
Submitted:
07 May 2024
Posted:
08 May 2024
You are already at the latest version
Abstract
Abstract: (1) Background: We determined the prevalence of hepatitis B virus (HBV) amongst people without human immunodeficiency virus (HIV) in rural and peri-urban areas in Botswana. (2) Methods: We screened hepatitis B surface antigen (HBsAg) from archived plasma samples of people without HIV (n=2135) randomly selected from the Botswana Combination Prevention Program (BCPP) (2013–2018). We sequenced 415 bp of the surface region using BigDye sequencing chemistry. (3) Results: The median age of participants was 31 (IQR: 24–46) and 64% (1360/2135) were female. HBsAg prevalence was 4.0% (86/2135) [95% CI: 3.3 – 4.9]) and ranged between 0%–9.2%. Older participants (>35 years) had increased odds of HBsAg positivity (OR: 1.94; 95% CI: [1.32–2.86]; p = 0.001. Thirteen samples were sequenced and 7 (53.8%) were genotype A, 3 (23.1%) were genotype D and genotype E each. Clinically significant mutations were identified in the surface region, but no classic drug resistance mutations were identified. (4) Conclusions: We report an HBV prevalence of 4.0% (95% CI 3.3 – 4.9) among people without HIV in rural and pe-ri-urban communities in Botswana with varying rates in different communities. A comprehen-sive national HBV program is required in Botswana to guide HBV prevention, testing and management.
Keywords:
Hepatitis B Virus
; HBV
; prevalence
; genotypes
; mutations
; HIV negative
; Africa
1. Introduction
Approximately 254 million people worldwide are infected with chronic hepatitis B virus infections (CHB), with most cases prevalent in the Western Pacific region and sub-Saharan Africa, where 96.8 million and 64.7 million people are affected, respectively[1]. In Botswana, hepatitis B surface antigen (HBsAg) prevalence ranging between 1.1% – 10.6% has been reported in blood donors, pregnant women and mostly in people with human immunodeficiency virus (PWH) [2,3,4,5,6,7,8]. Most of these studies were in urban areas except for the largest HBV study in PWH in the country which was conducted in rural and peri-urban areas [4]. This study reported an overall prevalence of 8% with varying prevalence rates between communities some differing by as much as 10-fold (2.1%-22.1%) [4]. Several studies reported similar HBV prevalence rates in PWH and those without, while others reported differences [7,9,10].
HBV is classified into 10 genotypes (A-I) and a putative genotype J based on a nucleotide divergency of >7.5% with more than 35 subgenotypes [11,12,13]. Genotypes differ by geographic distribution and by disease progression [13,14,15]. In Africa including in Botswana genotypes A, D and E have been reported [14,16,17]. Subgenotype A1 predominantly found in Africa has been linked with increased chronicity and faster progression to hepatocellular carcinoma [14,15].
One of the barriers to elimination especially in Africa is the lack of national data hence most of national HBV prevalence rates have been extrapolated from the little data available within the region [10,18,19]. In Botswana there is no prevalence data of HBV at the population level among people without HIV therefore, we aimed to establish the HBV prevalence amongst people without HIV across rural and peri-urban areas in Botswana.
2. Materials and Methods
2.1. Study Population
Archived plasma samples from people without HIV who were enrolled in the Botswana Combination Prevention Project (BCPP) were used. BCPP was a pair-matched, cluster-randomized trial conducted in 30 communities (15 pairs matched according to size, pre-existing health services, population age structure, and geographical location, including proximity to urban areas) around Botswana from Oct 30, 2013, to Nov 24, 2018 [20]. BCPP enrolled 12610 participants aged 16-64 years, of whom 9014 were without HIV [21]. A total of 2135 samples from people without HIV were randomly selected for characterization in this study ensuring proportional distributions amongst the sites. Out of 30 BCPP communities, 2 were not sampled (Otse and Molapowabojang) because there were stored samples available. Samples were stored at -80°C prior to testing.
2.2. Ethical Approval
Approval for the study was sought from the Institute of Advanced Medical Advanced Research and Training (IAMRAT) approval number UI/EC/22/0001 and the Human Research Development Committee (HRDC) at the Botswana Ministry of Health (MoH), HRDC number 01028. Participants provided written informed consent for their samples to be used in the parent study as well as subsequent studies.
2.3. Serological Assays
2.3.1. Validation of the HBsAg Dilution Potocol
Briefly, archived plasma samples from people with HIV (PWH) from the Botswana Combination Prevention Project (BCPP) were used. Twenty-four HBsAg positive samples with known optical densities (ODs) were used. The samples were divided into three categories: high positive (OD >4.9, n=6), medium positives (OD range 2.0-4.9, n= 9) and low positives (OD range 0-1.9, n= 9) based on their ODs. Serial dilutions of 1:10, 1:100, 1:1000 and 1:10 0000 of sample: phosphate buffered saline (PBS) were performed. HBsAg was then screened in the samples with the various dilutions using the Murex HBsAg Version 3 enzyme-linked immunosorbent assay (ELISA) kit (Murex Biotech, Dartford, UK) following manufacturer’s instructions. The results showed that all high and medium positives remain positive from the 1:10 to the 1:10 000 dilutions. The low positives were all positive at the 1:10 dilution, however (4/9) 44% of low positives lost HBsAg at the 1:100 dilution and an additional (3/9) 33% lost the HBsAg positivity by the 1:1000 dilution. These results show that regardless of the OD, 10-1 dilution remains positive.
2.3.2. HBsAg Screening
Plasma samples had insufficient volumes, so they were diluted using the previously validated in-house protocol shown above. Briefly, prior to HBsAg screening, samples were diluted using PBS at a 10-fold dilution which detects 100% of samples with positive HBsAg serology (HBsAg+) including those with low optical densities. After dilution, the samples were screened for HBsAg using the Murex HBsAg Version 3 enzyme-linked immunosorbent assay (ELISA) kit (Murex Biotech, Dartford, UK). HBsAg+ samples were subsequently repeated for confirmation following the manufacturer’s guidelines.
2.4. Amplification and Sequencing
Total nucleic acid was extracted from 200uL of HBsAg+ plasma samples diluted with PBS at 10-fold using an inhouse validated protocol. We used the QIAamp DNA Blood (QIAGEN, Hilden, Germany) following the manufacturer’s instructions with an elution volume of 30μL [22]. A 415 bp fragment of the surface/polymerase gene was amplified by semi-nested PCR with two rounds of amplification using Platinum Taq DNA Polymerase High Fidelity kit according to the manufacturer (Invitrogen, Waltham, USA), using primers and thermal cycling conditions as described previously [17,23]. Sequencing was done using big dye sequencing chemistry. Briefly, the master mix for sequencing PCR consisted of 3µl Sequencing Buffer (5X), 1µl BigDye ®Terminator v3.1, 0.2 µl of 2 µM Primer, 1µl purified PCR product, and 4.8µl in a 10µl reaction mix. The cycling conditions were 5 minutes denaturing at 94°C, and then 30 cycles of denaturing for 45 seconds at 94°C, annealing for 30 seconds at 50 °C, and elongation at 72 °C for 90 seconds, with extension at 72 °C for 10 minutes using HBV840 (5′-GTTTAAATGTATACCCAAAGAC-3′;nt840-861) and HBV381 (5′-TGCGGCGTTTTATCATCTTCCT-3′; nt381-402 ) primers for first round. Second round cycling conditions were denaturation at 94°C for 5 minutes and then 30 cycles of denaturation at 94°C for 45 seconds, annealing at 55°C for 30 seconds, elongation at 72°C for 60 seconds, and extension at 72°C for 10 minutes using HBV381 and HBV801 (5′-CAGCGGCATAAAGGGACTCAAG-3′ nt801-822;) primers. Sequencing reactions were setup using big dye sequencing chemistry and two primers, HBV 381 and HBV 801. ZR DNA Sequencing Clean-up Kit (Zymo, Irvine, CA, USA) was used for sequencing clean-up according to the manufacturer’s protocol. Sanger sequencing was performed on an ABI 3130xl genetic analyzer (Applied Biosystems, Foster City, CA).
2.5. Data Analyses
Manual editing of sequences was carried out using Sequencher v5.0 software (Gene Codes Corp., Ann Arbor, MI, USA). Consensus sequences were generated from the overlapping sequences. Next, consensus sequences generated were viewed, trimmed and aligned with previously generated Botswana reference sequences from GenBank using AliView alignment viewer [24]. Sequences were then uploaded to the Geno2Pheno online database (http://www.geno2Pheno.org) for genotypic, drug resistance and escape mutations analyses. Sequences were further uploaded to a second online database, the Stanford HBVseq database, (http://hivdb.stanford.edu/HBV/HBVseq/development/HBVseq.html) to confirm genotype and drug resistance mutations. We constructed a maximum-likelihood tree using the best fitting model of nucleotide substitution [TVMe+R2] using IQTREE with 1000 bootstrap replicates [25].
2.6. Statistical Analysis
Predictors associated with HBsAg positivity were determined using both univariate and multivariable logistic regression, adjusting for clustering by community. Stata version 14.0 (Stata Corp, College Station, TX, USA) was used for statistical analysis and p-values<0.05 were considered statistically significant.
3. Results
3.1. HBV Prevalence
The median age of all participants was 31 (IQR: 24 – 46) and of these, 63.7% (1360/2135) [95% CI: 61.6 – 65.7] were female. We report an overall HBsAg prevalence of 4.0% (86/2135) [95% CI: 3.3 – 4.9]. Univariate logistic regression analysis shows that participants 35 years and older have increased odds of HBsAg positivity (OR: 1.94; 95% CI: [1.32–2.86]; p = 0.001) (Table 1, supplementary Figure S1). We observed reduced odds of HBsAg positivity among people with secondary education (OR: 0.37; 95% CI: [0.17–0.81]; p = 0.012) and those with higher education levels (OR: 0.35; 95% CI: [0.14–0.88 p = 0.025) compared to those with non-formal education. There was a trend towards more inconsistent condom use in the HBsAg+ group compared to the HBsAg- group. There were no significant independent predictors of HBsAg as shown in the multivariate logistic regression in Table 1.
3.2. HBV Genetic Diversity
Out of the 86 HBsAg+ samples, n= 13 (15%) were amplified and all of them (100%) were successfully sequenced. The online databases and the phylogenetic analysis revealed that HBV genotype A, n=7 (54%) was found to be the most predominant circulating HBV genotype while the remaining 6 sequences were equally shared between genotypes D (23%) and E (23%). The 13 newly generated sequences were used to construct a maximum-likelihood tree and the available online sequences from Botswana were used to confirm genotypes and determine clustering patterns of these sequences with reference sequences. The tree shows that the sequences generated from our study clustered with reference sequences from Botswana (Figure 2).
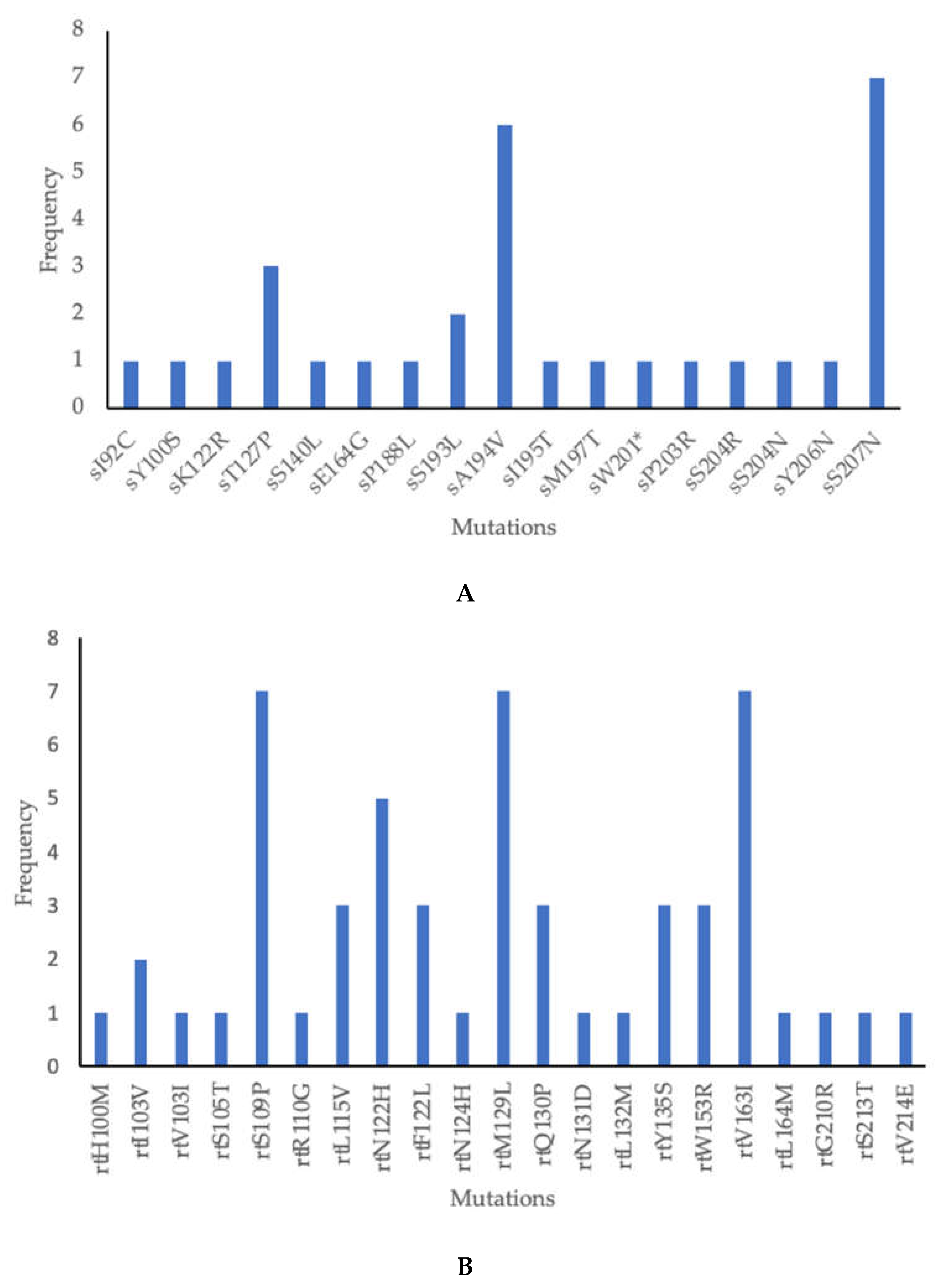
As depicted in Table 2 below, 38 mutations were found in 12/13 isolates (92.3%) showing amino acid substitution in the HBV surface (s) gene and the overlapping reverse transcriptase (rt) region. Amongst these, 17 were in the s region, 21 in the rt region. Only two mutations were novel. Clinically significant mutations included escape mutations as well as those associated with disease progression and OBI were identified in this study (Table 3). The most common mutations in the s region were sS207N and sA194V found in 7 (54%) and 6 (46%) participants respectively, Figure 3A. The most common mutations in the rt region were rtS109P, rtM129L, and rtV163I found in 7 (54%) each. These 3 mutations appeared together in 6 participants, Figure 3B.
4. Discussion
To our knowledge, the current study is the largest study to report HBV prevalence in people without HIV in rural and peri-urban communities in Botswana. The relationship between the prevalence of HBV and participant socioeconomic demographics was explored. We report a 4.0% HBV prevalence which is higher than approximately 1% previously reported in pregnant women without HIV in Botswana and South Africa but similar to a serosurvey in South Africa [3,10,42]. The differences in these prevalence rates might be due to differences in geographic regions, cultural practices, access to healthcare, vaccination coverage as the study on pregnant women was conducted in urban areas while the current study was in peri-urban and rural areas. Differences in HBV prevalence rates even in the same country have been reported before including in Botswana [4,43]. Notably, this prevalence is lower than that reported in PWH in the same communities which was 8% overall [4]. Higher rates of HBV prevalence in PWH compared with people without HIV have been reported before in South Africa [10] while some studies including a multicenter study which included participants from Botswana, South Africa, Kenya, Malawi, India, Thailand and Brazil reported similar rates [3,7,44]. The differences might be due to differences in risk factors found in different places as well as the level of immunosuppression in the HIV infected groups. HIV associated immunosuppression has been associated with HBV reactivation resulting in a higher HBsAg positivity [45,46].
In this study, there were varying HBV prevalence rates between communities, with the highest prevalence recorded in Nata (9.2%) while some places such as Tati, Tsetsebjwe, Mmadinare and Mathangwane reported no HBV cases. Differences in rates between communities have been reported before including in PWH in the same communities [4]. Interestingly the HBV prevalence spread is different between PWH and people without HIV in these communities [4]. For example, Nata had the highest HBV prevalence in people without HIV whereas it had the 9th highest prevalence out of 30 communities in a study on HBV prevalence in PWH. In our current study, Tati and Tsetsebjwe also reported no cases of HBV in people without HIV but had the 3rd and 12th highest HBV prevalence in the same study of PWH respectively [4]. The differences between the places might be due to cultural practices. It is also possible that the current study randomly selected HBsAg negative participants as only a subset of samples was screened for HBV.
Older age and secondary education were predictors of HBsAg positivity in univariate analysis but there were no predictors of HBsAg positivity in multivariate analysis. Other studies showed no association with education but showed an association with age [10,45]. The South African study also did not show relationship status as well as the number of lifetime partners as predictors of HBsAg positivity similar to the current study [10]. Most studies reported more HBV in males ,however, for the current study, sex was not a predictor of HBsAg positivity [4,45,47,48]. In a study in PWH in the same communities, the male sex and northern geographic region were independent predictors of HBsAg positivity unlike in the current study. In the said study with PWH being older was only a predictor in univariate analysis similar to the current study [4]. Predictors of HBsAg vary across studies which might be due to lifestyle practices, genotypes as well as host genetics [10,48].
The distribution of the genotypes across peri-urban and rural areas around Botswana will aid in furthering our understanding of the prevalent HBV genotypes amongst people without HIV. Similar to the existing literature on the HBV genotypes circulating in sub-Saharan Africa, the HBV genotypes A (54%), D (23%) and E (23%) were also found to be prevalent in Botswana [16,22]. Genotypes A, D and E from this study clustered together with sequences of known HBV genotypes from Botswana suggesting local transmission. Information on the distribution of HBV genotypes across rural and peri-urban areas in Botswana is limited.
This study identified mutations, some of which were of clinical significance. In the surface region sA194V and sS207N were the predominant mutations as reported in a study from Kenya [49]. A194V has not been characterized but it has been reported in many studies while sS207N is linked to immune escape [39,40,41]. Other mutations identified which were associated with immune escape were sK122R, sT127P and sS140L [29,30,31,32,33,34]. Other mutations with clinical significance were sY100S, sK122R, sM197T, sW201*, sS204R, and sS204N which impair the production and secretion of HBsAg, associated with low HBV viral load and OBI [26,27,28,29,30,35,36,38]. In the polymerase region, the most common mutations were rtS109P, rtM129L and rtV163I found in 7 participants each similar to other studies which reported these mutations in genotype A and indeed these mutations were present in genotype A in this study [50,51,52]. These 3 mutations appeared together in 6 genotype A participants, rtS109P was also found in genotype E. We report no classic drug resistance-associated mutations such as rtL80I/V, rtV173L, rtL180M, rtA181S, rtA194T, rtS202I, rtM204V/I, rtN236T and rtM250L/V associated with resistance against lamivudine, telbivudine, adefovir, entecavir and tenofovir in this study similar to other studies [22,52,53]. The rtY135S mutation was mostly seen in treatment non-responders and some studies classified it under drug resistance mutations against lamivudine, telbivudine, adefovir and entecavir but its characterization is not yet concluded [54,55,56,57]. This contrasts with a study in PWH in the same communities which reported a high prevalence of drug resistance mutations against lamivudine [58]. These results might suggest that there were no drug resistance mutations transmitted between PWH and those without HIV. We identified uncharacterized mutations which have been reported in other studies (sE164G, sP188L, sS193L, sA194V, sI195T, sY206N), rtI103V, rtV103I, rtS105T, rtS109P, rtR110G, rtL115V (only found in genotype D), rtN122H, rtF122L, rtN124H, rtQ130P, rtN131D, rtL132M, rtW153R, rtV163I, rtL164M, rtS213T, rtG210R, and rtV214E) [22,50,51,55,59,60,61,62,63,64,65,66,67,68,69,70,71]. We also reported novel mutations in this study, sI92C and rtH100M all found in. genotype E.
This study bears the strength of being the largest study on HBV in people without HIV in understudied rural and peri-urban areas in Botswana. The limitations of the study include insufficient sample volumes which necessitated the use of a dilution protocol which however can be applied in future studies. The effects of insufficient sample volumes were that we could not test for more HBV biomarkers and diluting the samples might have affected the sequencing success, which was low in our study. The community level HBV prevalence should be taken with caution as the study was not powered to determine the prevalence at community level.
5. Conclusions
We report an HBV prevalence of 4.0% among people without HIV in rural and peri-urban communities in Botswana with varying rates in different communities and the presence of clinically relevant mutations. Therefore, integration of general care, management, and advocacy in HBV regardless of HIV status is urgently needed in Botswana to accelerate reductions in the prevalence of HBV among the general population. Surveillance of clinically relevant mutations is recommended to guide prevention strategies. Future studies screening for further HBV biomarkers are needed to further characterize HBV in people without HIV in Botswana.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, MA, SG; Data curation, TM, BBP, GM, WTC, BP, LNB, DD, KB, LMD, LO, TR; Formal analysis, MA, TM, BBP, GM, WTC, BP, TR, SM, SG; Funding acquisition, MA; Methodology, MA, BBP, GM, WTC, BP, LNB, KB, SM, SG Resources, COA, SM, SG; Supervision, MA, COA, SG; Validation, MA, BBP, GM, WTC, SM, SG, Writing – original draft, MA, TM; Writing – review &; editing, MA, TM, BBP, GM, COA, WTC, BP, LNB, DD, KB, LMD, LO, TR, SM, SG. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Wellcome Trust (grant number 218770/Z/19/Z) and Bill & Melinda Gates Foundation (INV-033558). B. B. P. and S. G. are supported by the National Institutes of Health (NIH) Common Fund, award number U41HG006941 (H3ABioNet). H3ABioNet is an initiative of the Human Health and Heredity in Africa Consortium (H3Africa) program of the African Academy of Science. B. B. P., R. M., and S. M. are also supported by Trials of Excellence in Southern Africa (TESAIII), which is part of the EDCTP2 program supported by the European Union (grant number CSA2020NoE-3104 TESAIII). S. L., R. S. and S.M. received support from the NIH (award numbers K24 AI131928, K24 AI131924 and K43 TW012350, respectively). W. T. C., S. M., and S. G. are partly supported through the Sub-Saharan African Network for TB/HIV Research Excellence (SANTHE 2.0) from the Bill & Melinda Gates Foundation (INV-033558). S. G. and B.B.P are supported by the Fogarty International Center at the US National Institutes of Health (D43 TW009610). S.G and W.T.C are supported partly by NIH (award number 1G11TW012503-01). The views and opinions in this manuscript are solely those of the authors and do not represent the official position of the funding agencies. The funders had no role in the design, conduct of the study and decision to publish the results.
Data Availability Statement
The datasets [generated/analyzed] for this study are available upon request.
Acknowledgments
The authors thank the Botswana Prevention Combination Project study participants, Dikgosi and other community leaders, the clinic staff, District Health Management Teams, and Community Health Facilities at study sites; the ‘Ya Tsie’ Study Team at the Botswana Harvard Health Partnership, the Harvard T. H. Chan School of Public Health, the Centers for Disease Control and Prevention (CDC) Botswana, CDC Atlanta, and the Botswana Ministry of Health. The authors also thank those who served on the Ya Tsie Community Advisory Board, Laboratory Staff, and Management of Botswana Harvard HIV Reference Laboratory.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- WHO, Global hepatitis report 2024: action for access in low- and middle-income countries. 2024, World Health Organization: Geneva.
- Anderson, M., et al., Slow CD4(+) T-Cell Recovery in Human Immunodeficiency Virus/Hepatitis B Virus-Coinfected Patients Initiating Truvada-Based Combination Antiretroviral Therapy in Botswana. Open Forum Infect Dis, 2016. 3(3): p. ofw140.
- Mbangiwa, T., et al., Chronic and Occult Hepatitis B Virus Infection in Pregnant Women in Botswana. Genes (Basel), 2018. 9(5).
- Phinius, B.B., et al., High Prevalence of Hepatitis B Virus Infection Among People With HIV in Rural and Periurban Communities in Botswana. Open Forum Infect Dis, 2023. 10(1): p. ofac707.
- Wester, C.W., et al., Serological evidence of HIV-associated infection among HIV-1-infected adults in Botswana. Clin Infect Dis, 2006. 43(12): p. 1612-5.
- Patel, P., et al., Prevalence of hepatitis B and hepatitis C coinfections in an adult HIV centre population in Gaborone, Botswana. Am J Trop Med Hyg, 2011. 85(2): p. 390-4.
- Greer, A.E., et al., Comparison of Hepatitis B Virus Infection in HIV-Infected and HIV-Uninfected Participants Enrolled in a Multinational Clinical Trial: HPTN 052. J Acquir Immune Defic Syndr, 2017. 76(4): p. 388-393.
- Choga, W.T., et al., Molecular characterization of hepatitis B virus in blood donors in Botswana. Virus Genes, 2018.
- Adewumi, M.O., et al., HBV Infection Among HIV-Infected Cohort and HIV-Negative Hospital Attendees in South Western Nigeria. Afr J Infect Dis, 2015. 9(1): p. 14-7.
- Samsunder, N., et al., Seroprevalence of hepatitis B virus: Findings from a population-based household survey in KwaZulu-Natal, South Africa. Int J Infect Dis, 2019. 85: p. 150-157.
- Kramvis, A., Genotypes and genetic variability of hepatitis B virus. Intervirology, 2014. 57(3-4): p. 141-50.
- Kramvis, A., M. Kew, and G. Francois, Hepatitis B virus genotypes. Vaccine, 2005. 23(19): p. 2409-23.
- Norder, H., et al., Genetic diversity of hepatitis B virus strains derived worldwide: genotypes, subgenotypes, and HBsAg subtypes. Intervirology, 2004. 47(6): p. 289-309.
- Kramvis, A., Molecular characteristics and clinical relevance of African genotypes and subgenotypes of hepatitis B virus. S Afr Med J, 2018. 108(8b): p. 17-21.
- Kramvis, A., The clinical implications of hepatitis B virus genotypes and HBeAg in pediatrics. Rev Med Virol, 2016. 26(4): p. 285-303.
- Choga, W.T., et al., Molecular characterization of hepatitis B virus in blood donors in Botswana. Virus Genes, 2019. 55(1): p. 33-42.
- Anderson, M., et al., Molecular characterisation of hepatitis B virus in HIV-1 subtype C infected patients in Botswana. BMC Infect Dis, 2015. 15: p. 335.
- Sonderup, M.W. and C.W. Spearman, Global Disparities in Hepatitis B Elimination-A Focus on Africa. Viruses, 2022. 14(1).
- WHO, Global health sector strategy on viral hepatitis 2016–2021. Towards ending viral hepatitis 2016: https://iris.who.int/bitstream/handle/10665/246177/WHO-HIV-2016.06-eng.pdf?sequence=1.
- Makhema, J., et al., Universal Testing, Expanded Treatment, and Incidence of HIV Infection in Botswana. N Engl J Med, 2019. 381(3): p. 230-242.
- Gaolathe, T., et al., Botswana’s progress toward achieving the 2020 UNAIDS 90-90-90 antiretroviral therapy and virological suppression goals: a population-based survey. Lancet HIV, 2016. 3(5): p. e221-30.
- Anderson, M., et al., Molecular Characterization of Near Full-Length Genomes of Hepatitis B Virus Isolated from Predominantly HIV Infected Individuals in Botswana. Genes (Basel), 2018. 9(9).
- Ma, Y., et al., Genotyping the hepatitis B virus with a fragment of the HBV DNA polymerase gene in Shenyang, China. Virol J, 2011. 8: p. 315.
- Larsson, A., AliView: a fast and lightweight alignment viewer and editor for large datasets. Bioinformatics, 2014. 30(22): p. 3276-8.
- Nguyen, L.T., et al., IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol Biol Evol, 2015. 32(1): p. 268-74.
- Svicher, V., et al., Novel HBsAg markers tightly correlate with occult HBV infection and strongly affect HBsAg detection. Antiviral Res, 2012. 93(1): p. 86-93.
- Mello, F.C., et al., Expression of Hepatitis B Virus Surface Antigen Containing Y100C Variant Frequently Detected in Occult HBV Infection. Hepat Res Treat, 2011. 2011: p. 695859.
- Biswas, S., D. Candotti, and J.P. Allain, Specific amino acid substitutions in the S protein prevent its excretion in vitro and may contribute to occult hepatitis B virus infection. J Virol, 2013. 87(14): p. 7882-92.
- Wang, J., et al., The sK122R mutation of hepatitis B virus (HBV) is associated with occult HBV infection: Analysis of a large cohort of Chinese patients. J Clin Virol, 2020. 130: p. 104564.
- Wang, H., et al., Novel hepatitis B virus surface antigen mutations associated with occult genotype B hepatitis B virus infection affect HBsAg detection. J Viral Hepat, 2020. 27(9): p. 915-921.
- Martin, C.M., et al., Mutations associated with occult hepatitis B virus infection result in decreased surface antigen expression in vitro. J Viral Hepat, 2012. 19(10): p. 716-23.
- Luongo, M., et al., Acute hepatitis B caused by a vaccine-escape HBV strain in vaccinated subject: sequence analysis and therapeutic strategy. J Clin Virol, 2015. 62: p. 89-91.
- Ijaz, S., R.B. Ferns, and R.S. Tedder, A ‘first loop’ linear epitope accessible on native hepatitis B surface antigen that persists in the face of ‘second loop’ immune escape. J Gen Virol, 2003. 84(Pt 2): p. 269-275.
- Osasona, O.G., et al., Patterns of hepatitis b virus immune escape and pol/rt mutations across clinical cohorts of patients with genotypes a, e and occult hepatitis b infection in Nigeria: A multi-centre study. Virulence, 2023. 14(1): p. 2218076.
- Mirabelli, C., et al., Specific mutations in the C-terminus domain of HBV surface antigen significantly correlate with low level of serum HBV-DNA in patients with chronic HBV infection. J Infect, 2015. 70(3): p. 288-98.
- Zhou, H., et al., Sequence analysis and functional characterization of full-length hepatitis B virus genomes from Korean cirrhotic patients with or without liver cancer. Virus Res, 2017. 235: p. 86-95.
- Thi Cam Huong, N., et al., Mutations in the HBV PreS/S gene related to hepatocellular carcinoma in Vietnamese chronic HBV-infected patients. PLoS One, 2022. 17(4): p. e0266134.
- Kalinina, T., et al., A dominant hepatitis B virus population defective in virus secretion because of several S-gene mutations from a patient with fulminant hepatitis. Hepatology, 2001. 34(2): p. 385-94.
- Weinberger, K.M., et al., High genetic variability of the group-specific a-determinant of hepatitis B virus surface antigen (HBsAg) and the corresponding fragment of the viral polymerase in chronic virus carriers lacking detectable HBsAg in serum. J Gen Virol, 2000. 81(Pt 5): p. 1165-74.
- Zheng, X., et al., Mutant hepatitis B virus surface antigens (HBsAg) are immunogenic but may have a changed specificity. Virology, 2004. 329(2): p. 454-64.
- Coppola, N., et al., Clinical significance of hepatitis B surface antigen mutants. World J Hepatol, 2015. 7(27): p. 2729-39.
- Joseph Davey, D., et al., Low prevalence of hepatitis B virus infection in HIV-uninfected pregnant women in Cape Town, South Africa: implications for oral pre-exposure prophylaxis roll out. BMC Infect Dis, 2022. 22(1): p. 719.
- Matthews, P.C., et al., HLA-A is a Predictor of Hepatitis B e Antigen Status in HIV-Positive African Adults. J Infect Dis, 2016. 213(8): p. 1248-52.
- Amponsah-Dacosta, E., et al., Evidence for a change in the epidemiology of hepatitis B virus infection after nearly two decades of universal hepatitis B vaccination in South Africa. J Med Virol, 2014. 86(6): p. 918-24.
- Msomi, N., et al., High incidence and persistence of hepatitis B virus infection in individuals receiving HIV care in KwaZulu-Natal, South Africa. BMC Infect Dis, 2020. 20(1): p. 847.
- Phinius, B.B., et al., Incidence of hepatitis B virus infection among human immunodeficiency virus-infected treatment naive adults in Botswana. Medicine (Baltimore), 2020. 99(9): p. e19341.
- Yuen, M.F., et al., Prognostic determinants for chronic hepatitis B in Asians: therapeutic implications. Gut, 2005. 54(11): p. 1610-4.
- Brown, R., P. Goulder, and P.C. Matthews, Sexual Dimorphism in Chronic Hepatitis B Virus (HBV) Infection: Evidence to Inform Elimination Efforts. Wellcome Open Res, 2022. 7: p. 32.
- King, R.E., et al., Identification and Characterization of Hepatitis B Virus Immune Escape Mutants in Kenya. American Journal of Molecular Biology,, 2023. 13(ISSN Online: 2161-6663.
- ISSN Print: 2161-6620): p. 1-17.
- Mathew, A., et al., Hepatitis B virus genotypes and drug resistance mutations circulating in blood donors in Beira, Mozambique. PLoS One, 2023. 18(2): p. e0281855.
- Ismail, A.M., et al., Antiviral resistance mutations and genotype-associated amino acid substitutions in treatment-naive hepatitis B virus-infected individuals from the Indian subcontinent. Intervirology, 2012. 55(1): p. 36-44.
- Koyaweda, G.W., et al., Detection of circulating hepatitis B virus immune escape and polymerase mutants among HBV-positive patients attending Institut Pasteur de Bangui, Central African Republic. Int J Infect Dis, 2020. 90: p. 138-144.
- Pacheco, S.R., et al., Genotyping of HBV and tracking of resistance mutations in treatment-naive patients with chronic hepatitis B. Infect Drug Resist, 2017. 10: p. 201-207.
- Mahmood, M., et al., Molecular Analysis of Hepatitis B Virus Reverse Transcriptase Domain for Mutations Associated with Viral Resistance in Pakistani Patients. Viral Hepat J, 2021. 27(3): p. 153-158.
- Singla, B., et al., Response to potent anti-HBV agents in chronic hepatitis B and combined effect of HBV reverse transcriptase mutations. Gene, 2015. 567(1): p. 22-30.
- Gohar, M., et al., Phylogenetic Analysis and Emerging Drug Resistance against Different Nucleoside Analogues in Hepatitis B Virus Positive Patients. Microorganisms, 2023. 11(11).
- Tan, J., et al., Tenofovir monotherapy is effective in hepatitis B patients with antiviral treatment failure to adefovir in the absence of adefovir-resistant mutations. J Hepatol, 2008. 48(3): p. 391-8.
- Phinius, B.B., et al., High Prevalence of Hepatitis B Virus Drug Resistance Mutations to Lamivudine among People with HIV/HBV Coinfection in Rural and Peri-Urban Communities in Botswana. Viruses, 2024. 16(4).
- Bubonja-Sonje, M., et al., Prevalence of occult hepatitis B virus infection and characterisation of hepatitis B surface antigen mutants among adults in western Croatia. Ann Hepatol, 2024. 29(1): p. 101156.
- Oluyinka, O.O., et al., Occult Hepatitis B Virus Infection in Nigerian Blood Donors and Hepatitis B Virus Transmission Risks. PLoS One, 2015. 10(7): p. e0131912.
- Wang, L., et al., Sequence analysis of the S gene region in HBV DNA from patients positive for both HBsAg and HBsAb tests. Hepatol Res, 2010. 40(12): p. 1212-8.
- Logoida, M., et al., Comparison of Two Diagnostic Methods for the Detection of Hepatitis B Virus Genotypes in the Slovak Republic. Pathogens, 2021. 11(1).
- Hosseini, S.Y., et al., Association of HBsAg mutation patterns with hepatitis B infection outcome: Asymptomatic carriers versus HCC/cirrhotic patients. Ann Hepatol, 2019. 18(4): p. 640-645.
- Kuhns, M.C., et al., Molecular and Serological Characterization of Hepatitis B Virus (HBV)-Positive Samples with Very Low or Undetectable Levels of HBV Surface Antigen. Viruses, 2021. 13(10).
- Malve, B., et al., Impact of deletions and mutations in Hepatitis B virus envelope proteins on serological profile and clinical evolution. Virus Res, 2017. 238: p. 141-147.
- Ijaz, S., et al., Investigating the impact of hepatitis B virus surface gene polymorphism on antigenicity using ex vivo phenotyping. J Gen Virol, 2012. 93(Pt 11): p. 2473-2479.
- Enriquez-Navarro, K., et al., Identification of mutations in the S gene of hepatitis B virus in HIV positive Mexican patients with occult hepatitis B virus infection. Ann Hepatol, 2020. 19(5): p. 507-515.
- Choga, W.T., et al., In Silico Prediction of Human Leukocytes Antigen (HLA) Class II Binding Hepatitis B Virus (HBV) Peptides in Botswana. Viruses, 2020. 12(7).
- Panigrahi, R., et al., Characterization of antiviral resistance mutations among the Eastern Indian Hepatitis B virus infected population. Virol J, 2013. 10: p. 56.
- Makondo, E., T.G. Bell, and A. Kramvis, Genotyping and molecular characterization of hepatitis B virus from human immunodeficiency virus-infected individuals in southern Africa. PLoS One, 2012. 7(9): p. e46345.
- Quiros-Roldan, E., et al., Evidence of long-term suppression of hepatitis B virus DNA by tenofovir as rescue treatment in patients coinfected by HIV. Antivir Ther, 2008. 13(3): p. 341-8.
Figure 1.
HBsAg prevalence by community among people without HIV from the BCPP cohort.
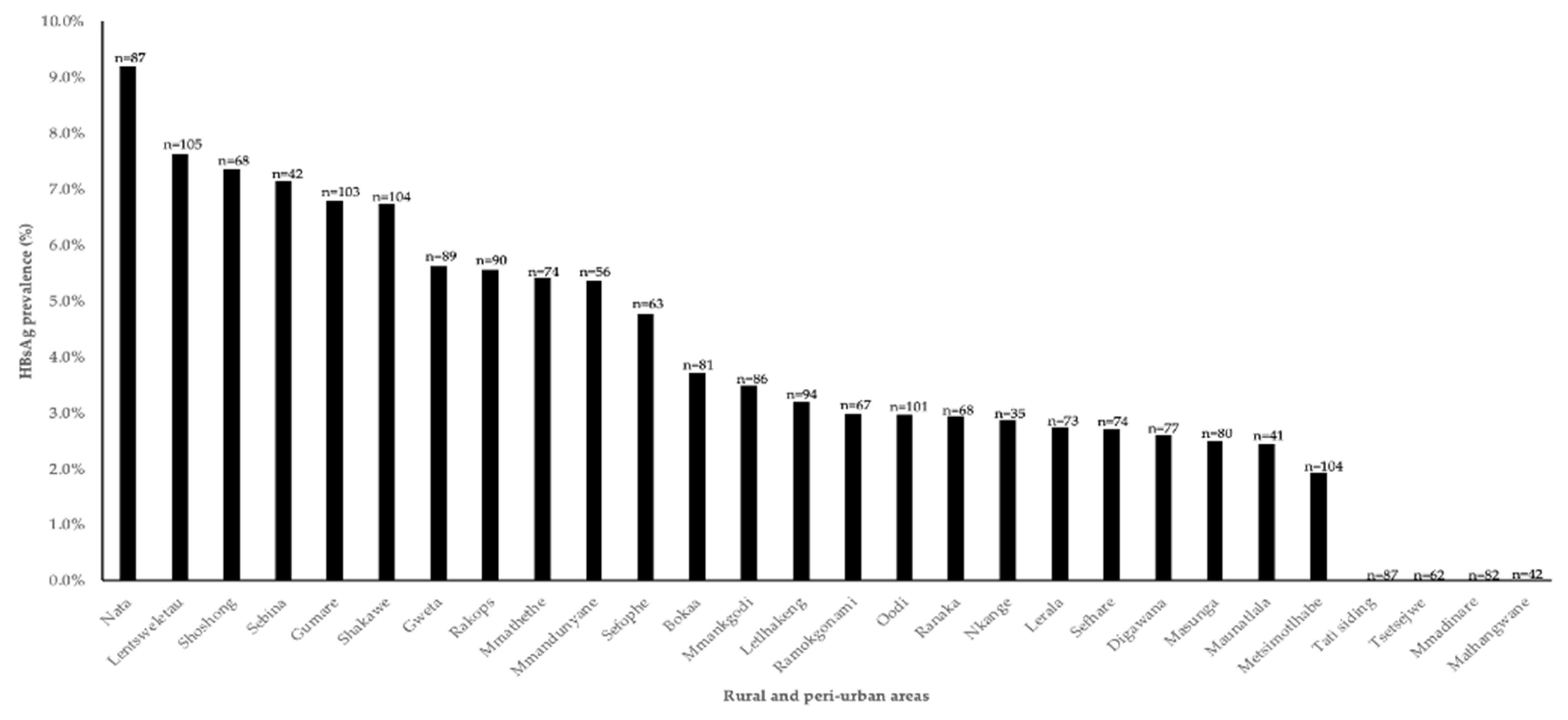
Figure 2.
Phylogenetic analysis of HBV strains obtained from HIV-uninfected participants living in rural and peri-urban areas in Botswana. A phylogenetic analysis of a 415 bp fragment of the HBV from different insolates with seven HBV A, three HBV D and three HBV E sequences clustered with HBV genotype Botswana reference. Study sequences are denoted by the name HBV followed by a number and the references begin with an accession number followed by a letter denoting the genotype. Genotype A is coloured red, D is coloured blue and E is coloured green.
Figure 2.
Phylogenetic analysis of HBV strains obtained from HIV-uninfected participants living in rural and peri-urban areas in Botswana. A phylogenetic analysis of a 415 bp fragment of the HBV from different insolates with seven HBV A, three HBV D and three HBV E sequences clustered with HBV genotype Botswana reference. Study sequences are denoted by the name HBV followed by a number and the references begin with an accession number followed by a letter denoting the genotype. Genotype A is coloured red, D is coloured blue and E is coloured green.
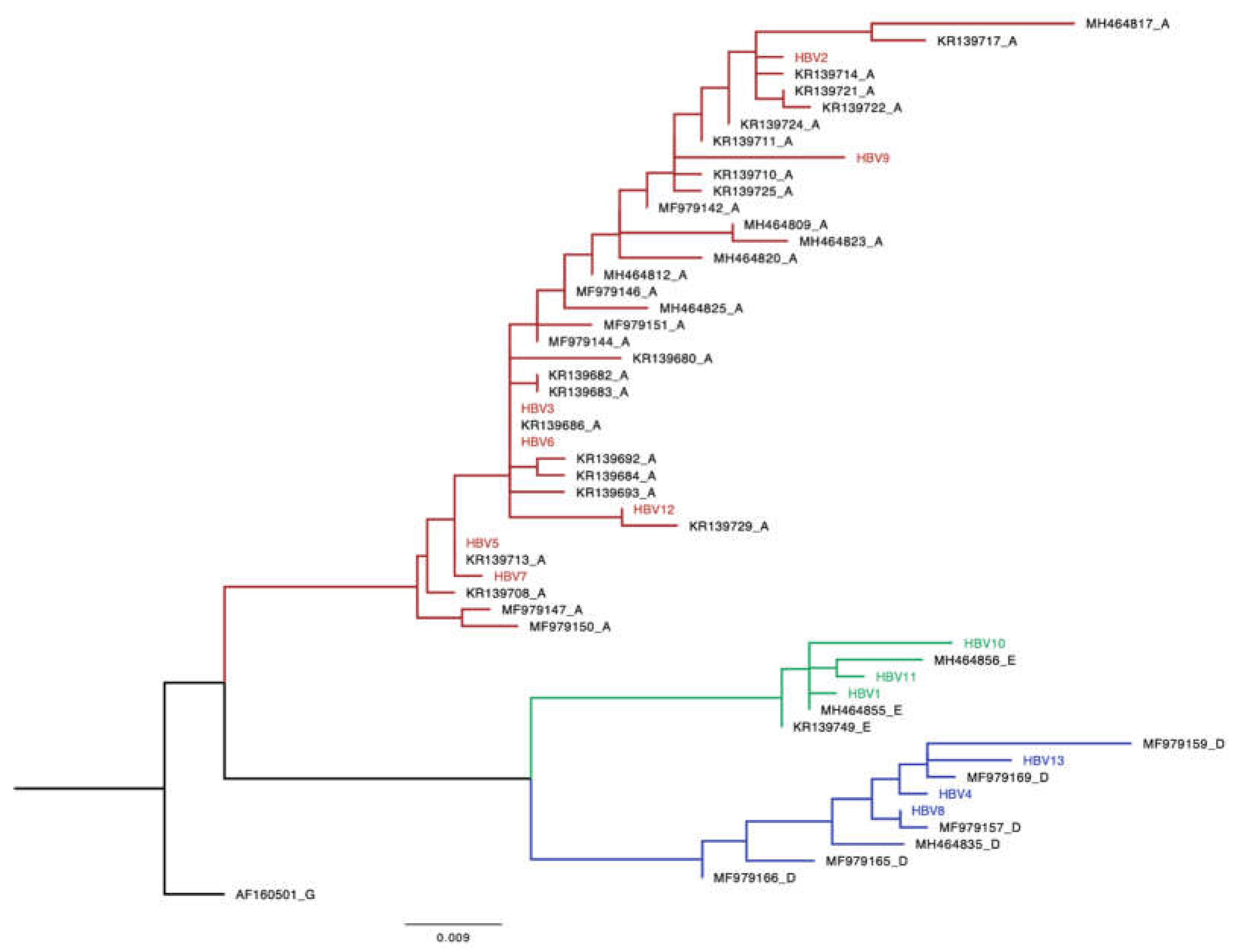
Figure 3.
A. Frequency of surface region mutations. B. Frequency of rt region mutations.
| Mutation | Genotype | Clinical significance |
| rtH100M | E | Not characterized |
| rtI103V | A | Not characterized |
| rtV103I | D | Not characterized |
| rtS105T | A | Not characterized |
| rtS109P | A, E | Not characterized |
| rtR110G | A | Not characterized |
| rtL115V | D | Not characterized |
| rtN122H | A | Not characterized |
| rtF122L | D | Not characterized |
| rtN124H | A | Not characterized |
| rtM129L | A | Not characterized |
| rtQ130P | D | Not characterized |
| rtN131D | A | Not characterized |
| rtL132M | E | Not characterized |
| rtY135S | D | Partly linked to drug resistance |
| rtW153R | A | Not characterized |
| rtV163I | A | Not characterized |
| rtL164M | D | Not characterized |
| rtS213T | D | Not characterized |
| rtG210R | D | Not characterized |
| rtV214E | D | Not characterized |
| sI92C | E | Not characterized |
| sY100S | D | Impair production and secretion of HBsAg, associated with OBI [26,27,28] |
| sK122R | A | Immune escape, Associated with OBI [29,30,31] |
| sT127P | D | Immune escape [32,33] |
| sS140L | E | Immune escape [34] |
| sE164G | D | Not characterized |
| sP188L | E | Not characterized |
| sS193L | A | Not characterized |
| sA194V | A | Not characterized |
| sI195T | A | Not characterized |
| sM197T | A | Correlate with low HBV viral load [35] |
| sW201* | D | Reduces intracellular HBsAg [36] |
| sP203R | A | Associated with HCC [37] |
| sS204R | D | Reduces HBsAg secretion [38] |
| sS204N | A | Correlate with low HBV viral load [35] |
| sY206N | D | Not characterized |
| sS207N | A | Immune escape [39,40,41] |
| Sample ID | Genotype | Mutation rt | Mutations |
|---|---|---|---|
| MA1 | E | - | - |
| MA2 | A | rtS109P, rtM129L, rtW153R, rtV163I | sS193L, sS207N |
| MA3 | A | rtS109P, rtN122H, rtM129L, rtV163I | sA194V, sS207N |
| MA4 | D | rtL115V, rtF122L, rtQ130P, rtY135S, rtG210R | sT127P, sW201* |
| MA5 | A | rtI103V, rtS109P, rtN122H, rtM129L, rtW153R, rtV163I | sA194V, sS207N |
| MA6 | A | rtS109P, rtN122H, rtM129L, rtV163I | sA194V, sS207N |
| MA7 | A | rtI103V, rtS109P, rtN122H, rtM129L, rtW153R, rtV163I | sA194V, sP203R, sS207N |
| MA8 | D | rtV103I, rtL115V, rtF122L, rtQ130P, rtY135S, rtS213T | sT127P, sS204R |
| MA9 | A | rtS105T, rtR110G, rtM129L, rtV163I | sS193L, sA194V, sI195T, sM197T, sS204N, Ss207N |
| MA10 | E | rtH100M, rtS109P | sI92C, sS140L |
| MA11 | E | rtL132M | sP188L |
| MA12 | A | rtS109P, rtN122H, rtN124H, rtM129L, rtN131D, rtV163I | sK122R, sA194V, sS207N |
| MA13 | D | rtL115V, rtF122L, rtQ130P, rtY135S, rtL164M, rtV214E | sY100S, sT127P, sE164G, sY206N |

Table 1.
Univariate and multivariate analysis of demographic factors associated with HBsAg positivity.
Table 1.
Univariate and multivariate analysis of demographic factors associated with HBsAg positivity.
| Characteristic | HBsAg + (n=86) | HBsAg – (n=2049) | Univariate logistic regression | Multivariate logistic regression | ||
|---|---|---|---|---|---|---|
| OR (95% CI) | p-value | aOR(95%CI) | p-value | |||
| Sex, n (%) | ||||||
| Female | 49 (57.0) | 1311 (64.0) | Ref | Ref | ||
| Male | 37 (43.0) | 739 (36.0) | 1.34 (0.73-2.46) | 0.34 | 1.41 (0.77-2.58) | 0.27 |
| Age category, n (%) | ||||||
| <35 | 38 (44.2) | 1242 (60.6) | Ref | Ref | ||
| ≥35 | 48 (55.8) | 807 (39.4) | 1.94 (1.32-2.86) | 0.001 | 1.40 (1.76-2.58) | 0.28 |
| Marital status, n (%) | ||||||
| Single or never married | 58 (67.4) | 1557 (76.0) | Ref | Ref | ||
| Married | 26 (30.2) | 419 (20.5) | 1.67 (1.15-2.41) | 0.007 | 1.15 (0.68-1.92) | 0.61 |
| Widowed/Divorced/separated | 2 (2.3) | 73 (3.5) | 0.74 (0.9-2.87) | 0.66 | 0.43 (0.10-1.82) | 0.25 |
| Region, n (%) | ||||||
| South | 30 (34.9) | 760 (37.1) | Ref | |||
| Central | 23 (26.7) | 639 (31.2) | 0.91 (0.51-1.62) | 0.75 | ||
| North | 33 (38.4) | 650 (31.7) | 1.29 (0.74-2.24) | 0.38 | ||
| Number of lifetime sexual partners (n=1952) | ||||||
| <10 | 68 (82.9) | 1608 (86.0) | Ref | |||
| ≥10 | 14 (17.1) | 262 (14.0) | 1.26(0.64-2.48) | 0.50 | ||
| Education level, n (%) | ||||||
| Non-formal | 19 (22.1) | 216 (10.6) | Ref | Ref | ||
| Primary | 19 (22.1) | 335 (16.4) | 0.64 (0.29-1.42) | 0.28 | 0.64 (0.28-1.46) | 0.29 |
| Secondary | 37 (43.0) | 1133 (55.5) | 0.37 (0.17-0.81) | 0.012 | 0.45 (0.16-1.22) | 0.12 |
| Higher than senior secondary | 11 (12.8) | 359 (17.5) | 0.35 (0.14-0.88) | 0.025 | 0.41 (0.14-1.17) | 0.10 |
| Employment | ||||||
| Unemployed | 54 (62.8) | 1444 (70.5) | Ref | |||
| Employed | 32 (37.2) | 605 (29.5) | 1.41(0.88-2.31) | 0.17 | ||
| Circumcised (n=775) | ||||||
| No | 29 (78.4) | 495 (67.1) | Ref | |||
| Yes | 8 (21.6) | 241 (32.7) | 0.57 (0.29-1.12) | 0.10 | ||
| Inconsistent condom use(n=1632) | ||||||
| Yes | 48 (73.9) | 943 (60.2) | Ref | |||
| No | 17 (26.2) | 624 (39.8) | 0.54 (0.28-1.04) | 0.06 | ||
| Age at First Sex | ||||||
| <18 | 22 (31.9) | 487 (29.7) | Ref | |||
| ≥18 | 47 (68.1) | 1155 (70.34) | 0.90 (0.58-1.40) | 0.64 | ||
Abbreviations: HBsAg; hepatitis B surface antigen, OR; Odds ratio, aOR; adjusted Odds ratio, HBsAg+; hepatitis B surface antigen positive, HBsAg-; hepatitis B surface antigen negative.
Table 2.
HBV Mutations.
| Sample ID | Genotype | Mutation rt | Mutations |
|---|---|---|---|
| MA1 | E | - | - |
| MA2 | A | rtS109P, rtM129L, rtW153R, rtV163I | sS193L, sS207N |
| MA3 | A | rtS109P, rtN122H, rtM129L, rtV163I | sA194V, sS207N |
| MA4 | D | rtL115V, rtF122L, rtQ130P, rtY135S, rtG210R | sT127P, sW201* |
| MA5 | A | rtI103V, rtS109P, rtN122H, rtM129L, rtW153R, rtV163I | sA194V, sS207N |
| MA6 | A | rtS109P, rtN122H, rtM129L, rtV163I | sA194V, sS207N |
| MA7 | A | rtI103V, rtS109P, rtN122H, rtM129L, rtW153R, rtV163I | sA194V, sP203R, sS207N |
| MA8 | D | rtV103I, rtL115V, rtF122L, rtQ130P, rtY135S, rtS213T | sT127P, sS204R |
| MA9 | A | rtS105T, rtR110G, rtM129L, rtV163I | sS193L, sA194V, sI195T, sM197T, sS204N, Ss207N |
| MA10 | E | rtH100M, rtS109P | sI92C, sS140L |
| MA11 | E | rtL132M | sP188L |
| MA12 | A | rtS109P, rtN122H, rtN124H, rtM129L, rtN131D, rtV163I | sK122R, sA194V, sS207N |
| MA13 | D | rtL115V, rtF122L, rtQ130P, rtY135S, rtL164M, rtV214E | sY100S, sT127P, sE164G, sY206N |
Abbreviation: rt: reverse transcriptase, s: surface. Bold =novel, *: stop codon.
Table 3.
Clinical significance of identified HBV mutations.
| Mutation | Genotype | Clinical significance |
| rtH100M | E | Not characterized |
| rtI103V | A | Not characterized |
| rtV103I | D | Not characterized |
| rtS105T | A | Not characterized |
| rtS109P | A, E | Not characterized |
| rtR110G | A | Not characterized |
| rtL115V | D | Not characterized |
| rtN122H | A | Not characterized |
| rtF122L | D | Not characterized |
| rtN124H | A | Not characterized |
| rtM129L | A | Not characterized |
| rtQ130P | D | Not characterized |
| rtN131D | A | Not characterized |
| rtL132M | E | Not characterized |
| rtY135S | D | Partly linked to drug resistance |
| rtW153R | A | Not characterized |
| rtV163I | A | Not characterized |
| rtL164M | D | Not characterized |
| rtS213T | D | Not characterized |
| rtG210R | D | Not characterized |
| rtV214E | D | Not characterized |
| sI92C | E | Not characterized |
| sY100S | D | Impair production and secretion of HBsAg, associated with OBI [26,27,28] |
| sK122R | A | Immune escape, Associated with OBI [29,30,31] |
| sT127P | D | Immune escape [32,33] |
| sS140L | E | Immune escape [34] |
| sE164G | D | Not characterized |
| sP188L | E | Not characterized |
| sS193L | A | Not characterized |
| sA194V | A | Not characterized |
| sI195T | A | Not characterized |
| sM197T | A | Correlate with low HBV viral load [35] |
| sW201* | D | Reduces intracellular HBsAg [36] |
| sP203R | A | Associated with HCC [37] |
| sS204R | D | Reduces HBsAg secretion [38] |
| sS204N | A | Correlate with low HBV viral load [35] |
| sY206N | D | Not characterized |
| sS207N | A | Immune escape [39,40,41] |
Abbreviation: rt: reverse transcriptase, s: surface. Bold =novel, *: stop codon.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



