
Submitted:
08 May 2024
Posted:
09 May 2024
You are already at the latest version
Abstract
Omsk hemorrhagic fever virus (OHFV) is a member of the tick-borne encephalitis virus (TBEV) complex of the Flaviviridae family. Currently, there is no data on cross-reactivity of antibodies to the NS1 proteins of OHFV and TBEV. Such data are of major interest for monitoring viral encephalitis of unknown etiology due to the increasing geographical distribution of OHFV. In this study, recombinant OHFV NS1 protein was produced using Escherichia сoli expression system and purified. The recombinant OHFV NS1 protein was recognized by specific mice immune ascetic fluids to the native OHFV NS1 protein. Western blot analysis and ELISA of recombinant NS1 proteins of OHFV and TBEV were used to study the cross-reactivity of antibodies from immune ascites fluid obtained from OHFV-infected mice and mAbs against TBEV NS1. Anti-TBEV NS1 mouse monoclonal antibodies (mAbs) have been shown not to be cross-reactive to OHFV NS1 protein. Sera from patients with confirmed tick-borne encephalitis (TBE) were examined by ELISA using recombinant OHFV NS1 and TBEV NS1 proteins as antigens. It was shown for the first time that cross-reactive antibodies to OHFV NS1 protein were not detected in the sera of TBE patients whereas the sera contained antibodies to TBEV NS1 protein.
Keywords:
NS1 protein
; Omsk hemorrhagic fever virus
; OHFV
; tick-borne encephalitis virus
; TBEV
; Fla-viviruses
; Flavivirus diagnosis.
1. Introduction
Omsk hemorrhagic fever (OHF) is a zoonotic infectious disease that was registered only in several regions of Russian Siberia until 2022 [1]. The infection is caused by the Omsk hemorrhagic fever virus (OHFV, Orthoflavivirus omskense), which contains a single-stranded (+) RNA genome [2]. OHFV belongs to the flavivirus complex of tick-borne encephalitis virus (TBEV) of the Flaviviridae family. During the incubation period, infected individuals develop nonspecific flu-like symptoms, followed by the appearance of primary specific symptoms, including headache, cough, nausea, chills and muscle pain, as well as gastrointestinal symptoms, subconjunctival bleeding, nasal, gingival and uterine bleeding [3]. Skin hemorrhages then appear, accompanied by fever (39 °C – 40 °C). In addition, the disease leads to skin hyperesthesia and a rash on the upper part of the body. In 30-50% of patients, encephalitic symptoms and meningeal triad are manifested. Diagnosis of this infection is symptomatic and, in the absence of hemorrhagic manifestations, this pathology remains undiagnosed [3]. OHFV is transmitted to humans by bites of ticks from the Dermacentor genus (D. reticulatus and D. marginatus) and Ixodes genus (I. persulcatus), by direct contacts with infected animals, and by airborne or foodborne transmission [4].
In 2022, data on the widespread of OHFV across the territory of the Republic of Kazakhstan appeared. The virus was found in three regions, including Alma-Ata and West Kazakhstan Oblast, which do not border the regions endemic for OHFV in Russian Siberia and are 1000 km or more away from them [1]. For this reason, the spread of OHFV in new areas endemic for TBEV may lead to complications in serologic surveillance of seasonal TBEV activity and diagnosis of patients infected with TBEV. In this regard, it is required to develop test systems for the diagnosis of OHF, including retrospective epidemiological analysis of patients with clinical signs of spring-summer tick-borne meningoencephalitis, in whom TBEV or antibodies to it were not detected.
For the detection of OHFV, as for other flaviviruses, the use of RT-PCR based methods are limited due to the peculiarities of the OHFV life cycle in the human body [5]. In this regard, the diagnosis of OHFV is currently based on clinical features and the presence of cross-reactivity between antibodies against the glycoprotein E of TBEV and OHFV excludes the use of existing serologic kits for differential diagnosis of these infections [6,7]. An additional difficulty for the differential diagnosis of OHF and TBE is the possibility of hemorrhagic forms of TBE, which have been described previously [8].
It is known that the most suitable target for differential serologic diagnosis of flavivirus infections is the non-structural flavivirus protein NS1 [6,9,10]. NS1 is a highly conserved protein with a molecular mass of approximately 46 kDa, which is detected in the serum of flavivirus-infected persons [6,9,11]. Initially, NS1 protein is synthesized intracellularly in a monomeric form; then, it forms a dimeric form upon maturation in the endoplasmic reticulum and is transported to the surface of infected cells [12]. The hexameric form of the NS1 protein of flaviviruses is secreted and circulates in the bloodstream of infected individuals [12]. The intracellular form of NS1 plays a role in viral genome replication and virion maturation, whereas the extracellular hexameric form is capable of triggering various immunologic responses, including the formation of anti-NS1 antibodies [13]. In recent years, NS1 protein has been used as an antigen for early diagnosis of mosquito-borne flavivirus infections by ELISA, as it is detected in infected individuals on day 6-8 [6,9,10]. Both antibodies to this protein and the antigen itself in the serum of an infected person may appear earlier than the immune response to protein E [14]. In addition, test systems based on the NS1 antigen are currently used for epidemiologic diagnosis, including differential diagnosis of the Saint Louis encephalitis and West Nile viruses [15].
The possibility of using the NS1 protein as an ELISA antigen for differential diagnosis of tick-borne flavivirus infections has not been previously investigated. In addition, cross-reactivity of antibodies against the NS1 protein of TBEV and sera from TBE-infected donors to the NS1 protein of OHFV has not been studied. In this study, recombinant OHFV NS1 protein was produced in Escherichia coli and purified. Monoclonal antibodies (mAbs) against TBEV NS1 did not bind the recombinant OHFV NS1 protein, whereas mAbs against the native OHFV NS1 recognized it. In addition, cross-reactive antibodies against OHFV NS1 were not found in the sera of patients with confirmed TBE.
2. Materials and Methods
2.1. Sera from Patients
Sera from patients (54% males and 46% females) hospitalized in Novosibirsk Infectious Diseases Clinical Hospital No. 1 between April and September 2017-2019 with TBE and sera from healthy volunteers were used in this study. These serum samples have been previously characterized for the presence of anti-TBEV antibodies [16]. Patient selection criteria were as follows: (i) history of tick bites; (ii) presence of symptoms of infectious diseases; (iii) patient age more than 18 years. All patients underwent appropriate medical examinations, including serological detection of TBE. Volunteers were young healthy adults without chronic diseases (including autoimmune diseases) who had not been infected and/or hospitalized for at least six months. All patients and volunteers participating in this study gave informed consent. The study was approved by the ethical committee of the Novosibirsk Infectious Diseases Clinical Hospital No. 1. Sera were stored at -70 °C. These serum samples have been previously characterized for the presence of anti-TBEV protein E antibodies by ELISA and for TBEV RNA by RT-PCR [16].
2.2. Phylogenetic Analyses of the OHFV NS1 Gene and in silico Structural Analysis of NS1 protein of the OHFV Strain 186-1964
The OHFV NS1 gene was amplified by reverse transcription reaction using OT-M-MuLV-RH reagent kit (Biolabmix, Novosibirsk, Russia) with random hexaprimers; total RNA isolated from the inactivated brain lysate of BALB/c mouse infected with OHFV, strain 186-1964 was used as a template. PCR fragments encoding the NS1 protein of OHFV (1 - 352 aa) was obtained using primers Start_NS1_OHFV_59_pET_U26 5'-CCGTTGGATCCGACGTTGGATGTGCTGTGTGGACACTGA-3' and End_NS1_OHFV_58_pET_L21 5'-GGCTTTGAATTCCCAGCCACCACCACCATCGAGCGCGCAC-3' and cDNA as a matrix. For phylogenetic analyses, the sequence of the NS1 gene of OHFV, strain 186-1964 was determined and aligned with those of other OHFV strains and several TBEV strains including the European, Far-Eastern, and Siberian subtypes via the MUSCLE algorithm in MEGA X [17]. The maximum likelihood phylogenetic tree was constructed using TN93+G+I substitution model with 1000 bootstrap iterations in MEGA X.
The putative three-dimensional (3D) structure of OHFV NS1 protein was predicted using the AlphaFold2 algorithm [18] (https://colab.research.google.com/github/sokrypton/ColabFold/blob/main/AlphaFold2.ipynb, accessed September 11-14, 2023). The UCSF Chimera molecular visualizer, version 1.15 was used to visualize ribbon and surface representations OHFV NS1 protein [19].
2.3. Construction of the Plasmid Encoding the OHFV NS1 Protein
Bacterial strains E. coli XL1-Blue and E. coli BL21 (DE3) that were used to construct and produce the target proteins were obtained from the Collection of Extremophile Microorganisms and Type Cultures (CEMTC) of ICBFM SB RAS.
The obtained PCR fragment containing the OHFV NS1 gene and the expression plasmid pET-32a(+)were cleaved by the restriction endonucleases BamHI and EcoRI (Sibenzyme, Novosibirsk, Russia) and combined in a ligation reaction. E. coli XL1-Blue cells (recA1, endA1, gyrA96, thi, hsdR17 (rK-, mK+), supE44, relA1, lac, [F′, proAB+, laclqZΔM15, Tn10 (Tetr)]) were transformed with the resulting ligation products and seeded on LB-agar with ampicillin at a dose of 50 μg/ml and cultured. Individual colonies of E. coli cells containing plasmid pET-32a(+)_NS1OHFV were tested by PCR using the same primers. PCR amplification conditions were as follows: 5 min at 95 °C, followed by 30 cycles of 30 s at 95 °C, 20 s at 56 °C, 1.5 min at 72 °C, and a final elongation of 6 min at 72 °C. Obtained PCR products were assessed by electrophoresis in 1% agarose gel. The accuracy of the insertion of the gene encoding the OHFV NS1 protein was confirmed by Sanger sequencing using primers pETseq-dir 5'-TGCTAGTTAGTAGTATTGCTCAGCG-3' and pETseq-rev 5'-GGTTCTGGTTCTGGTTCTGGTTCTGGCCATA-3'. The resulting plasmid was pET-32a(+)_NS1OHFV, which encodes the OHFV NS1 protein with a His-tag at the C-terminus.
In the recombinant expression plasmid pET-32a(+)_NS1OHFV, the NS1 gene is located directly after the Trx coding sequence in the same open reading frame. Meanwhile, the sequence encoding the 6His-tag, which is required for purification using metal-chelate chromatography, is located directly after the gene. Thus, the expressed recombinant NS1 protein contains a Trx tag at the N-terminus, a 6His tag at the C-terminus, and a thrombin cleavage site between Trx and NS1 sequences.
2.4. Optimization of the OHFV NS1 Expression
E. coli strain BL21 (DE3) cells (F-, ompT, hsdSB (rB-, mB-), dcm, gal, λ(DE3), pLysS, Cmr) were transformed with the plasmid pET-32a(+)_NS1OHFV, cultured in LB medium in the presence of ampicillin at a concentration of 50 μg/ml. When cell culture reached OD600 = 0.5, the isopropyl β-D-1-thiogalactropyranoside (IPTG) at a concentration of 100 μM was added. After induction, cells were grown at 30 °C overnight. Production of recombinant OHFV NS1 protein in cell lysates was assessed using 12.5% PAGE.
To select the optimal conditions for OHFV NS1 protein production, different IPTG concentrations (1 μM, 10 μM, 100 μM, and 1000 μM) were used to induce the transcription of the target gene when cell culture reached OD600 = 0.5 and the cells were cultivated at different temperatures varying from 12 °C to 30 °C. At 4 h and 11 h after induction, cells were centrifuged at 3000 g for 10 min. The cell pellet was resuspended in 50 mM Tris-HCl, pH 8.0, and disrupted using the ultrasonic homogenizer Sonopuls hd 2070 (Bandelin, Berlin, Germany) for 2 minutes at 30% amplitude, with 10 second of sonication and 10 second of rest period. The obtained suspension was centrifuged at 16,000 g for 10 minutes, after which the supernatant containing soluble cytoplasmic proteins, was transferred to a new tube. The precipitate containing the inclusion body fraction was dissolved in 50 mM Tris-HCl, pH 8.0. The resulting cell fractions were analyzed by 12.5% PAGE under reducing conditions.
2.5. Purification of the Recombinant OHFV NS1 Protein
E. coli BL21 (DE3)/pET-32a(+)_NS1OHFV cells were cultivated under the previously selected conditions and centrifuged at 9000 g for 30 minutes. The precipitate was dissolved in a wash buffer containing 1 mM phenylmethylsulfonyl fluoride (PMSF), 2 M urea, 20 mM Tris-HCl and 500 mM NaCl, sonicated for 15 minutes as described previously, and centrifuged at 15,000 g for 10 minutes. Next, the precipitate was dissolved in the inclusion body solubilizing buffer (pH 7.4, 20 mM Tris-HCl, 6 M urea, 1 mM β-ME, 5 mM imidazole, 500 mM NaCl), sonicated as described above, and centrifuged at 12,000 g for 15 min. Recombinant OHFV NS1 protein was purified from inclusion bodies using Ni-NTA Sepharose (Qiagen, Venlo, The Netherlands), according to the manufacturer's protocol. The column containing Ni-NTA agarose was equilibrated with a buffer B (3 M urea, 500 mM NaCl, 20 mM Tris-HCl, 1 mM β-ME, pH 8.0). The supernatant containing solubilized inclusion bodies was applied to the column and washed with the buffer B. The recombinant OHFV NS1 protein was eluted with buffer B, containing 500 mM imidazole, and concentrated using an Amicon Ultra-4 centrifuge filter unit (Millipore, Burlington, MA, USA) with a cut-off threshold of 10 kDa. The recombinant OHFV NS1 protein was consistently dialyzed against three refolding buffers (300 mM NaCl, 200 mM sucrose, 1 mM reduced glutathione (GSH), 0.2 mM oxidized glutathione (GSSG) and 0.1% Triton X-100, pH 7.5.) containing consistently decreasing concentrations of urea (2 M, 0.5 M, and 0 M urea, respectively) and imidazole (250 mM, 100 mM and 20 mM, respectively). Finally, the purified recombinant OHFV NS1 protein was dialyzed in a storage buffer (50 mM Tris-HCl, pH 8.0, containing 10 mM imidazole, 300 mM NaCl, and 200 mM sucrose). The purified recombinant OHFV NS1 protein was analyzed using 12.5% PAGE. Protein purity was determined using Gel Doc XR+ gel documentation system (Bio-Rad, Hercules, CA, USA) with ImageLab 3.0 software (Bio-Rad, Hercules, CA, USA). Protein concentration was evaluated using the Qubit protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA) on a Qubit 4 fluorometer (Thermo Fisher Scientific, Waltham, MA, USA). The obtained proteins were stored at a concentration of at least 0.5 mg/ml.
2.6. Western Blot Analysis
The lysates of E. coli cells producing the OHFV NS1 protein were separated using 12.5% PAGE and then transfer to a nitrocellulose membrane (Bio-Rad, Hercules, CA, USA). After blocking the nonspecific binding sites with 3% bovine serum albumin solution (BSA, Amresco, Solon, OH, USA), the membrane was incubated with sera obtained from TBE patients at a dilution of 1:800 (N=26), or with 0.02 mg anti-TBEV NS1 protein monoclonal antibodies (N=5). The membrane was then incubated with Anti-Mouse IgG (Fc specific)–Peroxidase conjugated polyclonal antibodies produced in rabbits (Biosan, Novosibirsk, Russia). Immune complexes were revealed using 4-chloro-1-naphthol (Applichem, Darmstadt, Germany). Immune ascitic pooled fluids obtained from OHFV-infected mice was used as a positive control.
2.7. ELISA
For indirect ELISA, 1 μg/ml of anti-His tag mAb.H1 (Biolabmix, Novosibirsk, Russia) was sorbed into the wells of 96-well polystyrene plates (Greiner, Kremsmünster, Austria); then, after blocking the non-specific binding sites with 5% skim milk solution, 1 μg of OHFV NS1 protein or TBEV NS1 protein (as a positive control) was added. After incubation, sera obtained from TBE-positive patients at a dilution of 1:500 (N=26) or individual mAb against TBEV NS1 protein (N=5) at a concentration of 10 μg/ml were added. After washing, wells were incubated with Anti-Mouse IgG (Fc specific) HRP conjugated polyclonal antibodies produced in rabbits (Biosan, Novosibirsk, Russia) or Anti-Human IgG (Fc specific) HRP conjugated mAb X-53 (Biosan, Novosibirsk, Russia) for one hour at 37 °C. Immune complexes were revealed using tetramethylbenzidine-3,3,5,5 (TMB, Applichem, Solon, OH, Germany). Absorbance was measured at a wavelength of 450 nm using a microplate reader (Bio-Rad, Hercules, CA, USA).
To determine the cutoff level for ELISA with sera from TBE patients, sera obtained from conditionally healthy donors who had not previously had TBE were used. The mean optical density level of the control sera plus one standard deviation was taken as cutoff. The ELISA signal levels of sera from TBE patients above the cutoff were considered positive.
2.8. Statistics
Statistical analysis was carried out using one-way ANOVA using the Statistica 10 software package (StatSoft Inc., Tulsa, OK, USA).
3. Results
3.1. Phylogenetic Analysis and in silico Modelling of the OHFV NS1 Protein
Sequencing of the gene encoding OHFV NS1 protein, strain 186-1964, and subsequent phylogenetic analysis showed that the sequence of this gene clustered with NS1 gene sequences of previously described OHFV strains (Figure 1). The nucleotide identity (NI) of the investigated NS1 gene with the closest published strains of OHFV Kubrin and OHFV Bogoluvovska was 98.2% (19 nucleotide substitutions). This resulted in only three amino acid (aa) substitutions, two of which did not change the class of the aa residue (Figure 2A). Expectedly, the sequence similarity of the NS1 gene sequence of OHFV 186-1964 to the corresponding TBEV genes was lower and the NI value was 80.1% (210 nucleotide substitutions, Figure 2B).
It is known that the flaviviral NS1 protein contains three domains: the β-roll domain, 1-29 aa; the wing domain, 30-180 aa; and the β-ladder domain, 181-352 aa [20]. It was shown that "β-roll" participates in NS1 dimerization and is hidden inside the NS1 dimer, while aa residues from other domains are exposed on the surface of the NS1 dimer. Notably, all aa residues, which are different between OHFV NS1 and TBEV NS1 are located predominantly in the wing and β-ladder domains (21 and 15 aa, respectively) exposed on the surface of the NS1 dimer (Figure 2B).
To analyze the arrangement of aa substitutions on the surface of NS1 OHFV, the 3D structures of the OHFV NS1 and TBEV NS1 proteins were predicted using the AlfaFold2 algorithm [18]. The modeling results (Figure 3) indicated that the OHFV NS1 protein has a structure similar to the resolved structures of the NS1 proteins of Zika virus, West Nile virus, Japanese encephalitis virus, and Dengue virus, for which the 3D structures were previously obtained [21,22,23]. The NS1 proteins of OHFV form a homodimer, each consisting of three domains. As shown above, all differences in aa residues are in domains 2 and 3, which are marked in red in Figure 3. Almost all of the differing aa are located on the surface of the OHFV NS1 protein and may form different epitopes for antibodies than TBEV NS1. Thus, this analysis indicates that the OHFV NS1 protein could potentially be used for the differential diagnosis of OHF and TBE.
NS1 is an important flavivirus protein that plays a central role in efficient viral RNA replication. This protein accumulates intracellularly and is secreted from infected cells. Soluble hexamer of the flaviviral NS1 protein circulates in serum and other body fluids and therefore can be detected. Since antibodies against this protein are produced during, they can be used for serologic diagnostics.
3.2. Production of the OHFV NS1 Protein in E. coli Cells
In the recombinant expression plasmid pET-32a(+)_NS1OHFV, the NS1 gene (GenBank accession number PP525060) is located directly after the Trx coding sequence in the same open reading frame. Meanwhile, the sequence encoding the 6His-tag, which is required for purification using metal-chelate chromatography, is located directly after the gene. Thus, the expressed recombinant NS1 protein contains a Trx tag at the N-terminus, a 6His tag at the C-terminus, and a thrombin cleavage site between Trx and NS1 sequences.
To obtain recombinant OHFV NS1 protein, E. coli BL21 (DE3) cells were transformed with the obtained plasmid pET-32a(+)_NS1OHFV and induced. The lysates of induced BL21 (DE3) / pET-32a(+)_NS1OHFV cells were fractionated using 12.5% SDS- PAGE. The electrophoretic mobility of the obtained recombinant OHFV NS1 corresponded to the theoretically predicted ~ 58 kDa, and also corresponded to the electrophoretic mobility of recombinant TBEV NS1 obtained previously using a similar plasmid vector [24]. The OHFV NS1 protein was found to be located in the inclusion body fraction (Figure 4).
The production of the recombinant OHFV NS1 protein in either E. coli cells or eukaryotic expression systems has not been previously described. To increase the solubility of the recombinant protein, we optimized the cultivation conditions similar to those previously used to obtain soluble TBEV NS1 protein in E. coli [24]. For this purpose, E. coli BL21 (DE3) cells transformed with the pET-32a(+)-NS1OHFV plasmid were grown at various IPTG concentrations, temperature, and duration of cultivation after induction. The results showed that induction of the transformed cell culture with 0.01 mM IPTG for 11 h at 12 °C resulted in the maximum yield of soluble OHFV NS1 protein. Induction of the transformed culture with 0.1 mM IPTG for 11 h at 30 °C resulted in maximum yield of OHFV NS1 protein; however, almost all of the NS1 protein was localized in the inclusion body fraction (Figure 5). The yield of soluble OHFV NS1 protein was low, so refolded protein from the inclusion body fraction was obtained for further studies.
3.3. OHFV NS1 Protein Isolation and Purification
Protein purification was performed using Ni-NTA agarose from the inclusion body fraction, similar to the previously described protocol for the TBEV NS1 [14]. Buffers with increasing concentrations of urea were used to isolate the OHFV NS1 protein from inclusion bodies (Figure 6). The maximum dissolved protein content was observed in a wash buffer containing 6 M urea (Figure 6). The NS1 protein was purified by metal-chelate chromatography on Ni-NTA agarose under denaturing conditions, bound protein was eluted with buffer A containing 500 mM imidazole (Figure 6). The OHFV NS1 protein was dialyzed with a refolding buffer containing reduced (GSH) and oxidized (GSSG) glutathione and sucrose to promote the acquisition of the correct conformation of the OHFV NS1. The productivity was 80 mg of purified OHFV NS1 protein per 1 liter of cell culture. The purity of the resulting purified OHFV NS1 protein, assessed after PAGE was about 90%. The electrophoretic mobility of the purified recombinant OHFV NS1 protein was ~58 kDa.
It has been shown that mAb.His1 against the C-terminal His-tag (Biosan, Russia) reveal in the lysate of E. coli BL21 (DE3) / pET-32a(+)-NS1OHFV cells a protein with a molecular mass of about 58 kDa, which corresponds to the recombinant OHFV NS1 protein (Figure 7). The TBEV NS1 protein, also possessing a C-terminal His-tag [14], which was previously shown to be detected by the mAb.His1, was used as an antigen control.
3.4. Evaluation of the Antigenic Properties of the Recombinant OHFV NS1 Protein
The antigenic properties of the obtained OHFV NS1 protein were evaluated using ELISA. For this purpose, immune ascites fluids (anti-OHFV IAF) were used that were obtained from mice independently infected with two different OHFV strains (OHFV P-15-2213 and OHFV Oz-31_Kd_10866) [4,25]. The NS1 protein of TBEV was used as a negative control antigen. Serial dilutions of two anti-OHFV IAFs were tested for their binding to the recombinant OHFV NS1 and TBEV NS1 proteins using ELISA. Both anti-OHFV IAFs bound recombinant OHFV NS1 protein at a dilution of at least 1:100,000 (1:4,500,000 for the strain P-15-2213), whereas they revealed recombinant TBEV NS1 protein at a dilution of 1:500 (Figure 8). Thus, it was demonstrated that the obtained recombinant OHFV NS1 protein retained its antigenic properties after refolding.
Western blot analysis indicated that both anti-OHFV IAFs and the anti-His-tag mAb.His1 revealed a protein with a molecular mass of approximately 58 kDa, which corresponds to the recombinant OHFV NS1 protein (Figure 7). None of the IAFs, unlike the mAb.His1, detected recombinant TBEV NS1 protein. This is probably due to the small number of similar epitopes on the surface of OHFV NS1 and TBEV NS1 (Figure 7). So, after refolding, the resulting recombinant OHFV NS1 protein retained its conformation epitopes recognizable by specific antibodies against the native OHFV NS1.
The ability of TBEV NS1-specific mAbs FVN-NS1-3, FVN-NS1-6, FVN-NS1-44, FVN-NS1-290 and FVN-NS1-299 to bind the recombinant OHFV NS1 protein was tested. These mAbs have been previously obtained and characterized [24] and all the mAbs did not demonstrate. In parallel experiments, equal concentrations of OHFV NS1 and TBEV NS1 proteins were detected by serial dilutions of the mAb. It was shown that these mAbs recognized only TBEV NS1 and did not bind NS1 OHFV. The obtained data indicate that the epitopes that were bound by mAbs FVN-NS1-3, FVN-NS1-6, FVN-NS1-44, FVN-NS1-290 and FVN-NS1-299 were absent on the OHFV NS1 surface (Figure 9). Thus, it was confirmed that purified OHFV NS1 protein is retained its antigenic properties and its antigenic structure is different from that of TBEV NS1, which provides the possibility of using the recombinant OHFV NS1 for differential diagnosis of TBE and OHV infection.
3.5. Examination of Sera from Patients with TBE for Cross-Reactivity with the OHFV NS1 protein
A panel of sera obtained from patients with confirmed TBE was examined for the ability to detect OHFV NS1. At the first step, the cutoff was determined. For this purpose, a panel of sera (N = 96) obtained from presumptively healthy volunteers who had no TBE and OHF previously (group 1) was tested. OHFV NS1 and TBEV NS1 were used as antigens. The cutoff value was 0.494. Then, 26 sera obtained from patients with confirmed TBE (group 2) were tested for antibodies to TBEV NS1. Since two serum samples showed low signal (below cutoff), 24 TBEV NS1-positive sera were selected for further analysis. These positive sera were tested by ELISA for the presence of cross-reactive antibodies to the OHFV NS1 protein. The result indicated that only in one serum the signal in ELISA was above the cutoff level, in another one the antibody level was equal to the cutoff, corresponding to a questionable result (Figure 10). Thus, the specificity of ELISA using recombinant the recombinant OHFV NS1 protein for differential diagnosis of TBE and OHFV was at least 90%.
4. Discussion
OHFV is a rare diagnosed virus; the first recorded outbreak was observed in Siberia region of USSR between 1945 and 1958, with 972 confirmed cases and more than 1500 cases of suspected OHF. Until 1998, more than 300 additional cases were recorded in the Tyumen, Kurgan, Omsk and Novosibirsk regions of the Russian Federation. After 1998, no new cases of OHF have been reported in Russia and other countries, which is probably due to both the lack of specific diagnostics and the probable mild course of the disease [12]. Another likely reason for the absence of new OHF cases is routine TBEV vaccinations. It is known that protective immunity against TBEV can also protect a vaccinated person from OHFV [3]. In 2022, OHF was unexpectedly detected in the new regions of the Republic of Kazakhstan, located more than 1000 km from OHF-endemic areas in Russia. In this regard, it is required to strengthen the epidemiological control of OHFV.
The correct diagnostics of Flavivirus infections is difficult, especially in regions where more than one flavivirus are circulating. Cross-reactivity of antibodies to the E glycoprotein of closely related mosquito-borne flaviviruses, such as Dengue virus and Zika virus, is known to cause low specificity of ELISA and serological tests based on the detection of E glycoprotein or antibodies to it [26]. Because of the significant aa differences in the NS1 proteins of these viruses, several ELISA kits have been developed for the detection of antibodies to the NS1 protein of Zika virus and such test systems showed high specificity [26,27,28,29]. In addition, sera from patients with previous infections caused by the Dengue virus, West Nile virus, yellow fever virus, and Japanese encephalitis virus have been shown to show little or no detectable cross-reactivity to Zika virus NS1, in contrast to ELISAs based on the detection of antibodies to the Zika virus glycoprotein E [26]. The development of tests for detecting the NS1 protein of the Dengue virus in serum has made it possible to easily and affordably diagnose patients with Dengue fever in the acute phase [30]. However, there are currently no clinical tests for the detection of the NS1 protein of other flaviviruses, as it has previously been shown that NS1 secretion in flavivirus infection is low [31,32]. Nevertheless, the time interval at which the NS1 protein of West Nile virus can be detected in serum is similar to the time interval for the detection of genomic RNA of this virus by RT-PCR.
The data obtained in this work also confirm the promising use ELISA kits based on the NS1 proteins of flaviviruses from the TBEV complex for differential diagnosis, including monitoring of viral encephalitis of unknown etiology. The development of more sensitive methods of NS1 protein detection, capable of detecting it even in low concentrations, may also allow the use of such test systems for the diagnosis of flaviviruses, including those from the tick-borne encephalitis group.
5. Conclusions
Currently, there is no reliable serological diagnosis to differentiate each virus of the Flavivirus family is optimal, and the introduction of NS1-based serology can provide more accurate diagnosis and better epidemiologic support for the global spread of the flaviviruses.
In this work, the refolded recombinant OHFV NS1 protein was obtained for the first time, which retains antigenic properties similar to the native OHFV NS1 protein. It was indicated that IAFs from OHFV-infected mice did not recognize the TBEV NS1 protein and mAbs to TBEV NS1 did not have cross-reactivity to OHFV NS1. In addition, it was shown for the first time that no cross-reactive antibodies to the NS1 protein of OHFV were detected in the sera of TBE patients containing antibodies to the NS1 protein of TBEV. Thus, this study indicated that the OHFV NS1 protein has an antigenic structure different from TBEV NS1 and could be potentially used as an antigen for the differential diagnosis of TBE and OHF.
Author Contributions
Conceptualization, Y.A.K. and A.L.M.; methodology, B.I.K., Y.A.K. and A.L.M.; software, B.I.K., Y.A.K. and A.L.M.; validation, Y.A.K., and B.I.K.; formal analysis, B.I.K., Y.A.K. and A.L.M.; investigation, B.I.K., Y.A.K., G.S.C., V.V.Y., E.I.K. and A.L.M.; resources, Y.A.K., N.V.T.; data curation, Y.A.K. and B.I.K.; writing—original draft preparation, B.I.K., Y.A.K., N.V.T. and A.L.M.; writing—review and editing, A.L.M. and N.V.T.; visualization, B.I.K. and A.L.M.; supervision, A.L.M.; project administration, Y.A.K. and N.V.T.; funding acquisition, Y.A.K. and N.V.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant number 22-74-10103, https://rscf.ru/project/22-74-10103/. Escherichia coli strain was obtained from the Collection of Extremophile Microorganisms and Type Cultures of ICBFM SB RAS, which is supported by the Ministry of Education and Science, Project No. 121031300043-8.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the ethical committee of the Novosibirsk Infectious Diseases Clinical Hospital No. 1 (protocol code №3, date of approval 15.07.2016). The animal study protocol was approved by the Bioethical Committee of the N.N.Vorozhtsov Novosibirsk Institute of Organic Chemistry (Resolution Number P-05-08.2022-14, date of approval 05.08.2022).
Informed Consent Statement
“Informed consent was obtained from all patients and volunteers involved in the study.”
Data Availability Statement
Not applicable.
Acknowledgments
The authors express their gratitude to Nikita A. Dubovitsky, junior researcher oof Laboratory of Molecular Epidemiology and Virus Biodiversity, FIC FTM, for his invaluable contribution to the study of OHFV phylogeny and invaluable advice for the design of phylogenetic trees.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wagner, E., Shin, A., Tukhanova, N., Turebekov, N., Nurmakhanov, T., Sutyagin, V., Berdibekov, A., Maikanov, N., Lezdinsh, I., Shapiyeva, Z., Shevtsov, A., Freimüller, K., Peintner, L., Ehrhardt, C., Essbauer, S. First Indications of Omsk Haemorrhagic Fever Virus beyond Russia. Viruses 2022, 14(4), 754. [CrossRef]
- Lin, D., Li, L., Dick, D., Shope, R.E., Feldmann, H., Barrett, A.D., Holbrook, M.R. Analysis of the complete genome of the tick-borne flavivirus Omsk hemorrhagic fever virus. Virology 2003, 313(1), 81–90. [CrossRef]
- Růžek, D., Yakimenko, V.V., Karan, L.S., Tkachev, S.E. Omsk haemorrhagic fever. Lancet (London, England). 2010, 376(9758), 2104–2113. [CrossRef]
- Karan, L.S., Ciccozzi, M., Yakimenko, V.V., Presti, A.L., Cella, E., Zehender, G, Rezza, G., Platonov, A. E. The deduced evolution history of Omsk hemorrhagic fever virus. Journal of Medical Virology 2014, 86(7), 1181–1187. [CrossRef]
- Földvári, G., Široký, P., Szekeres, S., Majoros, G., Sprong, H. Dermacentor reticulatus: a vector on the rise. Parasites & Vectors. 2016, 9, 314. [CrossRef]
- Fisher, R., Lustig, Y., Sklan, E.H., Schwartz, E. The Role of NS1 Protein in the Diagnosis of Flavivirus Infections. Viruses 2023, 15(2), 572. [CrossRef]
- Tsekhanovskaya, N.A., Matveev, L.E., Rubin, S.G., Karavanov, A.S., Pressman, E.K. Epitope analysis of tick-borne encephalitis (TBE) complex viruses using monoclonal antibodies to envelope glycoprotein of TBE virus (persulcatus subtype). Virus Research 1993, 30(1), 1–16. [CrossRef]
- Ternovoi, V.A., Kurzhukov, G.P., Sokolov, Y.V., Ivanov, G.Y., Ivanisenko, V.A., Loktev, A.V., Ryder, R.W., Netesov, S.V., Loktev, V.B. Tick-Borne Encephalitis with Hemorrhagic Syndrome, Novosibirsk Region, Russia, 1999. Emerging Infectious Diseases. 2003, 9(6), 743–746. [CrossRef]
- Muller, D.A., Young, P.R. The flavivirus NS1 protein: molecular and structural biology, immunology, role in pathogenesis and application as a diagnostic biomarker. Antiviral Research 2013, 98(2), 192–208. [CrossRef]
- Chong, H.Y., Leow, C.Y., Abdul Majeed, A.B., Leow, C.H. Flavivirus infection-A review of immunopathogenesis, immunological response, and immunodiagnosis. Virus Research 2019, 274, 197770. [CrossRef]
- Rastogi, M., Sharma, N., Singh, S.K. Flavivirus NS1: a multifaceted enigmatic viral protein. Virology Journal 2016, 13, 131. [CrossRef]
- Crooks, A.J., Lee, J.M., Easterbrook, L.M., Timofeev, A.V., Stephenson, J.R. The NS1 protein of tick-borne encephalitis virus forms multimeric species upon secretion from the host cell. J. Gen. Virol. 1994, 75, 3453-3460. [CrossRef]
- Carpio, K.L., Barrett, A.D.T. Flavivirus NS1 and Its Potential in Vaccine Development. Vaccines (Basel) 2021, 9(6), 622. [CrossRef]
- Oceguera, L.F., Patiris, P.J., Chiles, R.E., Busch, M.P., Tobler, L.H., Hanson, C.V. Flavivirus Serology by Western Blot Analysis. The American Journal of Tropical Medicine and Hygiene 2007, 77(1), 159–163. [CrossRef]
- Lorch, M. S., Collado, M. S., Argüelles, M. H., Rota, R. P., Spinsanti, L. I., Lozano, M. E., Goñi, S. E. Production of recombinant NS1 protein and its possible use in encephalitic flavivirus differential diagnosis. Protein Expression and Purification 2019, 153, 18–25. [CrossRef]
- Igolkina, Y., Rar, V., Krasnova, E., Filimonova, E., Tikunov, A., Epikhina, T., Tikunova, N. Occurrence and clinical manifestations of tick-borne rickettsioses in Western Siberia: First Russian cases of Rickettsia aeschlimannii and Rickettsia slovaca infections. Ticks and Tick-borne Diseases 2022, 13. 101927. 10.1016/j.ttbdis.2022.101927.
- Kumar, S., Stecher, G., Li, M., Knyaz, C., Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol. 2018, 35(6), 1547-1549. [CrossRef]
- Mirdita, M., Schütze, K., Moriwaki, Y., Heo, L., Ovchinnikov, S., Steinegger, M. ColabFold: making protein folding accessible to all. Nature Methods 2022, 19(6), 679–682. [CrossRef]
- Pettersen, E.F., Goddard, T.D., Huang, C.C., Couch, G. S., Greenblatt, D. M., Meng, E. C., Ferrin, T. E. UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem. 2004, 25(13), 1605-1612. [CrossRef]
- Akey, D.L., Brown, W.C., Jose, J., Kuhn, R.J., Smith, J.L. Structure-guided insights on the role of NS1 in flavivirus infection. BioEssays: news and reviews in molecular, cellular and developmental biology 2015, 37(5), 489–494. [CrossRef]
- Brown, W. C., Akey, D. L., Konwerski, J. R., Tarrasch, J. T., Skiniotis, G., Kuhn, R. J., Smith, J. L. Extended surface for membrane association in Zika virus NS1 structure. Nature Structural & Molecular Biology 2016, 23(9), 865–867. [CrossRef]
- Akey, D. L., Brown, W. C., Dutta, S., Konwerski, J., Jose, J., Jurkiw, T. J., DelProposto, J., Ogata, C. M., Skiniotis, G., Kuhn, R. J., Smith, J. L. Flavivirus NS1 Structures Reveal Surfaces for Associations with Membranes and the Immune System. Science 2014, 343(6173), 881–885. [CrossRef]
- Edeling, M.A., Diamond, M.S., Fremont, D.H. Structural basis of Flavivirus NS1 assembly and antibody recognition. Proceedings of the National Academy of Sciences 2014, 111(11), 4285–4290. [CrossRef]
- Andrey M, Yana K, Olga G, Bogdana K, Sergey T, Lyudmila E, et al. Tick-borne encephalitis nonstructural protein NS1 expressed in E. coli retains immunological properties of the native protein. Protein Expression and Purification 2022, 191, 106031. [CrossRef]
- Kovalev, S.Y., Mazurina, E.A., Yakimenko, V.V. Molecular variability and genetic structure of Omsk hemorrhagic fever virus, based on analysis of the complete genome sequences. Ticks and Tick-borne Diseases 2021, 12(2), 101627. [CrossRef]
- Steinhagen, K., Probst, C., Radzimski, C., Schmidt-Chanasit, J., Emmerich, P., van Esbroeck, M., Schinkel, J., Grobusch, M. P., Goorhuis, A., Warnecke, J. M., Lattwein, E., Komorowski, L., Deerberg, A., Saschenbrecker, S., Stöcker, W., and Schlumberger, W. Serodiagnosis of Zika virus (ZIKV) infections by a novel NS1-based ELISA devoid of cross-reactivity with dengue virus antibodies: a multicohort study of assay performance, 2015 to 2016. Euro Surveillance: Bulletin Europeen Sur Les Maladies Transmissibles = European Communicable Disease Bulletin 2016, 21(50), 30426. [CrossRef]
- Pasquier, C., Joguet, G., Mengelle, C., Chapuy-Regaud, S., Pavili, L., Prisant, N., Izopet, J., Bujan, L., and Mansuy, J. M. Kinetics of anti-ZIKV antibodies after Zika infection using two commercial enzyme-linked immunoassays. Diagnostic Microbiology and Infectious Disease 2018, 90(1), 26–30. [CrossRef]
- Lustig, Y., Zelena, H., Venturi, G., Van Esbroeck, M., Rothe, C., Perret, C., Koren, R., Katz-Likvornik, S., Mendelson, E., and Schwartz, E. Sensitivity and Kinetics of an NS1-Based Zika Virus Enzyme-Linked Immunosorbent Assay in Zika Virus-Infected Travelers from Israel, the Czech Republic, Italy, Belgium, Germany, and Chile. Journal of Clinical Microbiology 2017, 55(6), 1894–1901. [CrossRef]
- Kadkhoda, K., Gretchen, A., Racano, A. Evaluation of a commercially available Zika virus IgM ELISA: specificity in focus. Diagnostic Microbiology and Infectious Disease 2017, 88(3), 233–235. [CrossRef]
- Fuchs, I., Bin, H., Schlezinger, S., Schwartz, E. NS1 antigen testing for the diagnosis of dengue in returned Israeli travelers. Journal of Medical Virology 2014, 86(12), 2005–2010. [CrossRef]
- Macdonald, J., Tonry, J., Hall, R.A., Williams, B., Palacios, G., Ashok, M.S., et al. NS1 protein secretion during the acute phase of West Nile virus infection. Journal of Virology 2005, 79(22), 13924–13933. [CrossRef]
- Chung, K.M., Diamond, M.S. Defining the levels of secreted non-structural protein NS1 after West Nile virus infection in cell culture and mice. Journal of Medical Virology 2008, 80(3), 547–556. [CrossRef]
Figure 1.
The phylogram constructed by the NJ method based on nucleotide sequences encoding OHFV NS1 and TBEV NS1 proteins, TN93+G+I substitution model with 1000 bootstrap iterations in MEGA X. Significant bootstrap values (>70%) are shown on the nodes. The sequence determined in this study is highlighted by a triangle.
Figure 1.
The phylogram constructed by the NJ method based on nucleotide sequences encoding OHFV NS1 and TBEV NS1 proteins, TN93+G+I substitution model with 1000 bootstrap iterations in MEGA X. Significant bootstrap values (>70%) are shown on the nodes. The sequence determined in this study is highlighted by a triangle.

Figure 2.
Condensed alignment based on the aa sequence of the NS1 protein OHFV strain 186-1964, obtained in this study, and NS1 proteins of 10 strains of OHFV (a) and protein TBEV NS1 strain 205 (b).
Figure 2.
Condensed alignment based on the aa sequence of the NS1 protein OHFV strain 186-1964, obtained in this study, and NS1 proteins of 10 strains of OHFV (a) and protein TBEV NS1 strain 205 (b).
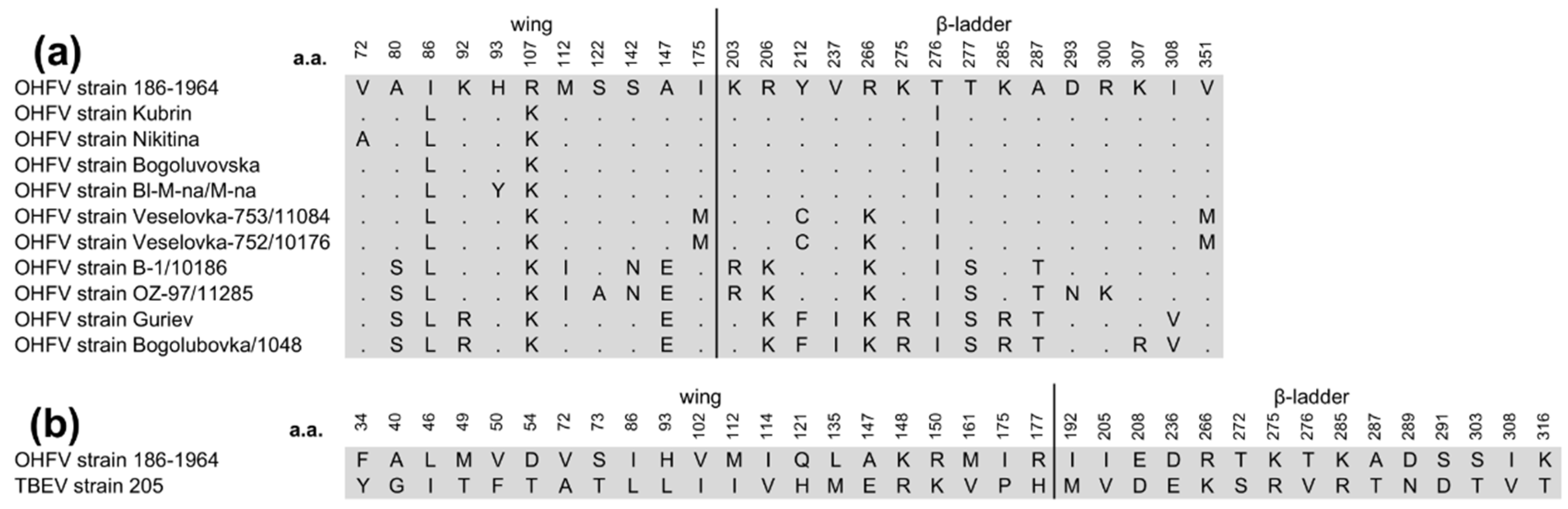
Figure 3.
Predicted 3D structures of the OHFV NS1 and TBEV NS1 protein dimers obtained using the AlphaFold2 algorithm. Domains 1, 2, and 3 are marked in green, yellow, and blue, respectively. Aa residues that differ between OHFV NS1 and TBEV NS1 proteins are highlighted in red. (a) - top view of the OHFV NS1 protein dimer; (b) top view of the TBEV NS1 protein dimer; (c) - bottom view of the OHFV NS1 protein dimer; (b) - bottom view of the TBEV NS1 protein dimer. The molecular coordinates of the putative OHFV NS1 protein were displayed using the UCSF Chimera, version 1.15.
Figure 3.
Predicted 3D structures of the OHFV NS1 and TBEV NS1 protein dimers obtained using the AlphaFold2 algorithm. Domains 1, 2, and 3 are marked in green, yellow, and blue, respectively. Aa residues that differ between OHFV NS1 and TBEV NS1 proteins are highlighted in red. (a) - top view of the OHFV NS1 protein dimer; (b) top view of the TBEV NS1 protein dimer; (c) - bottom view of the OHFV NS1 protein dimer; (b) - bottom view of the TBEV NS1 protein dimer. The molecular coordinates of the putative OHFV NS1 protein were displayed using the UCSF Chimera, version 1.15.
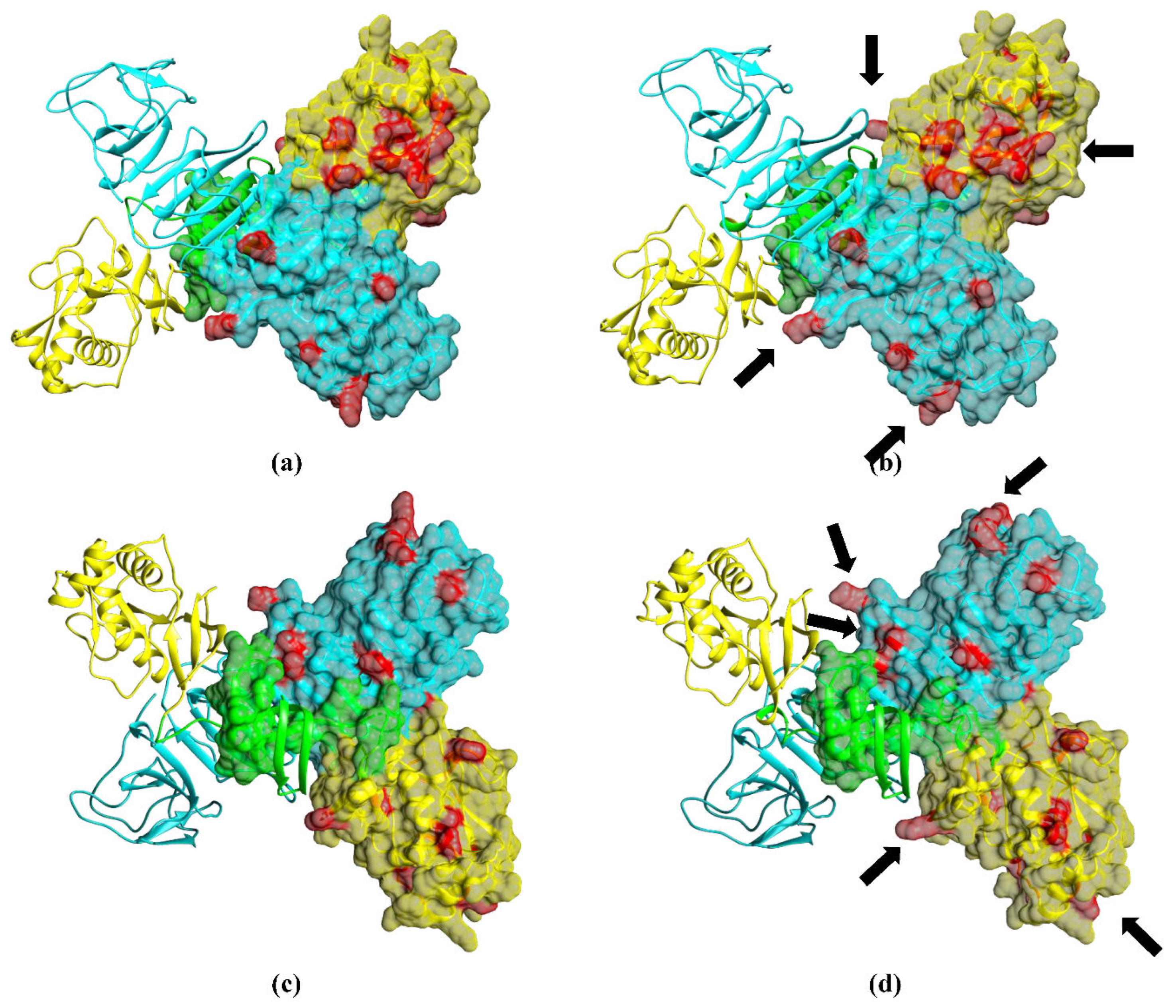
Figure 4.
SDS-PAGE of lysates of E. coli BL21 (DE3) cells transformed with the plasmid pET-32a(+)_NS1OHFV (1), cytoplasmic fraction (2) and inclusion body fraction (3). The BL21 (DE3) cells transformed with pET-32a(+) plasmid were used as control (1). Purified TBEV NS1 protein [24] was applied as a positive control (5). M - molecular mass marker of proteins.
Figure 4.
SDS-PAGE of lysates of E. coli BL21 (DE3) cells transformed with the plasmid pET-32a(+)_NS1OHFV (1), cytoplasmic fraction (2) and inclusion body fraction (3). The BL21 (DE3) cells transformed with pET-32a(+) plasmid were used as control (1). Purified TBEV NS1 protein [24] was applied as a positive control (5). M - molecular mass marker of proteins.
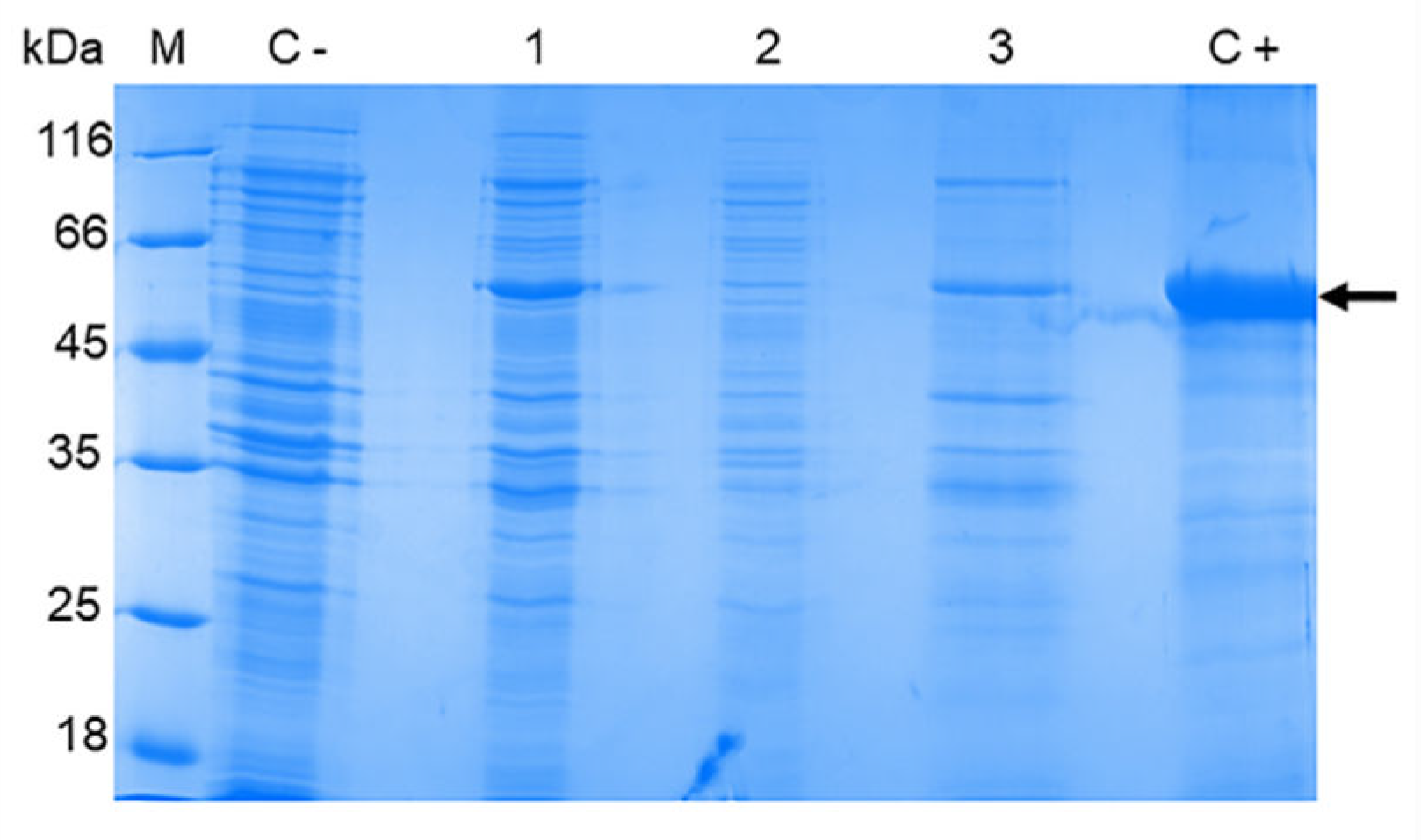
Figure 5.
SDS-PAGE of lysates of E. coli BL21 (DE3) / pET-32a(+)-NS1OHFV cells producing Trx-OHFV NS1 protein after optimization of induction conditions using different concentrations of IPTG; cultured for 1, 2, 4 and 16 hours at 18 °C and 30 °C. 1 - cells induced with 0.001 mM IPTG; 2 - cells induced with 0.01 mM IPTG; 3 - precipitate of cells induced with 0.1 mM IPTG; 4 - lysate of cells induced with 1 mM IPTG. M – PageRuler™ Unstained Protein Ladder (#26613, ThermoFischer scientific, Waltham, MA, USA).
Figure 5.
SDS-PAGE of lysates of E. coli BL21 (DE3) / pET-32a(+)-NS1OHFV cells producing Trx-OHFV NS1 protein after optimization of induction conditions using different concentrations of IPTG; cultured for 1, 2, 4 and 16 hours at 18 °C and 30 °C. 1 - cells induced with 0.001 mM IPTG; 2 - cells induced with 0.01 mM IPTG; 3 - precipitate of cells induced with 0.1 mM IPTG; 4 - lysate of cells induced with 1 mM IPTG. M – PageRuler™ Unstained Protein Ladder (#26613, ThermoFischer scientific, Waltham, MA, USA).
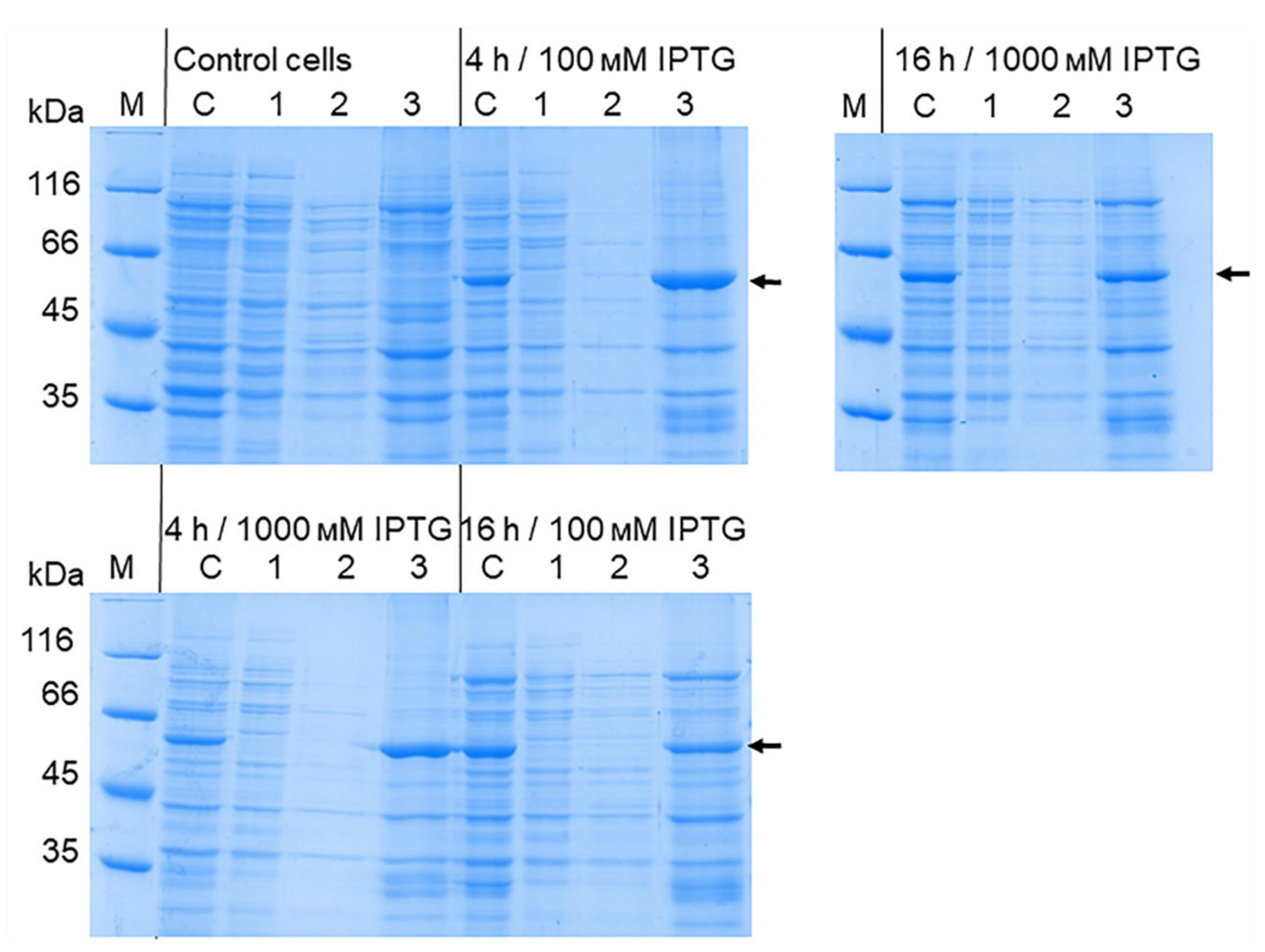
Figure 6.
SDS-PAGE of E. coli BL21 (DE3) / pET-32a(+)-NS1OHFV cell lysates cultured under different conditions and their different fractions. M - protein molecular weight marker, 1- initial cell lysates, 2 – soluble cytoplasm, 3 - fraction of inclusion bodies after refolding in 2 M urea, 4 - fraction of inclusion bodies after refolding in 6 M urea.
Figure 6.
SDS-PAGE of E. coli BL21 (DE3) / pET-32a(+)-NS1OHFV cell lysates cultured under different conditions and their different fractions. M - protein molecular weight marker, 1- initial cell lysates, 2 – soluble cytoplasm, 3 - fraction of inclusion bodies after refolding in 2 M urea, 4 - fraction of inclusion bodies after refolding in 6 M urea.
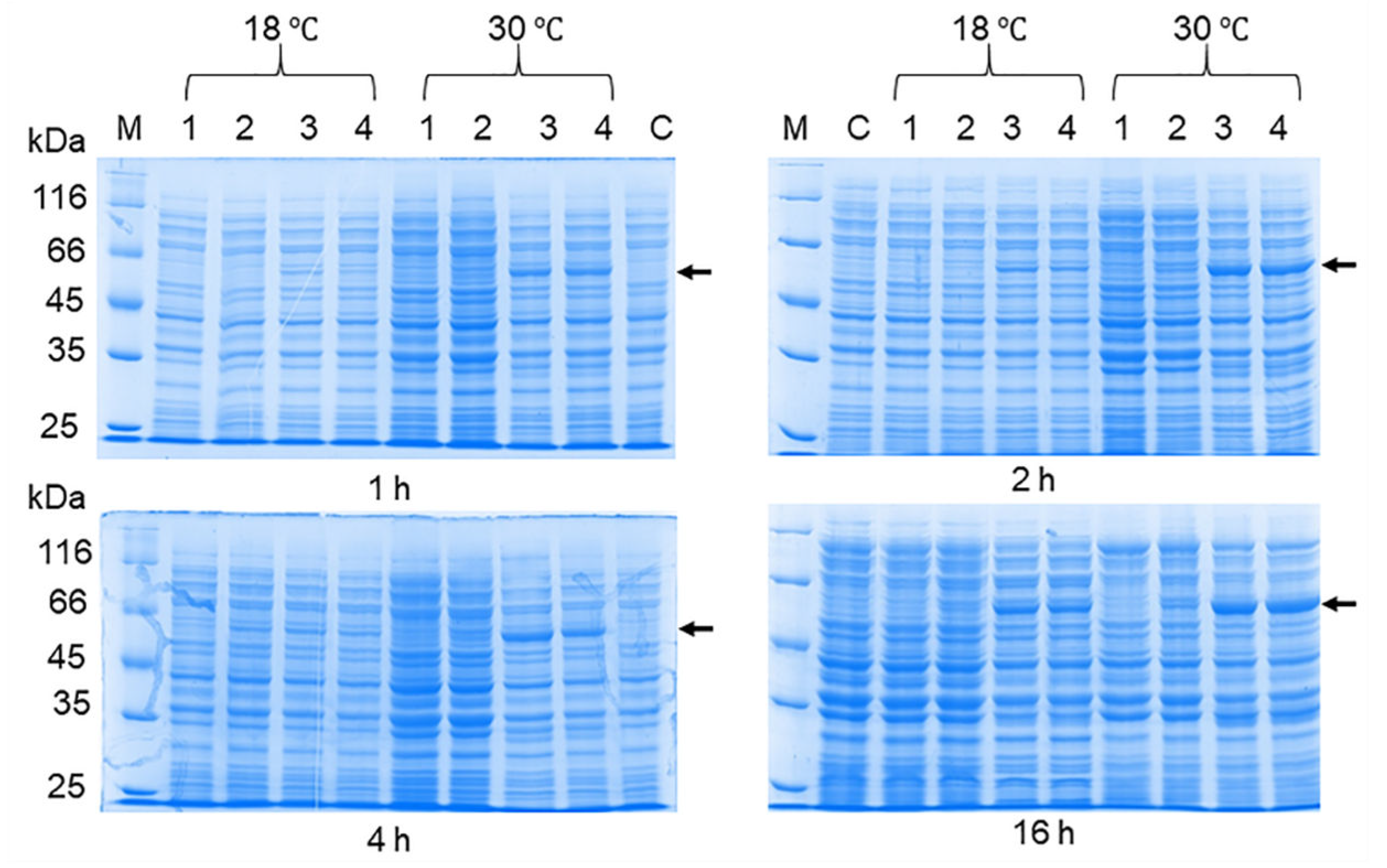
Figure 7.
Western blot analysis if the recombinant OHFV NS1 (lines 1-3) and TBEV NS1 (lines 4-6) proteins bound by the mAb.His1 (lines 1,4) against the C-terminal His-tag and immune ascites fluids from mice infected with OHFV P-15-2213 (lines 2,5) and OHFV Oz-31_Kd_10866 (lines 3,6).
Figure 7.
Western blot analysis if the recombinant OHFV NS1 (lines 1-3) and TBEV NS1 (lines 4-6) proteins bound by the mAb.His1 (lines 1,4) against the C-terminal His-tag and immune ascites fluids from mice infected with OHFV P-15-2213 (lines 2,5) and OHFV Oz-31_Kd_10866 (lines 3,6).

Figure 8.
ELISA serial dilution of immune ascetic fluids from mice infected with OHFV strains P-15-2213 and Oz-31_Kd_10866 to bind recombinant OHFV NS1 and OHFV NS1 proteins.
Figure 8.
ELISA serial dilution of immune ascetic fluids from mice infected with OHFV strains P-15-2213 and Oz-31_Kd_10866 to bind recombinant OHFV NS1 and OHFV NS1 proteins.
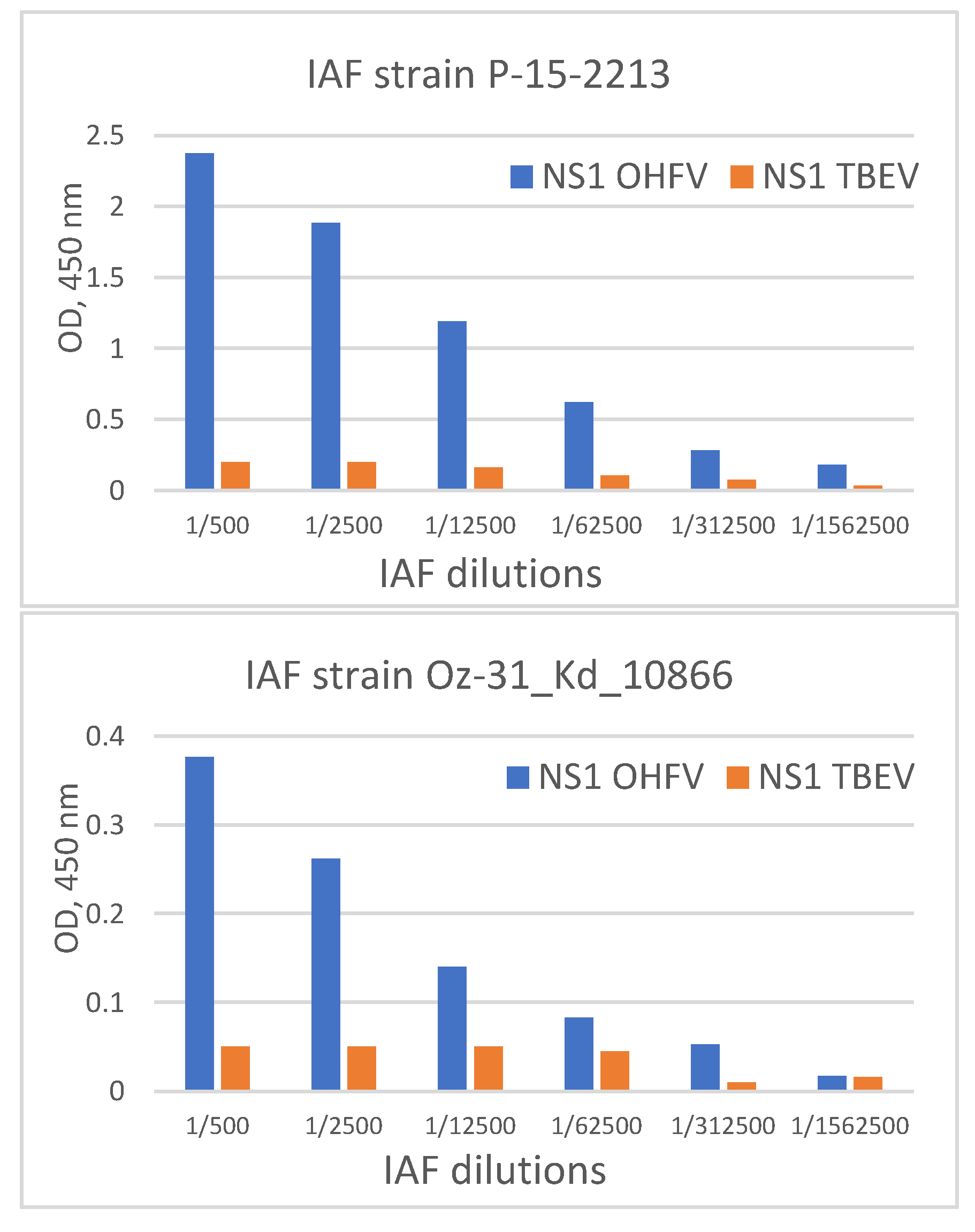
Figure 9.
ELISA of serial dilutions of FVN-NS1-44, FVN-NS1-290 and FVN-NS1-299 monoclonal antibodies specific to TBEV NS1 to bind of recombinant OHFV NS1 and OHFV NS1 proteins.
Figure 9.
ELISA of serial dilutions of FVN-NS1-44, FVN-NS1-290 and FVN-NS1-299 monoclonal antibodies specific to TBEV NS1 to bind of recombinant OHFV NS1 and OHFV NS1 proteins.
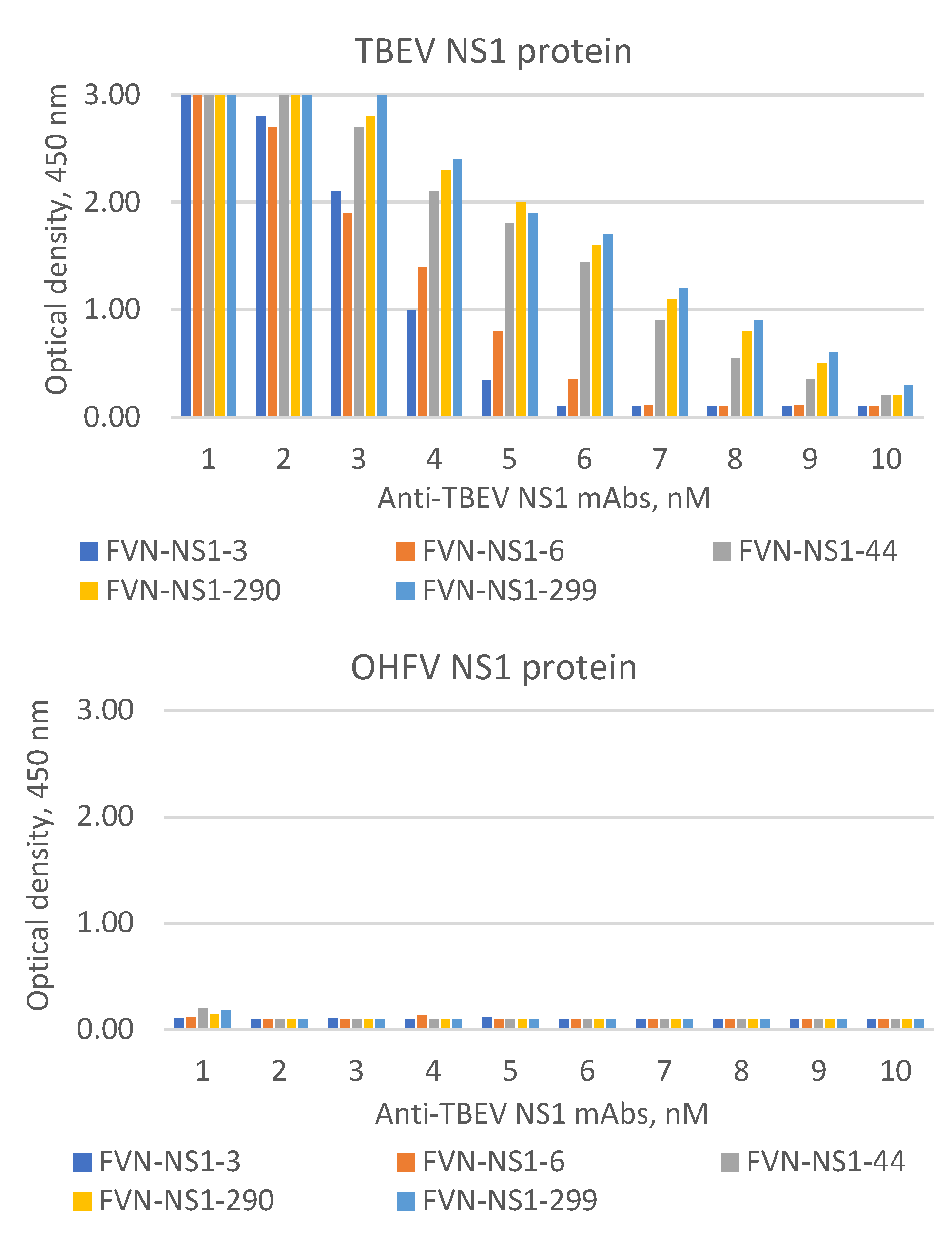
Figure 10.
ELISA of human sera for antibodies binding TBEV NS1 and OHFV NS1 proteins. * - ρ < 0.05, (one-way ANOVA). Symbols: blue column - sera from conditionally healthy donors without TBE and OHF (group 1), green column - sera from patients with clinically confirmed TBE (group 2). Red line - cutoff level. * - ρ < 0.05, (one-way ANOVA).
Figure 10.
ELISA of human sera for antibodies binding TBEV NS1 and OHFV NS1 proteins. * - ρ < 0.05, (one-way ANOVA). Symbols: blue column - sera from conditionally healthy donors without TBE and OHF (group 1), green column - sera from patients with clinically confirmed TBE (group 2). Red line - cutoff level. * - ρ < 0.05, (one-way ANOVA).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



