
Submitted:
06 May 2024
Posted:
08 May 2024
You are already at the latest version
Abstract
Spontaneous tumours in higher plants can develop depending on plant genotype without any pathogen invasion. Spontaneous tumour formation on the taproots is consistently observed in certain inbred lines of radish (Raphanus sativus var. radicula Pers.). In this work, we have sequenced the genomes of two closely related radish inbred lines that differ in their ability to spontaneously form tumours. We have identified numerous single nucleotide variants (amino acid substitutions, insertions or deletions) that are likely to be associated with the spontaneous tumour formation. Furthermore, we performed the identification of CLE and WOX genes the genomic sequences of the radish inbred lines and, as a result, identified two unique radish CLE genes which probably encode proteins with multiple CLE domains. The results obtained provide a basis for investigating the mechanisms of plant tumour formation and also for future genetic and genomic studies of radish.
Keywords:
spontaneous tumours
; Raphanus sativus
; inbred lines
; genomic sequence
; single nucleotide variants
1. Introduction
Tumour formation is a pathological process that results from the uncontrolled proliferation of a group of cells. Tumours occur in virtually all multicellular organisms and are represented by newly formed tissues whose cells are actively proliferating. In animals, a system of proto-oncogenes and tumour suppressor genes forms a complex network which systemically controls the rate of cell division, growth and differentiation at the level of the whole organism. Disruption of such control, both under the influence of environmental factors and due to genomic instability, leads to the development of tumour growth. Higher plants contain functional orthologues of many mammalian tumour suppressors and oncogenes, but mutations in these genes in plants have not led to tumour formation, suggesting a very different principle of organization of the systemic control of cell division and differentiation in plants [1,2,3].
Most examples of plant tumours arise as a result of interactions with a variety of pathogens and phytophages, from bacteria and viruses to nematodes and arthropods [4]. More rarely, spontaneous tumours develop in the plants with specific genotypes (mutants, interspecific hybrids, inbred lines) in the absence of pathogens, making them more similar to animal tumours [1,4]. The exact causes of spontaneous tumour formation have only been studied in Arabidopsis mutants that have defects in cell-to-cell adhesion due to loss of function of enzymes involved in the biosynthesis or modification of cell wall components [5,6,7,8]. The study of these mutants has revealed one of the unknown aspects of the systemic control of cell division in plants, bringing cell adhesion to the fore [1]. At the same time, not all tumor mutants of Arabidopsis and other plants have impaired cell adhesion. Studying other examples of plant tumours could help identify other systemic regulators of cell division in higher plants.
The objects of our research are spontaneous tumours of the inbred lines of European cherry radish (Raphanus sativus var. radicula Pers.) (Figure 1A). The genetic collection of radish inbred lines has been maintained at St. Petersburg State University (SPbSU) since the 1960s by selfing individual plants, and now contains thirty three highly inbred lines, originated from four radish cultivars. Eleven radish inbred lines stably form tumours on the taproots of plants at the flowering stage ([9,10], Figure 1).
As with most examples of spontaneous tumours in plants, the mechanism triggering tumour formation in the radish inbred lines is unknown. Tumours on radish taproots originate from the pericycle and cambium as callus-like structure and later acquire features of secondary differentiation, such as vasculature, and meristematic foci similar to root apical meristems due to auxin maxima and WOX5 expression [11]. The RNA-seq of radish tumours compared to lateral roots revealed the differential expression of the more than 1600 genes [12]. Most of the pathways upregulated in radish tumours were associated with the control of cell division, showing extreme activation of this process in the tumour tissue [12].
In the present work, we have sequenced the genomes of two closely related radish inbred lines 18 and 19 that contrastingly differ in their ability to form tumours ([9], Figure 1). In genetic crosses between these two lines, this trait was inherited as a monogenic recessive [13], providing an opportunity to identify a specific gene that regulates spontaneous tumour formation.
As a result, a number of SNVs (InDels and SNPs) was revealed in the tumour radish line. Among these, we found more than a hundred SNVs in the CDS of protein-coding genes that are thought to lead to changes in protein structure ("stop lost" / "stop gained" or a frameshift) or in the position 1-20 of 5’-UTR that could severely influence the translation efficiency [14]. Many of the genes with such SNVs in the tumour-forming line are homologs of Arabidopsis genes, which are involved in cell cycle regulation, cytoskeleton organisation, meristem development and phytohormone homeostasis. Among them, we selected 108 SNVs which are in the homozygous state in the tumour radish line. The presence of the selected InDels and SNPs in the radish tumour line was verified by sequencing the amplicons of the corresponding gene regions in the radish lines 18 and 19.
To search for the association of SNVs with spontaneous tumour formation, we performed the sequencing of 40 SNV-containing gene regions in seven tumour and fourteen non-tumour radish lines of the SPbSU genetic collection. As a result, we found that the RsERF018 gene contains the CAG insertion in the 5’-UTR close to start codon in most tumour radish lines and only two non-tumour lines, which allows us to propose it as a candidate regulator of spontaneous tumour formation.
Based on genome assemblies’ data of two radish inbred lines, we carried out the identification and chromosomal location of the genes belonging to the CLE and WOX families which are known to be master regulators of meristem identity and stem cell homeostasis. Among them, we identified new, previously uncharacterised radish CLE genes which are likely to encode proteins with multiple CLE domains. Homologs of such a group of CLEs are absent in Arabidopsis, but have been identified in Brassica napus [15].
The sequencing of the genome of the tumour radish line may be a step towards identifying new mechanisms underlying the spontaneous tumour formation in higher plants.
2. Results
2.1. Assessment of the Assembly Quality of the Genomes of Two Radish Inbred Lines
To compare the genomic DNA sequences of tumour radish line 19 and non-tumour radish line 18, we performed a hybrid chromosome-level assembly using a combination of data obtained by Illumina and Oxford Nanopore sequencing methods.
As a result of the assembly quality assessment using the BUSCO programme (https://busco.ezlab.org/), it was shown that the number of single copies of nuclear genes was greater than 92.2% for line 19 and 91,1% for line 18. The number of duplicated sequences was 6.4% for line 19 and 5.7% for line 18, and the overall assembly quality index was greater than 98.6 % for line 19 and 96,8 % for line 18, indicating a low content of fragmented or incomplete sequences and no contamination by sequences from other phylogenetic taxa (Figure 2). The assembly parameter values obtained using the Quast programme indicated that the genome size of line 18 was 492,907,896 bp with N50 = 12750, and genome size of line 19 was 480,234,765 bp, and N50 = 13846043. These parameters are comparable to the characteristics of reference radish genomes [16,17,18].
A BUSCO analysis of the genome assemblies of lines 18 and 19 has shown the quality indicators as 93.8 and 98.9, respectively. Thus, the assemblies quality of the radish lines genomes acquired in this work is not inferior to those available in the NCBI database (https://www.ncbi.nlm.nih.gov/datasets/genome/?taxon=3725; available on 23.01.2024).
2.2. Identification of SNVs in the Protein-Coding Genes of Tumour Radish Line
When analysing the genome sequences of tumour and non-tumour radish lines, we have identified a large number of SNVs (514083 InDels and 2260270 SNPs) in tumour line 19 (Table 1, Suppl. Fig.1). Among them, 35399 InDels and 688148 SNPs were located in the CDS of protein-coding genes or in position -1-20 of 5’-UTR. Of these, 21698 InDels and 9451 were likely to result in the altered translation of the corresponding proteins due to frameshift, loss of start or stop codon, gain of start codon, or decrease of the translation efficiency due to the changes in the 5’-UTR close to start [14].
Among the genes with these SNVs, we selected 240 InDels and 135 SNPs in the genes related to GO probably associated with the control of plant cell proliferation: related to regulation of cell growth (GO:0008283, GO:0007346, GO:0010564, GO:0000278, GO:0051726, GO:0006261, GO:0042023, GO:0000910, GO:0000911, GO:0000226, GO:0009828, GO:0009505, GO:0009825), meristem activity (GO:0010014, GO:0010075, GO:0009933), phytohormone signaling (GO:0009736, GO:0009690, GO:0009686, GO:0045487, GO:0009734, GO:0009733, GO:0009735, GO:00097390, gene expression regulation (GO:0003700, GO:0006306, GO:0034968, GO:0051567), and organogenesis (GO:0048364, GO:0048527, GO:0090451).
Among the genes belonging to these GO pathways, 72 genes with InDels and 36 genes with SNPs were in the homozygous state in the radish line 19. Of these 72 InDels, 57 resulted in frameshift, 9 - in frameshift and loss of start codon, 5 - in frameshift and gain of stop codon, and 1 – in change of the 5’-UTR near start codon. For 36 SNPs, 23 resulted in stop codon gain, 10 - in stop codon loss, and 3 - in start codon loss. We performed the chromosomal location of genes with such SNVs (Figure 3 and Figure 4). More detailed information on these genes is presented in Supplementary Tables 1 and 2.
It can be assumed that the abovementioned SNVs could lead to loss of function of the corresponding protein-coding genes in the tumour radish line, and thus each of these SNVs could cause tumour formation. The effects of loss-of-function mutations in some of these genes on plant development have also been described for their homologs in Arabidopsis (Suppl. Tables 1, 2).
2.3. Search for the Presence of Identified SNVs in the Tumour and Non-Tumour Lines of the Radish Genetic Collection
To search for probable candidate regulators of spontaneous tumour formation among genes containing selected SNVs in the line 19, we amplified the corresponding gene regions of several other tumour (12, 13, 14, 16, 20, 21, 32) and non-tumour (3, 5, 6, 8, 9, 23, 25, 26, 27, 28, 29, 30, 37, 39) lines of the radish genetic collection.
As a result, the presence of the same SNV in most tumour lines was confirmed for the RsERF018 gene (Figure 4). As for the other 39 genes, in some of them SNVs were only identified in line 19, or there was a polymorphism that was not associated with the tumour formation trait.
The RsERF018 gene, whose homolog in Arabidopsis controls response to ethylene and cambium cell division [19], contains a CAG insertion just upstream of the start codon of the gene in the tumour lines 12, 13, 14, 19, 20 and 21, and also in non-tumour line 26 and 27, whereas no insertion was detected in the tumour lines 16, 32 and in most non-tumour lines (Figure 4). According to data obtained in Arabidopsis, this kind of changes in the positions -1-20 of the 5’-UTR dramatically decreases the efficiency of translation [20].
The RsERF018 gene needs to be further investigated as a possible regulator of spontaneous tumour formation.
2.4. Identification and Chromosomal Localisation of WOX and CLE Genes in the Obtained Genome Assemblies of Inbred Radish Lines
Meristem regulators are known to be involved in the control of the plant cell division plan, and they was shown to participate in the development of numerous examples of plant tumours [4]. The balance between cell division and differentiation in various plant meristems is under control of WOX-CLAVATA system, a highly conserved regulatory module [21], which consists of CLAVATA3/EMBRYO SURROUNDING REGION-related (CLE) peptides, the protein kinase receptors that bind CLEs, and the targets of CLE action, the WUSCHEL-RELATED HOMEOBOX (WOX) homeodomain transcription factors [22,23,24].
We carried out the identification of the radish CLE and WOX family genes in our genome assemblies of 18 and 19 radish lines (Figure 5, Suppl. Fig. 2, 3). A total of 52 RsCLE genes and 24 RsWOX genes were found. All 24 RsWOX genes were previously identified [25]. Among RsCLE genes, 16 RsCLEs were identified in our previous work [26], and other RsCLE genes were annotated in the reference radish genome [16]. The chromosomal location of RsWOX and RsCLE genes (Figure 5) revealed the clusters of closely located RsCLEs on 2, 4 and 9 radish chromosomes.
It is important to note that the genes RsWOX2, RsWOX14 and RsCLE7 were among those in which SNVs in the CDS, which probably leads to loss of function, were identified in tumour radish line 19 (Suppl. Figure 6, Suppl. Tables 1, 2). At the same time, these SNVs were only confirmed in tumour line 19 and not in other radish tumour lines.
2.4. Identification of Radish CLE Genes Likely to Encode Proteins with Multiple CLE Domains
Among all the RsCLE genes identified in this work (Figure 7), we have found two unique RsCLEs of unknown function, which probably encode proteins with multiple CLE domains. We then found the same genes in the radish reference genome, where they had not been described as CLE genes and named in the NCBI database as actin-binding protein wsp1-like (LOC108807713) and proline-rich receptor-like protein kinase PERK10 (LOC108858878). We have uploaded the sequences of these genes found in our assemblies to the NCBI database (Submission ID: 2791313, GenBank numbers PP236904.1 and PP236905.1) under the names RsCLEm1 and RsCLEm2 (“RsCLE multidomain”).
3. Discussion
To date, radish genome sequencing has previously been carried out for several Asian and European cultivars and isolates [16,17,27,28,29,30]. The Rs1.0 genome, which is a radish reference genome, was based on the chromosome sequences of R. sativus of the Korean cultivar WK10039 [16].
In our work, we sequenced the genomes of two closely related radish inbred lines which differ in their ability to form spontaneous tumours [9,10,11,12,13]. This is the first attempt to sequence the genome of plants with spontaneous tumours formation.
To date, the most well-studied examples of spontaneous tumours in higher plants are several monogenic mutants of Arabidopsis [5,31,32,33] and one of Nicotiana tabacum [34], which form tumours on different organs of seedlings. In most cases, tumours in these mutants are the result of loss of function of pectin metabolism genes, which are involved in cell wall formation and cell adhesion [5,6,7,8]. The discovery of such mutants showed that cell adhesion is one of the mechanisms that systemically regulate cell proliferation in the plant body. However, cell adhesion is not the only mechanism of such systemic regulation. In Arabidopsis there are also tumour-forming mutants with loss of function of the other the genes whose association with tumour development is much less obvious, such as the gene encoding the immunophilin family protein [31], the tyrosine phosphatase-like protein [35], and the chromatin remodelling factor [36]. Thus, the identification of plant genes whose loss of function leads to spontaneous tumour formation will help to identify new systemic mechanisms for cell division control in higher plants.
In our work, we have identified numerous SNVs, including those in the CDS or position -1-20 of the 5’UTR of protein-coding genes, that distinguish the tumour radish line from the relative non-tumour line. Therefore, we can assume that certain SNVs could be inducers of spontaneous tumour formation. According to data on transcriptome analysis of the roots and spontaneous tumours in the radish inbred line, all 108 genes with loss-of-function SNVs in the tumour line 19, were expressed in radish taproots [12]. Moreover, five genes with such SNVs identified in this study were among the DEGs: the expression levels of the cell cycle regulator RsPCNA1 and the gene of unknown function LOC108817684 were increased in the tumours, whereas the expression levels of the radish homologs of the auxin response gene RsSAUR32, the ethylene response cambium-associated genes RsERF018 and RsERF019, and also the RsLRR-RK gene encoding receptor-like protein kinase were decreased [12].
Due to the large number of SNVs identified, it is currently not possible to make clear assumptions about the role of each SNV in spontaneous tumour formation. Additional testing for the presence of the identified SNVs in tumour and non-tumour radish lines revealed that a CAG insertion at position -1 of 5’-UTR of the RsERF018 gene was present in the seven out of eight tumour radish lines tested and absent in the thirteen out of fifteen non-tumour lines. Without the insertion, this region contained an AAA sequence just before the start codon, which should result in high translation efficiency [20]. Therefore, an insertion of a CAG between the start and the AAA region (Figure 4) should result in a considerable decrease in the amount of the translated protein, as had been shown in Arabidopsis [20].
In this work, we also characterised and chromosomally localised gene genes of the WOX and CLE families in the genomic sequences of radish lines from the SPbSU genetic collection. Among the RsWOX and RsCLE genes, the loss-of-function SNVs were detected in the RsWOX14, RsWOX2 and RsCLE7 genes in line 19 (Suppl. Figure 6).
In Arabidopsis, the WOX14 gene is a regulator of cambium and xylem balance and acts redundantly with WOX4 [37]. The WOX2 is known to be a regulator of early embryogenesis and a callus formation [38]. The CLE7 gene in Arabidopsis also functions as a regulator of callus formation and regeneration [39]. Since, according to our previous data, spontaneous tumours on radish taproots originate from the cambium and develop as undifferentiated callus-like structures [11], these genes are perspective candidates for tumour regulators. However, the results on these were not very encouraging, as our data show that none of the corresponding SNVs were found in the sequences of these genes in other radish tumour lines studied.
The genes RsWOX14, RsWOX2 and RsCLE7 are represented by a single copy in the radish genome, but homozygosity for loss-of-function mutations in them does not result in reduced viability of the radish line 19. According to available data, a single mutation in each of these genes in Arabidopsis did not cause any serious developmental abnormalities in the mature plants [37].
Analysis of the genomes of radish lines also allowed us to identify two RsCLE genes, RsCLEm1 and RsCLEm2, which are likely to encode proteins with multiple CLE domains and a unique CLE domain composition (Figure 7). There are no identified homologs of these genes in Arabidopsis, but they are related to the B. napus CLEm genes, which encode multidomain CLE proteins that function as light stimulators of shoot apical meristem activity [15]. The RsCLEms contain eight tandem CLE domain sequences and are close to BnCLEm3, whose product contains five nearly identical tandem CLE domains [15].
Thus, in addition to identifying SNVs probably associated with tumours, the sequencing of the radish inbred lines allowed the identification of novel CLE family genes.
4. Materials and Methods
4.1. Plant Material
Closely related lines 18 and 19 of the R. sativus genetic collection were used in this study. Both lines originated from a single self-pollinated plant of the Saxa cultivar [9] and are now represented by the progeny of approximately 40-45 generations of inbreeding, indicating an extremely low level of heterozygosity.
Saxa (cat. № 9464454 in the State Register of Selection Achievements Admitted for Use (National List).) is the cultivar of European radish with round red taproot, which was obtained at the Federal Scientific Centre for Vegetable Growing (Moscow Region, Russia) by the method of mass selection from a sample originating from Central Europe. This radish variety has not yet been subjected to genome sequencing.
4.2. Genomic DNA Isolation, Library Preparation and Sequencing
Total DNA was isolated from 50 of 7-day old etiolated radish seedlings of inbred lines according to an unpublished protocol approved by the Laboratory of Plant-Microbial Interactions of All-Russia Research Institute for Agricultural Microbiology (ARRIAM).
DNA sequencing of line 19 was performed using Oxford Nanopore technology in the Core Centrum “Genomic Technologies, Proteomics and Cell Biology” at the ARRIAM using a MinION device (Oxford Nanopore, Cambridge, UK). The genome assembly of line 19 was performed using the Canu v.1.7.1 tool (https://github.com/marbl/canu/releases). Sequencing of line 19 was also performed on the Illumina HiSeq2500 sequencer at the Centre of Molecular and Cellular Technologies of Saint Petersburg State University Research Park. The NEBNext® Ultra™ DNA Library Prep Kit for Illumina (New England Biolabs) was used for library construction. Dual barcoding was performed using the NEBNext® Ultra™ DNA Index Prep Kit for Illumina and NEBNext® Multiplex Oligos® Illumina® (Dual Index Primers Set 1). To improve the quality of the genome assemblies, the data were refined to correct possible errors in the Pilon v.1.22 tool (https://github.com/broadinstitute/pilon/releases) based on data obtained by two sequencing technologies (Illumina and Nanopore).
DNA sequencing of line 18 was performed on an Illumina at the Centre of Molecular and Cellular Technologies of Saint Petersburg State University Research Park using HiSeq2500 sequencer. The NEBNext® Ultra™ DNA Library Prep Kit for Illumina (New England Biolabs) was used for library construction. Dual barcoding was performed using the NEBNext® Ultra™ DNA Index Prep Kit for Illumina and NEBNext® Multiplex Oligos® Illumina® (Dual Index Primers Set 1). Line 18 genome assembly was performed using the SOAPdenovo v.2.04 tool (https://github.com/aquaskyline/SOAPdenovo2) with maximal read length = 150, average insert size =100, cutoff of pair number for a reliable connection = 5).
4.3. Bioinformatic Processing of the Sequencing Results
For each assembly, MultiQC v.1.12 [40] and Trimmomatic v.0.40 with the HEADCROP:15 and CROP:140 options [41] programmes were used for quality control and read correction, respectively. The assemblies were indexed using the bowtie2 tool (https://github.com/BenLangmead/bowtie2). Assemblies of two chromosome-level genomes were generated using the Ragtag tool (https://github.com/malonge/RagTag) and the chromosome-level reference radish genome GCA_019703475.1 (https://www.ncbi.nlm.nih.gov/data-hub/genome/GCA_019703475.1/).
Annotation of the genomes of lines 18 and 19 was performed using the Augustus Gene Prediction Tool (https://github.com/Gaius-Augustus/Augustus) with –species=arabidopsis parameter.
Alignment of the line 19 sequences to the line 18 genome assembly and vice versa was performed using the bowtie2 program (https://github.com/BenLangmead/bowtie2), and identification of candidate genes and differences in the structure of these genes in different radish lines was performed using the SnpEff [42], SnpSift [42] and GATK with the HaplotypeCaller option (https://gatk.broadinstitute.org/hc/en -us/articles/360037225632-HaplotypeCaller), SelectVariants --select-type SNP or --select-type INDEL options programmes.
Visualisation of the sequence alignment for assembly and checking for the presence of InDels and SNPs in silico was performed in the IGV genome browser (https://igv.org/).
Sequences for the RsWOX genes were searched in the radish genome assemblies represented in the NBCI database (https://www.ncbi.nlm.nih.gov/datasets/genome/?taxon=3725; available on 23.01.2024) using the blastP, blastN and tblastN algorithms of the NCBI database (https://blast.ncbi.nlm.nih.gov/Blast.cgi), based on the nucleotide and amino acid sequences of A. thaliana and R. sativus genes and proteins.
The phylogenetic tree of radish CLE protein sequences was constructed based on the alignment of R. sativus CLE protein amino acid sequences in MEGA7 software (https://www.megasoftware.net/) using the Muscle algorithm [43] by Neighbour joining [44] with default parameters and bootstrap 1000 [45]; the tree was visualised using iTOL software (https://itol.embl.de/).
Nucleotide and amino acid sequences were analysed using the following programmes: ApE (https://jorgensen.biology.utah.edu/wayned/ape/, v.3.1.0), SnapGENE (https://www.snapgene.com/; v.6.0.2), UGENE (http://ugene.net/ru/; v.33), MEGA7 (https://www.megasoftware.net/; v. 10.2). Signal motifs were predicted with the SignalP-6.0 tool (https://services.healthtech.dtu.dk/service.php?SignalP).
The location of genes on radish chromosomes was visualised using the MapChart 2.32 software ((https://www.wur.nl/en/show/mapchart.htm).
The search for domains in proteins and their visualisation was performed using the MEME online tool (https://meme-suite.org/meme/tools/meme).
5. Conclusions
By sequencing the genomes of related tumour and non-tumour radish lines, it was possible to identify a number of candidate genes for the role of regulators of spontaneous tumours. Further study of the relationship between the identified genes and tumour formation could increase our knowledge of the role of different pathways involved in the systemic regulation of plant cell division. In addition, this work analysed the radish WOX and CLE family genes and identified new, previously unknown CLE genes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, I.D. and L.L.; methodology, X.K., A.A, E.G., L.D., M.G., V.T., N.G.; software, X.K., A.A., L.D.; validation, X.K., A.A., L.D; formal analysis, X.K.; investigation, X.K.; resources, X.K.; data curation, X.K.; writing—original draft preparation, X.K.; writing—review and editing, I. D. and V.T.; visualization, K.K. and M.G.; supervision, I.D.; project administration, L.L.; funding acquisition, L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Ministry of Science and Higher Education of the Russian Federation in accordance with the agreement № 075-15-2022-322 dated 22.04.2022 on providing a grant in the form of subsidies from the federal budget of the Russian Federation. The grant was provided for state support for the creation and development of a World-class Scientific Center “Agrotechnologies for the Future”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article. Raw sequence data are uploaded as BioProject submission SUB13927602.
Acknowledgments
We thank Alexey Masharskii, Anna Romanovich (Centre of Molecular and Cellular Technologies of Saint Petersburg State University Research Park) for DNA sequencing. We are also grateful to Paulina A. Pavlova and Tessa C. Kovalevski for their helpful suggestions during bioinformatics data processing. We also express our sincere gratitude to Maria Lebedeva for her advice and critical reading. The article is dedicated to the 300th anniversary of St. Petersburg State University.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| InDel | Insertion or Deletion |
| SNP | Single Nucleotide Polymorphism |
| SNV | Single Nucleotide Variant. |
References
- Doonan, J.H.; Sablowski, R. Walls around Tumours - Why Plants Do Not Develop Cancer. Nat Rev Cancer 2010, 10, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Harashima, H.; Sugimoto, K. Integration of Developmental and Environmental Signals into Cell Proliferation and Differentiation through RETINOBLASTOMA-RELATED 1. Curr Opin Plant Biol 2016, 29, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Komaki, S.; Sugimoto, K. Control of the Plant Cell Cycle by Developmental and Environmental Cues. Plant Cell Physiol 2012, 53, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Dodueva, I.E.; Lebedeva, M.A.; Kuznetsova, K.A.; Gancheva, M.S.; Paponova, S.S.; Lutova, L.L. Plant Tumors: A Hundred Years of Study. Planta 2020, 251, 82. [Google Scholar] [CrossRef]
- Krupková, E.; Immerzeel, P.; Pauly, M.; Schmülling, T. The TUMOROUS SHOOT DEVELOPMENT2 Gene of Arabidopsis Encoding a Putative Methyltransferase Is Required for Cell Adhesion and Co-Ordinated Plant Development. Plant J 2007, 50, 735–750. [Google Scholar] [CrossRef] [PubMed]
- Paredez, A.R.; Persson, S.; Ehrhardt, D.W.; Somerville, C.R. Genetic Evidence That Cellulose Synthase Activity Influences Microtubule Cortical Array Organization. Plant Physiol 2008, 147, 1723–1734. [Google Scholar] [CrossRef] [PubMed]
- Krupková, E.; Schmülling, T. Developmental Consequences of the Tumorous Shoot Development1 Mutation, a Novel Allele of the Cellulose-Synthesizing KORRIGAN1 Gene. Plant Mol Biol 2009, 71, 641–655. [Google Scholar] [CrossRef] [PubMed]
- Iwai, H.; Masaoka, N.; Ishii, T.; Satoh, S. A Pectin Glucuronyltransferase Gene Is Essential for Intercellular Attachment in the Plant Meristem. Proc Natl Acad Sci U S A 2002, 99, 16319–16324. [Google Scholar] [CrossRef]
- Buzovkina, I.S.; Lutova, L.A. The Genetic Collection of Radish Inbred Lines: History and Prospects. Russ J Genet 2007, 43, 1181–1192. [Google Scholar] [CrossRef]
- Narbut, S.I. [Genetic tumor in a radish obtained by inbreeding]. Vestn Leningr Univ Biol 1967, 3, 144–149. [Google Scholar]
- Lebedeva (Osipova), M.A.; Tvorogova, V.E.; Vinogradova, A.P.; Gancheva, M.S.; Azarakhsh, M.; Ilina, E.L.; Demchenko, K.N.; Dodueva, I.E.; Lutova, L.A. Initiation of Spontaneous Tumors in Radish (Raphanus Sativus): Cellular, Molecular and Physiological Events. Journal of Plant Physiology 2015, 173, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Tkachenko, A.; Dodueva, I.; Tvorogova, V.; Predeus, A.; Pravdina, O.; Kuznetsova, K.; Lutova, L. Transcriptomic Analysis of Radish (Raphanus Sativus L.) Spontaneous Tumor. Plants (Basel) 2021, 10, 919. [Google Scholar] [CrossRef] [PubMed]
- Matveeva, T.V.; Frolova, N.V.; Smets, R.; Dodueva, I.E.; Buzovkina, I.S.; Van Onckelen, H.; Lutova, L.A. Hormonal Control of Tumor Formation in Radish. J Plant Growth Regul 2004, 23, 37–43. [Google Scholar] [CrossRef]
- Jackson, R.J.; Hellen, C.U.T.; Pestova, T.V. THE MECHANISM OF EUKARYOTIC TRANSLATION INITIATION AND PRINCIPLES OF ITS REGULATION. Nat Rev Mol Cell Biol 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Khan, M.H.U.; Yang, Y.; Zhu, K.; Li, H.; Zhu, M.; Amoo, O.; Khan, S.U.; Fan, C.; Zhou, Y. Identification and Comprehensive Analysis of the CLV3/ESR-Related (CLE) Gene Family in Brassica Napus L. Plant Biol (Stuttg) 2020, 22, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.-M.; Kim, N.; Ahn, B.O.; Oh, M.; Chung, W.-H.; Chung, H.; Jeong, S.; Lim, K.-B.; Hwang, Y.-J.; Kim, G.-B.; et al. Elucidating the Triplicated Ancestral Genome Structure of Radish Based on Chromosome-Level Comparison with the Brassica Genomes. Theor Appl Genet 2016, 129, 1357–1372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, T.; Wang, J.; Wang, P.; Qiu, Y.; Zhao, W.; Pang, S.; Li, X.; Wang, H.; Song, J.; et al. Pan-Genome of Raphanus Highlights Genetic Variation and Introgression among Domesticated, Wild, and Weedy Radishes. Mol Plant 2021, 14, 2032–2055. [Google Scholar] [CrossRef] [PubMed]
- Shirasawa, K.; Hirakawa, H.; Fukino, N.; Kitashiba, H.; Isobe, S. Genome Sequence and Analysis of a Japanese Radish (Raphanus Sativus) Cultivar Named ‘Sakurajima Daikon’ Possessing Giant Root. DNA Research 2020, 27, dsaa010. [Google Scholar] [CrossRef] [PubMed]
- Etchells, J.P.; Provost, C.M.; Turner, S.R. Plant Vascular Cell Division Is Maintained by an Interaction between PXY and Ethylene Signalling. PLoS Genet 2012, 8, e1002997. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, G.; Jeon, E.; Sohn, E. ju; Lee, Y.; Kang, H.; Lee, D. wook; Kim, D.H.; Hwang, I. The Immediate Upstream Region of the 5′-UTR from the AUG Start Codon Has a Pronounced Effect on the Translational Efficiency in Arabidopsis Thaliana. Nucleic Acids Res 2014, 42, 485–498. [Google Scholar] [CrossRef]
- Kuznetsova, K.; Efremova, E.; Dodueva, I.; Lebedeva, M.; Lutova, L. Functional Modules in the Meristems: “Tinkering” in Action. Plants (Basel) 2023, 12, 3661. [Google Scholar] [CrossRef] [PubMed]
- Tvorogova, V.E.; Krasnoperova, E.Y.; Potsenkovskaia, E.A.; Kudriashov, A.A.; Dodueva, I.E.; Lutova, L.A. [What Does the WOX Say? Review of Regulators, Targets, Partners]. Mol Biol (Mosk) 2021, 55, 362–391. [Google Scholar] [CrossRef]
- Song, X.-F.; Hou, X.-L.; Liu, C.-M. CLE Peptides: Critical Regulators for Stem Cell Maintenance in Plants. Planta 2021, 255, 5. [Google Scholar] [CrossRef]
- Poliushkevich, L.O.; Gancheva, M.S.; Dodueva, I.E.; Lutova, L.A. Receptors of CLE Peptides in Plants. Russ J Plant Physiol 2020, 67, 1–16. [Google Scholar] [CrossRef]
- Aliaga Fandino, A.C.; Kim, H.; Rademaker, J.D.; Lee, J.-Y. Reprogramming of the Cambium Regulators during Adventitious Root Development upon Wounding of Storage Tap Roots in Radish (Raphanus Sativus L.). Biol Open 2019, 8, bio039677. [Google Scholar] [CrossRef]
- Gancheva, M.S.; Dodueva, I.E.; Lebedeva, M.A.; Tvorogova, V.E.; Tkachenko, A.A.; Lutova, L.A. Identification, Expression, and Functional Analysis of CLE Genes in Radish (Raphanus Sativus L.) Storage Root. BMC Plant Biol, 1. [CrossRef]
- Kitashiba, H.; Li, F.; Hirakawa, H.; Kawanabe, T.; Zou, Z.; Hasegawa, Y.; Tonosaki, K.; Shirasawa, S.; Fukushima, A.; Yokoi, S.; et al. Draft Sequences of the Radish (Raphanus Sativus L.) Genome. DNA Res 2014, 21, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Jeong, Y.-M.; Jeong, S.; Kim, G.-B.; Baek, S.; Kwon, Y.-E.; Cho, A.; Choi, S.-B.; Kim, J.; Lim, W.-J.; et al. Identification of Candidate Domestication Regions in the Radish Genome Based on High-Depth Resequencing Analysis of 17 Genotypes. Theor Appl Genet 2016, 129, 1797–1814. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Qiu, Y.; Wang, H.; Wang, P.; Zhang, X.; Li, C.; Song, J.; Gui, W.; Shen, D.; et al. SSR-Sequencing Reveals the Inter- and Intraspecific Genetic Variation and Phylogenetic Relationships among an Extensive Collection of Radish (Raphanus) Germplasm Resources. Biology (Basel) 2021, 10, 1250. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Y.; Dong, J.; Zhang, W.; Tang, M.; Zhang, W.; Wang, K.; Chen, Y.; Zhang, X.; He, Q.; et al. A Chromosome-Level Genome Assembly of Radish (Raphanus Sativus L.) Reveals Insights into Genome Adaptation and Differential Bolting Regulation. Plant Biotechnol J 2023, 21, 990–1004. [Google Scholar] [CrossRef]
- Smyczynski, C.; Roudier, F.; Gissot, L.; Vaillant, E.; Grandjean, O.; Morin, H.; Masson, T.; Bellec, Y.; Geelen, D.; Faure, J.-D. The C Terminus of the Immunophilin PASTICCINO1 Is Required for Plant Development and for Interaction with a NAC-like Transcription Factor. J Biol Chem 2006, 281, 25475–25484. [Google Scholar] [CrossRef]
- Sieberer, T.; Hauser, M.-T.; Seifert, G.J.; Luschnig, C. PROPORZ1, a Putative Arabidopsis Transcriptional Adaptor Protein, Mediates Auxin and Cytokinin Signals in the Control of Cell Proliferation. Curr Biol 2003, 13, 837–842. [Google Scholar] [CrossRef]
- Frémont, N.; Riefler, M.; Stolz, A.; Schmülling, T. The Arabidopsis TUMOR PRONE5 Gene Encodes an Acetylornithine Aminotransferase Required for Arginine Biosynthesis and Root Meristem Maintenance in Blue Light. Plant Physiol 2013, 161, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.H. The Inheritance of Genetic Tumors in Nicotiana Hybrids. Journal of Heredity 1988, 79, 277–283. [Google Scholar] [CrossRef]
- Bellec, Y.; Harrar, Y.; Butaeye, C.; Darnet, S.; Bellini, C.; Faure, J.-D. Pasticcino2 Is a Protein Tyrosine Phosphatase-like Involved in Cell Proliferation and Differentiation in Arabidopsis. Plant J 2002, 32, 713–722. [Google Scholar] [CrossRef]
- Mao, Y.; Pavangadkar, K.A.; Thomashow, M.F.; Triezenberg, S.J. Physical and Functional Interactions of Arabidopsis ADA2 Transcriptional Coactivator Proteins with the Acetyltransferase GCN5 and with the Cold-Induced Transcription Factor CBF1. Biochim Biophys Acta 2006, 1759, 69–79. [Google Scholar] [CrossRef]
- Etchells, J.P.; Provost, C.M.; Mishra, L.; Turner, S.R. WOX4 and WOX14 Act Downstream of the PXY Receptor Kinase to Regulate Plant Vascular Proliferation Independently of Any Role in Vascular Organisation. Development 2013, 140, 2224–2234. [Google Scholar] [CrossRef]
- Hassani, S.B.; Trontin, J.-F.; Raschke, J.; Zoglauer, K.; Rupps, A. Constitutive Overexpression of a Conifer WOX2 Homolog Affects Somatic Embryo Development in Pinus Pinaster and Promotes Somatic Embryogenesis and Organogenesis in Arabidopsis Seedlings. Front Plant Sci 2022, 13, 838421. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Wang, X.; Ishida, T.; Grienenberger, E.; Zheng, Q.; Wang, J.; Zhang, Y.; Chen, W.; Chen, M.; Song, X.-F.; et al. A Group of CLE Peptides Regulates de Novo Shoot Regeneration in Arabidopsis Thaliana. New Phytol 2022, 235, 2300–2312. [Google Scholar] [CrossRef] [PubMed]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A Program for Annotating and Predicting the Effects of Single Nucleotide Polymorphisms, SnpEff. Fly (Austin) 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Molecular Biology and Evolution 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. CONFIDENCE LIMITS ON PHYLOGENIES: AN APPROACH USING THE BOOTSTRAP. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Dexter, R.; Qualley, A.; Kish, C.M.; Ma, C.J.; Koeduka, T.; Nagegowda, D.A.; Dudareva, N.; Pichersky, E.; Clark, D. Characterization of a Petunia Acetyltransferase Involved in the Biosynthesis of the Floral Volatile Isoeugenol. Plant J 2007, 49, 265–275. [Google Scholar] [CrossRef]
- Randall, R.S.; Miyashima, S.; Blomster, T.; Zhang, J.; Elo, A.; Karlberg, A.; Immanen, J.; Nieminen, K.; Lee, J.-Y.; Kakimoto, T.; et al. AINTEGUMENTA and the D-Type Cyclin CYCD3;1 Regulate Root Secondary Growth and Respond to Cytokinins. Biol Open 2015, 4, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Cromer, L.; Jolivet, S.; Horlow, C.; Chelysheva, L.; Heyman, J.; De Jaeger, G.; Koncz, C.; De Veylder, L.; Mercier, R. Centromeric Cohesion Is Protected Twice at Meiosis, by SHUGOSHINs at Anaphase I and by PATRONUS at Interkinesis. Curr Biol 2013, 23, 2090–2099. [Google Scholar] [CrossRef] [PubMed]
- Kono, A.; Umeda-Hara, C.; Lee, J.; Ito, M.; Uchimiya, H.; Umeda, M. Arabidopsis D-Type Cyclin CYCD4;1 Is a Novel Cyclin Partner of B2-Type Cyclin-Dependent Kinase. Plant Physiol 2003, 132, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Day, R.C.; Müller, S.; Macknight, R.C. Identification of Cytoskeleton-Associated Genes Expressed during Arabidopsis Syncytial Endosperm Development. Plant Signal Behav 2009, 4, 883–886. [Google Scholar] [CrossRef]
- Pradillo, M.; Knoll, A.; Oliver, C.; Varas, J.; Corredor, E.; Puchta, H.; Santos, J.L. Involvement of the Cohesin Cofactor PDS5 (SPO76) During Meiosis and DNA Repair in Arabidopsis Thaliana. Front Plant Sci 2015, 6, 1034. [Google Scholar] [CrossRef]
- Lin, C.; Choi, H.-S.; Cho, H.-T. Root Hair-Specific EXPANSIN A7 Is Required for Root Hair Elongation in Arabidopsis. Mol Cells 2011, 31, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xu, L.; Lin, H.; Cao, J. Two Expansin Genes, AtEXPA4 and AtEXPB5, Are Redundantly Required for Pollen Tube Growth and AtEXPA4 Is Involved in Primary Root Elongation in Arabidopsis Thaliana. Genes (Basel) 2021, 12, 249. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, Y.; Chang, J.; Zheng, F.; Pei, H.; Yi, Y.; Chang, C.; Dong, C.-H. Regulatory Function of Arabidopsis Lipid Transfer Protein 1 (LTP1) in Ethylene Response and Signaling. Plant Mol Biol 2016, 91, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, J.; Valdivia, E.R.; Fraga, P.; Iglesias, N.; Revilla, G.; Zarra, I. Soluble and Membrane-Bound β-Glucosidases Are Involved in Trimming the Xyloglucan Backbone. Plant Physiol 2017, 173, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Coculo, D.; Lionetti, V. The Plant Invertase/Pectin Methylesterase Inhibitor Superfamily. Front Plant Sci 2022, 13, 863892. [Google Scholar] [CrossRef]
- Kong, Z.; Ioki, M.; Braybrook, S.; Li, S.; Ye, Z.-H.; Julie Lee, Y.-R.; Hotta, T.; Chang, A.; Tian, J.; Wang, G.; et al. Kinesin-4 Functions in Vesicular Transport on Cortical Microtubules and Regulates Cell Wall Mechanics during Cell Elongation in Plants. Mol Plant 2015, 8, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Heyndrickx, K.S.; Vandepoele, K. Systematic Identification of Functional Plant Modules through the Integration of Complementary Data Sources. Plant Physiol 2012, 159, 884–901. [Google Scholar] [CrossRef] [PubMed]
- Dangarh, P.; Pandey, N.; Vinod, P.K. Modeling the Control of Meiotic Cell Divisions: Entry, Progression, and Exit. Biophys J 2020, 119, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, T.J.; Deeks, M.J.; Wang, P.; Hussey, P.J. The Evolution of the Actin Binding NET Superfamily. Front Plant Sci 2014, 5, 254. [Google Scholar] [CrossRef]
- Qian, J.; Chen, Y.; Xu, Y.; Zhang, X.; Kang, Z.; Jiao, J.; Zhao, J. Interactional Similarities and Differences in the Protein Complex of PCNA and DNA Replication Factor C between Rice and Arabidopsis. BMC Plant Biol 2019, 19, 257. [Google Scholar] [CrossRef]
- Pfluger, J.; Wagner, D. Histone Modifications and Dynamic Regulation of Genome Accessibility in Plants. Curr Opin Plant Biol 2007, 10, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Breuer, C.; Stacey, N.J.; West, C.E.; Zhao, Y.; Chory, J.; Tsukaya, H.; Azumi, Y.; Maxwell, A.; Roberts, K.; Sugimoto-Shirasu, K. BIN4, a Novel Component of the Plant DNA Topoisomerase VI Complex, Is Required for Endoreduplication in Arabidopsis. Plant Cell 2007, 19, 3655–3668. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.A.; Gubler, F. The Arabidopsis GAMYB-like Genes, MYB33 and MYB65, Are microRNA-Regulated Genes That Redundantly Facilitate Anther Development. Plant Cell 2005, 17, 705–721. [Google Scholar] [CrossRef]
- Cao, X.; Yang, K.-Z.; Xia, C.; Zhang, X.-Q.; Chen, L.-Q.; Ye, D. Characterization of DUF724 Gene Family in Arabidopsis Thaliana. Plant Mol Biol 2010, 72, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.M.; Masiero, S.; Nobre, M.S.; Costa, M.L.; Solís, M.-T.; Testillano, P.S.; Sprunck, S.; Coimbra, S. Differential Expression Patterns of Arabinogalactan Proteins in Arabidopsis Thaliana Reproductive Tissues. J Exp Bot 2014, 65, 5459–5471. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Wang, X.; Lyu, M.; Siligato, R.; Eswaran, G.; Vainio, L.; Blomster, T.; Zhang, J.; Mähönen, A.P. Cytokinins Initiate Secondary Growth in the Arabidopsis Root through a Set of LBD Genes. Curr Biol 2021, 31, 3365–3373. [Google Scholar] [CrossRef]
- Dong, L.; Wang, Z.; Liu, J.; Wang, X. AtSK11 and AtSK12 Mediate the Mild Osmotic Stress-Induced Root Growth Response in Arabidopsis. Int J Mol Sci 2020, 21, 3991. [Google Scholar] [CrossRef]
- Johnson, K.L.; Kibble, N.A.J.; Bacic, A.; Schultz, C.J. A Fasciclin-like Arabinogalactan-Protein (FLA) Mutant of Arabidopsis Thaliana, Fla1, Shows Defects in Shoot Regeneration. PLoS One 2011, 6, e25154. [Google Scholar] [CrossRef]
- Mantegazza, O.; Gregis, V.; Mendes, M.A.; Morandini, P.; Alves-Ferreira, M.; Patreze, C.M.; Nardeli, S.M.; Kater, M.M.; Colombo, L. Analysis of the Arabidopsis REM Gene Family Predicts Functions during Flower Development. Ann Bot 2014, 114, 1507–1515. [Google Scholar] [CrossRef]
- Shimotohno, A.; Heidstra, R.; Blilou, I.; Scheres, B. Root Stem Cell Niche Organizer Specification by Molecular Convergence of PLETHORA and SCARECROW Transcription Factor Modules. Genes Dev 2018, 32, 1085–1100. [Google Scholar] [CrossRef]
- Chung, K.; Sakamoto, S.; Mitsuda, N.; Suzuki, K.; Ohme-Takagi, M.; Fujiwara, S. WUSCHEL-RELATED HOMEOBOX 2 Is a Transcriptional Repressor Involved in Lateral Organ Formation and Separation in Arabidopsis. Plant Biotechnol (Tokyo) 2016, 33, 245–253. [Google Scholar] [CrossRef]
- Zlobin, N.; Lebedeva, M.; Monakhova, Y.; Ustinova, V.; Taranov, V. An ERF121 Transcription Factor from Brassica Oleracea Is a Target for the Conserved TAL-effectors from Different Xanthomonas Campestris Pv. Campestris Strains. Mol Plant Pathol 2021, 22, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Horák, J.; Brzobohatý, B.; Lexa, M. Molecular and Physiological Characterisation of an Insertion Mutant in the ARR21 Putative Response Regulator Gene from Arabidopsis Thaliana. Plant Biol. 2003, 5, 245–254. [Google Scholar] [CrossRef]
- Cui, H.; Kong, D.; Liu, X.; Hao, Y. SCARECROW, SCR-LIKE 23 and SHORT-ROOT Control Bundle Sheath Cell Fate and Function in Arabidopsis Thaliana. Plant J 2014, 78, 319–327. [Google Scholar] [CrossRef]
- Lakehal, A.; Dob, A.; Rahneshan, Z.; Novák, O.; Escamez, S.; Alallaq, S.; Strnad, M.; Tuominen, H.; Bellini, C. ETHYLENE RESPONSE FACTOR 115 Integrates Jasmonate and Cytokinin Signaling Machineries to Repress Adventitious Rooting in Arabidopsis. New Phytol 2020, 228, 1611–1626. [Google Scholar] [CrossRef]
- Jorgensen, R.A.; Dorantes-Acosta, A.E. Conserved Peptide Upstream Open Reading Frames Are Associated with Regulatory Genes in Angiosperms. Front Plant Sci 2012, 3, 191. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Beers, E.P. Alternative Splicing of Myb-Related Genes MYR1 and MYR2 May Modulate Activities through Changes in Dimerization, Localization, or Protein Folding. Plant Signal Behav 2013, 8, e27325. [Google Scholar] [CrossRef]
- Sun, L.; Song, L.; Zhang, Y.; Zheng, Z.; Liu, D. Arabidopsis PHL2 and PHR1 Act Redundantly as the Key Components of the Central Regulatory System Controlling Transcriptional Responses to Phosphate Starvation1. Plant Physiol 2016, 170, 499–514. [Google Scholar] [CrossRef]
- Sun, Z.; Guo, T.; Liu, Y.; Liu, Q.; Fang, Y. The Roles of Arabidopsis CDF2 in Transcriptional and Posttranscriptional Regulation of Primary MicroRNAs. PLoS Genet 2015, 11, e1005598. [Google Scholar] [CrossRef]
- Zubo, Y.O.; Blakley, I.C.; Yamburenko, M.V.; Worthen, J.M.; Street, I.H.; Franco-Zorrilla, J.M.; Zhang, W.; Hill, K.; Raines, T.; Solano, R.; et al. Cytokinin Induces Genome-Wide Binding of the Type-B Response Regulator ARR10 to Regulate Growth and Development in Arabidopsis. Proc Natl Acad Sci U S A 2017, 114, E5995–E6004. [Google Scholar] [CrossRef]
- Lilay, G.H.; Castro, P.H.; Campilho, A.; Assunção, A.G.L. The Arabidopsis bZIP19 and bZIP23 Activity Requires Zinc Deficiency - Insight on Regulation From Complementation Lines. Front Plant Sci 2018, 9, 1955. [Google Scholar] [CrossRef]
- Fang, Q.; Wang, Q.; Mao, H.; Xu, J.; Wang, Y.; Hu, H.; He, S.; Tu, J.; Cheng, C.; Tian, G.; et al. AtDIV2, an R-R-Type MYB Transcription Factor of Arabidopsis, Negatively Regulates Salt Stress by Modulating ABA Signaling. Plant Cell Rep 2018, 37, 1499–1511. [Google Scholar] [CrossRef]
- Steffen, J.G.; Kang, I.-H.; Portereiko, M.F.; Lloyd, A.; Drews, G.N. AGL61 Interacts with AGL80 and Is Required for Central Cell Development in Arabidopsis. Plant Physiol 2008, 148, 259–268. [Google Scholar] [CrossRef]
- Verelst, W.; Twell, D.; de Folter, S.; Immink, R.; Saedler, H.; Münster, T. MADS-Complexes Regulate Transcriptome Dynamics during Pollen Maturation. Genome Biol 2007, 8, R249. [Google Scholar] [CrossRef]
- Gao, J.; Wang, T.; Liu, M.; Liu, J.; Zhang, Z. Transcriptome Analysis of Filling Stage Seeds among Three Buckwheat Species with Emphasis on Rutin Accumulation. PLoS One 2017, 12, e0189672. [Google Scholar] [CrossRef]
- Nakamura, M.; Katsumata, H.; Abe, M.; Yabe, N.; Komeda, Y.; Yamamoto, K.T.; Takahashi, T. Characterization of the Class IV Homeodomain-Leucine Zipper Gene Family in Arabidopsis. Plant Physiol 2006, 141, 1363–1375. [Google Scholar] [CrossRef]
- Zhang, Z.-L.; Ogawa, M.; Fleet, C.M.; Zentella, R.; Hu, J.; Heo, J.-O.; Lim, J.; Kamiya, Y.; Yamaguchi, S.; Sun, T. Scarecrow-like 3 Promotes Gibberellin Signaling by Antagonizing Master Growth Repressor DELLA in Arabidopsis. Proc Natl Acad Sci U S A 2011, 108, 2160–2165. [Google Scholar] [CrossRef]
- Torti, S.; Fornara, F. AGL24 Acts in Concert with SOC1 and FUL during Arabidopsis Floral Transition. Plant Signal Behav 2012, 7, 1251–1254. [Google Scholar] [CrossRef]
- Bao, M.; Bian, H.; Zha, Y.; Li, F.; Sun, Y.; Bai, B.; Chen, Z.; Wang, J.; Zhu, M.; Han, N. miR396a-Mediated Basic Helix-Loop-Helix Transcription Factor bHLH74 Repression Acts as a Regulator for Root Growth in Arabidopsis Seedlings. Plant Cell Physiol 2014, 55, 1343–1353. [Google Scholar] [CrossRef]
- Horstman, A.; Willemsen, V.; Boutilier, K.; Heidstra, R. AINTEGUMENTA-LIKE Proteins: Hubs in a Plethora of Networks. Trends Plant Sci 2014, 19, 146–157. [Google Scholar] [CrossRef]
- Shibata, M.; Breuer, C.; Kawamura, A.; Clark, N.M.; Rymen, B.; Braidwood, L.; Morohashi, K.; Busch, W.; Benfey, P.N.; Sozzani, R.; et al. GTL1 and DF1 Regulate Root Hair Growth through Transcriptional Repression of ROOT HAIR DEFECTIVE 6-LIKE 4 in Arabidopsis. Development 2018, 145, dev159707. [Google Scholar] [CrossRef]
- Hidayati, N.A.; Yamada-Oshima, Y.; Iwai, M.; Yamano, T.; Kajikawa, M.; Sakurai, N.; Suda, K.; Sesoko, K.; Hori, K.; Obayashi, T.; et al. Lipid Remodeling Regulator 1 (LRL1) Is Differently Involved in the Phosphorus-depletion Response from PSR1 in Chlamydomonas Reinhardtii. Plant J 2019, 100, 610–626. [Google Scholar] [CrossRef]
- Lu, W.; Deng, F.; Jia, J.; Chen, X.; Li, J.; Wen, Q.; Li, T.; Meng, Y.; Shan, W. The Arabidopsis Thaliana Gene AtERF019 Negatively Regulates Plant Resistance to Phytophthora Parasitica by Suppressing PAMP-triggered Immunity. Mol Plant Pathol 2020, 21, 1179–1193. [Google Scholar] [CrossRef]
- Li, X.; Cheng, Y.; Wang, M.; Cui, S.; Guan, J. Weighted Gene Coexpression Correlation Network Analysis Reveals a Potential Molecular Regulatory Mechanism of Anthocyanin Accumulation under Different Storage Temperatures in “Friar” Plum. BMC Plant Biol 2021, 21, 576. [Google Scholar] [CrossRef]
- Cutcliffe, J.W.; Hellmann, E.; Heyl, A.; Rashotte, A.M. CRFs Form Protein-Protein Interactions with Each Other and with Members of the Cytokinin Signalling Pathway in Arabidopsis via the CRF Domain. J Exp Bot 2011, 62, 4995–5002. [Google Scholar] [CrossRef]
- Zhang, S.; Xie, M.; Ren, G.; Yu, B. CDC5, a DNA Binding Protein, Positively Regulates Posttranscriptional Processing and/or Transcription of Primary microRNA Transcripts. Proc Natl Acad Sci U S A 2013, 110, 17588–17593. [Google Scholar] [CrossRef]
- Plesch, G.; Störmann, K.; Torres, J.T.; Walden, R.; Somssich, I.E. Developmental and Auxin-Induced Expression of the Arabidopsis Prha Homeobox Gene. Plant J 1997, 12, 635–647. [Google Scholar] [CrossRef]
- Liu, Z.-W.; Shao, C.-R.; Zhang, C.-J.; Zhou, J.-X.; Zhang, S.-W.; Li, L.; Chen, S.; Huang, H.-W.; Cai, T.; He, X.-J. The SET Domain Proteins SUVH2 and SUVH9 Are Required for Pol V Occupancy at RNA-Directed DNA Methylation Loci. PLoS Genet 2014, 10, e1003948. [Google Scholar] [CrossRef]
- Martinez-Garcia, M.; White, C.I.; Franklin, F.C.H.; Sanchez-Moran, E. The Role of Topoisomerase II in DNA Repair and Recombination in Arabidopsis Thaliana. Int J Mol Sci 2021, 22, 13115. [Google Scholar] [CrossRef]
- Lyu, G.; Li, D.; Li, S. Bioinformatics Analysis of BBX Family Genes and Its Response to UV-B in Arabidopsis Thaliana. Plant Signal Behav 2020, 15, 1782647. [Google Scholar] [CrossRef]
- Nibau, C.; Gibbs, D.J.; Bunting, K.A.; Moody, L.A.; Smiles, E.J.; Tubby, J.A.; Bradshaw, S.J.; Coates, J.C. ARABIDILLO Proteins Have a Novel and Conserved Domain Structure Important for the Regulation of Their Stability. Plant Mol Biol 2011, 75, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Araya, T.; von Wirén, N.; Takahashi, H. CLE Peptides Regulate Lateral Root Development in Response to Nitrogen Nutritional Status of Plants. Plant Signal Behav 2014, 9, e29302. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, C.; Hepworth, S.R.; Ma, C.; Li, H.; Li, J.; Wang, S.-M.; Yin, H. SAUR15 Interaction with BRI1 Activates Plasma Membrane H+-ATPase to Promote Organ Development of Arabidopsis. Plant Physiol 2022, 189, 2454–2466. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-E.; Kim, Y.-S.; Yoon, H.-K.; Park, C.-M. Functional Characterization of a Small Auxin-up RNA Gene in Apical Hook Development in Arabidopsis. Plant Science 2007, 172, 150–157. [Google Scholar] [CrossRef]
- Qiu, T.; Chen, Y.; Li, M.; Kong, Y.; Zhu, Y.; Han, N.; Bian, H.; Zhu, M.; Wang, J. The Tissue-Specific and Developmentally Regulated Expression Patterns of the SAUR41 Subfamily of Small Auxin up RNA Genes: Potential Implications. Plant Signal Behav 2013, 8, e25283. [Google Scholar] [CrossRef]
- Sakai, M.; Sakamoto, T.; Saito, T.; Matsuoka, M.; Tanaka, H.; Kobayashi, M. Expression of Novel Rice Gibberellin 2-Oxidase Gene Is under Homeostatic Regulation by Biologically Active Gibberellins. J Plant Res 2003, 116, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.; Horák, J.; Chaban, C.; Mira-Rodado, V.; Witthöft, J.; Elgass, K.; Grefen, C.; Cheung, M.-K.; Meixner, A.J.; Hooley, R.; et al. The Histidine Kinase AHK5 Integrates Endogenous and Environmental Signals in Arabidopsis Guard Cells. PLoS One 2008, 3, e2491. [Google Scholar] [CrossRef] [PubMed]
- Schott-Verdugo, S.; Müller, L.; Classen, E.; Gohlke, H.; Groth, G. Structural Model of the ETR1 Ethylene Receptor Transmembrane Sensor Domain. Sci Rep 2019, 9, 8869. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Furlan, C.; Campos, R.; Toth, J.N.; Van Norman, J.M. Distinct Mechanisms Orchestrate the Contra-Polarity of IRK and KOIN, Two LRR-Receptor-Kinases Controlling Root Cell Division. Nat Commun 2022, 13, 235. [Google Scholar] [CrossRef]
- Diepold, A.; Li, G.; Lennarz, W.J.; Nürnberger, T.; Brunner, F. The Arabidopsis AtPNG1 Gene Encodes a Peptide: N-Glycanase. Plant J 2007, 52, 94–104. [Google Scholar] [CrossRef]
- Ronceret, A.; Guilleminot, J.; Lincker, F.; Gadea-Vacas, J.; Delorme, V.; Bechtold, N.; Pelletier, G.; Delseny, M.; Chabouté, M.-E.; Devic, M. Genetic Analysis of Two Arabidopsis DNA Polymerase Epsilon Subunits during Early Embryogenesis. Plant J 2005, 44, 223–236. [Google Scholar] [CrossRef]
- Vatamaniuk, O.K.; Mari, S.; Lang, A.; Chalasani, S.; Demkiv, L.O.; Rea, P.A. Phytochelatin Synthase, a Dipeptidyltransferase That Undergoes Multisite Acylation with Gamma-Glutamylcysteine during Catalysis: Stoichiometric and Site-Directed Mutagenic Analysis of Arabidopsis Thaliana PCS1-Catalyzed Phytochelatin Synthesis. J Biol Chem 2004, 279, 22449–22460. [Google Scholar] [CrossRef]
- Abrahams, S.; Cavet, G.; Oakenfull, E.A.; Carmichael, J.P.; Shah, Z.H.; Soni, R.; Murray, J.A. A Novel and Highly Divergent Arabidopsis Cyclin Isolated by Complementation in Budding Yeast. Biochim Biophys Acta 2001, 1539, 1–6. [Google Scholar] [CrossRef]
- d’Erfurth, I.; Jolivet, S.; Froger, N.; Catrice, O.; Novatchkova, M.; Simon, M.; Jenczewski, E.; Mercier, R. Mutations in AtPS1 (Arabidopsis Thaliana Parallel Spindle 1) Lead to the Production of Diploid Pollen Grains. PLoS Genet 2008, 4, e1000274. [Google Scholar] [CrossRef]
- Van Damme, D.; De Rybel, B.; Gudesblat, G.; Demidov, D.; Grunewald, W.; De Smet, I.; Houben, A.; Beeckman, T.; Russinova, E. Arabidopsis α Aurora Kinases Function in Formative Cell Division Plane Orientation. Plant Cell 2011, 23, 4013–4024. [Google Scholar] [CrossRef]
- Romanowski, A.; Furniss, J.J.; Hussain, E.; Halliday, K.J. Phytochrome Regulates Cellular Response Plasticity and the Basic Molecular Machinery of Leaf Development. Plant Physiol 2021, 186, 1220–1239. [Google Scholar] [CrossRef]
- Draeger, C.; Ndinyanka Fabrice, T.; Gineau, E.; Mouille, G.; Kuhn, B.M.; Moller, I.; Abdou, M.-T.; Frey, B.; Pauly, M.; Bacic, A.; et al. Arabidopsis Leucine-Rich Repeat Extensin (LRX) Proteins Modify Cell Wall Composition and Influence Plant Growth. BMC Plant Biol 2015, 15, 155. [Google Scholar] [CrossRef]
- Farquharson, K.L. Mirror, Mirror on the Wall: A Role for AGP18 in Functional Megaspore Selection. Plant Cell 2013, 25, 1190. [Google Scholar] [CrossRef]
- Li, S.; Ge, F.-R.; Xu, M.; Zhao, X.-Y.; Huang, G.-Q.; Zhou, L.-Z.; Wang, J.-G.; Kombrink, A.; McCormick, S.; Zhang, X.S.; et al. Arabidopsis COBRA-LIKE 10, a GPI-Anchored Protein, Mediates Directional Growth of Pollen Tubes. Plant J 2013, 74, 486–497. [Google Scholar] [CrossRef]
- Dharmasiri, N.; Dharmasiri, S.; Weijers, D.; Lechner, E.; Yamada, M.; Hobbie, L.; Ehrismann, J.S.; Jürgens, G.; Estelle, M. Plant Development Is Regulated by a Family of Auxin Receptor F Box Proteins. Dev Cell 2005, 9, 109–119. [Google Scholar] [CrossRef]
- Cho, E.J.; Choi, S.H.; Kim, J.H.; Kim, J.E.; Lee, M.H.; Chung, B.Y.; Woo, H.R.; Kim, J.-H. A Mutation in Plant-Specific SWI2/SNF2-Like Chromatin-Remodeling Proteins, DRD1 and DDM1, Delays Leaf Senescence in Arabidopsis Thaliana. PLoS One 2016, 11, e0146826. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.S.; Khadka, J.; Domb, K.; Zemach, A.; Grafi, G. CMT3 and SUVH4/KYP Silence the Exonic Evelknievel Retroelement to Allow for Reconstitution of CMT1 mRNA. Epigenetics Chromatin 2018, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Fernández, R.; Barrero-Sicilia, C.; Carrillo-Barral, N.; Oñate-Sánchez, L.; Carbonero, P. Arabidopsis Thaliana bZIP44: A Transcription Factor Affecting Seed Germination and Expression of the Mannanase-Encoding Gene AtMAN7. Plant J 2013, 74, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Hagen, G.; Guilfoyle, T. Auxin-Responsive Gene Expression: Genes, Promoters and Regulatory Factors. Plant Mol Biol 2002, 49, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, L.; De Clercq, I.; Narsai, R.; Xu, Y.; Hartmann, A.; Claros, D.L.; Custovic, E.; Lewsey, M.G.; Whelan, J.; et al. ANAC017 Coordinates Organellar Functions and Stress Responses by Reprogramming Retrograde Signaling. Plant Physiol 2019, 180, 634–653. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-A.; Hoai, T.N.T.; Park, H.-J.; Zhao, M.; Twell, D.; Honys, D.; Park, S.-K. MYB81, a Microspore-Specific GAMYB Transcription Factor, Promotes Pollen Mitosis I and Cell Lineage Formation in Arabidopsis. Plant J 2020, 101, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, G.; Li, C.; Zhang, C.; Cui, L.; Ai, G.; Wang, X.; Zheng, F.; Zhang, D.; Larkin, R.M.; et al. NF-Y Plays Essential Roles in Flavonoid Biosynthesis by Modulating Histone Modifications in Tomato. New Phytol 2021, 229, 3237–3252. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, X.; Yang, B.; Xu, S.; Wei, X.; Zhao, P.; Niu, F.; Sun, M.; Wang, C.; Cheng, H.; et al. WRKY55 Transcription Factor Positively Regulates Leaf Senescence and the Defense Response by Modulating the Transcription of Genes Implicated in the Biosynthesis of Reactive Oxygen Species and Salicylic Acid in Arabidopsis. Development 2020, 147, dev189647. [Google Scholar] [CrossRef] [PubMed]
- Seyfferth, C.; Wessels, B.; Jokipii-Lukkari, S.; Sundberg, B.; Delhomme, N.; Felten, J.; Tuominen, H. Ethylene-Related Gene Expression Networks in Wood Formation. Front Plant Sci 2018, 9, 272. [Google Scholar] [CrossRef]
- Paul, P.; Simm, S.; Mirus, O.; Scharf, K.-D.; Fragkostefanakis, S.; Schleiff, E. The Complexity of Vesicle Transport Factors in Plants Examined by Orthology Search. PLoS One 2014, 9, e97745. [Google Scholar] [CrossRef]
- Radoeva, T.; Lokerse, A.S.; Llavata-Peris, C.I.; Wendrich, J.R.; Xiang, D.; Liao, C.-Y.; Vlaar, L.; Boekschoten, M.; Hooiveld, G.; Datla, R.; et al. A Robust Auxin Response Network Controls Embryo and Suspensor Development through a Basic Helix Loop Helix Transcriptional Module[OPEN]. Plant Cell 2019, 31, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Wolff, P.; Weinhofer, I.; Seguin, J.; Roszak, P.; Beisel, C.; Donoghue, M.T.A.; Spillane, C.; Nordborg, M.; Rehmsmeier, M.; Köhler, C. High-Resolution Analysis of Parent-of-Origin Allelic Expression in the Arabidopsis Endosperm. PLoS Genet 2011, 7, e1002126. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Beltran, E.; Elander, P.H.; Dalman, K.; Dayhoff, G.W.; Moschou, P.N.; Uversky, V.N.; Crespo, J.L.; Bozhkov, P.V. Tudor Staphylococcal Nuclease Is a Docking Platform for Stress Granule Components and Is Essential for SnRK1 Activation in Arabidopsis. EMBO J 2021, 40, e105043. [Google Scholar] [CrossRef]
- Domínguez-Figueroa, J.; Carrillo, L.; Renau-Morata, B.; Yang, L.; Molina, R.-V.; Marino, D.; Canales, J.; Weih, M.; Vicente-Carbajosa, J.; Nebauer, S.G.; et al. The Arabidopsis Transcription Factor CDF3 Is Involved in Nitrogen Responses and Improves Nitrogen Use Efficiency in Tomato. Front Plant Sci 2020, 11, 601558. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Sotomayor, P.; Chávez Montes, R.A.; Silvestre-Vañó, M.; Herrera-Ubaldo, H.; Greco, R.; Pablo-Villa, J.; Galliani, B.M.; Diaz-Ramirez, D.; Weemen, M.; Boutilier, K.; et al. Altered Expression of the bZIP Transcription Factor DRINK ME Affects Growth and Reproductive Development in Arabidopsis Thaliana. Plant J 2016, 88, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Mouchel, C.F.; Briggs, G.C.; Hardtke, C.S. Natural Genetic Variation in Arabidopsis Identifies BREVIS RADIX, a Novel Regulator of Cell Proliferation and Elongation in the Root. Genes Dev 2004, 18, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Showalter, A.M.; Keppler, B.; Lichtenberg, J.; Gu, D.; Welch, L.R. A Bioinformatics Approach to the Identification, Classification, and Analysis of Hydroxyproline-Rich Glycoproteins. Plant Physiol 2010, 153, 485–513. [Google Scholar] [CrossRef]
- Choi, J.; Strickler, S.R.; Richards, E.J. Loss of CRWN Nuclear Proteins Induces Cell Death and Salicylic Acid Defense Signaling. Plant Physiol 2019, 179, 1315–1329. [Google Scholar] [CrossRef]
- Xun, Q.; Wu, Y.; Li, H.; Chang, J.; Ou, Y.; He, K.; Gou, X.; Tax, F.E.; Li, J. Two Receptor-like Protein Kinases, MUSTACHES and MUSTACHES-LIKE, Regulate Lateral Root Development in Arabidopsis Thaliana. New Phytol 2020, 227, 1157–1173. [Google Scholar] [CrossRef]
- Sun, Y.; Qiao, Z.; Muchero, W.; Chen, J.-G. Lectin Receptor-Like Kinases: The Sensor and Mediator at the Plant Cell Surface. Front Plant Sci 2020, 11, 596301. [Google Scholar] [CrossRef]
- Stührwohldt, N.; Ehinger, A.; Thellmann, K.; Schaller, A. Processing and Formation of Bioactive CLE40 Peptide Are Controlled by Posttranslational Proline Hydroxylation. Plant Physiol 2020, 184, 1573–1584. [Google Scholar] [CrossRef]
- Kosarev, P.; Mayer, K.F.X.; Hardtke, C.S. Evaluation and Classification of RING-Finger Domains Encoded by the Arabidopsis Genome. Genome Biol 2002, 3, RESEARCH0016. [Google Scholar] [CrossRef] [PubMed]
- D’Ippólito, S.; Arias, L.A.; Casalongué, C.A.; Pagnussat, G.C.; Fiol, D.F. The DC1-Domain Protein VACUOLELESS GAMETOPHYTES Is Essential for Development of Female and Male Gametophytes in Arabidopsis. Plant J 2017, 90, 261–275. [Google Scholar] [CrossRef]
- Giuntoli, B.; Lee, S.C.; Licausi, F.; Kosmacz, M.; Oosumi, T.; van Dongen, J.T.; Bailey-Serres, J.; Perata, P. A Trihelix DNA Binding Protein Counterbalances Hypoxia-Responsive Transcriptional Activation in Arabidopsis. PLoS Biol 2014, 12, e1001950. [Google Scholar] [CrossRef]
Figure 1.
Spontaneous tumour formation in inbred radish lines: (a). taproots of related lines 19 (left) and 18 (right) contrasting in the tumour formation trait; (b). a family tree of the radish genetic collection showing the origin of the inbred lines; tumour lines 10, 11, 12, 13, 14, 16, 19, 20, 21, 32, 34 are marked in red; the squares indicate the intended progeny of each radish lineage. Different boxes represent lines of diverse cultivars. The sector that includes lines originating from the Saxa cultivar is highlighted in green.
Figure 1.
Spontaneous tumour formation in inbred radish lines: (a). taproots of related lines 19 (left) and 18 (right) contrasting in the tumour formation trait; (b). a family tree of the radish genetic collection showing the origin of the inbred lines; tumour lines 10, 11, 12, 13, 14, 16, 19, 20, 21, 32, 34 are marked in red; the squares indicate the intended progeny of each radish lineage. Different boxes represent lines of diverse cultivars. The sector that includes lines originating from the Saxa cultivar is highlighted in green.
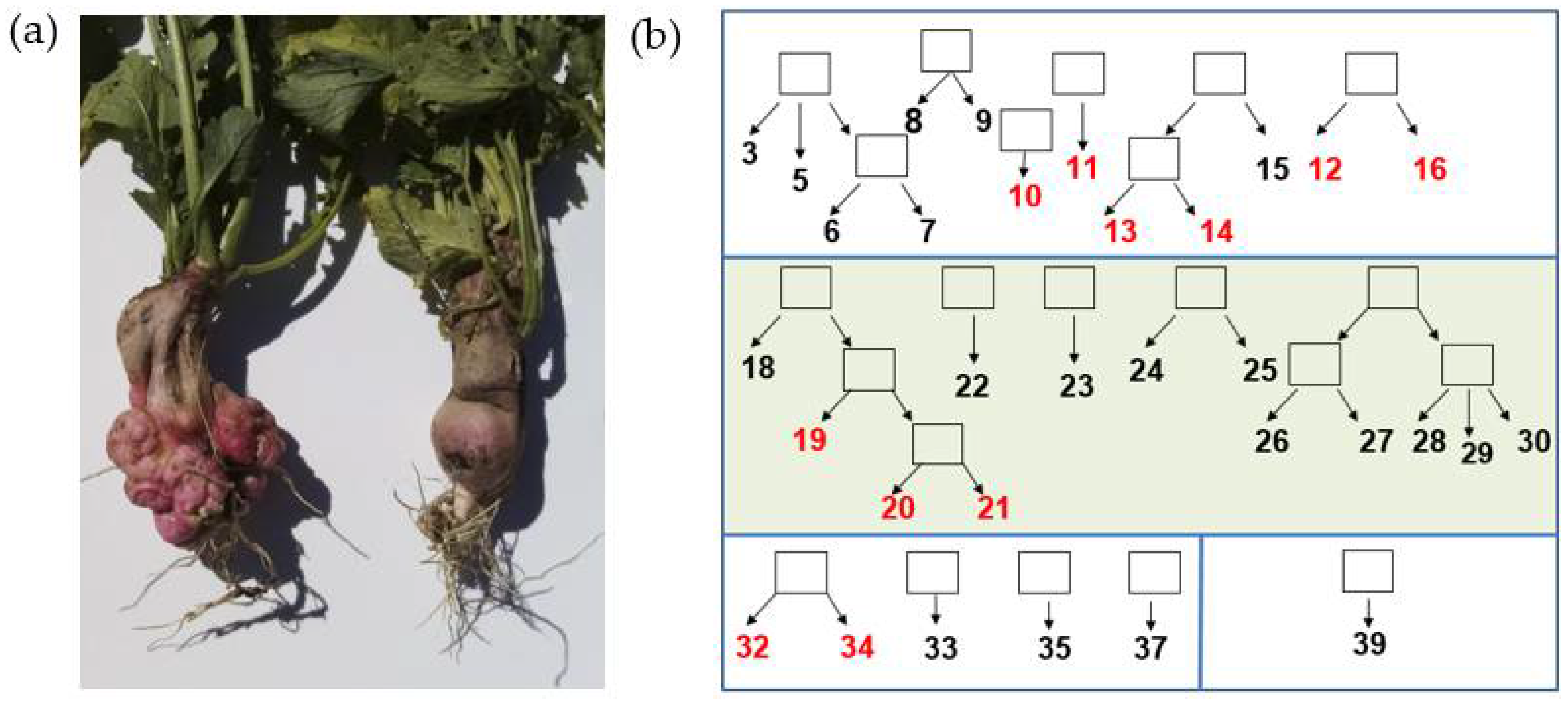
Figure 2.
Comparative characteristics of the genomes of radish lines 18 and 19 sequenced in this work and radish reference genome (GCA_019705955.1). The analysis was carried out using the BUSCO programme.
Figure 2.
Comparative characteristics of the genomes of radish lines 18 and 19 sequenced in this work and radish reference genome (GCA_019705955.1). The analysis was carried out using the BUSCO programme.
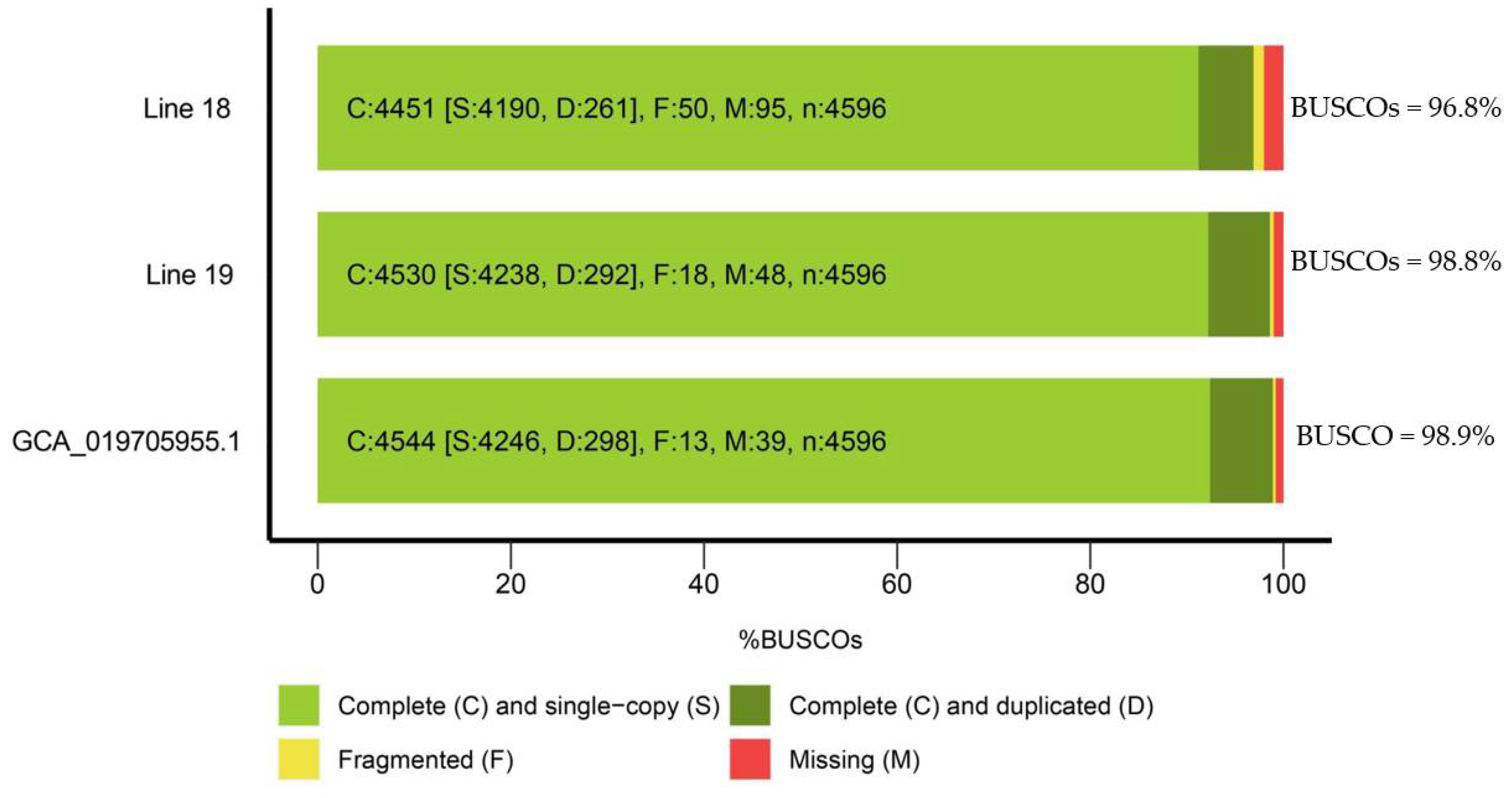
Figure 3.
Chromosomal location of radish genes with (a). InDels or (b). SNPs identified in the tumour line 19 compared to the non-tumour line 18 performed using the MapChart 2.32 software (https://www.wur.nl/en/show/mapchart.htm).
Figure 3.
Chromosomal location of radish genes with (a). InDels or (b). SNPs identified in the tumour line 19 compared to the non-tumour line 18 performed using the MapChart 2.32 software (https://www.wur.nl/en/show/mapchart.htm).
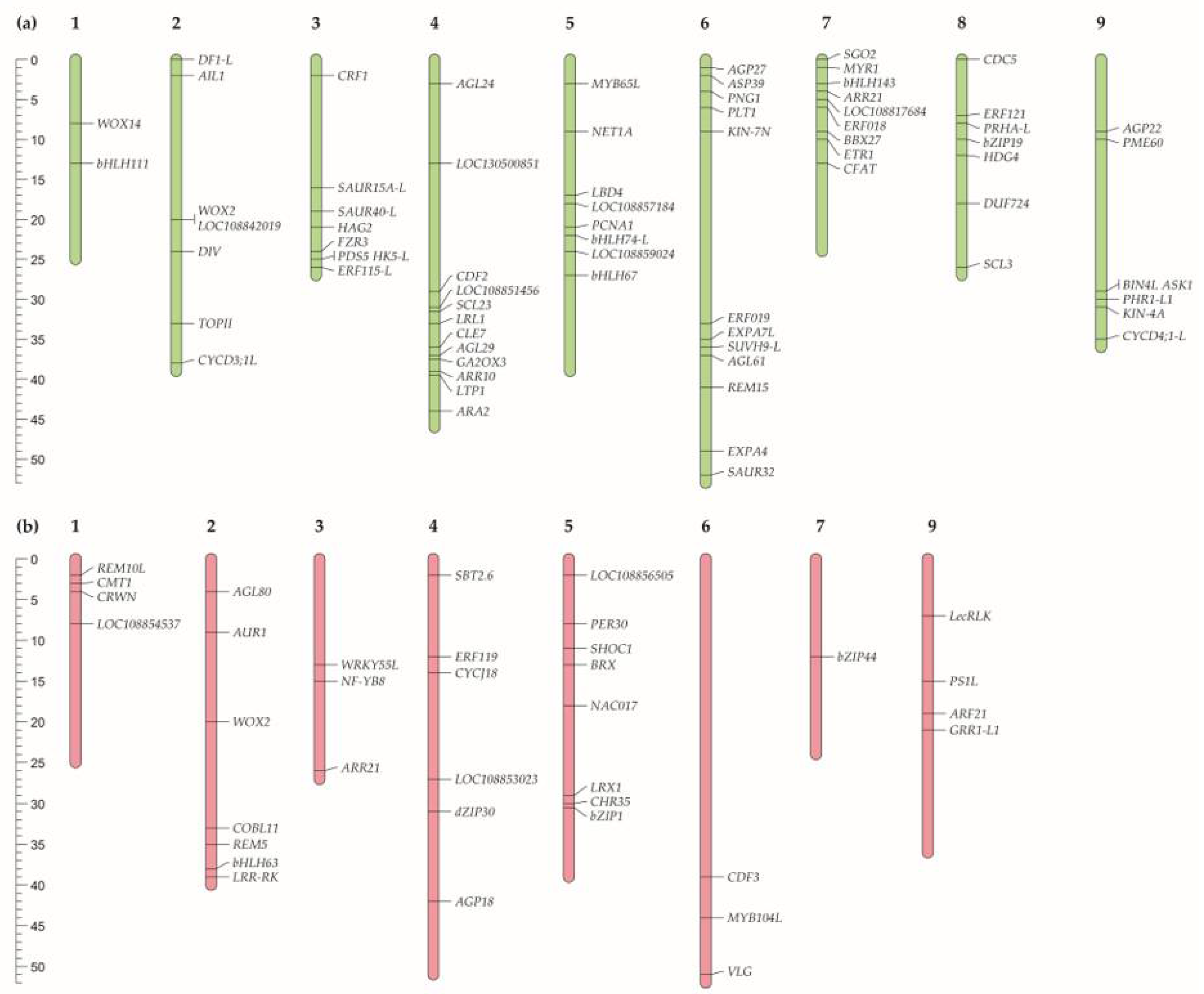
Figure 4.
Schematic representation of the insertion (marked by an asterisk), which was detected in the RsERF018 gene. a). The scheme of a ERF18 gene. The insertion is located in the border of the 5’-UTR and the start codon. b). 5‘UTR insertion of the RsERF018 gene in radish lines and its possible consequences. The amino acid content of the protein synthesised during translation of the normal sequence is marked in black, and the protein synthesised during translation in the case of CAG insertion is marked in white. Radish tumour lines are highlighted in red. .
Figure 4.
Schematic representation of the insertion (marked by an asterisk), which was detected in the RsERF018 gene. a). The scheme of a ERF18 gene. The insertion is located in the border of the 5’-UTR and the start codon. b). 5‘UTR insertion of the RsERF018 gene in radish lines and its possible consequences. The amino acid content of the protein synthesised during translation of the normal sequence is marked in black, and the protein synthesised during translation in the case of CAG insertion is marked in white. Radish tumour lines are highlighted in red. .

Figure 5.
Chromosomal location of radish (a) CLE and (b) WOX family genes performed using the MapChart 2.32 software (https://www.wur.nl/en/show/mapchart.htm).
Figure 5.
Chromosomal location of radish (a) CLE and (b) WOX family genes performed using the MapChart 2.32 software (https://www.wur.nl/en/show/mapchart.htm).
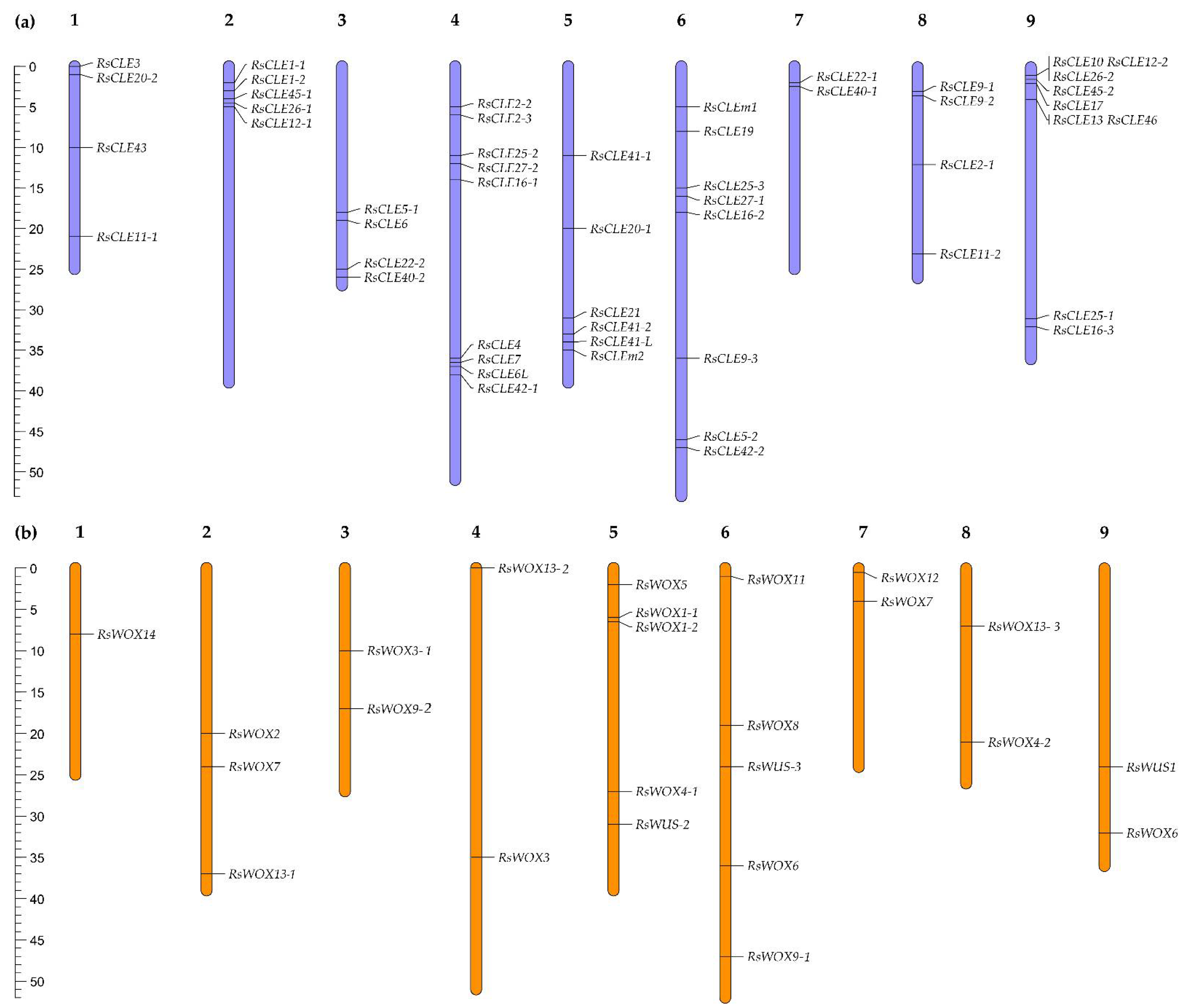
Figure 6.
Radish CLE genes (RsCLEs) family. (a). Phylogenetic tree of RsCLE genes constructed using the Neighbour-joining algorithm. The colour indicates RsCLEm1 and RsCLEm2 genes which encode proteins with multiple CLE domains; (b). CLE domain consensus sequences of all RsCLE peptides identified in radish.
Figure 6.
Radish CLE genes (RsCLEs) family. (a). Phylogenetic tree of RsCLE genes constructed using the Neighbour-joining algorithm. The colour indicates RsCLEm1 and RsCLEm2 genes which encode proteins with multiple CLE domains; (b). CLE domain consensus sequences of all RsCLE peptides identified in radish.
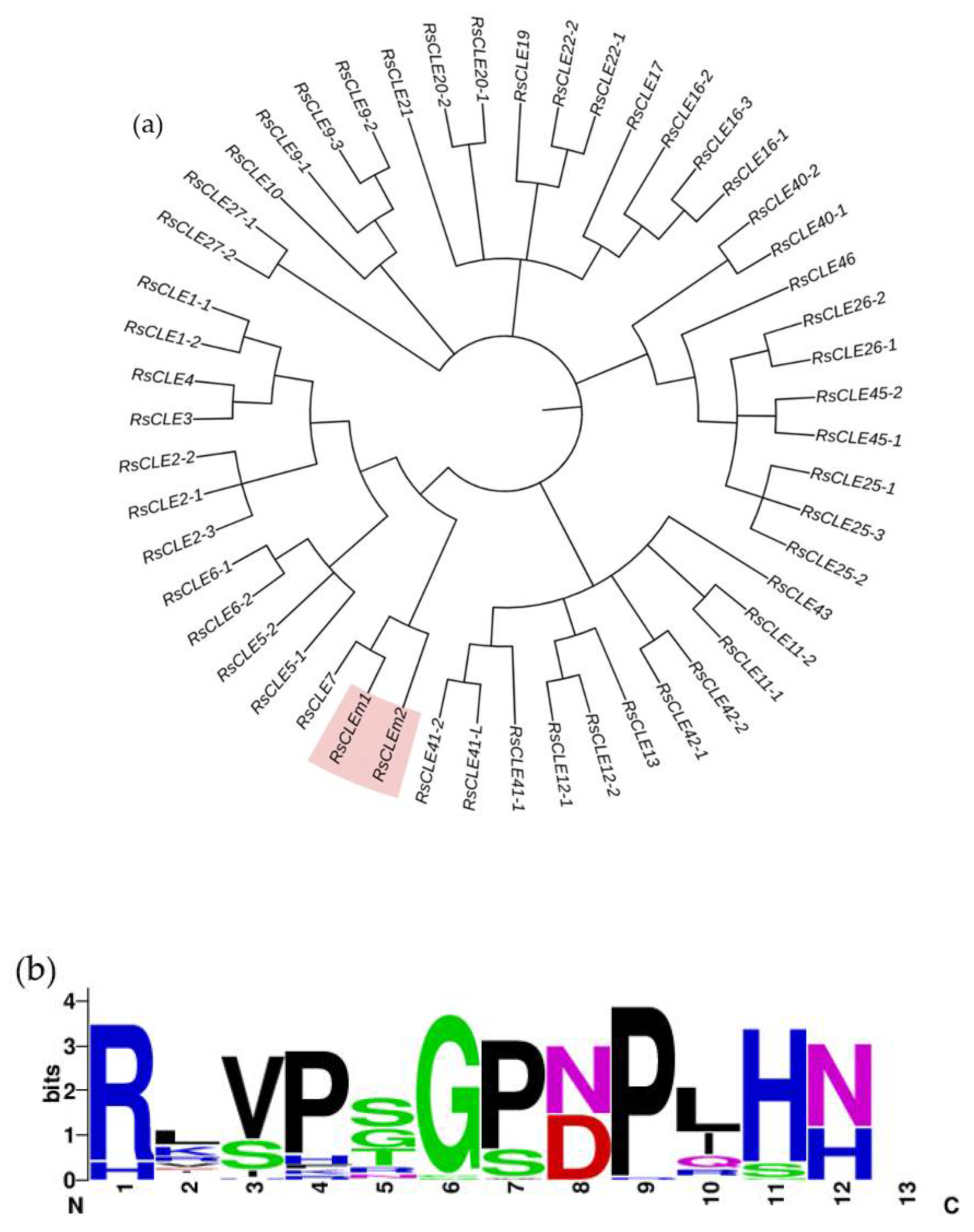
Figure 7.
The proteins with multiple CLE domains probably encoded by RsCLEm genes. (a) A representation of the domain organization of RsCLEm proteins, including the positions of signal peptide (SP) and CLE domains. Signal motifs were predicted with the SignalP-6.0 tool (https://services.healthtech.dtu.dk/service.php?SignalP). Identical sequences of CLE domains are marked with the same colour. (b) CLE domain consensus sequences of Raphanus sativus and Brassica napus. (c) Sequence alignment of the putative 12-amino acid CLE domain sequences encoded by CLEm genes of Raphanus sativus and Brassica napus. (d) Phylogenetic analysis of BnCLEm and RsCLEm peptides.
Figure 7.
The proteins with multiple CLE domains probably encoded by RsCLEm genes. (a) A representation of the domain organization of RsCLEm proteins, including the positions of signal peptide (SP) and CLE domains. Signal motifs were predicted with the SignalP-6.0 tool (https://services.healthtech.dtu.dk/service.php?SignalP). Identical sequences of CLE domains are marked with the same colour. (b) CLE domain consensus sequences of Raphanus sativus and Brassica napus. (c) Sequence alignment of the putative 12-amino acid CLE domain sequences encoded by CLEm genes of Raphanus sativus and Brassica napus. (d) Phylogenetic analysis of BnCLEm and RsCLEm peptides.
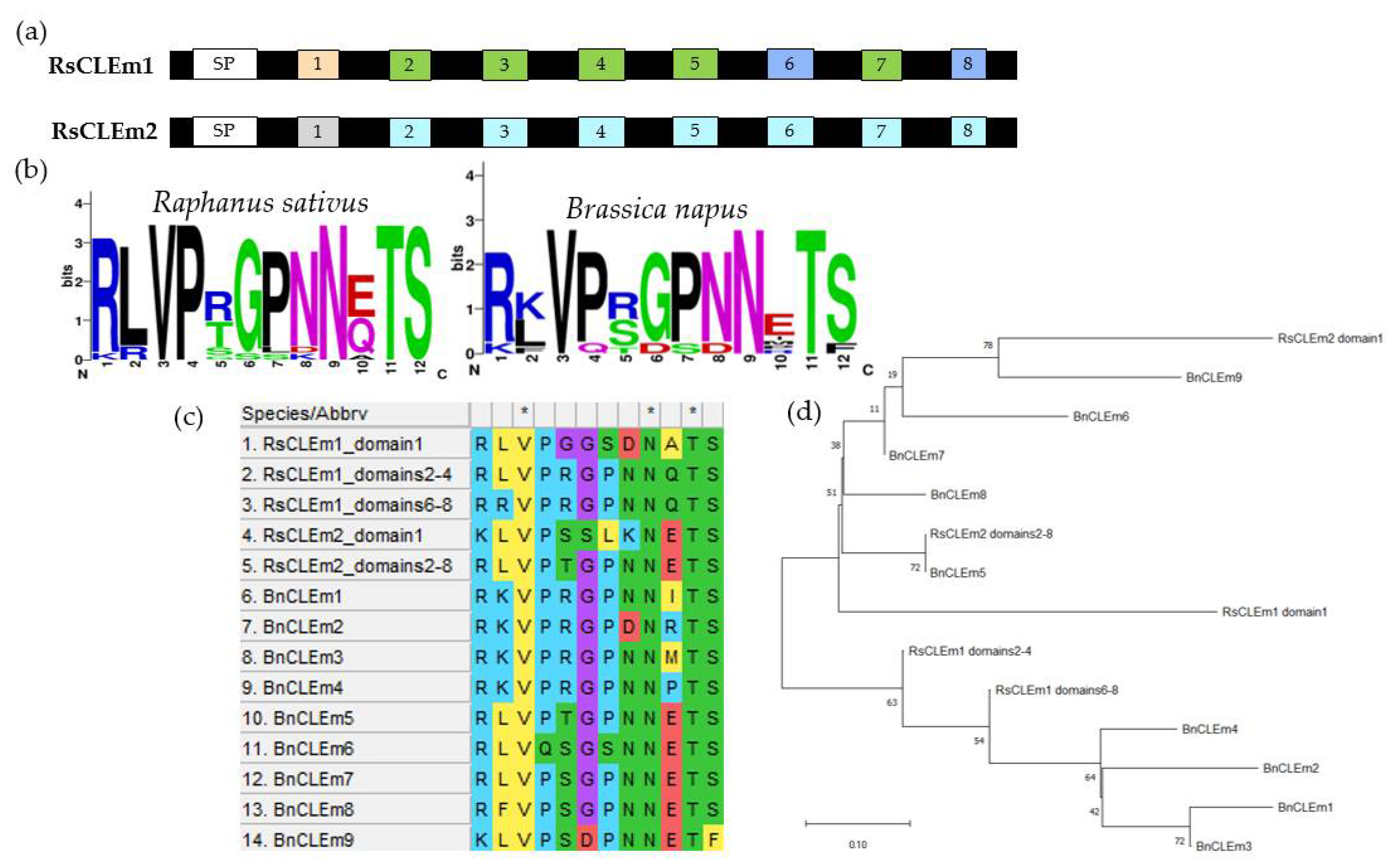

Table 1.
Number of SNVs (InDels, SNPs) identified in tumour radish line and their probable impacts on gene structure (high, low, moderate, modifier). For SNPs, there is information on their number in different functional classes (missense, nonsense, silent). Data were obtained with SnpEff tool.
Table 1.
Number of SNVs (InDels, SNPs) identified in tumour radish line and their probable impacts on gene structure (high, low, moderate, modifier). For SNPs, there is information on their number in different functional classes (missense, nonsense, silent). Data were obtained with SnpEff tool.
| Variant | Impact | Functional class | |||
|---|---|---|---|---|---|
| Type | Total | Type | Number | Type | Number |
| SNP | 2260270 | HIGH | 9451 | MISSENSE | 292963 |
| LOW | 432159 | NONSENSE | 5618 | ||
| MODERATE | 291254 | SILENT | 393274 | ||
| MODIFIER | 4334386 | ||||
| INDEL | 514083 | HIGH | 12234 | ||
| LOW | 15755 | ||||
| MODERATE | 15497 | ||||
| MODIFIER | 1227445 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



