
You are currently viewing a beta version of our website. If you spot anything unusual, kindly let us know.
Preprint
Article
Effects of Adding Sphingomonas Z392 to Drinking Water on Growth Performance, Intestinal Histological Structure and Microbial Community of Broiler Chickens
Altmetrics
Downloads
74
Views
34
Comments
0
A peer-reviewed article of this preprint also exists.



This version is not peer-reviewed
Abstract
Probiotics have emerged as a prominent alternative to antibiotics in antimicrobial-free broiler farming. To assess the effect of the probiotics Sphingomonas sp. Z392 isolated and identified by this this laboratory on broiler growth, we conducted a study to examine the changes in broiler body weight, EPI, intestinal histological structure and gut microbiota. The results showed that supplementation of broiler drinking water with 4 × 105 CFU/mL of Sphingomonas resulted in an increase in the relative abundance of Lactobacillus, Bacteroides, Lachnospiraceae, Aminobacterium, Oribacterium, Christensenellaceae, Faecalibacterium, Barnesiella, Ruminococcus, Parabacteroides, Phascolarctobacterium, Butyricicoccaceae, and Caproiciproducens, which were reported to be positively correlated with the digestion and absorption of broiler chickens. The relative abundance was increased of Odoribacter, Alistipes, Parabacteroides, and Rikenellaceae, which were reported to be negatively correlated with the occurrence of intestinal diseases. The relative abundance was decreased of Campylobacter, Shigella Castellani, Bilophila, Campylobacter, Clostridia, and Anaerotruncus, which were reported positively correlated with the occurrence of intestinal diseases. At the same time, it was increased in the integrity of small intestinal villus morphology, the number of goblet cells in small intestinal epithelial cells, the health of mitochondria in the cytoplasm of jejunal villous epithelial cells, and the number of lysosomes in the cytoplasm of goblet cells in small intestinal epithelium, ileal villous epithelial cells, mitochondria in the cytoplasm of large intestinal villous epithelial cells, the VH/CD of the ileum, digestive, absorption, and defense capabilities. Especially the final weight was increased by 4.33% and the EPI increased by 10.10%. Therefore, the supplementation of broiler drinking water with Sphingomonas generated better economic benefits for the broiler chickens.
Keywords:
Subject: Biology and Life Sciences - Animal Science, Veterinary Science and Zoology
1. Introduction
In the past 50 years, the poultry industry has made significant progress through breed improvement, scientific management, and advances in nutritional science, particularly in the use of feed additives, which have made outstanding contributions to the success of current broiler production [1]. With increasing concern for food safety and animal welfare, the selection and application of feed additives have become more refined and scientific [2]. Various additives such as antibiotics, probiotics, oligosaccharides, enzymes, and organic acids are widely used in poultry feed [3,4,5]. Antibiotics, as additives, have been widely favored for their effectiveness in preventing diseases and promoting weight gain [6]. But due to the emergence of drug-resistant strains and the resultant concern related to public health, China has now banned the use of antibiotics in feed additives for chicken farming. This change has driven the emergence of alternative approaches to replace antibiotics. Among them, probiotics are widely used as substitutes for antibiotics due to their unique advantages [7]. They can increase the number of beneficial microorganisms in the intestine, promote host intestinal health, and assist in the digestion and absorption of nutrients by producing hydrolytic enzymes [8]. They can also enhance immunity by regulating the gut microbiota and reduce the colonization of pathogenic bacteria in the intestine, thereby improving the structure of the gut microbiota [6,9]. In broiler farming, microorganisms such as Lactobacillus, Lactobacillus bulgaricus, Lactobacillus acidophilus, Lactobacillus casei, Lactobacillus salivarius, Streptococcus thermophilus, Enterococcus faecalis, Lactobacillus plantarum, fecal coliform, Bifidobacterium, Aspergillus oryzae and brewing yeast lactis have been widely used [6,10,11,12,13,14]. These probiotics not only help improve intestinal health and feed utilization, but also promote the growth and development of broilers and improve meat quality.
Sphingomonas belongs to the class of Proteobacteria α-4 subclasses, Gram negative bacteria. The characteristics of the colony are as follows: yellow, round (diameter 2-3 mm), neat edges, central protrusion, smooth and glossy surface, moist, and sticky texture. The cellular characteristics include short rod-shaped cells, lacking spores, and having unilateral polar flagella.
In our preliminary research, Sphingomonas Z392 was isolated from chicken cecum using a selective culture medium with ammonium nitrogen as the sole nitrogen source. It could use ammonium nitrogen in the intestine as a nitrogen source to convert it into nitrite nitrogen and further convert nitrite nitrogen into nitrate nitrogen, acting as a comammox bacterium [15].
In order to investigate the effect of Sphingomonas Z392 on the growth of broiler chickens, this study explored the role and mechanism of this strain in promoting growth in broiler chickens through changes in body weight, EPI, intestinal histological structure, and gut microbiota structure, providing reference for the application of Sphingomonas Z392 in broiler production.
2. Materials and Methods
2.1. Experimental Design and Animal Breeding Management
600 one-day-old Kebao broiler chickens weighing 42 ± 2 g provided by Henan Longhua Animal Husbandry Co., Ltd. were randomly divided into two groups: control group (CK) and experimental group (T). Each group had 3 replicates, with 100 broiler chickens raised in each replicate. The experimental group of broiler chickens added Z392 4.0 × 105 CFU/mL of Sphingomonas to their drinking water. The feeding period for broiler chickens was 42 days, and the feeding management was carried out in accordance with the "Kebao Broiler Feeding Management Manual", using indoor cage breeding, free feeding and drinking. Drinking water was added 6 times a day, with bacterial agents added according to the amount of water consumed each time.
The use of experimental animals had been approved by the Ethics Committee of Experimental Animals of Huanghuai University and strictly follows the provisions of the "Ethical Review of Experimental Animals of Huanghuai University" (Permit number: 202009220005).
The feeding management of Kebao broilers was divided into three stages: 1-18 day old, 19-33 day old, and 34-42 day old. The nutritional composition of the feed in each stage was detailed in Table 1.
2.2. Weight Changes of Broiler Chickens in Each Group
At the age of 1 day, 5 broiler chickens were randomly selected from all the two groups, and their weight was measured using an electronic scale and recorded.
At the ages of 7 d, 14 d, 21 d, 28 d, 35 d, and 42 d, 5 chickens were randomly selected from each group, and their weight was measured using an electronic scale and recorded.
2.3. European Production Efficiency Factor for Each Group of Broilers
Sold age, the survival rate, sold weight, and feed conversion ratio were calculated, and the European production efficiency factor (EPI) for broiler farming was calculated. EPI = [survival rate × sold weight (kg)] / (feed conversion ratio × sold age) × 10000 [16].
2.4. Observation Methods of Intestinal Microstructure Tissue
After 42 d broiler chickens were euthanized by bleeding, the duodenum, jejunum, ileum, cecum, and rectum were collected in a timely manner. After being fixed with formalin, they were embedded in paraffin, sliced, hematoxylin-eosin (H&E) staining, and sealed. The changes in the intestinal microstructure were observed under a microscope and photographed. Case Viewer 2.4 browsing software was used for analysis and observation.
2.5. Observation Method of Intestinal Ultra Microscopic Structure Tissue
After 42 d, the broiler chickens were euthanized by bleeding, the duodenum, jejunum, ileum, cecum, and rectum were collected in a timely manner. After being fixed with the fixative for electron microscopy, dehydrated at room temperature, resin penetration and embedding, polymerized, thinly sliced, stained, and observed under transmission electron microscopy (TEM) (HT7800, Hitachi Production Co., Ltd) for image analysis.
2.6. Sequencing of Gut Microbiota
After 42 d broiler chickens were euthanized by bleeding, 5 intestinal segments duodenum (du), jejunum (je), ileum (il), cecum (ce), and rectum (re) were collected in a timely and sterile manner, each with a length of 3 cm. After ligation at both ends, they were sent to Wuhan Servicebio Technology Co., Ltd. for testing. Based on the Illumina platform, the genome of the gut microbiota was sequenced and bioinformatics analysis was performed using a two end sequencing (PE250) method. The gut microbiota of broiler chickens was analyzed and determined using 16s rDNA sequencing technology.
2.7. Methods of Data Statistical Analysis
The experimental data were sorted out by Microsoft Excel 2013, and the mean and standard deviation were calculated, and draw line and bar charts. SPSS 14.0 software was used for analysis of variance.
3. Results
3.1. Changes in Body Weight of Broiler Chicken in Each Group
Take the average weight of broiler chickens aged 1, 7, 14, 21, 28, 35, and 42 days in each group, and create a graph of broiler weight (g) - day (d) (Figure 1). The average weight in test (T) group at selected time points was significantly higher than that in control (CK) group (p<0.05).
Note: Different letters at the end of the line chart indicate significant differences between the two groups of data (p<0.05).
3.2. Comparison of EPI between Test and Control Groups of Broilers
During the experiment, the EPI of broilers in each group were detailed in Table 2. Both groups of broiler chickens have European indices above 300, achieving profitability. Briefly, addition of 4.0 × 105 CFU/mL of Sphingomonas Z392 to drinking water can increase the final weight by 4.33% and the EPI by 10.10%, resulting in better economic benefits for broilers.
3.3. Changes in Intestinal Microstructure
After paraffin sectioning and H&E staining of various intestinal tissues, the results were observed under a binocular biological microscope, as shown in Figure S1-S3. In the experimental group, the integrity of the small intestinal villi enhanced, and the number of goblet cells in the epithelial cells of the small intestine increased. However, in the control group, the small intestinal villi of the broiler chickens became shorter and fragmented, and the villi did not show their inherent morphology, especially the duodenum and jejunum, which were most severely damaged. There were no visible differences in the folds and villi of the cecum and rectum in each group of broiler chickens.
3.4. Changes in the Ratio of Small intestinal Intestinal Villus Height to Crypt Depth
After measuring the villus height (VH) and crypt depth (CD) of the small intestine using Case Viewer 2.4 browsing software, the ratio of villus height to crypt depth (VH/CD) of each segment of the small intestine was calculated, as shown in Figure 3. There was no significant difference in VH/CD between the duodenum and jejunum groups (p>0.05). The VH/CD of the ileum in the experimental group (6.62 ± 0.06) was significantly higher than that in the control group (3.25 ± 0.55) (p<0.05).
3.5. Changes in the Ultramicroscopic Structure of the Intestine
After preparing ultra-thin sections from different intestinal segments and staining, they were observed under transmission electron microscopy, as shown in Figure S4-S8. In the test group, addition of 4 × 105 CFU/mL of Sphingomonas Z392 into the drinking water of broiler chickens reduced the degree of mitochondrial swelling in the cytoplasm of jejunal villous epithelial cells and increased the number of goblet cells in the small intestine epithelium, lysosomes in villous epithelial cells in the ileum, and mitochondria in villous epithelial cells in the large intestine.
3.616. S rRNA Gene Sequence Statistics
After Illumina NovaSeq sequencing, the 16S rRNA gene sequence data from fecal samples of the duodenum, jejunum, ileum, cecum, and rectum from the two groups of chickens were presented in Table 3. Following quality control, the removal of chimeric sequences and those out of the target range, the average number of 16S rRNA gene sequences retained for analysis in each intestinal segment of the CK group was 62,608 in the duodenum, 69890 in the jejunum, 66047 in the ileum, 60924 in the cecum, and 69109 in the rectum. The average number of 16S rRNA gene sequences retained for analysis in each intestinal segment of the T group was 52233 in the duodenum, 55390 in the jejunum, 62781 in the ileum, 67302 in the cecum, and 53054 in the rectum.
3.7. Alpha Diversity Analysis
From Figure 5 and Table 4, it can be observed that, at a similarity level of 97%, the bioinformatics analysis of OTU showed no significant difference in the Simpson index and Shannon index between the T and the CK groups (p>0.05). This finding suggests a high degree of consistency in the diversity of gut microbiota between the CK and T groups.
3.8. NMDS Non Metric Multidimensional Scale Analysis
By using NMDS non metric multidimensional scale analysis on the bacterial communities at the genus level in 5 different parts, the results were shown in Figures 4-10. There is a significant overlap between the microbial communities of the CK and T groups in the duodenum, indicating that the differences in microbial structure and composition between the CK and T groups in the duodenum are relatively small. However, the microbiota of the CK and T groups in the jejunum, ileum, cecum, and rectum did not cross indicating significant differences in the gut microbiota structure and composition between the CK and T groups in the jejunum, ileum, cecum, and rectum.
Figure 6.
Non metric multidimensional scale analysis of NMDS in different intestines of different groups of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Figure 6.
Non metric multidimensional scale analysis of NMDS in different intestines of different groups of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
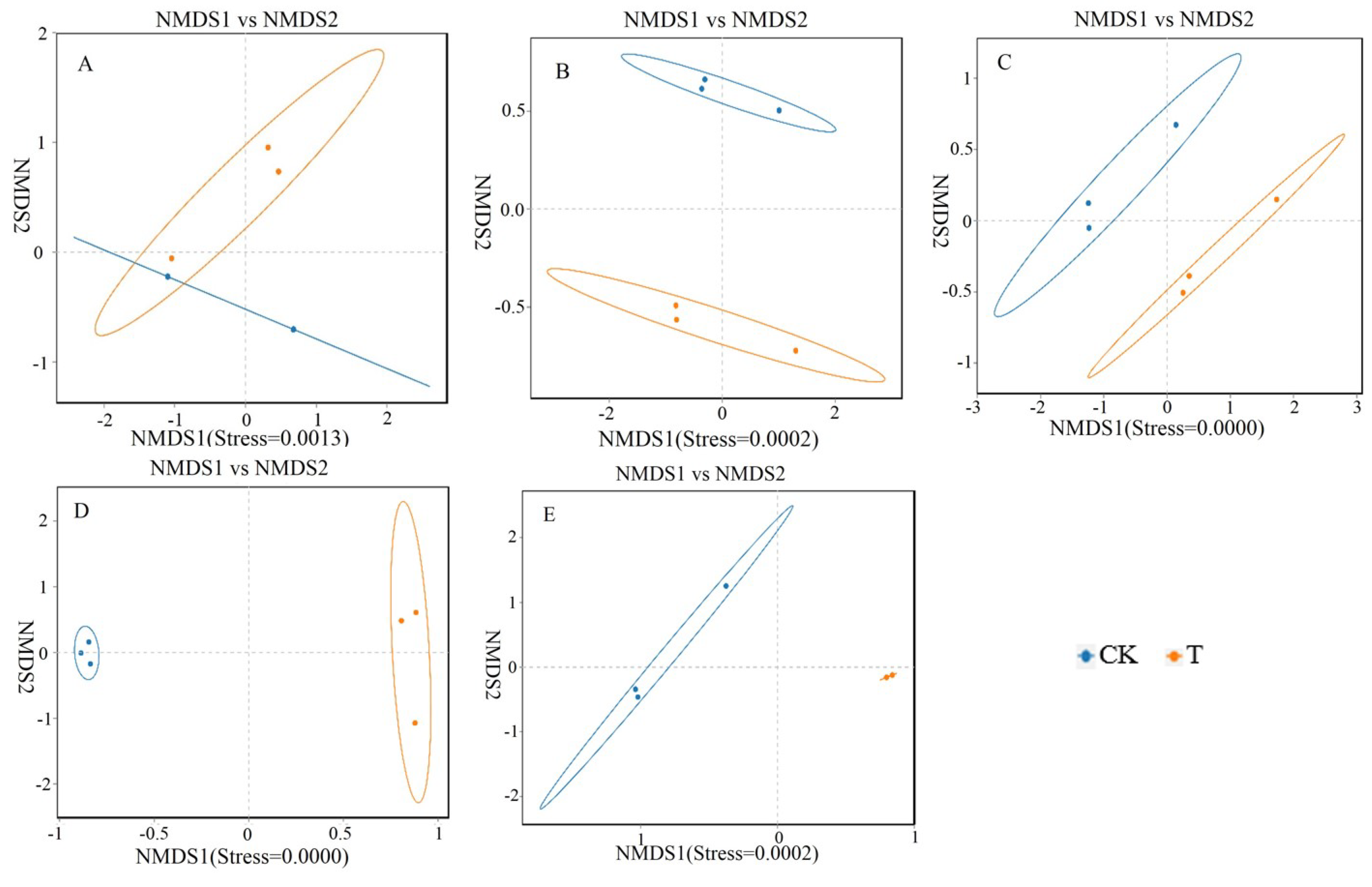
Note: 1 A, B, C, D, and E are the results of non metric multidimensional scale analysis of the duodenum, empty field, ileum, cecum, and rectum, respectively.
2. The appearance of obvious cross indicates that there is no significant difference in microbial structure and composition between groups, while the absence of cross indicates significant differences in microbial structure and composition between groups.
3. Each point in the figure represents a sample; Different colors represent different groups; The elliptical circle represents a 95% confidence ellipse (i.e., if there are 100 samples in the sample group, 95 will fall within it). When the stress is less than 0.1, it can be considered a good sorting; When the stress is less than 0.05, it has good representativeness. It is generally believed that when the Stress is less than 0.2, it indicates that NMDS analysis has a certain level of reliability. The closer the sample is on the coordinate map, the higher the similarity.
3.9. Analysis of Microbial Diversity at Phylum Level
The classification results of gut microbiota in various intestinal sites across different experimental groups are presented in Table 5 and Figure 7. At the phylum level, a total of 47 phyla are involved in the gut microbiota of the five parts. To facilitate the observation of relationships and species composition among samples, a bar chart of species distribution was constructed, based on the abundance of each species within each sample. At the phylum level, there are 10 phyla with higher relative abundance, including Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria, Acidobacteriota, Chloroflexi, Cyanobacteria, Desulfobacteria, Gemmatimonadota, and unclassified phyla.

Table 5.
Classification and statistical results of microbial flora in different intestinal segments based on various classification levels of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 5.
Classification and statistical results of microbial flora in different intestinal segments based on various classification levels of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Samples | Kindom | Phylum | Class | Order | Family | Genus | Species | |
| CK | du | 2±0 | 27±2.52 | 50±14.01 | 107±35 | 177±73.93 | 254±126.89 | 276±143.62 |
| je | 2±0 | 21±3.21 | 35±11.02 | 71±25.51 | 109±47.18 | 149±71.77 | 160±72.92 | |
| il | 2±0.58 | 28±6.08 | 55±15.59 | 127±49.39 | 226±95.31 | 381±177.79 | 417±187.16 | |
| ce | 1±0.58 | 13±5.77 | 22±10.97 | 45±20.5 | 83±38.73 | 149±55.18 | 178±55.07 | |
| re | 2±0.58 | 25±6.66 | 51±20.21 | 115±43.04 | 195±75.92 | 306±115.77 | 345±125.3 | |
| T | du | 2±0 | 25±3.21 | 50±11.24 | 109±32.08 | 170±69.46 | 252±129.69 | 269±140.88 |
| je | 2±0 | 27±3.79 | 51±5.51 | 107±13.05 | 169±33.08 | 248±67.1 | 264±74.57 | |
| il | 2±0 | 24±2.08 | 50±5.29 | 119±10.79 | 200±10.26 | 310±22.5 | 348±25.42 | |
| ce | 2±0 | 17±5 | 29±11.14 | 63±25.79 | 103±30.12 | 175±43.27 | 209±44.6 | |
| re | 1±0 | 12±6.08 | 20±10.97 | 46±19.35 | 80±27.73 | 143±38.4 | 173±41.88 | |
Note: The numbers in the table represents the total number of Reads covered by the sample at that level.
Figure 7.
The content of microbial communities in different intestinal segments of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water at the phylum level.
Figure 7.
The content of microbial communities in different intestinal segments of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water at the phylum level.
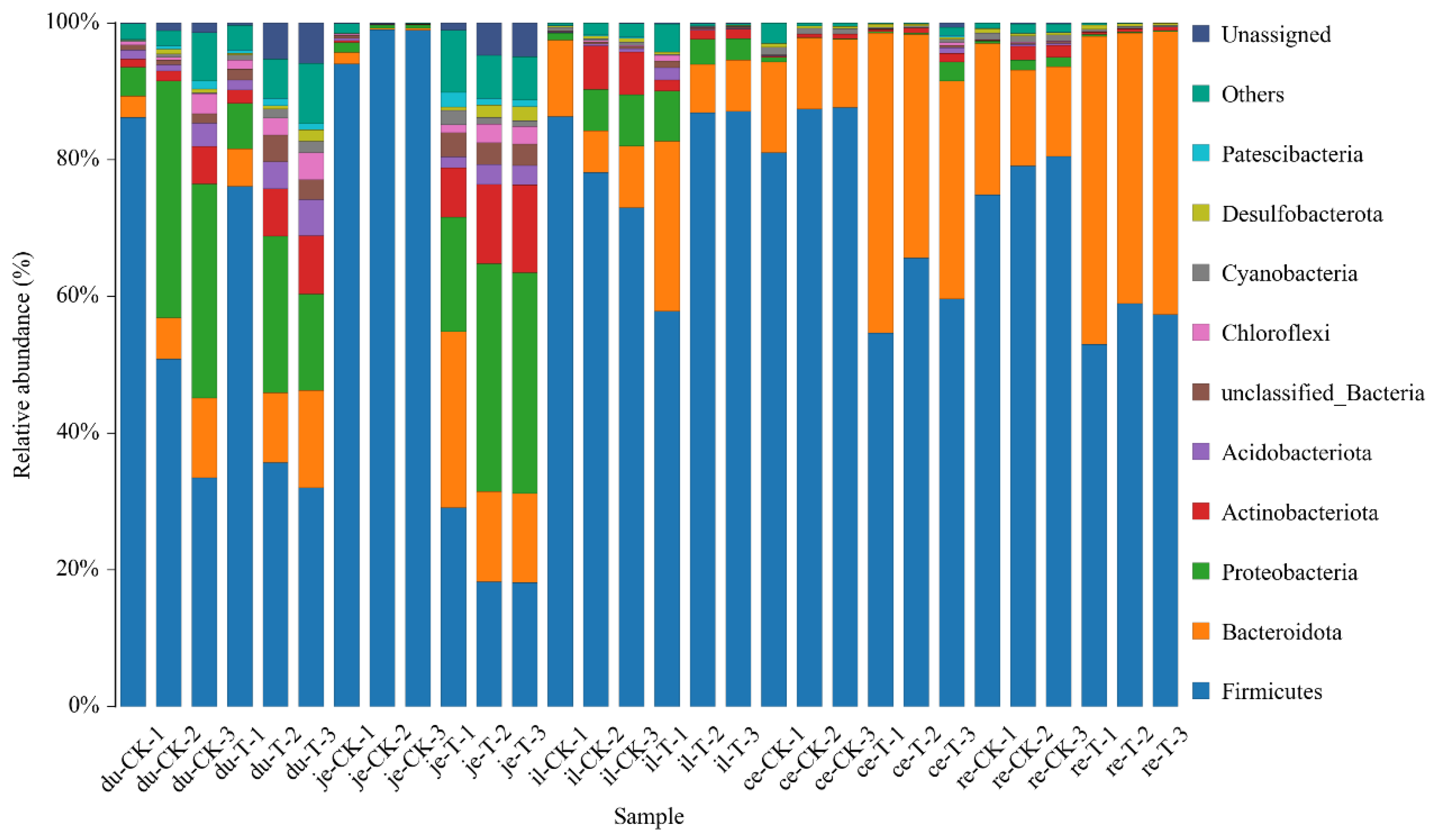
Note: The horizontal axis represents the sample name; The vertical axis represents the relative abundance percentage (%). Different colors represent different species; Stacked columns represent the top 10 taxonomic groups with relative abundance at each taxonomic level.
3.10. Diversity Analysis of Microbial Communities at the Genus Level
To facilitate the observation of relationships between the samples and composition of microbial species, a figure depicting the distribution of microbial species for each sample was constructed, based on the abundance of each microbial species within each sample (Figure 8). There were 10 genera with higher relative abundance, including Alistipes, Bacteroides, Christensenellaceae R_7_group, Faecalibacterium, Lactobacillus, Limosillactobacillus, Ruminococcus_torques_group, unclassified Lachnospiraceae, unclassified Oscillospiraceae, and others, all of which exhibit intergroup differences.
Note: The horizontal axis represents the sample name; The vertical axis represents the relative abundance percentage (%). Different colors represent different species; Stacked columns represent the top 10 taxonomic groups with relative abundance at each taxonomic level.
3.10.1. Differential Analysis of Duodenal Microbiota at the Genus Level
The content of bacterial strains in the duodenum and the changes in different groups are shown in Table 6. Compared with the CK group, in the T group, the relative abundance were increased of Actobacillus, Bacteroides, Lachnospiraceae, and Aminobacterium, which were positively correlated with digestion and absorption. On the other hand, the relative abundance were increased of Odoribacter, which was negatively correlated with the occurrence of intestinal diseases.
3.10.2. Differential Analysis on Jejunum Microbiota at the Genus Level
The content of bacterial strains in the jejunum and the changes in different groups are shown in Table 7. Compared with the CK group, in the T group, the relative abundance were increased of Bacteroides, Lachnospiraceae, Oribacterium, Christensenellaceae, Proteiniphilum, Faecalibacterium, Barnesiella, Ruminococcus, Phascolarctobacterium, and Butyricicoccaceae, which were positively correlated with digestion and absorption. The relative abundance was increased of Alistipes and Paraacteroides distasonis, which was negatively correlated with the occurrence of intestinal diseases. But the abundance of Staphylococcus in the jejunum was also significantly increased.
3.10.3. Differential analysis on ileum microbiota at the genus level
The content of bacterial strains in the ileum and the changes in different groups are shown in Table 8. Compared to the CK group, the T group exhibited an increase in the relative abundance of Bacteroides, Aminobacterium, and Proteiniphilum, which were positively correlated with digestion and absorption; the relative abundance was increased of Parabacteroides, which was negatively correlated with the occurrence of intestinal diseases; the relative abundance was reduced of Campylobacter, Escherichia, and Bilophila, which was positively correlated with the occurrence of intestinal diseases.
3.10.4. Differential Analysis of Cecal Microbiota at the Genus Level
The content of bacterial strains in the cecal and the changes in different groups are shown in Table 9. Compared with the CK group, in the T group, the relative abundance were increased of Barnesiella, Oribacterium, Christensenellaceae, and Caproiciproducens, which were positively correlated with digestion and absorption; the relative abundance was increased of Rikenellaceae, which was negatively correlated with the occurrence of intestinal diseases; the relative abundance was reduced of Clostridia, Anaerotruncus, and Helicobacter, which was positively correlated with the occurrence of intestinal diseases.
3.10.5. Differential Analysis on Rectal Microbiota at the Genus Level
The content of bacterial strains in the rectal and the change folds in different groups are shown in Table 10. Compared with the CK group, in the T group, the relative abundance was increased of Bacteroides, Oribacterium, and Ruminococcus, which were positively correlated with digestion and absorption; the relative abundance was increased of Rikenellaceae, which was negatively correlated with the occurrence of intestinal diseases; the relative abundance was reduced of Clostridia, which was positively correlated with the occurrence of intestinal diseases.
4. Discussion
4.1. Effects of Sphingomonas on the Histological Structure of Intestine
The gastrointestinal tract (GIT) serves as the primary site for digestion and absorption, and defense against harmful foreign microorganisms. Its function correlates positively with organelle numbers and health status. The villi in the small intestine play a crucial role in nutrient absorption, and the height, integrity, and cell count of these villi significantly affect small intestine digestive and absorption functions [17]. After adding 4 × 105 CFU/mL of Sphingomonas Z392 to drinking water, broiler chickens exhibited increased integrity of small intestinal villi, an augmented number of goblet cells in small intestinal epithelial cells, enhanced mitochondrial health in the cytoplasm of jejunal villous epithelial cells, and elevated counts of lysosomes in the cytoplasm of goblet cells in the small intestinal epithelium, ileal villous epithelial cells, and mitochondria in the cytoplasm of large intestinal villous epithelial cells. The VH/CD of the ileum significantly increased (p<0.05).
VH, CD, and VH/CD serve as vital indicators for measuring intestinal nutrient digestion and absorption abilities, as well as its health status [18]. The villus's edges secrete various digestive enzymes; thus, higher VH indicates stronger enzyme secretion capacity and better digestion and absorption abilities. CD reflects cell generation rates, with shallower crypts indicating increased cell maturation rates and enhanced intestinal secretion function. A higher VH/CD value signifies stronger intestinal digestive and absorption capacities, while a decrease indicates weakened abilities [19].
Previous research indicates that adding probiotics to the diet can protect the gastrointestinal tract from excessive feeding, increase VH and VH/CD in broiler chickens, and reduce CD [20]. The results of this experiment demonstrate that adding Sphingomonas to broiler chickens' drinking water significantly increased the VH/CD ratio of the ileum and enhanced its digestive, absorption, and defense abilities. Moreover, the addition of Sphingomonas Z392 to drinking water led to a notable increase in intestinal probiotics such as Lactobacillus, which would activate cell mitosis and induce intestinal epithelial cell proliferation by secreting short-chain fatty acids (SCFAs), resulting in increased intestinal villus length [21,22].
4.2. Effects of Sphingomonas on Gut Microbiota
Studying the relative abundance and diversity of the gut microbiota is crucial for understanding the impact of beneficial bacteria [23]. In this experiment, the Shannon curve's horizontal width for multiple samples in each group was relatively large, indicating high species abundance in the samples. Additionally, the curve appeared relatively flat, suggesting a relatively uniform distribution of species across each group of samples. The coverage index for each group was 1, indicating that the different intestinal sequencing results in this experiment accurately reflected the true microbial composition of the samples.
The degree of intestinal health is determined by indicators such as microbial content and morphology, with the distribution of microorganisms varying among different intestinal segments [24]. The gut microbiota plays a crucial role in maintaining animal health, immunity, and production performance, and probiotics have been shown to improve the microecological environment of broiler intestines [23]. The relative research previously indicated that adding Bacillus amyloliquefaciens to the diet can increase the relative abundance of Bacteroidetes, Butyricicoccaceae, Faecalibacterium, Heliobacillus, Lactobacillus, Parabacteroides, and Ruminococcus in the gut of broiler chickens [25].
This experiment found that adding 4 × 105 CFU/mL Sphingomonas Z392 to the drinking water of broiler chickens increased the relative abundance of microorganisms, including actobacillus in the duodenum; Bacteroides in the duodenum, jejunum, ileum, and rectum; Lachnospiraceae in the duodenum and jejunum; Aminobacterium in the duodenum and ileum; Oribacterium in the jejunum, cecum, and rectum; Christensenellaceae of bacteria in the jejunum and cecum; Proteiniphilum in the jejunum and ileum; Faecalibacterium in the jejunum; Barnesiella in the jejunum and cecum; Ruminococcus in the jejunum and rectum; Phascolarctobacterium in the jejunum; Butyricicoccaceae in the jejunum; Caproiciproducens in the cecum. The above microorganisms could promote the digestion and absorption ability of broiler chickens. Furthermore, the microorganisms with reduced relative abundance include Odoribacter in the duodenum, Alistipes in the jejunum, Parabacteroides in the jejunum, ileum, and rectum, and Rikenellaceae in the cecum and rectum. These microorganisms could inhibit the occurrence of intestinal diseases [26,27].
The microorganisms Actobacillus, Lachnospiraceae, Oribacterium, Christensenellaceae, Faecalibacterium, Ruminococcus, and Caproiciproducens belong to Firmicutes, which can hydrolyze starch and other sugars to produce butyrate and other secretes short-chain fatty acids (SCFAs), such as acetate, propionate, butyrate, and lactate [28,29]. SCFAs play multiple roles in the intestine. They not only provide energy to the body, but also reduce the types of harmful bacteria, stimulate the proliferation and differentiation of intestinal epithelial villous cells, increase the height of villi, expand the contact area between villi and chyme, and improve the digestion and absorption ability of broilers [29]. In addition, the increase of beneficial microorganisms such as Actobacillus in the intestine also helps to maintain the integrity of the intestinal structure, promote metabolism, and thereby plays an important protective role as the first line of defense against pathogenic bacteria [30].
In addition, Alistipes, Odoribacter, Rikenellaceae, Bacteroides, and Parabacteroides belong to Bacteroidetes. They primarily digest grain feed and secrete mucin to protect and lubricate the intestines, exerting an anti-inflammatory effect [31,32,33]. Bacteroidetes can also enhance the disease resistance of broiler chickens by stimulating the immune system, increasing macrophage phagocytosis, and resisting the colonization of pathogenic bacteria [32]. The lithocholic acid secreted by Odoribacter promotes fat digestion and absorption, regulates blood lipids, promotes cell proliferation, exhibits anti-inflammatory properties, and protects the gastric mucosa. Additionally, the lithocholic acid produced by Odoribacter demonstrates excellent anti-inflammatory and antibacterial activities. Even in small amounts, it effectively eliminates pathogenic microorganisms such as Clostridium and Enterococcus faecalis, reduces inflammation levels, and regulates the body's immunity [34]. Bacteroides, a core member of the gut microbiota, regulates the host mucosal immune system, reduces inflammation, participates in carbon metabolism, and secretes SCFAs such as acetate and propionate [35]. Rikenellaceae in the intestine plays a certain role in protecting the body's health and alleviating diseases [36].
Aminobacterium Bacteroides, a core member of the gut microbiota, regulates the host mucosal immune system, reduces inflammation, participates in carbon metabolism, and secretes short-chain fatty acids (SCFAs) such as acetate and propionate [37]. Phascolarctobacterium produces SCFAs, including acetate and propionate, reduces inflammation, and protects the intestinal mucosal barrier by decreasing levels of lipopolysaccharide (LPS)-binding proteins and C-reactive protein (CRP) [38]. Barnesiella is associated with bile acid production and plays a crucial role in fat metabolism [39]. Proteiniphilum contributes to the conversion of acetic acid to butyric acid [40]. Butyricicoccaceae produces digestive enzymes in the intestine that break down starch, protein, and cellulose, promoting the digestion and absorption of these nutrients. Furthermore, butyric acid, the main metabolite of this genus, promotes the regeneration and repair of intestinal epithelial cells. Therefore, Butyricicoccaceae significantly influences the intestinal microbiota's structure, inhibits pathogenic bacteria, and promotes the growth of beneficial bacteria such as Lactobacillus [41].
Probiotics not only enhance nutrient digestion and absorption, improve the breeding environment, but also reduce pathogenic microorganisms and increase the relative abundance of anti-inflammatory microorganisms, thereby enhancing the body's resistance to infections [25]. This study revealed that adding Sphingomonas Z392 to the drinking water of broiler chickens decreased the relative abundance of microorganisms such as Campylobacter in the jejunum and ileum, Shigella Castellani in the ileum, Bilophila in the ileum, Clostridia in the cecum and rectum, and Anaerotruncus in the cecum. These microorganisms are positively correlated with intestinal diseases [26,27].
But after adding Sphingomonas to the drinking water of broiler chickens, the relative abundance of Staphylococcus was increased in the jejunum, which can easily cause gastrointestinal diseases. Therefore, when using Sphingomonas, it is necessary to use other microorganisms in combination to inhibit the proliferation of potential harmful bacteria, ensuring the health and breeding benefits of broilers.
4.3. Growth Promotion of Sphingomonas spp.
In this study, the relative abundance of microorganisms positively correlated with digestion and absorption and negatively correlated with intestinal diseases in broiler chickens was significantly increased by adding Sphingomonas spp. to drinking water during the feeding process. These microorganisms promote the digestion and absorption of nutrients in chyme by secreting digestive enzymes such as amylase, protease, glycosidase, and cellulase, or produce SCFAs, which reduce the types of harmful bacteria and stimulate the proliferation and differentiation of intestinal epithelial villous cells. This results in a high increase in villi, expanding the contact area between villi and chyme, and improving the digestion and absorption ability of broiler chickens [29]. Alternatively, secreted bile acids can promote fat digestion and absorption, regulate blood lipids, promote cell proliferation, exhibit anti-inflammatory properties, and protect the gastric mucosa. Meanwhile, the addition of these strains significantly reduced the degree of inflammation in the small intestine, ensuring its basic digestive and absorption functions, and promoting the digestion of nutrients such as sugars, proteins, fats, and inorganic salts. Additionally, the number of goblet cells in the small intestine villous epithelium increases, playing an important role in maintaining intestinal health and promoting nutrient absorption. At the same time, the degree of mitochondrial swelling in the villous epithelial cells of the jejunum decreases, indicating a decrease in their degree of damage, thereby ensuring the digestive and absorption functions of the jejunum. Importantly, the VH/CD of the ileum significantly increased (p<0.05), and the number of lysosomes in the villous epithelial cells increased. This significantly improved the digestive and decomposition ability of the ileum, facilitating further absorption of nutrients in the ileum. Lactobacillus and Clostridium butyricum also significantly affect the growth, development, and feed utilization of broiler chickens by regulating small intestinal VH and CD [6,42,43].
Additionally, due to the relatively short intestine of chickens, the digestion of chyme in the small intestine is not complete, so the cecum plays an important role in the chicken's digestive system [44]. This study found that the number of mitochondria in cecal villous epithelial cells increased under the action of Sphingomonas, which promoted the absorption of nutrients by the cecum.
After adding 4.0 × 105 CFU/mL of Sphingomonas Z392 to the drinking water of broiler chickens, the improvement of intestinal histology and microbial community structure not only enhanced the absorption function of the overall digestive tract of broiler chickens, but also improved the utilization rate of feed and the growth performance of broiler chickens, resulting in an increase of 4.33% in slaughter weight and 10.10% in the EPI, resulting in better economic benefits for broiler chickens.
In practical applications, adding an appropriate amount of Sphingomonas Z392 to the drinking water of broiler chickens could significantly increase their final weight, increase the EPI, and thus bring significant economic benefits. These results indicate that Sphingomonas Z392 has broad application prospects in broiler feeding and is expected to provide new strategies for the sustainable development of the broiler industry.
5. Conclusions
Addition of 4 × 105 CFU/mL of Sphingomonas sp. Z392 to drinking water improved intestinal histology and flora structure, the function of the digestion and enhanced absorption chickens and feed utilization and resulted in a 4.33% increase in the final weight of broilers and a 10.10% increase in the EPI, which led to better economic benefits for broilers.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
W. M. C. and G. A. Z. designed the whole and wrote the final manuscript. Z. J. and Z. S. Q. carried out all experiments, data collection, and manuscript editing. W. G. L., X. H. L., G. Y. N. and L. C. Y. participated in data analysis.
Funding
This work was supported by Key R & D projects of Ningxia Hui Autonomous Region (2021BEE02032), the Science and Technology Development Plan Project of Henan province of China (172102110201 and 212102110005), the Science and Technology Innovation Demonstration Project for High Quality Agricultural Development and Ecological Protection of the Ningxia Academy of Agriculture and Forestry Sciences (NGSB-2021-12-05).
Institutional Review Board Statement
The use of experimental animals had been approved by the Ethics Committee of Experimental Animals of Huanghuai University and strictly follows the provisions of the "Ethical Review of Experimental Animals of Huanghuai University" (Permit number: 202009220005).
Data Availability Statement
1. Experimental results of intestinal ultra microstructure
Extract code: dueg
2. Experimental results of 16S rRNA sequencing of gut microbiota
Extraction code: q0u
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gado, A. R.; Ellakany, H. F.; Elbestawy, A. R.; El-Hack, M. E. A.; Mahgoub, S. A.; Herbal Medicine Additives as Powerful Agents to Control and Prevent Avian Influenza Virus in Poultry – A Review. Ann. Anim. Sci. 2019, 19(4): 905-935. [CrossRef]
- Farag, M. R.; Alagawany. M. The role of Astragalus membranaceus as immunomodulator in poultry. World's Poultry Science Journal. 2019, 75: 1-12. [CrossRef]
- Bin-Jumah, M.; El-Hack, M. E. A.; Abdelnour, S. A.; Hendy, Y. A.; Aleya, L. Potential use of chromium to combat thermal stress in animals: A review. Science of The Total Environment. 2019, 707: 135996. [CrossRef]
- Elgeddawy, S. A.; Shaheen, H. M.; El-Sayed, Y. S.; Elaziz, M. A.; Darwish, A.; Samak, D.; Batiha, G. E.; Mady, R. A.; Bin-Jumah, M.; Allam, A. A. Effects of the dietary inclusion of a probiotic or prebiotic on florfenicol pharmacokinetic profile in broiler chicken. Journal of animal physiology and animal nutrition. 2020, 104(2): 549-557. [CrossRef]
- Hussein, E. O. S.; Suliman, G. M.; Alowaimer, A. N.; Ahmed, S. H.; Swelum, A. A. Growth, carcass characteristics, and meat quality of broilers fed a low-energy diet supplemented with a multienzyme preparation. Poultry Science. 2020, 99(4): 1988-1994. [CrossRef]
- Abd El-Hack, M. E.; El-Saadony, M. T.; Shafi, M. E.; Qattan, S. Y. A.; Batiha, G. E.; Khafaga, A. F.; Abdel-Moneim, A. E.; Alagawany, M. Probiotics in poultry feed: A comprehensive review. J. Anim. Physiol. Anim. Nutr (Berl). 2020, 104: 1835-1850. [CrossRef]
- Nuria, V. S.; Yanath, B.; Ruth, R.; Eric, A.; Djamel, D. Benefits and Inputs From Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters During Food-Animal Production. Frontiers in microbiology. 2019, 10: 57. [CrossRef]
- M'sadeq, S. A.; Wu, S.; Swick, R. A.; Choct, M. Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide. Animal Nutrition. 2015: 1(1): 1-11. [CrossRef]
- Wu, Y.; Wang, B.; Zeng, Z.; Liu, R.; Tang, L.; Gong, L.; Li, W. Effects of probiotics Lactobacillus plantarum 16 and Paenibacillus polymyxa 10 on intestinal barrier function, antioxidative capacity, apoptosis, immune response, and biochemical parameters in broilers. Poult. Sci. 2019, 98: 5028-5039. [CrossRef]
- Hossain, M.; Begum, M.; Kim, I. Effect of Bacillus subtilis, Clostridium butyricum and Lactobacillus acidophilus endospores on growth performance, nutrient digestibility, meat quality, relative organ weight, microbial shedding and excreta noxious gas emission in broilers. Veterinární Medicína. 2015, 60(2): 77-86. [CrossRef]
- Liao, X. D.; Ma, G.; Cai, J.; Fu, Y.; Yan, X. Y.; Wei, X. B.; Zhang, R. J. Effects of Clostridium butyricum on growth performance, antioxidation, and immune function of broilers. Poultry Science. 2015, 94: 662-667. [CrossRef]
- Mi, J.; Chen, X.; Liao, X. Screening of single or combined administration of 9 probiotics to reduce ammonia emissions from laying hens. Poultry Science. 2019, 98: 3977-3988. [CrossRef]
- Shehata, A. A.; Tarabees, R.; Basiouni, S.; ElSayed, M. S.; Gaballah, A.; Krueger, M. Effect of a Potential Probiotic Candidate Enterococcus faecalis-1 on Growth Performance, Intestinal Microbiota, and Immune Response of Commercial Broiler Chickens. Probiotics and Antimicrobial Proteins. 2020, 12(2): 451-460. [CrossRef]
- Zhou, Y.; Zeng, Z.; Xu, Y.; Ying, J.; Wang, B.; Majeed, M.; Majeed, S.; Pande, A.; Li, W. Application of Bacillus coagulans in Animal Husbandry and Its Underlying Mechanisms. Animals (Basel). 2020, 10(3): 454. [CrossRef]
- Wang, M. C.; Wang, Y.; W, G. L.; Xu, L. L.; Li, E. Z. Isolation and identification of Sphingomonas sp. from chicken cecum and its ammonia-degrading activity. Journal of Biotech Research. 2021, 12: 65-73. https://btsjournals.com/assets/2021v12p65-73.pdf.
- Wang, M. C.; Tang, J. Z.; Wang, G. L.; Guo, Y. N.; Guo, L.; Li, E. Z. Isolation and identification of an acid-tolerant Lactobacillus species from chicken intestine and its application. Journal of Biotech Research. 2022, 13: 142-151. https://www.btsjournals.com/assets/2022v13p142-151.pdf.
- Hanafy, B. G.; Abumandour, M. M. A.; Kandyle, R.; Bassuoni, N. F. Ultrastructural characterization of the intestine of the Eurasian common moorhen using scanning electron microscopy and light microscopy. Microsc. Res. Tech. 2022, 85(1): 106-116. [CrossRef]
- Aliverdi-Nasab, K.; Zhandi, M.; Yousefi, A. R.; Zahedi, V.; Rafieian-Naeini, H. R. The effect of acidifier supplementation on egg production performance and intestinal histology of Japanese quail (Coturnix japonica). Vet. Med. Sci. 2023, 9(1): 263-271. [CrossRef]
- Li, G.; Wang, X.; Liu, Y.; Gong, S.; Yang, Y.; Wang, C.; Wang, H.; He, D. Bile acids supplementation modulates lipid metabolism, intestinal function, and cecal microbiota in geese. Front Microbiol. 2023, 14: 1185218. [CrossRef]
- Belote, B. L.; Soares, I.; Sanches, A. W. D.; de Souza, C.; Scott-Delaunay, R.; Lahaye, L.; Kogut, M. H.; Santin, E. Applying different morphometric intestinal mucosa methods and the correlation with broilers performance under Eimeria challenge. Poult. Sci. 2023, 102(9): 102849. [CrossRef]
- Bai, S. P.; Wu, A. M.; Ding, X. M.; Lei, Y.; Bai, J.; Zhang, K. Y. Effects of probiotic-supplemented diets on growth performance and intestinal immune characteristics of broiler chickens. Poultry Science, 2013, 92: 663-670. [CrossRef]
- Samanya, M.; Yamauchi, K. E. Histological alterations of intestinal villi in chickens fed dried Bacillus subtilis var. natto. Comparative biochemistry and physiology, Part A. Molecular and integrative physiology, 2002, 133(1): 95-104. [CrossRef]
- Kridtayopas, C.; Rakangtong, C.; Bunchasak, C.; Loongyai, W. Effect of prebiotic and synbiotic supplementation in diet on growth performance, small intestinal morphology, stress, and bacterial population under high stocking density condition of broiler chickens. Poult Sci. 2019, 98(10): 4595-4605. [CrossRef]
- Elbaz A. M., Ashmawy ES, Ali SAM, Mourad DM, El-Samahy HS, Badri FB, Thabet HA. Effectiveness of probiotics and clove essential oils in improving growth performance, immuno-antioxidant status, ileum morphometric, and microbial community structure for heat-stressed broilers. Sci. Rep. 2023, 13(1): 18846. [CrossRef]
- Hong, Y.; Cheng, Y.; Li, Y.; Li, X.; Zhou, Z.; Shi, D.; Li, Z.; Xiao, Y. Preliminary Study on the Effect of Bacillus amyloliquefaciens TL on Cecal Bacterial Community Structure of Broiler Chickens. Biomed. Res. Int. 2019, 2019: 5431354. [CrossRef]
- Neveling, D. P.; Dicks, L. M. T. Probiotics: an Antibiotic Replacement Strategy for Healthy Broilers and Productive Rearing. Probiotics Antimicrob Proteins. 2021, 13: 1-11. [CrossRef]
- Xu, D.; Wu, Q.; Liu, W.; Hu, G.; Meng, H.; Wang, J. Therapeutic efficacy and underlying mechanisms of Gastrodia elata polysaccharides on dextran sulfate sodium-induced inflammatory bowel disease in mice: Modulation of the gut microbiota and improvement of metabolic disorders. Int. J. Biol. Macromol. 2023, 248: 125919. [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F. M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms. 2020, 8(4): 573. [CrossRef]
- Wielen, P. W. V. D.; Biesterveld, S.; Notermans, S.; Hofstra, H.; Knapen, F. V. Role of Volatile Fatty Acids in Development of the Cecal Microflora in Broiler Chickens during Growth. Applied and Environmental Microbiology. 2000, 66: 2536-2540. [CrossRef]
- Oviedo-Rondón, E. O.; Hume, M. E.; Hernández, C.; Clemente-Hernández, S. Intestinal microbial ecology of broilers vaccinated and challenged with mixed Eimeria species, and supplemented with essential oil blends. Poultry Science. 2006, 85(5): 854-860. [CrossRef]
- Parker, B. J.; Wearsch, P. A.; Veloo, A. C. M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria With Emerging Implications to Inflammation, Cancer, and Mental Health. Front Immunol. 2020, 11: 906. [CrossRef]
- Wang, C.; Zhao, J.; Zhang, H.; Lee, Y. K.; Zhai, Q.; Chen, W. Roles of intestinal bacteroides in human health and diseases. Crit. Rev. Food Sci. Nutr. 2021, 61: 3518-3536. [CrossRef]
- Zafar, H.; Saier, M.H. Jr. Gut Bacteroides species in health and disease. Gut Microbes. 2021, 13(1): 1-20. [CrossRef]
- Sato, Y.; Atarashi, K.; Plichta, D. R.; Arai, Y.; Sasajima, S.; Kearney, S. M.; Suda, W.; Takeshita, K.; Sasaki, T.; Okamoto, S.; et al. Novel bile acid biosynthetic pathways are enriched in the microbiome of centenarians. Nature. 2021, 599(7885): 458-464. [CrossRef]
- Cui Y, Zhang L, Wang X, Yi Y, Shan Y, Liu B, Zhou Y, Lü X. Roles of intestinal Parabacteroides in human health and diseases. FEMS Microbiol Lett. 2022,369(1): fnac072. [CrossRef]
- Pin Viso, N.; Redondo, E.; Díaz Carrasco, J. M.; Redondo, L.; Sabio, Y.; Garcia, J.; Fernández Miyakawa, M.; Farber, M. D. Geography as non-genetic modulation factor of chicken cecal microbiota. PLoS One. 2021, 16(1): e0244724. [CrossRef]
- Shilova, S. A.; Khrenova, M. G.; Matyuta, I. O.; Nikolaeva, A. Y.; Rakitina, T. V.; Klyachko, N. L.; Minyaev, M. E.; Boyko, K. M.; Popov, V. O.; Bezsudnova, E. Y. To the Understanding of Catalysis by D-Amino Acid Transaminases: A Case Study of the Enzyme from Aminobacterium colombiense. Molecules. 2023, 28(5): 2109. [CrossRef]
- Zhou, X.; Zhang, Z.; Huang, F.; Yang, C.; Huang, Q. In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed. Molecules. 2020, 25(19): 4354. [CrossRef]
- Zhuang, P.; Li, H.; Jia, W.; Shou, Q.; Zhu, Y.; Mao, L.; Wang, W.; Wu, F.; Chen, X.; Wan, X.; Wu, Y.; Liu, X.; Li, Y.; Zhu, F.; He, L.; Chen, J.; Zhang, Y.; Jiao, J. Eicosapentaenoic and docosahexaenoic acids attenuate hyperglycemia through the microbiome-gut-organs axis in db/db mice. Microbiome. 2021, 9(1): 185. [CrossRef]
- Yu, P.; Tu, W.; Wu, M.; Zhang, Z.; Wang, H. Pilot-scale fermentation of urban food waste for volatile fatty acids production: The importance of pH. Bioresour Technol. 2021, 332: 125116. [CrossRef]
- Xiao, X.; Guo, K.; Liu, J.; Liu, Y.; Yang, C.; Xu, Y.; Deng, B. The Effect of Sodium Alginate-Coated Nano-Zinc Oxide on the Growth Performance, Serum Indexes and Fecal Microbial Structure of Weaned Piglets. Animals (Basel). 2023, 14(1): 146. [CrossRef]
- Lv, J.; Guo, L.; Chen, B.; Hao, K.; Ma, H.; Liu, Y.; Min, Y. Effects of different probiotic fermented feeds on production performance and intestinal health of laying hens. Poult. Sci. 2022, 101: 101570. [CrossRef]
- Yang, T.; Du, M.; Zhang, J.; Ahmad, B.; Cheng, Q.; Wang, X.; Abbas, Z.; Tong, Y.; Li, J.; Zhou, Y.; Zhang, R.; Si, D. Effects of Clostridium butyricum as an Antibiotic Alternative on Growth Performance, Intestinal Morphology, Serum Biochemical Response, and Immunity of Broilers. Antibiotics (Basel). 2023, 12(3): 433. [CrossRef]
- Yadav, S.; Jha, R. Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. Journal of Animal Science and Biotechnology. 2019, 10: 2. [CrossRef]
Figure 1.
Weight changes of broiler chickens in each group.
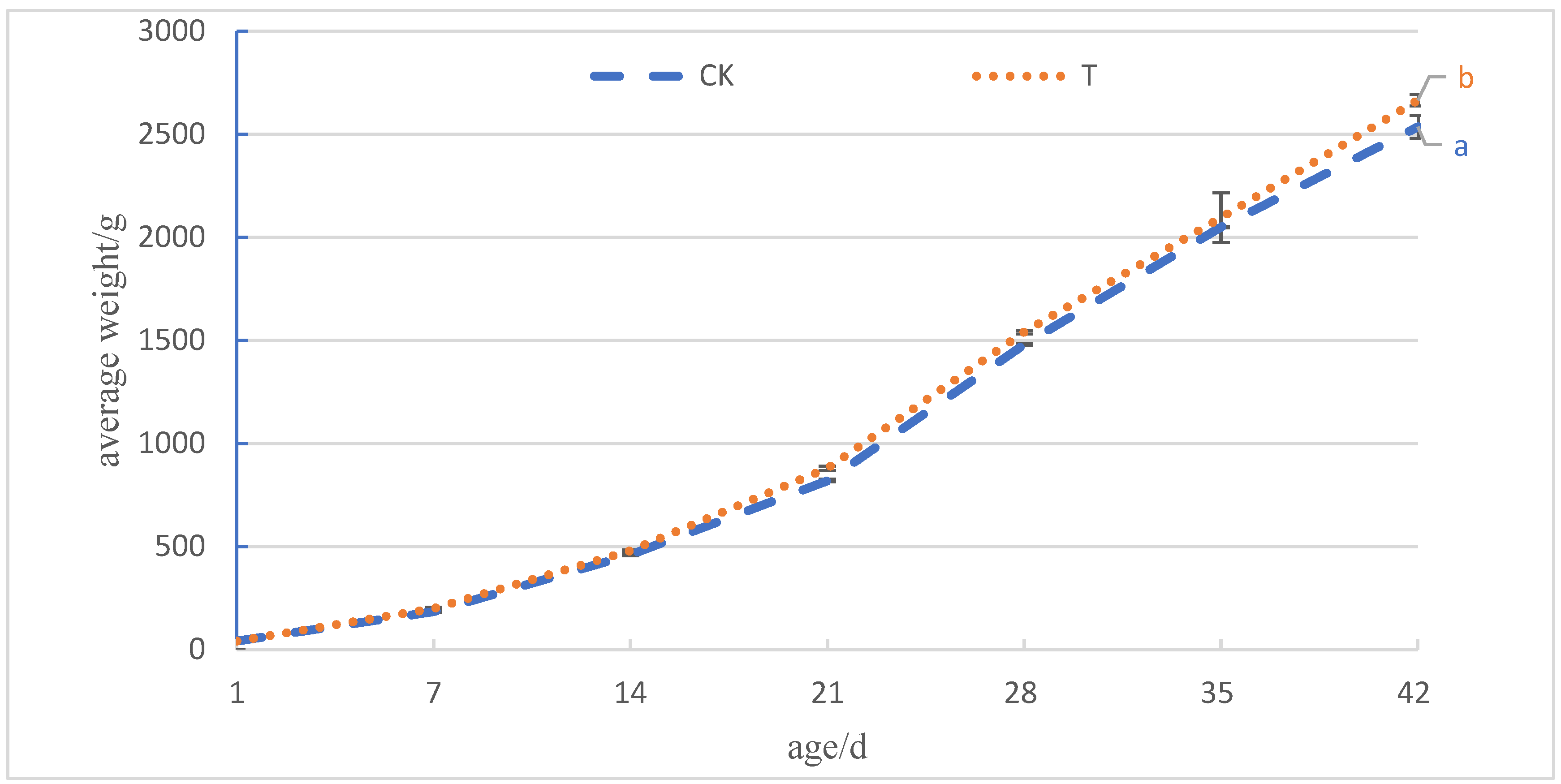
Figure 3.
The ratio of small intestine villus height to crypt depth of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water. Note: Having the same letter on the same data column indicates no significant difference (p>0.05); The absence of identical letters indicates significant differences (p<0.05).
Figure 3.
The ratio of small intestine villus height to crypt depth of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water. Note: Having the same letter on the same data column indicates no significant difference (p>0.05); The absence of identical letters indicates significant differences (p<0.05).
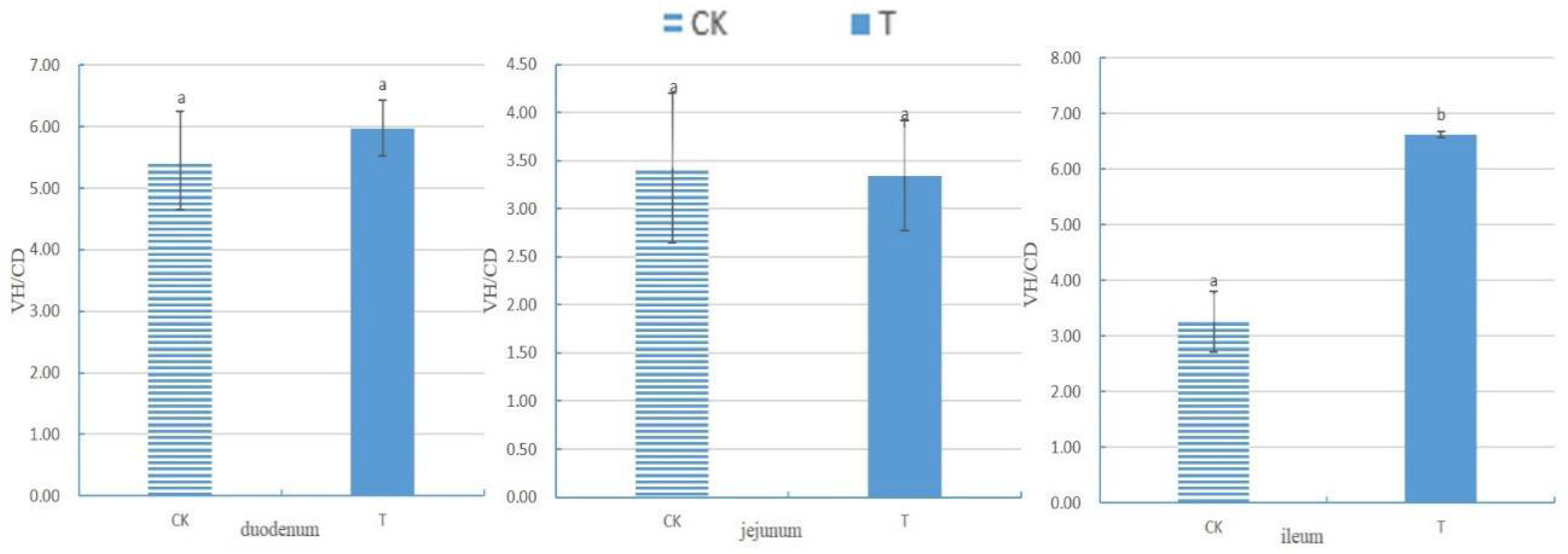
Figure 5.
Multi samples shannon curves for the broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water. Note: The horizontal axis represents the number of sequencing samples randomly selected from a certain sample, and the vertical axis represents the Shannon index index. As the sequencing quantity increases, more species are discovered, and even after species saturation, increasing the number of samples does not reveal new features.
Figure 5.
Multi samples shannon curves for the broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water. Note: The horizontal axis represents the number of sequencing samples randomly selected from a certain sample, and the vertical axis represents the Shannon index index. As the sequencing quantity increases, more species are discovered, and even after species saturation, increasing the number of samples does not reveal new features.
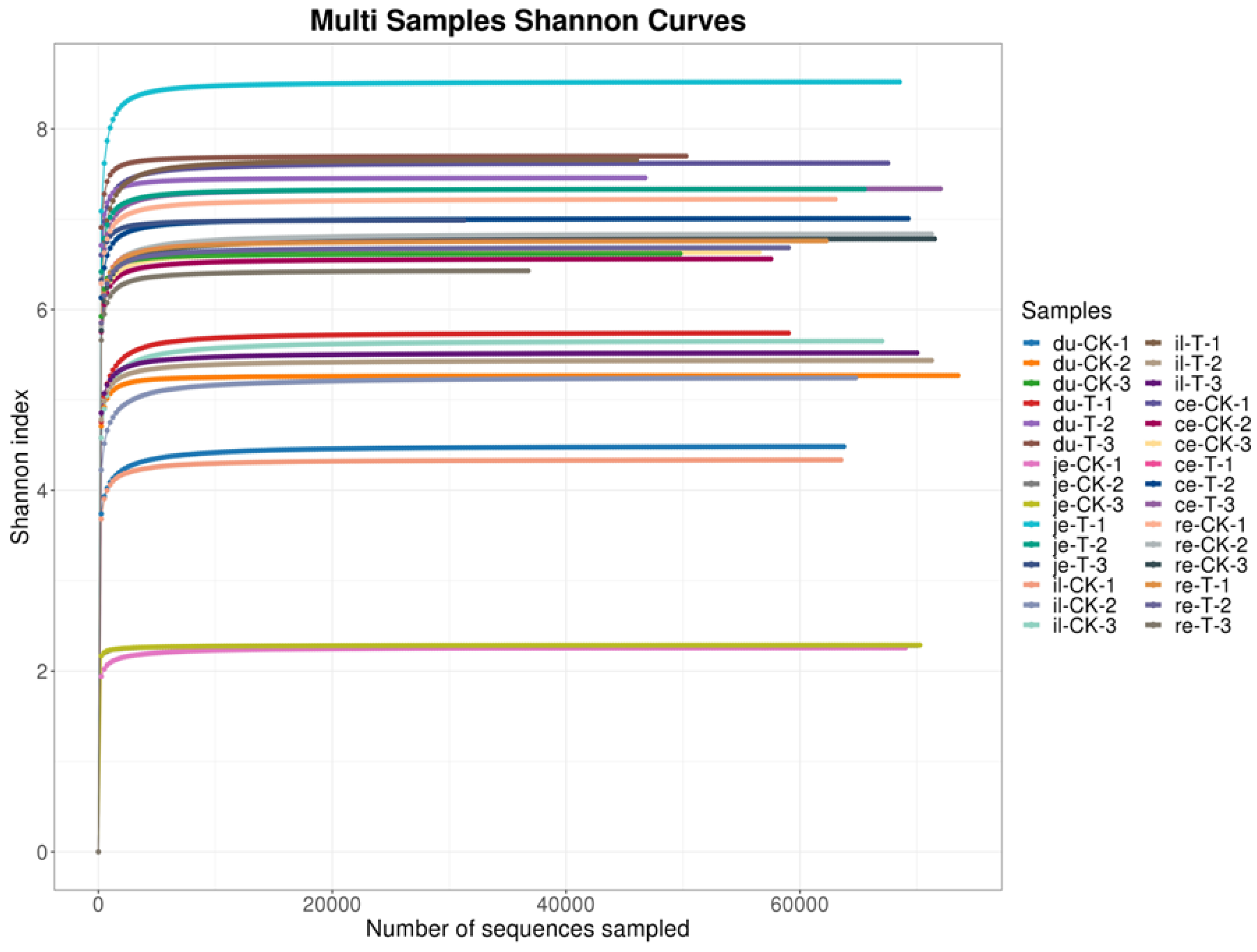
Figure 8.
The content of different genera levels of microbial flora in different intestinal segments of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Figure 8.
The content of different genera levels of microbial flora in different intestinal segments of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
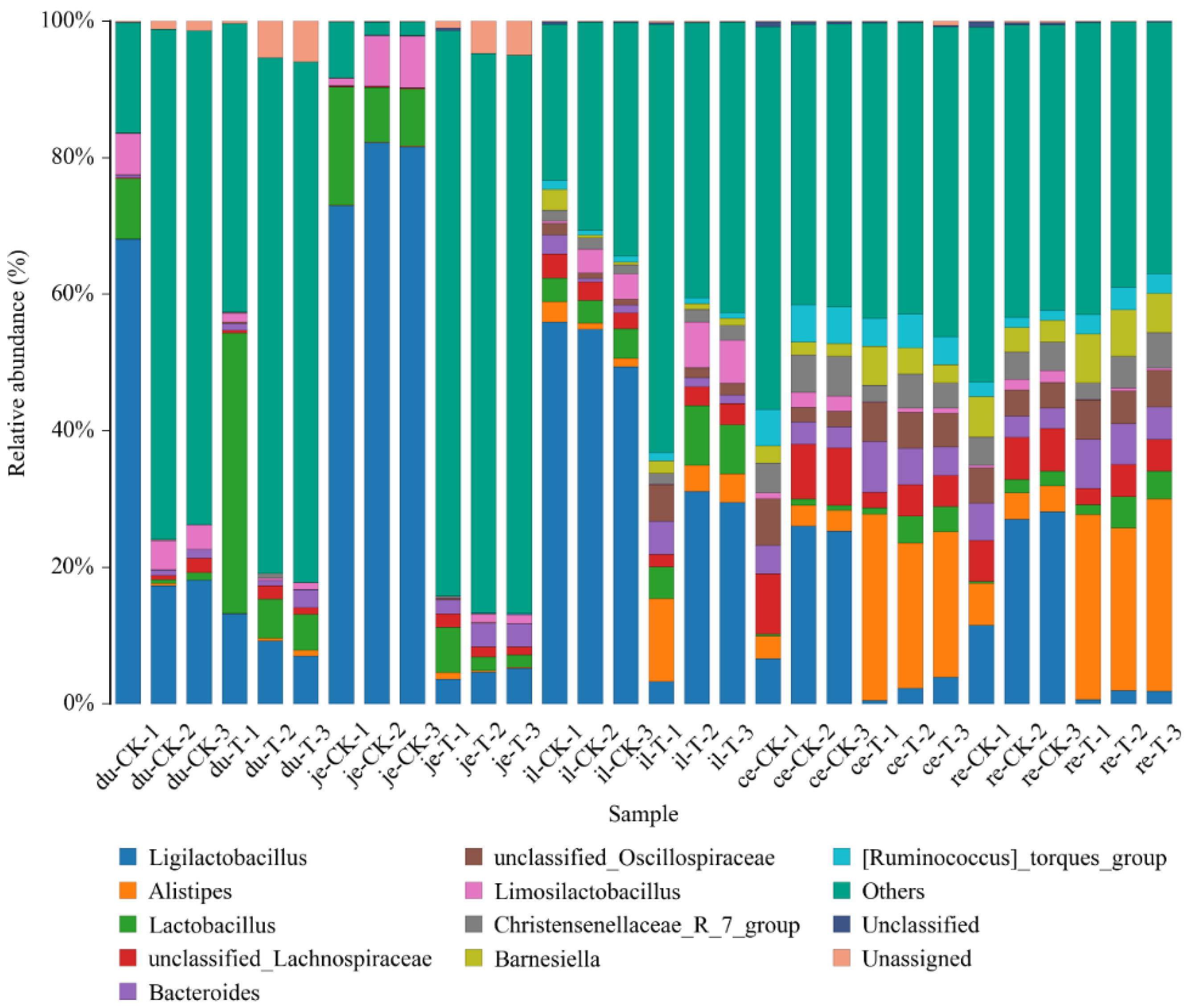
Table 1.
Nutrient composition of feed for Kebao broiler chickens at different stages (%).
| Feeding stage (Day) | Water | Crude protein | Crude fat | Crude fiber |
Crude ash |
Calcium | Total phosphorus | Sodium chloride | Methionine |
|---|---|---|---|---|---|---|---|---|---|
| 1-18 | 12 | 22 | 4 | 3 | 5 | 0.8 | 0.65 | 0.54 | ≤0.9 |
| 19-33 | 12 | 20 | 5 | 3 | 5 | 0.8 | 0.55 | 0.38 | ≤0.9 |
| 34-42 | 12 | 19 | 7 | 3 | 5 | 0.7 | 0.6 | 0.37 | ≤0.9 |
Table 2.
EPI of broiler chickens in each group.
| Sold age/d | Sold weight/kg | Survival rate | Feed conversion ratio | EPI | |
|---|---|---|---|---|---|
| CK | 42 | 2.54±0.006a | 0.96±0.006 a | 1.59±0.012a | 362.65±3.47a |
| T | 42 | 2.65±0.01b | 0.98±0.006b | 1.54±0.006b | 399.28±1.63b |
Note: The same letter on the right shoulder of the data in the same column indicates no significant difference; (p>0.05); different letters indicate significant differences (p<0.05).
Table 3.
Statistics of sample sequencing data processing results for broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 3.
Statistics of sample sequencing data processing results for broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| intestinal segment | raw reads | clean reads | denoised reads | merged reads | non chimeric read | |
|---|---|---|---|---|---|---|
| CK | du | 69584±13639.24 | 63662±12142.49 | 63583±12127.45 | 63109±12058.75 | 62608±12002.32 |
| je | 79087±1484.33 | 73337±2068.34 | 73278±2057.47 | 73062±2111.19 | 69890±643.3 | |
| il | 79965±87.37 | 75332±205.46 | 74958±275.43 | 71065±740.99 | 66047±1952.38 | |
| ce | 79953±195.22 | 73727±2624.85 | 73449±2531.63 | 68091±1050.93 | 60924±5916.03 | |
| re | 80060±268.34 | 73906±2927.62 | 73701±2957.73 | 71129±3951.46 | 69109±4771.54 | |
| T | du | 58835±7561.23 | 53492±6656.76 | 53425±6664.03 | 53043±6616.83 | 52233±6351.19 |
| je | 64566±26465.03 | 57869±22754.19 | 57758±22691.23 | 57221±22541.65 | 55390±20704.27 | |
| il | 71205±15305.56 | 65995±15788.91 | 65858±15748.33 | 64743±15233 | 62781±14352.12 | |
| ce | 79934±65.73 | 74194±2152.66 | 73969±2236.57 | 70843±4343.02 | 67302±6587.09 | |
| re | 67939±21015.84 | 62847±18892.92 | 62600±18770.72 | 58192±16406.67 | 53054±14027.97 |
Note: Raw Reads is the number of raw reads obtained from sequencing; Clean Reads is the number of high-quality reads obtained after quality control of the original sequence; Denoised Reads is the number of Clean Reads after denoising; Merged Reads is the number of sequence entries obtained by concatenating Denoised Reads based on overlap; Non chimeric Reads is the final number of sequence entries after removing chimeras.
Table 4.
Alpha diversity analysis results for the broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 4.
Alpha diversity analysis results for the broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Sample | Feature | ACE | Chao1 | Simpson | Shannon | PD_whole_tree | Coverage |
|---|---|---|---|---|---|---|---|
| CK | 642±346.47 | 645±347.56 | 648±349.01 | 1±0.16 | 5±1.85 | 79±33.14 | 1 |
| T | 574±183.03 | 576±183.48 | 577±183.79 | 1±0.03 | 7±0.85 | 93±56.73 | 1 |
Table 6.
Change folds in the content of duodenal microbiota in the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 6.
Change folds in the content of duodenal microbiota in the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | T |
|---|---|---|
| Aminobacterium | 9±15.59 | 35±60.04 |
| Bacteroides | 484±224.89 | 755±508.83 |
| Lactobacillus | 2195±2958.18 | 9870±12485.8 |
| Lachnospiraceae_NK4A136_group | 537±459.08 | 562±335.41 |
| Odoribacter | 2±3.21 | 102±169.51 |
| Sphingomonas | 205±80.18 | 597±361.21 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Table 7.
Analysis of change folds in jejunal microbiota in different groups with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 7.
Analysis of change folds in jejunal microbiota in different groups with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | WT |
|---|---|---|
| Alistipes | 56±11.37 | 300±343.23 |
| Bacteroides | 54±20.66 | 1562±625.47 |
| Barnesiella | 15±13.45 | 20±3.5 |
| Campylobacter | 53±86.89 | 39±9.07 |
| Christensenellaceae_R_7_group | 16±1.15 | 82±84.61 |
| Faecalibacterium | 32±18.04 | 64±54.6 |
| Lachnospiraceae_NK4A136_grou | 78±24.76 | 914±503.41 |
| Phascolarctobacterium | 0±0 | 40±69.28 |
| Proteiniphilum | 20±8.33 | 232±200 |
| [Ruminococcus]_torques_group | 25±22.34 | 43±41.55 |
| Staphylococcus | 2±1.73 | 35±28.21 |
| Sphingomonas | 35±22.3 | 313±128.91 |
| unclassified_Butyricicoccaceae | 0±0 | 6±2.05 |
| unclassified_Oscillospiraceae | 16±1.15 | 82±84.61 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Table 8.
Change folds in ileal microbiota content in different groups of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 8.
Change folds in ileal microbiota content in different groups of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | WT |
|---|---|---|
| Aminobacterium | 2±4.04 | 18±31.18 |
| Bacteroides | 931±729.93 | 1353±775.55 |
| Bilophila | 58±58.04 | 39±13.58 |
| Campylobacter | 24±33.86 | 29±14.43 |
| Escherichia_Shigella | 63±46.61 | 156±122.19 |
| Parabacteroides | 572±586.62 | 305±20.53 |
| Proteiniphilum | 0±0 | 75±125.6 |
| Sphingomonadaceae | 42±37.24 | 10±16.74 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Table 9.
Change folds in the content of cecal microbiota in different groups with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 9.
Change folds in the content of cecal microbiota in different groups with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | WT |
|---|---|---|
| Anaerotruncus | 25±27.78 | 5±5.03 |
| Barnesiella | 1281±350.15 | 2652±705.11 |
| Caproiciproducens | 0±0 | 3±5.77 |
| Christensenellaceae_R_7_group | 2143±172.87 | 2522±1006.54 |
| Rikenella | 0±0 | 392±82.25 |
| Sphingomonadaceae | 3±5.2 | 0±0 |
| unclassified_Clostridia_UCG_014 | 2470±663.49 | 1549±293.3 |
| unclassified_Oscillospiraceae | 2387±1956.92 | 3533±107.68 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Table 10.
Change folds of rectal microbiota content in different groups of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 10.
Change folds of rectal microbiota content in different groups of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | WT |
|---|---|---|
| Rikenella | 5±4.16 | 377±163.69 |
| Ruminococcus | 1126±180.33 | 1589±476.63 |
| Sphingomonadaceae | 3±5.2 | 2±4.04 |
| unclassified_Clostridia_UCG_014 | 980±303.77 | 305±114.13 |
| unclassified_Oscillospiraceae | 112±20.53 | 256±103.01 |
| uncultured_Firmicutes_bacterium | 2609±715.17 | 3247±1414.46 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.
Submitted:
09 May 2024
Posted:
12 May 2024
You are already at the latest version
Alerts
A peer-reviewed article of this preprint also exists.
This version is not peer-reviewed
Submitted:
09 May 2024
Posted:
12 May 2024
You are already at the latest version
Alerts
Abstract
Probiotics have emerged as a prominent alternative to antibiotics in antimicrobial-free broiler farming. To assess the effect of the probiotics Sphingomonas sp. Z392 isolated and identified by this this laboratory on broiler growth, we conducted a study to examine the changes in broiler body weight, EPI, intestinal histological structure and gut microbiota. The results showed that supplementation of broiler drinking water with 4 × 105 CFU/mL of Sphingomonas resulted in an increase in the relative abundance of Lactobacillus, Bacteroides, Lachnospiraceae, Aminobacterium, Oribacterium, Christensenellaceae, Faecalibacterium, Barnesiella, Ruminococcus, Parabacteroides, Phascolarctobacterium, Butyricicoccaceae, and Caproiciproducens, which were reported to be positively correlated with the digestion and absorption of broiler chickens. The relative abundance was increased of Odoribacter, Alistipes, Parabacteroides, and Rikenellaceae, which were reported to be negatively correlated with the occurrence of intestinal diseases. The relative abundance was decreased of Campylobacter, Shigella Castellani, Bilophila, Campylobacter, Clostridia, and Anaerotruncus, which were reported positively correlated with the occurrence of intestinal diseases. At the same time, it was increased in the integrity of small intestinal villus morphology, the number of goblet cells in small intestinal epithelial cells, the health of mitochondria in the cytoplasm of jejunal villous epithelial cells, and the number of lysosomes in the cytoplasm of goblet cells in small intestinal epithelium, ileal villous epithelial cells, mitochondria in the cytoplasm of large intestinal villous epithelial cells, the VH/CD of the ileum, digestive, absorption, and defense capabilities. Especially the final weight was increased by 4.33% and the EPI increased by 10.10%. Therefore, the supplementation of broiler drinking water with Sphingomonas generated better economic benefits for the broiler chickens.
Keywords:
Subject: Biology and Life Sciences - Animal Science, Veterinary Science and Zoology
1. Introduction
In the past 50 years, the poultry industry has made significant progress through breed improvement, scientific management, and advances in nutritional science, particularly in the use of feed additives, which have made outstanding contributions to the success of current broiler production [1]. With increasing concern for food safety and animal welfare, the selection and application of feed additives have become more refined and scientific [2]. Various additives such as antibiotics, probiotics, oligosaccharides, enzymes, and organic acids are widely used in poultry feed [3,4,5]. Antibiotics, as additives, have been widely favored for their effectiveness in preventing diseases and promoting weight gain [6]. But due to the emergence of drug-resistant strains and the resultant concern related to public health, China has now banned the use of antibiotics in feed additives for chicken farming. This change has driven the emergence of alternative approaches to replace antibiotics. Among them, probiotics are widely used as substitutes for antibiotics due to their unique advantages [7]. They can increase the number of beneficial microorganisms in the intestine, promote host intestinal health, and assist in the digestion and absorption of nutrients by producing hydrolytic enzymes [8]. They can also enhance immunity by regulating the gut microbiota and reduce the colonization of pathogenic bacteria in the intestine, thereby improving the structure of the gut microbiota [6,9]. In broiler farming, microorganisms such as Lactobacillus, Lactobacillus bulgaricus, Lactobacillus acidophilus, Lactobacillus casei, Lactobacillus salivarius, Streptococcus thermophilus, Enterococcus faecalis, Lactobacillus plantarum, fecal coliform, Bifidobacterium, Aspergillus oryzae and brewing yeast lactis have been widely used [6,10,11,12,13,14]. These probiotics not only help improve intestinal health and feed utilization, but also promote the growth and development of broilers and improve meat quality.
Sphingomonas belongs to the class of Proteobacteria α-4 subclasses, Gram negative bacteria. The characteristics of the colony are as follows: yellow, round (diameter 2-3 mm), neat edges, central protrusion, smooth and glossy surface, moist, and sticky texture. The cellular characteristics include short rod-shaped cells, lacking spores, and having unilateral polar flagella.
In our preliminary research, Sphingomonas Z392 was isolated from chicken cecum using a selective culture medium with ammonium nitrogen as the sole nitrogen source. It could use ammonium nitrogen in the intestine as a nitrogen source to convert it into nitrite nitrogen and further convert nitrite nitrogen into nitrate nitrogen, acting as a comammox bacterium [15].
In order to investigate the effect of Sphingomonas Z392 on the growth of broiler chickens, this study explored the role and mechanism of this strain in promoting growth in broiler chickens through changes in body weight, EPI, intestinal histological structure, and gut microbiota structure, providing reference for the application of Sphingomonas Z392 in broiler production.
2. Materials and Methods
2.1. Experimental Design and Animal Breeding Management
600 one-day-old Kebao broiler chickens weighing 42 ± 2 g provided by Henan Longhua Animal Husbandry Co., Ltd. were randomly divided into two groups: control group (CK) and experimental group (T). Each group had 3 replicates, with 100 broiler chickens raised in each replicate. The experimental group of broiler chickens added Z392 4.0 × 105 CFU/mL of Sphingomonas to their drinking water. The feeding period for broiler chickens was 42 days, and the feeding management was carried out in accordance with the "Kebao Broiler Feeding Management Manual", using indoor cage breeding, free feeding and drinking. Drinking water was added 6 times a day, with bacterial agents added according to the amount of water consumed each time.
The use of experimental animals had been approved by the Ethics Committee of Experimental Animals of Huanghuai University and strictly follows the provisions of the "Ethical Review of Experimental Animals of Huanghuai University" (Permit number: 202009220005).
The feeding management of Kebao broilers was divided into three stages: 1-18 day old, 19-33 day old, and 34-42 day old. The nutritional composition of the feed in each stage was detailed in Table 1.
2.2. Weight Changes of Broiler Chickens in Each Group
At the age of 1 day, 5 broiler chickens were randomly selected from all the two groups, and their weight was measured using an electronic scale and recorded.
At the ages of 7 d, 14 d, 21 d, 28 d, 35 d, and 42 d, 5 chickens were randomly selected from each group, and their weight was measured using an electronic scale and recorded.
2.3. European Production Efficiency Factor for Each Group of Broilers
Sold age, the survival rate, sold weight, and feed conversion ratio were calculated, and the European production efficiency factor (EPI) for broiler farming was calculated. EPI = [survival rate × sold weight (kg)] / (feed conversion ratio × sold age) × 10000 [16].
2.4. Observation Methods of Intestinal Microstructure Tissue
After 42 d broiler chickens were euthanized by bleeding, the duodenum, jejunum, ileum, cecum, and rectum were collected in a timely manner. After being fixed with formalin, they were embedded in paraffin, sliced, hematoxylin-eosin (H&E) staining, and sealed. The changes in the intestinal microstructure were observed under a microscope and photographed. Case Viewer 2.4 browsing software was used for analysis and observation.
2.5. Observation Method of Intestinal Ultra Microscopic Structure Tissue
After 42 d, the broiler chickens were euthanized by bleeding, the duodenum, jejunum, ileum, cecum, and rectum were collected in a timely manner. After being fixed with the fixative for electron microscopy, dehydrated at room temperature, resin penetration and embedding, polymerized, thinly sliced, stained, and observed under transmission electron microscopy (TEM) (HT7800, Hitachi Production Co., Ltd) for image analysis.
2.6. Sequencing of Gut Microbiota
After 42 d broiler chickens were euthanized by bleeding, 5 intestinal segments duodenum (du), jejunum (je), ileum (il), cecum (ce), and rectum (re) were collected in a timely and sterile manner, each with a length of 3 cm. After ligation at both ends, they were sent to Wuhan Servicebio Technology Co., Ltd. for testing. Based on the Illumina platform, the genome of the gut microbiota was sequenced and bioinformatics analysis was performed using a two end sequencing (PE250) method. The gut microbiota of broiler chickens was analyzed and determined using 16s rDNA sequencing technology.
2.7. Methods of Data Statistical Analysis
The experimental data were sorted out by Microsoft Excel 2013, and the mean and standard deviation were calculated, and draw line and bar charts. SPSS 14.0 software was used for analysis of variance.
3. Results
3.1. Changes in Body Weight of Broiler Chicken in Each Group
Take the average weight of broiler chickens aged 1, 7, 14, 21, 28, 35, and 42 days in each group, and create a graph of broiler weight (g) - day (d) (Figure 1). The average weight in test (T) group at selected time points was significantly higher than that in control (CK) group (p<0.05).
Note: Different letters at the end of the line chart indicate significant differences between the two groups of data (p<0.05).
3.2. Comparison of EPI between Test and Control Groups of Broilers
During the experiment, the EPI of broilers in each group were detailed in Table 2. Both groups of broiler chickens have European indices above 300, achieving profitability. Briefly, addition of 4.0 × 105 CFU/mL of Sphingomonas Z392 to drinking water can increase the final weight by 4.33% and the EPI by 10.10%, resulting in better economic benefits for broilers.
3.3. Changes in Intestinal Microstructure
After paraffin sectioning and H&E staining of various intestinal tissues, the results were observed under a binocular biological microscope, as shown in Figure S1-S3. In the experimental group, the integrity of the small intestinal villi enhanced, and the number of goblet cells in the epithelial cells of the small intestine increased. However, in the control group, the small intestinal villi of the broiler chickens became shorter and fragmented, and the villi did not show their inherent morphology, especially the duodenum and jejunum, which were most severely damaged. There were no visible differences in the folds and villi of the cecum and rectum in each group of broiler chickens.
3.4. Changes in the Ratio of Small intestinal Intestinal Villus Height to Crypt Depth
After measuring the villus height (VH) and crypt depth (CD) of the small intestine using Case Viewer 2.4 browsing software, the ratio of villus height to crypt depth (VH/CD) of each segment of the small intestine was calculated, as shown in Figure 3. There was no significant difference in VH/CD between the duodenum and jejunum groups (p>0.05). The VH/CD of the ileum in the experimental group (6.62 ± 0.06) was significantly higher than that in the control group (3.25 ± 0.55) (p<0.05).
3.5. Changes in the Ultramicroscopic Structure of the Intestine
After preparing ultra-thin sections from different intestinal segments and staining, they were observed under transmission electron microscopy, as shown in Figure S4-S8. In the test group, addition of 4 × 105 CFU/mL of Sphingomonas Z392 into the drinking water of broiler chickens reduced the degree of mitochondrial swelling in the cytoplasm of jejunal villous epithelial cells and increased the number of goblet cells in the small intestine epithelium, lysosomes in villous epithelial cells in the ileum, and mitochondria in villous epithelial cells in the large intestine.
3.616. S rRNA Gene Sequence Statistics
After Illumina NovaSeq sequencing, the 16S rRNA gene sequence data from fecal samples of the duodenum, jejunum, ileum, cecum, and rectum from the two groups of chickens were presented in Table 3. Following quality control, the removal of chimeric sequences and those out of the target range, the average number of 16S rRNA gene sequences retained for analysis in each intestinal segment of the CK group was 62,608 in the duodenum, 69890 in the jejunum, 66047 in the ileum, 60924 in the cecum, and 69109 in the rectum. The average number of 16S rRNA gene sequences retained for analysis in each intestinal segment of the T group was 52233 in the duodenum, 55390 in the jejunum, 62781 in the ileum, 67302 in the cecum, and 53054 in the rectum.
3.7. Alpha Diversity Analysis
From Figure 5 and Table 4, it can be observed that, at a similarity level of 97%, the bioinformatics analysis of OTU showed no significant difference in the Simpson index and Shannon index between the T and the CK groups (p>0.05). This finding suggests a high degree of consistency in the diversity of gut microbiota between the CK and T groups.
3.8. NMDS Non Metric Multidimensional Scale Analysis
By using NMDS non metric multidimensional scale analysis on the bacterial communities at the genus level in 5 different parts, the results were shown in Figures 4-10. There is a significant overlap between the microbial communities of the CK and T groups in the duodenum, indicating that the differences in microbial structure and composition between the CK and T groups in the duodenum are relatively small. However, the microbiota of the CK and T groups in the jejunum, ileum, cecum, and rectum did not cross indicating significant differences in the gut microbiota structure and composition between the CK and T groups in the jejunum, ileum, cecum, and rectum.
Figure 6.
Non metric multidimensional scale analysis of NMDS in different intestines of different groups of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Figure 6.
Non metric multidimensional scale analysis of NMDS in different intestines of different groups of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Note: 1 A, B, C, D, and E are the results of non metric multidimensional scale analysis of the duodenum, empty field, ileum, cecum, and rectum, respectively.
2. The appearance of obvious cross indicates that there is no significant difference in microbial structure and composition between groups, while the absence of cross indicates significant differences in microbial structure and composition between groups.
3. Each point in the figure represents a sample; Different colors represent different groups; The elliptical circle represents a 95% confidence ellipse (i.e., if there are 100 samples in the sample group, 95 will fall within it). When the stress is less than 0.1, it can be considered a good sorting; When the stress is less than 0.05, it has good representativeness. It is generally believed that when the Stress is less than 0.2, it indicates that NMDS analysis has a certain level of reliability. The closer the sample is on the coordinate map, the higher the similarity.
3.9. Analysis of Microbial Diversity at Phylum Level
The classification results of gut microbiota in various intestinal sites across different experimental groups are presented in Table 5 and Figure 7. At the phylum level, a total of 47 phyla are involved in the gut microbiota of the five parts. To facilitate the observation of relationships and species composition among samples, a bar chart of species distribution was constructed, based on the abundance of each species within each sample. At the phylum level, there are 10 phyla with higher relative abundance, including Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria, Acidobacteriota, Chloroflexi, Cyanobacteria, Desulfobacteria, Gemmatimonadota, and unclassified phyla.
Table 5.
Classification and statistical results of microbial flora in different intestinal segments based on various classification levels of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 5.
Classification and statistical results of microbial flora in different intestinal segments based on various classification levels of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Samples | Kindom | Phylum | Class | Order | Family | Genus | Species | |
| CK | du | 2±0 | 27±2.52 | 50±14.01 | 107±35 | 177±73.93 | 254±126.89 | 276±143.62 |
| je | 2±0 | 21±3.21 | 35±11.02 | 71±25.51 | 109±47.18 | 149±71.77 | 160±72.92 | |
| il | 2±0.58 | 28±6.08 | 55±15.59 | 127±49.39 | 226±95.31 | 381±177.79 | 417±187.16 | |
| ce | 1±0.58 | 13±5.77 | 22±10.97 | 45±20.5 | 83±38.73 | 149±55.18 | 178±55.07 | |
| re | 2±0.58 | 25±6.66 | 51±20.21 | 115±43.04 | 195±75.92 | 306±115.77 | 345±125.3 | |
| T | du | 2±0 | 25±3.21 | 50±11.24 | 109±32.08 | 170±69.46 | 252±129.69 | 269±140.88 |
| je | 2±0 | 27±3.79 | 51±5.51 | 107±13.05 | 169±33.08 | 248±67.1 | 264±74.57 | |
| il | 2±0 | 24±2.08 | 50±5.29 | 119±10.79 | 200±10.26 | 310±22.5 | 348±25.42 | |
| ce | 2±0 | 17±5 | 29±11.14 | 63±25.79 | 103±30.12 | 175±43.27 | 209±44.6 | |
| re | 1±0 | 12±6.08 | 20±10.97 | 46±19.35 | 80±27.73 | 143±38.4 | 173±41.88 | |
Note: The numbers in the table represents the total number of Reads covered by the sample at that level.
Figure 7.
The content of microbial communities in different intestinal segments of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water at the phylum level.
Figure 7.
The content of microbial communities in different intestinal segments of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water at the phylum level.
Note: The horizontal axis represents the sample name; The vertical axis represents the relative abundance percentage (%). Different colors represent different species; Stacked columns represent the top 10 taxonomic groups with relative abundance at each taxonomic level.
3.10. Diversity Analysis of Microbial Communities at the Genus Level
To facilitate the observation of relationships between the samples and composition of microbial species, a figure depicting the distribution of microbial species for each sample was constructed, based on the abundance of each microbial species within each sample (Figure 8). There were 10 genera with higher relative abundance, including Alistipes, Bacteroides, Christensenellaceae R_7_group, Faecalibacterium, Lactobacillus, Limosillactobacillus, Ruminococcus_torques_group, unclassified Lachnospiraceae, unclassified Oscillospiraceae, and others, all of which exhibit intergroup differences.
Note: The horizontal axis represents the sample name; The vertical axis represents the relative abundance percentage (%). Different colors represent different species; Stacked columns represent the top 10 taxonomic groups with relative abundance at each taxonomic level.
3.10.1. Differential Analysis of Duodenal Microbiota at the Genus Level
The content of bacterial strains in the duodenum and the changes in different groups are shown in Table 6. Compared with the CK group, in the T group, the relative abundance were increased of Actobacillus, Bacteroides, Lachnospiraceae, and Aminobacterium, which were positively correlated with digestion and absorption. On the other hand, the relative abundance were increased of Odoribacter, which was negatively correlated with the occurrence of intestinal diseases.
3.10.2. Differential Analysis on Jejunum Microbiota at the Genus Level
The content of bacterial strains in the jejunum and the changes in different groups are shown in Table 7. Compared with the CK group, in the T group, the relative abundance were increased of Bacteroides, Lachnospiraceae, Oribacterium, Christensenellaceae, Proteiniphilum, Faecalibacterium, Barnesiella, Ruminococcus, Phascolarctobacterium, and Butyricicoccaceae, which were positively correlated with digestion and absorption. The relative abundance was increased of Alistipes and Paraacteroides distasonis, which was negatively correlated with the occurrence of intestinal diseases. But the abundance of Staphylococcus in the jejunum was also significantly increased.
3.10.3. Differential analysis on ileum microbiota at the genus level
The content of bacterial strains in the ileum and the changes in different groups are shown in Table 8. Compared to the CK group, the T group exhibited an increase in the relative abundance of Bacteroides, Aminobacterium, and Proteiniphilum, which were positively correlated with digestion and absorption; the relative abundance was increased of Parabacteroides, which was negatively correlated with the occurrence of intestinal diseases; the relative abundance was reduced of Campylobacter, Escherichia, and Bilophila, which was positively correlated with the occurrence of intestinal diseases.
3.10.4. Differential Analysis of Cecal Microbiota at the Genus Level
The content of bacterial strains in the cecal and the changes in different groups are shown in Table 9. Compared with the CK group, in the T group, the relative abundance were increased of Barnesiella, Oribacterium, Christensenellaceae, and Caproiciproducens, which were positively correlated with digestion and absorption; the relative abundance was increased of Rikenellaceae, which was negatively correlated with the occurrence of intestinal diseases; the relative abundance was reduced of Clostridia, Anaerotruncus, and Helicobacter, which was positively correlated with the occurrence of intestinal diseases.
3.10.5. Differential Analysis on Rectal Microbiota at the Genus Level
The content of bacterial strains in the rectal and the change folds in different groups are shown in Table 10. Compared with the CK group, in the T group, the relative abundance was increased of Bacteroides, Oribacterium, and Ruminococcus, which were positively correlated with digestion and absorption; the relative abundance was increased of Rikenellaceae, which was negatively correlated with the occurrence of intestinal diseases; the relative abundance was reduced of Clostridia, which was positively correlated with the occurrence of intestinal diseases.
4. Discussion
4.1. Effects of Sphingomonas on the Histological Structure of Intestine
The gastrointestinal tract (GIT) serves as the primary site for digestion and absorption, and defense against harmful foreign microorganisms. Its function correlates positively with organelle numbers and health status. The villi in the small intestine play a crucial role in nutrient absorption, and the height, integrity, and cell count of these villi significantly affect small intestine digestive and absorption functions [17]. After adding 4 × 105 CFU/mL of Sphingomonas Z392 to drinking water, broiler chickens exhibited increased integrity of small intestinal villi, an augmented number of goblet cells in small intestinal epithelial cells, enhanced mitochondrial health in the cytoplasm of jejunal villous epithelial cells, and elevated counts of lysosomes in the cytoplasm of goblet cells in the small intestinal epithelium, ileal villous epithelial cells, and mitochondria in the cytoplasm of large intestinal villous epithelial cells. The VH/CD of the ileum significantly increased (p<0.05).
VH, CD, and VH/CD serve as vital indicators for measuring intestinal nutrient digestion and absorption abilities, as well as its health status [18]. The villus's edges secrete various digestive enzymes; thus, higher VH indicates stronger enzyme secretion capacity and better digestion and absorption abilities. CD reflects cell generation rates, with shallower crypts indicating increased cell maturation rates and enhanced intestinal secretion function. A higher VH/CD value signifies stronger intestinal digestive and absorption capacities, while a decrease indicates weakened abilities [19].
Previous research indicates that adding probiotics to the diet can protect the gastrointestinal tract from excessive feeding, increase VH and VH/CD in broiler chickens, and reduce CD [20]. The results of this experiment demonstrate that adding Sphingomonas to broiler chickens' drinking water significantly increased the VH/CD ratio of the ileum and enhanced its digestive, absorption, and defense abilities. Moreover, the addition of Sphingomonas Z392 to drinking water led to a notable increase in intestinal probiotics such as Lactobacillus, which would activate cell mitosis and induce intestinal epithelial cell proliferation by secreting short-chain fatty acids (SCFAs), resulting in increased intestinal villus length [21,22].
4.2. Effects of Sphingomonas on Gut Microbiota
Studying the relative abundance and diversity of the gut microbiota is crucial for understanding the impact of beneficial bacteria [23]. In this experiment, the Shannon curve's horizontal width for multiple samples in each group was relatively large, indicating high species abundance in the samples. Additionally, the curve appeared relatively flat, suggesting a relatively uniform distribution of species across each group of samples. The coverage index for each group was 1, indicating that the different intestinal sequencing results in this experiment accurately reflected the true microbial composition of the samples.
The degree of intestinal health is determined by indicators such as microbial content and morphology, with the distribution of microorganisms varying among different intestinal segments [24]. The gut microbiota plays a crucial role in maintaining animal health, immunity, and production performance, and probiotics have been shown to improve the microecological environment of broiler intestines [23]. The relative research previously indicated that adding Bacillus amyloliquefaciens to the diet can increase the relative abundance of Bacteroidetes, Butyricicoccaceae, Faecalibacterium, Heliobacillus, Lactobacillus, Parabacteroides, and Ruminococcus in the gut of broiler chickens [25].
This experiment found that adding 4 × 105 CFU/mL Sphingomonas Z392 to the drinking water of broiler chickens increased the relative abundance of microorganisms, including actobacillus in the duodenum; Bacteroides in the duodenum, jejunum, ileum, and rectum; Lachnospiraceae in the duodenum and jejunum; Aminobacterium in the duodenum and ileum; Oribacterium in the jejunum, cecum, and rectum; Christensenellaceae of bacteria in the jejunum and cecum; Proteiniphilum in the jejunum and ileum; Faecalibacterium in the jejunum; Barnesiella in the jejunum and cecum; Ruminococcus in the jejunum and rectum; Phascolarctobacterium in the jejunum; Butyricicoccaceae in the jejunum; Caproiciproducens in the cecum. The above microorganisms could promote the digestion and absorption ability of broiler chickens. Furthermore, the microorganisms with reduced relative abundance include Odoribacter in the duodenum, Alistipes in the jejunum, Parabacteroides in the jejunum, ileum, and rectum, and Rikenellaceae in the cecum and rectum. These microorganisms could inhibit the occurrence of intestinal diseases [26,27].
The microorganisms Actobacillus, Lachnospiraceae, Oribacterium, Christensenellaceae, Faecalibacterium, Ruminococcus, and Caproiciproducens belong to Firmicutes, which can hydrolyze starch and other sugars to produce butyrate and other secretes short-chain fatty acids (SCFAs), such as acetate, propionate, butyrate, and lactate [28,29]. SCFAs play multiple roles in the intestine. They not only provide energy to the body, but also reduce the types of harmful bacteria, stimulate the proliferation and differentiation of intestinal epithelial villous cells, increase the height of villi, expand the contact area between villi and chyme, and improve the digestion and absorption ability of broilers [29]. In addition, the increase of beneficial microorganisms such as Actobacillus in the intestine also helps to maintain the integrity of the intestinal structure, promote metabolism, and thereby plays an important protective role as the first line of defense against pathogenic bacteria [30].
In addition, Alistipes, Odoribacter, Rikenellaceae, Bacteroides, and Parabacteroides belong to Bacteroidetes. They primarily digest grain feed and secrete mucin to protect and lubricate the intestines, exerting an anti-inflammatory effect [31,32,33]. Bacteroidetes can also enhance the disease resistance of broiler chickens by stimulating the immune system, increasing macrophage phagocytosis, and resisting the colonization of pathogenic bacteria [32]. The lithocholic acid secreted by Odoribacter promotes fat digestion and absorption, regulates blood lipids, promotes cell proliferation, exhibits anti-inflammatory properties, and protects the gastric mucosa. Additionally, the lithocholic acid produced by Odoribacter demonstrates excellent anti-inflammatory and antibacterial activities. Even in small amounts, it effectively eliminates pathogenic microorganisms such as Clostridium and Enterococcus faecalis, reduces inflammation levels, and regulates the body's immunity [34]. Bacteroides, a core member of the gut microbiota, regulates the host mucosal immune system, reduces inflammation, participates in carbon metabolism, and secretes SCFAs such as acetate and propionate [35]. Rikenellaceae in the intestine plays a certain role in protecting the body's health and alleviating diseases [36].
Aminobacterium Bacteroides, a core member of the gut microbiota, regulates the host mucosal immune system, reduces inflammation, participates in carbon metabolism, and secretes short-chain fatty acids (SCFAs) such as acetate and propionate [37]. Phascolarctobacterium produces SCFAs, including acetate and propionate, reduces inflammation, and protects the intestinal mucosal barrier by decreasing levels of lipopolysaccharide (LPS)-binding proteins and C-reactive protein (CRP) [38]. Barnesiella is associated with bile acid production and plays a crucial role in fat metabolism [39]. Proteiniphilum contributes to the conversion of acetic acid to butyric acid [40]. Butyricicoccaceae produces digestive enzymes in the intestine that break down starch, protein, and cellulose, promoting the digestion and absorption of these nutrients. Furthermore, butyric acid, the main metabolite of this genus, promotes the regeneration and repair of intestinal epithelial cells. Therefore, Butyricicoccaceae significantly influences the intestinal microbiota's structure, inhibits pathogenic bacteria, and promotes the growth of beneficial bacteria such as Lactobacillus [41].
Probiotics not only enhance nutrient digestion and absorption, improve the breeding environment, but also reduce pathogenic microorganisms and increase the relative abundance of anti-inflammatory microorganisms, thereby enhancing the body's resistance to infections [25]. This study revealed that adding Sphingomonas Z392 to the drinking water of broiler chickens decreased the relative abundance of microorganisms such as Campylobacter in the jejunum and ileum, Shigella Castellani in the ileum, Bilophila in the ileum, Clostridia in the cecum and rectum, and Anaerotruncus in the cecum. These microorganisms are positively correlated with intestinal diseases [26,27].
But after adding Sphingomonas to the drinking water of broiler chickens, the relative abundance of Staphylococcus was increased in the jejunum, which can easily cause gastrointestinal diseases. Therefore, when using Sphingomonas, it is necessary to use other microorganisms in combination to inhibit the proliferation of potential harmful bacteria, ensuring the health and breeding benefits of broilers.
4.3. Growth Promotion of Sphingomonas spp.
In this study, the relative abundance of microorganisms positively correlated with digestion and absorption and negatively correlated with intestinal diseases in broiler chickens was significantly increased by adding Sphingomonas spp. to drinking water during the feeding process. These microorganisms promote the digestion and absorption of nutrients in chyme by secreting digestive enzymes such as amylase, protease, glycosidase, and cellulase, or produce SCFAs, which reduce the types of harmful bacteria and stimulate the proliferation and differentiation of intestinal epithelial villous cells. This results in a high increase in villi, expanding the contact area between villi and chyme, and improving the digestion and absorption ability of broiler chickens [29]. Alternatively, secreted bile acids can promote fat digestion and absorption, regulate blood lipids, promote cell proliferation, exhibit anti-inflammatory properties, and protect the gastric mucosa. Meanwhile, the addition of these strains significantly reduced the degree of inflammation in the small intestine, ensuring its basic digestive and absorption functions, and promoting the digestion of nutrients such as sugars, proteins, fats, and inorganic salts. Additionally, the number of goblet cells in the small intestine villous epithelium increases, playing an important role in maintaining intestinal health and promoting nutrient absorption. At the same time, the degree of mitochondrial swelling in the villous epithelial cells of the jejunum decreases, indicating a decrease in their degree of damage, thereby ensuring the digestive and absorption functions of the jejunum. Importantly, the VH/CD of the ileum significantly increased (p<0.05), and the number of lysosomes in the villous epithelial cells increased. This significantly improved the digestive and decomposition ability of the ileum, facilitating further absorption of nutrients in the ileum. Lactobacillus and Clostridium butyricum also significantly affect the growth, development, and feed utilization of broiler chickens by regulating small intestinal VH and CD [6,42,43].
Additionally, due to the relatively short intestine of chickens, the digestion of chyme in the small intestine is not complete, so the cecum plays an important role in the chicken's digestive system [44]. This study found that the number of mitochondria in cecal villous epithelial cells increased under the action of Sphingomonas, which promoted the absorption of nutrients by the cecum.
After adding 4.0 × 105 CFU/mL of Sphingomonas Z392 to the drinking water of broiler chickens, the improvement of intestinal histology and microbial community structure not only enhanced the absorption function of the overall digestive tract of broiler chickens, but also improved the utilization rate of feed and the growth performance of broiler chickens, resulting in an increase of 4.33% in slaughter weight and 10.10% in the EPI, resulting in better economic benefits for broiler chickens.
In practical applications, adding an appropriate amount of Sphingomonas Z392 to the drinking water of broiler chickens could significantly increase their final weight, increase the EPI, and thus bring significant economic benefits. These results indicate that Sphingomonas Z392 has broad application prospects in broiler feeding and is expected to provide new strategies for the sustainable development of the broiler industry.
5. Conclusions
Addition of 4 × 105 CFU/mL of Sphingomonas sp. Z392 to drinking water improved intestinal histology and flora structure, the function of the digestion and enhanced absorption chickens and feed utilization and resulted in a 4.33% increase in the final weight of broilers and a 10.10% increase in the EPI, which led to better economic benefits for broilers.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
W. M. C. and G. A. Z. designed the whole and wrote the final manuscript. Z. J. and Z. S. Q. carried out all experiments, data collection, and manuscript editing. W. G. L., X. H. L., G. Y. N. and L. C. Y. participated in data analysis.
Funding
This work was supported by Key R & D projects of Ningxia Hui Autonomous Region (2021BEE02032), the Science and Technology Development Plan Project of Henan province of China (172102110201 and 212102110005), the Science and Technology Innovation Demonstration Project for High Quality Agricultural Development and Ecological Protection of the Ningxia Academy of Agriculture and Forestry Sciences (NGSB-2021-12-05).
Institutional Review Board Statement
The use of experimental animals had been approved by the Ethics Committee of Experimental Animals of Huanghuai University and strictly follows the provisions of the "Ethical Review of Experimental Animals of Huanghuai University" (Permit number: 202009220005).
Data Availability Statement
1. Experimental results of intestinal ultra microstructure
Extract code: dueg
2. Experimental results of 16S rRNA sequencing of gut microbiota
Extraction code: q0u
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gado, A. R.; Ellakany, H. F.; Elbestawy, A. R.; El-Hack, M. E. A.; Mahgoub, S. A.; Herbal Medicine Additives as Powerful Agents to Control and Prevent Avian Influenza Virus in Poultry – A Review. Ann. Anim. Sci. 2019, 19(4): 905-935. [CrossRef]
- Farag, M. R.; Alagawany. M. The role of Astragalus membranaceus as immunomodulator in poultry. World's Poultry Science Journal. 2019, 75: 1-12. [CrossRef]
- Bin-Jumah, M.; El-Hack, M. E. A.; Abdelnour, S. A.; Hendy, Y. A.; Aleya, L. Potential use of chromium to combat thermal stress in animals: A review. Science of The Total Environment. 2019, 707: 135996. [CrossRef]
- Elgeddawy, S. A.; Shaheen, H. M.; El-Sayed, Y. S.; Elaziz, M. A.; Darwish, A.; Samak, D.; Batiha, G. E.; Mady, R. A.; Bin-Jumah, M.; Allam, A. A. Effects of the dietary inclusion of a probiotic or prebiotic on florfenicol pharmacokinetic profile in broiler chicken. Journal of animal physiology and animal nutrition. 2020, 104(2): 549-557. [CrossRef]
- Hussein, E. O. S.; Suliman, G. M.; Alowaimer, A. N.; Ahmed, S. H.; Swelum, A. A. Growth, carcass characteristics, and meat quality of broilers fed a low-energy diet supplemented with a multienzyme preparation. Poultry Science. 2020, 99(4): 1988-1994. [CrossRef]
- Abd El-Hack, M. E.; El-Saadony, M. T.; Shafi, M. E.; Qattan, S. Y. A.; Batiha, G. E.; Khafaga, A. F.; Abdel-Moneim, A. E.; Alagawany, M. Probiotics in poultry feed: A comprehensive review. J. Anim. Physiol. Anim. Nutr (Berl). 2020, 104: 1835-1850. [CrossRef]
- Nuria, V. S.; Yanath, B.; Ruth, R.; Eric, A.; Djamel, D. Benefits and Inputs From Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters During Food-Animal Production. Frontiers in microbiology. 2019, 10: 57. [CrossRef]
- M'sadeq, S. A.; Wu, S.; Swick, R. A.; Choct, M. Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide. Animal Nutrition. 2015: 1(1): 1-11. [CrossRef]
- Wu, Y.; Wang, B.; Zeng, Z.; Liu, R.; Tang, L.; Gong, L.; Li, W. Effects of probiotics Lactobacillus plantarum 16 and Paenibacillus polymyxa 10 on intestinal barrier function, antioxidative capacity, apoptosis, immune response, and biochemical parameters in broilers. Poult. Sci. 2019, 98: 5028-5039. [CrossRef]
- Hossain, M.; Begum, M.; Kim, I. Effect of Bacillus subtilis, Clostridium butyricum and Lactobacillus acidophilus endospores on growth performance, nutrient digestibility, meat quality, relative organ weight, microbial shedding and excreta noxious gas emission in broilers. Veterinární Medicína. 2015, 60(2): 77-86. [CrossRef]
- Liao, X. D.; Ma, G.; Cai, J.; Fu, Y.; Yan, X. Y.; Wei, X. B.; Zhang, R. J. Effects of Clostridium butyricum on growth performance, antioxidation, and immune function of broilers. Poultry Science. 2015, 94: 662-667. [CrossRef]
- Mi, J.; Chen, X.; Liao, X. Screening of single or combined administration of 9 probiotics to reduce ammonia emissions from laying hens. Poultry Science. 2019, 98: 3977-3988. [CrossRef]
- Shehata, A. A.; Tarabees, R.; Basiouni, S.; ElSayed, M. S.; Gaballah, A.; Krueger, M. Effect of a Potential Probiotic Candidate Enterococcus faecalis-1 on Growth Performance, Intestinal Microbiota, and Immune Response of Commercial Broiler Chickens. Probiotics and Antimicrobial Proteins. 2020, 12(2): 451-460. [CrossRef]
- Zhou, Y.; Zeng, Z.; Xu, Y.; Ying, J.; Wang, B.; Majeed, M.; Majeed, S.; Pande, A.; Li, W. Application of Bacillus coagulans in Animal Husbandry and Its Underlying Mechanisms. Animals (Basel). 2020, 10(3): 454. [CrossRef]
- Wang, M. C.; Wang, Y.; W, G. L.; Xu, L. L.; Li, E. Z. Isolation and identification of Sphingomonas sp. from chicken cecum and its ammonia-degrading activity. Journal of Biotech Research. 2021, 12: 65-73. https://btsjournals.com/assets/2021v12p65-73.pdf.
- Wang, M. C.; Tang, J. Z.; Wang, G. L.; Guo, Y. N.; Guo, L.; Li, E. Z. Isolation and identification of an acid-tolerant Lactobacillus species from chicken intestine and its application. Journal of Biotech Research. 2022, 13: 142-151. https://www.btsjournals.com/assets/2022v13p142-151.pdf.
- Hanafy, B. G.; Abumandour, M. M. A.; Kandyle, R.; Bassuoni, N. F. Ultrastructural characterization of the intestine of the Eurasian common moorhen using scanning electron microscopy and light microscopy. Microsc. Res. Tech. 2022, 85(1): 106-116. [CrossRef]
- Aliverdi-Nasab, K.; Zhandi, M.; Yousefi, A. R.; Zahedi, V.; Rafieian-Naeini, H. R. The effect of acidifier supplementation on egg production performance and intestinal histology of Japanese quail (Coturnix japonica). Vet. Med. Sci. 2023, 9(1): 263-271. [CrossRef]
- Li, G.; Wang, X.; Liu, Y.; Gong, S.; Yang, Y.; Wang, C.; Wang, H.; He, D. Bile acids supplementation modulates lipid metabolism, intestinal function, and cecal microbiota in geese. Front Microbiol. 2023, 14: 1185218. [CrossRef]
- Belote, B. L.; Soares, I.; Sanches, A. W. D.; de Souza, C.; Scott-Delaunay, R.; Lahaye, L.; Kogut, M. H.; Santin, E. Applying different morphometric intestinal mucosa methods and the correlation with broilers performance under Eimeria challenge. Poult. Sci. 2023, 102(9): 102849. [CrossRef]
- Bai, S. P.; Wu, A. M.; Ding, X. M.; Lei, Y.; Bai, J.; Zhang, K. Y. Effects of probiotic-supplemented diets on growth performance and intestinal immune characteristics of broiler chickens. Poultry Science, 2013, 92: 663-670. [CrossRef]
- Samanya, M.; Yamauchi, K. E. Histological alterations of intestinal villi in chickens fed dried Bacillus subtilis var. natto. Comparative biochemistry and physiology, Part A. Molecular and integrative physiology, 2002, 133(1): 95-104. [CrossRef]
- Kridtayopas, C.; Rakangtong, C.; Bunchasak, C.; Loongyai, W. Effect of prebiotic and synbiotic supplementation in diet on growth performance, small intestinal morphology, stress, and bacterial population under high stocking density condition of broiler chickens. Poult Sci. 2019, 98(10): 4595-4605. [CrossRef]
- Elbaz A. M., Ashmawy ES, Ali SAM, Mourad DM, El-Samahy HS, Badri FB, Thabet HA. Effectiveness of probiotics and clove essential oils in improving growth performance, immuno-antioxidant status, ileum morphometric, and microbial community structure for heat-stressed broilers. Sci. Rep. 2023, 13(1): 18846. [CrossRef]
- Hong, Y.; Cheng, Y.; Li, Y.; Li, X.; Zhou, Z.; Shi, D.; Li, Z.; Xiao, Y. Preliminary Study on the Effect of Bacillus amyloliquefaciens TL on Cecal Bacterial Community Structure of Broiler Chickens. Biomed. Res. Int. 2019, 2019: 5431354. [CrossRef]
- Neveling, D. P.; Dicks, L. M. T. Probiotics: an Antibiotic Replacement Strategy for Healthy Broilers and Productive Rearing. Probiotics Antimicrob Proteins. 2021, 13: 1-11. [CrossRef]
- Xu, D.; Wu, Q.; Liu, W.; Hu, G.; Meng, H.; Wang, J. Therapeutic efficacy and underlying mechanisms of Gastrodia elata polysaccharides on dextran sulfate sodium-induced inflammatory bowel disease in mice: Modulation of the gut microbiota and improvement of metabolic disorders. Int. J. Biol. Macromol. 2023, 248: 125919. [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F. M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms. 2020, 8(4): 573. [CrossRef]
- Wielen, P. W. V. D.; Biesterveld, S.; Notermans, S.; Hofstra, H.; Knapen, F. V. Role of Volatile Fatty Acids in Development of the Cecal Microflora in Broiler Chickens during Growth. Applied and Environmental Microbiology. 2000, 66: 2536-2540. [CrossRef]
- Oviedo-Rondón, E. O.; Hume, M. E.; Hernández, C.; Clemente-Hernández, S. Intestinal microbial ecology of broilers vaccinated and challenged with mixed Eimeria species, and supplemented with essential oil blends. Poultry Science. 2006, 85(5): 854-860. [CrossRef]
- Parker, B. J.; Wearsch, P. A.; Veloo, A. C. M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria With Emerging Implications to Inflammation, Cancer, and Mental Health. Front Immunol. 2020, 11: 906. [CrossRef]
- Wang, C.; Zhao, J.; Zhang, H.; Lee, Y. K.; Zhai, Q.; Chen, W. Roles of intestinal bacteroides in human health and diseases. Crit. Rev. Food Sci. Nutr. 2021, 61: 3518-3536. [CrossRef]
- Zafar, H.; Saier, M.H. Jr. Gut Bacteroides species in health and disease. Gut Microbes. 2021, 13(1): 1-20. [CrossRef]
- Sato, Y.; Atarashi, K.; Plichta, D. R.; Arai, Y.; Sasajima, S.; Kearney, S. M.; Suda, W.; Takeshita, K.; Sasaki, T.; Okamoto, S.; et al. Novel bile acid biosynthetic pathways are enriched in the microbiome of centenarians. Nature. 2021, 599(7885): 458-464. [CrossRef]
- Cui Y, Zhang L, Wang X, Yi Y, Shan Y, Liu B, Zhou Y, Lü X. Roles of intestinal Parabacteroides in human health and diseases. FEMS Microbiol Lett. 2022,369(1): fnac072. [CrossRef]
- Pin Viso, N.; Redondo, E.; Díaz Carrasco, J. M.; Redondo, L.; Sabio, Y.; Garcia, J.; Fernández Miyakawa, M.; Farber, M. D. Geography as non-genetic modulation factor of chicken cecal microbiota. PLoS One. 2021, 16(1): e0244724. [CrossRef]
- Shilova, S. A.; Khrenova, M. G.; Matyuta, I. O.; Nikolaeva, A. Y.; Rakitina, T. V.; Klyachko, N. L.; Minyaev, M. E.; Boyko, K. M.; Popov, V. O.; Bezsudnova, E. Y. To the Understanding of Catalysis by D-Amino Acid Transaminases: A Case Study of the Enzyme from Aminobacterium colombiense. Molecules. 2023, 28(5): 2109. [CrossRef]
- Zhou, X.; Zhang, Z.; Huang, F.; Yang, C.; Huang, Q. In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed. Molecules. 2020, 25(19): 4354. [CrossRef]
- Zhuang, P.; Li, H.; Jia, W.; Shou, Q.; Zhu, Y.; Mao, L.; Wang, W.; Wu, F.; Chen, X.; Wan, X.; Wu, Y.; Liu, X.; Li, Y.; Zhu, F.; He, L.; Chen, J.; Zhang, Y.; Jiao, J. Eicosapentaenoic and docosahexaenoic acids attenuate hyperglycemia through the microbiome-gut-organs axis in db/db mice. Microbiome. 2021, 9(1): 185. [CrossRef]
- Yu, P.; Tu, W.; Wu, M.; Zhang, Z.; Wang, H. Pilot-scale fermentation of urban food waste for volatile fatty acids production: The importance of pH. Bioresour Technol. 2021, 332: 125116. [CrossRef]
- Xiao, X.; Guo, K.; Liu, J.; Liu, Y.; Yang, C.; Xu, Y.; Deng, B. The Effect of Sodium Alginate-Coated Nano-Zinc Oxide on the Growth Performance, Serum Indexes and Fecal Microbial Structure of Weaned Piglets. Animals (Basel). 2023, 14(1): 146. [CrossRef]
- Lv, J.; Guo, L.; Chen, B.; Hao, K.; Ma, H.; Liu, Y.; Min, Y. Effects of different probiotic fermented feeds on production performance and intestinal health of laying hens. Poult. Sci. 2022, 101: 101570. [CrossRef]
- Yang, T.; Du, M.; Zhang, J.; Ahmad, B.; Cheng, Q.; Wang, X.; Abbas, Z.; Tong, Y.; Li, J.; Zhou, Y.; Zhang, R.; Si, D. Effects of Clostridium butyricum as an Antibiotic Alternative on Growth Performance, Intestinal Morphology, Serum Biochemical Response, and Immunity of Broilers. Antibiotics (Basel). 2023, 12(3): 433. [CrossRef]
- Yadav, S.; Jha, R. Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. Journal of Animal Science and Biotechnology. 2019, 10: 2. [CrossRef]
Figure 1.
Weight changes of broiler chickens in each group.
Figure 3.
The ratio of small intestine villus height to crypt depth of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water. Note: Having the same letter on the same data column indicates no significant difference (p>0.05); The absence of identical letters indicates significant differences (p<0.05).
Figure 3.
The ratio of small intestine villus height to crypt depth of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water. Note: Having the same letter on the same data column indicates no significant difference (p>0.05); The absence of identical letters indicates significant differences (p<0.05).
Figure 5.
Multi samples shannon curves for the broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water. Note: The horizontal axis represents the number of sequencing samples randomly selected from a certain sample, and the vertical axis represents the Shannon index index. As the sequencing quantity increases, more species are discovered, and even after species saturation, increasing the number of samples does not reveal new features.
Figure 5.
Multi samples shannon curves for the broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water. Note: The horizontal axis represents the number of sequencing samples randomly selected from a certain sample, and the vertical axis represents the Shannon index index. As the sequencing quantity increases, more species are discovered, and even after species saturation, increasing the number of samples does not reveal new features.
Figure 8.
The content of different genera levels of microbial flora in different intestinal segments of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Figure 8.
The content of different genera levels of microbial flora in different intestinal segments of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 1.
Nutrient composition of feed for Kebao broiler chickens at different stages (%).
| Feeding stage (Day) | Water | Crude protein | Crude fat | Crude fiber |
Crude ash |
Calcium | Total phosphorus | Sodium chloride | Methionine |
|---|---|---|---|---|---|---|---|---|---|
| 1-18 | 12 | 22 | 4 | 3 | 5 | 0.8 | 0.65 | 0.54 | ≤0.9 |
| 19-33 | 12 | 20 | 5 | 3 | 5 | 0.8 | 0.55 | 0.38 | ≤0.9 |
| 34-42 | 12 | 19 | 7 | 3 | 5 | 0.7 | 0.6 | 0.37 | ≤0.9 |
Table 2.
EPI of broiler chickens in each group.
| Sold age/d | Sold weight/kg | Survival rate | Feed conversion ratio | EPI | |
|---|---|---|---|---|---|
| CK | 42 | 2.54±0.006a | 0.96±0.006 a | 1.59±0.012a | 362.65±3.47a |
| T | 42 | 2.65±0.01b | 0.98±0.006b | 1.54±0.006b | 399.28±1.63b |
Note: The same letter on the right shoulder of the data in the same column indicates no significant difference; (p>0.05); different letters indicate significant differences (p<0.05).
Table 3.
Statistics of sample sequencing data processing results for broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 3.
Statistics of sample sequencing data processing results for broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| intestinal segment | raw reads | clean reads | denoised reads | merged reads | non chimeric read | |
|---|---|---|---|---|---|---|
| CK | du | 69584±13639.24 | 63662±12142.49 | 63583±12127.45 | 63109±12058.75 | 62608±12002.32 |
| je | 79087±1484.33 | 73337±2068.34 | 73278±2057.47 | 73062±2111.19 | 69890±643.3 | |
| il | 79965±87.37 | 75332±205.46 | 74958±275.43 | 71065±740.99 | 66047±1952.38 | |
| ce | 79953±195.22 | 73727±2624.85 | 73449±2531.63 | 68091±1050.93 | 60924±5916.03 | |
| re | 80060±268.34 | 73906±2927.62 | 73701±2957.73 | 71129±3951.46 | 69109±4771.54 | |
| T | du | 58835±7561.23 | 53492±6656.76 | 53425±6664.03 | 53043±6616.83 | 52233±6351.19 |
| je | 64566±26465.03 | 57869±22754.19 | 57758±22691.23 | 57221±22541.65 | 55390±20704.27 | |
| il | 71205±15305.56 | 65995±15788.91 | 65858±15748.33 | 64743±15233 | 62781±14352.12 | |
| ce | 79934±65.73 | 74194±2152.66 | 73969±2236.57 | 70843±4343.02 | 67302±6587.09 | |
| re | 67939±21015.84 | 62847±18892.92 | 62600±18770.72 | 58192±16406.67 | 53054±14027.97 |
Note: Raw Reads is the number of raw reads obtained from sequencing; Clean Reads is the number of high-quality reads obtained after quality control of the original sequence; Denoised Reads is the number of Clean Reads after denoising; Merged Reads is the number of sequence entries obtained by concatenating Denoised Reads based on overlap; Non chimeric Reads is the final number of sequence entries after removing chimeras.
Table 4.
Alpha diversity analysis results for the broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 4.
Alpha diversity analysis results for the broiler chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Sample | Feature | ACE | Chao1 | Simpson | Shannon | PD_whole_tree | Coverage |
|---|---|---|---|---|---|---|---|
| CK | 642±346.47 | 645±347.56 | 648±349.01 | 1±0.16 | 5±1.85 | 79±33.14 | 1 |
| T | 574±183.03 | 576±183.48 | 577±183.79 | 1±0.03 | 7±0.85 | 93±56.73 | 1 |
Table 6.
Change folds in the content of duodenal microbiota in the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 6.
Change folds in the content of duodenal microbiota in the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | T |
|---|---|---|
| Aminobacterium | 9±15.59 | 35±60.04 |
| Bacteroides | 484±224.89 | 755±508.83 |
| Lactobacillus | 2195±2958.18 | 9870±12485.8 |
| Lachnospiraceae_NK4A136_group | 537±459.08 | 562±335.41 |
| Odoribacter | 2±3.21 | 102±169.51 |
| Sphingomonas | 205±80.18 | 597±361.21 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Table 7.
Analysis of change folds in jejunal microbiota in different groups with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 7.
Analysis of change folds in jejunal microbiota in different groups with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | WT |
|---|---|---|
| Alistipes | 56±11.37 | 300±343.23 |
| Bacteroides | 54±20.66 | 1562±625.47 |
| Barnesiella | 15±13.45 | 20±3.5 |
| Campylobacter | 53±86.89 | 39±9.07 |
| Christensenellaceae_R_7_group | 16±1.15 | 82±84.61 |
| Faecalibacterium | 32±18.04 | 64±54.6 |
| Lachnospiraceae_NK4A136_grou | 78±24.76 | 914±503.41 |
| Phascolarctobacterium | 0±0 | 40±69.28 |
| Proteiniphilum | 20±8.33 | 232±200 |
| [Ruminococcus]_torques_group | 25±22.34 | 43±41.55 |
| Staphylococcus | 2±1.73 | 35±28.21 |
| Sphingomonas | 35±22.3 | 313±128.91 |
| unclassified_Butyricicoccaceae | 0±0 | 6±2.05 |
| unclassified_Oscillospiraceae | 16±1.15 | 82±84.61 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Table 8.
Change folds in ileal microbiota content in different groups of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 8.
Change folds in ileal microbiota content in different groups of chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | WT |
|---|---|---|
| Aminobacterium | 2±4.04 | 18±31.18 |
| Bacteroides | 931±729.93 | 1353±775.55 |
| Bilophila | 58±58.04 | 39±13.58 |
| Campylobacter | 24±33.86 | 29±14.43 |
| Escherichia_Shigella | 63±46.61 | 156±122.19 |
| Parabacteroides | 572±586.62 | 305±20.53 |
| Proteiniphilum | 0±0 | 75±125.6 |
| Sphingomonadaceae | 42±37.24 | 10±16.74 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Table 9.
Change folds in the content of cecal microbiota in different groups with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 9.
Change folds in the content of cecal microbiota in different groups with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | WT |
|---|---|---|
| Anaerotruncus | 25±27.78 | 5±5.03 |
| Barnesiella | 1281±350.15 | 2652±705.11 |
| Caproiciproducens | 0±0 | 3±5.77 |
| Christensenellaceae_R_7_group | 2143±172.87 | 2522±1006.54 |
| Rikenella | 0±0 | 392±82.25 |
| Sphingomonadaceae | 3±5.2 | 0±0 |
| unclassified_Clostridia_UCG_014 | 2470±663.49 | 1549±293.3 |
| unclassified_Oscillospiraceae | 2387±1956.92 | 3533±107.68 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Table 10.
Change folds of rectal microbiota content in different groups of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
Table 10.
Change folds of rectal microbiota content in different groups of the chicken with (T) and without (CK) Sphingomonas Z392 in drinking water.
| Genus | CK | WT |
|---|---|---|
| Rikenella | 5±4.16 | 377±163.69 |
| Ruminococcus | 1126±180.33 | 1589±476.63 |
| Sphingomonadaceae | 3±5.2 | 2±4.04 |
| unclassified_Clostridia_UCG_014 | 980±303.77 | 305±114.13 |
| unclassified_Oscillospiraceae | 112±20.53 | 256±103.01 |
| uncultured_Firmicutes_bacterium | 2609±715.17 | 3247±1414.46 |
Note: The numbers in the table represent the total number of Reads of the bacterial genus.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.
Effects of Lactobacillus reuteri and Streptomyces coelicolor on Growth Performance of Broiler Chickens
Sarayu Bhogoju
et al.
Microorganisms,
2021
Combination of Bacillus licheniformis and Salinomycin: Effect on the Growth Performance and GIT Microbial Populations of Broiler Chickens
Jacek Trela
et al.
Animals,
2020
Impact of Dietary Sodium Butyrate and Salinomycin on Performance and Intestinal Microbiota in a Broiler Gut Leakage Model
Mohammad Naghizadeh
et al.
Animals,
2022



MDPI Initiatives
Important Links
© 2024 MDPI (Basel, Switzerland) unless otherwise stated