Submitted:
09 May 2024
Posted:
10 May 2024
You are already at the latest version
Abstract
Drug-induced liver injury (DILI) stands as a significant cause of drug attrition. To mitigate clinical DILI risks, assessing drugs using human liver models is crucial since animal studies may fall short due to species-specific liver pathway variations. Cell-based preclinical hepatotoxicity testing is often pertinent. In the present study, cells from a human liver cancer line (HepG2, HepaRG) were cultured in both formats of 2D and 3D spheroids to explore their responses to drugs. Liver-specific marker expressions across cell lines and culture formats were also examined to assess disparities in drug-induced liver injury marker expressions. In cytotoxicity assays, HepG2 cells exhibited heightened sensitivity to amiodarone HCl, while HepaRG cells showed greater sensitivity to acetaminophen. Generally, 2D cultures were more responsive to drugs than 3D cultures. Consistency was observed between cytotoxicity assays and liver injury markers AST and ALT. Variations were also noted in liver cell marker secretion, with albumin and urea concentrations declining post-drug treatment, particularly with higher baseline expression in HepaRG cells and 3D cultures. Additionally, mRNA expression levels of CYP enzymes involved in hepatocellular drug metabolism were compared following treatment with enzyme inducers. CYP1A2 and CYP2C9 were not expressed in HepG2 cells. On the other hand, CYP3A4 was expressed higher in 3D culture than in 2D culture. HepaRG cells exhibited significantly increased expression of all three enzymes post-treatment. Notably, enzyme expression was notably higher in 3D cultures than in 2D cultures. Collectively, these findings suggest that HepaRG cells and 3D cultures hold promise for evaluating DILI during early-stage drug development.
Keywords:
2D and 3D cultures
; HepG2
; HepaRG
; liver injury markers
; CYP450
1. Introduction
DILI stands as a leading cause of preclinical or clinical drug attrition and acute liver failures [1]. Nearly 100 marketed drugs can potentially induce various types of liver injury, ranging from cell necrosis to fibrosis, or a combination thereof [2]. The severity of DILI can fluctuate based on factors such as the patient’s age, gender, genetics, and underlying medical conditions. Unfortunately, the requirement by regulatory bodies such as the Food and Drug Administration (FDA) for live animal testing during preclinical drug development can only detect less than half of human DILIs primarily due to species-specific differences in drug pathways and an inability to accurately replicate human genetics and disease backgrounds [3]. Such challenges inherent in animal-based drug screening have underscored the increasing importance of in vitro human liver cultures over the past decade or so, with in vitro human liver culture becoming a cornerstone of drug discovery [4]. The process of drug discovery has historically relied on both in vitro and in vivo methods to assess the safety of compounds, with recent emphasis on high-throughput and cost-effective approaches such as cell-based screening assays. Traditionally, these in vitro cell-based assays are performed using 2D monolayer cell cultures, which poorly mimic in vivo conditions. 2D monolayer cultures lack numerous biological functions, such as cell-cell and cell-substrate contact. These features lead to alterations in cell signaling pathways and reduction or loss of enzymes involved in the metabolism of foreign substances [5,6]. It is increasingly recognized that cells cultured in 3D environments more closely mimic normal cell function by mimicking in vivo structures of natural tissues and organs [7,8,9]. In addition, studies have shown that hepatocytes cultured in 3D form have similar structure and function to liver [10]. Therefore, tremendous effort has been put into the development of a variety of 3D cell models, which hold the promise for applications in drug discovery, cancer cell biology, stem cell research, safety studies, and many other cell-based analyses to bridge the traditional 2D monolayer cell culture models and whole-animal systems. HepG2 cells remain popular due to their ease of handling and widespread use as one of the oldest and most well-known hepatic cell lines [11]. Previous studies have shown that culturing HepG2 cells for at least 72 hours is sufficient metabolic capacity for in vitro toxicity studies [12,13]. However, HepG2 cells are known to have low activities of several metabolic enzymes and low expression levels of genes encoding cytochrome P450 enzymes that are essential in drug metabolism [12]. On the other hand, HepaRG cells derived from HCC express functional liver markers including various CYP and phase II enzymes, nuclear receptors, and membrane transporters. Additionally, HepaRG cells have been proven to be excellent surrogates for primary human hepatocytes, particularly in CYP3A4 induction studies [14]. In this study, we cultured HepG2 and HepaRG cells and exposed them to hepatotoxicants ADR and AAP to compare their responses. Moreover, we cultured cells in both formats of 2D and 3D spheroids to compare cytotoxicity and hepatic characteristics following drug treatment, aiming to identify which cells and culture types would be the most suitable for drug screening studies.
2. Materials and Methods
2.1. Chemicals
Amiodarone HCl (ADR) and acetaminophen (AAP) were purchased from Sigma-Aldrich (St. Louis, MO, USA). These hepatotoxicants were selected based on published information regarding their hepatotoxic potential and mechanism of action [15,16,17]. CYP inducers omeprazole and rifampin were also purchased from Sigma-Aldrich (St. Louis, MO, USA) [18]. All chemicals were initially dissolved in dimethyl sulfoxide (DMSO, Sigma-Aldrich, USA) to obtain concentrated stock solutions. They were then diluted with culture medium before use. The non-treated control group was incubated with a culture medium including a final concentration of 0.1% DMSO.
2.2. Cell Culture and CYP450 Enzyme Induction
The human Hepatocellular carcinoma cell line (HepG2) was purchased from ATCC (VA, USA). HepG2 cells were cultured in Eagle’s minimum essential medium (EMEM, ATCC, USA) supplemented with 10% fetal bovine serum (FBS, Merk Millipore, USA) and 1% penicillin and streptomycin (Millipore, USA). HepG2 cells were cultured in T-75 cell culture flask (Corning, USA) at 37℃ in a 5% CO2 humidified incubator. Cryopreserved HepaRG cells (Gibco, Germany) were purchased from Thermo Fisher Scientific and maintained according to the manufacturer’s instructions. HepaRG cells were cultured in William’s E medium (Gibco, USA) added with HepaRG™ Thaw, Plate & General Purpose Medium supplement (Gibco, USA) and 1% penicillin and streptomycin (Millipore, USA). HepaRG cells were cultured in T-75 cell culture flasks (Corning, USA) at 37℃ in a 5% CO2 humidified incubator. When HepG2 and HepaRG cells reached about 80% confluency, cells were trypsinized (0.25% Trypsin-EDTA, Gibco, USA) for 5 min, added with medium to neutralize trypsin, and centrifuged at 1,100 rpm for 5 min at 4℃. The supernatant was removed and cells were re-suspended in medium. These cells were seeded at a density of 1.5×104 cells/well in a total volume of 100 μl in ultra-low attachment 96-well plates (BD Bioscience, USA) and incubated at 37℃ in a 5% CO2 humidified incubator. For CYP enzyme induction, Cells were cultured in ultra-low attachment 96-well plates. Omeprazole was used as a CYP1A2 inducer. Rifampicin was used as an inducer for CYP2C9 and CYP3A4. When cell density reached more than 80%, cells were incubated according to the condition listed in Table 1 to induce the expression of CYP enzymes.
2.3. In Vitro Cell Viability Assay
CellTiter-Glo® 2D Cell Viability Assay and CellTiter-Glo® 3D Cell Viability Assay (Promega, USA) were used to measure ATP as an indicator of viability and generate a luminescent readout reported to be more sensitive than other methods [19,20]. These assays work by lysing cells with their proprietary lytic component. They differ solely by their lytic capabilities. Promega has demonstrated that 3D cell viability assay kit has a higher ability than 2D cell viability assay kit to lyse cells within small spheroids. Therefore, the 3D cell viability assay kit was hypothesized to work better in our studies. Briefly, in 2D assays, 50 μl of PBS was added to each well followed by the addition of 50 μl CellTiter-Glo reagent which was prepared according to the manufacturer’s protocol. These contents were mixed in a plate shaker for 2 min to induce cell lysis and then incubated for 10 min at room temperature. The cellular lysate was then transferred to a white opaque 96-well plate. Luminescent signal was measured using a luminometer (FlexStation III; Molecular Device, USA). In 3D assays, plates were mixed vigorously for 5 min to induce cell lysis before incubating at room temperature for 25 min to stabilize the luminescent signal. Finally, 50 μl of the solution was transferred to a white opaque 96-well plate to be measured with a luminometer as a relative luminescence unit (RLU) with an integration time of 1 sec.
2.4. Lactate Dehydrogenase (LDH), Aspartate Aminotransferase (AST), and Aspartate Aminotransferase (AST) Assays for Detecting Liver Injury Markers
To measure enzyme leaking due to cellular damage, lactate dehydrogenase (LDH) activities were estimated using a LDH cytotoxicity detection kit (Takara Bio, Japan) according to the manufacturer’s instruction. In brief, HepG2 and HepaRG cells were seeded into 96-well cell culture plates (1.5×104 cells/ml) with culture medium and exposed to hepatotoxicants. Following a 24 hr treatment period, cells were used to determine lactate dehydrogenase (LDH) activities according to the manufacturer’s instructions. Absorbance was read on a luminometer (FlexStation III) at a wavelength of 490 nm. Cell-free wells were used to eliminate absorption by nanoparticles in the background.
Serum aspartate aminotransferase (AST) and transaminases alanine transaminase (ALT) levels were measured using commercial kits specific for each test following the manufacturer’s instruction. Briefly, ALT level was measured using an ALT activity assay kit (Sigma-Aldrich, USA). Serum samples were added to cell-free wells containing the reaction mix. AST level was measured using an AST activity assay kit (Sigma-Aldrich, USA). After incubated with reaction mix for 5 min at 37℃, absorbance was read on a luminometer (FlexStation III) at a wavelength of 570 nm for ALT and 450 nm for AST every 5 min for up to 30 min. After completing the measurement, values were calculated by subtracting the starting measurement from the last measurement.
2.5. Albumin and Urea Secretion
HepG2 and HepaRG cells were seeded into 96-well cell culture plates (1.5×104 cells/ml) with culture medium. On the following day, cells were treated with serial concentrations of ADR and AAP for 24 hr. Supernatants collected from each well were centrifuged at 3,000 x g for 5 min to remove floating cells. Supernatants were then stored at -20 ℃ until assay. Albumin and urea amounts in culture medium were estimated using an Albumin Human ELISA kit (Abnova, USA) and a Urea assay kit (Cell Biolabs, CA, USA), respectively, according to each manufacturer’s instructions. Absorbance was read on a luminometer (FlexStation III) at a wavelength of 450 nm for ALB and 630 nm for Urea. Albumin and urea amounts were calculated using each standard curve and normalized according to cell count (1.5×104 cell/ml).
2.6. Quantitative RT-PCR Analysis of CYP450 Enzyme mRNA
RNA was extracted from cells using the RNeasy Mini Kit (Qiagen, Germany). The concentration and purity of RNAs were assessed with an ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). cDNA synthesis was performed using 1 μg of the total RNA with an Omniscript RT kit (Qiagen, Germany). PCR cycling conditions consisted of denaturation at 95℃ for 10 sec, annealing at 45℃ to 64℃ for 10 sec, and extension at 72℃ for 20 sec for up to 40 cycles. After reverse transcription of RNA into cDNA, expression levels of CYP1A2, CYP2C9, and CYP3A4 were measured by qPCR. Primers were used to amplify targeted genes using a CFX Real-Time PCR Detection System (Bio-Rad, USA). The mRNA level of each test gene (CYP1A2, CYP2C9, and CYP3A4) was normalized to β-actin according to the following formula: △CT = CT (test gene) – CT (β-actin). Thereafter, relative mRNA level of each gene was calculated using the △△CT method: △△CT (test gene) = △CT (test gene in the treated sample) - △CT (test gene in the untreated sample). The fold change of mRNA level was expressed as the relative expression of 2-△△CT. A fold induction greater than 2 relative to the vehicle control and at least 20% relative to the positive control was defined as an induction. Sequences of primer sets are listed in Table 2.
2.7. Statistical Analysis
Experimental data were analyzed by two-way analysis of variance (ANOVA) followed by Bonferroni posttest using GraphPad Prism software version 5.0. Data were considered significant at p < 0.05 (*). TC-50 value was calculated using GraphPad Prism software (logarithmic transformation of X-values and nonlinear regression sigmoidal dose response analysis with variable slope-with bottom and top constraints set at 0 and 100, respectively). Values were given ± 95% confidence intervals.
3. Result
3.1. Cytotoxicities of ADR and AAP to HepG2 and HepaRG Cells Depending on Culture Type
Cytotoxicities of ADR and AAP were evaluated based on cell viability using two different cell culture types (2D and 3D cultures) and compared with those of HepG2 and HepaRG cells without ADR or AAP treatment. ADR and AAP decreased cell viability of both cell lines and culture types in a dose-dependent manner (Figure 1a,b). However, different responses were seen depending on the cell culture type. For both cells, cell viability was rapidly decreased in 2D culture, showing significant differences from 3D culture. The 50% inhibitory concentration (IC50) value of ADR was found to be 44.85 μM for HepG2 2D, 65.47 μM for HepaRG 2D, 55.64 μM for HepG2 3D, and 109.38 μM for HepaRG 3D. The IC50 value of AAP was determined to be 14.95 mM for HepG2 2D, 7.11 mM for HepaRG 2D, 48.43 mM for HepG2 3D, and 26.19 mM for HepaRG 3D (Table 3). Overall, 2D cultures were found to be more sensitive to drugs than 3D cultures.
3.2. Detection of Drug-Induced Hepatic Injury Markers
To measure the expression of LDH, AST and ALT (liver injury markers), HepG2 cells and HepaRG cells were cultured in 2D and 3D and then treated with hepatotoxicants ADR and AAP. Activities of LDH (Figure 2a,b), AST (Figure 2c,d), and ALT (Figure 2e,f) were increased by treatment with ADR or AAP in a dose-dependent manner in both cell lines and culture types. However, different responses were seen depending on the type of cell culture. It was confirmed that their activities in both cells were increased more in 2D culture than in 3D culture in a drug concentration-dependent manner. HepG2 cells showed higher expression of liver injury factors after ADR treatment than HepaRG cells and 2D cultures showed higher expression of these factors than 3D cultures. HepaRG cells showed higher expression of these factors after AAP treatment than HepG2 cells and 2D cultures showed higher expression of factors than 3D cultures. These results were consistent with results of the cell viability assay.
3.3. Analysis of Hepatocyte Specific Factors ALB and Urea Secretion
To determine effects of cell culture type and treatment with hepatotoxicants on expression levels of liver specific markers, differences in ALB and urea secretion between 2D and 3D cultures were analyzed. Each result was based on the control of HepG2 2D culture. Quantitative analysis of albumin showed that its expression levels were higher in 3D cultures and HepaRG cells than in 2D cultures and HepG2 cells, respectively. In addition, after culturing each cell, cells were treated with ADR or AAP to analyze changes in albumin and urea secretion depending on the drug concentration. As a result, it was found that albumin secretion was reduced depending on the concentration of each drug (Figure 3a,b). Urea secretion analysis gave the same results as albumin secretion (Figure 3c,d).
3.4. Compare Expression Levels CYP450 Enzymes after Drugs Treatment
We treated each cell with a CYP enzyme inducer and then examined gene expression level of the enzyme to determine which cells and culture types had higher expression levels of the CYP450 enzyme known to be involved in drug metabolism in the liver. Each result was based on the control of HepG2 2D culture. Expression levels of CYP1A2, CYP2C9, and CYP3A4 enzymes in HepG2 and HepaRG cells were found to show significant differences after treatment with drugs. In HepaRG cells, expression levels of CYP1A2, CYP2C9, and CYP3A4 were increased after treatment with inducers. Compared with HepaRG cells, HepG2 cells had relatively lower levels of CYP1A2, CYP2C9, and CYP3A4. Additionally, significant differences were seen between 2D and 3D cultures. There was no difference in the expression of CYP1A2 and CYP2C9 in 2D and 3D cultures of HepG2 cells. However, CYP3A4 had higher expression levels in 3D cultures than in 2D cultures. In HepaRG cells, all three enzymes CYP1A2, CYP2C9, and CYP3A4 were found to have higher expression levels in 3D cultures than in 2D cultures. Activities of CYP1A2 in 2D and 3D cultures of HepaRG were significantly (p < 0.05) increased after treatment with 25 μM of omeprazole (Figure 4a). Activities of CYP2C9 and CYP3A4 were significantly (p < 0.05) increased after treatment with 10 μM rifampin (Figure 4b,c). In HepG2 cell 2D and 3D cultures, the activity of CYP3A4 was significantly (p < 0.05) increased after treatment with rifampin at 10 μM (Figure 4c). But, CYP1A2 and CYP2C9 were not increased (p < 0.05) in 2D and 3D cultures of HepG2 cells (Figure 4a,b).
4. Discussion
Authors While the current gold standard for drug metabolism and toxicology studies is the use of primary human liver cells (PHHs) [21], their short lifespan, high cost, limited availability and the observed inter-donor variability severely restrict their use in high throughput in vitro drug screening. Therefore, the immortalised hepatic lines HepG2 and HepaRG were used in this study. While HepG2 cells have many limitations in 2D culture, including loss of liver-specific structure and functionality [21], the ease of handling compared to PHH, coupled with a stable phenotype and lack of donor variability [21,22], offers many advantages for standard operating procedures (SOPs) development in preclinical drug screening. On the other hand, culturing HepG2 cells in a 3D environment has been shown to restore some of the characteristics lost when cultured in a 2D environment. In addition, 3D cultured HepG2 cells has a reduced proliferation rate, self-organise and differentiate into 3D spheroids that restore lost liver structure and function [23,24]. HepaRG cells are a hepatocellular carcinoma-derived human cell line expressing functional liver markers such as several cytochrome P450 (CYP) and phase II enzymes, nuclear receptors and membrane transporters. HepaRG cells have been shown to be an excellent surrogate for primary human hepatocytes in cytochrome P450 3A4 (CYP3A4) induction studies. Therefore, we cultured cells in 2D and 3D cell culture type using HepG2 and HepaRG cells, which are most commonly used for hepatotoxicity studies. We also compared the response to drug treatment by each cell and culture type. Cell viability was measured for both 2D and 3D spheroid viability using CellTiter-Glo® ATP quantification. In parallel with the assessment of viability by quantification of ATP, the LDH assay was used to determine the cytotoxicity induced by ADR and AAP. The LDH assay avoids the problems of having to dissociate the spheroid structure as required by other methods of assessing membrane integrity, such as neutral red or trypan blue, because it is supernatant based, with the cytosolic enzyme LDH being released into the culture supernatant by compromised plasma membranes. When evaluated using these assays, we observed decreased viability or increased cytotoxicity induction after treatment with ADR and AAP in both culture types. When comparing the responsiveness to the drugs in each cell type, HepG2 cells were more sensitive to ADR treatment, while HepaRG cells were more sensitive to AAP treatment. Changes in cell viability based on cell culture type were also observed. For both HepG2 and HepaRG cells, it was observed that 2D cultures were more sensitive to drugs than 3D cultures. These results were also observed as a result of cytotoxicity induction. The expression of LDH increased in a drug concentration-dependent manner, with 2D cultures being more sensitive than 3D cultures. AST and ALT are markers of liver injury that increase when cells are damaged [25]. Two well-known markers of liver function are albumin and urea. Albumin is a major protein synthesized in the liver and plays an important role in maintaining tumor pressure, transporting various molecules and regulating pH balance. Urea is a waste product formed in the liver during protein metabolism. The liver's ability to metabolize ammonia into urea is crucial for maintaining nitrogen balance in the body [26,27]. HepG2 and HepaRG cells were cultured in 2D and 3D forms and each cell was treated with ADR and AAP to determine the extent of liver damage. Similar to cell viability and toxicity tests, the expression of AST and ALT increased in a drug concentration-dependent manner. Similar results were observed for the secretion of ALB and Urea. Secretion decreased in a concentration-dependent manner and expression was significantly reduced in 2D cultures. In addition, HepaRG cells had higher basal expression compared to HepG2 cells when tested under the same conditions. In the same cells, expression of ALB and urea was higher in cells cultured in a 3D cultures. The hepatotoxicants ADR and AAP cause mitochondrial dysfunction [28] and liver failure [29]. ADR and AAP has been reported to cause hepatotoxicity and decrease cell viability in HepG2 [30,31] and HepaRG [32,33] cells. These findings are consistent with our results which showed decreased cell viability, increased expression of liver injury markers, and decreased liver function after drug treatment. The evaluation of DILI using hepatocytes relies on an accurate understanding of the characteristics of the cells and drugs, which is difficult to achieve. There are studies that show different liver cells have different responses to the same drug [34]. Alternatively, studies have shown that when a single cell is treated with different drugs and shows varying sensitivity to each drug [17]. Therefore, it is important to study liver toxicity using a variety of cells and drugs, as we have done in our study. A number of critical enzymes play an important role in cellular metabolism under disease conditions or in the metabolism of drugs. Cytochrome P450 enzymes have been extensively studied since their identification because of their involvement in the metabolism of various substrates, including therapeutic agents and components of biological processes, in humans [31,35]. In particular, over 95% of oxidative and reductive reactions are reported to be catalysed by CYPs [31]. They mediate the activation/inactivation of exogenous substrates, including drugs, xenobiotics, food additives or endogenous compounds such as steroids, eicosanoids or fatty acids [37,38]. In particular, CYP1A2, CYP2C9 and CYP3A4 enzymes are known to be the most involved in drug metabolism in human hepatocytes. CYP1A2 is a member of the cytochrome P450 enzyme family primarily found in the liver, although it is also present in other tissues. This enzyme is responsible for the metabolism of various substrates, including caffeine, theophylline, certain drugs, and environmental toxins like polycyclic aromatic hydrocarbons found in cigarette smoke. CYP1A2 has been extensively studied due to its role in drug metabolism and its potential impact on drug efficacy and toxicity [39]. CYP2C9 is another important cytochrome P450 enzyme predominantly expressed in the liver, although it is also found in other tissues. CYP2C9 substrate specificity and catalytic properties have been extensively studied to understand its role in drug metabolism and pharmacokinetics [40]. CYP3A4 is one of the most abundant cytochrome P450 enzymes in the liver and intestine, playing a pivotal role in the metabolism of a wide range of drugs. It metabolizes approximately 50% of clinically used drugs, including statins, immunosuppressants, antivirals, and many others. Due to its broad substrate specificity and significant contribution to drug metabolism, CYP3A4 is a major determinant of drug clearance and efficacy [41]. In this study, we compared the mRNA expression of CYP enzymes in HepG2 and HepaRG cells in 2D and 3D cultures. The expression of each enzyme was induced by treating the cells with known standard inducers. HepG2 cells showed no expression of CYP1A2 and CYP2C9 enzymes in both 2D and 3D cultures. On the other hand, the expression of CYP3A4 was increased by inducer treatment. It also showed higher expression in 3D culture. As demonstrated in various studies, HepG2 cells normally express CYP1A2 [42]. However, there is evidence that while caffeine induces CYP1A2 expression in rat hepatocytes, it may not have the same effect in human HepG2 cells [43]. HepG2 cells generally express CYP1A2, but the expression level and regulatory factors can vary depending on the experimental conditions and stimuli. There is also evidence that HepG2 cells have low expression of CYP2C9, an important enzyme involved in drug metabolism [44,45,46]. This low expression affects their suitability as a model for studying hepatotoxicity and drug metabolism[47,48]. It is important to consider this limitation when using HepG2 cells as a model system for drug metabolism research. In contrast to HepG2 cells, HepaRG cells showed expression of CYP1A2, CYP2C9 and CYP3A4 enzymes in both 2D and 3D cultures. Furthermore, we found that all three enzymes were more highly expressed in 3D cultures than in 2D cultures. Interestingly, the expression of all enzymes was much higher than in HepG2 cells. HepaRG cells are a widely used model for drug metabolism studies due to their ability to express a variety of cytochrome P450 enzymes [49]. They respond to typical inducers by inducing a variety of cytochrome P450 enzymes, including CYP1A2, CYP2C9, and CYP3A4 enzymes, which are essentially involved in human drug metabolism [50]. HepaRG cells are known to be metabolically similar to human adult hepatocytes, making them useful for drug metabolism and toxicity studies [51]. These findings corroborate our results that HepaRG cells have a higher capacity to induce CYP enzymes compared to HepG2 cells. Furthermore, our results are consistent with studies showing increased mRNA levels of CYP1A2, CYP3A4 [52,53] and increased expression of drug metabolising enzymes including CYP2C9 in HepaRG 3D cultures [54]. In conclusion, the development of in vitro models that can predict DILI at the preclinical stage for drug development is becoming increasingly important. Therefore, the selection of in vitro models that can study a clearer and more diverse range of drug responses is important. In this study, HepG2 cells and HepaRG cells, which are often used in drug screening were compared, and each cell was cultured in 2D and 3D forms to evaluate them as in vitro models for drug development. The results of the study showed that the sensitivity of the cells varied depending on the type of cell, culture type and type of drug. In addition, HepaRG cells with higher expression of CYP enzymes involved in human drug metabolism are more suitable for drug metabolism evaluation, and 3D culture is more suitable as a model for drug metabolism evaluation than 2D culture. However, the results of CYP enzymes need to be validated by further experiments on metabolic mechanisms through enzyme activity measurement or LC-MS/MS metabolite.
Author Contributions
Nam-Ju and Byung-Suk contributed to the design and implementation of the research. Ji-Hyun Bang, Hee Yi, Hyun-Ok Ku and Seon-Jong Yun contributed to the analysis and interpretation of the results. Nam-Ju took the lead in writing the manuscript with input from Byung-Suk, Ji-Hyun Bang, Hee Yi, Hyun-Ok Ku and Seon-Jong Yun. All authors provided critical feedback that helped shape the research and approved the submitted manuscript version.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Dataset available on request the authors.
Acknowledgments
The project was supported by research funds (B-1543073-2022-24-01) from Animal and Plant Qurantine Agency, Republic of Korea.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
References
- Kaplowitz, N. Idiosyncratic drug hepatotoxicity. Nature reviews Drug discovery 2005, 4, 489–49. [Google Scholar] [CrossRef]
- Abboud, G.; Kaplowitz, N. Drug-induced liver injury. Drug safety 2007, 30, 277–294. [Google Scholar] [CrossRef]
- Olson, H.; Betton, G.; Robinson, D.; Thomas, K.; Monro, A.; Kolaja, G.; Lilly, P.; Sanders, J.; Sipes, G.; Bracken, W. Concordance of the toxicity of pharmaceuticals in humans and in animals. Regulatory toxicology and pharmacology 2000, 32, 56–67. [Google Scholar] [CrossRef]
- Lin, C.; Khetani, S.R. Advances in engineered liver models for investigating drug-induced liver injury. BioMed research international 2016, 2016. [Google Scholar] [CrossRef]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-dimensional cell culture systems and their applications in drug discovery and cell-based biosensors. Assay and drug development technologies 2014, 12, 207–218. [Google Scholar] [CrossRef]
- Wrzesinski, K.; J Fey, S. From 2D to 3D-a new dimension for modelling the effect of natural products on human tissue. Current Pharmaceutical Design 2015, 21, 5605–5616. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, S.-T. High-throughput 3-D cell-based proliferation and cytotoxicity assays for drug screening and bioprocess development. Journal of biotechnology 2011, 151, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Fey, S.J.; Wrzesinski, K. Determination of drug toxicity using 3D spheroids constructed from an immortal human hepatocyte cell line. Toxicological sciences 2012, 127, 403–411. [Google Scholar] [CrossRef]
- Gunness, P.; Mueller, D.; Shevchenko, V.; Heinzle, E.; Ingelman-Sundberg, M.; Noor, F. 3D organotypic cultures of human HepaRG cells: a tool for in vitro toxicity studies. Toxicological sciences 2013, 133, 67–78. [Google Scholar] [CrossRef]
- Bell, C.C.; Hendriks, D.F.; Moro, S.M.; Ellis, E.; Walsh, J.; Renblom, A.; Fredriksson Puigvert, L.; Dankers, A.C.; Jacobs, F.; Snoeys, J. Characterization of primary human hepatocyte spheroids as a model system for drug-induced liver injury, liver function and disease. Scientific reports 2016, 6, 25187. [Google Scholar] [CrossRef]
- Hart, S.N.; Li, Y.; Nakamoto, K.; Subileau, E.-a.; Steen, D.; Zhong, X.-b. A comparison of whole genome gene expression profiles of HepaRG cells and HepG2 cells to primary human hepatocytes and human liver tissues. Drug metabolism and disposition 2010, 38, 988–994. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.J.; Edvardsson, A. Validation of a multiparametric, high-content-screening assay for predictive/investigative cytotoxicity: evidence from technology transfer studies and literature review. Chemical Research in Toxicology 2017, 30, 804–829. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.J.; Irwin, W.; Diaz, D.; Howard-Cofield, E.; Krejsa, C.M.; Slaughter, M.R.; Gao, B.; Kaludercic, N.; Angeline, A.; Bernardi, P. High concordance of drug-induced human hepatotoxicity with in vitro cytotoxicity measured in a novel cell-based model using high content screening. Archives of toxicology 2006, 80, 580–604. [Google Scholar] [CrossRef] [PubMed]
- McGinnity, D.F.; Zhang, G.; Kenny, J.R.; Hamilton, G.A.; Otmani, S.; Stams, K.R.; Haney, S.; Brassil, P.; Stresser, D.M.; Riley, R.J. Evaluation of multiple in vitro systems for assessment of CYP3A4 induction in drug discovery: human hepatocytes, pregnane X receptor reporter gene, and Fa2N-4 and HepaRG cells. Drug metabolism and disposition 2009, 37, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lechon, M.; Lahoz, A.; Gombau, L.; Castell, J.; Donato, M. In vitro evaluation of potential hepatotoxicity induced by drugs. Current pharmaceutical design 2010, 16, 1963–1977. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, L.; Pinto, S.; Donato, M.T.; Lahoz, A.; Castell, J.V.; O’Connor, J.E.; Gómez-Lechón, M.J. Development of a multiparametric cell-based protocol to screen and classify the hepatotoxicity potential of drugs. Toxicological Sciences 2012, 127, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Tham, N.T.; Hwang, S.-R.; Bang, J.-H.; Yi, H.; Park, Y.-I.; Kang, S.-J.; Kang, H.-G.; Kim, Y.-S.; Ku, H.-O. High-content analysis of in vitro hepatocyte injury induced by various hepatotoxicants. Journal of Veterinary Science 2019, 20, 34. [Google Scholar] [CrossRef]
- Huang, S.M.; Temple, R.; Throckmorton, D.; Lesko, L. Drug interaction studies: study design, data analysis, and implications for dosing and labeling. Clinical Pharmacology & Therapeutics 2007, 81, 298–304. [Google Scholar]
- Kleijn, A.; Kloezeman, J.; Balvers, R.; Kaaij, M.v.d.; Dirven, C.; Leenstra, S.; Lamfers, M. A systematic comparison identifies an ATP-based viability assay as most suitable read-out for drug screening in glioma stem-like cells. Stem cells international 2016, 2016. [Google Scholar] [CrossRef]
- Crouch, S.; Kozlowski, R.; Slater, K.; Fletcher, J. The use of ATP bioluminescence as a measure of cell proliferation and cytotoxicity. Journal of immunological methods 1993, 160, 81–88. [Google Scholar] [CrossRef]
- LeCluyse, E.L.; Witek, R.P.; Andersen, M.E.; Powers, M.J. Organotypic liver culture models: meeting current challenges in toxicity testing. Critical reviews in toxicology 2012, 42, 501–548. [Google Scholar] [CrossRef]
- Gomez-Lechon, M.; Donato, M.; Lahoz, A.; Castell, J. Cell lines: a tool for in vitro drug metabolism studies. Current drug metabolism 2008, 9, 1–11. [Google Scholar] [CrossRef]
- Wrzesinski, K.; Magnone, M.C.; Hansen, L.V.; Kruse, M.E.; Bergauer, T.; Bobadilla, M.; Gubler, M.; Mizrahi, J.; Zhang, K.; Andreasen, C.M. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology research 2013, 2, 163–172. [Google Scholar] [CrossRef]
- Bokhari, M.; Carnachan, R.J.; Cameron, N.R.; Przyborski, S.A. Culture of HepG2 liver cells on three dimensional polystyrene scaffolds enhances cell structure and function during toxicological challenge. Journal of anatomy 2007, 211, 567–576. [Google Scholar] [CrossRef]
- McGill, M.R. The past and present of serum aminotransferases and the future of liver injury biomarkers. EXCLI journal 2016, 15, 817. [Google Scholar]
- Spinella, R.; Sawhney, R.; Jalan, R. Albumin in chronic liver disease: structure, functions and therapeutic implications. Hepatology international 2016, 10, 124–132. [Google Scholar] [CrossRef]
- Nagamani, S.C.; Ali, S.; Izem, R.; Schady, D.; Masand, P.; Shneider, B.L.; Leung, D.H.; Burrage, L.C. Biomarkers for liver disease in urea cycle disorders. Molecular genetics and metabolism 2021, 133, 148–156. [Google Scholar] [CrossRef]
- Yan, M.; Huo, Y.; Yin, S.; Hu, H. Mechanisms of acetaminophen-induced liver injury and its implications for therapeutic interventions. Redox biology 2018, 17, 274–283. [Google Scholar] [CrossRef]
- Essrani, R.; Mehershahi, S.; Essrani, R.K.; Ravi, S.J.K.; Bhura, S.; Sudhakaran, A.; Hossain, M.; Mehmood, A. Amiodarone-induced acute liver injury. Case Reports in Gastroenterology 2020, 14, 87–90. [Google Scholar] [CrossRef]
- Behrends, V.; Giskeødegård, G.F.; Bravo-Santano, N.; Letek, M.; Keun, H.C. Acetaminophen cytotoxicity in HepG2 cells is associated with a decoupling of glycolysis from the TCA cycle, loss of NADPH production, and suppression of anabolism. Archives of toxicology 2019, 93, 341–353. [Google Scholar] [CrossRef]
- Zahno, A.; Brecht, K.; Morand, R.; Maseneni, S.; Török, M.; Lindinger, P.W.; Krähenbühl, S. The role of CYP3A4 in amiodarone-associated toxicity on HepG2 cells. Biochemical pharmacology 2011, 81, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Allard, J.; Bucher, S.; Massart, J.; Ferron, P.-J.; Le Guillou, D.; Loyant, R.; Daniel, Y.; Launay, Y.; Buron, N.; Begriche, K. Drug-induced hepatic steatosis in absence of severe mitochondrial dysfunction in HepaRG cells: proof of multiple mechanism-based toxicity. Cell Biology and Toxicology 2021, 37, 151–175. [Google Scholar] [CrossRef]
- McGill, M.R.; Yan, H.M.; Ramachandran, A.; Murray, G.J.; Rollins, D.E.; Jaeschke, H. HepaRG cells: a human model to study mechanisms of acetaminophen hepatotoxicity. Hepatology 2011, 53, 974–982. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-J.; Lee, H.-M.; Park, Y.-I.; Yi, H.; Lee, H.; So, B.; Song, J.-Y.; Kang, H.-G. Chemically induced hepatotoxicity in human stem cell-induced hepatocytes compared with primary hepatocytes and HepG2. Cell biology and toxicology 2016, 32, 403–417. [Google Scholar] [CrossRef]
- Tham, N.T.; Hwang, S.-R.; Bang, J.-H.; Yi, H.; Park, Y.-I.; Kang, S.-J.; Kang, H.-G.; Kim, Y.-S.; Ku, H.-O. High-content analysis of in vitro hepatocyte injury induced by various hepatotoxicants. Journal of Veterinary Science 2019, 20, 34. [Google Scholar] [CrossRef]
- Capdevila, J.; Harris, R.; Falck, J. Microsomal cytochrome P450 and eicosanoid metabolism. Cellular and Molecular Life Sciences 2002, 59, 780–789. [Google Scholar] [CrossRef]
- Nebert, D.W.; Dalton, T.P. The role of cytochrome P450 enzymes in endogenous signalling pathways and environmental carcinogenesis. Nature Reviews Cancer 2006, 6, 947–960. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Human cytochrome P450 enzymes. Cytochrome P450: structure, mechanism, and biochemistry. 2015, 523-785.
- Zhou, S.-F.; Wang, B.; Yang, L.-P.; Liu, J.-P. Structure, function, regulation and polymorphism and the clinical significance of human cytochrome P450 1A2. Drug metabolism reviews 2010, 42, 268–354. [Google Scholar] [CrossRef]
- Wang, B.; Wang, J.; Huang, S.-Q.; Su, H.-H.; Zhou, S.-F. Genetic polymorphism of the human cytochrome P450 2C9 gene and its clinical significance. Current drug metabolism 2009, 10, 781–834. [Google Scholar] [CrossRef]
- Guengerich, F.P. Cytochrome P-450 3A4: regulation and role in drug metabolism. Annual review of pharmacology and toxicology 1999, 39, 1–17. [Google Scholar] [CrossRef]
- Xie, C.; Pogribna, M.; Word, B.; Lyn-Cook Jr, L.; Lyn-Cook, B.D.; Hammons, G.J. In vitro analysis of factors influencing CYP 1A2 expression as potential determinants of interindividual variation. Pharmacology Research & Perspectives 2017, 5, e00299. [Google Scholar]
- Washio, I.; Maeda, M.; Sugiura, C.; Shiga, R.; Yoshida, M.; Nonen, S.; Fujio, Y.; Azuma, J. Cigarette smoke extract induces CYP2B6 through constitutive androstane receptor in hepatocytes. Drug metabolism and disposition 2011, 39, 1–3. [Google Scholar] [CrossRef]
- Horn, J.R.; Hansten, P.D. Get to know an enzyme: CYP1A2. Pharmacy Times 2007, 73, 76. [Google Scholar]
- Tolosa, L.; Gómez-Lechón, M.J.; Pérez-Cataldo, G.; Castell, J.V.; Donato, M.T. HepG2 cells simultaneously expressing five P450 enzymes for the screening of hepatotoxicity: identification of bioactivable drugs and the potential mechanism of toxicity involved. Archives of toxicology 2013, 87, 1115–1127. [Google Scholar] [CrossRef]
- Jover, R.; Bort, R.; Gómez-Lechón, M.J.; Castell, J.V. Re-expression of C/EBPα induces CYP2B6, CYP2C9 and CYP2D6 genes in HepG2 cells. FEBS letters 1998, 431, 227–230. [Google Scholar] [CrossRef]
- Sekretarska, J.; Szczepaniak, J.; Sosnowska, M.; Grodzik, M.; Kutwin, M.; Wierzbicki, M.; Jaworski, S.; Bałaban, J.; Daniluk, K.; Sawosz, E. Influence of selected carbon nanostructures on the CYP2C9 enzyme of the P450 cytochrome. Materials 2019, 12, 4149. [Google Scholar] [CrossRef]
- Negoro, R.; Tasaka, M.; Deguchi, S.; Takayama, K.; Fujita, T. Generation of HepG2 Cells with High Expression of Multiple Drug-Metabolizing Enzymes for Drug Discovery Research Using a PITCh System. Cells 2022, 11, 1677. [Google Scholar] [CrossRef]
- Gerets, H.; Tilmant, K.; Gerin, B.; Chanteux, H.; Depelchin, B.; Dhalluin, S.; Atienzar, F. Characterization of primary human hepatocytes, HepG2 cells, and HepaRG cells at the mRNA level and CYP activity in response to inducers and their predictivity for the detection of human hepatotoxins. Cell biology and toxicology 2012, 28, 69–87. [Google Scholar] [CrossRef]
- Kanebratt, K.P.; Andersson, T.B. HepaRG cells as an in vitro model for evaluation of cytochrome P450 induction in humans. Drug metabolism and disposition 2008, 36, 137–145. [Google Scholar] [CrossRef]
- Ueyama, T.; Tsuji, S.; Sugiyama, T.; Tada, M. Fluorometric evaluation of CYP3A4 expression using improved transgenic HepaRG cells carrying a dual-colour reporter for CYP3A4 and CYP3A7. Scientific Reports 2017, 7, 2874. [Google Scholar] [CrossRef]
- Mandon, M.; Huet, S.; Dubreil, E.; Fessard, V.; Le Hégarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports 2019, 9, 10548. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, Y.; Kawai, K.; Kanaki, T.; Yamazaki, H.; Chesné, C.; Guguen-Guillouzo, C.; Suemizu, H. Functional polymer-dependent 3D culture accelerates the differentiation of HepaRG cells into mature hepatocytes. Hepatology Research 2016, 46, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Ott, L.M.; Ramachandran, K.; Stehno-Bittel, L. An automated multiplexed hepatotoxicity and CYP induction assay using HepaRG cells in 2D and 3D. SLAS DISCOVERY: Advancing Life Sciences R&D 2017, 22, 614–625. [Google Scholar]
Figure 1.
Cytotoxicities of ADR and AAP to HepG2 and HepaRG cells. HepG2 and HepaRG cells treated with ADR (0, 10, 17.8, 31.7, 56.4, 100 μM) and AAP (0, 1, 3, 10, 30 mM) for 24 hr were incubated for 2D and 3D cultures and then used to compare cytotoxicities of ADR and AAP. Cell viability was estimated via CellTiter-Glo® luminescence assay after treatment with serial concentrations of ADR (a) and AAP (b). Values are presented as mean ± SD of three replicates (n = 3). *, p < 0.05; **, p < 0.01 (significantly different from the control).
Figure 1.
Cytotoxicities of ADR and AAP to HepG2 and HepaRG cells. HepG2 and HepaRG cells treated with ADR (0, 10, 17.8, 31.7, 56.4, 100 μM) and AAP (0, 1, 3, 10, 30 mM) for 24 hr were incubated for 2D and 3D cultures and then used to compare cytotoxicities of ADR and AAP. Cell viability was estimated via CellTiter-Glo® luminescence assay after treatment with serial concentrations of ADR (a) and AAP (b). Values are presented as mean ± SD of three replicates (n = 3). *, p < 0.05; **, p < 0.01 (significantly different from the control).
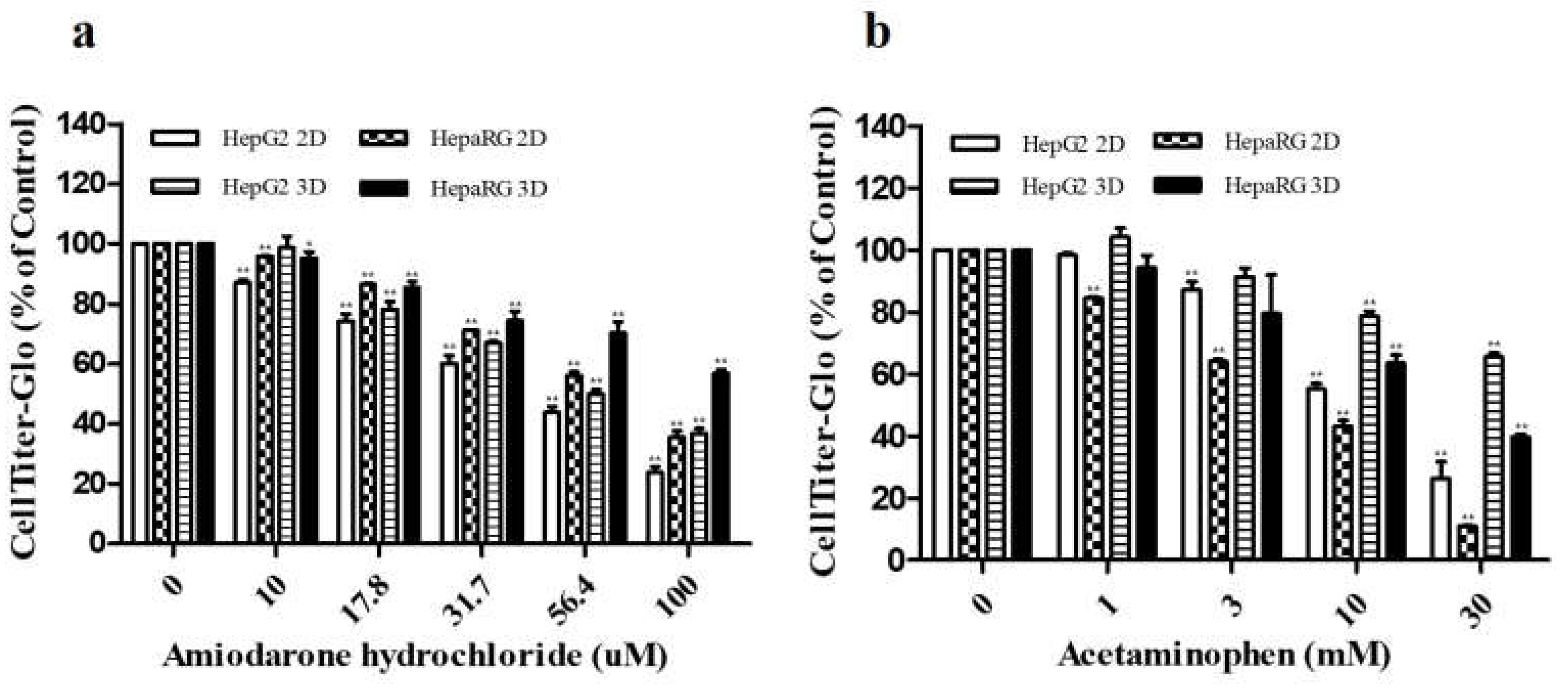
Figure 2.
Confirmation of in vitro drug-induced hepatotoxicity at 24 hr post-treatment. Cellular damage was estimated using LDH activities after treatment with ADR (c) and AAP (d). To compare liver injury markers ALT and AST, HepG2 and HepaRG cells treated with ADR (0, 10, 17.8, 31.7, 56.4, 100 μM) (a) and AAP (0, 1, 3, 10, 30 mM) (b) for 24 hr were cultured in 2D and 3D. Values are presented as mean ± SD of three replicates (n = 3). *, p < 0.05; **, p < 0.01 (significantly different from the control).
Figure 2.
Confirmation of in vitro drug-induced hepatotoxicity at 24 hr post-treatment. Cellular damage was estimated using LDH activities after treatment with ADR (c) and AAP (d). To compare liver injury markers ALT and AST, HepG2 and HepaRG cells treated with ADR (0, 10, 17.8, 31.7, 56.4, 100 μM) (a) and AAP (0, 1, 3, 10, 30 mM) (b) for 24 hr were cultured in 2D and 3D. Values are presented as mean ± SD of three replicates (n = 3). *, p < 0.05; **, p < 0.01 (significantly different from the control).
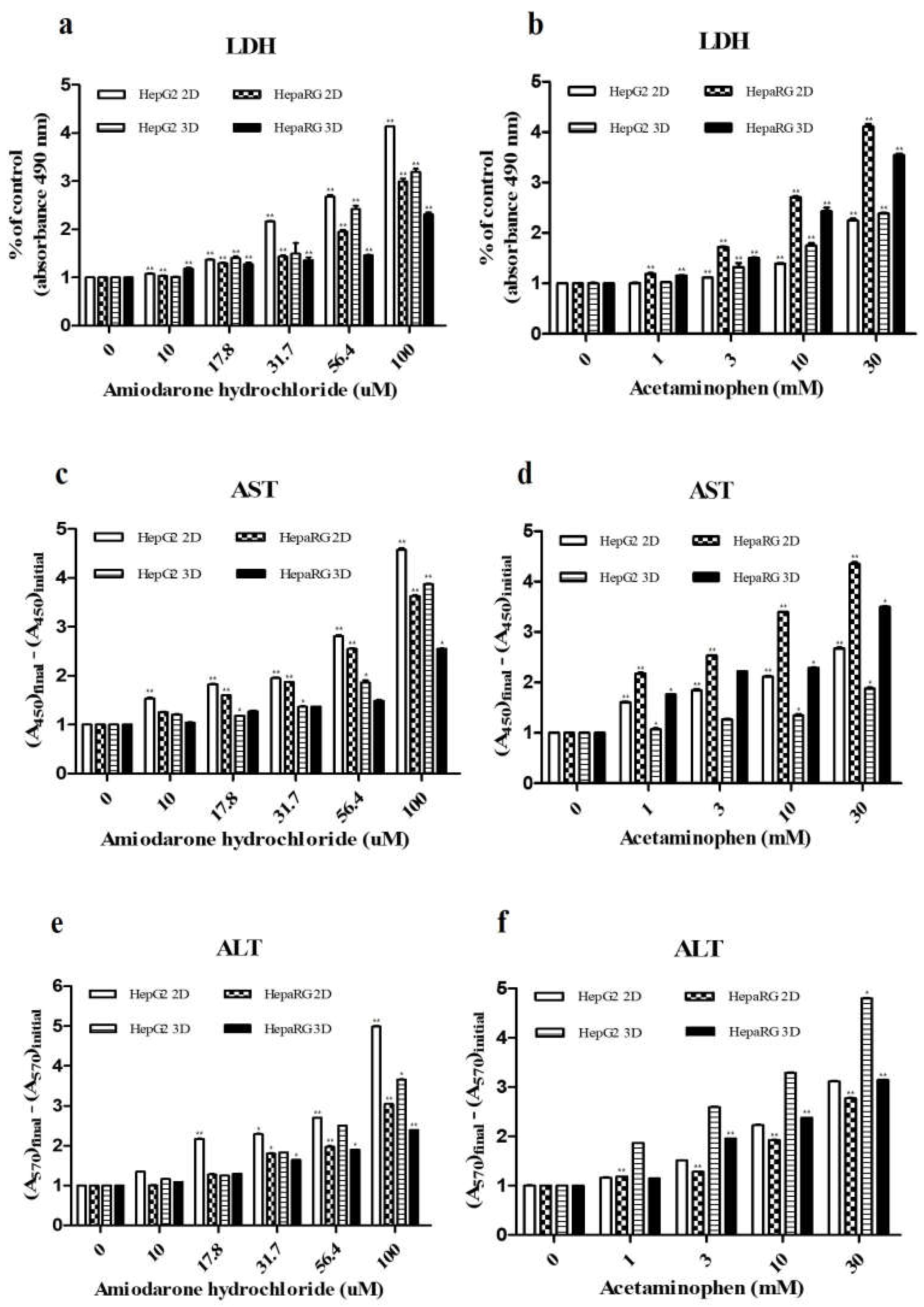
Figure 3.
Effects of ADR and AAP on secretion of ALB and urea. Amounts of ALB and urea after treatment with ADR (0, 10, 17.8, 31.7, 56.4, 100 uM) (a, c) and AAP (0, 1, 3, 10, 30 mM) (b, d) were evaluated. Values are presented as mean ± SD of three replicates (n = 3). *, p < 0.05; **, p < 0.01 (significantly different from the control).
Figure 3.
Effects of ADR and AAP on secretion of ALB and urea. Amounts of ALB and urea after treatment with ADR (0, 10, 17.8, 31.7, 56.4, 100 uM) (a, c) and AAP (0, 1, 3, 10, 30 mM) (b, d) were evaluated. Values are presented as mean ± SD of three replicates (n = 3). *, p < 0.05; **, p < 0.01 (significantly different from the control).
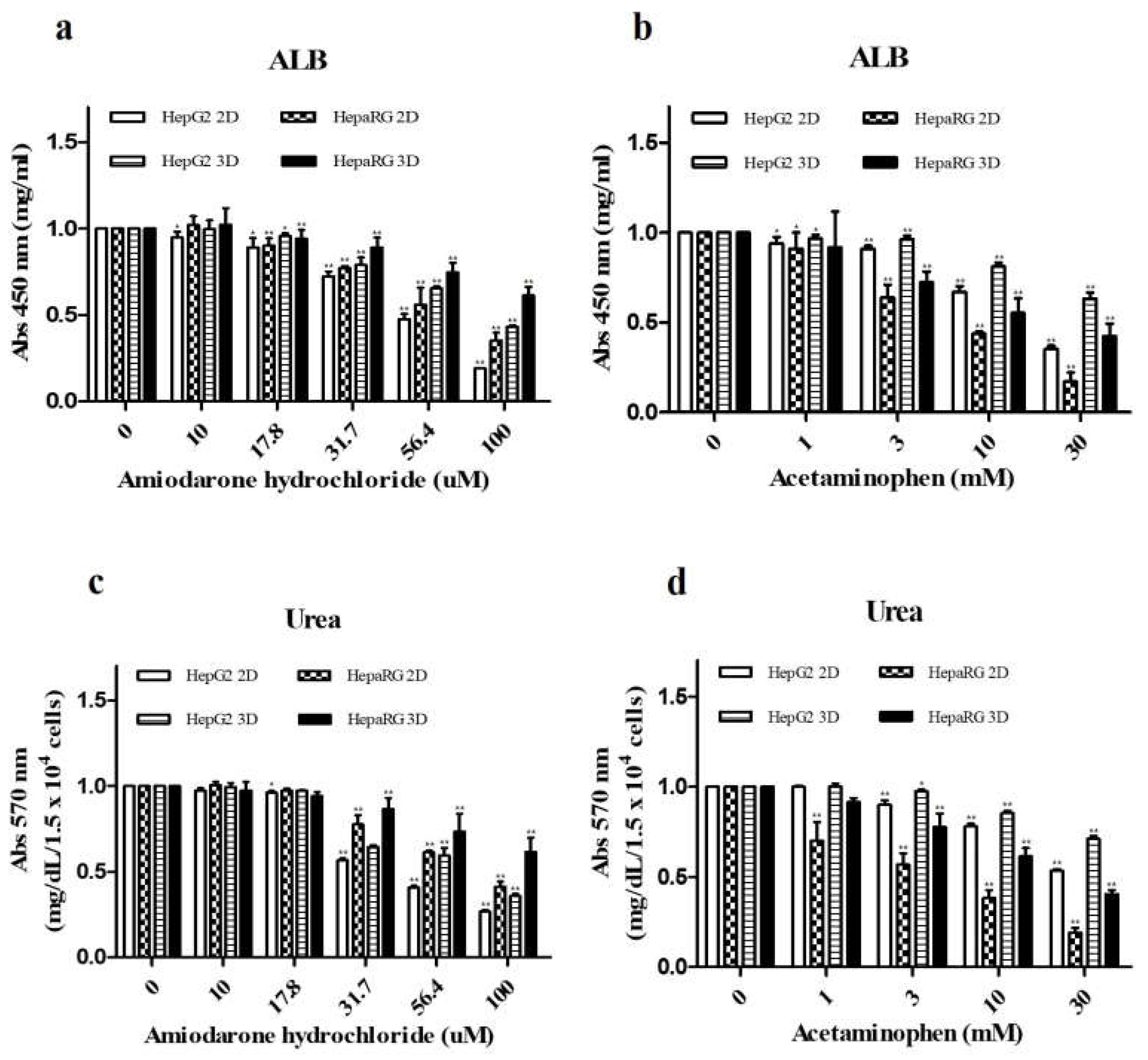
Figure 4.
Comparison of CYP enzyme mRNA expression in HepG2 and HepaRG cells with 2D and 3D cultures. HepG2 and HepaRG cells treated with omeprazole (0, 25, 50, 100 µM) and rifampin (0, 10, 25, 50 µM) for 48 hr were cultured in 2D or 3D type. Expression levels of CYP1A2 (a), CYP2C9 (b), and CYP3A4 (c) mRNAs were evaluated after treatment with omeprazole or rifampin. Concentration-dependent responses such as increases of mRNA levels for each cell types were compared to the untreated group (DMSO) of HepG2 2D control. Values are expressed as mean ± SD of three replicates (n = 3). *, p < 0.05; **, p < 0.01 (significantly different from the control).
Figure 4.
Comparison of CYP enzyme mRNA expression in HepG2 and HepaRG cells with 2D and 3D cultures. HepG2 and HepaRG cells treated with omeprazole (0, 25, 50, 100 µM) and rifampin (0, 10, 25, 50 µM) for 48 hr were cultured in 2D or 3D type. Expression levels of CYP1A2 (a), CYP2C9 (b), and CYP3A4 (c) mRNAs were evaluated after treatment with omeprazole or rifampin. Concentration-dependent responses such as increases of mRNA levels for each cell types were compared to the untreated group (DMSO) of HepG2 2D control. Values are expressed as mean ± SD of three replicates (n = 3). *, p < 0.05; **, p < 0.01 (significantly different from the control).

Table 1.
This is a table. Tables should be placed in the main text near to the first time they are cited.
Table 1.
This is a table. Tables should be placed in the main text near to the first time they are cited.
| CYP enzyme | Inducer | Final Concentration (μM) | Incubation time |
|---|---|---|---|
| CYP1A2 | Omeprazole | 0, 25, 50, 100 | |
| CYP2C9 | Refampin | 0, 10, 25, 50 | 48 hr |
| CYP3A4 | Refampin | 0, 10, 25, 50 |
Table 2.
Primer Sequence.
| Primersets | Sequences | |
| CYP1A2 | Forward | 5‘-CACCATGGCATTGTCCCAGTCTG-3‘ |
| Reverse | 5‘-CCGCGGTCAGTTGATGGAGAAGCGC-3‘ | |
| CYP2C9 | Forward | 5‘-CAGATCTGCAATAATTTTTCTC-3‘ |
| Reverse | 5‘-CTTTCAATAGTAAATTCAGATG-3‘ | |
| CYP3A4 | Forward | 5‘-CTCTCATCCCAGACTTGGCCA-3‘ |
| Reverse | 5‘-ACAGGCTGTTGACCATCATAAAAG-3‘ | |
| β-actin | Forward | 5‘-CAAAGGCGAGGCTCTGTG-3‘ |
| Reverse | 5‘-CCGAAAGTTGCCTTTTATGG-3‘ | |
Table 3.
This is a table. Tables should be placed in the main text near to the first time they are cited.
Table 3.
This is a table. Tables should be placed in the main text near to the first time they are cited.
| Hepatotoxicants | ||
| Celltype | AmiodaroneHClIC50(μM) | AcetaminophenIC50(mM) |
| HepG2 2D | 44.85 | 14.95 |
| HepaRG 2D | 65.47 | 7.11 |
| HepG2 3D | 55.64 | 48.43 |
| HepaRG 3D | 109.38 | 26.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



