
Submitted:
09 May 2024
Posted:
10 May 2024
You are already at the latest version
Abstract
In recent years, long periods of drought and heat waves have become increasingly frequent, causing forest dieback phenomena that make stands more sensitive to biotic stressors. How trees may respond to extreme climatic events and which metabolites are involved under stress conditions is still not clear. In this study, we analysed by SPME how dieback (D) and non-dieback (ND) Hungarian oak trees from the San Paolo Albanese site respond to these climatic dynamics, in terms of volatile organic compounds (VOCs). For each group of trees, three wood samples were taken, each divided into four sub-samples with 5 growth rings, subjected to SPME and increase in basal area (BAI) analysis of the last 20 years. Dieback trees had a lower amount of leaves, and this condition may translate into less photosynthesis, less organic matter production and lower reserves of carbohydrates available for growth. Indeed, D trees showed lower radial increases and a lower content of aldehydes, terpenes, and fatty acids than ND trees, indicating a better health of ND trees compared to D trees. While D trees showed a reduction in terpenes, such as α-pinene, γ-eudesmol, and cyperene (with significant insecticidal activity), a reduction in aromatic aldehydes, such as furfural and 5-methylfurfural, and an increase in silanols (with antimicrobial function). Considering the different compounds’ contents between D and ND trees, our study could be useful for detecting bio-indicators to identify an early warning signal of dieback phenomena.
Keywords:
climate change
; drought
; forest dieback
; Hungarian oak
; growth ring
; SPME
; VOC
1. Introduction
Anthropogenic emissions of greenhouse gases are significantly changing the global climate (IPCC, 2022). The increased frequency of climatic anomalies, such as long periods of drought and heatwaves cause physiological stress in vegetation, making forests more susceptible to dieback phenomena [1,2,3].
The increase in forest mortality associated with global climate change is of great concern [4,5]. Forest decline has been attributed to interconnected abiotic and biotic factors. Triggers include extreme weather (drought, temperature extremes, high irradiance), often followed by attacks of defoliating insects, fungi and pathogens, but air pollution, such as nitrogen deposition, can also affect forest health [6]. The response of forests to such phenomena is very complex and can also be affected by local conditions, vegetation type, and general site characteristics (elevation, slope and exposure) [7]. The combination of drought and excess nitrogen reduces the concentration of allelochemicals in plant leaves [6], that are secondary metabolites produced by plants which can have a defensive function thanks to their repellent and/or toxic action against harmful defoliators and pathogens. These substances include phenolic compounds and their derivatives (e.g., tannins against defoliating or herbivorous insects), whose reduction in the leaves can make trees more susceptible to insect attack. Thus, biotic and abiotic factors are interconnected: defoliation, favoured by drought and reduced allelochemicals, causes a decrease in leaf area and a consequent reduction in photosynthetic activity and in the supply of carbohydrates to the plants [8,9]. In order to understand the mechanisms that cause carbon depletion in underground structures, some authors studied the dynamics of metabolites with progressive drought in forest species: tree carbon fluxes revealed that in roots, fructose, glucose, sucrose, non-structural carbohydrates (NSC), starch and the osmoprotectant proline increased during drought, while in the phloem there was only an accumulation of sucrose and in leaves no accumulation [10,11,12]. Based on this, it seems that trees initially damaged by drought give priority to root assimilates, probably to promote root function after the drought event [10]. The reduction of carbon in the belowground sink is caused by the persistence and frequency of drought conditions over time, like a prolonged stress that does not allow the storage of additional reserve substances, thus causing the phenomenon of carbon starvation [11,12,13].
The content of carbon, aquaporins (which facilitate water flow in the roots), glucose, sucrose and other reserve substances could also be associated with a further response of vegetation to extreme weather, namely xylem filling after embolism. Some forest species die because of embolism, while others tolerate high levels of embolism and subsequently recover hydraulic function during rehydration [14]. Current knowledge about this physiological response suggests that sugars and ions released into the ducts cause osmotic gradients by channelling the flow of water through parenchyma, phloem and xylem. However, frequent drought events with embolism phenomena can cause chronic stress and thus an inability to reverse embolism [14]. These processes are still not very clear and require further studies to better understand how plants can respond to harsh conditions and which substances/metabolites are involved in forest resilience or recovery after periods of stress.
Therefore, in order to have more accurate information on the dynamics that are affecting a stand of Pollino National Park (Basilicata Region, Southern Italy), which are already hit by dieback phenomena, in this study we propose the evaluation of the effects of climate change on volatile organic compounds (VOCs) and plant secondary compounds (PSCs) in Mediterranean oak forests affected by dieback phenomena.
We hypothesize that declining trees because of extreme drought conditions occurred during the last two decades [15,16] stress showed considerable variations in VOCs and PSCs that had made the trees more susceptible to the action of pests. Here, we assessed the presence or absence of metabolites (e.g., phenolic compounds and their derivatives) useful for warding off biotic attacks or the presence or absence of substances useful for enabling the vegetation to cope with stress conditions. These processes are still poorly understood, so further studies are needed to fully understand which metabolites are involved in the response to dieback phenomena.
2. Materials and Methods
2.1. Study Site
The study was carried out in a Q. frainetto Ten. high forest that manifested dieback phenomena. The stand was an even-aged high forest with an average breast-height diameter of 40 cm and a mean age of about 145 years [17]. This stand is located in the Pollino National Park in the municipality of San Paolo Albanese within the Basilicata region in southern Italy (40° 1' 16.90" N; 16° 20' 27.57" E) with an elevation of 1030 m a.s.l. and an average slope of 25-30%. According to Pavari phyto-climatic classification, the forest belongs to the Castanetum belt, typical of mesophilic oak woods. The site has a sandy-clayy soil and a Mediterranean climate, with hot, dry summers and wet, mild winters; the average annual temperature is 16.4 °C and annual rainfall is 742 mm. Q. frainetto stand has shown symptoms of dieback since the early 2000s (shoot desiccation, significant growth decline, high mortality rate) with 80% of the trees showing crown transparency levels >50%[17,18].
2.2. Field Sampling and Dendrochronological Data
At the analysed site, two groups of trees were identified through visual assessment (Figure S1): decaying plants (hereafter D trees) with crown transparency >50% and non-decaying plants (hereafter ND trees) with crown transparency <50% [17,19]. For each group (D and ND), three plants were sampled by extracting wood cores at breast height (1.30 m) using a 1 cm Pressler drill, and the diameter of each plant was measured using a tree caliper. The samples taken were coded and stored in a refrigerator at 4 °C, in order to avoid alterations that could affect subsequent analyses. Each sample was set up on special supports and smoothed so that the annual growth rings were visible (Figure S2). The analysis involved the study of the last 20 years of growth by using a dendrocronograph (LINTAB6, Rinntech). Each growth ring was measured with an accuracy of 1/1000 mm using the TSAPwin software (Rinntech). By the measured tree ring widths (TRW), growth histories from 2002 to 2021 were obtained for each tree group (D and ND). These chronologies were used to calculate the basal area increment (BAI) by applying the following formula:
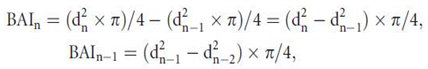 where d2n is the diameter squared of the year n, while d2n -1 is the diameter squared of the previous year n-1 and so on d2n-1 - d2n-2....
where d2n is the diameter squared of the year n, while d2n -1 is the diameter squared of the previous year n-1 and so on d2n-1 - d2n-2....
The calculation of BAI was performed without detrending the radial increments, as the analyzed trees were adults, with an age of approximately 145 years [17] therefore with a growth quite parallel to the x-axis not influenced by age trends [20]. Using the raw data, we stayed as close as possible to the source data and thus made fewer assumptions. Calculating BAI relative to TRW allowed us to reduce the geometric effect of radial growth increments of cores taken at 1.30 m [21]. The average BAI was calculated for each ND and D tree class and the average BAI increment for each analysed period was obtained.
2.3. Sample Preparation
The last 20 rings (20 years, from 2021 to 2002) of each sample were divided into 4 sections of 5 growth rings per each. For each sample, 4 Eppendorf tubes were used, each containing 5 growth rings, i.e. 2002/2006 - 2007/2011 - 2012/2016 - 2017/2021. Thus, a total of 24 sections were obtained from the 6 samples analysed (3 D and 3 ND), and their wood material was placed in sterile Eppendorf tubes. The samples were then dried at 50 °C overnight, each sample was added 5 ml of distilled water and was analysed by SPME (solid-phase micro extraction).
2.4. Statistical Analyses
With the aid of the SPSS software version 11.0 (SPSS, Chicago, Illinois, USA), an analysis of variance and comparison of treatment means (one-way ANOVA) was conducted in order to investigate the impact of trees (D and ND) on the peak area of various volatile chemicals detected in the cores of oak trees. All series of data set (D and ND) were statistically analysed to evaluate the significance of the results.
2.5. Solid Phase Micro Extraction Analyses
Solid Phase Micro Extraction (SPME) was used to determine the VOCs chemical composition of the 24 samples. An aliquot of 200 mg of sample was placed in vials and incubated at 50 °C for 15 min to promote volatile compounds in the headspace. Extractions were realized by immersing the fiber in the headspace for 30 min and then withdrawn into the needle and transferred to a GC/MS system [22]. A 50/30-µm DVB/CAR/PDMS module (57328-U, Supelco, Milan, Italy) was employed to determine volatile organic compounds (VOCs).
Analyses were accomplished with an HP 6890 Plus gas chromatograph (Agilent) equipped with a PhenomenexZebron ZB-5 MS capillary column (30-m × 0.25-mm i.d. × 0.25 µm FT) (Agilent, Milan, Italy). An HP 5973 mass selective detector (Agilent) was utilized with helium at 0.8 mL min−1 as the carrier gas. A splitless injector was maintained at 250 °C and the detector at 230 °C. The oven was held at 40 °C for 2 min, then gradually warmed, 8 °C min−1, up to 250 °C and held for 10 min. Tentatively identification of components was based on mass spectra and NIST 11 library comparison. Single VOC peak was considered as identified when its experimental spectrum matched with the one in the library with a score over 90% and if the retention time was in agreement with the reported retention index [22]. The analyses were performed in triplicate.
3. Results and Discussion
3.1. Dendrochronological Analysis
The differences between the growth rates of ND and D trees were evident (Figure 1 and S2). The BAI of ND plants for each 5-year period analysed showed increases of about 7-8 cm2. Instead, in D plants the increases in BAI, over the 20 years analysed, was always below 5 cm2, except for the period 2007/2011, when the value was almost 5 cm2. These observations further confirm the difference in the health status of the two groups observed by visual inspection done by Colangelo et al. (2017a) (D trees with crown transparency >50% and ND trees with crown transparency <50%) [17]. Certainly, the increases in BAI were very low for both groups over the analized period but, the lower growth rate in D trees compared to ND trees is an unequivocal sign of decay phenomena [15]. Indeed, periods of drought can favour embolism, leading to physiological stress and consequent canopy self-thinning. A very low BAI rate could be attributable to differences in the amount of available crown/leaves between D and ND trees. In fact, dieback trees have a lower amount of leaves and this condition may translate into less photosynthesis, less organic matter production and lower reserves of carbohydrates available for their growth [23]. In the last decades there have been many years characterized by drought and heat waves in the Mediterranean environment [7,17,24], such as 2001, 2008, 2012, 2013, 2017 which may have eroded the carbohydrate reserves of plants causing a greater dependence of decaying plants to recent climatic conditions [25].
3.2. SPME Analysis
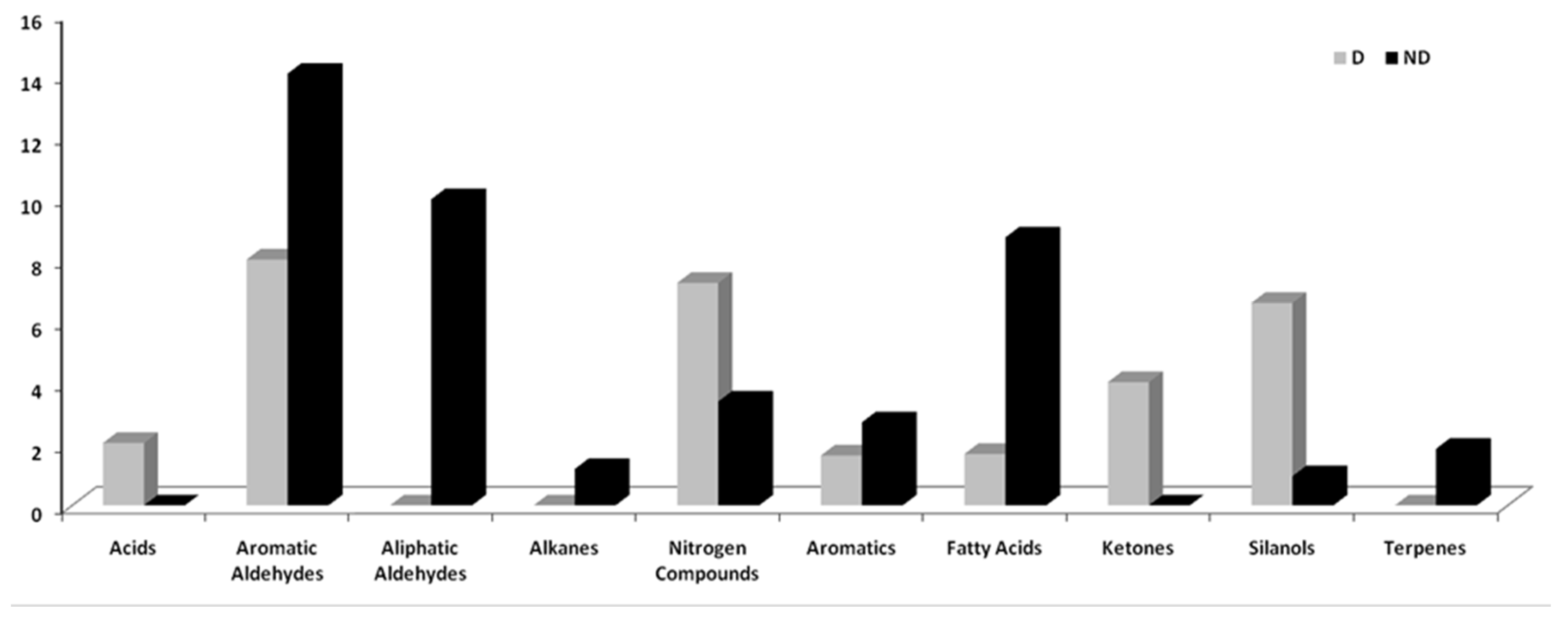
Plant secondary compounds (PSCs) have a high chemical and structural diversity and include non-volatile or volatile organic compounds (VOCs). These compounds have specific physiological and ecological functions in plants’ adaptation [26,27]. PSCs can help trees to adapt to climate change and to the attack of invasive insects and pathogens. In higher plants, terpenoids (30,000), alkaloids (21,000), and phenolic compounds (8,000) are the most diverse PSC groups [28,29,30]. Table 1 shows the total percentage areas of VOCs, both for decaying (1D, 2D, 3D) and non-decaying (1ND, 2ND, 3ND) wood material in the four five-year periods 2002/2006 - 2007/2011 - 2012/2016 – 2017/2021. The compounds have been separated into different classes of organic compounds, to better evaluate the variations: acids, aldehydes, alkanes, nitrogen compounds, aromatic compounds, fatty acids (FA), ketones, silanols, and terpenes.
As shown in Figure 2, over the years, a notable reduction of aliphatic and aromatic aldehydes, alkanes, aromatic compounds, terpenes, and fatty acids was observed in D plants compared to ND plants; while the same plants showed an increase in acids, nitrogen compounds, ketones, and silanols.
Aldehydes can be involved in signaling pathways and are often associated with stress responses in plants. The higher content of aldehydes in ND trees, especially aromatic ones like furfural and 5-methylfurfural, suggests a more effective stress response mechanism, aiding these trees in coping with adverse conditions [23].
Alkanes are typically part of the waxy cuticle, serving as a protective barrier against environmental stressors. The absence or presence of alkanes might reflect changes in the composition of the protective surface, potentially impacting the tree's ability to resist drought and pathogens.
Aromatic compounds can serve as defense mechanisms against herbivores and pathogens. The reduction in aromatic aldehydes in D trees may indicate a weakened defense mechanism, making them more susceptible to stressors like pests and diseases [31].
Terpenes are a diverse class of compounds with various functions, including defense against herbivores and pathogens. The reduction in terpenes in D trees indicates a reduced capacity to defend against stressors, such as pests and diseases [32].
Fatty acids (FA) are fundamental components of lipids and are involved in energy storage and membrane structure. The higher content of fatty acids in ND trees suggests they have more energy reserves for growth and stress response, which is essential for maintaining good physiological status; they are essential for cell membranes’ stability and fluidity, for plant metabolism, and indicate a better health status of the plants. Among the multiple roles of FA, in fact, they have structural functions as constituents of phospholipids which are the “building blocks” of cell membranes. As part of neutral lipids, FA serves as storage materials in cells and FA derivatives are involved in cell signalling [33]. At the moment, their role in the regulation of the cellular membrane fluidity and their participation in complex processes, such as proliferation, differentiation, secretion, migration, invasion and phagocytosis, highlights the importance of these molecules for the maintenance and adaptation of plants in the forests [33].
Acids can play multiple roles in plant metabolism. Organic acids are involved in various biochemical pathways, such as the citric acid cycle (Krebs cycle) and as components of amino acids. A higher presence of acids in D trees could indicate a response to metabolic stress, to compensate for the potential impact on energy production and nutrient availability [34].
Nitrogen compounds are essential for plant growth and are typically found in amino acids, proteins, and nucleic acids. The presence or absence of certain nitrogen compounds could indicate changes in nitrogen availability and utilization, which are critical for plant development and response to stress [35].
Ketones can be involved in energy metabolism. The presence of ketones may indicate changes in energy metabolism due to stress, but the specific role in this context would require further investigation.
Silanols are organic compounds containing silicon and can have antimicrobial properties. An increase in silanols in D trees may be part of their defense mechanism against pathogens due to the absence of healthier alternatives [36].
Previous studies explain how embolism of xylem vessels, caused by drought, with a discontinuity of hydraulic flows, induces branch desiccation, leaves yellowing and generalised necrosis, causing a decrease in leaf area and a consequent reduction in photosynthetic activity [14]. Plants, to limit the loss of water through transpiration, close the stomata during drought, resulting in a poor supply of CO2. The main consequence of carbon starvation and the reduction of photosynthesis is a reduction in carbohydrate synthesis and supply to the plants [8,37]. This would explain why in D plants, a reduction of compounds deriving from carbohydrates, such as aldehydes, terpenoids, terpenes, and phenolic compounds occurred. In fact, terpenoids and some terpenes originate directly from the glycolysis of glucose through the mevalonate pathway or the methyleritritol phosphate (MEP) pathway [38], while phenolic compounds originate from the shikimic acid pathway, which is related to the metabolism of carbohydrates and aromatic amino acids [29].
Climate stress could have determined a reduction of compounds such as oxime methoxy-phenyl and benzaldehyde 4-hydroxy-3,5-dimethoxy (or syringaldehyde) with important anti-microbial and antifungal activities [39,40], and of terpenes, such as α-pinene, γ-eudesmol, and cyperene, which have significant insecticidal activity [41,42,43] (Table 1 and Table 2). Such decreases in compounds thus explain part of the deterioration of D plants compared to ND plants, because the reduction of these substances in the leaves could have made trees more sensitive to insect attacks and parasites. To compensate, D plants defended themselves by increasing compounds, such as silanediol dimethyl, an important antimicrobial, and diethyltoluamide, which has a marked insecticidal activity [44,45].
According to Martinez et al. (1996) [46], the decrease of these compounds is a consequence of uncontrolled decomposition of several compounds (for example, lignin), which causes the formation of much less reactive compounds. At the same time plants, especially those progressing towards dieback, under stress conditions, may release mixtures of volatile organic compounds (like silanols) that indirectly protect the plant by attracting natural enemies of the herbivore [47]. Recent studies suggest that these VOCs can also prime nearby crop plants for enhanced induction of defence upon future insect attack [48]. Thus, these differences could be bio-indicators of early warning signals of an onset of the dieback state [49].

Table 1.
Area (% ± 0.03) of compounds found in SPME analysis of dieback (D) and non-dieback (ND) Q. frainetto Ten. Trees every 5 years, for the past 20 years (I:2002-2006; II:2007-2011; III:2012-2016; IV:2017-2021).
Table 1.
Area (% ± 0.03) of compounds found in SPME analysis of dieback (D) and non-dieback (ND) Q. frainetto Ten. Trees every 5 years, for the past 20 years (I:2002-2006; II:2007-2011; III:2012-2016; IV:2017-2021).
| Class of Compounds | Compound | 1D | 2D | 3D | |||||||||
| Acids | Acetic acid | I | II | III | IV | I | II | III | IV | I | II | III | IV |
| 2.75 | 1.35 | 1.37 | 1.27 | 1.37 | 1.83 | 4.15 | 3.42 | 2.83 | 1.35 | 1.38 | 1.28 | ||
| Aromatic Aldehydes |
Sinapaldehyde | 1.27 | 4.17 | 1.42 | |||||||||
| Coniferaldehyde | |||||||||||||
| Syringaldehyde | 4.15 | 3.15 | 2.05 | 1.90 | 3.95 | 4.00 | 2.60 | 2.43 | 1.90 | 1.82 | 1.72 | 1.63 | |
| Vanillin | 6.88 | 0.92 | 10.95 | 2.95 | 18.90 | 6.53 | 2.68 | 8.08 | |||||
| Aliphatic Aldehydes |
3-Methylbutanal | ||||||||||||
| Decanal | |||||||||||||
| Furfural | |||||||||||||
| Heptanal | |||||||||||||
| Nonanal | |||||||||||||
| Octanal | |||||||||||||
| Alkanes | Eicosane | ||||||||||||
| Nitrogen Compounds |
N-p-Bromophenylselenoacetamide | 1.05 | |||||||||||
| Diethyltoluamide | 6.30 | 4.75 | 4.48 | 20.70 | 20.10 | 6.52 | 11.58 | 11.38 | |||||
| Dimethyl palmitamine | |||||||||||||
| Aromatics | 1,1,3-Trimethyl-3-phenyl-2H-indene | ||||||||||||
| 2,6-Di-tert-butylphenol | |||||||||||||
| Methyl N-hydroxybenzimidate | 1.05 | 0.83 | 2.45 | 1.27 | 1.35 | 2.88 | 4.70 | ||||||
| 2,6-Dimethoxyphenol | 2.05 | 3.25 | |||||||||||
| Xylene | |||||||||||||
| Fatty acids | Pentanoic acid | 0.60 | |||||||||||
| Nonanoic acid | 0.37 | ||||||||||||
| Tetradecanoic acid | |||||||||||||
| Hexadecanoic acid | 2.62 | 2.75 | 2.15 | 2.45 | |||||||||
| Hexanedioic acid | 2.22 | 2.03 | 1.98 | ||||||||||
| Octadecanoic acid | 1.55 | 1.32 | |||||||||||
| Ketones | 4-Oxo-β-dihydroionone | 12.37 | 3.75 | ||||||||||
| 3-Oxo-7,8-dihydro-α-ionone | 1.52 | 1.70 | 5.75 | 6.35 | 12.22 | 2.10 | 2.35 | ||||||
| Silanols | dimethylsilanediol | 8.22 | 9.33 | 7.43 | 7.05 | 3.43 | 3.72 | 7.03 | 5.48 | 5.50 | 7.02 | 7.63 | 7.28 |
| Terpenes | α-Bergamotene | ||||||||||||
| Cyperene | |||||||||||||
| γ-Eudesmol | |||||||||||||
| β-Panasinsene | |||||||||||||
| Pinocarvone | |||||||||||||
| α-Pinene | |||||||||||||
| Class of Compounds | Compound | 1ND | 2ND | 3ND | |||||||||
| I | II | III | IV | I | II | III | IV | I | II | III | IV | ||
| Acids | Acetic acid | ||||||||||||
| Aromatic Aldehydes |
Sinapaldehyde | ||||||||||||
| Coniferaldehyde | 17.13 | 20.38 | 12.40 | 8.35 | |||||||||
| Syringaldehyde | 14.45 | 13.20 | 12.20 | 6.40 | |||||||||
| Vanillin | 0.43 | 11.73 | 18.20 | 16.05 | 13.20 | 2.32 | 2.13 | ||||||
| Aliphatic Aldehydes |
3-Methylbutanal | 0.93 | |||||||||||
| Decanal | 0.58 | 16.45 | 3.27 | 4.77 | 4.27 | 2.13 | 1.77 | 7.92 | 5.57 | ||||
| Furfural | 0.52 | 2.07 | 0.70 | ||||||||||
| Heptanal | 1.02 | 2.05 | 2.65 | 2.45 | 4.40 | 0.97 | 4.25 | 2.43 | 1.88 | ||||
| Nonanal | 1.25 | 7.90 | 9.63 | 4.82 | 7.08 | ||||||||
| Octanal | 0.62 | 8.10 | 3.17 | 3.32 | |||||||||
| Alkanes | Eicosane | 2.67 | 7.20 | 4.28 | |||||||||
| Nitrogen Compounds |
N-p-Bromophenylselenoacetamide | ||||||||||||
| Diethyltoluamide | 5.87 | 8.47 | |||||||||||
| Dimethyl palmitamine | 5.93 | ||||||||||||
| Aromatics | 1,1,3-Trimethyl-3-phenyl-2H-indene | 0.52 | 0.43 | 0.48 | |||||||||
| 2,6-Di-tert-butylphenol | 2.27 | 1.73 | |||||||||||
| Methyl N-hydroxybenzimidate | 1.03 | 1.60 | 0.45 | 2.72 | 1.30 | 0.72 | |||||||
| 2,6-Dimethoxyphenol | 5.93 | 4.70 | 1.35 | ||||||||||
| Xylene | 0.53 | 4.72 | 1.93 | ||||||||||
| Fatty acids | Pentanoic acid | 0.30 | 4.05 | 2.87 | 2.87 | 1.93 | 3.55 | 2.40 | |||||
| Nonanoic acid | 1.88 | 4.90 | |||||||||||
| Tetradecanoic acid | 1.37 | 1.10 | 0.30 | 1.23 | |||||||||
| Hexadecanoic acid | 20.75 | 0.98 | 1.92 | 1.55 | 2.08 | 2.12 | 1.63 | ||||||
| Hexanedioic acid | 1.87 | 2.10 | 1.77 | 5.07 | 3.88 | ||||||||
| Octadecanoic acid | 17.57 | 1.40 | 1.60 | 9.63 | |||||||||
| Ketones | 4-Oxo-β-dihydroionone | ||||||||||||
| 3-Oxo-7,8-dihydro-α-ionone | |||||||||||||
| Silanols | dimethylsilanediol | 0.17 | 2.30 | 0.28 | 2.70 | 3.43 | 2.52 | ||||||
| Terpenes | α-Bergamotene | 0.60 | 1.08 | 0.98 | |||||||||
| Cyperene | 1,18 | 1.37 | 1.13 | ||||||||||
| γ-Eudesmol | 1.27 | 1.70 | |||||||||||
| β-Panasinsene | 1.80 | 0.43 | 1.62 | ||||||||||
| Pinocarvone | 1.02 | 1.40 | |||||||||||
| α-Pinene | 3.62 | 2.87 | |||||||||||
Table 2.
Average peak area of volatile compounds from dieback (D) and non-dieback (ND) oak wood determined by SPME.
Table 2.
Average peak area of volatile compounds from dieback (D) and non-dieback (ND) oak wood determined by SPME.
| Wood Type | Acids | Aromatic Aldehydes |
Aliphatic Aldehydes |
Alkanes | Nitrogen Compounds |
Aromatics | Fatty Acids | Ketones | Silanols | Terpenes | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| average | D | 2.0 | 4.5 | n.d. | n.d. | 9.7 | 2.2 | 1.8 | 5.3 | 6.6 | n.d. | |
| st. dev | 1.0 | 4.1 | 6.5 | 1.2 | 0.7 | 4.1 | 1.7 | |||||
| average | ND | n.d. | 10.6 | 3.8 | 4.7 | 6.8 | 1.9 | 3.6 | n.d. | 1.9 | 1.5 | |
| st. dev | 5.9 | 3.4 | 1.9 | 1.2 | 1.6 | 4.6 | 1.2 | 0.8 | ||||
| t Test (prob) | 0.008 | n.d. | n.d. | 0.502 | 0.649 | 0.220 | n.d. | 0.000 | n.d. | |||
| significance | ** | n.s. | n.s. | n.s. | *** | |||||||
Data given as area units of chromatograph peaks, mean of three analyses, nd, Not detected, Significance (* p < 0.05, ** p < 0.01, *** p < 0.001).
4. Conclusions
This study offers valuable insights into how Q. frainetto trees respond to the increasing climatic challenges of drought and heat waves. The lower growth rate in D trees compared to ND trees is an unequivocal sign of decay phenomena. In fact, D trees have a lower amount of leaves and this condition may translate into less photosynthesis, less organic matter production and lower reserves of carbohydrates produced by the trees available for growth. The observed differences in volatile organic compounds (VOCs) and plant secondary compounds (PSCs) between dieback (D) and non-dieback (ND) trees suggest that these compounds could serve as indicators of tree health and susceptibility to dieback phenomena. ND trees showed a higher content of aldehydes, terpenes and fatty acids, indicators of a better state of health than D trees. On the other hand, D trees contained less terpenes, like α-pinene, γ-eudesmol, and cyperene (having an insecticidal activity), and had low levels of aromatic aldehydes (i.e., furfural and 5-methylfurfural) and high silanols (these latter with an antimicrobial function). From a practical point of view, these findings have the potential to inform future forest management strategies, enabling the early detection of declining tree health. However, the study also underscores the complexity of tree responses to environmental stress, leaving room for future investigations to delve deeper into the precise roles of these compounds, explore underlying genetic and physiological mechanisms, and expand the scope to encompass multiple tree species and diverse regions. As climate change continues to pose threats to forests, this research paves the way for further studies and practical tools aimed at safeguarding our valuable tree populations and ecosystems.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Location of the San Poalo Albanese site in the Basilicata Region; a) an example of ND tree with crown transparency <50%; b) example of D tree with crown transparency >50%; Figure S2: Example of cross-sections of Q. frainetto wood. The arrows indicate the last 3 growth rings of (a) tree ND and of (b) tree D. The difference in annular growth between the two samples is evident.
Author Contributions
Conceptualization, writing, and original draft preparation by M.M., L.T., A.S., F.R. and S.S.P.I.; methodology, L.T., M.M. and S.S.P.I.; literature review, M.M., L.T., and M.D.; supervision, L.T. & F.R.; funding acquisition, F.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M., ... Cobb, N. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest ecology and management 2010, 259, 660–684. [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer; A., Barbati, A.; Garcia-Gonzalo, J.; ... Marchetti, M. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. Forest ecology and management 2010, 259, 698–709. [CrossRef]
- Gentilesca, T.; Camarero, J.J.; Colangelo, M.; Nolã, A.; Ripullone, F.; Nole, A. Drought-induced oak decline in the western Mediterranean region: an overview on current evidences, mechanisms and management options to improve forest resilience. IForest 2017, 10, 796–806. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; ... Woods, A.J. Climate change and forest diseases. Plant pathology 2011, 60, 133–149.
- Thomas, F.M.; Blank, R.; Hartmann, G. Abiotic and biotic factors and their interactions as causes of oak decline in Central Europe. Forest Pathology 2002, 32, 277–307. [Google Scholar] [CrossRef]
- Rita, A.; Camarero, J. J.; Nolè, A.; Borghetti, M.; Brunetti, M.; Pergola, N.; ... Ripullone, F. Global change biology 2020, 26, 851–863.
- Quentin, A.G.; Beadle, C.L.; O’grady, A.P.; Pinkard, E.A. Effects of partial defoliation on closed canopy Eucalyptus globulus Labilladière: growth, biomass allocation and carbohydrates. Forest Ecology and Management 2011, 261, 695–702. [Google Scholar] [CrossRef]
- Iqbal, N.; Masood, A.; Khan, N.A. Analyzing the significance of defoliation in growth, photosynthetic compensation and source-sink relations. Photosynthetica 2012, 50, 161–170. [Google Scholar] [CrossRef]
- Hagedorn, F.; Joseph, J.; Peter, M.; Luster, J.; Pritsch, K.; Geppert, U.; ... Arend, M. Recovery of trees from drought depends on belowground sink control. Nature plants 2016, 2, 1–5.
- Schmid, S.; Palacio, S.; Hoch, G. Growth reduction after defoliation is independent of CO2 supply in deciduous and evergreen young oaks. New Phytologist 2017, 214, 1479–1490. [Google Scholar] [CrossRef] [PubMed]
- Furze, M.E.; Wainwright, D.K.; Huggett, B.A.; Knipfer, T.; McElrone, A.J.; Brodersen, C.R. Ecologically driven selection of nonstructural carbohydrate storage in oak trees. New Phytologist 2021, 232, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Zeppel, M.J.; Anderegg, W.R.; Bloemen, J.; De Kauwe, M.G.; Hudson, P.; ... Nardini, A. Xylem embolism refilling and resilience against drought-induced mortality in woody plants: processes and trade-offs. Ecological research 2018, 33, 839–855. [CrossRef]
- Colangelo, M.; Camarero, J.J.; Borghetti, M.; Gazol, A.; Gentilesca, T.; Ripullone, F. Size matters a lot: drought-affected Italian oaks are smaller and show lower growth prior to tree death. Frontiers in Plant Science 2017, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Ripullone, F.; Camarero, J.J.; Colangelo, M.; Voltas, J. Variation in the access to deep soil water pools explains tree-to-tree differences in drought-triggered dieback of Mediterranean oaks. Tree Physiology 2020, 40, 591–604. [Google Scholar] [CrossRef]
- Colangelo, M.; Camarero, J.J.; Battipaglia, G.; Borghetti, M.; De Micco, V.; Gentilesca, T.; Ripullone, F. A multi-proxy assessment of dieback causes in a Mediterranean oak species. Tree physiology 2017, 37, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Cailleret, M.; Dakos, V.; Jansen, S.; Robert, E.M.; Aakala, T.; Amoroso, M.M.; ... Martinez-Vilalta, J. Early-warning signals of individual tree mortality based on annual radial growth. Frontiers in plant science 2019, 9, 1964. [CrossRef]
- Dobbertin, M. Tree growth as indicator of tree vitality and of tree reaction to environmental stress: a review. European Journal of Forest Research 2005, 124, 319–333. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: evidence of stress release by inter-specific facilitation. Plant biology 2013, 15, 483–495. [Google Scholar] [CrossRef]
- DeSoto, L.; Cailleret, M.; Sterck, F.; Jansen, S.; Kramer, K.; Robert, E.M.; ... Martínez-Vilalta, J. Low growth resilience to drought is related to future mortality risk in trees. Nature communications 2020, 11, 545. [CrossRef] [PubMed]
- Mecca, M.; Todaro, L.; Lo Giudice, V.; Lovaglio, T.; D’Auria, M. GC-MS and SPME techniques highlighted contrasting chemical behaviour in the water extractives of modified Castanea sativa mill. and Fagus sylvatica L. wood. Forests 2021, 12, 986. [Google Scholar] [CrossRef]
- Levanič, T.; Čater, M.; McDowell, N.G. Associations between growth, wood anatomy, carbon isotope discrimination and mortality in a Quercus robur forest. Tree physiology 2011, 31, 298–308. [Google Scholar] [CrossRef]
- Gentilesca, T.; Camele, I.N.; Colangelo, M.; Lauteri, M.; Lapolla, A.; Ripullone, F. Oak forest decline in southern Italy: the study case of Gorgoglione forest. In ATTI del Secondo Congresso Internazionale di Selvicoltura 2015 (Vol. 2, pp. 123–129). ACCADEMIA ITALIANA DI SCIENZE FORESTALI.
- Marques, I.G.; Campelo, F.; Rivaes, R.; Albuquerque, A.; Ferreira, M.T.; Rodríguez-González, P.M. Tree rings reveal long-term changes in growth resilience in Southern European riparian forests. Dendrochronologia 2018, 52, 167–176. [Google Scholar] [CrossRef]
- Duval, C.J.; Gourrat, K.; Perre, P.; Prida, A.; Gougeon, R.D. A HS–SPME–GC–MS analysis of IR heated wood: Impact of the water content on the depth profile of oak wood aromas extractability. Food research international 2013, 54, 277–284. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Virjamo, V.; Ghimire, R.P.; Blande, J.D.; Julkunen-Tiitto, R.; Kivimäenpää, M. Climate change effects on secondary compounds of forest trees in the northern hemisphere. Frontiers in plant science 2018, 9, 395200. [Google Scholar] [CrossRef]
- Wink, M. Introduction: biochemistry, physiology and ecological functions of secondary metabolites. Annual plant reviews volume 40: Biochemistry of plant secondary metabolism 2010, 1-19.
- Lindroth, R.L. Atmospheric change, plant secondary metabolites and ecological interactions. The ecology of plant secondary metabolites: from genes to global processes. Cambridge University Press 2012, Cambridge, 120-153.
- Lämke, J.S.; Unsicker, S.B. Phytochemical variation in treetops: causes and consequences for tree-insect herbivore interactions. Oecologia 2018, 187, 377–388. [Google Scholar] [CrossRef]
- Wasternack, C.; Strnad, M. Jasmonates: News on occurrence, biosynthesis, metabolism and action of an ancient group of signaling compounds. International journal of molecular sciences 2018, 19, 2539. [Google Scholar] [CrossRef]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Current opinion in plant biology 2006, 9, 297–304. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.C.; Caramujo, M.J. The various roles of fatty acids. Molecules 2018, 23, 2583. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Bykova, N.V. Role of organic acids in the integration of cellular redox metabolism and mediation of redox signalling in photosynthetic tissues of higher plants. Free Radical Biology and Medicine 2018, 122, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Kretovich, V.L. Molecular mechanisms of nitrogen assimilation by plants. Molecular mechanisms of nitrogen assimilation by plants 1980. [Google Scholar]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of silicon on plant–pathogen interactions. Frontiers in plant science 2017, 8, 255703. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; ... Yepez, E.A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New phytologist 2008, 178, 719–739. [CrossRef] [PubMed]
- Bartram, S.; Jux, A.; Gleixner, G.; Boland, W. Dynamic pathway allocation in early terpenoid biosynthesis of stress-induced lima bean leaves. Phytochemistry 2006, 67, 1661–1672. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.N.M.; Balakrishnan, R.S.; Shamsudeen, S.; Bahwani, S.A.; Adam, F. A concise review of the natural existance, synthesis, properties, and applications of syringaldehyde. BioResources 2012, 7, 4377–4399. [Google Scholar] [CrossRef]
- Sangeetha, C.; Krishnamoorthy, A.S.; Amirtham, D. Antifungal bioactive compounds from Chinese caterpillar fungus (Ophiocordyceps sinensis (Berk.) GH Sung et al.) against plant pathogens. Madras Agricultural Journal 2015, 102, 1. [Google Scholar]
- Ntalli, N.G.; Ferrari, F.; Giannakou, I.; Menkissoglu-Spiroudi, U. Synergistic and antagonistic interactions of terpenes against Meloidogyne incognita and the nematicidal activity of essential oils from seven plants indigenous to Greece. Pest management science 2011, 67, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Janaki, S.; Zandi-Sohani, N.; Ramezani, L.; Szumny, A. Chemical composition and insecticidal efficacy of Cyperus rotundus essential oil against three stored product pests. International Biodeterioration Biodegradation 2018, 133, 93–98. [Google Scholar] [CrossRef]
- Langsi, J.D.; Nukenine, E.N.; Oumarou, K.M.; Moktar, H.; Fokunang, C.N.; Mbata, G.N. Evaluation of the insecticidal activities of α-Pinene and 3-Carene on Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). Insects 2020, 11, 540. [Google Scholar] [CrossRef]
- Sfara, V.; Zerba, E.N.; Alzogaray, R.A. Fumigant insecticidal activity and repellent effect of five essential oils and seven monoterpenes on first-instar nymphs of Rhodnius prolixus. Journal of medical entomology 2014, 46, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Wahyuni, R.; Triadiati, T.; Falah, S. Induction of agarwood in Aquilaria malaccensis using nitrogen fertilizer and Fusarium solani. Jurnal Penelitian Kehutanan Wallacea 2018, 7, 165–171. [Google Scholar] [CrossRef]
- Martinez, R.G.; De La Serrana, H.L.G.; Mir, M.V.; Granados, J.Q.; Martinez, M.L. Influence of wood heat treatment, temperature and maceration time on vanillin, syringaldehyde, and gallic acid contents in oak wood and wine spirit mixtures. American Journal of Enology and Viticulture 1996, 47, 441–446. [Google Scholar] [CrossRef]
- Abbas, F.; O'Neill Rothenberg, D.; Zhou, Y.; Ke, Y.; Wang, H.C. Volatile organic compounds as mediators of plant communication and adaptation to climate change. Physiologia Plantarum 2022, 174, e13840. [Google Scholar] [CrossRef] [PubMed]
- Ton, J.; D'Alessandro, M.; Jourdie, V.; Jakab, G.; Karlen, D.; Held, M.; ... Turlings, T.C. Priming by airborne signals boosts direct and indirect resistance in maize. The Plant Journal 2007, 49, 16–26. [CrossRef]
- Camarero, J.J.; Sangüesa-Barreda, G.; Vergarechea, M. Prior height, growth, and wood anatomy differently predispose to drought-induced dieback in two Mediterranean oak speciesk. Annals of Forest Science 2016, 73, 341–351. [Google Scholar] [CrossRef]
Figure 1.
Average BAI of dieback (D) and non-dieback (ND) trees over five-year periods.
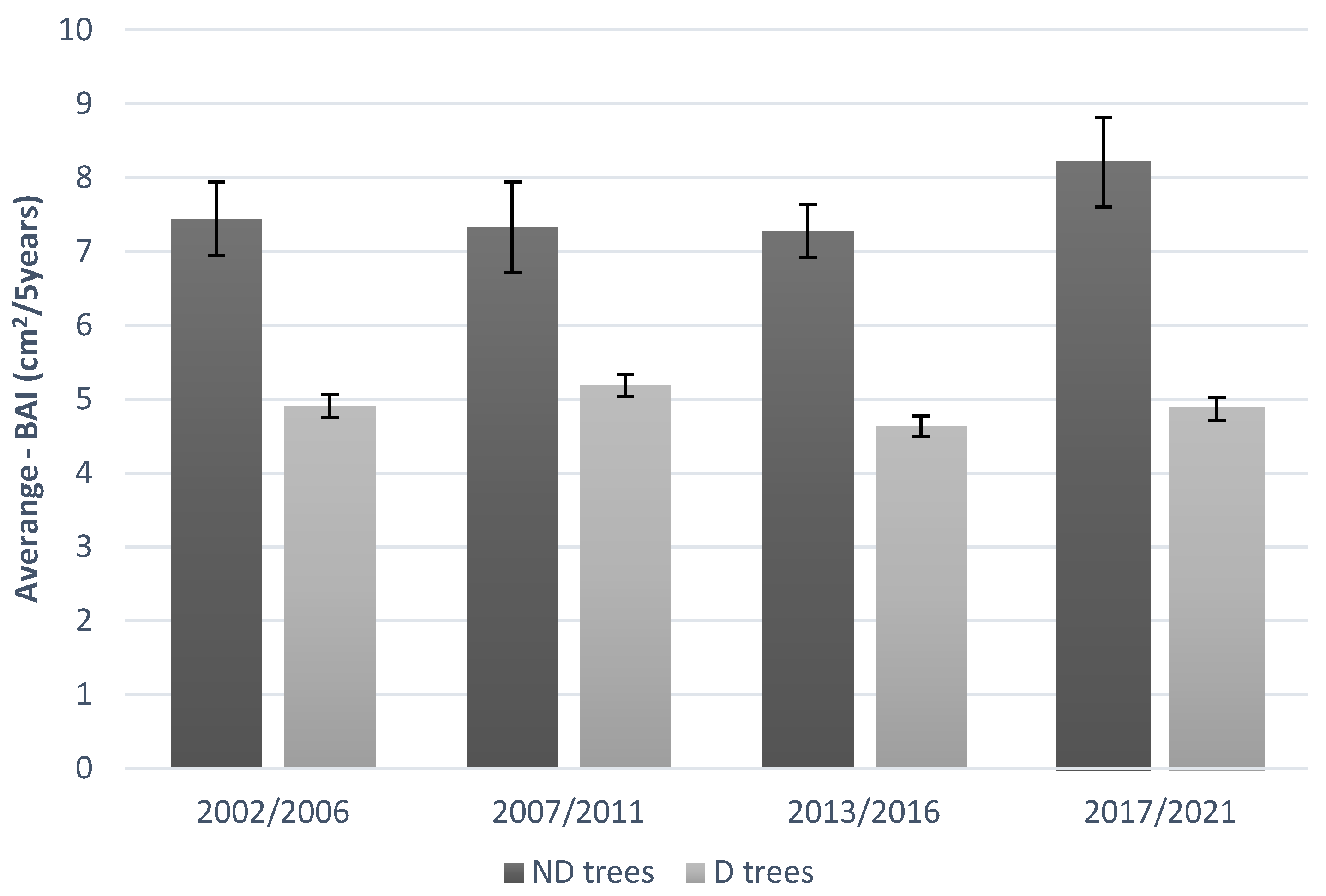
Figure 2.
Means of the total areas of the classes of compounds of dieback (D) and non-dieback (ND) trees.
Figure 2.
Means of the total areas of the classes of compounds of dieback (D) and non-dieback (ND) trees.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



