
Submitted:
08 May 2024
Posted:
09 May 2024
You are already at the latest version
Abstract
Understanding and controlling reproductive cycles and gonadal maturation in cultivated species are crucial in aquaculture. The sea urchin Paracentrotus lividus, known for its edible gonads, requires careful maturation control for both reproduction and commercialization. This study explores the impact of temperature, photoperiod, and diet on gonadal development to enhance year-round aquaculture practices. Using two independent cultivation systems, we manipulated environmental conditions to mimic different seasons. Sea urchins were exposed to natural or manipulated temperature and photoperiod conditions and fed either natural (Ulva spp.) or formulated diets. The gonadosomatic index (GI) and histological analysis assessed gonadal development. Results revealed a clear correlation between environmental conditions, diet, and gonadal maturation. Manipulated conditions accelerated maturation, with sea urchins showing advanced stages compared to natural conditions. Furthermore, sea urchins fed formulated diets exhibited higher GI values, indicating enhanced maturation. Histological analysis confirmed accelerated maturation, particularly in females. The study underscores the feasibility of controlling gonadal maturation through environmental manipulation and diet, enabling year-round marketable gonad production. Providing formulated diets rich in polyunsaturated fatty acids, notably docosahexaenoic acid (DHA), enhances commercial value. These findings optimize aquaculture practices for P. lividus, highlighting its adaptability to yield-maximizing production strategies. Understanding and controlling reproductive cycles and gonadal maturation in cultivated species are crucial in aquaculture. The sea urchin Paracentrotus lividus, known for its edible gonads, requires careful maturation control for both reproduction and commercialization. This study explores the impact of temperature, photoperiod, and diet on gonadal development to enhance year-round aquaculture practices. Using two independent cultivation systems, we manipulated environmental conditions to mimic different seasons. Sea urchins were exposed to natural or manipulated temperature and photoperiod conditions and fed either natural (Ulva spp.) or formulated diets. The gonadosomatic index (GI) and histological analysis assessed gonadal development. Results revealed a clear correlation between environmental conditions, diet, and gonadal maturation. Manipulated conditions accelerated maturation, with sea urchins showing advanced stages compared to natural conditions. Furthermore, sea urchins fed formulated diets exhibited higher GI values, indicating enhanced maturation. Histological analysis confirmed accelerated maturation, particularly in females. The study underscores the feasibility of controlling gonadal maturation through environmental manipulation and diet, enabling year-round marketable gonad production. Providing formulated diets rich in polyunsaturated fatty acids, notably docosahexaenoic acid (DHA), enhances commercial value. These findings optimize aquaculture practices for P. lividus, highlighting its adaptability to yield-maximizing production strategies.
Keywords:
aquaculture
; echinoderms
; reproduction
; histology
; diets
1. Introduction
The sea urchin is an echinoderm of growing interest as a marine resource. In some nations, such as Japan, sea urchin gonads, commonly known as "uni," are an integral component of local culture and cuisine. With almost 126 million residents, Japan is the greatest consumer of sea urchins in the world, consuming 80-90% of the world's sea urchin production [1]. Meanwhile, Chile, with about 6,435 km of coastline, is the largest supplier of sea urchins, with more than 55,000 tons per year [2]. The purple sea urchin Paracentrotus lividus is extensively dispersed in the Mediterranean Sea and throughout the northeastern Atlantic coast, from Scotland and Ireland to southern Morocco [3]. It is a commercially significant species with a strong market demand for its roe, notably in the Mediterranean Basin [4] and in other European non-Mediterranean locations [5,6]. Interest in echinoculture arose because of the growing demand and the unsustainability of capturing wild sea urchins [7,8,9,10]. Echinoculture, in addition to responding to the demand increase, has helped to relieve pressure from fishing in wild populations [11]. Good control of gonad development in sea urchin farming is extremely important to control the reproductive cycle of captive broodstock, but also to control the quality of the gonads when preparing them for collection and marketing for consumption. In Portugal, P. lividus reproduces yearly, with gonad maturity and gamete release occurring at the same time for both sexes. The autumn-winter season marked the gonads' maturity, and the spring/summer season was a single, extended spawning season that followed [12,13]. The Mediterranean Sea has a varied reproductive cycle, with a first vigorous spawning in spring and a second, less intense spawning in October [14]. Variations in environmental conditions, primarily seawater temperature, affects gonadal development and nutrient accumulation and also influences spawning duration, gametogenesis, and spawning are known to vary between geographical locations along a latitudinal gradient [12,15,16]. Nevertheless, the reproductive cycle might also vary significantly from year to year according to how the environment (primarily temperature and photoperiod, but also hydrodynamics) affects P. lividus population dynamics, reproductive biology, and secondary production [12]. The quality of the food also plays a major role in the development of the gonads, and can strongly influence the gonadosomatic indices and biochemical characteristics of the gonads [9,12,13,14,15,16,17]. The main objectives of this work were to understand the influence of two environmental parameters and feeding on the development and maturation of the gonads. To do this, two different seasonal periods were produced by adjusting the temperature and photoperiod. Additionally, two diets were tested: an inert feed formulated for sea urchins and a natural food (the macroalga Ulva spp.).
2. Materials and Methods
2.1. Spawning and Larval Rearing
The production of purple sea urchins (P. lividus) at Aquaculture Research Station (EPPO/IPMA) is carried out using a set of protocols drawn up by combining bibliographical research and the experience of the research team, as well as the specific characteristics of the EPPO's facilities.
2.1.1. Broodstock and Spawning
The wild broodstock were caught off the south coast of Portugal and acclimatized in fiberglass tanks, with open-water circuits, with temperature manipulation whenever necessary, with the manipulated temperature, especially to avoid temperatures above 25 ºC during the summer period. The broodstock was fed with macroalgae (Ulva spp.) produced in earthed ponds at EPPO facilities. Spawning was induced by injecting 1 ml of 0.5 M KCl into the coelom through the peristonial membrane. Fertilization was carried out at a ratio of 500 spermatozoa per 1 egg after determining the number of oocytes. Fertilization took place in 5-litre glass beakers filled with filtered, UV-disinfected water. The eggs were then hatched in 220-litre cylinder-conical fiber tanks, with no water renewal, soft aeration, and low light. Hatching occurred approximately 48 hours after fertilization.
2.1.2. Larval Rearing
The newborn larvae were kept in the same tanks, with an approximate density of 2250 larvae per liter. The tanks remained in a closed water system, with daily renewals of about 20% of the water in the tank. A 50-micron filter was installed at the water outlet to prevent larvae from escaping. The larvae were fed a mixture of microalgae in different proportions in the number of cells: Phaeodactylum tricornutum (50%), Isochrysis galbana Tiso (25%), Nannochloropsis oculate (25%). The amount provided varied depending on the stage of larval development, starting with 6,000 cells/ml to 30,000 cells/ml per day. The microalgae were produced at EPPO facilities, growing in 60-liter plastic bag photobioreactor at 20± 0.5 ºC with aeration, during the sea urchins' larval cultivation phase.
2.1.3. Post-Larvae and Juveniles’ Cultivation
Metamorphosis and settlement began between 20 and 30 days after hatching, approximately. To increase the surface available for fixation in the tanks, plastic pieces consisting of polyethylene terephthalate were suspended, in which natural biofilm was developed. During the first few weeks after settlement, the tanks were kept in a closed water system, with small daily renewals. In addition to the food provided by the biofilm, around 2 liters of Phaeodactylum tricornotum microalgae (roughly 6x109 cells) were added daily to each tank. With the progressive growth of the sea urchins, the supply of macroalgae (Ulva spp.) previously washed with fresh water was started. Once they were close to 5 mm in test diameter (a transfer-safe size), the juvenile sea urchins were transferred to 300-liter raceway fiber tanks with an open water system and aeration. The water temperature was kept at room temperature and cooled in the warmer months so as not to exceed 22ºC. From then on, the juveniles were fed exclusively on macroalgae Ulva spp. ad libitum. The tanks were cleaned before new food was supplied.
2.2. Experimental Design
In order to analyze the effect of temperature, photoperiod, and food type, four cultivation systems were created: NoM, U- No environmental manipulation and natural food (Ulva spp.); NoM, If- No environmental manipulation and inert feed; M, U- Manipulated environment and natural food (Ulva spp.); M, If - Manipulated environment and inert feed (M, If). Six rectangular fiberglass tanks with a volume of 110 L were used for this test. Three tanks were subjected to manipulated environmental conditions and three were subjected to natural environmental conditions. Plastic boxes measuring 0.57x0.38x0.08 m were placed in each tank, divided in half by a 3mm mesh plastic net. Each side of the box was subjected to different diets (Ulva spp. and inert food). Three replicates were created for each treatment (NoM,U, NoM, M, U and M, If). Each tank had an aeration system and open-water system with a renewal rate of approximately 112.5 L/h. The tanks were lit by halogen lamps with an average intensity at the water surface of 100 lux (Figure 1).
A total of 300 F1 purple sea urchins (P. lividus), approximately 3 years old, were used for this study with mean weight of 22.73 (±4.46) g. Fifty individuals were randomly placed in each box, 25 for each half of the box.
2.2.1. Environment Conditions
Two different environmental conditions were created, with three tanks in each system (Table 1). The “No environmental manipulation” (NoM) system was subjected to normal ambient temperatures for October to February (there was no temperature control). In this system, a photoperiod typical of southern Portugal (37.0265, -7.84045) was regulated for the three months in question (Oct-Feb). In the “Manipulated environment system”, the temperature was controlled using a system of chiller and heaters to recreate the temperature conditions of December to April, which means a two month anticipation. The photoperiod was regulated for the above-mentioned months (December-April).
2.2.2. Feeding
Two diets were used in this study. The natural diet (U) consisted of fresh macroalgae (Ulva spp.) produced in earthen ponds. The inert diet (If) consisted of pellets formulated and produced by the company SPAROS, R&D (Portugal), specifically for sea urchins. nutritional characteristics are described in Araújo et al. (2023). The sea urchins were fed once every 3-4 days based on a daily consumption for each urchin of 0.4 grams of Ulva spp. and 0.15 grams of inert food at the beginning of the trial. The amount of feed was adjusted every month based on the residual total biomass of each replicate.
2.3. Sampling
Biometric sampling was performed every month (5 sampling moments) measuring the total weight with a KERN PRS / PRJ precision and analytical balance of all sea urchins from the three replicate of each treatment (n= 25 each replicate). Five individuals were randomly selected from each treatment to collect the gonads, after recording their total weight. The gonads were weighted and the gonadosomatic index was calculated using the following equation:
Gonadosomatic Index (GSI) = (Gonad wet weight (g))/(Total wet weight (g))×100
Samples of each gonad were then placed in histological cassettes for later procedures and observation in the laboratory. The cassettes were maintained in 10% formaldehyde for 48h for fixation. Next, they were transferred to 70% ethanol for storage until paraffin histology.
2.4. Histology
Slide sections (4 µm thick) ( were prepared using a tissue processor (Model Citadel 2000, Thermo Scientific, Nanjing, China) and a microtome (Model Jung RM 2035, Leica Instruments mb, Etzlar, Germany). Haematoxylin and eosin were used to stain slides using an automatic slide Stainer (Model Shandon Varistain 24-4, Thermo Scientific, Nanjing, China). Images were visualized and photographed with the NDP View 2, and scanned with a Hamamatsu NanoZoomer (C13140-01). The different stages of maturation were classified according to Byrne’s (1990).
2.5. Statistical Analysis
A statistical analysis was carried out to compare the gonadosomatic indices of the sea urchins subjected to the four experimental conditions (NoM U- Non-manipulated conditions, feed with Ulva spp.; NoM If- Non-manipulated conditions, feed with formulated feeds; M U- Manipulated conditions, feed with Ulva spp; M If- Manipulated conditions, feed with formulated feeds), for both females and males, for each sampling time. For each point, normality was tested using the Shapiro-Wilk test and equality of variances was tested using the Brown-Forsythe test. If positive, One Way ANOVA was carried out; if negative, the data was subjected to Kruskal-Wallis One Way Analysis of Variance on Ranks. If there were significant differences (P>0.001), the data was subjected to All Pairwise Multiple Comparison procedures. Following ANOVA on Ranks, Dunn's Method was used. For One Way ANOVA, the Holm-Sidak method was used. Pairs of samples were considered statistically different when p<0.05. The analyses were carried out using the SIGMAPLOT 14.0 software developed by Systat Software, Inc
3. Results
3.1. Gonadosomatic Index
The variation in gonadosomatic indices was analyzed considering the different treatments and the different sexes. Starting with the male individuals, all treatments showed an increase in this index over the 4 months of work (Figure 2). It seems also clear that animals of both sexes fed inert food always had a higher gonadosomatic index. At each point, significant differences were seen in pairs of treatments (p<0.05). In all cases of significant differences, it was found that sea urchins fed inert food had higher gonadosomatic indices than those fed Ulva spp. In November, significant differences were found between sea urchins fed different diets under unmanipulated conditions (NoM U and NoM If). In December, these two treatments (NoM U and NoM If) maintained significant differences, and there were also differences between urchins fed different diets under manipulated conditions (M U and M If). In January, there were differences only between sea urchins in unmanipulated conditions fed on inert food (NoM If) and those in manipulated conditions fed on Ulva spp. (M U). In February there was an increase in the differences between the treatments. In this case, there were significant differences between all the pairs of treatments where feeding was different.
In the case of females, the differences were not so obvious. In November and December, the gonadosomatic indices for the four treatments were not statistically significant (P = 0.281 and P = 0.078, respectively). In the following months, the differences increased. In January, there were significant differences in sea urchins under manipulated conditions fed different diets (M U and M If). In February, the biggest difference was found in sea urchins in manipulated conditions fed Ulva spp. (M U) and individuals in unmanipulated conditions fed inert food (NoM If). As in the case of males, the differences were due to higher gonadosomatic index values in sea urchins fed inert food (If).
3.1. Gonad Development
As with the gonadosomatic index, gonad development was analyzed considering the condition of the test (manipulated and non-manipulated), the diet (Ulva spp. and inert food) and both sexes. Figure 3 and Figure 4 describe each stage for males and females, respectively.
The characteristics of the gonads and gametophytes were used to classify and quantify each individual in their respective stages of maturity, and this information is shown in Figure 5 and Figure 6.
Figure 5 shows the evolution of the maturation stages of the gonads of female sea urchins over the four months of the trial, for the four treatments: NoM U (non-manipulated conditions fed with Ulva spp.), NoM If (non-manipulated conditions fed with inert food), M U (manipulated conditions fed with Ulva spp.), M If (non- manipulated conditions fed with inert food. Keeping in mind that manipulation describes adjustments to temperature and photoperiod in relation to natural environmental conditions. For all four treatments, there was a notable evolution over the study period, always reaching the last stage of development, stage VI (spent stage). However, there were some differences in terms of speed of development. The greatest differences were found between urchins kept in manipulated environmental conditions compared to unmanipulated conditions.
When we compare the female sea urchins fed Ulva spp. but under different environmental conditions, we see a more rapid evolution from November onwards. The following month (December), under manipulated conditions, there was a higher percentage of sea urchins in stage II (growing stage), with some individuals already in stage III (premature stage). In January, 57.1% of the sea urchins in manipulated conditions were already in stage VI (spent stage), with no individuals in the same stage in non-manipulated conditions. This trend was also seen in females fed inert food. In December, in the manipulated conditions it was already possible to see individuals in stages II and III, unlike the situation in the non-manipulated conditions, where all the individuals observed were in stage I. In January, the female urchins in non-manipulated conditions were already at an advanced stage of maturation, with a mixture of females in stages II, III, IV and some in stage VI. For the same month, 70 per cent of the individuals in the manipulated situation fed the same inert food were already in stage VI. At the end of the trial, the percentage of individuals in stage VI was practically the same for both situations (66.6 per cent for NoM If and 71.4 per cent for M If). Although less emphasized, differences were also observed between sea urchins fed different diets under the same environmental conditions. In unmanipulated conditions, individuals in stage VI were found in January when fed inert food, which was not the case with sea urchins fed Ulva spp. In manipulated conditions, there was also an earlier onset of maturation in females fed inert food. However, these differences dissolve in January.
As with the females, the greatest differences were seen between the male sea-urchins kept in manipulated environmental conditions compared to unmanipulated conditions (figure 6). In the urchins fed Ulva spp., the stages of gonad maturation showed the greatest progression in December, with individuals already at stage III (premature stage). In January, 63 % of the sea urchins fed algae under manipulated conditions were already at stage VI (spent stage). In the same month, urchins fed the same food, but without manipulation of environmental conditions, still had no males in this stage. The following month, the males in the unmanipulated conditions reached the final stage (VI), with a large percentage (67%) of the manipulated ones already in the recovery stage (I). This situation is similar to that of the sea-urchins fed inert food, where it can be seen that in January 40% of the males were already in stage VI, and no individuals were observed in this stage in that month. When we compare the sea urchins under the same environmental conditions but with different feedings, we can also see some differences in terms of gonad maturation. In non-manipulated conditions, males started maturing earlier when fed inert food, but at the end of the trial the percentage of individuals in stage VI was very similar. Under manipulated conditions, the evolution was apparently superior in the animals fed on algae.
When comparing male and female individuals, females mature earlier than males, and this is particularly relevant in unmanipulated environmental conditions.
4. Discussion
In aquaculture, a good understanding of the maturation of the gonads of cultivated species are necessary. In the case of sea urchins, this control is important not only for reproductive purposes but also for the commercialization process, considering that the gonads are the edible part of sea urchins. In the natural environment, the species Paracentrotus lividus has a reproductive cycle that varies with the location of its population. In Portugal, it is considered a species with annual reproduction, with a fairly long spawning period that begins in mid-spring and ends in mid-summer [12,21,22]. On the northern coast of Spain, spawning occurs between March and September, potentially happening at two distinct times during this period, depending on the environmental characteristics of each year [23]. On the coast of Morocco, a more extended spawning period is observed, between March and October [24]. In higher latitudes, such as in Ireland, spawning begins later, in May and June, and ends in August or September [5]. In the Mediterranean Sea exhibits a varied reproductive cycle, with a primary vigorous spawning in spring and a secondary, less intense spawning in October [14]. Given the variability and reproductive adaptation of this species to geographically distinct conditions, it is considered feasible to control maturation and reproduction. By understanding the factors that influence the reproductive cycle, it may be possible to manipulate farming conditions in aquaculture so that the gonads mature and reproduce at any time of the year. In this way, it may be possible for a commercial aquaculture farm to have several spawning per year, thus increasing production.
According to the available literature, several factors influence maturation and the reproductive cycle in nature, the most commonly cited being temperature, photoperiod and food availability [5,12,23,24,25]. This work was therefore designed taking into account the influence of these three factors. Due to technical impossibility, the environmental parameters temperature and photoperiod were not analyzed in isolation, but were aggregated and their joint action considered as determining parameters of local seasonality. Therefore, two independent cultivation systems were created that recreate the environmental conditions in two seasonal periods. In the non- manipulated condition, the temperature and photoperiod conditions correspond to the natural conditions for the season. In the manipulated condition, the sea urchins were subjected to conditions that correspond to three months ahead of the actual month. Two different diets were used in these two conditions. A natural food (Ulva spp.) and a food formulated for sea urchins. The choice of these two diets comes from previous knowledge of their potential influence on gonad development [18].
4.1. Gonadossomatic Index
The general results of this study reflect the natural evolution of the reproductive cycle of this species, in this case the period after resting until spawning. In all treatments, an increase in the gonadosomatic index was observed between November and January, followed by a downward trend in February. In general, low GI indicates gamete release, while high GI indicates gonad maturity and the building of energy reserves [22,26]. Higher GI during the first few months of the year is indicative of growing during the winter and maturing (spring) stages of the gonadal development cycle, when the gonads are filled with spermatozoa and eggs. The index declines in late spring or early summer signaled the start of the spawning period [25,27].
The general results regarding the progression of the stages of gonad development also represent the natural tendency for this species. During the progression of gametogenesis there is an increase in the number of germ cells and an accumulation of nutrient reserves in the somatic cells, the nutritive phagocytes, at the beginning of gametogenesis. This is then visible through histological analysis of the gonads and an increase in the gonadosomatic index. In this study, we were able to see differences in the development of the gonads between the four treatments. Regarding gonadosomatic index, the most significant differences were between sea urchins fed different diets. Although the pattern of variation is very similar, the gonadosomatic index values are always higher for sea urchins fed inert food, with the differences being more significant in the last few months of the trial. Despite showing a similar pattern, the variation in males and females was slightly different, with a more significant increase in the first few months in the case of males, and then a greater increase in females from December onwards. The importance of feed in the gonadosomatic index has already been verified in other similar studies. It is known that the abundance of food in the wild has a positive correlation with the size of sea urchin gonads [22,29,30,31]. The nutritional quality of the food will also play a major role in the constitution and development of the gonads. Candeias-Mendes et al. (2020) found that sea urchins fed maize and Ulva spp. had gonadosomatic indices approximately three times higher than animals fed only algae, with a strong relationship between the fatty acid profile of the gonads and the feed provided. Lourenço et al. (2020) concluded that a feed with a protein content of 30% DM and 21 mg P kJ-1 promotes nutrient utilization and gonad growth in weight. Raposo et al. (2019) tested three different diets and found that animals fed maize and spinach had higher gonadosomatic indices, suggesting a direct relationship between lipid content and gonad size. Our study used two different diets in terms of presentation and nutritional characteristics: fresh Ulva spp. and a dry inert food presented in the form of disc-shaped pellets. Analyses of its fatty acid profile showed that the inert food had a higher content of the three types of fatty acids: saturated (SFA), monounsaturated (MUFA) and polyunsaturated (PUFA). The inert food had much higher levels of 22:6 ω3 (DHA) than the macroalgae, due to the enrichment of microalgae biomass (Schizochytrium), and a high content of this fatty acid was then found in the gonads of the sea urchins fed this food, showing the close relationship between the food supplied and the gonads [18].
4.2. Gonadal Maturity
The gonadosomatic index is a parameter that allows to obtain information on the development of the gonads, but it is through histological analysis that is possible to observe the development of gametogenesis and, therefore, the state of maturation of the gonads. This study used the maturation scale proposed by Byrne (1990), which has been widely used in studies of the reproductive cycle of the species P. lividus [12,23,24,25,28,33] . The development of the gonads observed in this study reflects the natural pattern of this species' reproductive cycle. In general, in November the vast majority of individuals were found to be in the recovery stage (stage I), which translates into the presence of developing germ cells, and primary oocytes and spermatocytes. In the following month, the differences between the treatments began to become more pronounced, revealing a tendency for the maturation of sea urchins exposed to the manipulation of temperature and photoperiod conditions to accelerate, especially for the females. In the following month (the non-manipulated situation corresponds to January, while the manipulated conditions correspond to April) a strong evolution of the gonads was observed, which was particularly noticeable again in the females subjected to manipulation of environmental conditions, where it was possible to observe a large percentage of individuals in stage VI (Spent stage). This stage corresponds to the post-spawning period, when the spent ovaries have thin ascinal walls and appear empty except for relict oocytes [5]. For the same month, most of the males were still in the lower stages of maturation. It is generally assumed that this species shows synchronous gonad maturation. Kahili et al. (2023) provides an example where synchronisation in the gametogenesis of males and females was verified, although several cases of some anticipation of females or males in terms of gonad maturation have been described. In Ouchene et al., 2021, an earlier maturation of the males was observed, and it was also possible to see that in the last month of laying most of them were already in stage VI. The same authors relate this difference to the environmental conditions that characterized that year's study. In the case of Machado et al. (2018), the reproductive cycle of two populations in different locations on the Portuguese coast was characterized, and a clear synchronism was observed in one population and an anticipation of females in the other population, which shows that environmental conditions may indeed play a role in the synchronism of gametogenesis.
In the last month of the study, there were individuals in stage VI in all the treatments, and it was also possible to observe that in the manipulated conditions there was already a significant percentage of animals in stage I (recovery stage), corresponding to individuals who were already restarting a new gametogenic cycle.
Overall, we can see that the manipulation of environmental conditions (temperature and photoperiod) influenced accelerating gonad maturation. Under the manipulated conditions, the animals were subjected to a cooling of the water and a reduction in the photoperiod in the first month of the trial, in order to reproduce the normal conditions corresponding to the month of January. It is known that lowering the water temperature promotes the storage of nutrients, while increasing it is associated with the stimulation of gametogenesis [5,31]. On the other hand, in relation to photoperiod, it is known that reducing the duration of daily light stimulates nutrient storage, with the shortest days of the year being associated with the onset of gametogenesis [25]. There are even studies that have shown that subjecting sea urchins to darkness promoted a strong stimulus for gonad growth [15]. In this study, manipulated conditions were created in which there was an early decrease in photoperiod and temperature in the animals under the manipulated conditions, with the minimum temperature and photoperiod values being recreated in the month of October. This manipulation then promoted the start of gametogenesis in these animals earlier than normal. In January, it was possible to observe animals already in stage VI (spend phase) in the males and females in the manipulated situation (there were also some females in the non-manipulated condition fed inert food, but in smaller quantities). This month in the manipulated condition corresponds to April, a time of year when spawning begins in several Iberian populations [12,23,25].
5. Conclusions
Controlling gonad maturation and reproduction is of great importance in production and commercialization of the gonads. Through this work it was possible to verify that by feeding with suitable inert formulated feeds and manipulating environmental conditions it is possible to create conditions that allow to get spawning all year round and marketable gonads to be obtained at different times of the year. Controlling temperature and photoperiod seems to be a viable strategy for controlling gonad maturation. Providing an inert food with a high PUFA content, especially DHA, could optimize the gonadosomatic index, thus enhancing the value of sea urchins at the time of marketing. These results show that this species of sea urchin, in addition to its commercial potential, has plastic characteristics that allow it to adapt to production strategies that optimize its commercial yield.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. Conceptualization, J.A., F.S., P.P.F.; methodology, J.A., F.S.,A.C.M. writing – original draft, J.A.; investigation, J.A.; data curation, J.A, I.C.; histology methods and analysis, A.G.; practical work, I.C., J.A.; formal analysis, J.A., I.C.; writing- review and editing, J.A., A.G., A.C.M., F.S., E.C.; supervision, E.C., F.S.; funding acquisition, PPF; resources, PPF.
Funding
This research was funded by Project INOVAQUA – Inovação, diversificação e desenvolvimento da aquacultura em Portugal (MAR-021.1.3-FEAMPA-00004) and project SAÚDE&AQUA II - Prevenção e bem-estar em aquacultura marinha: Apoio à identificação e controlo de patógenos em aquacultura (MAR-021.1.3-FEAMPA-00018).
Institutional Review Board Statement
The study was conducted according to the guidelines of the recommendations of the Federation of European Laboratory Animal Science Associations (FELASA) and the Portuguese legislation for Laboratory Animal Science (EU Directive 2010/63; Decreto-Lei nº 113/2013), and approved by IPMA’s Animal Welfare and Ethics Body (ORBEA), overseen by the National Authority for the use of live animals, also known as the Directorate-General for Food and Veterinary (DGAV).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sun, J.; Chiang, F.S. Use and Exploitation of Sea Urchins. In Echinoderm Aquaculture eds Brown, N.P., Eddy, S.D. 2015; 22–45. [Google Scholar]
- Carboni, S.; Vignier, J.; Chinatore, M.; Tocher, D.; Migaud, H. Effects of dietary microalgae on growth, survival and fatty acidcomposition of sea urchin Paracentrotus lividus throughout larval development. Aquac. 2012, 324–325, 250–258. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Verlaque, M. Ecology of Paracentrotus lividus, in: Lawrence, J.M. (Ed.) Edible sea urchins: biology and ecology. Developments in aquaculture and fisheries science. 2001 32: pp. 177–216.
- Régis, MB. Analyses des fluctuations des indices physiologiques chez deux khinoides (Paracentrotus lividus (Lmk) et Arbacia lixula L.) du Golfe de Marseille. Thkthys. 1979, 9, 167–181. [Google Scholar]
- Byrne, M. Annual reproductive cycles of the commercial sea urchin Paracentrotus lividus from an exposed intertidal and a sheltered subtidal habitat on the west coast of Ireland. Mar. Biol. 1990, 104, 275–289. [Google Scholar] [CrossRef]
- Barnes, D.; Crook, A. Quantifying behavioural determinants of the coastal European sea-urchin Paracentrotus lividus. Mar.Biol. 2001, 138, 1205–1212. [Google Scholar]
- Cyrus, M.D.; Bolton, J.J.; De Wet, L.; Macey, B.M. The development of a formulated feed containing Ulva (Chlorophyta) to promote rapid growth and enhanced production of high quality roe in the sea urchin Tripneustes gratilla (Linnaeus). Aquacult. Res. 2014, 45, 159–176. [Google Scholar] [CrossRef]
- Mendes, A.; Araújo, J.; Soares, F.; Pousão Ferreira, P. Produção de larvas e juvenis de ouriços-do-mar (Paracentrotus lividus) na estação piloto de piscicultura de Olhão (EPPO). Relat. Cient. Téc. IPMA 2018, Série digital nº20. [Google Scholar]
- Candeias-Mendes, A.; Araújo, J.; Santos, M.; Namora, M.; Soares, F.; Gomes, R.; Cardoso, C.; Afonso, C.; Bandarra, N.M.; Pousão-Ferreira, P. Growth, survival and fatty acids profile of sea urchins, Paracentrotus lividus juveniles fed with Ulva spp. and maize in aquaculture production. First results using G1 generation in Portugal. J. Aquac. Mar. Biol. 2020, 9, 208–214. [Google Scholar]
- Santos, P.; M. , Albano; P., Raposo; A.; Ferreira, S.M.F.; Costa, J.L.; Pombo, A. The effect of temperature on somatic and gonadal development of the sea urchin Paracentrotus lividus (Lamarck, 1816). Aquac. 2020, 528, 735487. [Google Scholar] [CrossRef]
- Ciriminna, L.; Signa, G.; Vaccaro, A.M.; Messina, C.M.; Mazzola, A.; Vizzini, S. Formulation of a new sustainable feed from food industry discards for rearing the purple sea urchin Paracentrotus lividus. Aquacult. Nutr. 2020, 26, 1046–1057. [Google Scholar] [CrossRef]
- Machado, I.; Moura, P.; Pereira, F.; Vasconcelos, P.; Gaspar, M.B. Reproductive cycle of the commercially harvested sea urchin (Paracentrotus lividus) along the western coast of Portugal. Invertebr. Biol. 2019, 138, 40–54. [Google Scholar] [CrossRef]
- Raposo, A.; Ferreira, S.; Ramos, R.; Anjos, C.; Baptista, T.; Tecelão, C.; M. , Santos; P.; Costa, J.; Pombo, A. Effect of three diets on the gametogenic development and fatty acid profile of Paracentrotus lividus (Lamarck, 1816) gonads. Aquac. Res. 2019, 50, 10–1111. [Google Scholar] [CrossRef]
- Vafidis, D.; Antoniadou, C.; Kyriakouli, K. Reproductive Cycle of the Edible Sea Urchin Paracentrotus lividus (Echinodermata: Echinoidae) in the Aegean Sea. Water 2019, 11, 1029. [Google Scholar] [CrossRef]
- McCarron, E.; Burnell, G.; Kerry, J.; Mouzakitis, G. An experimental assessment on the effects of photoperiod treatments on the somatic and gonadal growth of the juvenile European purple sea urchin Paracentrotus lividus. Aqua. Res. 2010, 41, 1072–1081. [Google Scholar] [CrossRef]
- Sellem, F.; Guillou, M. Reproductive biology of Paracentrotus lividus (Echinodermata: Echinoidea) in two contrasting habitats of northern Tunisia (south-east Mediterranean). J. Mar. Biol. Ass. 2007, UK 87, 763–767. [Google Scholar] [CrossRef]
- Cyrus, M.; Bolton, J.; De Wet, L.; Macey, B.M. The development of a Formulated Feed containing Ulva (Chlorophyta) to promote rapid growth and enhanced production of high-quality roe in the sea urchin Tripneustes gratilla (Linnaeus). Aquac. Res. 2012, 45, 159–176. [Google Scholar] [CrossRef]
- Araújo, J.; Loureiro, P.; Candeias-Mendes, A.; Gamboa, A.; Bandarra, N.; Cardoso, C.; Soares, F.; Dias, J. ; Pousão-Ferreira. The effect of a formulated feed on the body growth and gonads quality of purple sea urchin (Paracentrotus lividus) aquaculture produced. J. Aquac. Mar. Biol. 2023; 12, 11–18. [Google Scholar]
- Santos, P.M.; Ferreira, S.M.F.; Albano, P.; Raposo, A.; Costa, J.L.; Pombo, A. Can artificial diets be a feasible alternative for gonadal growth and maturation of the sea urchin Paracentrotus lividus (Lamarck, 1816)? J. World. Aquacult. Soc. 2020, 51, 463–487. [Google Scholar] [CrossRef]
- Schlosser, S.C. , Lupatsch, I. , Lawrence, J.M., Lawrence, A.L., & Shpigel, M. (2005). Protein and energy digestibility and gonad development of the European sea urchin Paracentrotus lividus (Lamarck) fed algal and prepared diets during spring and fall Aquac. Res. 2005, 36, 972–982. [Google Scholar]
- Rocha, F.; Baião, L.F.; Moutinho, S.; Reis, B.; Oliveira, A.; Arenas, F.; Maia, M.R.G.; Fonseca, A.J.M.; Pintado, M.; Valente, L.M.P. The effect of sex, season and gametogenic cycle on gonad yield, biochemical composition and quality traits of Paracentrotus lividus along the North Atlantic coast of Portugal. Sci. Re.p 2019, 28, 2994. [Google Scholar] [CrossRef]
- Nicolau, L.; Vasconcelos, P.; Machado, I.; Pereira, F.; Moura, P.; Carvalho, A.N.; Gaspar, M.B. Morphometric relationships, relative growth and roe yield of the sea urchin (Paracentrotus lividus) from the Portuguese coast, Reg. Stud. Ma.r Sci. 2022, 52, 102343. [Google Scholar]
- González-Irusta, J.; Cerio, F.; Canteras, J.C. Reproductive cycle of the sea urchin Paracentrotus lividus in the Cantabrian Sea (northern Spain): Environmental effects. J. Mar. Biol. Assoc. U. K. 2010, 90, 699–709. [Google Scholar] [CrossRef]
- Ouchene, H.; Boutgayout, H.; Hermas, J.; Benbani, A.; Oualid, J.A.; Elouizgani, H. Reproductive Cycle of Sea Urchin Paracentrotus lividus (Lamarck, 1816) from the South Coast of Morocco: Histology, Gonads Index, and Size at First Sexual Maturity. Arab. J. Sci. Eng. 2021, 46, 5393–5405. [Google Scholar] [CrossRef]
- De la Uz, S.; Carrasco, J.F.; Rodríguez, C. Temporal variability of spawning in the sea urchin Paracentrotus lividus from northern Spain Reg. Stud. Mar. Sci. 2018, 23, 2–7. [Google Scholar]
- Allain, J.Y. Structure des populations de Paracentrotus lividus (Lamarck) (Echinodermata, Echinoidea) soumises à la pêche sur les côtes Nord de Bretagne. Rev. Trav. Inst. Pêch. Marit 1975, 39, 171–212. [Google Scholar]
- Ouréns, R.; Fernández, L.; Freire, J. Geographic, population, and seasonal patterns in the reproductive parameters of the sea urchin Paracentrotus lividus. Mar. Biol. 2011, 158, 793–804. [Google Scholar] [CrossRef]
- Khaili, A.; Haroufi, O.; Bouzaidi, H.; Maroua, H.; Bouzoubaa, A.; Rharrass, A.; Essalmani, H. Reproductive cycle of the sea urchin Paracentrotus lividus (Lamarck, 1816) from the Moroccan western Mediterranean Sea: histology, gonadal index and size at first sexual maturity. EJABF 2023, 27, 745–771. [Google Scholar]
- Guetaff, M.; San Martin, G.A. Étude de la variabilité de l’indice gonadique de l’oursin comestible Paracentrotus lividus (Echinodermata: Echinoidea) en Mediterranée nord-occidental. Vie Milieu, 1995; 45, 129–137. [Google Scholar]
- Fernandez, C.; Boudouresque, C. Phenotypic plasticity of Paracentrotus lividus (Echinodermata: Echinoidea) in a lagoonal environment. Mar. Ecol. Prog. 1997, 152, 145–15. [Google Scholar] [CrossRef]
- Spirlet, C.; Grosjean, P.; Jangoux, M. Reproductive cycle of the echinoid Paracentrotus lividus. Invertebr. Reprod. Dev. 1998, 34, 69–81. [Google Scholar] [CrossRef]
- Lourenço, S.; Cunha, B.; Raposo, A.; Neves, M.; Santos, P.M.; Gomes, A.S.; Tecelão, C.; Ferreira, S.M.F.; Baptista, T.; Silvia, C.G.; Pombo, A. Somatic growth and gonadal development of Paracentrotus lividus (Lamarck, 1816) fed with diets of different ingredient sources. Aquac. 2021, 539. [Google Scholar] [CrossRef]
- Cirino, P.; Ciaravolo, M.; Paglialonga, A.; Toscano, A. Long-term maintenance of the sea urchin Paracentrotus lividus in culture. Aquac. Res. 2017, 7, 27–33. [Google Scholar] [CrossRef]
Figure 1.
Representative diagram of the system used for the test. On the left are three tanks (replicates) subjected to non-environmental manipulation (NoM). On the right are the tanks subjected to temperature manipulation and altered photoperiod (M). Each tank was divided in half and each half was subjected to different diets (U- Ulva spp., If- Inert feeds).
Figure 1.
Representative diagram of the system used for the test. On the left are three tanks (replicates) subjected to non-environmental manipulation (NoM). On the right are the tanks subjected to temperature manipulation and altered photoperiod (M). Each tank was divided in half and each half was subjected to different diets (U- Ulva spp., If- Inert feeds).
Figure 2.
Evolution of the gonadossomatic index of male (left) and female (right) sea urchins (Paracentrotus lividus) under different conditions (NoM U- Non-manipulated conditions, feed with Ulva spp.; NoM If- Non-manipulated conditions, feed with formulated feeds; M U- Manipulated conditions, feed with Ulva spp; M If- Manipulated conditions, feed with formulated feeds. Equal letters mean pairs of samples, with significant differences (P<0.05) for each month.
Figure 2.
Evolution of the gonadossomatic index of male (left) and female (right) sea urchins (Paracentrotus lividus) under different conditions (NoM U- Non-manipulated conditions, feed with Ulva spp.; NoM If- Non-manipulated conditions, feed with formulated feeds; M U- Manipulated conditions, feed with Ulva spp; M If- Manipulated conditions, feed with formulated feeds. Equal letters mean pairs of samples, with significant differences (P<0.05) for each month.
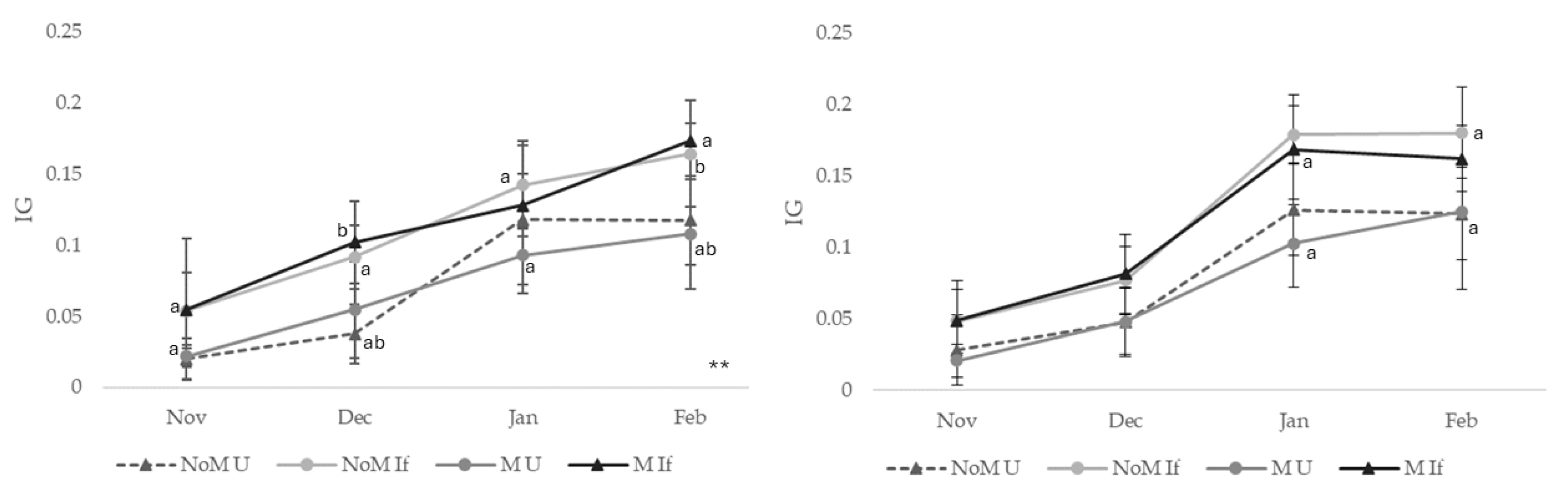
Figure 3.
Histology sections of testis in Paracentrotus lividus on different stages, through the trial: (a,b) Stage I: recovery stage may has a restored testis with relict spermatozoa (RS) and parallel displays of primary spermatocytes along the ascinal wall (PS) and nutritive phagocytes (NP). (c) Stage II: growing stage displays nutritive phagocytes (NP), primary spermatocytes (PS) along the ascinal wall and columns of spermatocytes (S) heading toward the center. (d) Stage III: The central region contains spermatozoa (S) and the peripheral region contains nutritive phagocytes (NF). (e) Stage V: The partially spawned stage presents spaces that have been vacated by the spawned spermatozoa. (f) Stage VI: spent stage presented large spaces devoid of content (L) with thin ascinal walls and nutritive phagocytes around the periphery. You can have relict spermatozoa. Scale bars: 250 µm.
Figure 3.
Histology sections of testis in Paracentrotus lividus on different stages, through the trial: (a,b) Stage I: recovery stage may has a restored testis with relict spermatozoa (RS) and parallel displays of primary spermatocytes along the ascinal wall (PS) and nutritive phagocytes (NP). (c) Stage II: growing stage displays nutritive phagocytes (NP), primary spermatocytes (PS) along the ascinal wall and columns of spermatocytes (S) heading toward the center. (d) Stage III: The central region contains spermatozoa (S) and the peripheral region contains nutritive phagocytes (NF). (e) Stage V: The partially spawned stage presents spaces that have been vacated by the spawned spermatozoa. (f) Stage VI: spent stage presented large spaces devoid of content (L) with thin ascinal walls and nutritive phagocytes around the periphery. You can have relict spermatozoa. Scale bars: 250 µm.
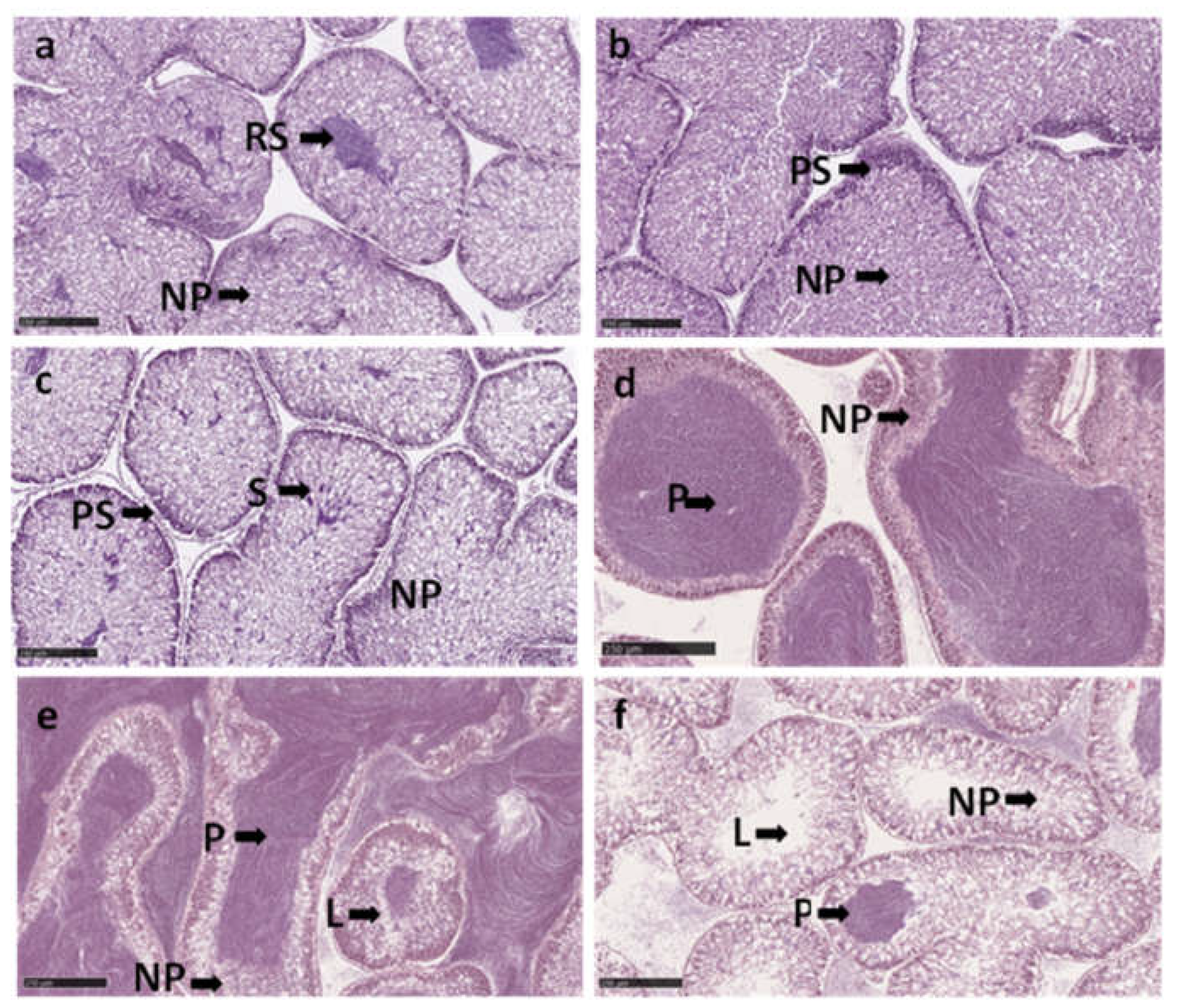
Figure 4.
Histology sections of ovaries in Paracentrotus lividus on different stages, throughout the trial: (a) Stage I: recovery stage ovary seems vacuole due to the formation of a mesh across the ascinus by nutritive phagocytes (NF). During the lysis process, the ovary may contain relyct ova (RO), which the phagocytes may absorb. And presents previtellogenic oocytes (PVO) along the acinal wall and, which range in diameter from 5 to 30 μm. (b) Stage II: growing stage represents the beginning of vitellogenesis. Vitllegenic oocytes (VO) increase in size (10 to 50μm diam) They are found connected on the ascinal wall, surrounded by nutritive phagocytes. (c) Stage III: premature stage, the vitellogeneses continue. The oocytes increase, ranging in size from 10 to 90 μm diameter, with an oval shape and migrate toward the ascinus's center. (d) Stage IV: mature stage, the ovaries contain a dense population of oocytes (O) that can grow up to 90 μm in size, and on the ascinal wall may exist some primary oocytes (PO) that range to 10 μm in size. (e) Stage V: partly spawned stage, the ovaries present space (L) emptied from the release ova. There are cases where oocytes are contained in the ovaries at all stages as mentioned in Stage III and Stage IV. As a result, the females from stages V may seem to be starting to spawn. (f) Stage VI: spent stage the ascinal wall of spend ovaries is thin and sometimes appears empty (L). Ovaries may be entirely devoid of oocytes or contain relyct ova (RO). Reabsorption and phagocytosis are applied to the remaining ova and oocytes. Scale bars: 250 µm.
Figure 4.
Histology sections of ovaries in Paracentrotus lividus on different stages, throughout the trial: (a) Stage I: recovery stage ovary seems vacuole due to the formation of a mesh across the ascinus by nutritive phagocytes (NF). During the lysis process, the ovary may contain relyct ova (RO), which the phagocytes may absorb. And presents previtellogenic oocytes (PVO) along the acinal wall and, which range in diameter from 5 to 30 μm. (b) Stage II: growing stage represents the beginning of vitellogenesis. Vitllegenic oocytes (VO) increase in size (10 to 50μm diam) They are found connected on the ascinal wall, surrounded by nutritive phagocytes. (c) Stage III: premature stage, the vitellogeneses continue. The oocytes increase, ranging in size from 10 to 90 μm diameter, with an oval shape and migrate toward the ascinus's center. (d) Stage IV: mature stage, the ovaries contain a dense population of oocytes (O) that can grow up to 90 μm in size, and on the ascinal wall may exist some primary oocytes (PO) that range to 10 μm in size. (e) Stage V: partly spawned stage, the ovaries present space (L) emptied from the release ova. There are cases where oocytes are contained in the ovaries at all stages as mentioned in Stage III and Stage IV. As a result, the females from stages V may seem to be starting to spawn. (f) Stage VI: spent stage the ascinal wall of spend ovaries is thin and sometimes appears empty (L). Ovaries may be entirely devoid of oocytes or contain relyct ova (RO). Reabsorption and phagocytosis are applied to the remaining ova and oocytes. Scale bars: 250 µm.
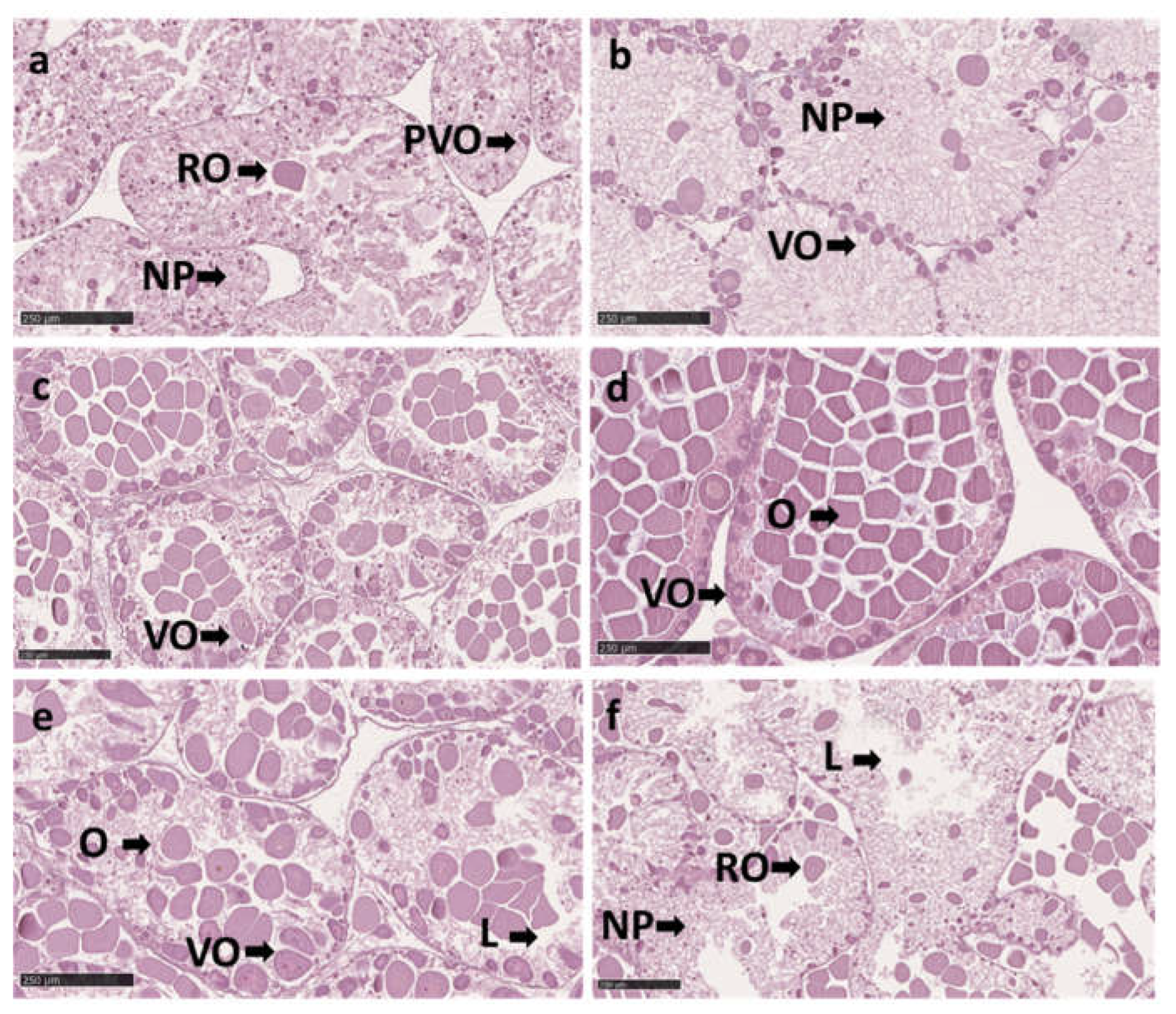
Figure 5.
Development of the gonads of female sea urchins (Paracentrotus lividus) under different conditions (non-manipulated NoM and manipulated M) and different feedings (Ulva spp. U and inert feed If). States of development: recovery stage (I), growing stage (II), premature stage (III), mature stage (IV), partly spawned stage (V), spent stage (VI), according to Byrne (1990).
Figure 5.
Development of the gonads of female sea urchins (Paracentrotus lividus) under different conditions (non-manipulated NoM and manipulated M) and different feedings (Ulva spp. U and inert feed If). States of development: recovery stage (I), growing stage (II), premature stage (III), mature stage (IV), partly spawned stage (V), spent stage (VI), according to Byrne (1990).
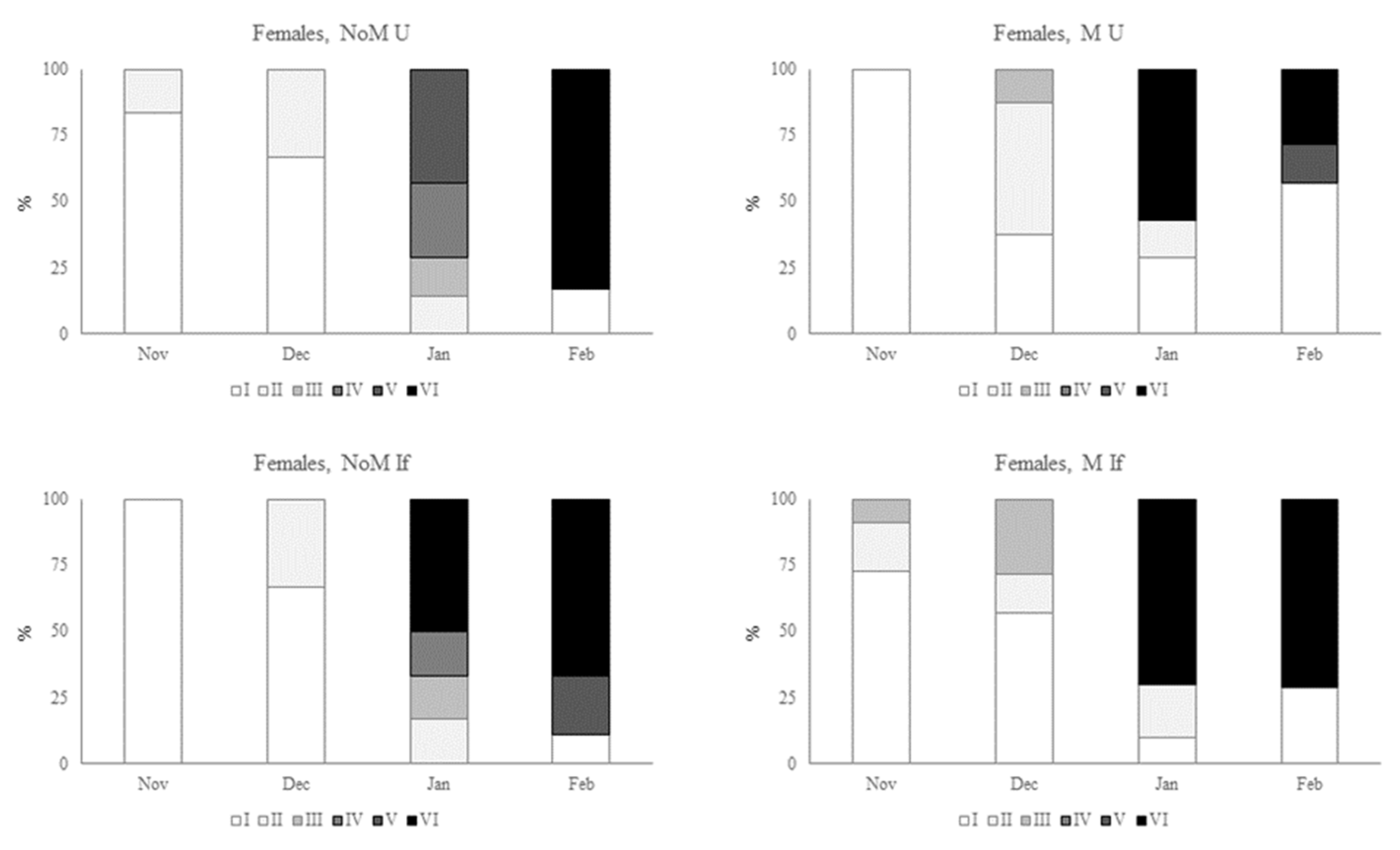
Figure 6.
Development of the gonads of male sea urchins (Paracentrotus lividus) under different conditions (non-manipulated NoM and manipulated M) and different feedings (Ulva spp. U and inert feed If). States of development: recovery stage (I), growing stage (II), premature stage (III), mature stage (IV), partly spawned stage (V), spent stage (VI), according to Byrne (1990).
Figure 6.
Development of the gonads of male sea urchins (Paracentrotus lividus) under different conditions (non-manipulated NoM and manipulated M) and different feedings (Ulva spp. U and inert feed If). States of development: recovery stage (I), growing stage (II), premature stage (III), mature stage (IV), partly spawned stage (V), spent stage (VI), according to Byrne (1990).
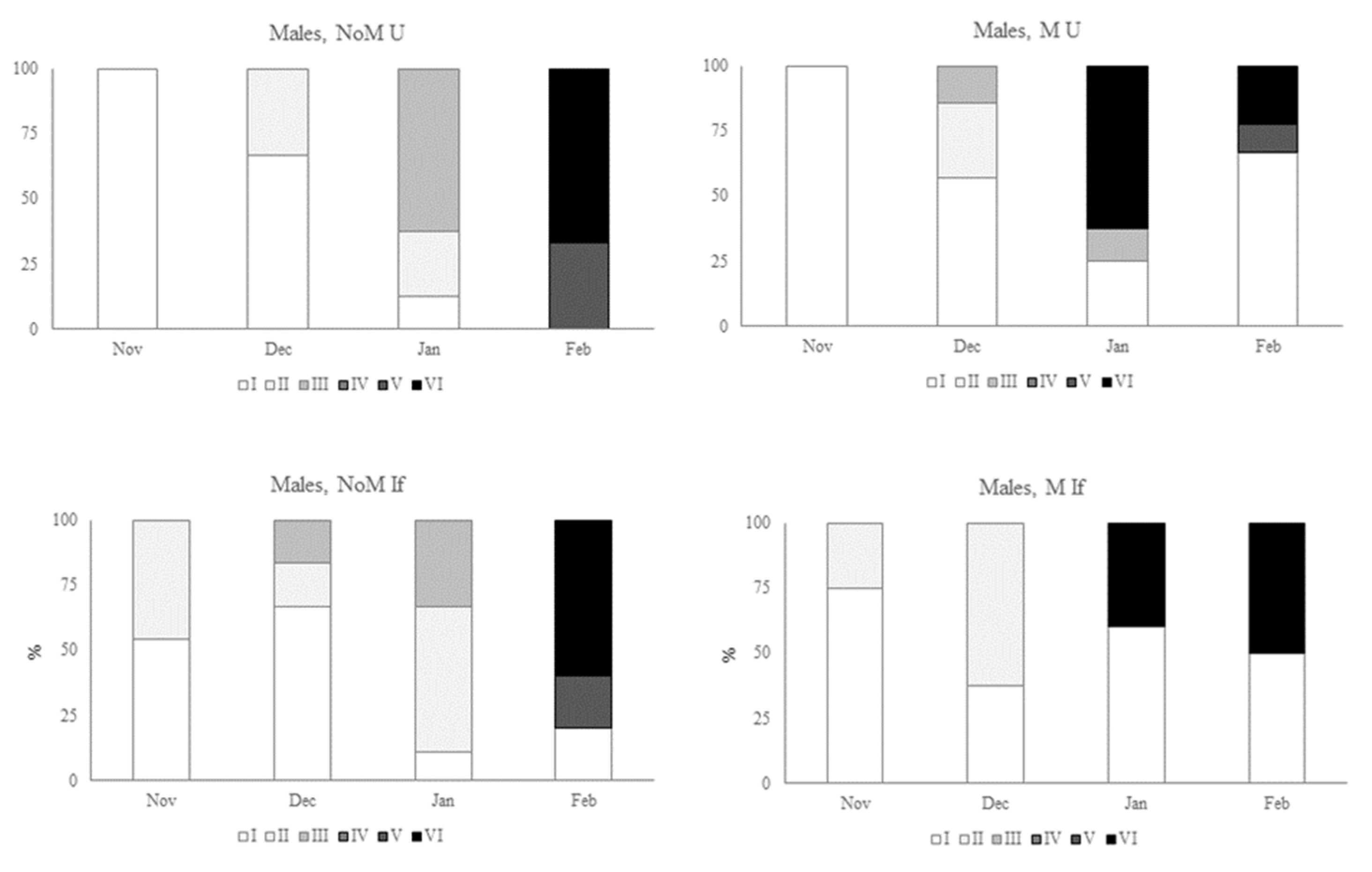

Table 1.
Photoperiod conditions (number of hours of light per day) and temperature (average monthly temperature) for each of the experimental situations (Manipulated and non-manipulated).
Table 1.
Photoperiod conditions (number of hours of light per day) and temperature (average monthly temperature) for each of the experimental situations (Manipulated and non-manipulated).
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



