The absorbance of each tube was measured at 540 nm (the absorbance of hemoglobin) using a UV-visible spectrophotometer against a blank containing PBS. Hemolysis percentage was calculated using the following equation (2) [51]:
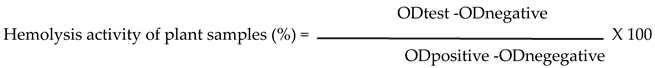
2.6. Statistical Analysis
When applicable, means of the various observations between groups (i.e. plant extracts) were compared by using ANOVA (for normally distributed data after the Shapiro-Wilk test) followed by Tukey’s HSD post hoc comparison at an alpha of 0.05. All statistical evaluations were performed with SPSS v28 (IBM Corp. Armonk, NY).
3. Results
As shown in Table 2, most abundant phytochemical compounds were found in the aqueous extracts from both plant species and in enzymatic extract from O. vulgare.
All plant extracts were rich in total phenolics with statistically significant differences among their concentration (Table 3) either in each one of the specie or between them (Figure 1). Among them, higher values were detected in oregano enzymatic extract (122.4±0.6 mg GAE/g), salvia aqueous extract (91.99±1.08 mg GAE/g), salvia enzymatic extract (90.42±0.5 mg GAE/g) and oregano 60% ethanolic-aqueous extract (88.44±0.6 mg GAE/g). Similarly, total flavonoids content was increased in salvia and oregano enzymatic extracts (64.75±0.65 mg CE/g and 48.83±1.17 mg CE/g respectively).
In concentrations above 100 μg/mL increased scavenging activity was observed in all extracts from both plant species (Table 4). However, the highest values were recorded again in enzymatic fractions with 86.43% and 87.14% for Salvia and Oregano respectively.
Similarly, increased plant extracts produced increased reducing power values in FRAP assay (Table 5) with plant extracts be comparable but now statistically significant with the values produced by the two reference compounds. However, in 25 μg/mL concentration, the absorbance of oregano enzymatic extract reached 1.93 which was higher than gallic and ascorbic acid.
The report on the antibiotic susceptibility patterns of the studied isolates depicts a varied landscape in terms of the recorded zone of inhibition: Amoxicillin (30 μg) produced zones of inhibition from 9 mm (P. intermedia) to 36 or 37 mm (S. salivarius, S. mutants). Amoxicilin with clavulanic acid from 24 – 35 mm, vancomycin (30 μg) 18 to 27 mm, imipenem (10 μg) 10 to 35 mm, erythromycin (15 μg) from 0 to 26 mm, clindamycin (2 μg ) from 0 to 29 mm, gentamycin (10 μg) from 0 to 266 mm, tetracycline (30 μg) from 0 to 28 mm, ciprofloxacin (5 μg) from 0 to 52 mm, metronidazole (5 μg) from 21 to 27 mm, cefuroxime (30 μg) from 10 to 30 mm and cefotaxime (30 μg) from 8 to 31 mm. Although most of the strains were susceptible or presented a medium susceptibility in common antibiotics, some pathogens (F. nucleatum, P. gingivalis, P. intermedia and P. micra) were proven multiresistant in amoxicillin, amoxicillin & clavulanic, vancomycin, imipenem, erythromycin, tetracycline and ciprofloxacin with MIC values ranging from 0.25 to 128 μg/mL and only P. micra against gentamycin reached 256 μg/mL The two out the three S. aureus isolates were resistant in 7 from 10 antibiotics and only the strain isolated from raw milk proven resistant in just 2 (amoxicillin and amoxicillin/clavulanic acid). In disk diffusion experiments against the oral pathogens, all plant extracts exhibited a variable antibacterial activity with zones of inhibition from 6 to 38 mm (Table 6 and Table 7). In general, the aqueous and ethanolic (E20) O. vulgare extracts were comparable with those of S. triloba while oregano 60% ethanolic-aqueous and S. triloba enzymatic extracts produced wider zones indicating their effectiveness. Most effective was the 60% ethanolic-aqueous extract from oregano in 100% disk content and similarly the 60% ethanolic-aqueous and the enzymatic extracts from S. triloba. As shown in Figure 2, concentrated plant extracts produced wider zones of inhibition from most of the antibiotics.
In general, low hemolytic activities (0.1% to 9.04%) were observed with plant extract concentrations up to 100 μg/mL for O blood type erythrocytes (Table 10). However, the largest activities were observed with Oregano E40 and E60 extracts as concentration further increased. When these two particular extracts were tested at their highest concentration of 1000 μg/mL, hemoglobin release reached to considerable percentages of appx. 55% and 83% respectively compared to Triton X-100 positive control. No similar activity was detected for the S. triloba extracts (Figure 5 and Figure 6).
4. Discussion
In our study, qualitative phytochemical assays revealed rich profiles in flavonoid, tannins, glycosides, and terpenoids in O. vulgare samples, present across all examined extracts. Alkaloids and steroids were exclusively detected in aqueous and enzymatic extracts, respectively [52]. Conversely, S. triloba exhibited similar findings, except for the absence of glycosides in enzymatic extracts and no steroids isolated from any extract. Alkaloids and anthracocyanins were exclusively isolated from the aqueous extract in S. triloba, while anthraquinones were only detected in O. vulgare's enzymatic extract. Aqueous extraction effectively retrieved most phytochemicals in both plants, with enzymatic extraction notably enhancing the appearance of certain compounds, particularly in oregano (Knez Hrnčič et al., 2020) [53]. These results reveal the multifaceted influence of extraction methods, analytical techniques, and plant characteristics on phytochemical profiles [54,55,56,57,58,59]. Additionally, O. vulgare essential oil contains a diverse array of terpenes, while S. triloba oil is rich in bicyclic oxygenated monoterpenes. Both plants' infusions and alcoholic extracts exhibit significant polyphenol and flavonoid content, along with di- and triterpenoids [60,61,62].
4.1. Pattern of Total Phenolics and Flavonoids Content
In O. vulgare, significant variability in total polyphenol content (TPC) was observed among different extracts, with the enzymatic extract showing the highest TPC followed by various ethanol extracts and finally, the aqueous extract, which had the lowest concentration. Similar trends were noted for flavonoid values. These TPC values are consistent with findings from other researchers. For instance, Jafari Khorsand et al. [63] reported TPC ranging from 20.3 to 35.5 mg GAE g−1 DW, while Yan et al. [64], noted values of 79–147 mg GAE g−1 DW in the hydro-methanolic extract of O. vulgare. Finally, Bower et al. reported a notable total phenolic content of 430 µg of GAE mg−1 DW for the methanolic extract of O. vulgare leaves [62]. Concerning S. triloba, higher TPC values were observed in the aqueous and enzymatic extracts compared to ethanol extracts. Researchers calculated TPC and total flavonoid content (TFC) in S. officinalis and S. triloba extracts, with values ranging from 193.50 ± 8.22 to 203.01 ± 7.85 GAE mg/g extract for TPC and 71.51 ± 1.88 to 78.84 ± 8.76 QE mg/g extract for TFC [65]. Furthermore, ultrasound-assisted extraction (UAE) extracts exhibited significantly higher values compared to pressurized liquid extraction (PLE) extracts for all photometric determinations. Aqueous methanol mixtures were found to be more effective for UAE extraction of phenolics and flavonoids compared to pure water or methanol [66].
4.2. Evaluation of Antioxidant Capacity: DPPH Assay and Ferric Reducing Assay Power (FRAP)
The DPPH assay is commonly used to evaluate the antioxidant properties of plant extracts, providing insights into their phenolic and flavonoid content [67]. Our study confirmed a dose-dependent scavenging activity, with both Salvia and Oregano showing enzymatic fractions exhibiting the highest activity. A cut-off concentration limit of 50 μg/mL marks the onset of scavenging activity toward DPPH, with percentages reaching 86.43% and 87.14% for Salvia and Oregano, respectively, at concentrations of 500 μg/mL (Table 4), respectively. This trend persisted across ethanolic and aqueous extracts, with the lowest activity observed in the latter. This indicates that variations in the free radical scavenging activity of extracts are strongly linked to both their chemical composition and the levels of total phenols and flavonoids, which are also concentration dependent [68,69]. High levels of Total Phenolic Content (TPC) and Total Flavonoid Content (TFC) were observed in Oregano' extracts, supporting previous research suggesting a correlation between antioxidant activity and phenolic/flavonoid levels [70,71]. In the case of S. triloba, high TPC and TFC levels were detected in both aqueous and enzymatic fractions, although aqueous fractions displayed lower DPPH radical neutralization. These variations may be attributed to the unique antioxidant composition within the plant [59]. Consistent with previous studies, Salvia extracts exhibited dose-dependent antioxidant activity in the DPPH test [59,65]. Research by Jan et al. [64] also demonstrated dose-dependent antioxidant activity in O. vulgare extracts, with higher IC50 values observed in wild accessions compared to cultivated ones. FRAP assay results mirrored these findings, with increased extract concentrations correlating with higher reducing power (Table 5). Notably, in our study, Oregano's enzymatic extract exhibited superior antioxidant activity compared to reference compounds. Overall, our findings emphasize the significant antioxidant potential of the extracts tested, irrespective of extraction methods. The capacity of constituents to reduce is a key indicator of their potential antioxidant activity. The Ferric Reducing Antioxidant Power (FRAP) assay relies on the transition from yellow to green, which reflects the sample's ability to reduce a Fe3+/ferricyanide complex to Fe2+. Subsequently, the percentage of Fe2+ reduced can be quantified by measuring the absorbance at 700 nm [72]. In conclusion, our research findings indicate that regardless of the differences in extraction methods, all types of extracts demonstrated significant antioxidant capacity.
4.3. Antimicrobial and Anti-Biofilm Activity of the Studied Extracts
4.3.1. O. vulgare Performance
While no distinct antibacterial pattern was discerned, all plant extracts exhibited variable antibacterial activity, with zones of inhibition ranging from 6 to 38.7 mm. The most effective extract was the 60% ethanolic-aqueous extract from oregano at 100% disk content. Compared to synthetic antibiotics, plant extracts generally produced wider zones of inhibition, particularly when compared to erythromycin. The analyzed extracts exhibited zones of inhibition against the pathogen S. aureus, methicillin and vancomycin, resistance ranging from approximately 15 to 20 mm. Notably, the Ethanol 60% extract displayed the strongest inhibitory properties, followed by the enzymatic extract, the Ethanol 40% extract, and the aqueous extract. Analyzing the inhibition zones for all tested microorganisms revealed that extracts with concentrations exceeding 50% exhibited satisfactory inhibition results, spanning approximately from 15.2 mm to 38.7 mm. Given the observed variation, it could be argued that the extracts possess antibiotic activity akin to pharmaceutical substances. However, since the extracts contain unidentified substances in unspecified concentrations, their effect might be synergistic or even antagonistic effect. In the latter scenario, there is a possibility of a potent substance being inhibited by another, which presents a significant limitation of this study. This presents a significant limitation of the study. All extracts in our study, showed effectiveness against the pathogens, with MIC values ranging from 0.4 to 12.5 μg/mL and MBC values from 3.1 to 50 μg/mL. Particularly, the enzymatic and 60% ethanolic-aqueous extracts of oregano displayed the lowest mean MIC values (ranging from 0.39±0 to 3.12±0 μg/mL) against the tested pathogens. Among the strains, MRSA isolated from poultry, P. gingivalis, F. nucleatum, P. intermedia, and P. micra were the most sensitive to oregano extracts. The MIC results complement the earlier findings on the minimum zone of inhibition, providing a more detailed assessment of the inhibitory potential of the extracts under study. Once again, the 60% ethanol extract stood out as a potent inhibitor, displaying strong inhibitory capacity even at minimal concentrations. Following closely was the enzymatic extract, while the aqueous extract ranked lowest among the four, although it exhibited a remarkably low concentration requirement for disruption. Ultimately, it becomes apparent that most extracts exhibited comparable or even stronger antimicrobial efficacy when contrasted with the effectiveness of conventional antibiotics.
The antibiofilm activity was assessed using a four-point scoring system based on the inhibition of biofilm formation: poor (0-50% inhibition), good (50-80% inhibition), very good (80-95% inhibition), and excellent (> 95% inhibition), and the results, along with the scores of the seven antibiotics, were depicted in Figure 3. In general, antibiotic substances displayed broad and potent antibiofilm efficacy against all pathogens, with vancomycin demonstrating the highest effectiveness. Nevertheless, extracts derived from O. vulgare, especially the enzymatic and 60% ethanol-aqueous extracts, showcased significant antibiofilm properties comparable to those of conventional pharmaceuticals. The aforementioned findings corroborate previous literature suggesting that ethanolic extracts demonstrate superior antimicrobial activity compared to aqueous extracts against a diverse array of oral pathogens, including Streptococcus, Enterococcus, and Lacticasei bacillus [21]. Moreover, researchers have documented that the methanol extract of O. vulgare effectively eradicated tested oral bacteria, exhibiting minimum bactericidal concentration (MBC) values ranging from 0.30 mg/ml for P. micra to 5.00 mg/ml for E. faecalis, S. mutans, and S. sobrinus. The most pronounced inhibitory effects were observed at an O. vulgare concentration of 0.30 mg/ml for P. gingivalis and P. micra, and 0.60 mg/ml for S. oralis, respectively [73]. Considering the commendable activity against biofilm production exhibited by both enzymatic and ethanol extracts, it is evident that these extracts hold promise and warrant further investigation.
4.3.2. S. tribola Performance
The study investigated the antimicrobial efficacy of ethanolic (40% & 60%), aqueous, and enzymatic extracts of S. triloba against both oral and non-oral pathogens. Our results highlight significant zones of inhibition, particularly with the ethanolic 60% extract, showing values such as 29.9 mm against MRSA, 30.9±0.3 mm against P. micra, and 28.9 mm against F. nucleatum. The ethanolic extract exhibited robust antibacterial activity, closely followed by the enzymatic extract. Regarding MIC values, the 60% ethanolic extract displayed exceptionally low values, with the 40% ethanolic extract in proximity. Notably, the aqueous extract showed significant variation among different microorganisms, with a MIC value as low as 0.8 for P. gingivalis. The findings regarding biofilm inhibition closely resemble those seen with the oregano plant, with both the enzymatic and 60% ethanolic extracts showing impressive outcomes, akin to antibiotics. Overall, all extracts demonstrate excellent antibiofilm activity against the tested isolates. Previous studies primarily focused on essential oils or highly concentrated ethanol extracts, whereas our investigation adopts a broader approach by encompassing various types of extracts and a range of pathogens from both oral and food sources. Although existing studies mainly examined Salvia officinalis, we aimed to align our experimental protocol as closely as possible. Popa et al. [74] for example, investigated the antimicrobial effects of volatile oils from S. officinalis against bacterial strains collected from individuals with oral diseases, with MIC values ranging between 0.00 and 45.9 mg/mL Other studies reported MIC values for essential oils from S. officinalis and S. triloba against foodborne bacteria, ranging from 0.3 to 10.0 mg/mL [75]. Additionally, inhibition zones against S. aureus were reported for various extracts from S. fruticosa, with MIC values ranging from 50 to 15 mg/mL [76]. Evaluation of the glycolic extract of S. officinalis against S. mutans and other bacterial species revealed effective elimination at a concentration of 50.0 mg/mL [77].
4.4. Hemolytic Activity
The in vitro hemolytic test was commonly used in the pharmaceutical industry to screen therapies throughout the early stages of clinical development [66,78]. The occurrence of erythrocyte hemolysis could be an indicator of cell oxidative damage that leads to cell death. Additionally, the lysis of erythrocyte membranes is a common side effect of chemotherapeutic drugs. Co-administration of plant-derived agents is among the potential strategies to mitigate chemotherapeutic-induced hemolysis [79]. The study conducted in vitro cytotoxicity testing based on the hemolysis of type “AB” human blood cells exposed to various concentrations of plant extracts from O. vulgare and S. triloba. Results were compared to negative and total hemolysis controls. Low hemolytic activities (0.1% to 9.04%) were observed with plant extract concentrations up to 100 μg/mL, but higher activities were noted with Oregano E40 and E60 extracts as concentrations increased, reaching approximately 55% and 83%, respectively, at 1000 μg/mL. Clear distinctions were observed in the hemolytic activity of O. vulgare extracts, with ethanolic and enzymatic extracts exhibiting higher activity compared to the aqueous extract. On one end of the spectrum are the ethanolic and enzymatic extracts, while on the other end lies the aqueous extract, which demonstrates minimal hemolytic activity even at a concentration of 1000 µg/mL, showing only 10% activity. Across all examined extracts, hemolytic activity remains below 10% up to a concentration of 100 µg/mL, with specific values noted as 5.4%, 6.04%, 7.03%, and 9.04% for the aqueous, E40, E60, and ENZ extracts, respectively. Researchers demonstrated minimal hemolytic activity, remaining below 30% at a concentration of 100 mg/mL, suggesting potential safety when testing the water-ethanol solvent mixture extract of Origanum grosii (O. grosii).
In our current investigation, S. triloba extracts displayed minimal hemolytic activity across all tested concentrations. These findings indicate that S. triloba extracts do not induce hemolysis of red blood cells, suggesting their non-toxic properties. Consistent with our results, Saleh et al. reported that the acetone extract of S. triloba exhibited minimal hemolytic effects, with values of 1.282% and 3.157% observed for concentrations of 54.51 µg/mL and 190 µg/mL, respectively, which closely aligns with our findings [80]. However, caution should be exercised when consuming potentially toxic substances such as plants to safeguard health. Additionally, based on the blood group system, it can be indirectly hypothesized that observed hemolysis in blood type “AB” may be due to the presence of antigen A or B on the cell membrane [50,81].
4.5. General Aspects
The oral cavity should be approached as a highly intricate multi-ecosystem hosting diverse microbial communities. Its distinctive anatomical features and connection to the external environment, primarily through dietary intake, render it a specialized ecological niche for the "oral cavity-dependent" microbiome [82]. Both internal and external factors influence the oral microbiota, maintaining a dynamic equilibrium. Disruption in this balance can alter the microbial composition, leading to oral and potentially systemic diseases. Amidst the current antimicrobial resistance crisis, there's a resurgence of interest in natural plant-derived products, which serve as abundant sources of bioactive substances primarily with antimicrobial properties. However, when these compounds also demonstrate additional biological actions such as antioxidant and anti-inflammatory effects, they epitomize a holistic and comprehensive approach to addressing various diseases. The current study attempted to investigate different types of extracts of two plants (wild ecotypes), O. vulgare and S. triloba, against a diverse range of isolates found in the oral cavity. These isolates include F. nucleatum, S. salivarius, S. mutans, P. gingivalis, P. micra, and P. intermedia. Additionally, the study examined isolates, such as multi-antibiotic-resistant S. aureus, which may enter the oral cavity through food sources. The significance of the mentioned microorganisms in both oral and systemic health cannot be overstated due to their distinct characteristics. F. nucleatum, regarded as a pivotal "bridge" organism, possesses the ability to adhere to various cell types, acting as a conduit for early colonizers such as Streptococcus [83,84]. While S. mutans does not act in isolation, it plays a primary role in dental caries development by forming biofilm-based dental plaque on tooth surfaces. This bacterium also produces multiple adhesive proteins that promote plaque formation and subsequent caries. Moreover, S. mutans collaborates with other acidogenic and aciduric species, thriving in the acidic, EPS-rich environments it helps establish, thereby exacerbating dental decay [85,86]. Furthermore, inflammation's role in dysbiosis is underscored by periodontitis-associated bacteria like P. gingivalis, which can modulate the host immune response [87]. Lastly, emerging research indicates an association between P. micra [88] and immune responses in colorectal cancer (CRC), while the influence of F. nucleatum and P. gingivalis in the carcinogenic process is considered undeniable [89,91]. Therefore, it's crucial to prioritize understanding and uncovering substances and mechanisms that operate within the triad of antioxidant, antibacterial, and anti-inflammatory actions, rather than solely concentrating on one pathway.
Food pathogens such as S. aureus, are an endless threat to public health, causing significant numbers of illness and deaths every year worldwide [91]. Food safety is thus an important issue for the food industry which is constantly seeking for methods to secure its products from hazardous microorganisms. Many of the various chemical substances that have been used as food preservatives are considered as detrimental to human health and include substances such as sodium nitrite [92]. Consumers are very reluctant to accept synthetic food preservatives while they are much more tolerant to natural substances [93,94]. Phytochemicals could be a viable alternative to this problem [95,96] and could be incorporated in the food matrix, added in the packaging material, or coat the food [97,98]. All historical civilizations have used phytochemicals for medical, cosmetic, and culinary purposes and this ethnopharmacological data has passed to us as a robust corpus of empirical knowledge to exploit [99]. Of course, phytochemicals should pass the safety tests as every other compound intended for the food industry.
In the backdrop of the preceding discussion, it's worth highlighting that research has uncovered the abundance of specific active ingredients in Oregano and Salvia plants, attributed to their anti-inflammatory, antioxidant, and antibacterial properties, whether they act individually or synergistically. These include tannins, flavonoids, triterpenes, and phenols such as rosmarinic acid, linalool, thymol, carvacrol, caffeic acid, carnosic acid, carnosol, salvianolic acid B, and scutellarin [49,100]. The analyzed samples revealed abundant bioactive compounds, such as phenolics and flavonoids, contributing to a robust antioxidant effect. Furthermore, they exhibited notable activity against biofilm formation and demonstrated significant antibacterial efficacy, frequently outperforming conventional antibiotics.
While the study provides insights into phytochemical composition and bioactivities, limitations include its focus on only two plant species, methodological constraints, and a lack of mechanistic insights. Future research should address these limitations through standardized methodologies and in-depth mechanistic studies using in vivo models or clinical trials. Despite limitations, the study highlights the antioxidant, antibacterial, and anti-biofilm properties of the extracts, suggesting their potential in oral care. However, sustainability concerns regarding habitat destruction and biodiversity loss due to extraction processes warrant further exploration of eco-friendly alternatives, including synthetic or bioengineered compounds [101,102].
5. Conclusions
1. The study reports on the multifactorial dependence of phytochemical isolation on factors such as extraction methods, analytical techniques, and plant species' unique characteristics. Despite variations in extraction methodologies, all types of extracts demonstrated significant antioxidant capacity, highlighting the potential health benefits of these plants.
2. Additionally, the extracts exhibited promising antibacterial activity against a diverse range of oral pathogens, including antibiotic-resistant strains, and showed efficacy in inhibiting oral biofilm formation. While the study has limitations, such as its focus on only two plant species and methodological constraints, it provides a foundation for future research to explore the mechanisms underlying the observed bioactivities and validate the efficacy and safety of plant-derived treatments through in vivo models or clinical trials.
3. The extracts from both plants showed an exceptional antibacterial action against the two S. aureus strains which originated from poultry meat and raw milk. This finding is essential for the future development of food preservatives based on these extracts.
Overall, the findings suggest that O. vulgare and S. triloba extracts contain bioactive compounds with potential therapeutic value, warranting further investigation in the field of natural product pharmacology and therapeutics.
Author Contributions
Conceptualization, M.A. and C.V; methodology, C.V., A.T., C.E., R.G ; software, A.A., A.D., N.V., K.Z. and G.R.; validation, C.E., K.Z., N.V., G.R. and A.A.; formal analysis, A.A., A.D., C.E., K.Z., G.R.; investigation, C.E., N.V.,A.D., G.R., K.Z., A.T., C.V.; resources, G.R., N.V., K.Z; data curation, A.A., G.R.; writing—original draft preparation, G.R., M.A., A.A., A.T., C.V. T.V; writing—review and editing, M.A., A.T., C.V.; visualization, C.V. and M.A.; supervision, M.A, C.V.; project administration A.T., C.V., T.V., and M.A.; funding acquisition, T.V. and C.V. All authors have read and agreed to the published version of the manuscript.
Funding
“This research received no external funding”
Conflicts of Interest
“The authors declare no conflicts of interest.”
References
- Antoniadou M, Rozos G, Vaiou N, Zaralis K, Ersanli C, Alexopoulos A, Tzora A, Varzakas T, Voidarou CC. The In Vitro Assessment of Antibacterial and Antioxidant Efficacy in Rosa damascena and Hypericum perforatum Extracts against Pathogenic Strains in the Interplay of Dental Caries, Oral Health, and Food Microbiota. Microorganisms. 2023 Dec 28;12(1):60. [CrossRef]
- Rezabakhsh A, Mahmoodpoor A, Soleimanpour H. Historical perspective of aspirin: A journey from discovery to clinical practice Ancient and modern history. J Cardiovasc Thorac Res. 2021;13(2):179-180. Epub 2021 Apr 17. [CrossRef]
- Elshafie HS, Camele I, Mohamed AA. A Comprehensive Review on the Biological, Agricultural and Pharmaceutical Properties of Secondary Metabolites Based-Plant Origin. Int J Mol Sci. 2023 Feb 7;24(4):3266. [CrossRef]
- Erb M, Kliebenstein DJ. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020 Sep;184(1):39-52. Epub 2020 Jul 7. [CrossRef]
- Buchanan BB, Gruissem W, Jones RL, 2000, Biochemistry and Molecular Biology of Plants. Rockville, MD: American Society of Plant Phys iologists. Bob B. Buchanan (Editor), Wilhelm Gruissem (Editor), Russell L. Jones (Editor).
- Lambers, Hans & Chapin III, F Stuart & Pons, Thijs. Ecological Biochemistry: Allelopathy and Defense Against Herbivores. 2008. [CrossRef]
- Alibi S, Crespo D, Navas J. Plant-Derivatives Small Molecules with Antibacterial Activity. Antibiotics (Basel). 2021 Feb 25;10(3):231. [CrossRef]
- Bajguz, A.; Piotrowska-Niczyporuk, A. Biosynthetic Pathways of Hormones in Plants. Metabolites 2023, 13, 884. [CrossRef]
- Soltani S, Shakeri A, Iranshahi M, Boozari M. A Review of the Phytochemistry and Antimicrobial Properties of Origanum vulgare L. and Subspecies. Iran J Pharm Res. 2021 Spring;20(2):268-285. [CrossRef]
- Liang J, Luo Y, Zhao H. Synthetic biology: putting synthesis into biology. Wiley Interdiscip Rev Syst Biol Med. 2011 Jan-Feb;3(1):7-20. [CrossRef]
- Caselli E, Fabbri C, D'Accolti M, Soffritti I, Bassi C, Mazzacane S, Franchi M. Defining the oral microbiome by whole-genome sequencing and resistome analysis: the complexity of the healthy picture. BMC Microbiol. 2020 May 18;20(1):120. [CrossRef]
- Pisano, M.; Giordano, F.; Sangiovanni, G.; Capuano, N.; Acerra, A.; D’Ambrosio, F. The Interaction between the Oral Microbiome and Systemic Diseases: A Narrative Review. Microbiol. Res. 2023, 14, 1862-1878. [CrossRef]
- Lamont RJ, Koo H, Hajishengallis G. The oral microbiota: dynamic communities and host interactions. Nat Rev Microbiol. 2018 16(12), 745-759. [CrossRef]
- Alibi, S.; Crespo, D.; Navas, J. Plant-Derivatives Small Molecules with Antibacterial Activity. Antibiotics 2021, 10, 231. [CrossRef]
- Giordano-Kelhoffer, B.; Lorca, C.; March Llanes, J.; Rábano, A.; del Ser, T.; Serra, A.; Gallart-Palau, X. Oral Microbiota, Its Equilibrium and Implications in the Pathophysiology of Human Diseases: A Systematic Review. Biomedicines 2022, 10, 1803. [CrossRef]
- Josh L Espinoza, Manolito Torralba, Pamela Leong, Richard Saffery, Michelle Bockmann, Claire Kuelbs, Suren Singh, Toby Hughes, Jeffrey M Craig, Karen E Nelson, Chris L Dupont, Differential network analysis of oral microbiome metatranscriptomes identifies community scale metabolic restructuring in dental caries, PNAS Nexus, Volume 1, Issue 5, November 2022, pgac239. [CrossRef]
- Varzakas, T.; Antoniadou, M. A Holistic Approach for Ethics and Sustainability in the Food Chain: The Gateway to Oral and Systemic Health. Foods 2024, 13, 1224. [CrossRef]
- Cugini C, Ramasubbu N, Tsiagbe VK, Fine DH. Dysbiosis From a Microbial and Host Perspective Relative to Oral Health and Disease. Front Microbiol. 2021 Mar 5;12:617485. [CrossRef]
- Kennedy DO, Wightman EL. Herbal extracts and phytochemicals: plant secondary metabolites and the enhancement of human brain function. Adv Nutr. 2011 Jan;2(1):32-50. [CrossRef]
- Chaachouay, N.; Zidane, L. Plant-Derived Natural Products: A Source for Drug Discovery and Development. Drugs Drug Candidates 2024, 3, 184-207. [CrossRef]
- Idir F, Van Ginneken S, Coppola GA, Grenier D, Steenackers HP, Bendali F. Origanum vulgare ethanolic extracts as a promising source of compounds with antimicrobial, anti-biofilm, and anti-virulence activity against dental plaque bacteria. Front Microbiol. 2022 Nov 2;13:999839. [CrossRef]
- Salam MA, Al-Amin MY, Salam MT, Pawar JS, Akhter N, Rabaan AA, Alqumber MAA. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare (Basel). 2023 Jul 5;11(13):1946. [CrossRef]
- Guefack MF, Ngangoue MO, Mbaveng AT, Nayim P, Kuete JRN, Ngaffo CMN, Chi GF, Ngameni B, Ngadjui BT, Kuete V. Antibacterial and antibiotic-potentiation activity of the constituents from aerial part of Donella welwitshii (Sapotaceae) against multidrug resistant phenotypes. BMC Complement Med Ther. 2022 Jul 20;22(1):194. [CrossRef]
- Fair, Richard & Tor, Yitzhak. (2014). Antibiotics and Bacterial Resistance in the 21st Century. Perspectives in medicinal chemistry. 6. 25-64. 10.4137/PMC.S14459.
- Aljeldah, M.M. Antimicrobial Resistance and Its Spread Is a Global Threat. Antibiotics 2022, 11, 1082. [CrossRef]
- Antimicrobial Resistance Collaborators. The burden of bacterial antimicrobial resistance in the WHO African region in 2019: a cross-country systematic analysis. Lancet Glob Health. 2024 Feb;12(2):e201-e216. [CrossRef]
- Luepke KH, Suda KJ, Boucher H, Russo RL, Bonney MW, Hunt TD, Mohr JF 3rd. Past, Present, and Future of Antibacterial Economics: Increasing Bacterial Resistance, Limited Antibiotic Pipeline, and Societal Implications. Pharmacotherapy. 2017 Jan;37(1):71-84. [CrossRef]
- Nasim N, Sandeep IS, Mohanty S. Plant-derived natural products for drug discovery: current approaches and prospects. Nucleus (Calcutta). 2022;65(3):399-411. Epub 2022 Oct 18. [CrossRef]
- Jadimurthy, R.; Jagadish, S.; Nayak, S.C.; Kumar, S.; Mohan, C.D.; Rangappa, K.S. Phytochemicals as Invaluable Sources of Potent Antimicrobial Agents to Combat Antibiotic Resistance. Life 2023, 13, 948. [CrossRef]
- Zhang QW, Lin LG, Ye WC. Techniques for extraction and isolation of natural products: a comprehensive review. Chin Med. 2018 Apr 17;13:20. [CrossRef]
- Cendrowski, A., Kraśniewska, K., Przybył, J.L., Zielińska, A., Kalisz, S. Antibacterial and Antioxidant Activity of Extracts from Rose Fruits (Rosa rugosa). Molecules 2020, 25, 1365. [CrossRef]
- Tiranakwit T, Puangpun W, Tamprasit K, Wichai N, Siriamornpun S, Srisongkram T, Weerapreeyakul N. Phytochemical Screening on Phenolic, Flavonoid Contents, and Antioxidant Activities of Six Indigenous Plants Used in Traditional Thai Medicine. Int J Mol Sci. 2023 Aug 30;24(17):13425. [CrossRef]
- Belew AA, W/Hana GG, Meshesha DS, Akele ML. Evaluation of Total Phenolic, Total Flavonoid Content And Antioxidant Activity of Rhus Vulgaris. Research Square; 2021. PPR:PPR437116. [CrossRef]
- Mabotja MB, Venter SL, Du Plooy CP, Kudanga T, Amoo SO. Phytochemical Content, Antioxidant, Alpha-Glucosidase Inhibitory and Antibacterial Activities of Spineless Cactus Pear Cultivars. Plants (Basel). 2021 Jun 28;10(7):1312. [CrossRef]
- Saeed N, Khan MR, Shabbir M. Antioxidant activity, total phenolic and total flavonoid contents of whole plant extracts Torilis leptophylla L. BMC Complement Altern Med. 2012, 16, 12:221. [CrossRef]
- Al-Nawas, B.; Karbach, J. S3 Leitlinie: Odontogene Infektionen. Online verfügbar unter.2017. Available Online at: https://wwwawmforg/uploads/tx_szleitlinien/007-006l_S3_Odontogene_Infektionen_2017-12pdf.
- Meinen, A, Reuss A, Willrich N.; Feig M, Noll I, Eckmanns T.; Al-Nawas B.; Markwart R. Antimicrobial Resistance and the Spectrum of Pathogens in Dental and Oral-Maxillofacial Infections in Hospitals and Dental Practices in Germany. Front Microbiol. 2021, 2, 12:676108. [CrossRef]
- Bhat, KG, Ingalagi P, Patil S, Patil S, Pattar G. Antimicrobial susceptibility pattern of oral gram negative anaerobes from Indian subjects. Anaerobe. 2021, 70, 102367. [CrossRef]
- LSI. Performance Standards for Antimicrobial Disk Susceptibility Tests: Approved Standard—CLSI Document M02-A11; Clinical & Laboratory Standards Institute: Wayne, PA, USA, 2015.
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: Approved Standard—Eighth Edition—CLSI Document M07-A7; Clinical & Laboratory Standards Institute: Wayne, PA, USA, 2006.
- Clinical and Laboratory Standards Institute (CLSI) Performance Standards for Antimicrobial Susceptibility Testing. 28th ed. Clinical and Laboratory Standards Institute; Wayne, PA, USA: 2018. CLSI Supplement M100.
- The European Committee on Antimicrobial Susceptibility Testing Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth microdilution. EUCAST Discussion Document E. Def 2003, 5.1. Clin. Microbiol. Infect. 2003, 9, 1–7. [CrossRef]
- Kowalska-Krochmal B, Dudek-Wicher R. The Minimum Inhibitory Concentration of Antibiotics: Methods, Interpretation, Clinical Relevance. Pathogens. 2021 Feb 4;10(2):165. [CrossRef]
- Veloso DJ, Abrão F, Martins CHG, Bronzato JD, Gomes BPFA, Higino JS, Sampaio FC. Potential antibacterial and anti-halitosis activity of medicinal plants against oral bacteria. Arch Oral Biol. 2020 Feb;110:104585. [CrossRef]
- Clinical Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial a Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard, 8th ed.; CLSI: Wayne, PA, USA, 2009; CLSI document M0-A8.
- Bagheri, R.; Bohlouli, S., Maleki Dizaj, S., Shahi, S., Memar, M.Y.; Salatin, S. The Antimicrobial and Anti-Biofilm Effects of Hypericum perforatum Oil on Common Pathogens of Periodontitis: An In Vitro Study. Clinics and Practice. 2022, 12, 1009–1019. [CrossRef]
- Ersanli, C.; Tzora, A.; Skoufos, I.; Fotou, K.; Maloupa, E.; Grigoriadou, K.; Voidarou, C.C.; Zeugolis, D.I. The Assessment of Antimicrobial and Anti-Biofilm Activity of Essential Oils against Staphylococcus aureus Strains. Antibiotics 2023, 12, 384. [CrossRef]
- Samreen, Qais FA, Ahmad I. Anti-quorum sensing and biofilm inhibitory effect of some medicinal plants against gram-negative bacterial pathogens: in vitro and in silico investigations. Heliyon. 2022 Oct 17;8(10):e11113. [CrossRef]
- Adeyemo RO, Famuyide IM, Dzoyem JP, Lyndy Joy M. Anti-Biofilm, Antibacterial, and Anti-Quorum Sensing Activities of Selected South African Plants Traditionally Used to Treat Diarrhoea. Evid Based Complement Alternat Med. 2022 Sep 28;2022:1307801. [CrossRef]
- Souza-Melo WO, Figueiredo-Júnior EC, Freire JCP, Costa BP, Lira AB, Freires IA, Cavalcanti YW, Lopes WS, Tavares JF, Pessôa HLF, Pereira JV. Phytochemistry, antifungal and antioxidant activity, and cytotoxicity of byrsonima gardneriana (A. Juss) extract. Arch Oral Biol. 2021 Mar;123:104994. [CrossRef]
- Sæbø, I.P.; Bjørås, M.; Franzyk, H.; Helgesen, E.; Booth, J.A. Optimization of the Hemolysis Assay for the Assessment of Cytotoxicity. Int. J. Mol. Sci. 2023, 24, 2914. [CrossRef]
- Jiang Z, Kempinski C, Chappell J. Extraction and Analysis of Terpenes/Terpenoids. Curr Protoc Plant Biol. 2016;1:345-358. Epub 2016 Jun 10. [CrossRef]
- Knez Hrnčič, M.; Cör, D.; Simonovska, J.; Knez, Ž.; Kavrakovski, Z.; Rafajlovska, V. Extraction Techniques and Analytical Methods for Characterization of Active Compounds in Origanum Species. Molecules 2020, 25, 4735. [CrossRef]
- Skoula M and Harborne JB. The taxonomy and chemistry of Origanum. Kintzios S., medicinal and aromatic plants–industrial profiles–oregan, The genera Origanum and Lipia, Taylor & Francis: London. (2002) Aug 29: 67-108.
- Lukas B, Schmiderer C and Novak J. Phytochemical diversity of Origanum vulgare L. subsp. vulgare (Lamiaceae) from Austria. Biochem. Syst. Ecol. 2013, 50: 106-13.
- Khan M, Khan ST, Khan NA, Mahmood A, AlKedhairy AA and Alkhathlan HZ. The composition of the essential oil and aqueous distillate of Origanum vulgare L. growing in Saudi Arabia and evaluation of their antibacterial activity. Arab. J. Chem. 2018, 11: 1189-200.
- Lukas B, Schmiderer C and Novak J. Essential oil diversity of European Origanum vulgare L.(Lamiaceae). Phytochemistry 2015, 119, 32-40.
- Timothy, Chris & Priya, V. Gayathri R, Gayathri R. Phytochemical analysis and total phenolic content of Origanum vulgare (oregano). Drug Invention Today. 2018, 10. 1903-1905).
- Mróz, M., Kusznierewicz, B. Phytochemical screening and biological evaluation of Greek sage (Salvia fruticosa Mill.) extracts. Sci Rep 2023, 13, 22309. [CrossRef]
- Al-Kalaldeh, J. Z., Abu-Dahab, R. & Aff, F. U. Volatile oil composition and antiproliferative activity of Laurus nobilis, Origanum syriacum, Origanum vulgare, and Salvia triloba against human breast adenocarcinoma cells. Nutr. Res. 2010, 30(4), 271–278. [CrossRef]
- Torun, M., Dincer, C., Topuz, A., Sahin-Nadeem, H. Ozdemir, F. Aqueous extraction kinetics of soluble solids, phenolics and favonoids from sage (Salvia fruticosa Miller) leaves. J. Food Sci. Technol. 2015, 52(5), 2797–2805. [CrossRef]
- Bower, A.M.; Real Hernandez, L.M.; Berhow, M.A.; de Mejia, E.G. Bioactive Compounds from Culinary Herbs Inhibit a Molecular Target for Type 2 Diabetes Management, Dipeptidyl Peptidase IV. J. Agric. Food Chem. 2014, 62, 6147–6158.
- Jafari Khorsand, G., Morshedloo, M.R., Mumivand, H. et al. Natural diversity in phenolic components and antioxidant properties of oregano (Origanum vulgare L.) accessions, grown under the same conditions. Sci Rep 12, 5813 (2022). [CrossRef]
- Yan, Feng & Azizi, Ali & Janke, Susanne & Schwarz, Madeleine & Zeller, Stefanie & Honermeier, Bernd. (2016). Antioxidant capacity variation in the oregano (Origanum vulgare L.) collection of the German National Genebank. Industrial Crops and Products. 92. 19–25. [CrossRef]
- Özüpek, Burçin; Pekacar, Sultan; Orhan, Didem Deliorman. Evaluation of Phytochemical Contents and Biological Activities of Salvia officinalis and Salvia triloba Grown with Organic Farming FABAD Journal of Pharmaceutical Sciences; Ankara 2023, 48, 1, 125-138. [CrossRef]
- Lantzouraki DZ, Tsiaka T, Soteriou N, Asimomiti G, Spanidi E, Natskoulis P, Gardikis K, Sinanoglou VJ, Zoumpoulakis P. Antioxidant Profiles of Vitis vinifera L. and Salvia triloba L. Leaves Using High-Energy Extraction Methodologies. J AOAC Int. 2020 Apr 1;103(2):413-421. [CrossRef]
- Zhang W, Chen H, Wang Z, Lan G, Zhang L. Comparative studies on antioxidant activities of extracts and fractions from the leaves and stem of Epimedium koreanum Nakai. J Food Sci Technol. 2013 Dec;50(6):1122-9. [CrossRef]
- Indumathy, R. & Aruna, A. Free radical scavenging activities, total phenolic and flavonoid content of Lepidium sativum (Linn.). International Journal of Pharmacy and Pharmaceutical Sciences. 2013, 5. 634-637.
- Kakouri, E., Trigas, P., Daferera, D., Skotti, E., Tarantilis, P.A., Kanakis, C. Chemical Characterization and Antioxidant Activity of Nine Hypericum Species from Greece. Antioxidants 2023, 12, 899. [CrossRef]
- Jan S, Rashid M, Abd Allah EF, Ahmad P. Biological Efficacy of Essential Oils and Plant Extracts of Cultivated and Wild E.cotypes of Origanum vulgare L. Biomed Res Int. 2020, 6, 2020:8751718. [CrossRef]
- Parra, C.; Muñoz, P.; Bustos, L.; Parra, F.; Simirgiotis, M.J.; Escobar, H. UHPLC-DAD Characterization of Origanum vulgare L. from Atacama Desert Andean Region and Antioxidant, Antibacterial and Enzyme Inhibition Activities. Molecules 2021, 26, 2100. [CrossRef]
- Gülçin, Ilhami. (2015). Fe3+–Fe2+ Transformation Method: An Important Antioxidant Assay. Methods in molecular biology (Clifton, N.J.). 1208. 233-46. [CrossRef]
- Hickl J, Argyropoulou A, Sakavitsi ME, Halabalaki M, Al-Ahmad A, Hellwig E. Mediterranean herb extracts inhibit microbial growth of representative oral microorganisms and biofilm formation of Streptococcus mutans. PLoS ONE 2018, 13(12): e0207574. [CrossRef]
- Popa M, Măruțescu L, Oprea E, Bleotu C, Kamerzan C, Chifiriuc MC, Grădișteanu Pircalabioru G. In Vitro Evaluation of the Antimicrobial and Immunomodulatory Activity of Culinary Herb Essential Oils as Potential Perioceutics. Antibiotics (Basel). 2020 Jul 21;9(7):428. [CrossRef]
- Delamare, Ana Paula & Moschen-Pistorello, Ivete & Artico, Liane & Atti-Serafini, Luciana & Echeverrigaray, Sergio. (2007). Antibacterial activity of the essential oils of Salvia officinalis L. and Salvia triloba L. cultivated in South Brazil. Food Chemistry. 100. 603-608. [CrossRef]
- Eltawaty, Salwa & Busba, Abdelnaser & Omare, Mohamed-Elfatih & Almagboul, Asha & Ahmed, Sanaa Osman Yagoub & Elramli, Asma & Elnour, Asim & Dafalla, Mohammed. (2021). ANTIMICROBIAL ACTIVITY OF LIBYAN SALVIA FRUTICOSA MIL AND MULTI- DRUG RESISTANT BACTERIA. 8. 416-423.
- de Oliveira JR, Vilela PGDF, Almeida RBA, de Oliveira FE, Carvalho CAT, Camargo SEA, Jorge AOC, de Oliveira LD. Antimicrobial activity of noncytotoxic concentrations of Salvia officinalis extract against bacterial and fungal species from the oral cavity. Gen Dent. 2019, 67(1):22-26.
- Amin, Ketan & Dannenfelser, Rose. (2006). In vitro hemolysis: Guidance for the pharmaceutical scientist. Journal of pharmaceutical sciences. 95. 1173-6. [CrossRef]
- Brewer, Susan. (2011). Natural Antioxidants: Sources, Compounds, Mechanisms of Action, and Potential Applications. Comprehensive Reviews in Food Science and Food Safety. 10. 221 - 247. [CrossRef]
- Saleh, N.A.M.; El-bary, R.B.E.-d.A.; Mpingirika, E.Z.; Essa, H.L.; El-Sayed, M.M.H.; Sherbetjian, M.S.; Elfandi, H.F.; Wahed, M.A.A.; Arafeh, R.; Amleh, A. Evaluating the Potential Anticancer Properties of Salvia triloba in Human-Osteosarcoma U2OS Cell Line and Ovarian Adenocarcinoma SKOV3 Cell Line. Appl. Sci. 2022, 12, 11545. [CrossRef]
- Daniels, G. The molecular genetics of blood group polymorphism. Transplant Immunology, 2005, 14(3-4 SPEC. ISS), 143–153. [CrossRef]
- Li X, Liu Y, Yang X, Li C, Song Z. The Oral Microbiota: Community Composition, Influencing Factors, Pathogenesis, and Interventions. Front. Microbiol. 2022, 13:895537. [CrossRef]
- Silbergleit M, Vasquez AA, Miller CJ, Sun J, Kato I. Oral and intestinal bacterial exotoxins: Potential linked to carcinogenesis. Prog Mol Biol Transl Sci. 2020, 171:131-193. [CrossRef]
- Sun CH, Li BB, Wang B, Zhao J, Zhang XY, Li TT, Li WB, Tang D, Qiu MJ, Wang XC, Zhu CM, Qian ZR. The role of Fusobacterium nucleatum in colorectal cancer: from carcinogenesis to clinical management. Chronic Dis Transl Med. 2019, 1;5(3):178-187. [CrossRef]
- Lemos JA, Palmer SR, Zeng L, Wen ZT, Kajfasz JK, Freires IA, Abranches J, Brady LJ. The Biology of Streptococcus mutans. Microbiol Spectr. 2019 Jan;7(1):10.1128/microbiolspec.GPP3-0051-2018. [CrossRef]
- Matsumoto-Nakano M. Role of Streptococcus mutans surface proteins for biofilm formation. Jpn Dent Sci Rev. 2018 Feb;54(1):22-29. [CrossRef]
- Lamont RJ, Koo H, Hajishengallis G. The oral microbiota: dynamic communities and host interactions. Nat Rev Microbiol. 2018 16(12), 745-759. [CrossRef]
- Löwenmark T, Li X, Löfgren-Burström A, Zingmark C, Ling A, Kellgren TG, Larsson P, Ljuslinder I, Wai SN, Edin S, Palmqvist R. Parvimonas micra is associated with tumour immune profiles in molecular subtypes of colorectal cancer. Cancer Immunol Immunother. 2022 Oct;71(10):2565-2575. [CrossRef]
- Pignatelli P, Nuccio F, Piattelli A, Curia MC. The Role of Fusobacterium nucleatum in Oral and Colorectal Carcinogenesis. Microorganisms. 2023 Sep 20;11(9):2358. [CrossRef]
- Kerdreux M, Edin S, Löwenmark T, Bronnec V, Löfgren-Burström A, Zingmark C, Ljuslinder I, Palmqvist R, Ling A. Porphyromonas gingivalis in Colorectal Cancer and its Association to Patient Prognosis. J Cancer. 2023 May 21;14(9):1479-1485. [CrossRef]
- Aladhadh, M. A Review of Modern Methods for the Detection of Foodborne Pathogens. Microorganisms 2023, 11, 1111. [CrossRef]
- Shakil MH, Trisha AT, Rahman M, Talukdar S, Kobun R, Huda N, Zzaman W. Nitrites in Cured Meats, Health Risk Issues, Alternatives to Nitrites: A Review. Foods. 2022 Oct 25;11(21):3355. [CrossRef]
- Redondo-Blanco S, Fernández J, López-Ibáñez S, Miguélez EM, Villar CJ, Lombó F. Plant Phytochemicals in Food Preservation: Antifungal Bioactivity: A Review. J Food Prot. 2020 Jan;83(1):163-171. [CrossRef]
- Dominguez LJ, Veronese N, Vernuccio L, Catanese G, Inzerillo F, Salemi G, Barbagallo M. Nutrition, Physical Activity, and Other Lifestyle Factors in the Prevention of Cognitive Decline and Dementia. Nutrients. 2021 Nov 15;13(11):4080. [CrossRef]
- Wong, Young & Chow, Yit-Lai. (2023). Exploring the potential of spice-derived phytochemicals as alternative antimicrobial agents. eFood. 5. [CrossRef]
- Teshome, S., Jaleta,A., Jemal, M., Abamecha, A., Gershe, S. Phytochemical screening and evaluation of antimicrobial activity of Pterolobium stellatum root extract. Pharm Pharmacol Int J. 2023;11(4):151‒156. https://medcraveonline.com/PPIJ/PPIJ-11-00413.pdf.
- Siddiqui SA, Khan S, Mehdizadeh M, Bahmid NA, Adli DN, Walker TR, Perestrelo R, Câmara JS. Phytochemicals and bioactive constituents in food packaging - A systematic review. Heliyon. 2023 Oct 24;9(11):e21196. [CrossRef]
- Bouarab Chibane, L., Degraeve, P., Ferhout, H., Bouajila, J. and Oulahal, N. (2019), Plant antimicrobial polyphenols as potential natural food preservatives. J. Sci. Food Agric., 99: 1457-1474. [CrossRef]
- Thakur, Monika & Singh, Karuna & Khedkar, Renu. (2020). Phytochemicals. [CrossRef]
- Lombrea A, Antal D, Ardelean F, Avram S, Pavel IZ, Vlaia L, Mut AM, Diaconeasa Z, Dehelean CA, Soica C, Danciu C. A Recent Insight Regarding the Phytochemistry and Bioactivity of Origanum vulgare L. Essential Oil. Int J Mol Sci. 2020 Dec 17;21(24):9653. [CrossRef]
- Prado L, Zalba M, Amato D. Sustainability in Oral Health: The Role of Natural Products. Molecules. 2020, 25(10):2267. [CrossRef]
- Souza LGA, Lopes LGC, Hill MP, Harries P, Eggersdorfer M, Marušić K. Sustainable Sourcing of Plant-Based Ingredients for Oral Health Care. Nutrients. 2021;13(3):1002. [CrossRef]
Figure 1.
Total phenolics and flavonoid content of O. vulgare and S. triloba extracts. Letters above bars indicate similarities or statistically significant differences within total phenolics or flavonoid results (Kruskal-Wallis, p<0.05 with Tukey’s HSD).
Figure 1.
Total phenolics and flavonoid content of O. vulgare and S. triloba extracts. Letters above bars indicate similarities or statistically significant differences within total phenolics or flavonoid results (Kruskal-Wallis, p<0.05 with Tukey’s HSD).
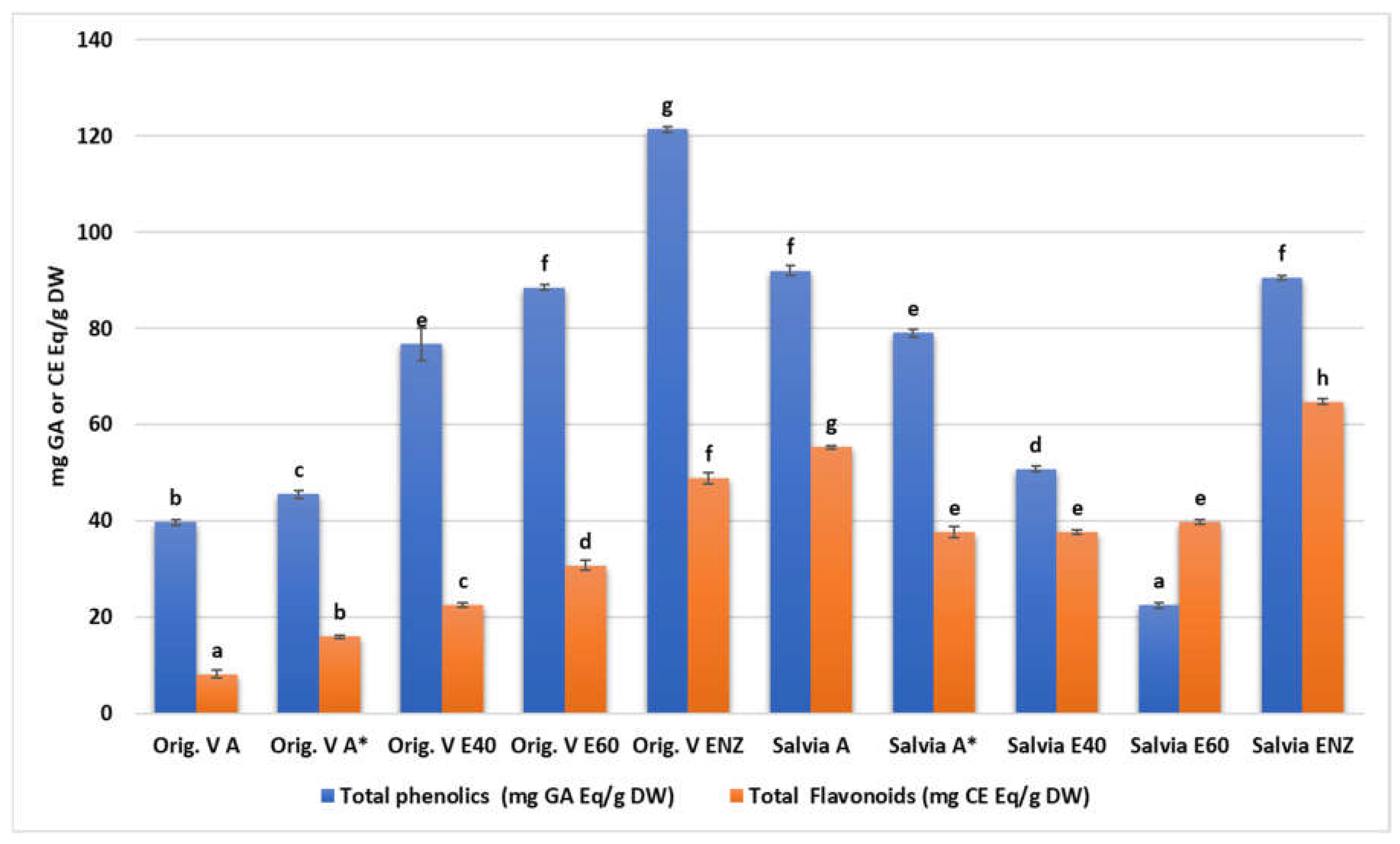
Figure 2.
Antimicrobial activity (agar diffusion in mm) of O. vulgare and S. triloba extracts (Aq, E40, E60, Enz) compared with common antibiotics. Values are means from the best responds (100% disk content) in each experiment.The antibacterial activity of the various plant extracts against oral pathogens as indicated by the minimum inhibitory concentration and minimum bactericidal concentration is presented in Table 8 for O. vulgare and in Table 9 for S. triloba. All extracts were effective against pathogens since MIC values ranged from 0.4 to 12.5 μg/mL and MBC values from 3.1 to 50 μg/mL. However, the oregano enzymatic and 60% ethanolic-aqueous extracts exhibited the lowest mean MIC values (0.39±0 to 3.12±0 μg/ml) against pathogens while the most effective were also with salvia extracts with mean values from 0.39±0 to 1.56±0 μg/mL. Most sensitive strains in oregano extracts were the MRSA isolated from poultry, P. gingivalis, F. nucleatum, P. intermedia and P. micra while in salvia extracts the MRSA (from raw poultry), S. mutans, S. salivarius, P. gingivalis, P. intermedia, P. micra and the reference strain S. aureus ATCC 12600 (Ref). As graphically depicted in Figure 3 most extracts presented a comparable or even stronger antimicrobial effectiveness when compared with the effectiveness of common antibiotics. .
Figure 2.
Antimicrobial activity (agar diffusion in mm) of O. vulgare and S. triloba extracts (Aq, E40, E60, Enz) compared with common antibiotics. Values are means from the best responds (100% disk content) in each experiment.The antibacterial activity of the various plant extracts against oral pathogens as indicated by the minimum inhibitory concentration and minimum bactericidal concentration is presented in Table 8 for O. vulgare and in Table 9 for S. triloba. All extracts were effective against pathogens since MIC values ranged from 0.4 to 12.5 μg/mL and MBC values from 3.1 to 50 μg/mL. However, the oregano enzymatic and 60% ethanolic-aqueous extracts exhibited the lowest mean MIC values (0.39±0 to 3.12±0 μg/ml) against pathogens while the most effective were also with salvia extracts with mean values from 0.39±0 to 1.56±0 μg/mL. Most sensitive strains in oregano extracts were the MRSA isolated from poultry, P. gingivalis, F. nucleatum, P. intermedia and P. micra while in salvia extracts the MRSA (from raw poultry), S. mutans, S. salivarius, P. gingivalis, P. intermedia, P. micra and the reference strain S. aureus ATCC 12600 (Ref). As graphically depicted in Figure 3 most extracts presented a comparable or even stronger antimicrobial effectiveness when compared with the effectiveness of common antibiotics. .
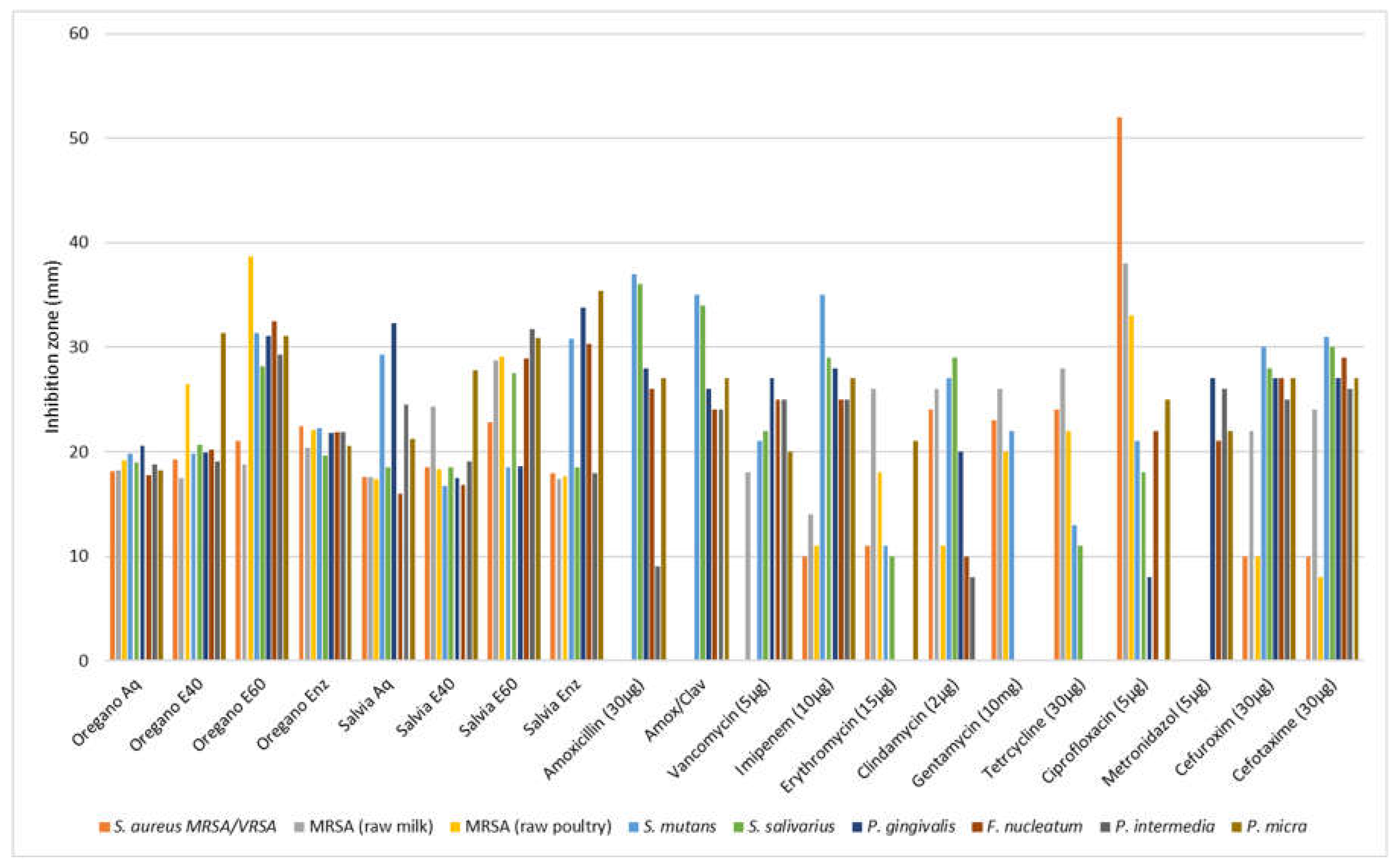
Figure 3.
Antimicrobial activity (mean MIC in μg/mL) of O. vulgare and S. triloba extracts (Aq, E40, E60, Enz) compared with common antibiotics. MIC values higher than 50 μg/mL are limited for best depiction to 50 μg/mLThe antibiofilm activity was assessed on a four-score scale, categorizing inhibition of biofilm formation as poor (0-50% inhibition), good (50-80% inhibition), very good (80-95% inhibition), and excellent (> 95% inhibition). Statistical analysis utilized medians or grouped medians instead of means. In Figure 4, the median scores of various herbal extracts and the scores of the 7 antibiotics are presented. Overall, antibiotics demonstrated broad and potent antibiofilm effectiveness against all pathogens tested, with vancomycin showing the highest efficacy. However, herbal extracts from both species, especially the enzymatic and 60% ethanol-aqueous extracts, exhibited significant antibiofilm properties comparable to those of conventional chemical drugs.
Figure 3.
Antimicrobial activity (mean MIC in μg/mL) of O. vulgare and S. triloba extracts (Aq, E40, E60, Enz) compared with common antibiotics. MIC values higher than 50 μg/mL are limited for best depiction to 50 μg/mLThe antibiofilm activity was assessed on a four-score scale, categorizing inhibition of biofilm formation as poor (0-50% inhibition), good (50-80% inhibition), very good (80-95% inhibition), and excellent (> 95% inhibition). Statistical analysis utilized medians or grouped medians instead of means. In Figure 4, the median scores of various herbal extracts and the scores of the 7 antibiotics are presented. Overall, antibiotics demonstrated broad and potent antibiofilm effectiveness against all pathogens tested, with vancomycin showing the highest efficacy. However, herbal extracts from both species, especially the enzymatic and 60% ethanol-aqueous extracts, exhibited significant antibiofilm properties comparable to those of conventional chemical drugs.
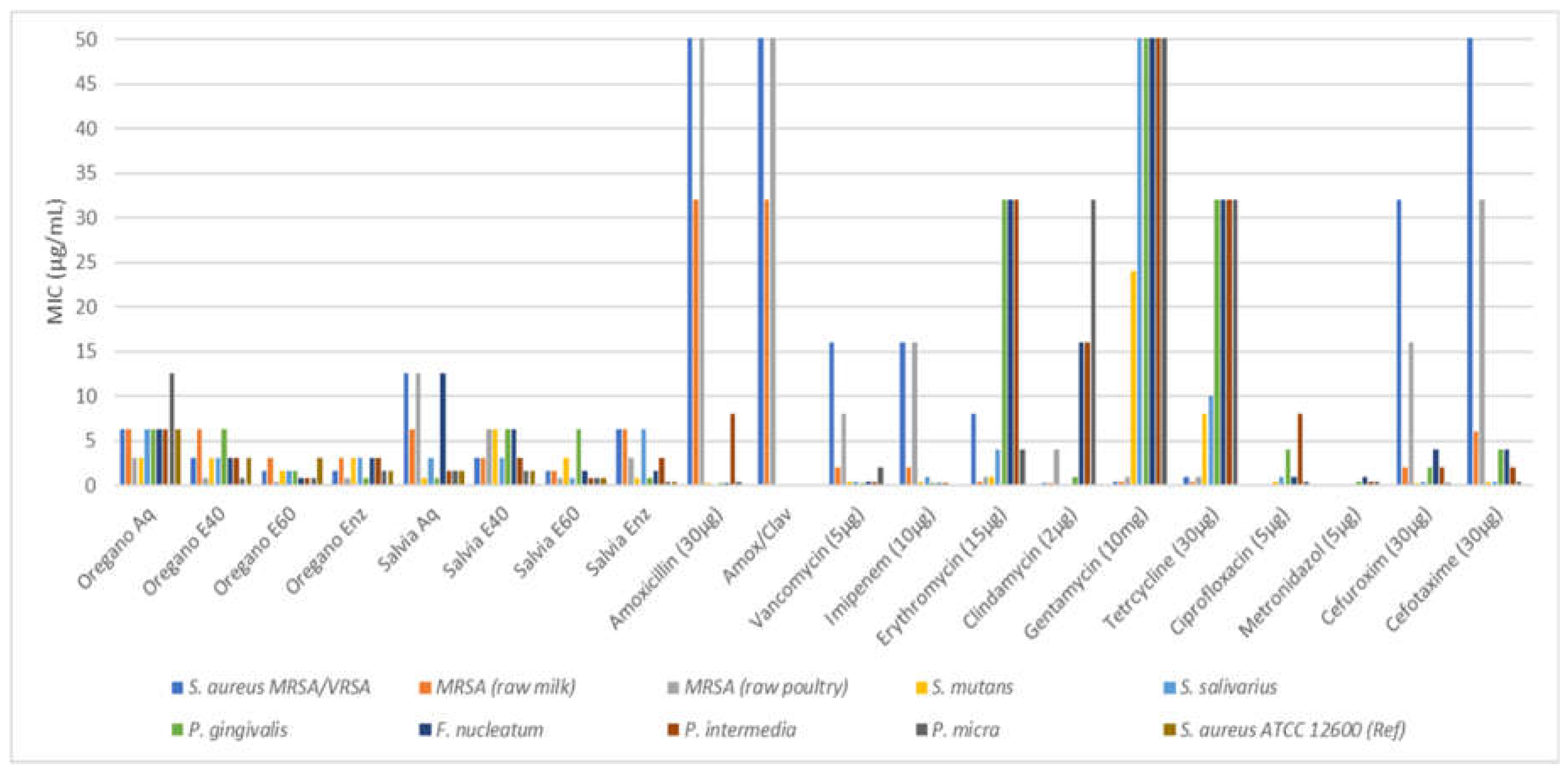
Figure 4.
Total median values (score 1: poor, 2: good, 3: very good, 4: excellent) of antibiofilm effectiveness of various types of extracts from the two herbs. Values close to 1 indicate poor biofilm inhibition (0-50%) while values close to 4 indicate excellent biofilm inhibition (>95%), Similar letters above bars indicate no significant difference in antibiofilm effectiveness between similar types of extracts among the various herbal species and antibiotics (Kruskal-Wallis, p<0.05).
Figure 4.
Total median values (score 1: poor, 2: good, 3: very good, 4: excellent) of antibiofilm effectiveness of various types of extracts from the two herbs. Values close to 1 indicate poor biofilm inhibition (0-50%) while values close to 4 indicate excellent biofilm inhibition (>95%), Similar letters above bars indicate no significant difference in antibiofilm effectiveness between similar types of extracts among the various herbal species and antibiotics (Kruskal-Wallis, p<0.05).
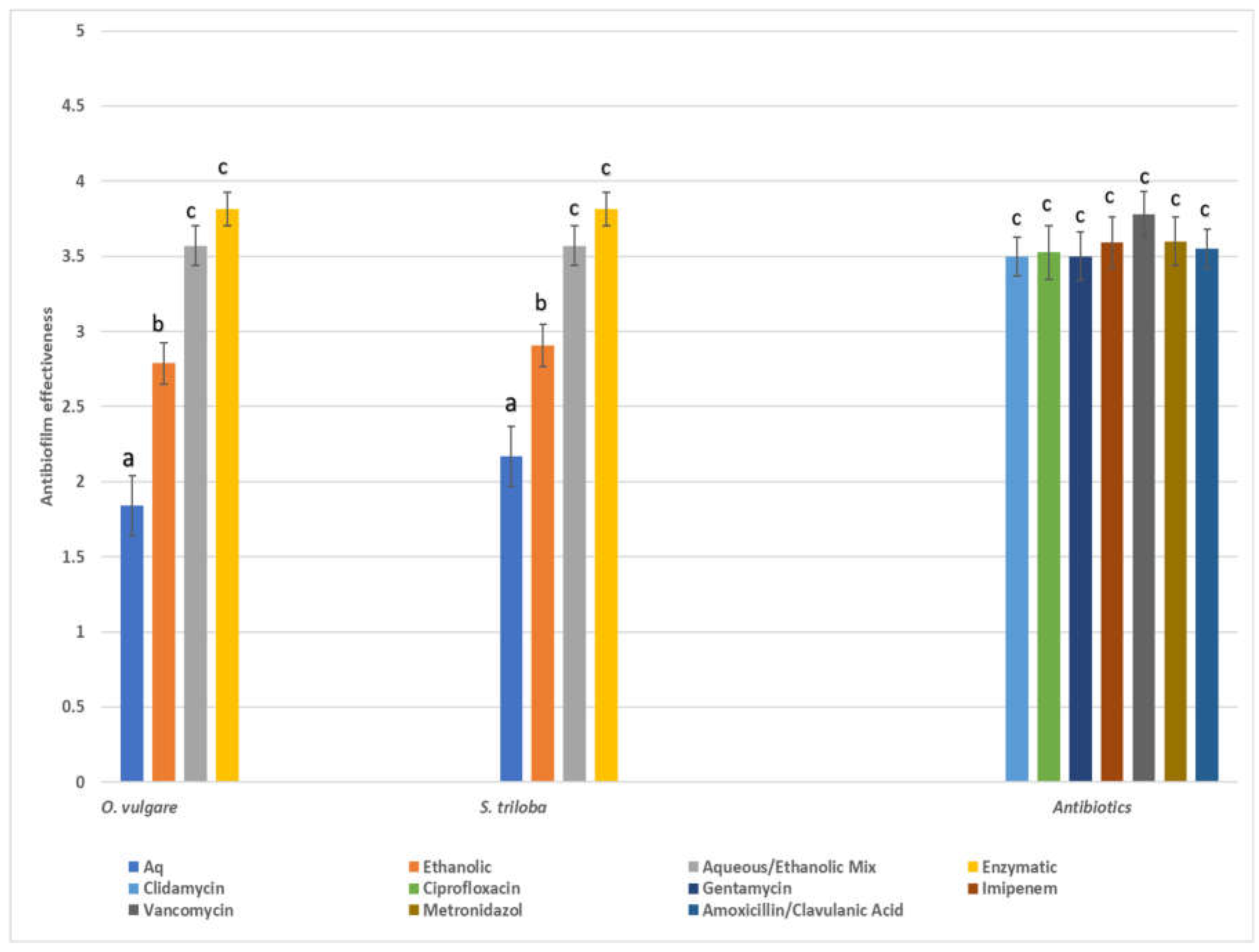
Figure 5.
Hemolytic activity of the various O. vulgare extracts in human blood type O erythrocytes compared to the negative and positive (Triton X-100) controls.
Figure 5.
Hemolytic activity of the various O. vulgare extracts in human blood type O erythrocytes compared to the negative and positive (Triton X-100) controls.
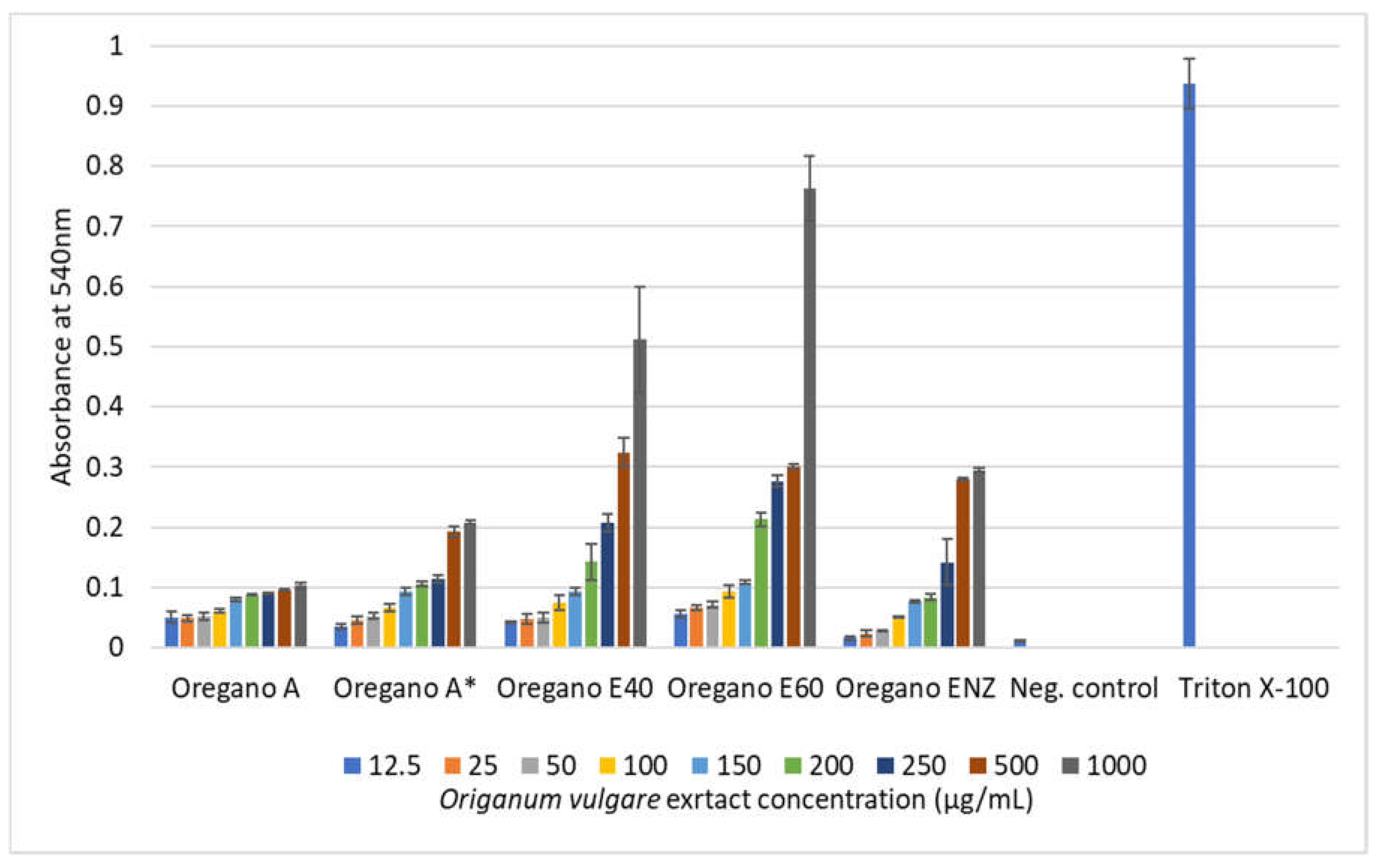
Figure 6.
Hemolytic activity of the various S. triloba extracts in human blood type O erythrocytes compared to the negative and positive (Triton X-100) controls.
Figure 6.
Hemolytic activity of the various S. triloba extracts in human blood type O erythrocytes compared to the negative and positive (Triton X-100) controls.
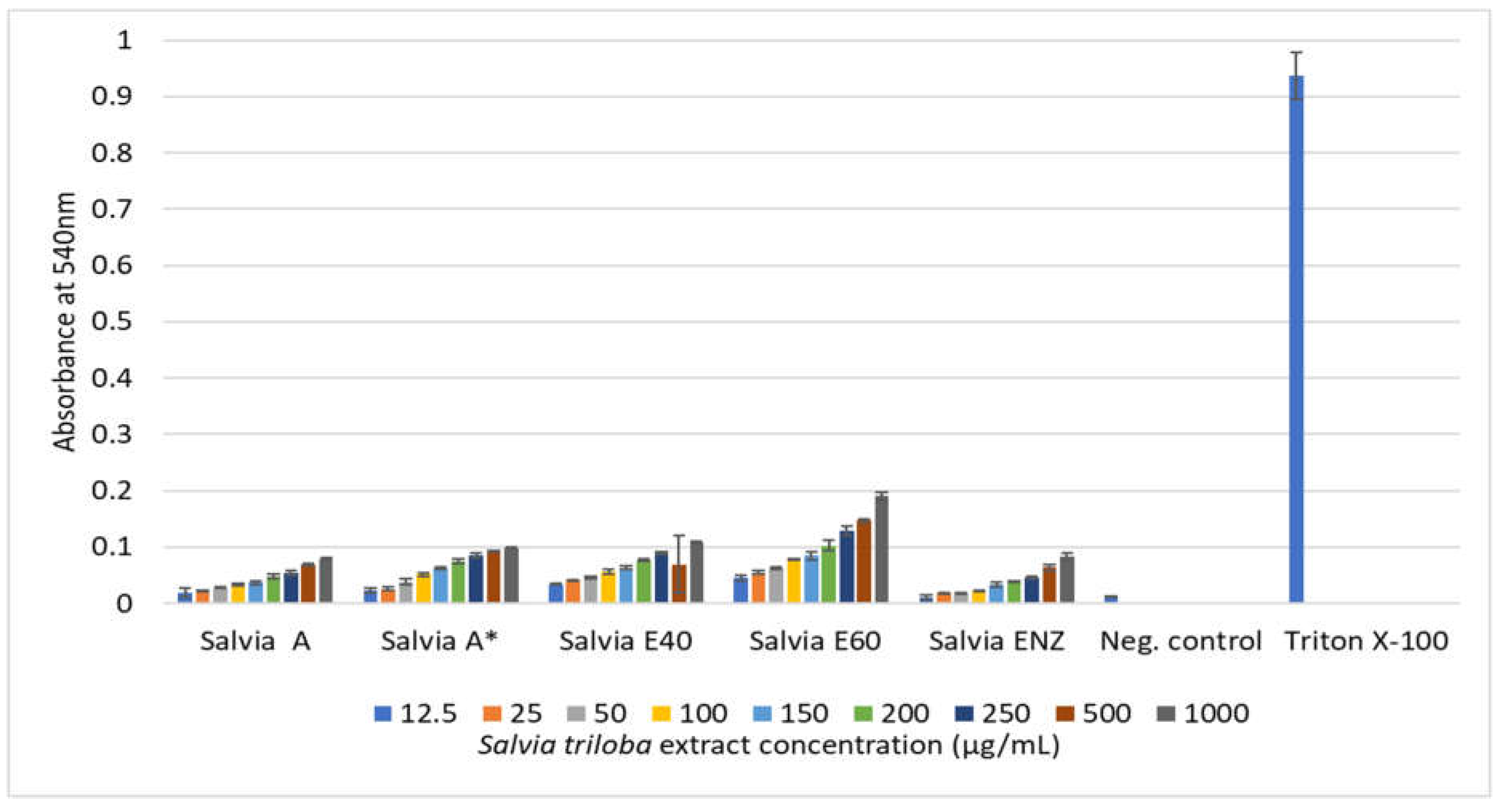

Table 1.
Strains of pathogenic bacteria used in the study.
| Isolate | Source | Characterization based on Gram staining |
Oxygen requirements for growth/ Growth conditions used in the present study |
| Streptococcus mutans | Oral cavity | Gram+ | Facultatively anaerobic/ Growth under anaerobic conditions |
| Streptococcus salivarius | Oral cavity | Gram+ | Facultatively anaerobic/ Growth under anaerobic conditions |
| Fusobacterium nucleatum | Oral cavity | Gram- | Obligate anaerobic / Growth under anaerobic conditions |
| Porphyromonas gingivalis | Oral cavity | Gram- | Obligate anaerobic / Growth under anaerobic conditions |
| Prevotella intermedia | Oral cavity | Gram- | Obligate anaerobic/ Growth under anaerobic conditions |
| Parvimonas micra | Oral cavity | Gram+ | Obligate anaerobic / Growth under anaerobic conditions |
| Staphylococcus aureus, Methicillin, and vancomycin resistant | Dental caries area | Gram+ | Best in the presence of oxygen-rich environment/ Growth under aerobic conditions |
| Methicillin-Resistant S. aureus | Raw milk | Gram+ | Best in the presence of oxygen-rich environment/ Growth under aerobic conditions |
| Methicillin-Resistant S. aureus | Poultry meat | Gram+ | Best in the presence of oxygen-rich environment/ Growth under aerobic conditions |
|
Staphylococcus aureus subsp. aureus |
Reference strain ATCC 6538 | ||
Table 2.
Phytochemical constituents present in individual plant extracts.
| Phytochemical compounds | Plants extracts | |||||||
|---|---|---|---|---|---|---|---|---|
| Origanum vulgare | Salvia triloba | |||||||
| A | Ε40 | Ε60 | ΕΝΖ | A | Ε40 | Ε60 | ΕΝΖ | |
| Alkaloids | + | - | - | + | + | - | - | - |
| Anthraquinones | - | - | - | + | + | - | - | - |
| Terpenoids (Salkowski’s test) | + | + | + | + | + | + | + | + |
| Steroids | + | - | - | + | - | - | - | - |
| Saponins | - | + | + | - | + | + | - | - |
| Flavonoids (alkaline reagent test) | + | + | + | + | + | + | + | + |
| Tannins (ferric chloride test) | + | + | + | - | + | + | + | + |
| Glycosides (Keller-Kiliani test) | + | + | + | + | + | + | + | - |
A—aqueous extract, E40—ethanolic extract (40% v/v aqueous ethanol), E60—ethanolic extract (60% v/v aqueous-ethanol), ENZ—enzymatic extract; +: positive detection, -: not detected.
Table 3.
Total phenolics and flavonoid content of O. vulgare and S. triloba plant extracts.
| Plant extracts | Total phenolics (mg gallic acid equivalent/g of dried sample) | Total Flavonoids (mg catechin equivalent per /g of dried sample) |
|---|---|---|
| Oregano A | 39.69±0.54b | 8.13±0.82a |
| Oregano A* | 45.46±0.8c | 15.9±0.38b |
| Oregano E40 | 76.68±3.34e | 22.56±0.48c |
| Oregano E60 | 88.44±0.6f | 30.74±0.98d |
| Oregano ENZ | 121.4±0.6g | 48.83±1.17f |
| Salvia A | 91.99±1.08f | 55.31±0.35g |
| Salvia A* | 79±0.8e | 37.58±1.15e |
| Salvia E40 | 50.76±0.6d | 37.56±0.47e |
| Salvia E60 | 22.44±0.6a | 39.67±0.44e |
| Salvia ENZ | 90.42±0.5f | 64.75±0.65h |
Values are the mean of three replicates (TPC). Different letters in columns denote statistical differences between the total phenolics or total flavonoids among the various extracts in each plant (Kruskal-Wallis, p<0.05 with Tukey’s HSD).
Table 4.
Percentage of neutralization of the DPPH Radical by Plants Extracts in the DPPH Assay.
| Plant extracts | Concentration (μg/ml) | |||||
|---|---|---|---|---|---|---|
| 1 | 10 | 50 | 100 | 300 | 500 | |
| Oregano A | 3,42±0,33bc | 10,57±0,13c | 33,34±0,58b | 38,55±0,21b | 52,24±0,22a | 59,32±0,85a |
| Oregano A* | 5,28±0,1d | 12,18±0,42d | 40,86±0,02e | 45,26±0,53c | 59,52±0,4b | 62,92±0,35b |
| Oregano E40 | 4,12±0,11cf | 12,19±0,23d | 50,46±0,34g | 70,48±0,07g | 74,5±0,39e | 77,33±0,11d |
| Oregano E60 | 12,37±0,28g | 18,06±0,19f | 55,24±0,21h | 76,79±0,22h | 80,7±0,17f | 81,36±0,25e |
| Oregano ENZ | 11,6±0,45ef | 14,32±0,34e | 57,99±0,22i | 78,77±0,17i | 82,44±0,42g | 86,43±0,42g |
| Salvia A | 11,5±0,39e | 14,12±0,12e | 39,53±0,45d | 60,53±0,33f | 82,21±0,32g | 84,24±0,29f |
| Salvia A* | 3,42±0,16bc | 9,36±0,27b | 32,38±0,44b | 48,43±0,34d | 80,13±0,29f | 82,38±0,44e |
| Salvia E40 | 2,63±0,2b | 10,22±0,19c | 35,27±0,22c | 48,36±0,39d | 72,37±0,26d | 74,49±0,42c |
| Salvia E60 | 1,37±0,41a | 6,33±0,25a | 22,38±0,41a | 30,31±0,23a | 54,97±0,44b | 60,53±0,41a |
| Salvia ENZ | 12,71±0,24g | 17,37±0,42f | 42,73±0,25f | 59,25±0,18e | 85,46±0,21h | 87,14±0,71g |
Values are the mean of three replicates. Different letters in columns denote statistical differences among the various extracts in each plant (Kruskal-Wallis, p<0.05 with Tukey’s HSD).
Table 5.
Reducing power of different concentrations of extracts from O. vulgare and S. triloba as evaluated by the FRAP assay.
Table 5.
Reducing power of different concentrations of extracts from O. vulgare and S. triloba as evaluated by the FRAP assay.
| Plant extracts | Concentration (μg/ml) | |||||
|---|---|---|---|---|---|---|
| 2.5 | 5 | 10 | 15 | 20 | 25 | |
| Oregano A | 0.15±0a | 0,21±0a | 0.31±0a | 0.57±0a | 0.77±0a | 0.85±0a |
| Oregano A* | 0.21±0c | 0,40±0c | 0.55±0f | 0.81±0c | 1.01±0c | 1.21±0d |
| Oregano E40 | 0.22±0d | 0,49±0g | 0.75±0h | 1.28±0h | 1.44±0g | 1.62±0f |
| Oregano E60 | 0.24±0c | 0,55±0h | 0.83±0i | 1.34±0i | 1.55±0h | 1.77±0g |
| Oregano ENZ | 0.23±0c | 0,58±0i | 0.99±0j | 1.48±0j | 1.65±0i | 1.93±0i |
| Salvia A | 0.24±0c | 0,37±0d | 0.44±0bc | 0.78±0d | 0.98±0d | 1.33±0c |
| Salvia A* | 0.18±0b | 0,32±0c | 0.54±0f | 0.71±0c | 0.86±0bc | 1.11±0c |
| Salvia E40 | 0.19±0b | 0,31±0c | 0.48±0d | 0.68±0b | 0.85±0b | 1.09±0b |
| Salvia E60 | 0.18±0b | 0,28±0b | 0.38±0b | 0.70±0c | 0.85±0b | 1.08±0b |
| Salvia ENZ | 0.26±0f | 0,38±0d | 0.51±0c | 0.88±0f | 1.31±0f | 1.76±0g |
| Reference | ||||||
| Gallic acid (GA) | 0.18±0.0b | 0.43±0f | 1.81±0k | 1.85±0k | 1.88±0j | 1.88±0h |
| Ascorbic acid (AA) | 0.19±0.0b | 0.42±0f | 0.68±0g | 1.12±0g | 1.65±0i | 1.88±0h |
Table 6.
Antibacterial activities (disk diffusion) of various concentrations (10, 20, 50 and 100%) of aqueous (A), ethanolic (E40 & E60) and enzymatic (Enz) extracts from O. vulgare against oral/food origin pathogens and a reference strain.
Table 6.
Antibacterial activities (disk diffusion) of various concentrations (10, 20, 50 and 100%) of aqueous (A), ethanolic (E40 & E60) and enzymatic (Enz) extracts from O. vulgare against oral/food origin pathogens and a reference strain.
| Pathogen | Disk Content (%) | A(mm) | E40(mm) | E60(mm) | Enz(mm) |
|---|---|---|---|---|---|
| S. aureus MRSA/VRSA | 10 | 6±0.1a | 7.5±0.5a | 9.7±0.6ab | 11.5±0.4d |
| 20 | 13.8±0.8ef | 10±0.6ab | 14.7±0.5b | 15±0.5e | |
| 50 | 15.7±0.5ab | 16.2±0.6b | 18.3±0.7a | 19.1±1.1b | |
| 100 | 18.1±0.3ab | 19.3±0.6bc | 21±0.1b | 22.4±0.1b | |
| MRSA (raw milk) | 10 | 6.5±0.3a | 8.4±0.3a | 10.8±0.6bc | 8.7±0.5ab |
| 20 | 10.8±0.3bc | 10.4±0.3ab | 15.1±0.4b | 15.3±0.9e | |
| 50 | 9.7±6.8a | 14.1±0.1a | 17.6±0.5a | 19±0.9b | |
| 100 | 18.2±0.4ab | 17.5±0.5a | 18.8±0.6a | 20.4±0.4ab | |
| MRSA (raw poultry) | 10 | 6.9±0.1a | 9.8±0.6bc | 12.2±0.2d | 8±0.5a |
| 20 | 11.5±0.4de | 13.2±0.6de | 18.5±0.3c | 15.1±0.4e | |
| 50 | 22.9±5.2b | 19.5±0.3d | 27.8±0.9d | 18.7±0.5b | |
| 100 | 19.2±0.4cd | 26.5±0.6d | 38.7±0.8f | 22.1±0.5b | |
| S. mutans | 10 | 9.6±0.7c | 8.1±0.3a | 12.1±0.8c | 9.5±0.4b |
| 20 | 12.6±0.4de | 11.2±0.6ab | 19.3±0.5de | 12.4±0.2bc | |
| 50 | 17.1±0.4ab | 17.3±0.6cd | 28.6±0.9d | 18.8±0.3b | |
| 100 | 19.8±0.5cd | 19.8±0.4bc | 31.4±0.6e | 22.3±2.7b | |
| S. salivarius | 10 | 6.2±0.2a | 8.1±0.2a | 10.7±0.5b | 7.7±0.6a |
| 20 | 9.8±0.3ab | 11.8±0.3bcd | 18.5±0.7cd | 11.5±0.5ab | |
| 50 | 15.2±0.4ab | 15.9±0.4b | 25±0.4c | 17±0.1b | |
| 100 | 19±0.3cd | 20.7±1.3c | 28.2±0.5c | 19.6±0.5ab | |
| P. gingivalis | 10 | 8.6±0.5bc | 10.7±0.5cd | 12.3±0.4d | 9.7±0.5bc |
| 20 | 13.5±0.4ef | 14.8±0.1e | 19.5±0.6de | 14.1±0.7de | |
| 50 | 18.2±0.5b | 19.5±0.3d | 29±0.3d | 19±0.3b | |
| 100 | 20.6±0.7d | 19.9±0.1bc | 31.1±0.2e | 21.8±0.4b | |
| F. nucleatum | 10 | 6.1±0.2a | 7.9±0.2a | 13.3±0.2d | 10.1±0.2c |
| 20 | 8.9±0.6a | 14.8±0.2e | 19.1±0.2de | 13.8±0.5cd | |
| 50 | 15.3±0.5ab | 18.1±0.7cd | 28.2±0.5d | 18.7±0.7b | |
| 100 | 17.8±0.5ab | 20.2±0.3c | 32.5±1.5ef | 21.9±0.3b | |
| P. intermedia | 10 | 6.1±0.1a | 8.6±0.4ab | 10.6±0.2b | 8.4±0.7a |
| 20 | 12.5±0.4ef | 12.4±0.5cd | 17.2±0.5c | 11.2±0.8a | |
| 50 | 16.8±0.5ab | 17.1±0.3bc | 21.9±1.8b | 18.1±0.2b | |
| 100 | 18.8±0.3abc | 19.1±0bc | 29.3±0.6b | 21.9±0.7b | |
| P. micra | 10 | 6.3±0.6a | 11.7±0.5d | 12.6±0.6d | 9.6±0.5b |
| 20 | 14.8±0.4f | 18.2±0.4f | 20±0.2e | 13.3±0.2cd | |
| 50 | 16.9±0.4ab | 24.2±1.1e | 27.9±0.4d | 18±0.2b | |
| 100 | 18.2±0.2ab | 31.4±0.4e | 31.1±0.1e | 20.6±0.4ab | |
| S. aureus ATCC 12600 (Ref) | 10 | 8.3±0.5b | 8.1±0.6a | 9.1±0.1a | 10.2±0.3c |
| 20 | 12.8±0.5de | 12.5±1.3c | 12.3±0.6a | 13±0.3bc | |
| 50 | 16.6±0.4ab | 17.4±0.6bc | 18±0.3a | 17.8±0.4ab | |
| 100 | 17.6±0.5a | 19.1±0.2bc | 18.7±0.5a | 19.2±0.4a |
Different letters in a row indicate significant differences among the various extracts for similar disk content (Kruskal-Wallis with Tukey’s HSD, p<0.05).
Table 7.
Antibacterial activities (disk diffusion) of various concentrations (10, 20, 50 and 100%) of aqueous (A), ethanolic (E40 & E60) and enzymatic (Enz) extracts from S. trilova against oral/food origin pathogens and a reference strain.
Table 7.
Antibacterial activities (disk diffusion) of various concentrations (10, 20, 50 and 100%) of aqueous (A), ethanolic (E40 & E60) and enzymatic (Enz) extracts from S. trilova against oral/food origin pathogens and a reference strain.
| Pathogen | Disk Content (%) | A(mm) | E40(mm) | E60(mm) | Enz(mm) |
|---|---|---|---|---|---|
| S. aureus MRSA/VRSA | 10 | 6.7±0.6ab | 10±0.7cd | 10.5±0.5abc | 6±0a |
| 20 | 11.3±0.4bc | 12.6±0.2cd | 15±0.4bc | 8.5±0.6a | |
| 50 | 13.4±0.4ac | 17.5±0.5cd | 18.9±0.5bc | 12.2±0.3a | |
| 100 | 17.6±0.3abc | 18.5±0.3bc | 22.8±2ab | 17.9±0.4a | |
| MRSA (raw milk) | 10 | 7.6±0.6ab | 12.5±0.4e | 12.6±0.3d | 7.4±0.5b |
| 20 | 11.9±0.7bc | 15±0.2e | 18.6±0.3ef | 12±0.4c | |
| 50 | 14.8±0.5bc | 18±0.3d | 23.2±0.6e | 14.6±0.2b | |
| 100 | 17.6±0.3abc | 24.3±0.4e | 28.7±0.6c | 17.4±0.4a | |
| MRSA (raw poultry) | 10 | 6.4±0.4a | 9.1±0.7ab | 11.4±0.7bcd | 7.8±0.5bc |
| 20 | 10.6±0.1bc | 10.6±0.2a | 16.2±0.6cd | 10.7±0.4b | |
| 50 | 14.9±0.2bc | 16.2±0.3bc | 20.6±0.5cd | 15.2±0.4bc | |
| 100 | 17.4±0.4bc | 18.3±0.6abc | 29.1±0.8c | 17.7±0.3a | |
| S. mutans | 10 | 10.8±0.8de | 8.6±0.3ab | 9.8±0.4a | 12.1±0.5e |
| 20 | 16.4±0.4f | 10.7±0.3ab | 11.9±0.5a | 17.1±0.2cd | |
| 50 | 20.7±0.8g | 15.2±0.3ab | 16.7±0.5ab | 21.5±0.7e | |
| 100 | 29.3±0.9f | 16.7±0.2a | 18.5±0.5a | 30.8±0.6b | |
| S. salivarius | 10 | 8.4±0.8bc | 10.8±0.6de | 11.7±0.4cd | 9.3±0.3d |
| 20 | 12.9±0.6de | 13.1±0.8d | 17.4±0.5de | 12.8±0.5b | |
| 50 | 16.4±0.5de | 16.3±0.5bc | 22.2±0.7de | 17.8±0.3d | |
| 100 | 18.5±0.6cd | 18.5±0.5bc | 27.5±5.5bc | 18.5±0.4a | |
| P. gingivalis | 10 | 11.2±0.8e | 7.6±0.5a | 10.2±0.2ab | 12.2±0.6e |
| 20 | 15.2±0.5f | 11.9±0.3c | 14±0.7b | 18.2±0.3de | |
| 50 | 20.6±0.5fg | 14.1±0.3a | 17±0.2ab | 28.1±1g | |
| 100 | 32.3±1.9g | 17.5±0.3ab | 18.6±0.4a | 33.8±1.2cd | |
| F. nucleatum | 10 | 6.1±0.1a | 9.8±0.4bcd | 11.9±0.6cd | 9.8±0.4d |
| 20 | 9.3±0.6a | 10.4±0.4a | 17.2±0.2de | 16.7±0.4c | |
| 50 | 12.4±0.6a | 13.9±0.2a | 24.1±1.4e | 19.6±0.8d | |
| 100 | 16±0.2b | 16.8±0.2a | 28.9±0.9c | 30.3±0.8b | |
| P. intermedia | 10 | 9.6±0.2cde | 8.3±0.6ab | 11.7±0.5cd | 9±0.1cd |
| 20 | 13.7±0.2e | 12±0.4c | 19±0.4f | 12.5±0.7b | |
| 50 | 19±0.2ef | 17.3±0.4cd | 28.4±0.6f | 16.7±0.4cd | |
| 100 | 24.5±0.5e | 19.1±0.8c | 31.7±0.6c | 17.9±0.1a | |
| P. micra | 10 | 9.5±0.6cd | 10.6±0.6cd | 11±0.5abc | 12±0.5e |
| 20 | 12.4±0.4de | 15.3±0.5e | 16.2±0.3cd | 19.3±0.5e | |
| 50 | 17.8±0.9de | 21.6±1.2e | 20.8±0.5de | 29.3±0.5f | |
| 100 | 21.2±0.9d | 27.8±1.1f | 30.9±0.3c | 35.4±1.1d | |
| S. aureus ATCC 12600 (Ref) | 10 | 9.4±0.5cd | 10.9±1.1de | 10.5±0.6abc | 11.1±0.1e |
| 20 | 12.9±0.5de | 14.9±0.1e | 18.1±0.6ef | 18±0.2de | |
| 50 | 17.5±0.6de | 18.3±1d | 26.7±0.7f | 22.2±1e | |
| 100 | 19.8±0.4cd | 21±0.4d | 30.7±0.4c | 31.8±2bc |
Different letters in a row indicate significant differences among the various extracts for similar disk content (Kruskal-Wallis with Tukey’s HSD, p<0.05).
Table 8.
Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC) of the various extracts (A-Aqueous, E40 & E60 – Ethanolic 40% and 60% and Enz-Enzymatic) from O. vulgare, against pathogens. Mean values (mg/mL) from three repetitions.
Table 8.
Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC) of the various extracts (A-Aqueous, E40 & E60 – Ethanolic 40% and 60% and Enz-Enzymatic) from O. vulgare, against pathogens. Mean values (mg/mL) from three repetitions.
| Minimum Inhibitory Concentration (mg/mL) |
Minimum Bactericidal Concentration (mg/mL) |
|||||||
|---|---|---|---|---|---|---|---|---|
| Pathogen | A | E40 | E60 | Enz | A | E40 | E60 | Enz |
| S. aureus MRSA/VRSA | 6.3±0b | 3.1±0b | 1.6±0b | 1.6±0b | 12.5±0b | 6.3±0c | 3.1±0b | 1.6±0a |
| MRSA (raw milk) | 6.3±0b | 6.3±0 | 3.1±0c | 3.1±0c | 12.5±0b | 12.5±0 | 12.5±0d | 6.3±0c |
| MRSA (raw poultry) | 3.1±0a | 0.8±0a | 0.4±0a | 0.8±0a | 6.3±0a | 1.6±0a | 1.6±0a | 1.6±0a |
| S. mutans | 3.1±0b | 3.1±0b | 1.6±0b | 3.1±0c | 6.3±0a | 12.5±0d | 3.1±0b | 6.3±0c |
| S. salivarius | 6.3±0b | 3.1±0b | 1.6±0b | 3.1±0c | 12.5±0b | 6.3±0c | 6.3±0c | 6.3±0c |
| P. gingivalis | 6.3±0b | 6.3±0c | 1.6±0b | 0.8±0a | 25±0c | 12.5±0d | 3.1±0b | 6.3±0c |
| F. nucleatum | 6.3±0b | 3.1±0b | 0.8±0a | 3.1±0c | 25±0c | 12.5±0d | 3.1±0b | 3.1±0b |
| P. intermedia | 6.3±0b | 3.1±0b | 0.8±0a | 3.1±0c | 25±0c | 12.5±0d | 6.3±0c | 12.5±0d |
| P. micra | 12.5±0c | 0.8±0a | 0.8±0a | 1.6±0b | 25±0c | 3.1±0b | 6.3±0c | 6.3±0c |
| S. aureus ATCC 12600 (Ref) | 6.3±0b | 3.1±0b | 3.1±0c | 1.6±0b | 12.5±0b | 12.5±0d | 12.5±0d | 6.3±0c |
Different superscript letters in a column indicate significant differences in MIC and MBC among the various extracts (ANOVA with Tukey’s HSD, p<0.05).
Table 9.
Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC) of the various extracts (A-Aqueous, E40 & E60 – Ethanolic 40% and 60% and Enz-Enzymatic) from S. triloba, against pathogens. Mean values (mg/mL) from three repetitions.
Table 9.
Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC) of the various extracts (A-Aqueous, E40 & E60 – Ethanolic 40% and 60% and Enz-Enzymatic) from S. triloba, against pathogens. Mean values (mg/mL) from three repetitions.
| Minimum Inhibitory Concentration (mg/mL) |
Minimum Bactericidal Concentration (mg/mL) |
|||||||
|---|---|---|---|---|---|---|---|---|
| Pathogen | A | E40 | E60 | Enz | A | E40 | E60 | Enz |
| S. aureus MRSA/VRSA | 12.5±0e | 3.1±0b | 1.6±0b | 6.3±0d | 50±0d | 12.5±0c | 6.3±0d | 25±0f |
| MRSA (raw milk) | 6.3±0d | 3.1±0b | 1.6±0b | 6.3±0d | 12.7±0.3b | 12.5±0c | 3.1±0c | 25±0f |
| MRSA (raw poultry) | 12.5±0e | 6.3±0c | 0.8±0a | 3.1±0c | 50±0d | 12.5±0c | 3.1±0c | 12.5±0e |
| S. mutans | 0.8±0a | 6.3±0c | 3.1±0c | 0.8±0a | 6.3±0a | 25±0d | 12.5±0d | 3.1±0c |
| S. salivarius | 3.1±0c | 3.1±0b | 0.8±0a | 6.3±0d | 12.7±0.3b | 6.3±0b | 1.6±0b | 12.5±0 |
| P. gingivalis | 0.8±0a | 6.3±0c | 6.3±0d | 0.8±0a | 6.3±0a | 25±0d | 12.5±0d | 1.6±0b |
| F. nucleatum | 12.5±0 | 6.3±0c | 1.6±0b | 1.6±0b | 50±0d | 12.5±0c | 3.1±0c | 1.6±0b |
| P. intermedia | 1.6±0b | 3.1±0b | 0.8±0a | 3.1±0c | 12.7±0.3b | 12.5±0c | 0.8±0a | 6.3±0d |
| P. micra | 1.6±0b | 1.6±0a | 0.8±0a | 0.4±0a | 6.3±0a | 3.1±0a | 0.8±0a | 1.6±0b |
| S. aureus ATCC 12600 (Ref) | 6.3±0d | 1.6±0a | 0.8±0a | 0.7±0.2a | 25±0c | 3.1±0a | 3.1±0c | 0.8±0a |
Different superscript letters in a column indicate significant differences in MIC and MBC among the various extracts (ANOVA with Tukey’s HSD, p<0.05).
Table 10.
Percentage (%) of hemolysis of human blood type “AB” erythrocytes treated with various O. vulgare and S. triloba extracts as compared to the positive control.
Table 10.
Percentage (%) of hemolysis of human blood type “AB” erythrocytes treated with various O. vulgare and S. triloba extracts as compared to the positive control.
| Concentration of each plant extract (μg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 12.5 | 25 | 50 | 100 | 150 | 200 | 250 | 500 | 1000 | |
| Oregano A | 4.35±11de | 4.21±0.61bc | 4.46±0.71cde | 5.45±0.31bc | 7.57±0.32c | 8.45±0.123c | 8.64±0.123bcd | 9.37±0.234ab | 10.1±0.54ab |
| Oregano A* | 2.74±0.51bc | 3.81±0.71bc | 4.54±0.612de | 6.04±0.72bcd | 9.04±0.73d | 10.43±0.534c | 11.34±0.74cde | 19.9±0.95c | 21.62±0.45cd |
| Oregano E40 | 3.55±0.11cd | 4.1±0.912bc | 4.17±0.912cd | 7.03±1.412cd | 9.07±0.712d | 14.42±3.323d | 21.51±1.63f | 34.39±2.64d | 54.92±9.75e |
| Oregano E60 | 4.94±0.61de | 6.07±0.41d | 6.62±0.51f | 9.04±1.11a | 10.68±0.31e | 22.14±1.22ce | 29.09±13g | 31.76±0.53d | 82.58±5.94f |
| Oregano ENZ | 0.55±0.41a | 1.43±0.51a | 1.94±0.11ab | 4.5±0.112b | 7.24±0.22c | 8.01±0.52 | 14.38±4.13e | 29.42±0.24d | 31.1±0.54 |
| Salvia A | 0.88±0.21ab | 1.32±0.112a | 1.9±0.123ab | 2.49±0.23a | 2.89±0.34a | 4.06±0.45ab | 4.76±0.65ab | 6.29±0.36a | 7.46±0.37a |
| Salvia A* | 1.39±0.41a | 1.61±0.31a | 3.04±0.72bc | 4.46±0.33b | 5.67±0.34b | 7.1±0.55bc | 8.23±0.56abc | 9.04±0.167a | 9.59±0.17ab |
| Salvia E40 | 2.6±0.31bc | 3.33±0.112b | 3.81±0.212cd | 4.9±0.412b | 5.74±0.4123b | 7.24±0.2123bc | 8.6±0.223bcd | 6.41±5.63a | 10.76±0.23ab |
| Salvia E60 | 3.77±0.31cde | 4.87±0.412cd | 5.63±0.22ef | 7.32±0.134de | 8.12±0.84cd | 10.06±15c | 12.99±0.96e | 14.89±0.47bc | 19.72±0.88bc |
| Salvia ENZ | 0.18±0.31a | 0.8±0.112a | 0.91±0.112a | 1.24±0.12a | 2.56±0.53a | 3.11±0.234a | 3.92±0.34a | 5.89±0.65a | 7.98±0.76a |
Similar superscript numbers indicate no statistical significant differences in rows and similar superscript letters indicate no statistical significant differences in columns (ANOVA with 95.0% Tukey HSD).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



