
Submitted:
04 May 2024
Posted:
06 May 2024
You are already at the latest version
Abstract
The aim was to identify if surface electromyography (sEMG) parameters are indicative of post-activation potentiation (PAP)/post-activation performance enhancement (PAPE), in terms of twitch potentiation and voluntary performance. Three databases were used in March 2024, with the following inclusion criteria: (a) original research, assessed in healthy human adults, (b) sEMG parameters were measured. The exclusion criteria were: (a) studies with no PAP/PAPE-protocol, (b) non-randomized control trials. The following data were extracted: study characteristics/demographics, PAP/PAPE-protocols, sEMG parameters, twitch/performance outcomes, and study findings. A modified physiotherapy evidence database (PEDro) scale was used for quality assessment. Fifteen randomized controlled trials (RCTs), with a total of 199 subjects were included. The M-wave amplitude (combined with a twitch torque outcome) was shown to generally be indicative of PAP. The sEMG amplitude (in some muscles) were found to be indicative of PAPE during ballistic movements, while a small decrease in the MdF (in certain muscles) were shown to reflect PAPE. Changes in the Hmax/Mmax ratio were found to contribute (temporally) to PAP, while the H-reflex amplitude was shown to be neither indicative of PAP nor PAPE. This review provides preliminarily findings suggesting that certain sEMG parameters could be indicative of PAP/PAPE. However, future research is warranted.
Keywords:
post-activation potentiation
; electromyography
; conditioning activities
; post-activation performance enhancement
1. Introduction
It is well documented that warming up prior to any physical activity may reduce the risk of musculoskeletal injuries [1,2] and can produce a noticeable improvement in athletic performance [3,4]. One of the mechanisms that has been of interested towards power-strength athletes and coaches, is the muscle potentiation effect that has been observed following warmups [5]. This enhancement, in response to voluntary muscle contractions, has been coined post-activation potentiation (PAP) and refers to a state where the contractile properties of the skeletal muscle (s) are acutely enhanced following a brief high-intensity contraction [5,6,7]. The PAP phenomenon was originally confirmed by measuring the maximum twitch force (or peak twitch torque [PTT]) that is evoked by supramaximal electrical stimulation [6,8], with PTT and maximal twitch rate of torque development (RTDTW), being the most common twitch outcomes of PAP, or also referred to as twitch potentiation [6,9,10]. Numerous mechanisms have been proposed to cause this potentiated twitch response or PAP effect, but currently it is believed that the primary mechanism for PAP is an increase expression of myosin regulatory light chain (MRLC) phosphorylation [5,11]. During maximal or near-maximum voluntary contractions, an increased influx of sarcoplasmic calcium (Ca2+) into the myoplasm upregulates the expression of skeletal muscle myosin light-chain kinases (skMLCK) [11,12]. This increased expression of skMLCK, phosphorylates the myosin subfragment-1 (S1) head closer to its joints with the subfragment-2 (S2) portion [12,13]. This sequentially augments the probability of a cross bridge to occur, improving myosin head’s mobility and allows potentiated fibers to improve their rate of force development (RFD), and this consequently enhances its contractility [5,11,12]. Practically, many sports coaches and athletes have therefore employed several different PAP strategies or conditioning activity (CA) protocols, with the aim of acutely enhancing voluntary force production and overall athletic performance [6,14]. However, a major limitation in the PAP literature is that the term has loosely been used to explain all acute improvements in voluntary performance, following different CA protocols, without a direct twitch verification test (i.e. observing an acute increased in PTT or RTDTW) [6,9,10]. As accumulating evidence indicate that acute improvements in voluntary performance can occur independent of any changes in PAP (by its classical definition), the term ‘post activation performance enhancement’ (PAPE) has in more recent years been used to describe the acute improvements in voluntary muscular performance following different CAs [6,15]. Common approaches to evaluate PAPE usually include observing acute improvements in plyometric performance, such as countermovement jump (CMJ) height [15,16] and squat jump (SJ) height [15,17], in addition to enhanced sprinting speed [18,19], RFD and peak power output (PPO) in different movements [15,17], following different CAs.
Further, while various CA protocols have been demonstrated to acutely enhance voluntary muscular performance, some interventional studies have actually reported a reduction in performance following different CA protocols [20,21], which has primarily been attributed to fatigue. Although there is no standard definition of fatigue [22], at the neuromuscular level, it has been defined as the failure to sustain a specified force output with a muscle or muscle group during exercise [23,24]. Numerous research instruments have been employed to distinguish between intramuscular (peripheral) and central (neural) processes that contributes to fatigue within the neuromuscular system, such as electrical stimulation [25], the interpolated twitch technique [26] and electromyography (EMG)/surface electromyography (sEMG) [27,28]. Among these research tools, sEMG has commonly been used as a convenient non-invasive tool to assess neuromuscular fatigue [28]. In particular, EMG spectral variables (e.g. mean power frequency [MPF] and median power frequency [MdF]) and the sEMG amplitude (e.g. mean absolute values [MAV] and root mean squared [RMS]) have commonly been evaluated [29,30,31], as changes usually occurs in these parameters during fatiguing muscular activity [29,31,32]. Although sEMG has extensively been used to indirectly estimate neuromuscular fatigue [30], there is conflicting findings regarding the use of different sEMG parameters for assessing neural mechanisms that could modulate the PAP/PAPE response [33,34].
Normally, muscle potentiation and fatigue coexist when performing any muscular activity [35]. However, the post-stimulus state will be affected by the net balance between these two factors. Depending on the dissipation of fatigue and the decay rate of potentiation following recovery, there may be a net potentiated effect, a net attenuated effect or unaltered state in comparison to the pre-stimulus state [5,35]. Further, experimental evidence suggests that the net balance between the PAP/PAPE response and neuromuscular fatigue are also influenced by training experience [36], the rest period [37] and the intensity of the CA [38] making the use of sEMG to provide insights into the neural mechanisms of PAP/PAPE more complex. Interestingly, assessing changes in PTT and muscle compound action potentials or M-waves (evoked via electrical stimulation and with the use of various sEMG channels), has in recent years been used as a method for confirming the presence of PAP [10,39,40]. The M-wave has commonly been used to examine peripheral properties of the neuromuscular system without the involvement of the central nervous system [41,42]. Further, importantly, eliciting the maximal M-wave (Mmax), via supramaximal nerve stimulation, activates all motor units of the pool, including the fast-twitch units [43], which are more responsive to MRLC phosphorylation (i.e. the primary mechanism of PAP) [6,44]. By assessing the M-wave, researchers can control for changes in neuromuscular propagation that could influence the PTT in the stimulated muscle (e.g. changes in sarcolemmal membrane excitability [10,45,46]. Furthermore, another common parameter elicited with electrical stimulation, and assessed with sEMG, is an electrically induced spinal reflex or H (Hoffmann) reflex [47]. The H-reflex measures the potency of synaptic transmission [47,48] and is frequently used to denote α-motoneurons excitability [48,49,50]. Still, our understanding of the H-reflex and its contribution to PAP/PAPE following different CAs protocols are currently limited, especially the ratio between the maximum H-wave (Hmax) amplitude and the Mmax amplitude (i.e. the EMG Hmax/Mmax ratio) [47], which is an index of the excitability of the motoneuron pool [43,47]. While accumulating data suggest that there may be different mechanisms that contributes to a potent PAP and PAPE response [6,8], data regarding the neural contribution of both PAP and PAPE is still limited [51,52].
Additionally, since there are several parameters of sEMG, such as the EMG amplitude (e.g., MAV and RMS), EMG spectral variables, (e.g., MPF and MdF), H-reflex parameters (e.g. amplitude and threshold [53]) and M-wave values (e.g. amplitude, area and duration [54]), it is unclear if some parameters of sEMG could provide better or worse insights to the neural mechanisms that could modulate the PAP/PAPE response. Having a greater understanding of how sEMG parameters relates to PAP/PAPE may help us reduce the incidence of musculoskeletal injuries in athletes and the general population, by providing us with new tools to construct better warmup guidelines. Clinically, this may also be of great importance for healthcare professionals rehabilitating patients with muscle weakness, a condition affecting millions of older adults worldwide [55]. Hence the aim of this systemic review was to identify if sEMG parameters are indicative of PAP/PAPE, in terms of twitch potentiation and voluntary performance. Since, an increase in sEMG amplitude in most instances is proportional with increasing muscle force [29] and increased MPF and MdF has been found to be indicative of higher proportion of type II fiber recruitment [56,57], two hypotheses were proposed: 1) the PAP/PAPE magnitude will be positively related to the sEMG amplitude of the working muscle group (s), and 2) short-term increases in the sEMG frequency variables of the working muscle group (s) will also be positively related to PAP/PAPE.
2. Materials and Methods
2.1. Information Sources and Search Strategy
This systematic review was carried out following the PRISMA (Preferred Reporting Items for Systematic reviews and Meta-analyses) guidelines [58]. The literature search was performed until April 2024 in three relevant electronic databases: PubMed, Web of Science and SCOPUS. The following keywords, in combination with Boolean operators (AND, OR) were used: "post activation potentiation", "post activation performance enhancement" and " electromyography". The advanced search in PubMed was used, with the following search entered in the query box: "electromyography"[All Fields] OR "EMG"[All Fields] OR "H-reflex"[All Fields] OR "M-wave*"[All Fields]) AND "post activation p*"[All Fields]. For Web of Science the advanced search query builder was used, with the following search: ((((ALL=("electromyography")) OR ALL=("EMG")) OR ALL=("H-reflex")) OR ALL=("M-wave*")) AND ALL= ("postactivation potentiation"). Lastly, the advanced search with all fields in SCOPUS was used, with the following search: {electromyography} OR {EMG} OR {H-reflex} OR {M-wave} AND {post activation potentiation}. The results were filtered by article/document type in each database (Clinical Trial in PubMed, Article in Web of Science, and Article in SCOPUS respectively). All studies were saved on Zotero (6.0.37) bibliographic reference manager [59].
2.2. Inclusion/Exclusion Criteria
The following inclusion criteria were applied: (a) the study was an original research article, and the PAP/PAPE-protocols were conducted in healthy human adults, and (b) sEMG recordings were an outcome variable of interest. The exclusion criteria were: (a) studies with no explicit PAP/PAPE protocol, (b) interventional studies with no control group or counterbalance.
2.3. Data Extraction
For all included articles, the following information were extracted: (a) study characteristics (author, year, and sample size); (b) subjects demographics (sex, age, and training status); (c) CA protocols (type of exercise and load); (d) sEMG parameters (e) twitch/performance measures (PAP and PAPE outcomes, respectively); (f) study findings.
2.4. Methodological Quality Evaluation
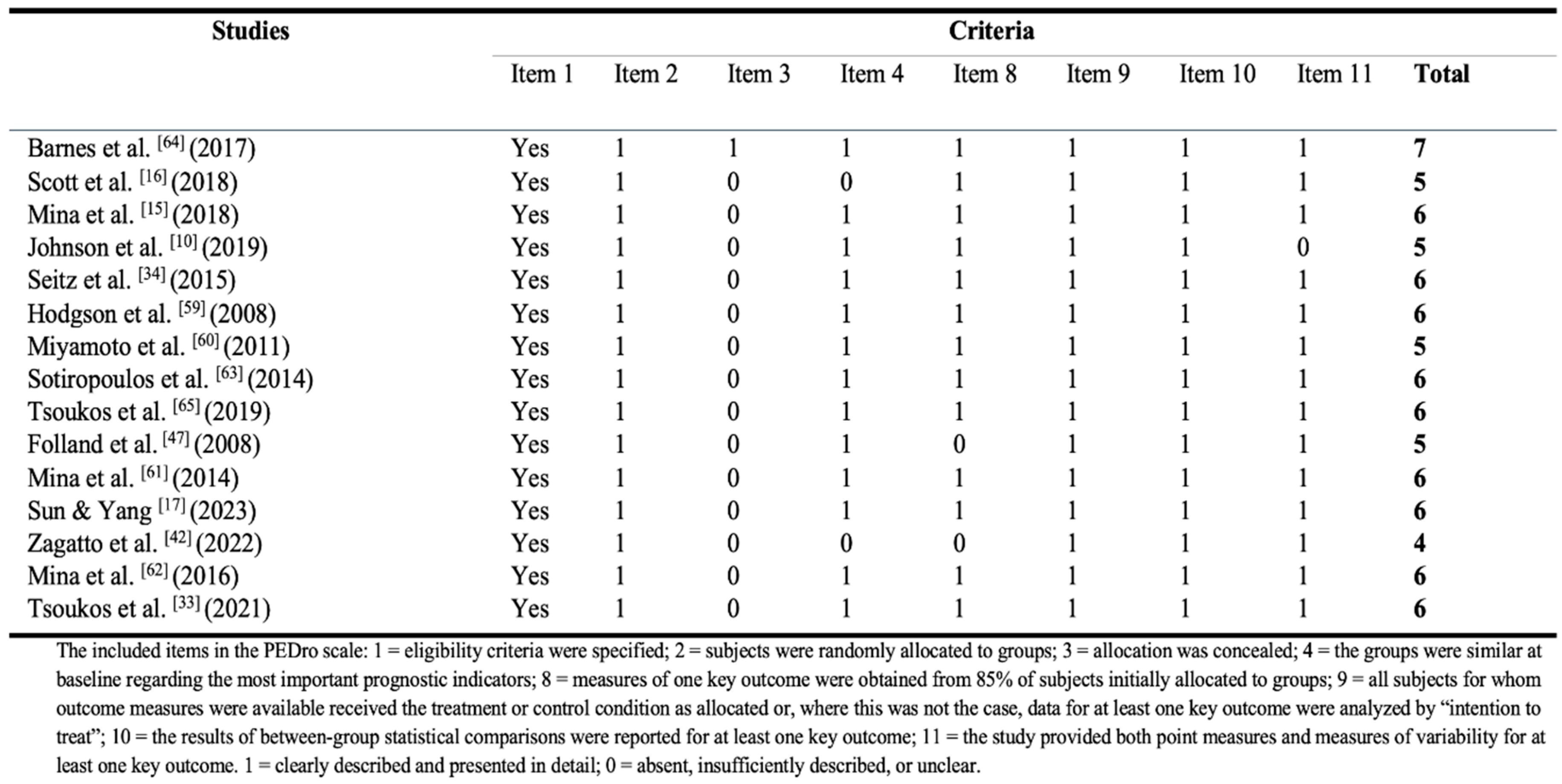
The modified Physiotherapy Evidence Database (PEDro) scale was used to evaluate the methodological quality of the included studies in the review (see Table 1), all of which were randomized control trials (RCTs). The items 5-7 were removed from the original PEDro scale, as blinding the subjects and investigators in supervised exercise interventions are not always feasible. This has been conducted in previous systematic reviews of exercise interventions [56,57]. Accordingly, this modified PEDro scale consisted of 8 items, and included eligibility criteria (item 1), randomization (item 2), concealed allocation (item 3), the groups were similar at baseline (item 4), more than 85% retention (item 8), intention-to-treat analysis (item 9), between-group comparison (item 10), and point measures and measures of variability (item 11). The highest score on this modified rating scale was 7, as the first item was not counted in the total score. The methodological quality was categorized as follows: poor quality (≤3 points), moderate quality (4 points), good quality (5 points), and excellent quality (6–7 points).
3. Results
3.1. Study Selection and Results of Literature Retrieval
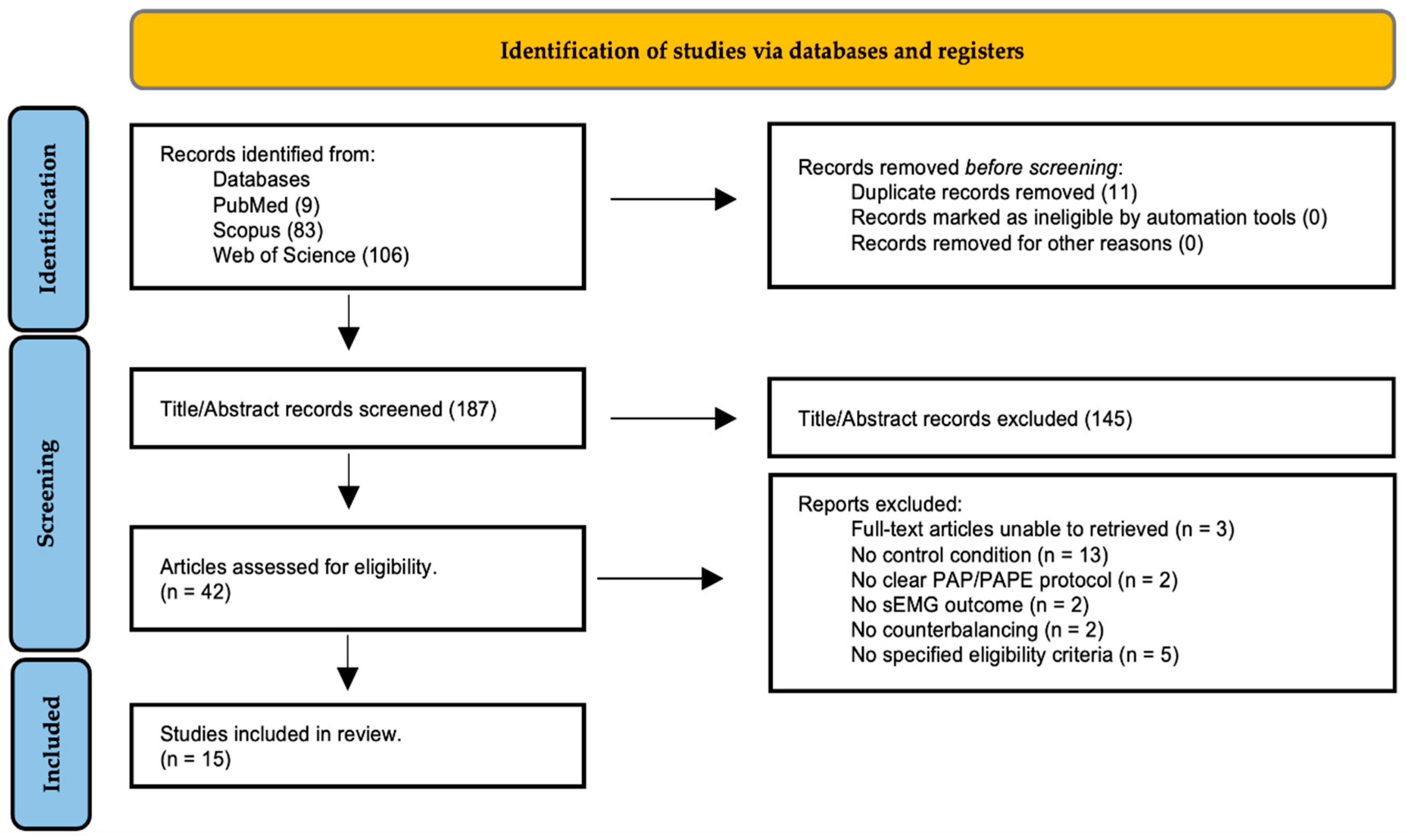
The search of PubMed, Web of Science and SCOPUS databases provided a total of 198 records. After deduplication, 187 records remained (see Figure 1) and 145 records were excluded based on their titles and abstracts. After reviewing the full text, fifteen underwent full data extraction and were included in the review. Among the included studies, twelve examined the sEMG amplitude, six the M-wave, two the MdF, two the H-reflex, and one the EMG Hmax/Mmax, ratio as sEMG parameters. The five most common twitch/performance measures to detect PAP/PAPE among the included studies was PTT, PPO, RFD and CMJ-height. The flow diagram is presented in Figure 1 and it illustrates the screening process and the main reasons for exclusion.
3.2. Characteristics of Studies Included.
All studies included in this review were RCTs and had a cross-over design [10,15,16,17,33,34,42,47,60,61,62,63,64,65,66]. 11 of 15 RCTs used separate two-way repeated-measures analyses of variances (ANOVAs) to evaluate the interaction between the time and different CA-protocols on sEMG and PAP/PAPE outcomes, respectively [10,15,16,17,33,34,42,47,61,64,65]. Additionally, three RCTs used separate multivariate analysis of variance (MANOVA) to determine the influence of different CA-protocol on several sEMG and PAP/PAPE outcomes, respectively [15,62,63], while three other RCTs used one-way repeated measures ANOVAs to compare the effect of time on different sEMG and PAP/PAPE outcomes, respectively [42,60,61]. The sample size in the studies varied from 8 [47] to 20 [16,42], with a total of 199 subjects included in this review. 14 of 15 studies used low (i.e. 30% to 40% of 1 repetition maximum [1RM] [17,66]), moderate (i.e. 60% to 70% of 1RM [65,66] or ∼70% of maximized mechanical-power output [Pmax] [64]) and high intensity (i.e. ≥80% of 1RM or ≥100% of Pmax, to isometric maximum voluntary contraction [iMVC]) resistance exercise as a CA [9,15,33,34,47,60,61,62,63,64,65]. The most common combinations of sEMG parameters and PAP/PAPE outcomes included, sEMG amplitude and PPO [15,16,17,64,65], M-wave and PTT [34,47,60,61], and sEMG amplitude and CMJ-height [15,16,17]. The remaining two studies [10,42] used a drop jump (DJ) protocol as a CA, with one study examining PTT and the M-wave [10], and another investigating repeated sprint ability (RSA) outcomes and RMS amplitude, M-wave and MdF, respectively [42]. Table 2. provides a full description of the included studies.
3.3. Quality of the Studies (Risk of Bias)
Results of the PEDro rating are presented in Table 1. The fifteen RCTs in this review had an average score of 6.07 (mean = 6.07 ± 1.27) and were therefore deemed as having ‘’excellent’’ methodological quality (on average). Still, most studies had methodological deficits concerning ‘’ concealed allocation’’ (14 of 15 studies). Moreover, five studies were discarded after assessing the quality, due to not having a specified eligibility criteria (see Figure 1), and consequently were deemed as having a high risk of bias.

Table 1.
PEDro Rating of the included studies.
3.4. Synthesis of Results
- Changes in sEMG parameters and twitch/performance outcomes between different CA protocols were examined. The evidence was summarized for conditions were sEMG parameters indicative of acute improvements in twitch/performance outcomes (PAP and PAPE respectively) in healthy athletic adults, but also incidents where they were deemed as unrelated.
Table 2.
The characteristics of the included studies.
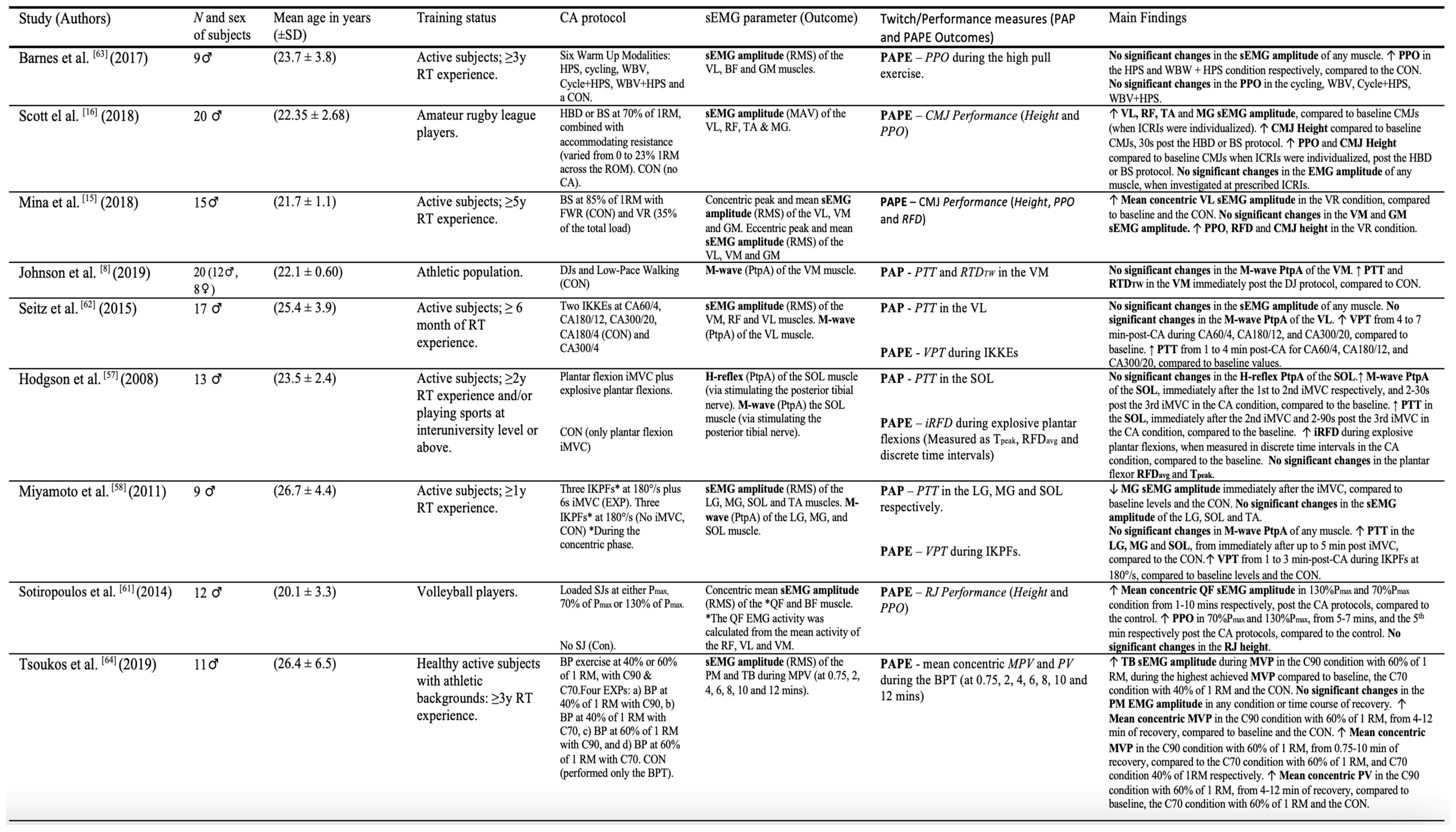
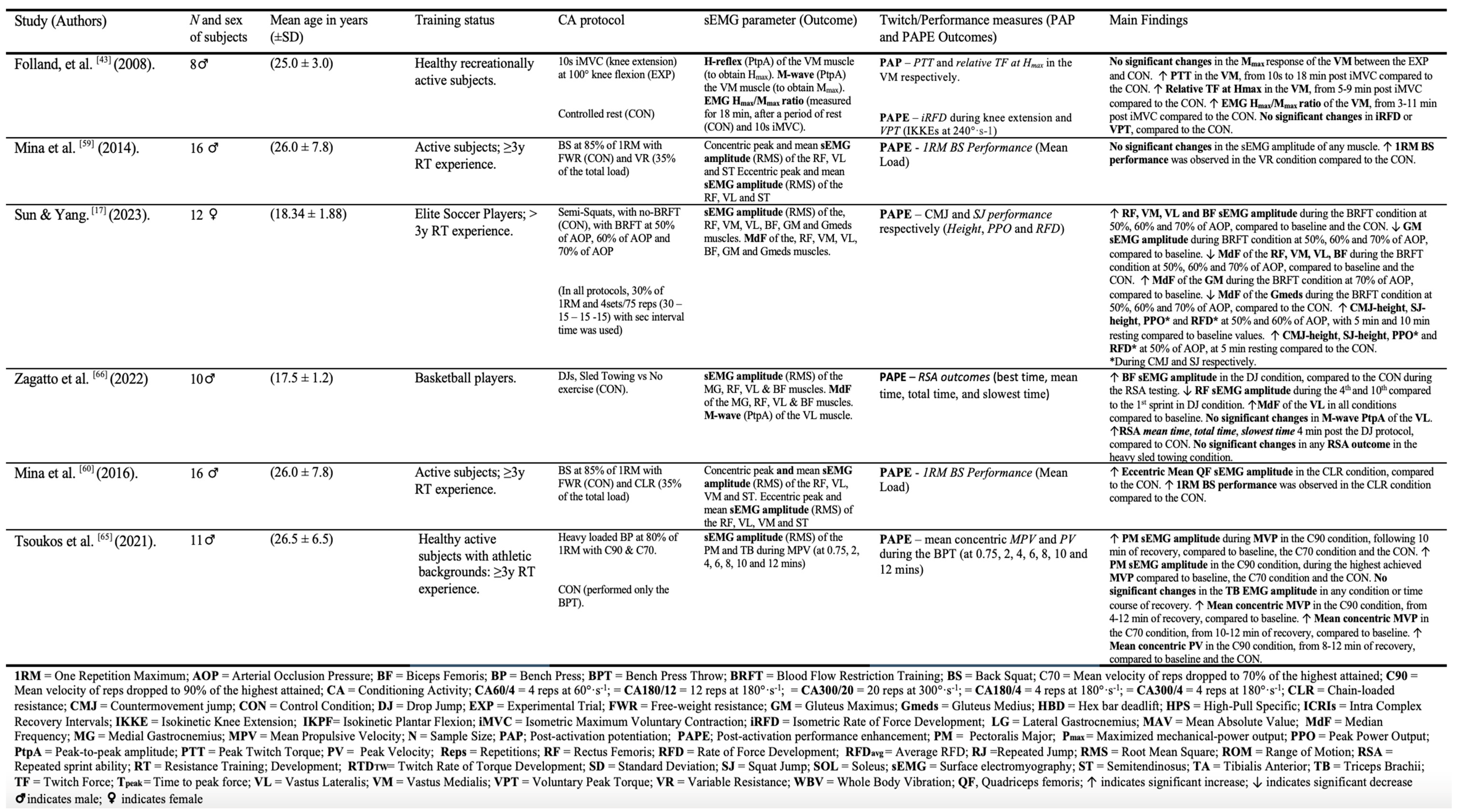
3.4.1. The sEMG Amplitude (RMS and MAV) and PAP/PAPE
Twelve studies examined the sEMG amplitude and the occurrence of PAP/PAPE [15,16,17,33,34,42,61,62,63,64,65,66]. Eight studies reported that both the sEMG amplitude (in different muscles) and certain PAP/PAPE outcomes (in particular PAPE) increased significantly [15,16,17,33,42,63,64,66] after performing either a series of DJs [42] or low to high intensity resistance exercise [15,16,17,33,63,64,66] as a CA (p < 0.05). One high quality RCT [17] reported that four sets of low intensity back squats (BS) with either 50%, 60% and 70% of arterial occlusion pressure (AOP), during blood flow restriction training (BFRT), were all effective for increasing the RMS amplitude in the vastus medialis (VM) (10.8% to 15.2%, p < 0.05), vastus lateralis (VL) (5.6% to 9.5%, p < 0.05), rectus femoris (RF) (5.9% to 8.3%, p < 0.05) and biceps femoris (BF) (9.6% to 25%, p < 0.05), compared to the control (BS with no AOP).
The highest RMS values was observed in the 70% AOP condition for all the muscles, followed by the 60% AOP condition. However, an acute increase in CMJ and SJ performance (i.e. PPO, RFD and vertical jump height respectively) was only noted in the 50% and 60% AOP group (5-10 min post the BS-BFRT protocol). Further, the greatest improvements in the CMJ-PPO (6.3%, p < 0.05), SJ-PPO (3.2%, p < 0.05), CMJ-RFD (2.4%, p < 0.05), SJ-RFD (2.3%, p < 0.05), CMJ-height (6.7%, p < 0.05) and SJ-height (6.4%, p < 0.05) was observed in the 50% AOP condition compared to the control (5 min post the BS-BFRT protocol). Interestingly, however, the gluteus maximus (GM) RMS amplitude decreased significantly (-6.6% to -4.9%, p <0.05) in all AOP groups compared to the first set (baseline), with the lowest and highest values observed in the 50% and 70% AOP group respectively. Contrarily, another high quality RCT by Mina and colleagues [15] reported a short-term increase in the VL RMS amplitude (27.5% to 33.4%, p <0.05), PPO (4.4% to 5.9%, p <0.05), RFD (12.9% to 19.1%, p <0.05) and CMJ-height (5.3% to 6.5%, p<0.05), 30sec to 12 min following a high intensity BS-protocol with variable resistance (VR), compared to pre-intervention values.
Noteworthy, the VL RMS amplitude was only significantly higher than the pre-intervention values when it was expressed as mean concentric RMS amplitude, as no significant differences was observed for peak concentric VL RMS amplitude (p >0.05) nor mean eccentric VL RMS amplitude (p >0.05). Although, interestingly, an earlier study by the same researchers [63] revealed that the mean eccentric RMS amplitude of the quadriceps femoris (QF) acutely increased (32.2%, p <0.01) after performing a chain-loaded resistance (CLR) BS-protocol (85% of 1RM, with 35% of the total load generated with chains), compared to a control (i.e. standard BS at 85% of 1RM). Of note, the same study found that the CLR-BS protocol acutely enhanced the subjects maximum strength (i.e. their 1RM load on the BS) (6.2%, p <0.05), compared to the control. However, similarly, the QF sEMG activity was not significant when it was expressed as peak concentric QF RMS amplitude (p >0.05), peak eccentric QF RMS amplitude (p >0.05), nor mean concentric QF RMS amplitude (p >0.05).
Contrariwise, nonetheless, another high quality RCT [64] found that the mean concentric QF RMS amplitude increased (7.0% to 19.3%, p <0.05), after performing a loaded SJ-CA protocol, with either moderate (i.e. 70% of Pmax) or high intensity (i.e. 130% of Pmax) compared to a control (no SJs). This higher QF activity was observed 1-10 mins post the respective CA in the 130%Pmax and 70%Pmax trial. Intriguingly, short-term improvements in PPO (during repeated jumps) were only noted 5 mins (13.9%, p <0.05) and 7 mins (8.2%, p <0.05) post the CA in the 130%Pmax and 70%Pmax condition respectively, which coincided with the highest RMS values in each respective condition, compared to the control. Contrarily, two other high quality RCTs [33,66] reported that the pectoralis major (PM) and triceps brachii (TB) RMS amplitude was only elevated when it was analyzed during the highest mean propulsive velocity (MVP) in the concentric phase of a bench press throw (BPT), although the intensity of the CA-protocol, the muscle used for EMG analysis and accumulated neuromuscular fatigue was shown to influence how they were related. Specifically, one RCT [66] investigated the effect of different bench press (BP) velocities (performed as fast as possible until the mean velocity dropped to 90% [C90] vs 70% [C70] of the fastest repetition) and different intensities (40% vs 60% of 1 RM). In this study, the authors demonstrated that the TB RMS amplitude was only significantly higher (28.6%, p <0.01) in the C90 condition (i.e. less accumulated neuromuscular fatigue) with 60% of 1 RM (i.e. moderate intensity), compared to baseline. In addition, this higher TB activity was accompanied with the greatest short-term improvements in the MVP (9.2%, p <0.01) and PV (7.5%, p <0.01) respectively, although no significant differences were observed for the PM RMS amplitude in this study (p >0.05). Contrariwise, in another study [33], the same authors observed that the PM RMS amplitude acutely increased (40% to 43.3%, p <0.01), 8-12 mins following a high intensity (i.e. 80% of 1RM) BP-C90 protocol, compared to baseline, and this coincided with the greatest acute improvements in the MVP (8.5%, p <0.01) and PV (6.0%, p <0.01) respectively. However, this time, no significant differences were observed for the TB RMS amplitude (p >0.05).
Furthermore, one RCT [16] in this review used the MAV to detect the sEMG amplitude, and in addition assessed the effect of individualized intra complex recovery intervals (ICRIs). This study found that the sEMG amplitude increased (in several lower body muscles) following either a hex bar deadlift (HBD) or BS-CA protocol respectively, with moderate intensity (i.e. 70% of 1RM) and an additional 23% accommodating resistance, compared to baseline. Specifically, an increase in the VL (20.4%, p < 0.01), BF (22.7%, p < 0.01), tibialis anterior (TA) (22.0%, p < 0.01) and GM (21.9%, p < 0.01) sEMG amplitude was noted, together with acute improvements in PPO (4.0%, p < 0.01) and CMJ-height (8.5%, p < 0.01) respectively when the ICRIs was individualized. Notably, these short-term increases in sEMG amplitude, PPO and CMJ-height disappeared when ICRIs was instead fixed and prescribed at either 30sec, 90sec or 180sec following the HBD and BS-protocol (p > 0.05).
Moreover, only one RCT by Zagatto and colleagues [42] used a DJ-CA protocol and assessed its effect on repeated sprint performance. In this study, a higher BF RMS amplitude (16.4%, p < 0.05) was reported, followed by significant improvements in RSA outcomes (mean time = -3.4%, p <0.05; slowest time = -3.8%, p <0.05; total time = -3.4%, p <0.05), compared to a control (no exercise condition). Furthermore, two moderate to good quality RCTs [42,61] in this review reported that the sEMG amplitude decreased (in different lower body muscles), while a PAP/PAPE effect was observed. Specifically, the same study by Zagatto and colleagues [42] also noted that the RF RMS amplitude decreased during the 4th (-24.9%, p =0.021) and 10th (-30.7%, p = 0.035) repeated sprint compared to the 1st sprint after the DJ protocol, while there were significant improvements in RSA outcomes.
Contrarily, in the other study [61], the medial and lateral gastrocnemius (MG and LG respectively), soleus (SOL) and TA muscle, and several performance outcomes was investigated following a 6-second plantar flexion iMVC. In this study, the MG RMS amplitude acutely decreased immediately following the iMVC (-20%, p <0.05), compared to the control (no iMVC condition), and this surprisingly coincided with the greatest short-term improvements in PTT (178.6%, p <0.05). Still, noteworthy, all twelve studies demonstrated that the PAP/PAPE outcomes increased (p <0.05), while there were no significant changes in the sEMG amplitude in some muscles (p > 0.05) [15,16,17,33,34,42,61,62,63,64,65,66]. Importantly, however, several studies used different normalizations procedures for the EMG signal. Still, the muscle of interest was most commonly normalized to the RMS values obtained during a 1 RM attempt [33,66] or squat isometric lifts [15,63] in this review.
3.4.2. The MdF and PAP/PAPE
Two studies examined the MdF and the occurrence of PAP/PAPE (specifically PAPE outcomes) [17,42]. One RCT [42] demonstrated that the MdF increased in the VL after a DJ protocol (9.3%, p <0.05), but no significant differences was observed compared to a sled towing trial or the control (no exercise) (p >0.05). Still, an improvement in RSA outcomes was noted in the DJ condition, compared to the control (p <0.05). Further, in another RCT [17], it was reported that four sets of BS with either 50%, 60% and 70% AOP reduced the Mdf of the VM (-17.4% to -10.0%, p <0.05), VL (-9.2% to -14.3%, p <0.05), RF (-11.9% to -4.0%, p <0.05), BF (-18.9% to -9.2%, p <0.05) and Gmed (-5.8% to -4.2%, p <0.05) respectively, compared to the control. The highest drop in MdF was observed in the 70% AOP condition in most muscles, followed by the 60% AOP condition. Still, short-term performance improvements were only observed in the 50% and 60% AOP conditions respectively, with the greatest effects observed in the 50% AOP condition compared to the control (5 min post the BS-BFRT protocol). Additionally, the MdF of the GM only increased (7.8%, p < 0.05) in the 70% AOP condition. Moreover, all studies in this review [17,42] demonstrated that various PAPE outcomes significantly increased (p<0.05), while there was no changes in the MdF in other muscles (p >0.05).
3.4.3. The M-Wave and PAP/PAPE
Six studies investigated the occurrence of PAP/PAPE and changes in the evoked M-wave response (peak to peak amplitude [PtpA]) obtained from either the VM [10,47], VL [34,42], soleus (SOL) [60], MG [61] or LG [61]. Five studies used motor nerve stimulation to obtain the M-wave [34,42,47,60,61], while one study used direct muscle stimulation (from the VL) in this review [10]. Nevertheless, all studies found that the M-wave PtpA remained unchanged, while different PAP/PAPE outcomes increased at certain time points (p < 0.05). Specifically, five RCTs [10,34,47,60,61] found that the M-wave remained constant while the PTT increased after performing a CA-protocol with either isokinetic knee extensions (2.6% to 4.9%, p <0.05) [34], plantar flexion iMVCs [60,61] (13.8% to 178.6%, p <0.05), DJs [10] (15% to 23%, p <0.001) or knee extension iMVC (6.7% to 66.6%, p <0.05) [47], compared to either baseline values or a control group. This PAP response occurred immediately (2-10sec) after the CA [10,47,60,61] and remained significant for up to 30sec to 18 mins [10,47,60,61] depending on the CA-protocol.
Further, one study reported a significantly higher RTDTW (32%, p <0.001), with no changes in the M-wave (p >0.05), immediately after a DJ-CA protocol [10], compared to a control (no DJs). In addition, three RCTs noted short-term improvements in either RSA performance outcomes (p <0.05) [42], or voluntary peak torque (2.1% to 6.1%, p <0.05) [34,61] compared to baseline, following a CA-protocol with DJs [42], plantar flexion iMVCs [61] or isokinetic knee extensions [34]. However, one RCT [60] observed a short-term increase in the M-wave PtpA of the SOL, immediately after three series of 5sec plantar flexion iMVCs (and explosive plantar flexions) compared to baseline, with increases at the 1st iMVC (8.7%, p < 0.05), 2nd iMVC (10.6%, p < 0.05) and 2-30sec post the last iMVC (5.5% to 12.2%, p < 0.05), although it rapidly dropped to non-significant values at 90sec post the last iMVC (p >0.05). Noteworthy, the PTT in the SOL was significantly higher at the 2nd iMVC (15.2%, p < 0.05) and 2-90sec post the last iMVC (13.8% to 18.8%, p < 0.05), but the values became non-significant at 150sec post the last iMVC (p >0.05). Interestingly, a short-term improvement in plantar flexor isometric RFD (iRFD) was also noted when the RFD profile was analyzed in discrete units of time (25.0% to 31.6%, p < 0.05) at 15-60sec post the CAs, although no significant changes was observed when the iRFD was expressed as time to peak force (Tpeak) or average RFD (RFDavg) (p > 0.05).
3.4.4. The H-Reflex and PAP/PAPE
Two studies evaluated the presence the PAP/PAPE and changes in the evoked H-reflex response (PtpA) obtained from either the VM [47] or the SOL [60]. Both studies used motor nerve stimulation to obtain the H-reflex and found that it remained unchanged (p >0.05), while different PAP/PAPE outcomes increased at certain time points (p<0.05). In particular, Hodgson and colleagues [60] observed a significant increase in the PTT of the SOL (13.8-18.8%, p <0.05) compared to baseline (2-90sec post iMVC), while the H-reflex remained constant. In the other study, Folland and colleagues [47] observed a significantly higher maximal twitch force (i.e., PTT) of the VM (66.6%, p <0.01) compared to a control condition (10sec post iMVC), while no significant changes occurred in any H-reflex parameters (p > 0.05).
3.4.5. The EMG Hmax/Mmax Ratio and PAP/PAPE
One RCT investigated how the EMG Hmax/Mmax ratio was related to the PAP/PAPE response [47]. This study found that the PTT acutely increased in the VM (6.7% to 66.6%, p < 0.05) compared to the control (10sec to 18 min post iMVC), followed by a greater EMG Hmax/Mmax ratio (23.2% to 42%, p < 0.01), 5-11 min post the iMVC. In addition, when the twitch force (evoked by Hmax stimulation) was expressed as a percentage of PTT, the twitch force was significantly higher (70.3% to 74.2%, p < 0.05) than the control (5-9 mins post the iMVC). Noteworthy, the greatest EMG Hmax/Mmax value (42%, p < 0.01) and the highest relative twitch force percentage (74.2%, p < 0.01) was both observed 5 min post the iMVC, although the highest mean value for the PTT occurred 10sec post iMVC. Still, this PAP response and the corresponding higher EMG Hmax/Mmax ratio did not coincide with any acute improvements in iRFD or voluntary peak torque outcomes (p>0.05). The Hmax and Mmax was obtained via motor nerve stimulation in this study.
4. Discussion
To our knowledge, this is the first systemic review that have evaluated if sEMG parameters are indicative of PAP/ PAPE, in terms of twitch potentiation and voluntary performance. The two hypotheses were the following: 1) the PAP/PAPE magnitude will be positively related to the sEMG amplitude of the working muscle group (s), and 2) short-term increases in the sEMG frequency variables of the working muscle group (s) will also be positively related to PAP/PAPE. This review revealed that increases in sEMG amplitude (RMS and MAV) may only be indicative of PAP/PAPE in some muscles, in particular acute increases in voluntary performance (i.e. PAPE), and this relation seems to be influenced by several factors, including 1) the velocity and intensity of the CA, 2) how the EMG data was normalized and recorded, 3) the time point in which the EMG response was analyzed and 4) accumulated neuromuscular fatigue. Most studies in this review that reported a simultaneous increase in sEMG amplitude (in some muscles) and short-term improvements in voluntary muscular performance, executed the CAs at maximal or near-maximal velocities, combined with moderate (60% of 1RM or at ∼70% of Pmax) [64,66] to high intensities (≥ 80% of 1 RM or ∼130% of Pmax, including DJs [15,16,33,42,63,64]). The higher sEMG amplitude (and the corresponding performance enhancement) occurred on average between 5-10 min post the CA [15,17,33,66], although one RCT [16] reported that individualized ICRIs may be needed to elicit any potentiating effect.
Nevertheless, many studies used different normalizations procedures for the EMG signal in this review, although the muscles of interest were most commonly normalized to the RMS values obtained during a 1 RM attempt [33,66] or squat isometric lifts [15,63]. Further, all the performance outcomes that subsequently was evaluated were ballistic in nature, and predominately performed concentrically, including PPO [15,16,64], RFD [15], CMJ height [15,16], MVP [33,66]. This suggest that assessing the sEMG amplitude may have some utility as an indicative measure of PAPE (at least in some muscles) during ballistic movements, focusing on maximum power development, while it generally may be a poor indicator of PAP outcomes (i.e. observing an acute increased in PTT or RTDTW), as one RCT in this review actually found that an acute increase in PTT coincided with a decreased MG RMS amplitude, and no myoelectrical changes in other calf muscles [61].
These findings support previous experimental studies inferring that the muscle potentiation effect is more prominent during peak muscle shortening speeds and high-speed concentric contractions, compared to isometric muscular activity [67,68]. However, it contradicts the importance of increased expression of MRLC phosphorylation (i.e. the primary mechanism of PAP) for voluntary performance enhancement at high velocities, and instead signify that other mechanisms, reflected by higher sEMG amplitude, may play a larger contributing role, at least when assessed during ballistic movements. Importantly, however, as it is well documented that the sEMG amplitude increases with higher muscle force but also with accumulated neuromuscular fatigue [29,31], it may be misleading to solely rely on acute increases in sEMG amplitude as an indicative measure of PAPE, without any direct performance assessment and/or evaluating other sEMG parameters.
Further, the second hypothesis in this review was that short-term increases in the sEMG frequency variables of the working muscle group (s) would be positively related to PAP/PAPE. This review revealed that an acute decrease in the MdF of the working muscles may (at least in some circumstances) be positively related to a PAP/PAPE response, when assessed in voluntary conditions (i.e. PAPE). While one good quality RCT in this review reported that the MdF increased in the VL muscle following a DJ-CA protocol [42], these Mdf changes was not distinguishable from a control (a standard warm up) and occurred independent of any voluntary performance improvements. This may partially contradict the notion that increases in MdF is indicative of a higher proportion of type II fiber recruitment [56,57] or simply signify that PAPE is unrelated to an acute increased recruitment of type II fibers (at least in some circumstances). Alternatively, it implies that some other mechanism related to an acute decrease in the MdF may be indicative of PAPE.
Typically, EMG spectral variables (i.e. MdF and MPF) tend to decrease over time during fatiguing muscular contractions [29], and this has been attributed to a decline in muscle fiber velocity [32,69], via a reduction in intracellular pH [70,71]. Importantly, however, this reduction in MdF have repeatedly been found to coincide with an increased sEMG amplitude during fatiguing muscular contractions, as this has been proposed to reflect increased motor unit recruitment to maintain a constant muscle force [29,31]. Further, as muscle potentiation and fatigue normally coexist when performing any muscular activity, this may imply that a smaller drop in MdF (relative to the rise in sEMG amplitude) may be indicative if an active muscle will be in a net potentiated or unpotentiated state. While a limitation with this review is that only two studies examined the occurrence of PAP/PAPE and MdF, this in part supports the findings from a good quality RCT in this review.
In this study, Sun and Yang [17] demonstrated that the MdF of several lower body muscles (incl. VM, VL, RF and BF) acutely decreased, while the RMS amplitude of the same muscles increased, after a BS-CA protocol combined with low intensity (i.e. 30% of 1RM) and BFRT at either 50%, 60% or 70% AOP respectively, compared to a control. However, short-term improvements in CMJ and SJ performance (i.e. PAPE) were only observed in the 50% and 60% AOP condition, respectively. Further, the greatest performance improvements and lowest absolute drop in MdF (and rise in the RMS amplitude) of the VM, VL, RF and BF, occurred in the 50% AOP BFRT. However, intriguingly, the MdF in the 50% AOP BFRT, was only lower (relative to the rise in the RMS amplitude) in the VM and RF muscle respectively, compared to the 70% AOP BFRT and the control, while the MdF of the VL and BF was in contrast lower (relative to the RMS amplitude level) in the 70% AOP condition, compared to the 50% AOP and the control (see subsection 3.4.1. and 3.4.2.). Additionally, the GM RMS amplitude decreased in all AOP groups compared to baseline, with the lowest and highest decline observed in the 50% and 70% AOP group respectively. However, the MdF of the GM only increased in the 70% AOP condition.
This implies that a smaller drop in MdF (relative to the rise in the sEMG amplitude) of a muscle/muscle group may (or may not) be indicative of a PAPE response, as it seems to be highly dependent on the muscle (at least with BFRT). Based on previous research, this may be related to differences in muscle fiber type composition [72], especially the distribution of type II fibers in each muscle [73,74] and/or simply signify that the muscle activation pattern was more favorable in the 50% AOP condition for plyometric performance. For instance, in a kinematic and electromyographic study by Bobbert & van Ingen Schenau [75] it was reported that the RF and VM sEMG amplitude increased during the initial part of the push-off phase of a vertical jump in skilled jumpers, and this coincided with higher net moments around the knees. Concurrently, the BF sEMG amplitude also decreased during the push-off phase, which was explained by lower net moments around the hip joint. This suggest that an acute increase in the sEMG amplitude (relative to the drop in the MdF) in the RF and VM muscle, in addition to a lower BF sEMG amplitude, may only be indicative of PAPE for vertical jumping, while different muscle activation pattern may be more favorable for other movements.
Further, while it is less certain how a reduction in the RMS amplitude and an increased MdF relates to fatigue within the neuromuscular system, a large decline in the MdF has consistently been shown to reflect neuromuscular fatigue in EMG research [29,31], which supports the findings in this review. Moreover, six studies included in this review [10,34,42,47,60,61] also investigated PAP/PAPE and changes in the evoked M-wave response (PtpA) obtained from either the VM, VL, SOL, MG and LG muscle. Intriguingly, all studies found that the evoked M-wave tended to remain the same, while significant improvements was observed in different PAP and PAPE outcomes. Noteworthy, the type of CA that was utilized, the muscle that was used for the M-wave assessment and the performance outcome that was used for pre- and post-analysis, did not significantly influence this relation. Intriguingly, however, one RCT [60] in this review observed a transient improvement in the evoked M-wave of the SOL, following three series of 5sec plantar flexion iMVCs. This M-wave enlargement or potentiation was noted immediately after the 1st and 2nd iMVC respectively, and 2-30sec post the last iMVC. Although, it rapidly dropped to non-significant values at 90sec post the last iMVC. Similarly, the same study found that the PTT in the SOL was significantly higher at the 2nd iMVC and 2-90sec post the last iMVC, but the values became non-significant at 150sec post the last iMVC. This infers that the action potentials that travels along the t-tubules to the muscle [10] may partially contribute to the initial development of PAP, suggesting that a transient raise in the evoked M-wave PtpA could be indicative of PAP. This short-lived M-wave potentiation (≤ 1 min) have been noted in previous studies [7,47] and has been attributed to mechanism in the fiber membrane’s Na+-k+ active transport [7,76], but have also simply been suggested to be an artifact via subtle movements of the electrodes [47]. Regardless, collectively, the M-wave PtpA may generally be indicative of PAP (assuming it is combined with a twitch torque outcome), as it has consistently been reported to be unaffected by various CA-protocols, and it has repeatedly been used to control for changes in neuromuscular propagation that could influence the potentiated twitch response (i.e. PAP) [10,39,40].
In contrast, whereas PAP tend to be highest immediately post a iMVC-CA protocol, and drop exponentially over time and sequentially disappear at 10-18 min post the CA [10,47,60,61], PAPE has generally been found to peak 5-10 min post the CA in most studies [6,15,17,33,66]. Therefore, any enhancement that the mechanisms related to the M-wave enlargement would temporally have on voluntary muscular performance would most likely be impaired by residual neuromuscular fatigue. This premise is supported by an experimental study by Hicks and colleagues [77], were the researchers noted that the M-waves values (area and PtpA) increased gradually during the first 2 min of the fatiguing protocol, and this coincided with a reduction in voluntary force. This suggest that assessment of the M-wave PtpA may mainly be applicable for PAP research (in addition to studying fatigue within the neuromuscular system), and that changes in the M-wave PtpA may generally not reflect PAPE. Still, one RCT included in this review actually reported that this M-wave enlargement (in the SOL) coincided with short-term increases in plantar flexor iRFD (i.e. PAPE), when it was analyzed in discrete units of time, compared to baseline (15-60sec post a iMVC-CA) [60]. This suggest that changes in neuromuscular propagation (e.g. sarcolemmal membrane excitability) may nevertheless partially contribute to the PAPE response, at least during brief explosive contractions. However, the same study [60] found that there were no significant differences in the iRFD when it was expressed as Tpeak and RFDavg. Thus, how the M-wave PtpA relates to PAPE still needs to be explored, as the PAPE response seems to be strongly influenced by how the PAPE outcome is expressed and analyzed. In addition, as no studies in this review assessed the area and duration of the M-wave, and most studies included in this review only stimulated the motor nerve to obtain the M-wave [34,42,47,60,61], how these M-wave values and direct muscle stimulation relates to PAPE (and PAP) also remains to be elucidated.
Furthermore, two RCTs included in this review also investigated how PAP and PAPE was related to the evoked H-reflex response (PtpA) obtained from either the VM [47] or SOL [60] muscle. Both studies reported that the H-reflex remained the same, while significant improvements was observed in different PAP and PAPE outcomes. This implies that PAP and PAPE may both be unrelated to spinal excitability, signifying that changes in the H-reflex response may generally not be indicative of PAP nor PAPE. These findings are supported by a more recent study from Iglesias-Soler and coworkers [53]. In this study, the authors observed a short-term improvement in voluntary explosive plantar flexion force (i.e. PAPE) after an iMVC-CA protocol, but no observed differences in any H-reflex parameters (incl. amplitude and threshold). Based on previous work, this may be related to the fact that the Hmax is elicited by submaximal nerve stimulation [43], and this has been found to primarily activate slow-twitch motor units [43,47]. In this regard, as MRLC phosphorylation has been observed to have less of an effect on slow-twitch skeletal muscles [6] and power-trained athletes tend to have lower amplitude of the Hmax potential compared to their endurance-trained counterparts [43], it would further suggest that changes in the H-reflex response may be unrelated to PAP, but also should have minimal or no effect on acute increases in voluntary force production (i.e., PAPE).
Lastly, one good quality RCT [47] included in this review also examined how PAP and PAPE was related to the EMG Hmax/Mmax ratio. In this study Folland and co-workers [47] found that the maximal twitch force (i.e., PTT) was significantly greater 10sec to 18 mins after the 10sec iMVC (i.e. a PAP response), compared to a control, and this was accompanied by a greater EMG Hmax/Mmax ratio (5-11 min post iMVC). Additionally, the same study found that when twitch force at Hmax was expressed as a percentage of PTT, the twitch force remained significantly higher than the control (5-9 mins post the iMVC). Noteworthy, the greatest EMG Hmax/Mmax value and the highest relative twitch force percentage was observed 5 min post the iMVC, although the highest mean value for the PTT occurred 10sec post iMVC.
This implies that the excitability of the motoneuron pool may temporally contribute to the evoked PAP response during twitch force assessment, signifying that the time point when the twitch response is analyzed impacts how twitch potentiation (i.e. PAP) relates to neurophysiological mechanisms. Based on the findings in this review, an increase in EMG Hmax/Mmax ratio may generally be indicative of a PAP response. However, the same study by Folland and colleagues [47] also found that a greater EMG Hmax/Mmax ratio was not accompanied with short-term improvements in iRFD or voluntary peak torque, indicating that the EMG Hmax/Mmax ratio may generally be a poor indicator of PAPE and thus have limited sports applications, at least for strength-power athletes.
This supports experimental findings reporting that EMG Hmax/Mmax ratios tend to be lower in athletes performing anaerobic compared to aerobic sports [43]. Intriguingly, power-trained athletes have also been found to have lower EMG Hmax/Mmax ratios than sedentary subjects [78], which provides further evidence that the EMG Hmax/Mmax ratio may generally be a poor indicator of PAPE. This highlights the inherent complexity with PAP/PAPE research. Still, there is a need for more high quality RCTs and control trials that explores how various CAs, sEMG parameters and twitch/performance outcomes interact, for us to construct better warmup guidelines to maximize muscular performance and minimize the incidence of musculoskeletal injuries. However, as the PAP/PAPE response has mainly been evaluated during dynamic movements, especially PAPE [6], it is currently unclear if CAs can also be utilized to induce short-term improvements in balance and stability of muscles, and if this can be measured effectively with sEMG. Further, in regard to injury prevention and musculoskeletal rehabilitation, this could have both sports and clinical applications.
4.1. Limitations and Recommendations for Future Research
The are several methodological limitations with the present study. This includes the small sample size in this review, and the lack of good quality RCTs on this topic. Additionally, it was found that several studies used different normalization procedure for the EMG signal and the subjects included in this review were restricted to healthy athletic populations. This, combined with the limited number of studies, may limit the generalizability of the findings. Hence, there is undoubtedly a need for higher quality studies on this topic. Future studies should for instance use more standardized normalization protocols but also examine how several different sEMG parameters are related to various CA protocols, to further improve our understanding of the underlying neurophysiology of PAP/PAPE. Moreover, how sEMG parameters are influenced by different muscle contraction mode (i.e. isometrics, concentrics and eccentrics) and how this relates to the PAP/PAPE response should also be explored. Lastly, how the M-wave, obtained via direct muscle stimulation, and different M-values (e.g. area and duration) relates to PAP/PAPE also remains to be elucidated.
5. Conclusions
This reviewed aimed to identify if sEMG parameters are indicative of post-activation potentiation PAP/PAPE, in terms of twitch potentiation and voluntary performance. The findings in this review showed that the M-wave amplitude (combined with a twitch torque outcome) may generally be indicative of PAP. The sEMG amplitude (in some muscles) were found to be indicative of PAPE during ballistic, high-power output movements, while a small decrease in the MdF (in certain muscles) were shown to reflect a PAPE response, although it generally seems to be a better tool for controlling excessive neuromuscular fatigue. On the contrary, changes in the EMG Hmax/Mmax ratio were found to contribute (temporally) to the evoked PAP response, while the H-reflex amplitude was shown to be neither indicative of PAP nor PAPE. This review provides preliminarily findings suggesting that certain sEMG parameters could be indicative of PAP/PAPE. However, due to the limited number of good to high quality RCTs on this topic, further research is warranted to re-examine how different sEMG parameters are related to the PAP/PAPE phenomenon. Having a greater understanding of how sEMG parameters relates to PAP/PAPE may help us reduce the incidence of musculoskeletal injuries in athletes and the general population, by providing us with new tools to construct better warmup guidelines. Clinically, this may also be of great importance for healthcare professionals rehabilitating patients with muscle weakness, a condition affecting millions of older adults worldwide. This review therefore highlights the potential value of understanding which parameters of sEMG can be used to detect PAP/PAPE.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, P.G. and P.V.T.; methodology, P.G., G.G., G.K.S., and P.V.T.; formal analysis; P.G., G.G., G.K.S., and P.V.T.; writing – original draft preparation, P.G.; writing—review and editing, P.G., G.G., G.K.S., and P.V.T.; visualization, P.G. and P.V.T.; supervision, P.V.T.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this review.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Safran, M.R.; Garrett, W.E.; Seaber, A.V.; Glisson, R.R.; Ribbeck, B.M. The role of warmup in muscular injury prevention. Am. J. Sports Med. 1988, 16, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Noonan, T.J.; Best, T.M.; Seaber, A.V.; Garrett, W.E. Thermal effects on skeletal muscle tensile behavior. Am. J. Sports Med. 1993, 21, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Shellock, F.G.; Prentice, W.E. Warming-Up and Stretching for Improved Physical Performance and Prevention of Sports-Related Injuries. Sports Med. 1985, 2, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Chang, C.-K.; Tseng, W.-C.; Chiu, C.-H.; Dai, X.; Ye, X. Acute Effects of Different Warm-up Protocols on Sports Performance in Elite Male Collegiate Handball Players. J. Strength Cond. Res. 2022, 36, 2262–2267. [Google Scholar] [CrossRef] [PubMed]
- McGowan, C.J.; Pyne, D.B.; Thompson, K.G.; Rattray, B. Warm-Up Strategies for Sport and Exercise: Mechanisms and Applications. Sports Med. Auckl. NZ 2015, 45, 1523–1546. [Google Scholar] [CrossRef] [PubMed]
- Blazevich, A.J.; Babault, N. Post-activation Potentiation Versus Post-activation Performance Enhancement in Humans: Historical Perspective, Underlying Mechanisms, and Current Issues. Front. Physiol. 2019, 10, 1359. [Google Scholar] [CrossRef]
- Hamada, T.; Sale, D.G.; MacDougall, J.D.; Tarnopolsky, M.A. Postactivation potentiation, fiber type, and twitch contraction time in human knee extensor muscles. J. Appl. Physiol. 2000, 88, 2131–2137. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H.B.; MacIntosh, B.R.; Dal Pupo, J. Does postactivation potentiation (PAP) increase voluntary performance? Appl. Physiol. Nutr. Metab. Physiol. Appl. Nutr. Metab. 2020, 45, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Gago, P.; Zoellner, A.; Cézar Lima da Silva, J.; Ekblom, M.M. Post Activation Potentiation and Concentric Contraction Performance: Effects on Rate of Torque Development, Neuromuscular Efficiency, and Tensile Properties. J. Strength Cond. Res. 2020, 34, 1600–1608. [Google Scholar] [CrossRef]
- Johnson, M.; Baudin, P.; Ley, A.L.; Collins, D.F. A Warm-Up Routine That Incorporates a Plyometric Protocol Potentiates the Force-Generating Capacity of the Quadriceps Muscles. J. Strength Cond. Res. 2019, 33, 380–389. [Google Scholar] [CrossRef]
- Zhi, G.; Ryder, J.W.; Huang, J.; Ding, P.; Chen, Y.; Zhao, Y.; Kamm, K.E.; Stull, J.T. Myosin light chain kinase and myosin phosphorylation effect frequency-dependent potentiation of skeletal muscle contraction. Proc. Natl. Acad. Sci. USA 2005, 102, 17519–17524. [Google Scholar] [CrossRef]
- Stull, J.T.; Kamm, K.E.; Vandenboom, R. Myosin light chain kinase and the role of myosin light chain phosphorylation in skeletal muscle. Arch. Biochem. Biophys. 2011, 510, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.; Dantzig, J.A.; Hollingworth, S.; Baylor, S.M.; Goldman, Y.E.; Mitchison, T.J.; Straight, A.F. A small-molecule inhibitor of skeletal muscle myosin II. Nat. Cell Biol. 2002, 4, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Maloney, S.J.; Turner, A.N.; Fletcher, I.M. Ballistic Exercise as a Pre-Activation Stimulus: A Review of the Literature and Practical Applications. Sports Med. 2014, 44, 1347–1359. [Google Scholar] [CrossRef] [PubMed]
- Mina, M.A.; Blazevich, A.J.; Tsatalas, T.; Giakas, G.; Seitz, L.B.; Kay, A.D. Variable, but not free-weight, resistance back squat exercise potentiates jump performance following a comprehensive task-specific warm-up. Scand. J. Med. Sci. Sports 2018, sms.13341. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.J.; Ditroilo, M.; Marshall, P. Effect of Accommodating Resistance on the Postactivation Potentiation Response in Rugby League Players. J. Strength Cond. Res. 2018, 32, 2510–2520. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Yang, T. Semi-Squat Exercises with Varying Levels of Arterial Occlusion Pressure during Blood Flow Restriction Training Induce a Post-Activation Performance Enhancement and Improve Vertical Height Jump in Female Football Players. J. Sports Sci. Med. 2023, 212–225. [Google Scholar] [CrossRef]
- Wyland, T.P.; Van Dorin, J.D.; Reyes, G.F.C. Postactivation Potentation Effects From Accommodating Resistance Combined With Heavy Back Squats on Short Sprint Performance. J. Strength Cond. Res. 2015, 29, 3115–3123. [Google Scholar] [CrossRef]
- Andersen, E.; Lockie, R.G.; Dawes, J.J. Relationship of Absolute and Relative Lower-Body Strength to Predictors of Athletic Performance in Collegiate Women Soccer Players. Sports (Basel) 2018, 6, 106. [Google Scholar] [CrossRef]
- Gossen, E.R.; Sale, D.G. Effect of postactivation potentiation on dynamic knee extension performance. Eur. J. Appl. Physiol. 2000, 83, 524–530. [Google Scholar] [CrossRef]
- Pearson, S.J.; Hussain, S.R. Lack of association between postactivation potentiation and subsequent jump performance. Eur. J. Sport Sci. 2014, 14, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, V.M.; DeLuca, J. Central fatigue: issues related to cognition, mood and behavior, and psychiatric diagnoses. PM R 2010, 2, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, A.; Behan, P.O. Fatigue and basal ganglia. J. Neurol. Sci. 2000, 179, 34–42. [Google Scholar] [CrossRef]
- Taylor, J.L.; Amann, M.; Duchateau, J.; Meeusen, R.; Rice, C.L. Neural Contributions to Muscle Fatigue: From the Brain to the Muscle and Back Again. Med. Sci. Sports Exerc. 2016, 48, 2294–2306. [Google Scholar] [CrossRef] [PubMed]
- Schillings, M.L.; Hoefsloot, W.; Stegeman, D.F.; Zwarts, M.J. Relative contributions of central and peripheral factors to fatigue during a maximal sustained effort. Eur. J. Appl. Physiol. 2003, 90, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Neyroud, D.; Cheng, A.J.; Bourdillon, N.; Kayser, B.; Place, N.; Westerblad, H. Muscle Fatigue Affects the Interpolated Twitch Technique When Assessed Using Electrically-Induced Contractions in Human and Rat Muscles. Front. Physiol. 2016, 7, 252. [Google Scholar] [CrossRef] [PubMed]
- Gandevia, S.C. Spinal and supraspinal factors in human muscle fatigue. Physiol. Rev. 2001, 81, 1725–1789. [Google Scholar] [CrossRef]
- Zhang, Q.; Iyer, A.; Lambeth, K.; Kim, K.; Sharma, N. Ultrasound Echogenicity as an Indicator of Muscle Fatigue during Functional Electrical Stimulation. Sensors 2022, 22, 335. [Google Scholar] [CrossRef]
- Guilherme H., Elcadi; Forsman, M.; Crenshaw, A.G. The relationship between oxygenation and myoelectric activity in the forearm and shoulder muscles of males and females. Eur. J. Appl. Physiol. 2011, 111, 647–658. [Google Scholar] [CrossRef]
- Toro, S.F.D.; Santos-Cuadros, S.; Olmeda, E.; Álvarez-Caldas, C.; Díaz, V.; San Román, J.L. Is the Use of a Low-Cost sEMG Sensor Valid to Measure Muscle Fatigue? Sensors 2019, 19, 3204. [Google Scholar] [CrossRef]
- Scano, A.; Pirovano, I.; Manunza, M.E.; Spinelli, L.; Contini, D.; Torricelli, A.; Re, R. Sustained fatigue assessment during isometric exercises with time-domain near infrared spectroscopy and surface electromyography signals. Biomed. Opt. Express 2020, 11, 7357–7375. [Google Scholar] [CrossRef] [PubMed]
- Ament, W.; Bonga, G.J.; Hof, A.L.; Verkerke, G.J. EMG median power frequency in an exhausting exercise. J. Electromyogr. Kinesiol. Off. J. Int. Soc. Electrophysiol. Kinesiol. 1993, 3, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Tsoukos, A.; Brown, L.E.; Terzis, G.; Veligekas, P.; Bogdanis, G.C. Potentiation of Bench Press Throw Performance Using a Heavy Load and Velocity-Based Repetition Control. J. Strength Cond. Res. 2021, 35, S72–S79. [Google Scholar] [CrossRef] [PubMed]
- Seitz, L.B.; Trajano, G.S.; Dal Maso, F.; Haff, G.G.; Blazevich, A.J. Postactivation potentiation during voluntary contractions after continued knee extensor task-specific practice. Appl. Physiol. Nutr. Metab. Physiol. Appl. Nutr. Metab. 2015, 40, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Johnson, P.W. Fatigue development in the finger flexor muscle differs between keyboard and mouse use. Eur. J. Appl. Physiol. 2014, 114, 2469–2482. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.Z.F.; Fry, A.C.; Weiss, L.W.; Schilling, B.K.; Brown, L.E.; Smith, S.L. Postactivation Potentiation Response in Athletic and Recreationally Trained Individuals. J. Strength Cond. Res. 2003, 17, 671. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.M.; Duncan, N.M.; Marin, P.J.; Brown, L.E.; Loenneke, J.P.; Wilson, S.M.C.; Jo, E.; Lowery, R.P.; Ugrinowitsch, C. Meta-Analysis of Postactivation Potentiation and Power: Effects of Conditioning Activity, Volume, Gender, Rest Periods, and Training Status. J. Strength Cond. Res. 2013, 27, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Seitz, L.B.; Trajano, G.S.; Haff, G.G.; Dumke, C.C.L.S.; Tufano, J.J.; Blazevich, A.J. Relationships between maximal strength, muscle size, and myosin heavy chain isoform composition and postactivation potentiation. Appl. Physiol. Nutr. Metab. 2016, 41, 491–497. [Google Scholar] [CrossRef]
- Baudry, S.; Duchateau, J. Postactivation potentiation in a human muscle: effect on the rate of torque development of tetanic and voluntary isometric contractions. J. Appl. Physiol. Bethesda Md 1985 2007, 102, 1394–1401. [Google Scholar] [CrossRef]
- Vargas, L.; Baratta, J.; Hu, X. Distribution of M-Wave and H-Reflex in Hand Muscles Evoked via Transcutaneous Nerve Stimulation: A Preliminary Report. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. IEEE Eng. Med. Biol. Soc. Annu. Int. Conf. 2021, 2021, 5897–5900. [Google Scholar] [CrossRef]
- Fukutani, A.; Miyamoto, N.; Kanehisa, H.; Yanai, T.; Kawakami, Y. Influence of the intensity of a conditioning contraction on the subsequent twitch torque and maximal voluntary concentric torque. J. Electromyogr. Kinesiol. Off. J. Int. Soc. Electrophysiol. Kinesiol. 2012, 22, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Zagatto, A.M.; Claus, G.M.; Dutra, Y.M.; De Poli, R.A.; Lopes, V.H.F.; Goodall, S.; Loturco, I.; Boullosa, D. Drop jumps versus sled towing and their effects on repeated sprint ability in young basketball players. BMC Sports Sci. Med. Rehabil. 2022, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Maffiuletti, N.A.; Martin, A.; Babault, N.; Pensini, M.; Lucas, B.; Schieppati, M. Electrical and mechanical H(max)-to-M(max) ratio in power- and endurance-trained athletes. J. Appl. Physiol. (1985) 2001, 90, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.L.; Stull, J.T. Myosin light chain phosphorylation in fast and slow skeletal muscles in situ. Am. J. Physiol. 1984, 247, C462–471. [Google Scholar] [CrossRef] [PubMed]
- Inns, T.B.; McCormick, D.; Greig, C.A.; Atherton, P.J.; Phillips, B.E.; Piasecki, M. Factors associated with electrical stimulation-induced performance fatigability are dependent upon stimulation location. Exp. Physiol. 2021, 106, 828–836. [Google Scholar] [CrossRef]
- Rodriguez-Falces, J.; Malanda, A.; Navallas, J.; Place, N. M-wave changes caused by brief voluntary and stimulated isometric contractions. Eur. J. Appl. Physiol. 2023, 123, 2087–2098. [Google Scholar] [CrossRef] [PubMed]
- Folland, J.P.; Wakamatsu, T.; Fimland, M.S. The influence of maximal isometric activity on twitch and H-reflex potentiation, and quadriceps femoris performance. Eur. J. Appl. Physiol. 2008, 104, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, R.M.; Ingersoll, C.D.; Hoffman, M.A. The hoffmann reflex: methodologic considerations and applications for use in sports medicine and athletic training research. J. Athl. Train. 2004, 39, 268–277. [Google Scholar] [PubMed]
- Racinais, S.; Girard, O.; Micallef, J.P.; Perrey, S. Failed excitability of spinal motoneurons induced by prolonged running exercise. J. Neurophysiol. 2007, 97, 596–603. [Google Scholar] [CrossRef]
- McNeil, C.J.; Butler, J.E.; Taylor, J.L.; Gandevia, S.C. Testing the excitability of human motoneurons. Front Hum Neurosci [Internet]. 2013 [cited 2023 Jun 13];7. Available from: http://journal.frontiersin.org/article/10.3389/fnhum.2013.00152/abstract. [CrossRef]
- Bergmann, J.; Kramer, A.; Gruber, M. Repetitive hops induce postactivation potentiation in triceps surae as well as an increase in the jump height of subsequent maximal drop jumps. PloS One 2013, 8, e77705. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Paternoster, F.K. Post-Activation-Performance Enhancement: Possible Contributing Factors. J. Sports Sci. Med. 2024, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Soler, E.; Paredes, X.; Carballeira, E.; Márquez, G.; Fernández-Del-Olmo, M. Effect of intensity and duration of conditioning protocol on post-activation potentiation and changes in H-reflex. Eur. J. Sport Sci. 2011, 11, 33–38. [Google Scholar] [CrossRef]
- Rodriguez-Falces, J.; Duchateau, J.; Muraoka, Y.; Baudry, S. M-wave potentiation after voluntary contractions of different durations and intensities in the tibialis anterior. J. Appl. Physiol. 2015, 118, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Larsson, S.C. Epidemiology of sarcopenia: Prevalence, risk factors, and consequences. Metabolism 2023, 144, 155533. [Google Scholar] [CrossRef]
- Kupa, E.J.; Roy, S.H.; Kandarian, S.C.; De Luca, C.J. Effects of muscle fiber type and size on EMG median frequency and conduction velocity. J. Appl. Physiol. Bethesda Md 1985 1995, 79, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Gerdle, B.; Karlsson, S.; Crenshaw, A.G.; Elert, J.; Fridén, J. The influences of muscle fibre proportions and areas upon EMG during maximal dynamic knee extensions. Eur. J. Appl. Physiol. 2000, 81, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 2021, n71. [Google Scholar] [CrossRef]
- Coar, J.T.; Sewell, J.P. Zotero: harnessing the power of a personal bibliographic manager. Nurse Educ. 2010, 35, 205–207. [Google Scholar] [CrossRef]
- Hodgson, M.J.; Docherty, D.; Zehr, E.P. Postactivation potentiation of force is independent of h-reflex excitability. Int. J. Sports Physiol. Perform. 2008, 3, 219–231. [Google Scholar] [CrossRef]
- Miyamoto, N.; Kanehisa, H.; Fukunaga, T.; Kawakami, Y. Effect of postactivation potentiation on the maximal voluntary isokinetic concentric torque in humans. J. Strength Cond. Res. 2011, 25, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Mina, M.A.; Blazevich, A.J.; Giakas, G.; Kay, A.D. Influence of variable resistance loading on subsequent free weight maximal back squat performance. J. Strength Cond. Res. 2014, 28, 2988–2995. [Google Scholar] [CrossRef] [PubMed]
- Mina, M.A.; Blazevich, A.J.; Giakas, G.; Seitz, L.B.; Kay, A.D. Chain-loaded variable resistance warm-up improves free-weight maximal back squat performance. Eur. J. Sport Sci. 2016, 16, 932–939. [Google Scholar] [CrossRef]
- Sotiropoulos, K.; Smilios, I.; Douda, H.; Christou, M.; Tokmakidis, S.P. Contrast Loading: Power Output and Rest Interval Effects on Neuromuscular Performance. Int. J. Sports Physiol. Perform. 2014, 9, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Barnes, M.J.; Petterson, A.; Cochrane, D.J. Effects of different warm-up modalities on power output during the high pull. J. Sports Sci. 2017, 35, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Tsoukos, A.; Brown, L.E.; Veligekas, P.; Terzis, G.; Bogdanis, G.C. Postactivation Potentiation of Bench Press Throw Performance Using Velocity-Based Conditioning Protocols with Low and Moderate Loads. J. Hum. Kinet. 2019, 68, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Caterini, D.; Gittings, W.; Huang, J.; Vandenboom, R. The effect of work cycle frequency on the potentiation of dynamic force in mouse fast twitch skeletal muscle. J. Exp. Biol. 2011, 214, 3915–3923. [Google Scholar] [CrossRef] [PubMed]
- Xeni, J.; Gittings, W.B.; Caterini, D.; Huang, J.; Houston, M.E.; Grange, R.W.; Vandenboom, R. Myosin light-chain phosphorylation and potentiation of dynamic function in mouse fast muscle. Pflugers Arch. 2011, 462, 349–358. [Google Scholar] [CrossRef]
- Arendt-Nielsen, L.; Mills, K.R. The relationship between mean power frequency of the EMG spectrum and muscle fibre conduction velocity. Electroencephalogr. Clin. Neurophysiol. 1985, 60, 130–134. [Google Scholar] [CrossRef]
- Komi, P.V.; Tesch, P. EMG frequency spectrum, muscle structure, and fatigue during dynamic contractions in man. Eur. J. Appl. Physiol. 1979, 42, 41–50. [Google Scholar] [CrossRef]
- Beretta-Piccoli, M.; D’Antona, G.; Barbero, M.; Fisher, B.; Dieli-Conwright, C.M.; Clijsen, R.; Cescon, C. Evaluation of central and peripheral fatigue in the quadriceps using fractal dimension and conduction velocity in young females. PloS One 2015, 10, e0123921. [Google Scholar] [CrossRef]
- Linssen, W.H.; Stegeman, D.F.; Joosten, E.M.; Merks, H.J.; ter Laak, H.J.; Binkhorst, R.A.; Notermans, S.L. Force and fatigue in human type I muscle fibres. A surface EMG study in patients with congenital myopathy and type I fibre predominance. Brain J. Neurol. 1991, 114 Pt 5, 2123–2132. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, M.; Houck, J.; Cuddeford, T.; Sharma, S.; Riley, N. Variations in the relationship between the frequency content of EMG signals and the rate of torque development in voluntary and elicited contractions. J. Electromyogr. Kinesiol. 2002, 12, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.K.; Timothy, G.F.; Yeow, C.H. Comparison of mean frequency and median frequency in evaluating muscle fiber type selection in varying gait speed across healthy young adult individuals. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. IEEE Eng. Med. Biol. Soc. Annu. Int. Conf. 2016, 2016, 1725–1728. [Google Scholar] [CrossRef]
- Bobbert, M.F.; van Ingen Schenau, G.J. Coordination in vertical jumping. J. Biomech. 1988, 21, 249–262. [Google Scholar] [CrossRef] [PubMed]
- McComas, A.J.; Galea, V.; Einhorn, R.W. Pseudofacilitation: a misleading term. Muscle Nerve 1994, 17, 599–607. [Google Scholar] [CrossRef]
- Hicks, A.; Fenton, J.; Garner, S.; McComas, A.J. M wave potentiation during and after muscle activity. J. Appl. Physiol. Bethesda Md 1985 1989, 66, 2606–2610. [Google Scholar] [CrossRef]
- Casabona, A.; Polizzi, M.C.; Perciavalle, V. Differences in H-reflex between athletes trained for explosive contractions and non-trained subjects. Eur. J. Appl. Physiol. 1990, 61, 26–32. [Google Scholar] [CrossRef]
Figure 1.
Flow diagram of the screening process.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



