
Submitted:
12 May 2024
Posted:
13 May 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Metabolic-associated fatty liver disease (MAFLD) is a risk factor for severe COVID-19. This study explores the potential influence of gut hormone receptor and immune response gene expression on COVID-19 outcomes in MAFLD patients. Methods: We investigated gene expression levels of AHR, FFAR2, FXR, and TGR5 in patients with MAFLD and COVID-19 compared to controls. We examined associations between gene expression and clinical outcomes. Results: COVID-19 patients displayed altered AHR expression, potentially impacting immune response and recovery. Downregulated AHR in MAFLD patients correlated with increased coagulation parameters. Elevated FFAR2 expression in MAFLD linked to specific immune cell populations and hospital stay duration. Significantly lower FXR expression was observed in both MAFLD and severe COVID-19. Conclusion: Our findings suggest potential modulatory roles for AHR, FFAR2, and FXR in COVID-19 and MAFLD.
Keywords:
COVID-19
; MAFLD
; AHR
; FXR
; Gene expression
1. Introduction
Since the initial detection of the Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in late 2019, the virus has had a significant global impact, with over 700 million confirmed cases and 7 million deaths attributed to COVID-19 [1]. Identifying factors that influence susceptibility to SARS-CoV-2 infection and understanding how pre-existing medical conditions (comorbidities) affect disease severity remain crucial areas of contemporary research [2]. These insights hold significant promise for the development of improved treatment strategies [3,4].
There is a substantial body of evidence demonstrating a high prevalence of metabolic and vascular disorders among individuals who succumb to COVID-19, with these comorbidities accounting for up to 50% of fatalities [5]. Several studies have specifically highlighted Metabolic Associated Fatty Liver Disease (MAFLD) as a potential independent risk factor for both SARS-CoV-2 acquisition and hospitalization during COVID-19 illness, even after controlling for other aspects of metabolic syndrome [6,7,8]. Furthermore, research suggests that MAFLD may contribute to altered gene expression patterns, potentially influencing the course of COVID-19 [9,10].
Within the realm of immunology, the aryl hydrocarbon receptor (AHR) is a ligand-activated transcription factor classified as a nuclear receptor [11]. This widely expressed protein is found in both innate and adaptive immune cells. HR plays a multifaceted role in physiological processes, including the regulation of immune responses, inflammatory pathways, and environmental adaptations [12].
Activation of indoleamine 2,3-dioxygenase 1 (IDO1) by immune cells leads to the release of kynurenine (Kyn), a tryptophan (Trp) metabolite. Functioning as an endogenous ligand, Kyn activates AHR, thereby potentially hindering protective immunity [13]. Studies have demonstrated that AHR activation suppresses the initiation of influenza virus-specific CD8+ T cells within the lung compartment [14]. Interestingly, metabolomic analysis of COVID-19 patients revealed a similar alteration in Trp metabolism, favoring the Kyn pathway [15].
Building on this, Giovannoni et al. (2021) proposed a potentially novel therapeutic approach for SARS-CoV-2 infection: AHR modulation [16]. Their research suggests that SARS-CoV-2 infection itself activates AHR, potentially promoting viral replication. In dendritic cells (DCs), AHR activity downregulates the expression of Major Histocompatibility Complex II (MHC II) molecules. Furthermore, AHR regulates the production of inflammatory cytokines such as IL-6, IL-12, IL-15, and IL-18, which are typically produced during DC differentiation [17].
Continuing the exploration of the AHR within the context of COVID-19 and MAFLD, research suggests a potential proviral and profibrotic role for AHR. Constitutive activation of AHR (CA-AHR) has been demonstrated to inhibit mitochondrial β-oxidation, increase adipose triglyceride lipase activity, decrease white adipose tissue fat mass, and promote hepatic oxidative stress [18]. Furthermore, studies employing transgenic mice with constitutively active human AHR exposed to a high-fat diet exhibited exacerbated steatosis, highlighting a potential link between AHR and hepatic fat accumulation [19]. Conversely, AHR knockout (KO) mice displayed improved insulin sensitivity and glucose tolerance on a standard chow diet [20]. Moreover, these AHR-deficient mice were protected from high-fat diet-induced steatosis, obesity, and inflammation.
The cumulative findings from these studies raise intriguing questions regarding the potential role of AHR as a proviral host factor in SARS-CoV-2 replication and MAFLD progression.
Previously classified as orphan receptors, FFARs are now recognized as G protein-coupled receptors (GPCRs) that mediate metabolic signaling in response to fatty acid (FA) activation. Obesity, type 2 diabetes mellitus (T2DM), and MAFLD are closely linked conditions, each contributing to the metabolic syndrome phenotype [21].
Among FFARs, FFAR2 demonstrates particularly widespread expression in adipocytes, enteroendocrine cells, pancreatic β-cells, and various immune cells such as macrophages and neutrophils [22,23]. his receptor plays a regulatory role in both lipid metabolism and glucose levels through its influence on hormone secretion and inflammatory processes [24]. While FFAR2 is not directly expressed in hepatocytes, its ability to modulate the intestinal microbiota inflammasome may hold implications for MAFLD progression [25]. his effect is likely attributable to the FFAR2 pathway's capacity to suppress inflammatory cytokines and mitigate chronic inflammation, both of which are beneficial for metabolic liver disease [26].
Shifting focus to COVID-19, the cytokine storm observed in this disease is triggered by the release of interleukin-6 (IL-6) from alveolar macrophages [27]. Interestingly, FFAR2 appears to play a role in regulating probiotic activity, which has been shown to reduce IL-6 levels in COVID-19 patients [28,29].
Further connecting the dots between bile acid metabolism and both COVID-19 and MAFLD, we can explore the roles of Farnesoid-X-receptors (FXR) and G protein bile acid-activated receptor 1 (GPBAR-1), also known as Takeda G-protein-coupled receptor 5 (TGR5). These represent the two most well-characterized receptors within the bile acid-activated receptor (BAR) family [30]. Interestingly, these receptors are also expressed at high levels in cells of the innate immune system, including monocytes/macrophages, dendritic cells (DCs), natural killer (NK) cells, and NKT cells [31,32,33].
Similar to FXR, TGR5 appears to exert counter-regulatory effects on the immune response. Activation of TGR5 promotes a shift in colonic macrophages from a pro-inflammatory M1 phenotype to an anti-inflammatory M2 phenotype [34]. Furthermore, TGR5 suppresses the expression of pro-inflammatory cytokines such as IFN-γ, IL-1β, IL-6, and TNF-α, while inducing the production of the anti-inflammatory cytokine IL-10 [35].
Clinical studies have revealed alterations in bile acid metabolism (specifically, the composition of bile acids) in COVID-19 patients as the disease progresses from non-severe/mild to severe stages [36,37]. Intestinal dysbiosis, an imbalance in the gut microbiome, is another common feature observed in COVID-19 patients [38]. The presence of SARS-CoV-2 RNA in fecal samples from infected individuals suggests the virus's potential to replicate within human enterocytes. This viral replication within the gut may contribute to the disruption of bile acid metabolism and diarrhea, a frequent symptom in COVID-19 [39].
A recent study by Stutz et al. (2022) suggests that elevated levels of fecal secondary bile acids are associated with improved clinical outcomes in COVID-19 patients [40]. This finding can be explained by the immunosuppressive activity of CD4+ regulatory T cells (Tregs). Deconjugated bile acids appear to influence these Tregs, leading to an increase in their numbers. Additionally, the action of these bile acids on DCs has been shown to reduce their immunostimulatory properties [41].
The cellular entry of SARS-CoV-2 is facilitated by the spike protein binding to the angiotensin-converting enzyme 2 (ACE2) receptor. Interestingly, BAs may influence ACE2 expression in various tissues through their interaction with FXR and GPBAR1 [42]. Brevini et al. demonstrated that biliary organoids cultured in the presence of cholic acid (CDCA), a primary bile acid, expressed ACE2. Conversely, the removal of CDCA from the culture medium resulted in a reduction of ACE2 expression [43]. Similarly, BA regulation of ACE2 via GPBAR1 has also been documented. A study by Biagioli et al. (2022) showed that in vivo activation of GPBAR1 increased the production of glucagon-like peptide-1 (GLP-1) in intestinal L cells, which in turn promoted colonic cell expression of ACE2 [44]. These findings suggest that the potential protective effects of bile acid receptors in COVID-19 patients may be mediated through modulation of ACE2 expression alongside influencing the immune response.
For patients with MAFLD, activation of bile acid receptors appears to play a beneficial role in reducing disease severity. FXR activation has been shown to decrease steatosis by inhibiting lipogenesis, reduce chemically induced hepatic inflammation and fibrosis, and maintain intestinal barrier integrity, thereby protecting the liver from inflammatory signals originating from gut bacteria [45,46]. Similarly, TGR5 activation has been demonstrated to mitigate high-fat diet-induced glucose intolerance, insulin resistance, and inflammation, while also protecting against lipopolysaccharide-induced inflammation [47,48].
Taken together, these findings suggest that AHR, FFAR2, FXR, and TGR5 may play significant roles in the progression of both COVID-19 and MAFLD. Targeting these genes represents a promising therapeutic strategy that has the potential to improve patient outcomes and reduce disease severity. The current study aims to investigate the specific roles of these genes in the co-morbid course of COVID-19 and MAFLD.
2. Materials and Methods
2.1. Sample Collection
This investigation recruited participants from Ternopil City Community Hospital №1. Following informed consent through a signed statement, blood samples were collected and stored at -80 °C until analysis. All procedures adhered to the Declaration of Helsinki and received ethical approval from the I. Horbachevsky Ternopil National Medical University Ethics Committee (protocol №74, dated October 13, 2023).
The study population included individuals of European ancestry (Ukrainian ethnicity) aged between 23 and 86 years. Participants were hospitalized between October 2022 and May 2023. Inclusion criteria comprised: confirmed COVID-19 diagnosis requiring hospitalization, no history of chronic diseases, and no antibiotic or probiotic use within the preceding 3 months. Exclusion criteria encompassed: pre-enrollment corticosteroid use; active serious bacterial infection upon admission; chronic liver disease; pregnancy; alcohol dependence; and HIV infection.
According to the National Institutes of Health (NIH) guidelines [49], COVID-19 severity was categorized into moderate, severe, and critical subgroups.
MAFLD diagnosis was established using the following criteria: presence T2DM, overweight/obesity, or demonstrable evidence of metabolic syndrome [50,51]. The hepatic steatosis index (HSI) was employed to evaluate the presence of hepatic steatosis. This scoring system incorporates body mass index, liver enzymes, and presence of diabetes to estimate the likelihood of liver fat accumulation [52].
2.2. Laboratory and Clinical Data
During the study, a comprehensive set of laboratory tests were performed as part of the routine clinical workup. This analysis encompassed hematological parameters including white blood cell count with differential, erythrocyte sedimentation rate, hematocrit, and platelet count. Coagulation parameters assessed were international normalized ratio (INR), prothrombin time (PT), activated partial thromboplastin time (aPTT), and fibrinogen. Liver function was evaluated through measurement of total bilirubin, alanine aminotransferase (ALT), and aspartate aminotransferase (AST). Renal function was assessed by measuring creatinine levels. Markers of cholestasis, specifically gamma-glutamyl transferase (GGT), were also included in the laboratory panel. C-reactive protein (CRP) served as an inflammatory marker. Finally, blood glucose levels were measured for all participants. Body mass index (BMI) was documented for each individual.
2.3. Gene Expression Analysis
2.3.1. RNA Extraction and cDNA Synthesis
Total RNA was extracted from the collected blood samples using a standard protocol using NucleoZOL (740404.200, Düren, Germany). The extracted RNA was dissolved in RNase-free water to obtain a concentration of 2 µg/µL.
Total RNA was isolated from collected blood samples using a commercially available NucleoZOL reagent (740404.200, Düren, Germany) following a standardized protocol. Extracted RNA was eluted in RNase-free water to achieve a final concentration of 2 µg/µL. Complementary DNA (cDNA) synthesis was subsequently performed using a RevertAid First Strand cDNA Synthesis Kit (K1621, Vilnius, Lithuania) according to the manufacturer's instructions.
2.3.2. Real-Time PCR Amplification
A Bio-Rad CFX 96 Real-Time PCR Detection System (185-5096, Bio-Rad, USA) was employed to quantify the expression levels of four target genes: AHR, FFAR2, FXR, and TGR5. Maxima SYBR Green/ROX qPCR Master Mix (2×) (K0221, Thermo Scientific) and gene-specific primers were utilized for the amplification reaction. Each reaction mixture contained 20 µL nuclease-free water, 0.5 µL of each forward and reverse primer, 2 µL of cDNA template, and 10 µL of 2× Maxima SYBR Green/ROX qPCR Master Mix. The PCR cycling conditions consisted of an initial denaturation step at 95 °C for 10 minutes, followed by 45 cycles of denaturation at 95 °C for 15 seconds, primer annealing at 60 °C for 40 seconds, and extension at 72 °C for 40 seconds.
The housekeeping gene Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was selected for normalization of target gene expression levels. The comparative Ct (2−ΔΔCt) method was employed to quantify the relative expression of target genes (AHR, FFAR2, FXR, and TGR5) normalized to the housekeeping gene. Ct values were first converted to relative expression values using a formula that compares the Ct value of the target gene to the Ct value of the housekeeping gene. These relative expression values were then transformed into Log2 values using the formula Log2 (relative expression).
2.4. Statistical Analysis
Patient characteristics and clinical data were rigorously assessed and presented using descriptive statistics. The Shapiro-Wilk test was employed to evaluate the normality of the data distribution. Given the absence of normality, medians and interquartile ranges were calculated for all variables to summarize their central tendency and dispersion.
Due to the non-normal distribution of the data, non-parametric statistical tests were utilized for subsequent analyses. The Mann-Whitney U test was implemented for comparisons between two independent groups. For comparisons involving three or more groups, the Kruskal-Wallis test, a non-parametric alternative to one-way ANOVA, was employed. Dunn's multiple comparison test was then conducted for post-hoc pairwise comparisons between groups.
All statistical tests were two-tailed with a significance level set at p-value less than 0.05. Spearman's rank correlation coefficient was calculated to assess the relationships between continuous variables within a correlation matrix. Principal component analysis (PCA) was implemented to identify factors associated with COVID-19 severity and the presence of MAFLD.
Statistical analyses were performed using commercially available software programs, including GraphPad Prism (version 8.4.3) and IBM SPSS Statistics (version 25).
3. Results
3.1. Сomparing Group Expression
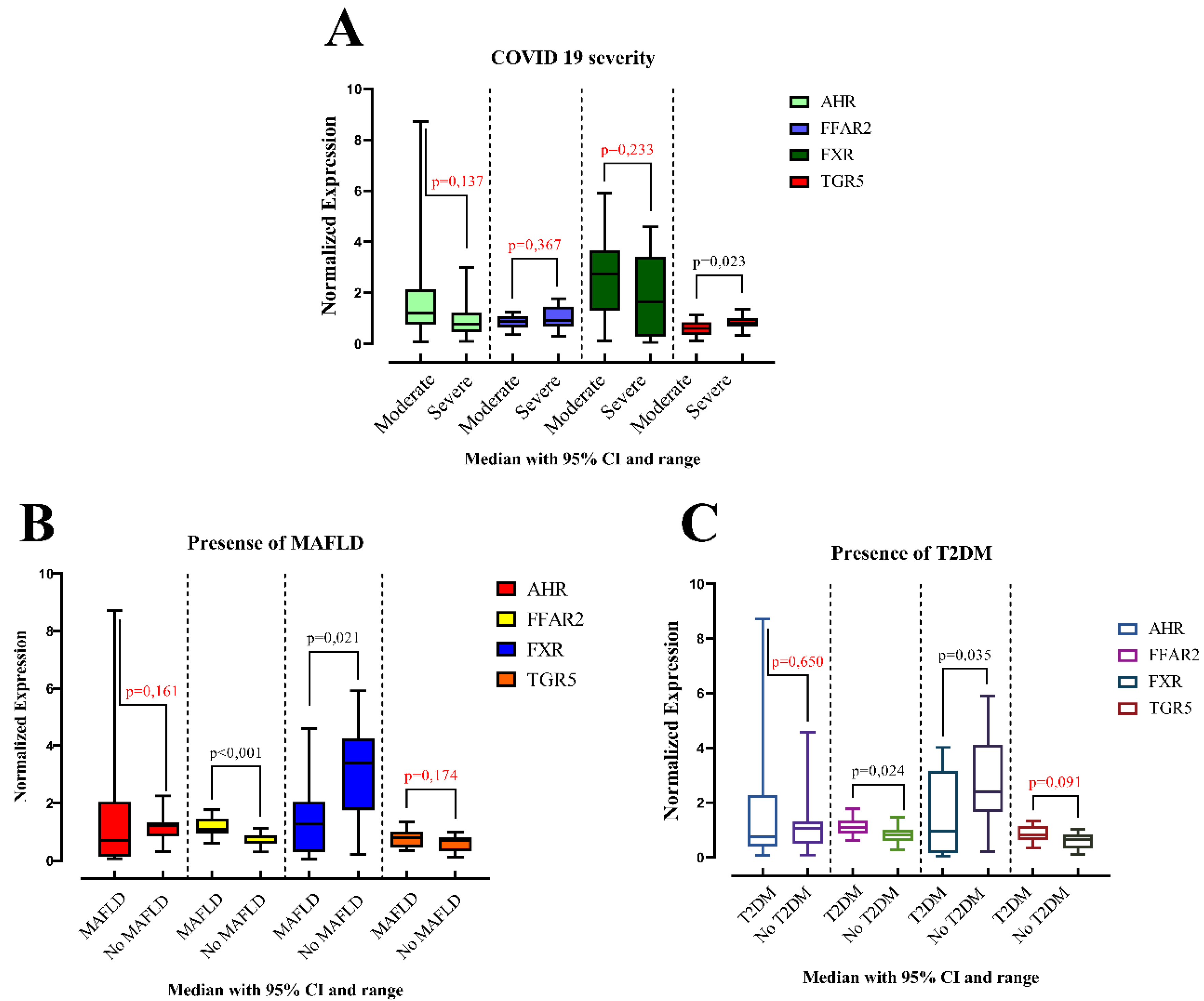
Our investigation into gene expression revealed no significant differences attributable to sex (male vs. female). However, compared to control subjects, individuals with MAFLD displayed a statistically significant decrease in FXR expression (p = 0.021) and a significant increase in FFAR2 expression (p < 0.001) (Figure 1). Expression levels of AHR and TGR5 remained unchanged in this patient group.
An analysis of COVID-19 severity revealed significantly lower TGR5 expression (p = 0.023) in patients with severe disease compared to those with moderate illness. Expression of AHR, FFAR2, and FXR did not exhibit statistically significant differences between these groups (Figure 1).
Interestingly, AHR expression was significantly lower in subjects with pneumonia (p = 0.001) compared to controls. Gene expression levels for all four genes were not significantly different between obese and non-obese subjects. Similarly, no significant differences were observed in gene expression for arterial hypertension or coronary heart disease.
Furthermore, gene expression levels for all four genes (AHR, FFAR2, FXR, and TGR5) were not statistically different between obese and non-obese subjects.
In contrast, individuals with T2DM displayed a significant increase in FFAR2 expression (p = 0.024) and a significant decrease in FXR expression (p = 0.035) compared to controls (Figure 1). Expression of AHR and TGR5 did not show statistically significant differences in this population. A detailed presentation of this data is provided in Table 1.
FFAR2 expression appears to be elevated in MAFLD and T2DM, while FXR expression is lowered. TGR5 expression is reduced in severe COVID-19 compared to moderate cases. AHR expression is lower in subjects with pneumonia. No significant associations were found between gene expression and obesity, arterial hypertension, or coronary heart disease.
Figure 1.
Comparison of the medians of two groups among patients: (A) with moderate and severe COVID-19; (B) with or without MAFLD; (C) with or without T2DM. Data are presented as medians for each group. Statistical comparisons were performed using the Mann-Whitney U test.
Figure 1.
Comparison of the medians of two groups among patients: (A) with moderate and severe COVID-19; (B) with or without MAFLD; (C) with or without T2DM. Data are presented as medians for each group. Statistical comparisons were performed using the Mann-Whitney U test.
We further investigated how gene expression levels within the MAFLD group affected clinical outcomes (Table 2). Patients were stratified based on down-regulated, unchanged, or up-regulated expression of specific genes.
In the non-MAFLD group, patients with down-regulated FXR gene expression exhibited altered laboratory values upon admission compared to those with unchanged FXR expression. They had significantly higher levels of band neutrophils (p = 0.045), INR (p = 0.038), PT (p = 0.021), and GGT (p = 0.008). Interestingly, these patients also presented with higher albumin levels at both admission (p = 0.045) and discharge (p = 0.004).
Conversely, patients with down-regulated FFAR2 gene expression displayed lower albumin levels at discharge (p = 0.018) compared to the unchanged FFAR2 group.
Within the MAFLD group, patients with up-regulated FFAR2 expression demonstrated a longer length of hospital stay (p = 0.001), higher leukocyte levels on admission (p = 0.010), and lower ALP (alkaline phosphatase) levels on admission and discharge (p < 0.02) compared to patients with unchanged FFAR2 expression. Patients with down-regulated FXR expression displayed opposite trends: shorter hospital stay (p = 0.033) and higher ALP levels on admission (p = 0.006) and discharge (p = 0.017) compared to those with unchanged FXR expression.
Additionally, patients with down-regulated AHR expression presented with elevated INR and PT levels upon admission compared to patients with unchanged and up-regulated AHR expression (p-values < 0.05).

Table 2.
Difference in laboratory outcomes in patients with downregulated, no change and up regulated gene expression.
Table 2.
Difference in laboratory outcomes in patients with downregulated, no change and up regulated gene expression.
| No-MAFLD cohort | ||||||
| Regulation of FXR gene expression | ||||||
| Down regulated (n = 5) | No change (n = 10) | p-Value a | ||||
| Band neutrophils, % (admission) | 9 (8.5–18) | 6 (4–10) | p = 0.045 | |||
| INR *, n (admission) | 1.1 (1.06–1.38) | 0.96 (0.865–1.06) | p = 0.038 | |||
| PT *, sec (admission) | 14.2 (13.5–16.6) | 12.5 (11.4–13.2) | p = 0.021 | |||
| GGT *, unit/l (admission) | 67 (49–127) | 31(19.1–42.5) | p = 0.008 | |||
| Albumin, g/l (admission) | 40 (35–40.5) | 45 (40–59.5) | p = 0.045 | |||
| Albumin, g/l (discharge) | 37 (33.5–40.5) | 47.5 (42.5–50) | p = 0.004 | |||
| Regulation of FFAR2 gene expression | ||||||
| No change (n = 12) | Down regulated (n = 3) | p-Value a | ||||
| Albumin, g/l (discharge) | 46 (41.5–49) | 35 (32–40) | p = 0.018 | |||
| Mafld cohort | ||||||
| Regulation of FFAR2 gene expression | ||||||
| No change (n = 11) | Up regulated (n = 4) | p-Value a | ||||
| Length of hospital stay, days | 11 (10–13) | 16.5 (15.3–17) | p = 0.001 | |||
| Leukocytes, 109/L (admission) | 7 (6.59–8.97) | 11.3 (10.4–14.6) | p = 0.010 | |||
| ALP *, mmol/L (admission) | 132 (107–271) | 89.5 (80.8–96.8) | p = 0.006 | |||
| ALP *, mmol/L (discharge) | 132 (106–195) | 90.5 (79–112) | p = 0.017 | |||
| Regulation of FXR gene expression | ||||||
| Down regulated (n = 12) | No change (n = 3) | p-Value a | ||||
| Length of hospital stay, days | 11.5 (10–13.8) | 17 (13–17) | p = 0.033 | |||
| ALP *, mmol/L (admission) | 130 (100–246) | 86 (79–103) | p = 0.031 | |||
| Regulation of AHR gene expression | ||||||
| Down regulated (n = 4) | No change (n = 8) | Up regulated (n = 3) | p-Value b | |||
| INR *, n (admission) | 1.17 (1.06–1.66) | 0.98 (0.9–1) | 0.99 (0.76–1.14) | p = 0.018 | ||
| PT *, sec (admission) | 14.8 (13.5–19.4) | 12.4(11.6–13.1) | 12.4 (10.12–14.9) | p = 0.033 | ||
a Mann Whitney test; b Kruskal-Wallis test. INR—International Normalized Ratio; PT—Prothrombin time; GGT—Gamma-glutamyltransferase; ALP—Alkaline phosphatase.
3.2. Relative Expression of AHR, FFAR2, FXR and TGR5 in COVID-19 Patients with and without MAFLD
This study investigated the messenger RNA (mRNA) expression levels of four genes (AHR, FFAR2, FXR, and TGR5) in patients with COVID-19, stratified by the presence or absence of metabolically associated fatty liver disease (MAFLD) (Figure 2). To account for potential confounding factors, relative normalized expression was calculated using the PCR method. Two control groups were employed: a group without COVID-19 for the COVID-19 without MAFLD group and a group with COVID-19 but without MAFLD for the COVID-19 with MAFLD group.
An initial analysis of gene expression was conducted in all COVID-19 patients (Figure 2). In the group without MAFLD (Figure 2, Panel A), a distinct pattern emerged. Fifteen patients displayed upregulation of the AHR gene, while TGR5 was downregulated in the same patients. Additionally, three and five patients from this group exhibited downregulation of FFAR2 and FXR genes, respectively.
Conversely, the group with MAFLD (Figure 2, Panel B) showed a different expression profile. Here, the FXR gene was downregulated in 12 patients, while the FFAR2 gene was upregulated in 4 patients. Interestingly, the AHR gene expression was mixed in this group, with upregulation observed in 3 patients and downregulation in 4 patients.
Volcano plot analysis was employed to further refine the findings by identifying statistically significant changes in gene expression (Figure 2, Panels C and D). This analysis takes into account both the fold-change and statistical significance of gene expression differences.
Within the group without MAFLD, significant differences were observed in a limited number of patients. Ten patients displayed significant up-regulation of AHR, while three and four patients showed significant down-regulation of FFAR2 and FXR, respectively.
The MAFLD group exhibited a more distinct expression pattern. Twelve patients displayed significant down-regulation of the FXR gene. Only two patients showed significant up-regulation of the AHR gene, while four patients had significant up-regulation of the FFAR2 gene. One patient in the MAFLD group displayed significant up-regulation of the TGR5 gene.
Compared to the control group, patients with COVID-19 exhibited significant changes in gene expression. There was a 21.3-fold up-regulation (CI: 16.0-26.5) of the AHR gene, a 1.85-fold down-regulation (CI: 1.42-2.28) of the FFAR2 gene, and a 15.7-fold down-regulation (CI: 1.42-2.28) of TGR5 gene.
Furthermore, the presence of MAFLD in COVID-19 patients resulted in a distinct expression pattern compared to controls without MAFLD. Patients with MAFLD displayed a 1.69-fold up-regulation (CI: 1.21-2.18) of the FFAR2 gene and an 11.7-fold downregulation (CI: 1.11-22.2) of the FXR gene compared to the control group.
3.3. Correlation Analysis of Genes Normalized Expression
This section explores the relationships between gene expression levels and various clinical parameters in COVID-19 patients without MAFLD. The analysis employed Spearman's rank correlation coefficient (r) to assess the strength and direction of the associations.
The normalized expression level of the AHR gene exhibited positive correlations with several parameters. These included SpO2 (admission oxygen saturation, r = 0.58, p = 0.001), general protein levels at both admission (r = 0.72, p = 0.003) and discharge (r = 0.73, p = 0.003), and albumin level at admission (r = 0.53, p = 0.043). Conversely, AHR expression showed negative correlations with APTT (activated partial thromboplastin time) at both admission (r = -0.52, p = 0.049) and discharge (r = -0.53, p = 0.042).
The normalized expression level of the FFAR2 gene displayed a positive correlation with the normalized expression level of the TGR5 gene (r = 0.56, p = 0.032). Additionally, it correlated positively with band neutrophils (r = 0.52, p = 0.049) and segmented neutrophils (r = 0.53, p = 0.044) at admission. However, FFAR2 expression exhibited a negative correlation with SpO2 at discharge (r = -0.80, p = 0.001), monocytes at admission (r = -0.71, p = 0.004), and ESR (erythrocyte sedimentation rate) at discharge (r = -0.60, p = 0.020).
The normalized expression level of the FXR gene showed a positive correlation with platelet level at admission (r = 0.55, p = 0.038). Conversely, it exhibited negative correlations with band neutrophils (r = -0.60, p = 0.021) at admission, PT (prothrombin time) at admission (r = -0.57, p = 0.030), and GGT at admission (r = -0.56, p = 0.034).
The normalized expression level of the TGR5 gene displayed a positive correlation with length of hospital stay (r = 0.66, p = 0.009) and QPT level at admission (r = 0.53, p = 0.045). However, it showed a negative correlation with SpO2 at discharge (r = -0.55, p = 0.034).
Figure 3.
Spearman correlation correlogram used for correlations between continuous data in COVID-19 patients with (A) without (B) MAFLD. Red: Strong negative correlation (r = -1.0). Blue: Strong positive correlation (r = 1.0).
Figure 3.
Spearman correlation correlogram used for correlations between continuous data in COVID-19 patients with (A) without (B) MAFLD. Red: Strong negative correlation (r = -1.0). Blue: Strong positive correlation (r = 1.0).
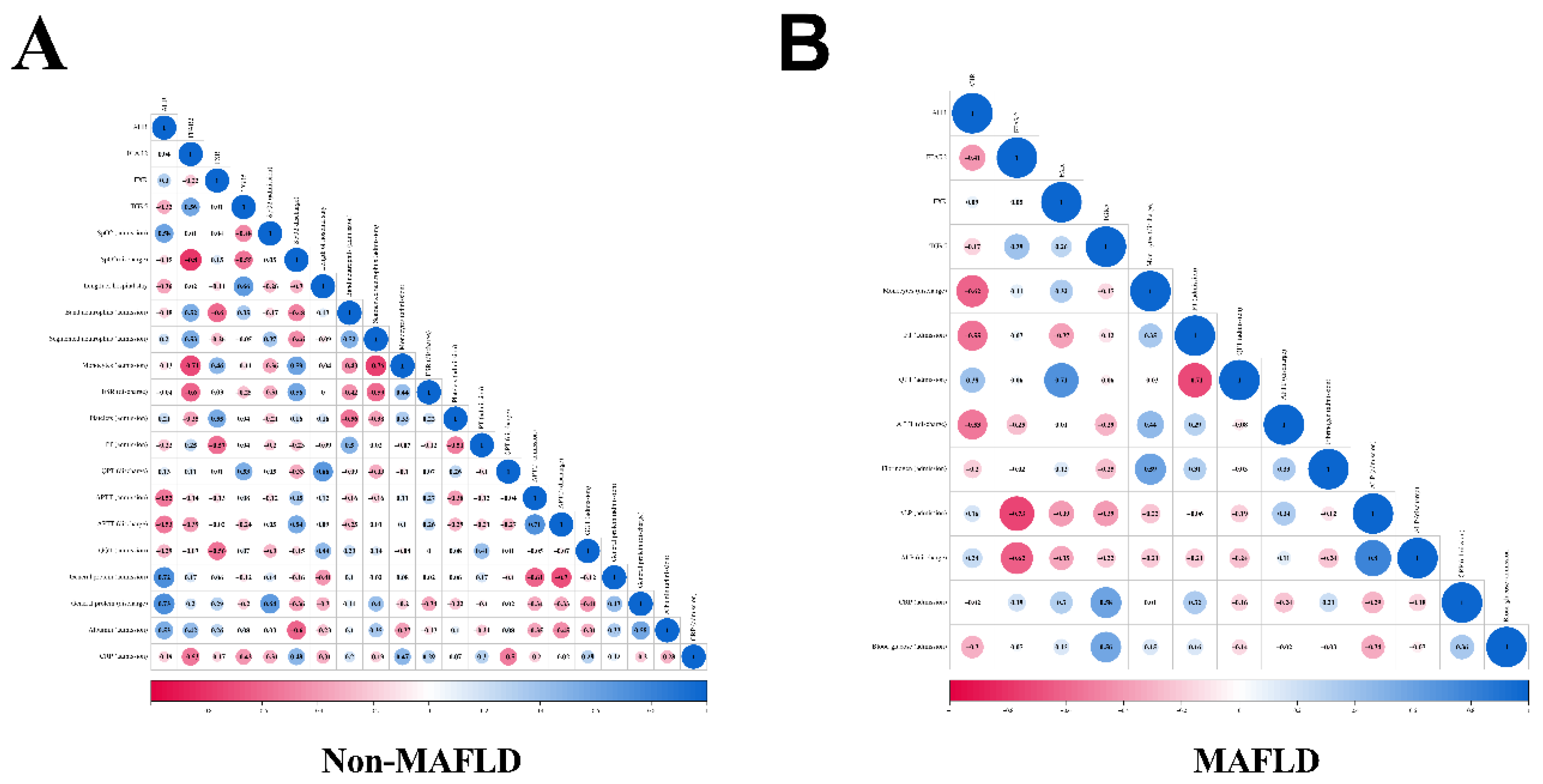
Next we analyzed relationships between gene expression levels and clinical parameters in COVID-19 patients with MAFLD.
The normalized expression level of the AHR gene exhibited negative correlations with monocyte count at discharge (r = -0.62, p = 0.016), PT at admission (r = -0.55, p = 0.0437), and APTT at admission (r = -0.53, p = 0.044).
The normalized expression level of the FFAR2 gene displayed a negative correlation with ALP (alkaline phosphatase) levels at both admission (r = -0.73, p = 0.003) and discharge (r = -0.62, p = 0.016).
The normalized expression level of the FXR gene showed a positive correlation with QPT (quick prothrombin time) at admission (r = 0.71, p = 0.004).
The normalized expression level of the TGR5 gene displayed positive correlations with CRP (C-reactive protein) at admission (r = 0.58, p = 0.026) and blood glucose levels (r = 0.56, p = 0.030).
3.4. Principal Component Analysis
We employed principal component analysis (PCA) to explore the underlying factors contributing to COVID-19 severity and the presence of MAFLD (Figure 4).
PCA identified two principal components (PCs) for COVID-19 severity, explaining a total of 74.8% of the variance (56.4% and 18.3% for PC1 and PC2, respectively). The Kaiser-Meyer-Olkin (KMO) measure of sampling adequacy was 0.747, indicating a suitable correlation matrix for PCA. Bartlett's test of sphericity yielded a significance level of p < 0.001, further supporting the applicability of PCA.
Figure 4.
Principal component analysis: (A) COVID-19 severity; (B) presence of MAFLD.
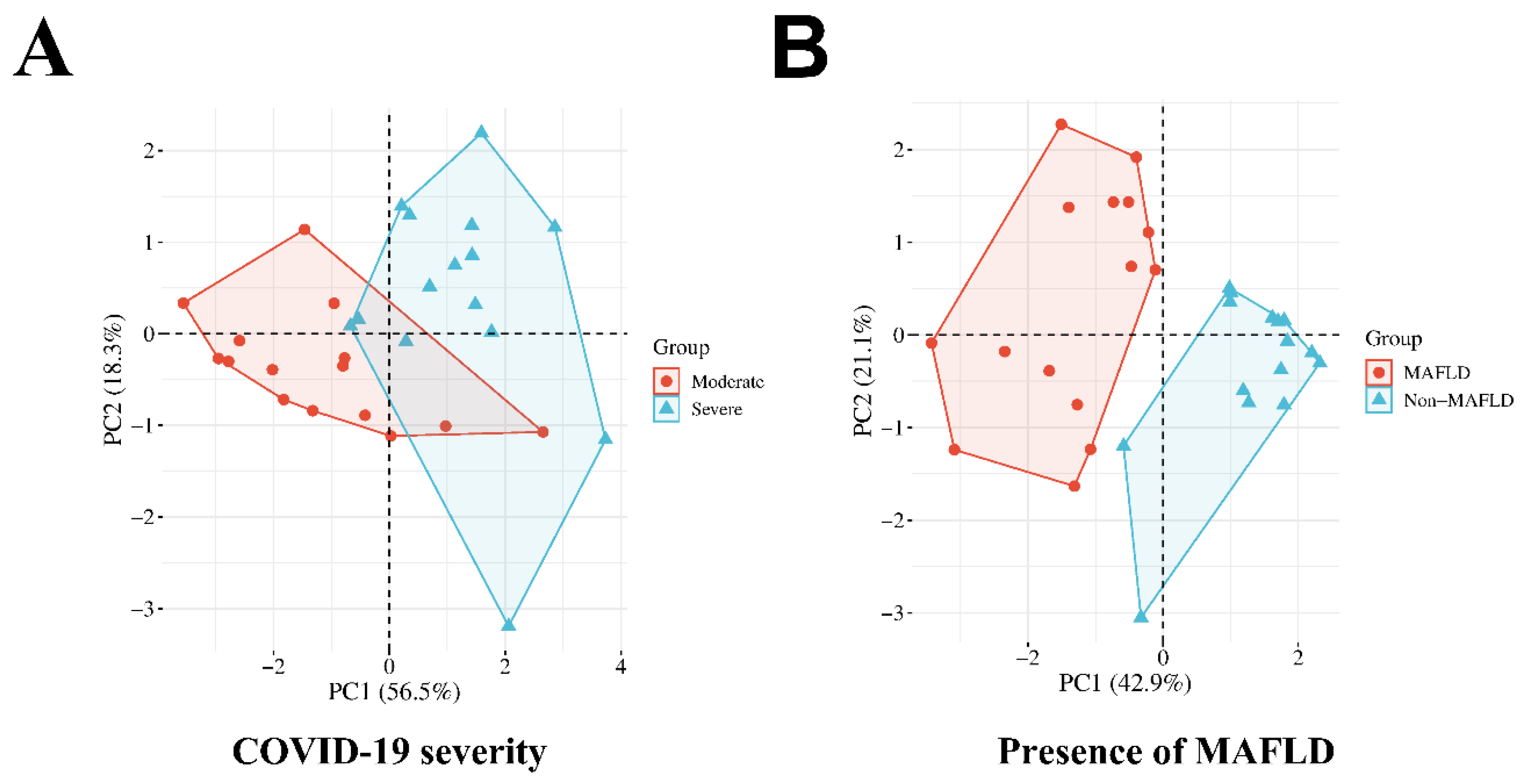
PC1 encompassed length of hospital stay, neutrophil-to-lymphocyte ratio (NLR), and lymphocyte count (discharge). PC2 comprised community-acquired pneumonia (CAP) status and oxygen saturation (SpO2) upon admission. Segmented neutrophils (discharge) were excluded due to cross-loadings on both PCs, hindering a clear attribution to either component (Table 3).
PCA identified two PCs for the presence of MAFLD, accounting for 64% of the total variance (42.9% and 21.1% for PC1 and PC2, respectively). The KMO measure was 0.588, signifying an adequate correlation matrix for PCA. Bartlett's test resulted in a significance level of p < 0.001, supporting the use of PCA.
PC1 included body mass index (BMI), normalized expression levels of FFAR2 and FXR, and the presence of T2DM. PC2 was constituted by SpO2 (admission) and the requirement for supplemental oxygen therapy.
Table 4.
Rotated Component Matrix for presence of MAFLD.
| Component | ||
| Factor 1 | Factor 2 | |
| BMI | 0.724 | 0.129 |
| FFAR2 Normalized Expression | 0.693 | 0.233 |
| FXR Normalized Expression | -0.635 | -0.085 |
| SpO2 (admission) | -0.18 | -0.914 |
| T2DM | 0.768 | 0.062 |
| The need for oxygen supply | 0.13 | 0.938 |
Extraction Method: Principal Component Analysis. Rotation Method: Varimax with Kaiser Normalization (significant loadings in bold). Rotation converged in 5 iterations.
4. Discussion
Our investigation explored potential associations between gene expression of AHR, FFAR2, FXR, and TGR5, and COVID-19 outcomes in patients with MAFLD. Prior research demonstrating MAFLD as a risk factor for severe COVID-19 presentations and ICU admissions [6,7]. However, the impact of MAFLD on mortality rates remains inconclusive, and explaining their interaction is still quite difficult. Elucidating the interplay between these factors poses a significant challenge. In this context, analyzing gene expression levels holds promise in uncovering novel insights into the combined course of COVID-19 and MAFLD.
Consistent with the AHR gene known role in regulating immune and inflammatory responses [12], our findings demonstrate altered AHR gene expression in COVID-19 patients. Giovannoni et al. (2021) previously reported AHR activation by coronaviruses and its impact on antiviral immunity in lung epithelial cells, further supporting this observation [16]. Our study corroborates these findings, with a 21.3-fold upregulation of AHR observed in COVID-19 patients compared to controls. Notably, AHR expression was also elevated in patients with pneumonia (p < 0.001), suggesting potential links to impaired local pulmonary immunity and increased susceptibility to infection. Interestingly, AHR level had positive correlations with SpO2 level at admission in COVID-19 patients.
While AHR expression remained largely unchanged in MAFLD patients, those with COVID-19 and MAFLD who displayed downregulated AHR exhibited higher INR and PT levels (coagulation parameters). These findings suggest a potential role for AHR in the complex interplay between COVID-19, MAFLD, and coagulopathy. However, further investigation is needed to elucidate the underlying mechanisms and potential therapeutic implications.
The FFAR2 gene, known to regulate lipid metabolism and glucose levels [24], may influence the course of MAFLD. While not directly expressed in liver cells (hepatocytes), FFAR2 is present in fat cells (adipocytes) and immune system cells like macrophages and neutrophils [22,23]. FFAR2 can modulate inflammation through both immune cell activation [24] and gut microbiota interactions [25]. Some studies suggest it can decrease the inflammatory marker IL-6 by regulating beneficial gut bacteria (probiotics) [28,29]. Our findings demonstrate higher normalized FFAR2 expression in MAFLD patients compared to controls, with similar observations in patients with type 2 diabetes mellitus (T2DM). However, no significant differences were observed between moderate and severe COVID-19 cases.
Within the MAFLD cohort, upregulated FFAR2 gene expression was associated with a longer hospital stay, higher white blood cell (leukocyte) count at admission, and lower alkaline phosphatase (ALP) levels. It's important to note that most patients did not exhibit upregulated FFAR2. In COVID-19 patients with MAFLD, FFAR2 expression showed positive correlations with segmented neutrophils and bands (immature neutrophils), indicating potential influence on specific immune cell populations. Interestingly, negative correlations were observed with monocytes. PCA analysis further identified FFAR2 as a factor associated with the presence of MAFLD.
Bile acids (BAs) have been proposed to modulate SARS-CoV-2 entry into host cells through FXR and GPBAR1 by regulating ACE2 expression in various tissues [42]. Additionally, BAs can suppress intestinal dendritic cell (DC) differentiation and activation via FXR [53,54], and regulate inflammatory responses through TGR5 activation, leading to decreased IFN-γ, IL-1β, IL-6, and TNF-α, and increased IL-10 [35]. FXR and TGR5 activation can also improve metabolic health by reducing steatosis (fatty liver) via inhibiting lipogenesis and decreasing hepatic inflammation [45], as well as by mitigating high-fat diet (HFD)-induced glucose intolerance and insulin resistance [46].
Our study revealed significantly lower normalized expression levels of FXR in patients with both MAFLD and T2DM, while TGR5 expression was lower in patients with severe COVID-19 compared to moderate cases. In COVID-19 patients, downregulated FXR expression correlated with higher levels of immature neutrophils (bands), INR (blood clotting parameter), PT (blood clotting parameter), and GGT (liver enzyme) upon admission, alongside a higher albumin level. Interestingly, within the MAFLD cohort, 12 out of 15 patients displayed a substantial (11.7-fold) downregulation of FXR, which was associated with a shorter hospital stay and a higher ALP level (another liver enzyme). Regarding TGR5, its normalized expression level in COVID-19 patients showed a positive correlation with hospital stay duration and a negative correlation with SpO2 (oxygen saturation) at discharge. Notably, changes in both FXR and TGR5 expression were identified as contributing factors associated with the presence of MAFLD in PCA analysis.
5. Limitations
Our study acknowledges several limitations. First, the relatively small sample size employed restricts the generalizability of our findings to a broader population. A larger, multicenter study would be necessary to confirm these observations and enhance their generalizability. Second, the monocentric design of this study inherently limits the population studied and may introduce selection bias. Ideally, future studies would incorporate participants from multiple centers to achieve a more representative sample.
6. Conclusions
This study investigated the relationships between gene expression of AHR, FFAR2, FXR, and TGR5, and COVID-19 outcomes in patients with MAFLD. Our findings highlight potential modulatory roles for these genes, particularly AHR and FXR, in both COVID-19 severity and MAFLD presentation. We observed altered expression of AHR in COVID-19 patients, potentially influencing immune responses and recovery. In MAFLD, downregulated AHR was associated with increased coagulation parameters. FFAR2 expression was elevated in MAFLD patients and correlated with specific immune cell populations and hospital stay duration. Interestingly, FXR expression was significantly lower in both MAFLD and severe COVID-19, potentially linking it to metabolic health and inflammatory processes. This study contributes to a growing understanding of the complex interplay between gene expression, MAFLD, and COVID-19 outcomes. By identifying potential gene targets, we pave the way for future research that could lead to improved clinical strategies for managing patients with these conditions.
Author Contributions
Conceptualization and writing—original draft preparation, M.B. and O.K.; writing—review and editing, O.B., V.O., and I.K.; supervision, O.K., and V.O.; project administration, V.O. and O.K.; visualization, I.K.; funding acquisition, M.B. and I.V. All authors have read and agreed to the published version of the manuscript.
Funding
RECOOP Grant #36—CSMC Senior Scientists (RCSS) “Comprehensive Analysis of Genetic Predictors for MAFLD Development in Patients with COVID-19”.
Institutional Review Board Statement
The study protocol met the requirements for biomedical research and was approved by the Local Ethics Committee of the I. Horbachevsky Ternopil National Medical University as protocol N74, dated 13 October2023.
Informed Consent Statement
All patients signed an informed consent for the study.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- COVID - Coronavirus Statistics - Worldometer. Available online: https://www.worldometers.info/coronavirus/ (accessed on 29 April 2024).
- Buchynskyi, M.; Oksenych, V.; Kamyshna, I.; Kamyshnyi, O. Exploring Paxlovid Efficacy in COVID-19 Patients with MAFLD: Insights from a Single-Center Prospective Cohort Study. Viruses 2024, 16, 112. [Google Scholar] [CrossRef] [PubMed]
- Buchynskyi, M.; Kamyshna, I.; Lyubomirskaya, K.; Moshynets, O.; Kobyliak, N.; Oksenych, V.; Kamyshnyi, A. Efficacy of Interferon Alpha for the Treatment of Hospitalized Patients with COVID-19: A Meta-Analysis. Front. Immunol. 2023, 14, 1069894. [Google Scholar] [CrossRef] [PubMed]
- Kamyshnyi, A.; Koval, H.; Kobevko, O.; Buchynskyi, M.; Oksenych, V.; Kainov, D.; Lyubomirskaya, K.; Kamyshna, I.; Potters, G.; Moshynets, O. Therapeutic Effectiveness of Interferon-A2b against COVID-19 with Community-Acquired Pneumonia: The Ukrainian Experience. Int. J. Mol. Sci. 2023, 24, 6887. [Google Scholar] [CrossRef] [PubMed]
- Steenblock, C.; Schwarz, P.E.H.; Ludwig, B.; Linkermann, A.; Zimmet, P.; Kulebyakin, K.; Tkachuk, V.A.; Markov, A.G.; Lehnert, H.; de Angelis, M.H.; et al. COVID-19 and Metabolic Disease: Mechanisms and Clinical Management. Lancet Diabetes Endocrinol. 2021, 9, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, P.J.; Váncsa, S.; Ocskay, K.; Dembrovszky, F.; Kiss, S.; Farkas, N.; Erőss, B.; Szakács, Z.; Hegyi, P.; Pár, G. Metabolic Associated Fatty Liver Disease Is Associated With an Increased Risk of Severe COVID-19: A Systematic Review With Meta-Analysis. Front. Med. 2021, 8, 626425. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Hussain, S.; Antony, B. Non-Alcoholic Fatty Liver Disease and Clinical Outcomes in Patients with COVID-19: A Comprehensive Systematic Review and Meta-Analysis. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Li, Y.; Cheng, B.; Zhou, T.; Gao, Y. Risk of Severe COVID-19 Increased by Metabolic Dysfunction-Associated Fatty Liver Disease: A Meta-Analysis. J. Clin. Gastroenterol. 2021, 55, 830. [Google Scholar] [CrossRef] [PubMed]
- Buchynskyi, M.; Oksenych, V.; Kamyshna, I.; Vari, S.G.; Kamyshnyi, A. Genetic Predictors of Comorbid Course of COVID-19 and MAFLD: A Comprehensive Analysis. Viruses 2023, 15, 1724. [Google Scholar] [CrossRef] [PubMed]
- Buchynskyi, M.; Kamyshna, I.; Oksenych, V.; Zavidniuk, N.; Kamyshnyi, A. The Intersection of COVID-19 and Metabolic-Associated Fatty Liver Disease: An Overview of the Current Evidence. Viruses 2023, 15, 1072. [Google Scholar] [CrossRef]
- Veldhoen, M.; Hirota, K.; Westendorf, A.M.; Buer, J.; Dumoutier, L.; Renauld, J.-C.; Stockinger, B. The Aryl Hydrocarbon Receptor Links TH17-Cell-Mediated Autoimmunity to Environmental Toxins. Nature 2008, 453, 106–109. [Google Scholar] [CrossRef]
- Neavin, D.R.; Liu, D.; Ray, B.; Weinshilboum, R.M. The Role of the Aryl Hydrocarbon Receptor (AHR) in Immune and Inflammatory Diseases. Int. J. Mol. Sci. 2018, 19, 3851. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.; Carbone, A.; Mazzoccoli, G. Aryl Hydrocarbon Receptor Role in Co-Ordinating SARS-CoV-2 Entry and Symptomatology: Linking Cytotoxicity Changes in COVID-19 and Cancers; Modulation by Racial Discrimination Stress. Biology 2020, 9, 249. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, B.P.; Roberts, A.D.; Neumiller, J.J.; Cundiff, J.A.; Woodland, D.L. Aryl Hydrocarbon Receptor Activation Impairs the Priming but Not the Recall of Influenza Virus-Specific CD8+ T Cells in the Lung. J. Immunol. 2006, 177, 5819–5828. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Stefanoni, D.; Reisz, J.A.; Nemkov, T.; Bertolone, L.; Francis, R.O.; Hudson, K.E.; Zimring, J.C.; Hansen, K.C.; Hod, E.A.; et al. COVID-19 Infection Alters Kynurenine and Fatty Acid Metabolism, Correlating with IL-6 Levels and Renal Status. JCI insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, F.; Li, Z.; Remes-Lenicov, F.; Dávola, M.E.; Elizalde, M.; Paletta, A.; Ashkar, A.A.; Mossman, K.L.; Dugour, A.V.; Figueroa, J.M.; et al. AHR Signaling Is Induced by Infection with Coronaviruses. Nat. Commun. 2021, 12, 5148. [Google Scholar] [CrossRef] [PubMed]
- Morelli, A.E.; Zahorchak, A.F.; Larregina, A.T.; Colvin, B.L.; Logar, A.J.; Takayama, T.; Falo, L.D.; Thomson, A.W. Cytokine Production by Mouse Myeloid Dendritic Cells in Relation to Differentiation and Terminal Maturation Induced by Lipopolysaccharide or CD40 Ligation. Blood 2001, 98, 1512–1523. [Google Scholar] [CrossRef]
- Lee, J.H.; Wada, T.; Febbraio, M.; He, J.; Matsubara, T.; Lee, M.J.; Gonzalez, F.J.; Xie, W. A Novel Role for the Dioxin Receptor in Fatty Acid Metabolism and Hepatic Steatosis. Gastroenterology 2010, 139, 653–663. [Google Scholar] [CrossRef]
- Lu, P.; Yan, J.; Liu, K.; Garbacz, W.G.; Wang, P.; Xu, M.; Ma, X.; Xie, W. Activation of Aryl Hydrocarbon Receptor Dissociates Fatty Liver from Insulin Resistance by Inducing Fibroblast Growth Factor 21. Hepatology 2015, 61, 1908–1919. [Google Scholar] [CrossRef]
- Wang, C.; Xu, C.-X.; Krager, S.L.; Bottum, K.M.; Liao, D.-F.; Tischkau, S.A. Aryl Hydrocarbon Receptor Deficiency Enhances Insulin Sensitivity and Reduces PPAR-α Pathway Activity in Mice. Environ. Health Perspect. 2011, 119, 1739–1744. [Google Scholar] [CrossRef]
- Lusis, A.J.; Attie, A.D.; Reue, K. Metabolic Syndrome: From Epidemiology to Systems Biology. Nat. Rev. Genet. 2008, 9, 819–830. [Google Scholar] [CrossRef]
- Kamp, M.E.; Shim, R.; Nicholls, A.J.; Oliveira, A.C.; Mason, L.J.; Binge, L.; Mackay, C.R.; Wong, C.H.Y. G Protein-Coupled Receptor 43 Modulates Neutrophil Recruitment during Acute Inflammation. PLoS One 2016, 11, e0163750. [Google Scholar] [CrossRef] [PubMed]
- McNelis, J.C.; Lee, Y.S.; Mayoral, R.; van der Kant, R.; Johnson, A.M.F.; Wollam, J.; Olefsky, J.M. GPR43 Potentiates β-Cell Function in Obesity. Diabetes 2015, 64, 3203–3217. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-Chain Fatty Acids Stimulate Glucagon-like Peptide-1 Secretion via the G-Protein-Coupled Receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Henao-Mejia, J.; Elinav, E.; Jin, C.; Hao, L.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J.; et al. Inflammasome-Mediated Dysbiosis Regulates Progression of NAFLD and Obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef]
- Liang, Y.; Liang, S.; Zhang, Y.; Deng, Y.; He, Y.; Chen, Y.; Liu, C.; Lin, C.; Yang, Q. Oral Administration of Compound Probiotics Ameliorates HFD-Induced Gut Microbe Dysbiosis and Chronic Metabolic Inflammation via the G Protein-Coupled Receptor 43 in Non-Alcoholic Fatty Liver Disease Rats. Probiotics Antimicrob. Proteins 2019, 11, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xie, J.; Zhao, L.; Fei, X.; Zhang, H.; Tan, Y.; Nie, X.; Zhou, L.; Liu, Z.; Ren, Y.; et al. Alveolar Macrophage Dysfunction and Cytokine Storm in the Pathogenesis of Two Severe COVID-19 Patients. EBioMedicine 2020, 57, 102833. [Google Scholar] [CrossRef]
- Pham, M.T.; Yang, A.J.; Kao, M.-S.; Gankhuyag, U.; Zayabaatar, E.; Jin, S.-L.C.; Huang, C.-M. Gut Probiotic Lactobacillus Rhamnosus Attenuates PDE4B-Mediated Interleukin-6 Induced by SARS-CoV-2 Membrane Glycoprotein. J. Nutr. Biochem. 2021, 98, 108821. [Google Scholar] [CrossRef]
- Kao, M.-S.; Yang, J.-H.; Balasubramaniam, A.; Traisaeng, S.; Jackson Yang, A.; Yang, J.J.; Salamon, B.P.; Herr, D.R.; Huang, C.-M. Colonization of Nasal Cavities by Staphylococcus Epidermidis Mitigates SARS-CoV-2 Nucleocapsid Phosphoprotein-Induced Interleukin (IL)-6 in the Lung. Microb. Biotechnol. 2022, 15, 1984–1994. [Google Scholar] [CrossRef]
- Kawamata, Y.; Fujii, R.; Hosoya, M.; Harada, M.; Yoshida, H.; Miwa, M.; Fukusumi, S.; Habata, Y.; Itoh, T.; Shintani, Y.; et al. A G Protein-Coupled Receptor Responsive to Bile Acids. J. Biol. Chem. 2003, 278, 9435–9440. [Google Scholar] [CrossRef]
- Mencarelli, A.; Renga, B.; Migliorati, M.; Cipriani, S.; Distrutti, E.; Santucci, L.; Fiorucci, S. The Bile Acid Sensor Farnesoid X Receptor Is a Modulator of Liver Immunity in a Rodent Model of Acute Hepatitis. J. Immunol. 2009, 183, 6657–6666. [Google Scholar] [CrossRef]
- Vavassori, P.; Mencarelli, A.; Renga, B.; Distrutti, E.; Fiorucci, S. The Bile Acid Receptor FXR Is a Modulator of Intestinal Innate Immunity. J. Immunol. 2009, 183, 6251–6261. [Google Scholar] [CrossRef] [PubMed]
- Biagioli, M.; Carino, A.; Fiorucci, C.; Marchianò, S.; Di Giorgio, C.; Bordoni, M.; Roselli, R.; Baldoni, M.; Distrutti, E.; Zampella, A.; et al. The Bile Acid Receptor GPBAR1 Modulates CCL2/CCR2 Signaling at the Liver Sinusoidal/Macrophage Interface and Reverses Acetaminophen-Induced Liver Toxicity. J. Immunol. 2020, 204, 2535–2551. [Google Scholar] [CrossRef] [PubMed]
- Fiorucci, S.; Carino, A.; Baldoni, M.; Santucci, L.; Costanzi, E.; Graziosi, L.; Distrutti, E.; Biagioli, M. Bile Acid Signaling in Inflammatory Bowel Diseases. Dig. Dis. Sci. 2021, 66, 674–693. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, R.; Takayama, T.; Yoneno, K.; Kamada, N.; Kitazume, M.T.; Higuchi, H.; Matsuoka, K.; Watanabe, M.; Itoh, H.; Kanai, T.; et al. Bile Acids Induce Monocyte Differentiation toward Interleukin-12 Hypo-Producing Dendritic Cells via a TGR5-Dependent Pathway. Immunology 2012, 136, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Valdés, A.; Moreno, L.O.; Rello, S.R.; Orduña, A.; Bernardo, D.; Cifuentes, A. Metabolomics Study of COVID-19 Patients in Four Different Clinical Stages. Sci. Rep. 2022, 12, 1650. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Yi, X.; Sun, Y.; Bi, X.; Du, J.; Zhang, C.; Quan, S.; Zhang, F.; Sun, R.; Qian, L.; et al. Proteomic and Metabolomic Characterization of COVID-19 Patient Sera. Cell 2020, 182, 59–72.e15. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.-Y.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut Microbiota Composition Reflects Disease Severity and Dysfunctional Immune Responses in Patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Durairajan, S.S.K.; Singh, A.K.; Saravanan, U.B.; Namachivayam, M.; Radhakrishnan, M.; Huang, J.D.; Dhodapkar, R.; Zhang, H. Gastrointestinal Manifestations of SARS-CoV-2: Transmission, Pathogenesis, Immunomodulation, Microflora Dysbiosis, and Clinical Implications. Viruses 2023, 15, 1231. [Google Scholar] [CrossRef]
- Stutz, M.R.; Dylla, N.P.; Pearson, S.D.; Lecompte-Osorio, P.; Nayak, R.; Khalid, M.; Adler, E.; Boissiere, J.; Lin, H.; Leiter, W.; et al. Immunomodulatory Fecal Metabolites Are Associated with Mortality in COVID-19 Patients with Respiratory Failure. Nat. Commun. 2022, 13, 1–11. [Google Scholar] [CrossRef]
- Campbell, C.; McKenney, P.T.; Konstantinovsky, D.; Isaeva, O.I.; Schizas, M.; Verter, J.; Mai, C.; Jin, W.-B.; Guo, C.-J.; Violante, S.; et al. Bacterial Metabolism of Bile Acids Promotes Generation of Peripheral Regulatory T Cells. Nature 2020, 581, 475–479. [Google Scholar] [CrossRef]
- Fiorucci, S.; Urbani, G.; Biagioli, M.; Sepe, V.; Distrutti, E.; Zampella, A. Bile Acids and Bile Acid Activated Receptors in the Treatment of Covid-19. Biochem. Pharmacol. 2023, 115983. [Google Scholar] [CrossRef]
- Brevini, T.; Maes, M.; Webb, G.J.; John, B.V.; Fuchs, C.D.; Buescher, G.; Wang, L.; Griffiths, C.; Brown, M.L.; Scott, W.E.; et al. FXR Inhibition May Protect from SARS-CoV-2 Infection by Reducing ACE2. Nature 2022, 615, 134–142. [Google Scholar] [CrossRef]
- Biagioli, M.; Marchianò, S.; Roselli, R.; Di Giorgio, C.; Bellini, R.; Bordoni, M.; Distrutti, E.; Catalanotti, B.; Zampella, A.; Graziosi, L.; et al. GLP-1 Mediates Regulation of Colonic ACE2 Expression by the Bile Acid Receptor GPBAR1 in Inflammation. Cells 2022, 11, 1187. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, L.; Mannaerts, I.; Schierwagen, R.; Govaere, O.; Klein, S.; Vander Elst, I.; Windmolders, P.; Farre, R.; Wenes, M.; Mazzone, M.; et al. FXR Agonist Obeticholic Acid Reduces Hepatic Inflammation and Fibrosis in a Rat Model of Toxic Cirrhosis. Sci. Rep. 2016, 6, 33453. [Google Scholar] [CrossRef]
- Inagaki, T.; Moschetta, A.; Lee, Y.-K.; Peng, L.; Zhao, G.; Downes, M.; Yu, R.T.; Shelton, J.M.; Richardson, J.A.; Repa, J.J.; et al. Regulation of Antibacterial Defense in the Small Intestine by the Nuclear Bile Acid Receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 3920–3925. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Xie, S.; Chi, Z.; Zhang, J.; Liu, Y.; Zhang, L.; Zheng, M.; Zhang, X.; Xia, D.; Ke, Y.; et al. Bile Acids Control Inflammation and Metabolic Disorder through Inhibition of NLRP3 Inflammasome. Immunity 2016, 45, 802–816. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-D.; Chen, W.-D.; Yu, D.; Forman, B.M.; Huang, W. The G-Protein-Coupled Bile Acid Receptor, Gpbar1 (TGR5), Negatively Regulates Hepatic Inflammatory Response through Antagonizing Nuclear Factor κ Light-Chain Enhancer of Activated B Cells (NF-ΚB) in Mice. Hepatology 2011, 54, 1421–1432. [Google Scholar] [CrossRef]
- Clinical Spectrum | COVID-19 Treatment Guidelines. Available online: https://www.covid19treatmentguidelines.nih.gov/overview/clinical-spectrum/ (accessed on 29 October 2023).
- Fouad, Y.; Waked, I.; Bollipo, S.; Gomaa, A.; Ajlouni, Y.; Attia, D. What’s in a Name? Renaming ‘NAFLD’ to ‘MAFLD.’ Liver Int. 2020, 40, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Sánchez, N.; Bugianesi, E.; Gish, R.G.; Lammert, F.; Tilg, H.; Nguyen, M.H.; Sarin, S.K.; Fabrellas, N.; Zelber-Sagi, S.; Fan, J.G.; et al. Global Multi-Stakeholder Endorsement of the MAFLD Definition. Lancet Gastroenterol. Hepatol. 2022, 7, 388–390. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, D.; Kim, H.J.; Lee, C.H.; Yang, J.I.; Kim, W.; Kim, Y.J.; Yoon, J.H.; Cho, S.H.; Sung, M.W.; et al. Hepatic Steatosis Index: A Simple Screening Tool Reflecting Nonalcoholic Fatty Liver Disease. Dig. Liver Dis. 2010, 42, 503–508. [Google Scholar] [CrossRef]
- Gadaleta, R.M.; Oldenburg, B.; Willemsen, E.C.L.; Spit, M.; Murzilli, S.; Salvatore, L.; Klomp, L.W.J.; Siersema, P.D.; van Erpecum, K.J.; van Mil, S.W.C. Activation of Bile Salt Nuclear Receptor FXR Is Repressed by Pro-Inflammatory Cytokines Activating NF-ΚB Signaling in the Intestine. Biochim. Biophys. Acta 2011, 1812, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Massafra, V.; Ijssennagger, N.; Plantinga, M.; Milona, A.; Ramos Pittol, J.M.; Boes, M.; van Mil, S.W.C. Splenic Dendritic Cell Involvement in FXR-Mediated Amelioration of DSS Colitis. Biochim. Biophys. Acta 2016, 1862, 166–173. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
This figure depicts the gene expression profile in patients with COVID-19. Venn diagram illustrating changes in gene regulation for patients with COVID-19 who: (A) do not have MAFLD; (B) have MAFLD. The volcano plot represents statistically significant alterations in the expression levels of the investigated genes (AHR, FFAR2, FXR and TGR5) for the group of fifteen COVID-19 patients: without MAFLD (C); with MAFLD (D).
Figure 2.
This figure depicts the gene expression profile in patients with COVID-19. Venn diagram illustrating changes in gene regulation for patients with COVID-19 who: (A) do not have MAFLD; (B) have MAFLD. The volcano plot represents statistically significant alterations in the expression levels of the investigated genes (AHR, FFAR2, FXR and TGR5) for the group of fifteen COVID-19 patients: without MAFLD (C); with MAFLD (D).
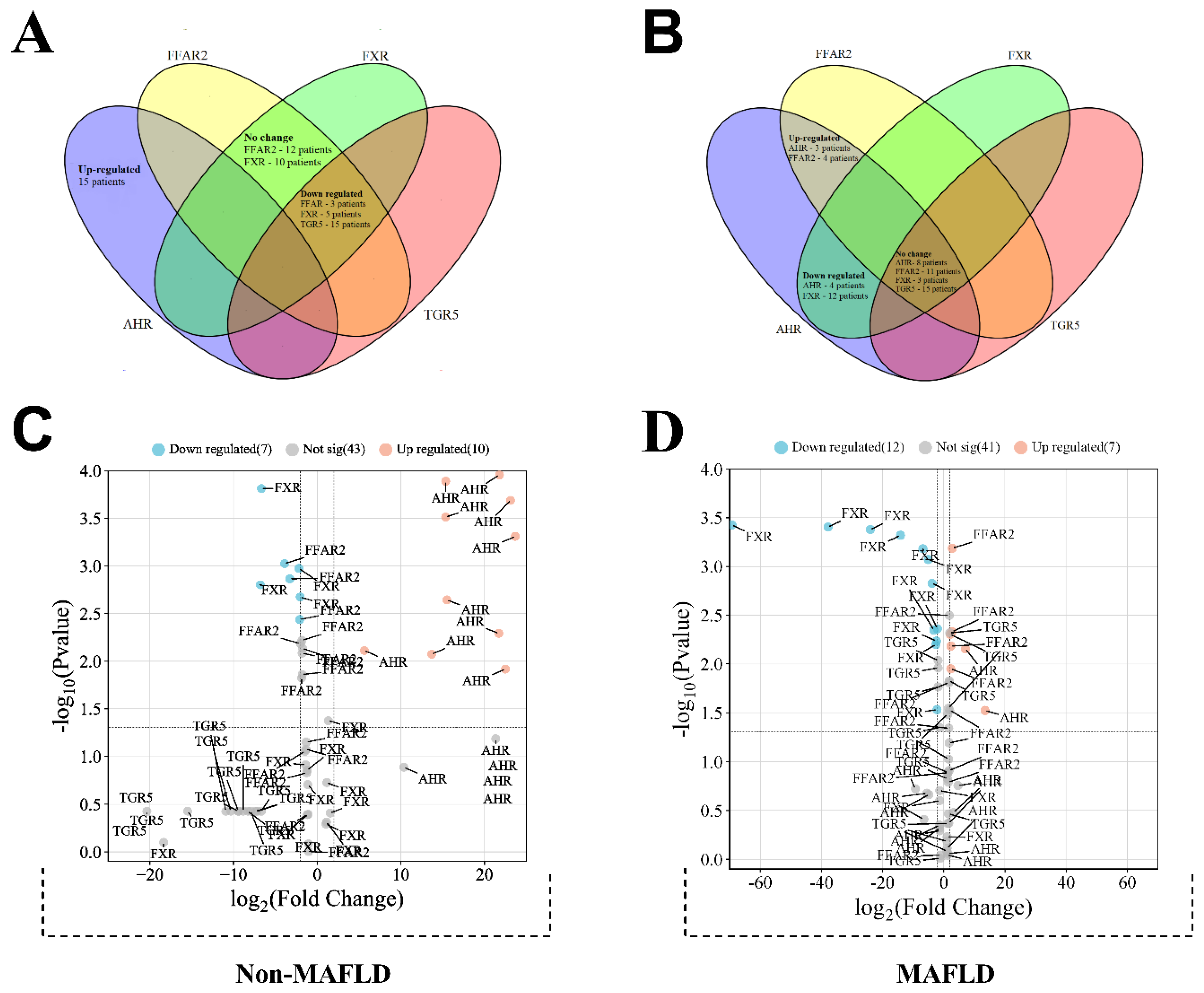
Table 1.
The expression levels of AHR, FFAR2, FXR, and TGR5 across various conditions.
| Normalized Expression (median, IQR a) | |||||
| AHR | FFAR2 | FXR | TGR5 | ||
| Sex | Male (n = 17) | 0.871 (0.519–1.53) | 0.938 (0.621–1.14) | 1.88 (0.863–3.77) | 0.798 (0.404–0.999) |
| Female (n = 13) | 1.22 (0.494–1.72) | 0.876 (0.656–1.1) | 1.92 (0.430–3.54) | 0.681 (0.387–0.848) | |
| p-Value b | p = 0.837 | p = 1.000 | p = 0.650 | p = 0.509 | |
| MAFLD | Presence (n = 15) | 0.715 (0.133–2.04) | 1.11 (0.938–1.46) | 1.3 (0.298–2.04) | 0.801 (0.46–1.03) |
| Absence (n = 15) | 1.22 (0.863–1.33) | 0.646 (0.584–0.88) | 3.41 (1.75–4.27) | 0.702 (0.325–0.832) | |
| p-Value | p = 0.161 | p < 0.001 | p = 0.021 | p = 0.174 | |
| COVID 19 severity | Moderate (15) | 1.20 (0.77–2.14) | 0.876 (0.64–1.07) | 2.74 (1.30–3.66) | 0.606 (0.344–0.846) |
| Severe (15) | 0.774 (0.456–1.22) | 0.928 (0.665–1.46) | 1.65 (0.298–3.41) | 0.801 (0.681–1.01) | |
| p-Value | p = 0.137 | p = 0.367 | p = 0.233 | p = 0.023 | |
| Presence of pneumonia | Presence (n = 16) | 0.649 (0.351–1.13) | 0.997 (0.7–1.4) | 1.7 (0.364–2.87) | 0.815 (0.484–1) |
| Absence (n = 14) | 1.53 (0.863–2.44) | 0.871 (0.636–0.999) | 3.19 (1.04–4.08) | 0.65 (0.348–0.812) | |
| p-Value | p = 0.001 | p = 0.193 | p = 0.093 | p = 0.179 | |
| Obesity | Presence (n = 10) | 0.847 (0.375–2.67) | 1.05 (0.868–1.23) | 1.86 (0.539–3.17) | 0.763 (0.348–1.05) |
| Absence (20) | 1.04 (0.616–1.32) | 0.835 (0.628–1.09) | 2.31 (0.707–3.92) | 0.703 (0.436–0.856) | |
| p-Value | p = 0.846 | p = 0.109 | p = 0.530 | p = 0.373 | |
| T2DM | Presence (n = 10) | 0.772 (0.396–2.28) | 1.12 (0.868–1.35) | 0.971 (0.160–3.17) | 0.833 (0.634–1.16) |
| Absence (n = 20) | 1.06 (0.519–1.32) | 0.835 (0.620–1.02) | 2.39 (1.67–4.12) | 0.671 (0.345–0.842) | |
| p-Value | p = 0.650 | p = 0.024 | p = 0.035 | p = 0.091 | |
| Arterial hypertension | Presence (n = 20) | 0.891 (0.552–1.25) | 0.953 (0.7–1.13) | 1.98 (0.698–3.5) | 0.774 (0.569–0.986) |
| Absence (n = 10) | 1.08 (0.124–1.86) | 0.761 (0.614–1.08) | 1.47 (0.608–4.48) | 0.581 (0.308–0.850) | |
| p-Value | p = 0.948 | p = 0.328 | p = 0.983 | p = 0.169 | |
| Coronary heart disease | Presence (n = 13) | 0.863 (0.607–1.67) | 0.987 (0.754–1.14) | 3.14 (0.237–3.77) | 0.705 (0.598–1.03) |
| Absence (n = 17) | 0.919 (0.386–1.53) | 0.876 (0.601–1.08) | 1.80 (0.727–3.54) | 0.702 (0.336–0.855) | |
| p-Value | p = 0.967 | p = 0.133 | p = 1.000 | p = 0.229 | |
a Median and interquartile range (IQR) were used to summarize the data. b Mann Whitney test.
Table 3.
Rotated Component Matrix for COVID-19 severity.
| Component | ||
| Factor 1 | Factor 2 | |
| Сommunity acquired pneumonia | 0.227 | 0.816 |
| SpO2 (admission) | -0.083 | -0.870 |
| Length of hospital stay (days) | 0.561 | 0 |
| Segmented neutrophils (discharge) | 0.868 | 0.411 |
| NLR (discharge) | 0.918 | 0.13 |
| Lymphocytes (discharge) | -0.903 | -0.307 |
Extraction Method: Principal Component Analysis. Rotation Method: Varimax with Kaiser Normalization (significant loadings in bold). Rotation converged in 3 iterations.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



