
Submitted:
11 May 2024
Posted:
13 May 2024
You are already at the latest version
Abstract
Biotic stressors pose serious threats to crop yield, jeopardizing food security and resulting in over US$220 billion per year losses to the agriculture industry. Plants activate innate defense mechanisms upon pathogen perception and invasion. The plant immune response comprises numerous concerted steps, including the recognition of invading pathogens, signal transduction, and activation of defensive pathways. However, pathogens have evolved various structures to evade plant immunity. Given these facts, genetic improvements of plants are required for sustainable disease management to ensure global food security. Advanced genetic technologies have provided new opportunities to revolutionize and boost plant disease resistance against devastating pathogens. Furthermore, targeting susceptibility (S) genes, such as OsERF922 and BnWRKY70, through CRISPR methodologies offers novel frontiers to disrupt the molecular compatibility of pathogens and introduce durable resistance against them in plants. Here, we provide a critical overview of advances in understanding disease resistance mechanisms. The review also critically examines management strategies under challenging environmental conditions and R gene-based plant genome engineering systems intending to enhance plant responses against emerging pathogens. This work underscores the transformative potential of modern genetic engineering practices in revolutionizing plant health and crop disease management while emphasizing the importance of responsible application to ensure sustainable and resilient agricultural systems.
Keywords:
Agriculture
; climate change
; pathogens
; resistance mechanisms
; genome engineering
1. Introduction
Agriculture is the most important sector as it provides raw materials for the feed and food industries. The world population is currently increasing by approximately 1.1% per year [1], and if this trend continues, the global population is projected to reach 9.9 billion by 2050 [2]. Consequently, global food production needs to increase by 70% to effectively meet the nutritional requirements of the growing population [3]. While climate-driven constraints threaten crop production worldwide [4]. At least half of the loss in agricultural production systems is due to biotic stresses imposed by pathogenic fungi, bacteria, viruses, and nematodes [5]. In some cases, new races of pests and diseases have resulted in global yield losses ranging from 10% to 40% [6], impacting the agriculture industry with annual losses exceeding US$220 billion [7]. For instance, emerging strains of wheat blast [8,9] and wheat stem rust fungus Ug99 [10] severely reduced wheat production by 15% in 2017 and 2018. Usually, chemical fungicides and pesticides have been extensively used to control plant diseases, but their excessive use has resulted in environmental pollution, harmful impacts on beneficial microorganisms, and the emergence of new pathogens [11]. Therefore, urgent efforts are required for the sustainable development of the agriculture system to address biotic stresses and increase food production.
Plant disease resistance is typically categorized into major resistance (qualitative) and partial resistance (quantitative) [12,13]. Major resistance in plants relies on the presence of resistance genes (R), which encode intracellular immune receptors such as nucleotide-binding leucine-rich repeats (NLRs) and immune receptors like receptor-like kinases. These cellular immune receptors detect specific avirulence (Avr) proteins/cognate effectors or conserved pathogenic molecules either directly or indirectly [14]. Consequently, most R genes confer pathogen race-specific "gene-for-gene resistance," which can be easily disrupted, as pathogens rapidly evolve to mutate the cognate Avr genes to escape host recognition [15]. In contrast, partial resistance is considered broad-spectrum and long-term resistance against numerous races of pathogens, controlled by quantitative trait loci (QTL) [16]. Crop improvement programs based on plant resistance genes/QTL need to be optimized and significantly studied to overcome the existing challenges posed by phytopathogens. The only feasible approach to address this alarming situation is to deploy innovative and advanced strategies or identify resistance mechanisms to combat biotic stresses. In recent decades, advancements in biotechnology have facilitated the understanding of plant-pathogen interactions and explored physiological and molecular defense mechanisms in plants to confer resistance against pathogens. For example, recent studies have concluded that the plant growth hormone salicylic acid (SA) activates systemic acquired resistance and regulates stomatal closure via the cell-specific transcription factor NTM1-LIKE 9 (NTL9) upon pathogen attack [17,18]. However, despite this progress in understanding disease resistance mechanisms, critical knowledge about plant-pathogen interactions remains incomplete. Exploring these gaps is crucial to further elucidate modern genetic engineering approaches aimed at revolutionizing crop disease resistance in agricultural systems.
The rapid expansion of genome-engineering techniques has enabled the investigation of the molecular basis of plant-pathogen interactions. Genome-engineering approaches have allowed us to explore new functions or enhance existing ones in crop plants [19,20]. Recently, targeting R-genes-mediated pathogen resistance using CRISPR-Cas9 has provided high-yield crop varieties and durable disease resistance against pathogens [21]. Liu et al. [22] reported that knocking out the GmTAP1 susceptibility gene in Glycine max enhanced resistance against Phytophthora sojae compared to wild-type plants. In another study, Zhou et al. [23] created targeted mutations in the ERF922 gene and observed strong resistance against bacterial blight disease in mutant rice plants compared to non-mutated plants. Given these facts, it is expected that R gene-based genome engineering is more powerful, cost-effective, and rapid than natural evolution. In this article, we summarize the progress made in the current understanding of pathogen resistance mechanisms, their implications for the agriculture sector, and the prospects of CRISPR-mediated genome engineering of R genes to improve plant resistance against different pathogens. Moreover, new strategies for improving durable disease resistance in crops against emerging pathogens are also proposed.
2. Host-Based Factors Affecting Pathogen-Intrusion
2.1. Immune Attenuation
The distribution of energy between defense and growth needs constant fine-tuning of the signaling process, including the immediate attenuation and activation of processes that significantly overlap. Rapid activation and engagement of broad immune signaling processes, such as mitogen-activated protein kinases (MAPK), are required against pathogen resistance following pathogen perception [24,25]. The MAPK cascade, one of the best-known immune signaling systems, also plays a crucial role in this process. In Arabidopsis thaliana, signaling facilitated by MAPK3/6, which triggers its own inhibitory process, provides a clear illustration of immune attenuation [26]. In an elegant and comprehensive analysis of MAPK phosphatases 1 (MKP1) dynamic activity, phosphorylation of MKP1 by MAPK6 is saturated for approximately 10 minutes after pathogen-associated molecular patterns (PAMP) treatment, leading to stabilization and elevation of MAPK1 protein levels [27] Besides MAPKs, MKPs might dephosphorylate various substrates involved in immune signaling, as evidenced by the fact that MKP1 regulates thousands of MKP6-independent pro-immune transcriptions within 90 minutes following flg22 elicitation in A. thaliana [28]. Furthermore, in line with the MKP1 function as a broad regulator of immune attenuation, previous studies have shown that MKP1 act as a negative regulator in defense against hemi-biotrophic pathogens (e.g. Pseudomonas syringae) in A. thaliana and biotrophic pathogens (e.g. Ralstonia solanacearum) in tomato [29].
Other than MKPs, protein phosphatases (i.e. PP2A/Cs) play a substantial role in reducing kinase activity during immune signaling, contributing to immune attenuation [30]. For instance, Jagodzik et al. [31] reported that groups of PP2Cs such as Hal1/2/3 interact with MAPK3/6 and inactivate flg22-mediated jasmonic acid (JA) signaling, a crucial virulence mechanism used by pathogens to control immune signaling in plants. The dephosphorylation of MAPK4/6 by Arabidopsis AP2C1 modulates SA and JA-associated immune signaling. In another study, Guo et al. [32] reported that protein phosphatase GhAP2C1 interacts with GhMPK4 and attenuates the immune response to enhance resistance against Fusarium oxysporum in cotton. Other kinases, such as pattern-triggered immunity (PTI) signaling regulators BAK1, CPK6, and BIK1, also modulate the activation of PP2A/Cs under biotic stress in plants [33]. Further exploration of the complex network of molecular responses mediated by the plant immune attenuation system can elucidate these mechanisms to develop robust strategies for long-term resistance against pathogens.
2.2. Developmental Stage
Plants experience dynamic changes in abiotic and biotic conditions throughout their life cycle, developing unique ecological niches for host-plant interactions. The developmental stage of growth and chronological age of a plant are significantly correlated with the strength of the plant immune system [34,35]. Immune signaling becomes more robust from early developmental stage to the reproductive period; thereafter, the strength of the immune system decreases due to host senescence [36]. A recent eco-genetic study showed two approaches adopted by the Arabidopsis natural population to enhance pathogen resistance: a shortened vegetative stage to promote fast reproduction and prevent pathogen infection and death, or a prolongation of the vegetative stage to obtain more energy for the development of a robust immune [35,37]. The strength of the immune system rises from early to late-developed organs due to the spatial scale process. For example, when rice plants were infected with Xanthomonas oryzae pv. oryzae, low disease symptoms were observed in adult leaves compared to juvenile leaves [38]. In another study, cauline leaves, adult rosettes, and juvenile rosettes from 8-week-old Arabidopsis plants showed variable resistance against Sclerotinia sclerotiorum [39]. Similar results were observed in tobacco plants inoculated with the S. sclerotiorum.
Moreover, the age-related robustness of the immune system depends on the functions of essential phytohormone signaling components, as evidenced by decreased resistance in cauline and adult leaves of hormone biosynthesis mutants abab1 (abscisic acid), jar1 (jasmonic acid), and sid1 (salicylic acid) compared to control plants [40]. However, the comprehensive molecular basis of developmental stage-associated host resistance remains unclear, and further studies are required to explore the underlying mechanisms of developmental stage-associated host resistance against pathogens.
3. Underlying Mechanisms to Counter Pathogen Attack
3.1. Physiological Mechanisms
3.1.1. Stomatal Immunity
Plants have developed sophisticated mechanisms to recognize microbial infections and prevent pathogen intrusion through controlled stomatal regulation. Numerous studies have investigated the role of stomata in plant innate immunity in pathogen resistance [41,42]. Stomata serves as the first line of defense against pathogens by regulating their opening and closing of guard cells. pathogen-associated molecular patterns (PAMPs) induce stomatal closure within 1 hour after pathogen recognition by host cells [43]. For example, wheat plants rapidly close their stomata upon detecting bacterial molecules via the FLS2 (flagellin sensing 2) receptor [44] Similarly, the application of elicitors such as chitosan or oligogalacturonic acid, associated with fungal invasion, stimulates stomatal closure in Asiatic dayflower and tomato under biotic stress condition [45], while a bacterial phytotoxin, syringomycin, induces stomatal closure in broad bean [46].
Stomatal closure is regulated by various pathways involving functional and regulatory components, including receptors, ion channels, secondary messengers, phosphatases, and protein kinases [47]. The elevated levels of SA following pathogen invasion stimulate the generation of secondary messenger ions such as Ca2+, NO, and reactive oxygen species (ROS) [48]. These secondary ion messengers further activate the SLAC1 (anion) and K+ in (potassium) ion channels to facilitate stomatal closure [49]. Moreover, abscisic acid (ABA) plays a vital role in promoting stomatal closure during pathogen invasion [50].
However, prolonged stomatal closure can reduce transpiration and photosynthesis, creating an aqueous apoplast that promotes pathogen colonization. Plant genomes encode thousands of peptides that regulate reproduction, development, and long- and short-distance signaling in plants [51,52]. Peptides such as PLANT SCREW UNRESPONSIVE RECEPTOR (NUT) and SMALL PHYTOCYTOKINES REGULATING DEFENSE AND WATER LOSS (SCREWs) secreted by plants regulate PAMP- and ABA-induced stomatal closure. SCREWs, sensed by NUT, function as immunoregulatory phytocytokines and recruit the SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE (SERK) co-receptor to modulate immune signaling. SCREWs stimulate NUT-dependent phosphorylation of ABI1 and ABI2, resulting in reduced activity of S-type ion channels and increased activity of ABI1 towards PAMP- and OST1-induced stomatal closure. Following initiation by pathogen infection and dehydration, SCREW-NUT signaling promotes apoplastic water loss and disrupts the pathogen-rich aqueous environment to prevent colonization [53].
3.1.2. Hypersensitive Response, Reactive Oxygen Species Burst, and cell Wall Modifications
Hypersensitive response (HR) is the most common immune response in plants, inducing programmed cell death (PCD) at the site of pathogen infection to prevent the further spread of infection to healthy tissues. Pathogen infection stimulates peroxidase activity, leading to the production of ROS [54,55]. NADPH oxidases, particularly RBOHD, play a main role in this process by triggering a burst in apoplastic H2O2 accumulation. RBOHD interacts with pattern recognition receptors (PRRs) and phosphorylates BIK1 to activate ROS production [56]. ROS stimulates PCD, and H2O2 spreads to adjacent cells, initiating the production of chemicals such as glutathione to prevent oxidative damage [57,58]. In addition to triggering HR, ROS creates an unfavorable environment for pathogen reproduction and survival [59]. Oxalate oxidases and amines are also involved in ROS generation, along with NADPH oxidases and peroxidases [60]. Barley and wheat produce oxalate oxidase, known as germins, which break down oxalic acid secreted by pathogens to enhance resistance. Transgenic plants with barley and wheat oxidase genes have shown improved resistance to S. sclerotiorum compared to control plants [61,62,63].
Pathogens are required to penetrate the cell wall to enter the host plant for successful infection by overcoming the physical barriers. The plant cell wall is reinforced with various polymers at sites of pathogen attack to inhibit intrusion [64]. Silicon (Si) is the most observed polymer deposited in the cell wall, along with lignin, callose, and extensins to prevent pathogen infection [65]. This deposition is supported by the upward translocation of Si through the apoplast, followed by Si deposition in the extracellular spaces of xylem vessels and leaf epidermal cells [66]. This process forms a thick layer of Si-cuticle complex in the membrane, providing mechanical resistance and enhancing plant resilience against pathogen attack. Si also forms complexes with organic compounds in epidermal cell walls, providing further strength and reducing disease intensity [67]. Abdelrhim et al. [68] reported that the application of Si improves resistance against Rhizoctonia solani in wheat plants by increasing extracellular adsorption in the spongy mesophyll and air spaces of the leaf. Si-mediated modifications in the cell wall contribute to increased plant physical resilience against pathogen penetration.
3.1.3. Phytohormones
Plants produce a diverse array of metabolites crucial for activating defense signaling against phytopathogen attacks (Figure 1). Among these metabolites, ABA, ethylene (ET), JA, and SA emerge as pivotal signaling molecules or phytohormones responsible for mediating defense responses across various pathogen lifecycles [69,70]. SA regulates local and systemic resistance responses to biotrophic and hemibiotrophic pathogens and interacts with numerous developmental and response functions [71]. SA and MAPK cascades can act upstream of each other, with SA activating MAPK cascades and some cascades stimulating SA activity [72,73]. SA transfers signals regarding pathogen presence through a complex signaling cascade, activating transcription factors to induce the expression of defense-related genes. After initiating a signaling cascade, SA mediates the reduction of disulfide bonds within the oligomeric protein nonexpressor of pathogenesis-related genes 1 (NPR1) by thioredoxins.This reduction process facilitates the translocation of NPR1 monomers from the cytosol into the nucleus. Within the nucleus, these monomers bind to the transcription factor TGA, specifically at the TGACG binding site, and upregulate the expression of resistance genes [74,75].
ET and JA play significant roles in conferring resistance against herbivorous insects and necrotrophic pathogens. These hormones stimulate the secretion of volatile compounds in response to caterpillar oral secretions [76]. ET synthesis increases upon perception of bacterial flagellin [77]. In the absence of ET, the F-box protein degrades the ethylene insensitive 3 (EIN3) transcription factor through proteasome and ubiquitination activity. ET inactivates the constitutive triple response1 (CTR1) protein and its receptors, reducing EIN2 and EIN3 expression and allowing upregulation of ET signaling and necrotroph resistance [78,79]. Pathogen interference also targets this pathway, with the XopD effector from Xanthomonas euvesicatoria bacterium desumoylating the transcription factor SIERF4 to disrupt hormone signaling, particularly inhibiting ET synthesis and resistance [80]. In addition to ET and JA, auxin and ABA are involved in plant resistance against pathogens. Stomata closure is linked to ABA signaling, regulating gas exchange, water loss, and preventing pathogen access to plant tissues [81]. Peptides can also act as plant hormones. Some studies have demonstrated that small peptide hormones like systemin are involved in systemic herbivory response, resulting in changes in gene expression, particularly in neighboring plants not exposed to biotic stress [82,83]. This suggests that plant hormones can facilitate communication among individual plants to enhance resistance.
3.2. Molecular Mechanisms Underpinning R Genes
3.2.1. Host Reprogramming by Passive Loss of Susceptibility
Mutations in cellular pathways represent a common strategy for host reprogramming, offering durable resistance against various pathogens. This form of susceptibility loss is typically governed by recessive traits, but sometimes dominant alleles are also involved. Genes associated with this phenomenon are termed adult plant resistance (APR) genes, as they confer resistance to plants in their later stages [84]. Moreover, APR genes commonly provide partial protection against a broad spectrum of pathogens [85]. It is important to note that the processes underlying this type of resistance are not used in adult plants, thus the term APR does not fully encompass this category. Furthermore, loss-of-interaction and perception-based mechanisms may specifically function in adult plants. For instance, recessive loss-of-function mlo (Mildew Locus O) alleles serve as key regulators of a senescence-associated loss-of-susceptibility mechanism in both monocots and dicots. MLO encodes a protein integral to membrane function, acting as a negative regulator of cell death under biotic stresses. Spontaneous cell death is associated with MLO loss-of-function alleles [86].
For instance, recessive loss-of-function mlo (Mildew Locus O) alleles serve as key regulators of a senescence-associated loss-of-susceptibility mechanism in both monocots and dicots. MLO encodes a protein integral to membrane function, acting as a negative regulator of cell death under biotic stresses. Spontaneous cell death is associated with MLO loss-of-function alleles [86]. In barley and A. thaliana, mlo co-expresses with PEN1/2/3 (or its barley orthologs), which are required to activate the immune response against powdery mildew disease. MLO serves as a negative regulator of the PN1/2/3 pathways, as these genes are essential for mlo-dependent immunity in both A. thaliana and barley [87]. Therefore, loss of the general cell death suppressor confers resistance by deregulating the PEN1/2/3 pathway in both monocots and dicots. This principle has been used to develop transgene-free tomatoes resistant to powdery mildew disease [88]. Similar to the mlo resistance mechanism, a loss-of-function mutation in the Pi21 gene, which encodes an HMA domain protein, functions to inhibit the plant defense response and confers recessive resistance against rice blast [89,90]. It is notable that HMA domain proteins serve as essential virulence targets for several pathogenic fungi. Effectors with unrelated sequences adopt a common fold and integrate with a domain of HMA proteins [91,92].
Therefore, analogous to pi21, the generation of loss-of-function alleles from genes encoding HMA domains confers resistance to many pathogenic fungi in different plant species. Similarly, the dominant R gene Lr67 in wheat provides partial adult resistance to stem rust, leaf rust, and powdery mildew, primarily due to mutations in a hexose transporter that differs from the susceptible allele by two amino acids. The heterodimerization of Lr67 with the susceptible allele product exhibits dominant negative effects, resulting in decreased glucose uptake and leaf tip necrosis [93,94]. The resistance conferred by these genes depends on the deregulation of early defense responses.
3.2.2. Disrupting Interaction with Host Key Targets
The disrupting interaction with key host susceptibility factors is a common mechanism underlying recessive R genes. Indeed, various identified R genes are involved in loss-of-interaction mechanisms to confer resistance against viruses [95]. Most of the known R genes act against potyviruses and encode 4E or 4G family mutant translation initiation factors, which are unable to interact with viral transcripts cap structures, thereby imparting resistance to potyviruses [96]. Moreover, about one-third of the protective R genes involved in resistance against bacterial blight disease are recessively inherited, and disrupting interaction with key host targets appears to be an underlying mechanism [97]. A recessive mutation in the promoter area of the xa27 allele prevents TALEAvrXa27 (Transcription activator-like effectors) from manipulation, resulting in the loss of susceptibility [98]. Similarly, a mutation in a single amino acid of the recessive xa5 gene encoding IIa OsTFIIAγ5 (gamma subunit of a transcription factor) restricts bacterial movement and confers resistance to different X. oryzae strains in the adult plant [99]. OsTFIIAg5 interacts directly with TALEs to complete their activity [100]. The mutation in Xa5 seems to affect the Xa5-TALEs interaction, thus showing a varying degree of resistance against many pathovars of X. oryzae [101,102].
3.2.3. NLRs Activation by Direct Intracellular Recognition of Effectors
The direct recognition of effectors is not limited to the cell surface, as various effectors have been reported to directly interact with nucleotide-binding leucine-rich repeat proteins (NLRs) to stimulate a defense response against pathogens [103]. For example, an effector of Hyaloperonospora Arabidopsis thaliana recognized1 (ATR1) interacts directly with the NLR Peronospora parasitica1 (RPP1), resulting in its recognition [104]. Numerous RPP1 alleles have different perceptions specific to ATR1 alleles, and the LRRs of RPP1 mediate this specificity [105]. Similarly, another effector, AVRL567 from Melampsora lini, binds directly to its cognate receptors L5, L6, and L7 NLRs, encoded by allelic genes [106,107]. These receptors recognize AvrL variants differently, and the LRR domain of the NLR determines this specificity [108,109]. Interestingly, cooperative polymorphism in other NLR domains also influences the perception of effectors, showing a perception mechanism in which intramolecular interactions compete with effector binding. The L5, L6, and L7 effectors exist in an “off” and “on” equilibrium state, and the binding of effectors triggers immune signaling by stabilizing them to the “on” state [107,110]. This equilibrium model might have a wider range of applications in NLR signaling. Indeed, 21 amino acid peptides from the viral movement protein (NSm21) are directly recognized by the NLR Sw-5b, and it has been explored that NSm21 binding disrupts the interaction between NB-ARC and the leucine-rich repeats (LRR) domain, triggering perception [111].
Variants of allelic effectors are perceived by allelic NLR-encoding genes in the case of L5/6/7 and RPP1. However, the perception of homologous effector proteins by homologous and/or allelic NLRs is not constant. For instance, barley and rice both have similar NLRs located within the R gene cluster. These NLRs can recognize distinct effectors from Blumeria graminis and M. oryzae, regardless of the differences in their genetic sequences [112,113]. Moreover, potato orthologs of two distinct NLR-encoding genes in tomato L2, which recognize a fungal effector, and Sw-5b, a viral effector, can also identify effector proteins from oomycetes [114,115]. The exact molecular mechanism by which highly similar NLRs detect sequence-unrelated effectors from different pathogens is still unraveled. In several cases, the C-terminal LRR region of the NLR has the specificity-determining region, while a mutation in a different location than the NLR is sufficient to alter the specificity [116,117]. Minor variations across various regions of the NLR appear to prompt its recognition of unrelated effectors. Moreover, the effectors might evolve distinct specificity to sequence-unrelated effectors by adopting a similar fold. Indeed, gaining a structural understanding of how these NLRs bind to effectors could offer more insights into the molecular mechanisms of direct perception [118,119].
3.2.4. Active Loss of Susceptibility
Active loss-of-susceptibility mechanisms encode host proteins that disarm pathogens by disrupting their lifecycle. These mechanisms are diverse and provide resistance against various pathogens. The active loss of susceptibility mechanisms triggers the generation of danger-associated molecular patterns (DAMPs) and PAMPs after pathogen recognition [120]. For example, the first cloned R gene, Hm1 encodes an NADPH-dependent reductase usually involved in detoxifying HC toxin in maize (Table 1) [121]. Cochliobolus carbonum race 1 (CCR1) is a key virulence factor of HC, causing ear mold and leaf light in maize. Hm1 orthologs exist in barley and the grass family and such orthologs play a significant role in providing non-host resistance against CCR1 [118]. Numerous active loss-of-susceptibility mechanisms are employed to develop resistance against viruses in plants. For instance, the Tm-2 gene product of tomato provides resistance against the Tomato mosaic virus and inhibits replication by binding to replication proteins [119]. Furthermore, the resistance genes Ty-1/Ty-3 in wild tomato encode γ-clade RNA-directed RNA polymerases. These enzymes initiate RNA-directed DNA methylation, offering protection against single-strand DNA geminiviruses [122].
3.2.5. TAL Effector-Dependent Expression of Executor Genes
Executor genes comprise a new class of R genes that are transcriptionally activated by TALEs produced by Xanthomonas species, conferring resistance against Xanthomonas strains carrying these TALEs. The TALEs secreted by Xanthomonas species act as important virulent factors that regulate the expression of susceptibility (S) genes for disease development [123,124]. Executor genes function as promoter traps, stimulating the transcription of immunity-related genes. The promoter of executor genes acts as a decoy, duplicating the promoter regions of susceptibility factors, thus activating the defense response [125]. Various executor genes have been identified and cloned in different plant species to confer resistance against different phytopathogens, such as in rice Xa10 [126], Xa23 [127], Xa27 [128], in pepper Bs4C-R [129], and Bs3/Bs3-E [130]. Previously reported executor genes (Bs4C-R, Xa10, Xa23, and Xa27) either encode a protein with many putative transmembrane domains or encode a protein with catalytic activity for flavin monooxygenase (Bs3 and Bs3-E). However, an improved understanding of the specificity of DNA binding to TALEs provides more insights into developing immunity against various Xanthomonas strains [131,132]. This approach was also employed to engineer RipTALs for resistance development against R. solanacearum in tomatoes [133].

Table 1.
List of cloned R genes against different disease in plants.
| Crop specie | Gene | Protein type | Disease | Pathogen | Reference |
|---|---|---|---|---|---|
| Barley | Stb6 | Receptor kinase | Septoria tritici blotch | Zymoseptoria tritici | [198] |
| Mla1 | NB-LRR | Powdery mildew | Blumeria graminis | [199] | |
| Mla6 | NB-LRR | Powdery mildew | B. graminis | [200] | |
| Rpg1 | Protein kinase | Stem rust | Puccinia graminis | [201] | |
| Wheat | Pm3 | NB-LRR | Powdery mildew | B. gramini | [202] |
| Lr10 | NB-LRR | Leaf rust | P. triticina | [83] | |
| Lr21 | NB-LRR | Leaf rust | P. triticina | [202] | |
| Maize | ZmTrxh | H-type thioredoxin | Lethal necrosis | Sugarcane mosaic virus | [203] |
| Rp1-D | NB-LRR | Leaf rust | Puccinia sorghi | [204] | |
| Rxo1 | NB-LRR | Bacterial streak | Xanthomonas oryzae | [205] | |
| Hm1 | HC toxin reductase | corn leaf blight | Cochliobolus carbonum | [121] | |
| Hm2 | HC toxin reductase | corn leaf blight | C. carbonum | [206] | |
| Rp3 | NB-LRR | Leaf rust | Puccinia sorghi | [57] | |
| qRfg1 | CCT domain gene | Gibberella stalk rot | Fusariumgraminearum | [208] | |
| qMdr9.02 | Lignin biosynthesis | Multiple | Multiple | [209] | |
| Rice | Piz-t | NB-LRR | Rice blast | Magnaporthe oryzae | [210] |
| Pi-ta | NB-LRR | Rice blast | M. oryzae | [211] | |
| Pi-b | NB-LRR | Rice blast | M. oryzae | [212] | |
| Pi-d2 | B-lectin receptor kinase | Rice blast | M. oryzae | [213] | |
| Pi9 | NB-LRR | Rice blast | M. oryzae | [214] | |
| RGA 5 | NB-LRR | Rice blast | M. oryzae | [215] | |
| Xa1 | NB-LRR | Bacterial blight | X. oryzae | [216] | |
| Xa5 | TFIIA Transcription factor | Bacterial blight | X. oryzae | [217] | |
| Xa7 | Executer R protein | Bacterial blight | X. oryzae | [218] | |
| Xa10 | Executer R protein | Bacterial blight | X. oryzae | [126] | |
| Xa21 | Receptor kinase | Bacterial blight | X. oryzae | [219] | |
| Xa23 | Executer R protein | Bacterial blight | X. oryzae | [127] | |
| Xa26 | Receptor kinase | Bacterial blight | X. oryzae | [220] | |
| Xa27 | No homolog | Bacterial blight | X. oryzae | [128] | |
| Arabidopsis thaliana | FLS2 | NB-LRR | Necrosis | Pseudomonas syringae | [221] |
| RPM1 | NB-LRR | Necrosis | Peronospora parasitica | [222] | |
| RSP2 | NB-LRR | Necrosis | P. syringae | [223] | |
| Tomato | Cf-2 | NB-LRR | Leaf mold | Cladosporium fulvum | [159] |
| Prf | NB-LRR | Necrosis | P. syringae | [157] |
3.3. Metagenomic Dynamics
Plants harbor a diverse assembly of microbial communities, which colonize the roots during the life cycle of host plants. These host-associated microbes confer beneficial traits to plants, such as disease suppression, nutrient uptake, and promotion of plant growth [134,135]. Moreover, microbes trigger defense mechanisms that enhance the resilience of host plants to pathogens, significantly increasing disease-combating efficiency [136]. Microbiomes also enhance the defense capabilities of plants by disrupting interactions between plants and pathogens, to increase disease resistance. Bacterial antagonists belonging to genera such as Enterobacter, Comamonas, Pantoea, and Microbacterium have shown significant biocontrol against rice blast disease and have stimulated the expression of genes involved in defense responses, such as OsPAD4, OsCEBiP, OsEDS1, and OsCERK1 in rice seedlings [137]. Similarly, root-associated microbes like Pantoea sp. EA106 and Pseudomonas sp. EA105 induces disease suppression in M. oryzae-inoculated rice plants by triggering ET and JA-induced systematic resistance [138]. However, advances in molecular biology have led to the development of omics techniques, such as metagenomics, which have recently gained importance in exploring the diversity of plant-microbe interactions. Metagenomics has the potential to target numerous unique signature loci in pathogen-affected plants. This approach has already been utilized in agriculture to identify novel genes, enzymes, and microbial communities involved in disease suppression [135,139].
A recent study conducted by Llontop et al. [140] found genes and key bacterial taxa, including 33,000 archaeal and bacterial species, involved in the suppression of fungal pathogens causing root diseases. Similarly, Mendes et al. [141] conducted a metagenomic analysis and showed that Fusarium oxysporum (Fox) infection led to increased microbial diversity, network complexity, and a higher proportion of the genera Flavobacterium, Bacillus, and Dyadobacter in the rhizosphere of the fox-resistant cultivar compared to the fox-susceptible cultivar. Moreover, unique functional traits such as the biosynthesis of antifungal genes, rhamnolipids, and phenazines, as well as protein secretion systems, were dominant in the rhizobia community of P. vulgaris. In another study, a comparative metagenomic analysis of resistant and susceptible varieties of tomatoes to R. solanacearum revealed the abundant presence of Flavobacterium in the resistant variety. When rhizosphere microbiota from the resistant variety were transplanted to the susceptible variety, reduced disease symptoms were observed compared to non-treated plants Kwak et al. [142]. These studies indicate that microbiome analysis through metagenomics provides a paradigm shift and insights into deciphering the role of the microbial community in protecting against different pathogens. Moreover, the knowledge gained from these aspects through metagenomics could be utilized to engineer rhizosphere or microbial consortia to enhance plant potential to withstand biotic stresses.
4. Strategies to Exploit Innate Immune Responses of the Host for Disease Resistance
4.1. Introgression of R Genes from Wild Species
The decline in crop diversity has increased the risk of modern crops to pathogen attacks, primarily due to the continuous pursuit of monoculture for high productivity [143]. Landraces and wild species serve as significant resources for novel R genes that can be introduced into modern varieties to combat emerging races of pathogens [144]. Hence, there is an imperative need to identify new R genes/alleles and introgressed them into modern varieties to obtain resistance. Indeed, various R genes, such as Xa21, Xa23, Xa27, Yr36, CcRpp1, and Fhb7, have been introgressed from landraces or wild species of plants into modern varieties to confer resistance against pathogens [127,128,145]. The Fhb7 gene, derived from wild relatives of wheat, confers resistance against Fusarium head blight [146], while CcRpp1, introgressed from a wild pigeon pea, showed strong resistance against soybean rust [147]. Similarly, in another study, the Rph gene was introgressed from wild barley carrying resistance to the leaf rust pathogen (Puccinia hordei) [148]. However, an efficient field trial platform is required to identify effective elite alleles and R genes involved in disease resistance for breeding programs. Natural-based selection for severely damaging crop pathogens must be developed for widespread screening of germplasm resources to develop resistance. Therefore, plants under high-pressure selection in fields are exposed to continuous infection by pathogens throughout plant growth to identify new resistance genes. Similarly, pigm-mediated resistance was confirmed using nursery trials in different locations over multiple years, inoculating with various isolates [149,150].
4.2. Identification and Acceleration of of R Gene Cloning
The identification and cloning of R genes are expensive and slow processes, primarily due to the preparation of libraries and artificial bacterial chromosomes. However, in recent years, advances in bioinformatics and genome sequencing technology have enabled the exploration of genomic regions linked with complex traits [144]. Moreover, high-throughput-based genotyping facilitates the detection of QTLs and genes associated with pathogen resilience. Mapping-by-sequencing is also gaining importance in identifying and cloning novel R genes in plants [151]. The genetic architecture of R genes in important crops has been identified through genome-wide association studies (GWAS) [152]. For example, R genes against rice blast disease, such as LABR_64 and the partial resistance gene LABR12, have been identified using GWAS analysis [153,154]. Similarly, scald resistance genes, including Rrs1, Rrs2, Rrs4, Rrs12, Rrs13, Rrs14, Rrs15, Rrs17, and Rsr18, have been identified in barley germplasm across different chromosomes using GWAS and mapped through different molecular markers [155,156,157].
Association genetics combined with gene enrichment sequencing (AgRenSeq) is another powerful tool for identifying NLR-like genes from landraces or wild species. AgRenSeq, which combines RenSeq with association mapping to achieve pan-genome variation in diverse crop germplasm, has been used to clone R genes such as SrTA1662, Sr33, and Sr45 in wheat. Mut-RenSeq, a technique that combines mutagenesis and RenSeq, was used to identify and isolate two resistance genes of stem rust, namely Sr45 and Sr22, in wheat [158]. Similarly, MutChromSeq combines chromosome flow sorting, EMS mutagenesis, and high-throughput sequencing to identify induced mutations in plants by comparing them with parental chromosomes [159]. Gao et al. [160] reported a new method targeting chromosome-based cloning by long-range assembly (TACCA). TACCA assembles complex genomes by combining Chicago long-range linkage with chromosome flow sorting. This technique has been employed to clone the Lr22a leaf rust-resistant gene in 4 months using ethyl methylsulfonate and marker information in wheat plants.
4.3. CRISPR-Cas9-Mediated Genome Engineering to Confer Disease Resistance
Conventional breeding plays a crucial role in improving plant resistance to pathogens, usually involving the growth and screening of large crop populations over multiple generations. However, identifying plants with the desired traits from such a vast population is labor-intensive and time-consuming [161]. The rapid advancement of genome engineering approaches in recent decades has enabled the modification of numerous components of the plant immune system to achieve long-term resistance against pathogens. Zinc finger nucleases (ZFNs), transcription-activator-like effector nucleases (TALENs), and CRISPR (clustered regularly interspaced short palindromic repeats)/associated Cas9 protein are widely used approaches for generating site-specific genetic mutations and developing disease-resistant plants [162]. Both ZFNs and TALENs rely on chimeric proteins consisting of non-specific cleavage nucleases and sequence-specific DNA binding [163,164]. However, their applications present certain challenges such as high cost, complex construction, and high off-target rates, which hinder their widespread use in agricultural research. In contrast, CRISPR/Cas9 is a technically simpler and time-saving method that can be deployed for genome engineering, offering promising prospects for improving plant disease resistance against dominant phytopathogens [161,165].
CRISPR-Cas9 has been extensively utilized for targeted mutagenesis in host plants to confer disease resistance in important agricultural crops [166]. For instance, CRISPR-Cas9 was successfully employed to confer resistance in wheat against powdery mildews by simultaneously targeting three MLO homologs, namely TAMLO-A, -B, and -D [167]. In another study, the knockout of the OsERF922 gene increased resistance against blast fungal pathogens in rice [168]. Moreover, CRISPR/Cas9-enabled mutagenesis of the eukaryotic translation initiation factor 4E (eIF4E) has been shown to enhance resistance to potyviruses in A. thaliana [169] and cucumber [170]. Similarly, targeted mutagenesis has also been employed to confer resistance against Phytophthora infestans by CRISPR/Cas9-mediated engineering of TaNFLX1, reducing post-infection pathogen proliferation in wheat [171]. In Brassica napus, the knockout of the BnWRKY70 gene increases host resistance against S. sclerotiorum [22].
CRISPR/Cas9 also offers significant potential for engineering S genes, which are essential for facilitating the infectious process of pathogens (see Figure 2). Disabling plant S genes through genome editing is a novel approach to confer broad-spectrum disease resistance in various economically important crops [172]. For instance, eIF4E is required for the cellular infection cycle of potyviruses. The 5′-terminal capped protein interacts with eIF4E to initiate the translation of viral proteins. CRISPR/Cas9-enabled mutagenesis of eIF4E has led to improved resistance to potyviruses in cassava [173]. Similarly, Mlo encodes a membrane-associated protein essential for the penetration of powdery mildew (PM) fungus into host epidermal cells. Mutation in Mlo through CRISPR-Cas9 has mediated resistance in tomato [174] and wheat [175]) via a non-transgenic system. Furthermore, a mutation introduced in DOWNY MILDEW RESISTANCE 6 (DMR6) through CRISPR/Cas9 in grapevines has conferred increased resistance to downy mildew disease [176]. Collectively, the studies illustrate the outstanding performance of CRISPR/Cas9 in developing disease-resistant crop varieties. Despite the remarkable achievements to date, challenges such as off-target effects remain to be addressed. However, potential future avenues for this technology in terms of design and application could be considered. Further identification of S genes in various plant species will pave the way for the long-term development of disease resistance using CRISPR-Cas9 technology.
4.4. Nanohyrbid-Induced Plant Disease Resistance
Traditional techniques such as crop cultivation management and plant breeding provide a solution to potentially ensure food security and safety, as well as to help plants cope with various stresses. However, these methods cannot effectively address the issues due to long process. The only practical solution to overcome this alarming situation is to employ innovative and advanced strategies or develop stress-resilient varieties to combat environmental challenges. The emergence of precise and modern genome engineering techniques, such as CRISPR-Cas9, has significantly increased the versatility of gene editing due to its superior efficacy in inducing targeted mutagenesis in the target genome [162,165]. The successful implementation of CRISPR/Cas9 for improving various agronomically important traits in diverse plant species reveals that this technology could become invaluable for crop improvement. However, these advanced genetic engineering techniques still face various challenges, including modularity, epigenetics, high complexity, standardization, difficulties in characterization, and the risk of accidental release into wild species. Moreover, genetic modification is an irreversible process, raising significant social concerns [177,178].
Nanohybrids, with their valuable and unique properties, have emerged as a stimulating branch of nanotechnology with a vast variety of applications in the agriculture sector. These nanohybrids endow augmented new properties such as tolerance against environmental stresses, artificial photosynthesis, and improved plant innate immunity [179,180]. Moreover, the advent of nanohybrids has also revolutionized the field of plant genetic engineering by enhancing plant resistance against pathogens without causing any harmful effects on the environment (see Figure 3). For example, silver nitrate coated with titanate nanotubes (AgTNTs) significantly inactivates the replication of Botrytis cinerea through the photoinactivation method. The sharp morphology and cytotoxicity of AgTNT trigger ROS production, which inhibits the further spread of conidia [181]. Similarly, Sidhu et al. [182] reported that MgO-sepiolite nanohybrids showed effective antifungal activity against devastating pathogens such as F. fujikuroi, and F. verticillioides, and Bipolaris oryzae compared to MgO NPs. MgO-sepiolite causes extensive damage to hyphae by breaking the cell wall and internal damage. In another study, Ranpariya et al. [183] synthesized silver-platinum nanohybrids (AgPtNHs) using Dioscorea bulbifera tuber extract and found that AgPtNHs exhibit strong antibacterial activity against Staphylococcus aureus and Pseudomonas aeruginosa, as well as significantly inhibit biofilm formation.
Given the remarkable structural diversity and composition of advanced synthetic materials, nanohybrid-based genome engineering is anticipated to be more powerful, cost-effective, and rapid compared to traditional genetic engineering or natural evolution. However, as information on nanohybrid-based genome engineering remains limited, further research in this area is necessary to deepen our understanding and strengthen our knowledge.
5. Challenges and Future Directions
Scientific communities and public institutions are investing considerable financial and human resources into the development of disease-resistant crops to sustain agricultural production. However, a considerable number of technical challenges are still to be solved.
5.1. Food Safety Issues
Agricultural production serves as the cornerstone of the modern economy. Both traditional and recently developed gene engineering methods show promise in addressing food security issues. However, concerns have arisen about food safety with the adoption of modern genetic engineering approaches [184]. Emphasizing food safety over the incorporation of new properties and traits is crucial. However, concerns exist regarding the ecological hazards and potential health risks associated with genetically modified crops intended for human consumption [178]. For instance, 'Starlink' maize, engineered to express Cry9c, was approved for industrial use and animal feed in the USA but not for human consumption due to elevated protein levels and potential immune system interactions [185]. Subsequently, Aventis discontinued 'Starlink' due to the detection of Cry protein residues in food products [186]. Moreover, certain studies have suggested potential health hazards such as chronic kidney diseases, increased tumor incidences, female mortality, and necrosis in male rats fed with transgenic maize known as 'Seralini' [187]. However, this study faced criticism for inappropriate statistical analysis and flawed experimental design, resulting in subsequent retraction. The research group later republished the study in 2014 with an extended format, underscoring the need for long-term feeding experiments to assess the safety of modified crops [188]. Further concerns have been raised about the potential horizontal transfer of antibiotic resistance marker genes to humans and gut bacteria in animals [189].
Pollen-mediated transfer of transgenes from transgenic crops to wild cultivars has been documented in various crops, including rice [190,191]. The introgression of transgenes from transgenic crops to conventional cultivars raises concerns about biodiversity loss and gene flow to weeds or their relatives, potentially leading to adverse effects such as the emergence of herbicide-resistant weeds and new weed species. For instance, 16 glyphosate-resistant weed species have been identified, developed due to modern engineering approaches [192]. Amaranthus tuberculatus and A. palmeri are among the most resistant weeds, significantly impacting the global economy [193]. Therefore, the development of proper testing methods is imperative to evaluate the safety of genetically modified crops and sustain the agricultural food production system in the future.
5.2. Nano Vehicle-Based Gene Delivery to Enhance Disease Resistance Traits
In recent years, CRISPR-Cas9 technology has made major revolutions in plant genome editing, enabling efficient and precise editing. However, some challenges are present about gene delivery methods. Conventional delivery techniques, such as Agrobacterium tumefaciens, protoplast transformation, biolistic, and liposome-mediated methods, have limitations including low transformation efficiencies, tissue damage, host-range restrictions, and complex protocols [194]. The rapid advancement of nanotechnology in recent decades has redirected plant genetic engineering towards nanoparticle-mediated gene transformation to overcome these challenges. Nanoparticles offer promising solutions due to their unique physicochemical properties, tunable size, and ability to penetrate biological barriers, thereby inducing desirable traits such as disease and pest resistance in plants [195]. For instance, Hajiahmadi et al. [196] used silicon nanoparticles (SiNPs) for delivering the cryIAb gene-containing plasmid into tomato plants by injecting the solution into the abaxial surface of tomato leaves. Subsequent bioassays and molecular analyses confirmed the expression of cryIAb, resulting in increased resistance of tomato plants against Tuta absoluta. Similarly, chitosan nanoparticles (Chitosan NPs) have been shown to enhance cargo delivery, leading to a significant increase in protection or durable resistance against pearl millet downy mildew in pearl millet [197].
5.3. Public Acceptance and Awareness
Modern agricultural practices have led to environmental degradation, land pollution, and ecosystem disruption, aggravated by the extensive use of chemical fertilizers. This unsustainable scenario in the food and agriculture sector requires innovative solutions. Recent research has unveiled the potential of nanohybrids to enhance disease resistance, improve agricultural inputs, and address agriculture-related challenges, thereby promoting plant resistance and food security. The prospective use of nanoscale agrochemicals, such as nanohybrid-based fertilizers, has transformed traditional agricultural practices, making them more sustainable and efficient. However, the application of nano-products in real-life scenarios raises concerns about exposure levels and potential toxicological effects on human health and the environment, highlighting the need for public awareness to alleviate confusion and stigmatization (from perceiving nano-products as either 'nano-good' or 'nano-bad'). To mitigate these risks, governmental and non-governmental organizations must implement regulatory frameworks to assess the ecotoxicological effects of nano-agrochemicals prior to their use in any sector. Moreover, addressing commercial potential, implementation barriers, and policy regulations about the use of nano-agrochemicals in agriculture is imperative to ensure their safe and effective integration into agricultural practices.
6. Conclusions
The knowledge of plant-pathogen interactions has experienced exponential growth in the 21st century, boosted by significant advancements in computational power and the emergence of molecular technologies. From the identification of (PAMPs to the implementation of robust defense mechanisms by plants, the ongoing arms race between hosts and pathogens underscores the intricate nature of these relationships. Furthermore, with the onset of climate change and evolving agricultural practices, new pathogen strains are emerging, requiring the urgent development of sustainable agricultural systems to address these challenges. However, by delving into the molecular mechanisms underlying plant-pathogen interactions, researchers can uncover fundamental biological processes that offer promising avenues for bolstering crop resilience against diseases. Leveraging this knowledge presents opportunities to devise innovative, targeted strategies for crop protection that are both environmentally sustainable and economically viable.
Author Contributions
Conceptualization, M.Z.; writing—original draft preparation, U.I.; writing—review and editing, C.Z; and S.S.; funding acquisition, M.Z.
Funding
This work is supported and funded by Grain Research & Development Corporation of Australia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflict of interest
The authors declare no conflicts of interest.
References
- Prospects, U.N. Highlights (ST/ESA/SER. A/423): United Nations, Department of Economic and Social Affairs, Population Division 2019. World Population.
- Anderson, R.; Bayer, P.E.; Edwards, D. Climate change and the need for agricultural adaptation. Curr. Opin. Plant. Biol. 2020, 56, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Bindraban, P.S.; Dimkpa, C.O.; Angle, S.; Rabbinge, R. Unlocking multiple public good services from balanced fertilizers. Food Secur. 2018, 10, 273–285. [Google Scholar] [CrossRef]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2022, 41, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Bai, Y.; Wei, Y.; Reiter, R.J.; Shi, H. Phytomelatonin as a central molecule in plant disease resistance. J. Exp. Bot 2022, 73, 5874–5885. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J.E.; Liu, H.; Trivedi, P. Climate change impacts on plant pathogens, food security and paths forward. Nat. Rev. Microbiol. 2023, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sadat, M.A.; Choi, J. Wheat blast: A new fungal inhabitant to Bangladesh threatening world wheat production. Plant Pathol. J. 2017, 33, 103. [Google Scholar] [CrossRef]
- Ceresini, P.C.; Castroagudin, V.L.; Rodrigues, F.A.; Rios, J.A.; Eduardo Aucique-Pérez, C.; Moreira, S.I.; Maciel, J.L.N. Wheat blast: Past, present, and future. Annu. Rev. Phytopathol. 2018, 56, 427–456. [Google Scholar] [CrossRef]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Bhavani, S.; Njau, P.; Govindan, V. The emergence of Ug99 races of the stem rust fungus is a threat to world wheat production. Annu. Rev. Phytopathol. 2011, 49, 465–481. [Google Scholar] [CrossRef]
- Intisar, A.; Ramzan, A.; Sawaira, T.; Kareem, A.T.; Hussain, N.; Din, M.I.; Iqbal, H. Occurrence, toxic effects, and mitigation of pesticides as emerging environmental pollutants using robust nanomaterials—A review. Chemosphere 2022, 293, 133538. [Google Scholar] [CrossRef]
- St. Clair, D.A. Quantitative disease resistance and quantitative resistance loci in breeding. Annu. Rev. Phytopathol. 2010, 48, 247–268. [Google Scholar] [CrossRef] [PubMed]
- Andersen, E.J.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M.P. Disease resistance mechanisms in plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef]
- Kourelis, J.; Van Der Hoorn, R.A. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell. 2018, 30, 285–299. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D. Plant pathogens and integrated defense responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Niks, R.E.; Qi, X.; Marcel, T.C. Quantitative resistance to biotrophic filamentous plant pathogens: Concepts, misconceptions, and mechanisms. Annu. Rev. Phytopathol. 2015, 53, 445–470. [Google Scholar] [CrossRef]
- Zheng, X.Y.; Zhou, M.; Yoo, H.; Pruneda-Paz, J.L.; Spivey, N.W.; Kay, S.A.; Dong, X. Spatial and temporal regulation of biosynthesis of the plant immune signal salicylic acid. Proc. Natl. Acad. Sci. 2015, 112, 9166–9173. [Google Scholar] [CrossRef]
- David, L.; Harmon, A.C.; Chen, S. Plant immune responses-from guard cells and local responses to systemic defense against bacterial pathogens. Plant Signal. Behav. 2019, 14, e1588667. [Google Scholar] [CrossRef]
- Hou, S.; Tsuda, K. Salicylic acid and jasmonic acid crosstalk in plant immunity. Essays. Biochem. 2022, 66, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Das, P.; Panda, D.; Xie, K.; Baig, M.J.; Molla, K.A. A detailed landscape of CRISPR-Cas-mediated plant disease and pest management. Plant Sci. 2022, 323, 111376. [Google Scholar] [CrossRef]
- Ganesh Bonagiri, D.D.G.; Kuri, A.; Kumar, V. CRISPR/cas9 gene editing tool for diseases resistant varieties. J. Pharm. Innov. 2022, 11, 2731–2738. [Google Scholar]
- Liu, T.; Ji, J.; Cheng, Y.; Zhang, S.; Wang, Z.; Duan, K.; Wang, Y. CRISPR/Cas9-mediated editing of GmTAP1 confers enhanced resistance to Phytophthora sojae in soybean. J Integr. Plant Biol. 2023, 65, 1609–1612. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, S.; Jiang, N.; Zhao, X.; Bai, Z.; Liu, J.; Yang, Y. Engineering of rice varieties with enhanced resistances to both blast and bacterial blight diseases via CRISPR/Cas9. Plant Biotechnol J. 2022, 20, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, L.; Macho, A.P.; Han, Z.; Hu, Z.; Zipfel, C.; Chai, J. Structural basis for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex. Science 2013, 342, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Xin, X.F. Pattern recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Li, P.; Lu, Y.J.; Chen, H.; Day, B. The lifecycle of the plant immune system. Cri. Rev. plant. Sci. 2020, 39, 72–100. [Google Scholar] [CrossRef] [PubMed]
- Park, H.C.; Song, E.H.; Nguyen, X.C.; Lee, K.; Kim, K.E.; Kim, H.S.; Chung, W.S. Arabidopsis MAP kinase phosphatase 1 is phosphorylated and activated by its substrate AtMPK6. Plant Cell. Rep. 2011, 30, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wan, Y.; Anderson, J.C.; Hou, J.; Islam, S.M.; Cheng, J.; Peck, S.C. Genetic dissection of Arabidopsis MAP kinase phosphatase 1-dependent PAMP-induced transcriptional responses. J. Exp. Bot. 2017, 68, 5207–5220. [Google Scholar] [CrossRef]
- Anderson, J.C.; Bartels, S.; Besteiro, M.A.G.; Shahollari, B.; Ulm, R.; Peck, S.C. Arabidopsis MAP Kinase Phosphatase 1 (AtMKP1) negatively regulates MPK6-mediated PAMP responses and resistance against bacteria. Plant. J. 2011, 67, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Withers, J.; Dong, X. Post-translational regulation of plant immunity. Curr. Opin. Plant. Biol. 2017, 38, 124–132. [Google Scholar] [CrossRef]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 2018, 9, 410346. [Google Scholar] [CrossRef]
- Guo, D.; Hao, C.; Hou, J.; Zhao, G.; Shan, W.; Guo, H.; Guo, X. The protein phosphatase GhAP2C1 interacts together with GhMPK4 to synergistically regulate the immune response to Fusarium oxysporum in cotton. Int. J. Mol. Sci. 2022, 23, 2014. [Google Scholar] [CrossRef]
- Segonzac, C.; Macho, A.P.; Sanmartín, M.; Ntoukakis, V.; Sánchez-Serrano, J.J.; Zipfel, C. Negative control of BAK 1 by protein phosphatase 2A during plant innate immunity. Plant Biotech. J. 2014, 33, 2069–2079. [Google Scholar] [CrossRef]
- Develey-Rivière, M.P.; Galiana, E. Resistance to pathogens and host developmental stage: A multifaceted relationship within the plant kingdom. New. Phytol. 2007, 175, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Yang, L. Time to fight molecular mechanisms of age-related resistance. Phytopathology 2019, 109, 1500–1508. [Google Scholar] [CrossRef]
- Häffner, E.; Konietzki, S.; Diederichsen, E. Keeping control: The role of senescence and development in plant pathogenesis and defense. Plants 2015, 4, 449–488. [Google Scholar] [CrossRef]
- Glander, S.; He, F.; Schmitz, G.; Witten, A.; Telschow, A.; de Meaux, J. Assortment of flowering time and immunity alleles in natural Arabidopsis thaliana populations suggests immunity and vegetative lifespan strategies coevolve. Gen. Biol. Evol. 2018, 10, 2278–2291. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Ding, X.; Cai, M.; Zhao, J.; Lin, Y.; Li, X.; Wang, S. The expression pattern of a rice disease resistance gene Xa3/Xa26 is differentially regulated by the genetic backgrounds and developmental stages that influence its function. Genetics 2007, 177, 523–533. [Google Scholar] [CrossRef]
- Paasch, B.C.; Sohrabi, R.; Kremer, J.M.; Nomura, K.; Cheng, Y.T.; Martz, J.; He, S.Y. A critical role of a eubiotic microbiota in gating proper immunocompetence in Arabidopsis. Nat. Plants. 2023, 9, 1468–1480. [Google Scholar] [CrossRef]
- Xu, Y.P.; Lv, L.H.; Xu, Y.J.; Yang, J.; Cao, J.Y.; Cai, X.Z. Leaf stage-associated resistance is correlated with phytohormones in a pathosystem-dependent manner. J. Integr. Plant. Biol. 2018, 60, 703–722. [Google Scholar] [CrossRef]
- Melotto, M.; Underwood, W.; He, S.Y. Role of stomata in plant innate immunity and foliar bacterial diseases. Annu. Rev. Phytopathol. 2008, 46, 101–122. [Google Scholar] [CrossRef]
- Rodrigues, O.; Shan, L. Stomata in a state of emergency: H2O2 is the target locked. Trends. Plant Sci. 2022, 27, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Amari, K.; Niehl, A. Nucleic acid-mediated PAMP-triggered immunity in plants. Curr. Opin. Virol. 2020, 42, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Battache, M.; Lebrun, M.H.; Sakai, K.; Soudière, O.; Cambon, F.; Langin, T.; Saintenac, C. Blocked at the stomatal gate, a key step of wheat Stb16q-mediated resistance to Zymoseptoria tritici. Front. Plant Sci. 2022, 13, 921074. [Google Scholar] [CrossRef]
- Lin, P.A.; Chen, Y.; Ponce, G.; Acevedo, F.E.; Lynch, J.P.; Anderson, C.T.; Felton, G.W. Stomata-mediated interactions between plants, herbivores, and the environment. Trends. plant Sci. 2022, 27, 287–300. [Google Scholar] [CrossRef]
- Mott, K.A.; Takemoto, J.Y. Syringomycin, a bacterial phytotoxin, closes stomata. Plant Physiol. 1989, 90, 1435–1439. [Google Scholar] [CrossRef]
- Assmann, S.M.; Jegla, T. Guard cell sensory systems: Recent insights on stomatal responses to light, abscisic acid, and CO2. Curr. Opin. Plant. Biol. 2016, 33, 157–167. [Google Scholar] [CrossRef]
- Qi, J.; Song, C.P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef]
- Sun, L.R.; Yue, C.M.; Hao, F.S. Update on roles of nitric oxide in regulating stomatal closure. Plant Signal. Behav. 2019, 14, e1649569. [Google Scholar] [CrossRef] [PubMed]
- Bharath, P.; Gahir, S.; Raghavendra, A.S. Abscisic acid-induced stomatal closure: An important component of plant defense against abiotic and biotic stress. Front. Plant. Sci. 2021, 12, 615114. [Google Scholar] [CrossRef]
- Takahashi, F.; Hanada, K.; Kondo, T.; Shinozaki, K. Hormone-like peptides and small coding genes in plant stress signaling and development. Cur. Opin. Plant. Biol. 2019, 51, 88–95. [Google Scholar] [CrossRef]
- Olsson, V.; Joos, L.; Zhu, S.; Gevaert, K.; Butenko, M.A.; De Smet, I. Look closely, the beauty may be small: Precursor-derived peptides in plants. Annu. Rev. Plant. Biol. 2019, 70, 153–186. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Hou, S.; Rodrigues, O.; Wang, P.; Luo, D.; Munemasa, S.; Shan, L. Phytocytokines signaling reopens stomata in plant immunity and water loss. Nature 2022, 605, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lu, Y.J.; Chen, H.; Day, B. The lifecycle of the plant immune system. Cri. Rev. plant. Sci. 2020, 39, 72–100. [Google Scholar] [CrossRef] [PubMed]
- Survila, M.; Davidsson, P.R.; Pennanen, V.; Kariola, T.; Broberg, M.; Sipari, N.; Palva, E.T. Peroxidase-generated apoplastic ROS impair cuticle integrity and contribute to DAMP-elicited defenses. Front. Plant. Sci. 2016, 7, 1945. [Google Scholar] [CrossRef] [PubMed]
- Kadota, Y.; Sklenar, J.; Derbyshire, P.; Stransfeld, L.; Asai, S.; Ntoukakis, V.; Zipfel, C. Direct regulation of the NADPH oxidase RBOHD by the PRR-associated kinase BIK1 during plant immunity. Mol. Cell. 2014, 54, 43–55. [Google Scholar] [CrossRef]
- Dat, J.F.; Pellinen, R.; Tom, Beeckman; Van De Cotte, B.; Langebartels, C.; Kangasjärvi, J.; Van Breusegem, F. Changes in hydrogen peroxide homeostasis trigger an active cell death process in tobacco. Plant J. 2003, 33, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Villalpando-Rodriguez, G.E.; Gibson, S.B. Reactive oxygen species (ROS) regulates different types of cell death by acting as a rheostat. Oxid. Med. Cell. Longe. 2021, 1155, 9912436. [Google Scholar] [CrossRef]
- Lehmann, S.; Serrano, M.; L’Haridon, F.; Tjamos, S.E.; Metraux, J.P. Reactive oxygen species and plant resistance to fungal pathogens. Phytochemistry 2015, 112, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Kärkönen, A.; Kuchitsu, K. Reactive oxygen species in cell wall metabolism and development in plants. Phytochemistry 2015, 112, 22–32. [Google Scholar] [CrossRef]
- Liu, F.; Wang, M.; Wen, J.; Yi, B.; Shen, J.; Ma, C.; Fu, T. Overexpression of barley oxalate oxidase gene induces partial leaf resistance to Sclerotinia sclerotiorum in transgenic oilseed rape. Plant Pathol. 2015, 64, 1407–1416. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Wang, Y.; He, H.; Niu, L.; Guo, D.; Dong, Y. Enhanced resistance to sclerotinia stem rot in transgenic soybean that overexpresses wheat oxalate oxidase. Transgenic Res. 2019, 28, 103–114. [Google Scholar] [CrossRef]
- Verma, R.; Kaur, J. Expression of barley oxalate oxidase confers resistance against Sclerotinia sclerotiorum in transgenic Brassica juncea cv Varuna. Transgenic Res. 2021, 30, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Bacete, L.; Melida, H.; Miedes, E.; Molina, A. Plant cell wall-mediated immunity: Cell wall changes trigger disease resistance responses. Plant. J. 2018, 93, 614–636. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Hausman, J.F.; Legay, S. Silicon and the plant extracellular matrix. Front. Plant Sci. 2016, 7, 463. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Greger, M. A review on Si uptake and transport system. Plants 2019, 8, 81. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Chen, S. Plant silicon-cell wall complexes: Identification, model of covalent bond formation and biofunction. Plant Physiol. Biochem. 2020, 155, 13–19. [Google Scholar] [CrossRef]
- Abdelrhim, A.S.; Mazrou, Y.S.; Nehela, Y.; Atallah, O.O.; El-Ashmony, R.M.; Dawood, M.F. Silicon dioxide nanoparticles induce innate immune responses and activate antioxidant machinery in wheat against Rhizoctonia solani. Plants 2021, 10, 2758. [Google Scholar] [CrossRef]
- Checker, V.G.; Kushwaha, H.R.; Kumari, P.; Yadav, S. Role of phytohormones in plant defense: Signalling and cross talk. Mol. Plant. Pathol. 2018, 3, 159–184. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, Q.; Wang, B.; Yuan, F. Roles of phytohormones and their signaling pathways in leaf development and stress responses. J. Agricul. Food Chem. 2021, 69, 3566–3584. [Google Scholar] [CrossRef]
- Vos, I.A.; Moritz, L.; Pieterse, C.M.; Van Wees, S.C. Impact of hormonal crosstalk on plant resistance and fitness under multi-attacker conditions. Front. Plant. Sci. 2015, 6, 639. [Google Scholar] [CrossRef]
- Wiermer, M.; Feys, B.J.; Parker, J.E. Plant immunity: The EDS1 regulatory node. Curr. Opin. Plant. Biol. 2005, 8, 383–389. [Google Scholar] [CrossRef]
- Saleem, M.; Fariduddin, Q.; Castroverde, C.D.M. Salicylic acid: A key regulator of redox signaling and plant immunity. Plant Physiol. Biochem. 2021, 168, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Tada, Y.; Spoel, S.H.; Pajerowska-Mukhtar, K.; Mou, Z.; Song, J.; Wang, C.; Dong, X. Plant immunity requires conformational charges of NPR1 via S-nitrosylation and thioredoxins. Science 2008, 321, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.I.; El-Shazly, H.H.; Badr, A. Role of salicylic acid in biotic and abiotic stress tolerance in plants. Plant Phenolics in Sustainable Agriculture 2020, 1, 533–554. [Google Scholar] [CrossRef]
- Hu, C.; Wei, C.; Ma, Q.; Dong, H.; Shi, K.; Zhou, Y.; Yu, J. Ethylene response factors 15 and 16 trigger jasmonate biosynthesis in tomato during herbivore resistance. Plant Physiol. 2021, 185, 1182–1197. [Google Scholar] [CrossRef]
- Denoux, C.; Galletti, R.; Mammarella, N.; Gopalan, S.; Werck, D.; De Lorenzo, G.; Dewdney, J. Activation of defense response pathways by OGs and Flg22 elicitors in Arabidopsis seedlings. Mol. Plant. 2008, 1, 423–445. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, Y.; Tu, Y.; Wang, Y.; Cheng, W.; Yang, Y. Overexpression of an EIN3-binding F-box protein2-like gene caused elongated fruit shape and delayed fruit development and ripening in tomato. Plant Sci. 2018, 272, 131–141. [Google Scholar] [CrossRef]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef]
- Kim, J.G.; Stork, W.; Mudgett, M.B. Xanthomonas type III effector XopD desumoylates tomato transcription factor SlERF4 to suppress ethylene responses and promote pathogen growth. Cell Host. Micr. 2013, 13, 143–154. [Google Scholar] [CrossRef]
- Bharath, P.; Gahir, S.; Raghavendra, A.S. Abscisic acid-induced stomatal closure: An important component of plant defense against abiotic and biotic stress. Front. Plant. Sci. 2021, 12, 615114. [Google Scholar] [CrossRef]
- Coppola, M.; Cascone, P.; Madonna, V.; Di Lelio, I.; Esposito, F.; Avitabile, C.; Corrado, G. Plant-to-plant communication triggered by systemin primes anti-herbivore resistance in tomatoes. Sci Rep. 2017, 7, 15522. [Google Scholar] [CrossRef] [PubMed]
- Feuillet, C.; Travella, S.; Stein, N.; Albar, L.; Nublat, A.; Keller, B. Map-based isolation of the leaf rust disease resistance gene Lr10 from the hexaploid wheat (Triticum aestivum L.) genome. Proc. Natl. Acad. Sci. 2003, 100, 15253–15258. [Google Scholar] [CrossRef] [PubMed]
- Dinglasan, E.; Periyannan, S.; Hickey, L.T. Harnessing adult-plant resistance genes to deploy durable disease resistance in crops. Essays. Biochem. 2022, 66, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.G.; Lagudah, E.S.; Spielmeyer, W.; Dodds, P.N. The past, present and future of breeding rust resistant wheat. Front. Plant. Sci. 2014, 5, 641. [Google Scholar] [CrossRef]
- Piffanelli, P.; Zhou, F.; Casais, C.; Orme, J.; Jarosch, B.; Schaffrath, U.; Schulze-Lefert, P. The barley MLO modulator of defense and cell death is responsive to biotic and abiotic stress stimuli. Plan Physiol. 2002, 129, 1076–1085. [Google Scholar] [CrossRef]
- Kusch, S.; Panstruga, R. Mlo-based resistance: A universal “weapon” to defeat powdery mildew disease. Mol. Plant-Micr. Interact. 2017, 30, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7, 1–6. [Google Scholar] [CrossRef]
- Fukuoka, S.; Saka, N.; Koga, H.; Ono, K.; Shimizu, T.; Ebana, K.; Yano, M. Loss of function of a proline-containing protein confers durable disease resistance in rice. Science 2009, 325, 998–1001. [Google Scholar] [CrossRef]
- Yang, J.; Fang, Y.; Wu, H.; Zhao, N.; Guo, X.; Mackon, E.; Li, R. Improvement of resistance to rice blast and bacterial leaf streak by CRISPR/Cas9-mediated mutagenesis of Pi21 and OsSULTR3; 6 in rice (Oryza sativa L.). Front. Plant Sci. 2023, 14, 1209384. [Google Scholar] [CrossRef]
- Maqbool, A.; Saitoh, H.; Franceschetti, M.; Stevenson, C.E.M.; Uemura, A.; Kanzaki, H.; Banfield, M.J. Structural basis of pathogen recognition by an integrated HMA domain in a plant NLR immune receptor. Elife 2015, 4, e08709. [Google Scholar] [CrossRef]
- de Guillen, K.; Ortiz-Vallejo, D.; Gracy, J.; Fournier, E.; Kroj, T.; Padilla, A. Structure analysis uncovers a highly diverse but structurally conserved effector family in phytopathogenic fungi. PLoS Pathog. 2015, 11, e1005228. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Foessel, S.A.; Singh, R.P.; Lillemo, M.; Huerta-Espino, J.; Bhavani, S.; Singh, S.; Lagudah, E.S. Lr67/Yr46 confers adult plant resistance to stem rust and powdery mildew in wheat. Theor. Appl. Genet. 2014, 127, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.W.; Herrera-Foessel, S.; Lan, C.; Schnippenkoetter, W.; Ayliffe, M.; Huerta-Espino, J.; Lagudah, E. A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 2015, 47, 1494–1498. [Google Scholar] [CrossRef] [PubMed]
- El-Orabey, W.M.; Hamwieh, A.; Ahmed, S.M. Molecular markers and phenotypic characterization of adult plant resistance genes Lr 34, Lr 46, Lr 67 and Lr 68 and their association with partial resistance to leaf rust in wheat. J. Genet. 2019, 98, 1–12. [Google Scholar] [CrossRef]
- Truniger, V.; Aranda, M.A. Recessive resistance to plant viruses. In Advances in Virus Research: Natural and Engineered Resistance to Plant Viruses, Part I. 2009, 75, 119–231. [Google Scholar] [CrossRef]
- Liu, Q.; Yuan, M.; Zhou, Y.A.N.; Li, X.; Xiao, J.; Wang, S. A paralog of the MtN3/saliva family recessively confers race-specific resistance to Xanthomonas oryzae in rice. Plant. Cell. Environ. 2011, 34, 1958–1969. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Huang, S.; Zhou, J.; Yang, B. Designer TAL effectors induce disease susceptibility and resistance to Xanthomonas oryzae pv. oryzae in rice. Mol. Plant. 2013, 6, 781–789. [Google Scholar] [CrossRef]
- Han, J.; Xia, Z.; Liu, P.; Li, C.; Wang, Y.; Guo, L.; Zhai, W. TALEN-based editing of TFIIAy5 changes rice response to Xanthomonas oryzae pv. Oryzae. Sci. Rep. 2020, 10, 2036. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ovenden, B.; Milgate, A. Recent insights into barley and Rhynchosporium commune interactions. Mol Plant. Pathol. 2020, 21, 1111–1128. [Google Scholar] [CrossRef]
- Huang, S.; Antony, G.; Li, T.; Liu, B.; Obasa, K.; Yang, B.; White, F.F. The broadly effective recessive resistance gene xa5 of rice is a virulence effector-dependent quantitative trait for bacterial blight. Plant J. 2016, 86, 186–194. [Google Scholar] [CrossRef]
- Hui, S.; Liu, H.; Zhang, M.; Chen, D.; Li, Q.; Tian, J.; Yuan, M. The host basal transcription factor IIA subunits coordinate for facilitating infection of TALEs-carrying bacterial pathogens in rice. Plant Sci. 2019, 284, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Lolle, S.; Stevens, D.; Coaker, G. Plant NLR-triggered immunity: From receptor activation to downstream signaling. Cur. Opin. Immun. 2020, 62, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Krasileva, K.V.; Dahlbeck, D.; Staskawicz, B.J. Activation of an Arabidopsis resistance protein is specified by the in-planta association of its leucine-rich repeat domain with the cognate oomycete effector. Plant Cell. 2010, 22, 2444–2458. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, A.D.; Goritschnig, S.; Staskawicz, B.J. Recognition and activation domains contribute to allele-specific responses of an Arabidopsis NLR receptor to an oomycete effector protein. PloS Pathog. 2015, 11, e1004665. [Google Scholar] [CrossRef] [PubMed]
- Bernoux, M.; Burdett, H.; Williams, S.J.; Zhang, X.; Chen, C.; Newell, K.; Dodds, P.N. Comparative analysis of the flax immune receptors L6 and L7 suggests an equilibrium-based switch activation model. Plant Cell. 2016, 28, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, X.; Rathjen, J.P.; Dodds, P.N. Direct recognition of pathogen effectors by plant NLR immune receptors and downstream signaling. Essays. Biochem. 2022, 66, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.I.A.; Guncˇar, G.; Forwood, J.K.; Teh, T.; Catanzariti, A.M.; Lawrence, G.J.; Kobe, B. Crystal structures of flax rust avirulence proteins AvrL567-A and-D reveal details of the structural basis for flax disease resistance specificity. Plant Cell. 2007, 19, 2898–2912. [Google Scholar] [CrossRef] [PubMed]
- Ravensdale, M.; Bernoux, M.; Ve, T.; Kobe, B.; Thrall, P.H.; Ellis, J.G.; Dodds, P.N. Intramolecular interaction influences binding of the Flax L5 and L6 resistance proteins to their AvrL567 ligands. PLoS Pathog. 2012, 8, e1003004. [Google Scholar] [CrossRef]
- Adachi, H.; Derevnina, L.; Kamoun, S. NLR singletons, pairs, and networks: Evolution, assembly, and regulation of the intracellular immunoreceptor circuitry of plants. Curr. Opin. Plant. Biol. 2019, 50, 121–131. [Google Scholar] [CrossRef]
- Zhu, M.; Jiang, L.; Bai, B.; Zhao, W.; Chen, X.; Li, J.; Tao, X. The intracellular immune receptor Sw-5b confers broad-spectrum resistance to tospoviruses through recognition of a conserved 21-amino acid viral effector epitope. Plant Cell. 2017, 29, 2214–2232. [Google Scholar] [CrossRef]
- Lu, X.; Kracher, B.; Saur, I.M.; Bauer, S.; Ellwood, S.R.; Wise, R.; Schulze-Lefert, P. Allelic barley MLA immune receptors recognize sequence-unrelated avirulence effectors of the powdery mildew pathogen. Proc. Natl. Acad. Sci. 2016, 113, E6486–E6495. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kou, Y.; Bao, J.; Li, Y.; Tang, M.; Zhu, X.; Zhou, B. Comparative genomics identifies the Magnaporthe oryzae avirulence effector AvrPi9 that triggers Pi9-mediated blast resistance in rice. New Phytol. 2015, 206, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Giannakopoulou, A.; Steele, J.F.; Segretin, M.E.; Bozkurt, T.O.; Zhou, J.; Robatzek, S.; Kamoun, S. Tomato I2 immune receptor can be engineered to confer partial resistance to the oomycete Phytophthora infestans in addition to the fungus Fusarium oxysporum. Mol. Plant-Micr. Inter. 2015, 28, 1316–1329. [Google Scholar] [CrossRef] [PubMed]
- Vossen, J.H.; van Arkel, G.; Bergervoet, M.; Jo, K.R.; Jacobsen, E.; Visser, R.G. The Solanum demissum R8 late blight resistance gene is a Sw-5 homologue that has been deployed worldwide in late blight resistant varieties. Theor. Appl. Genet. 2016, 129, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, F.; Nishimura, M.T. Structural, functional, and genomic diversity of plant NLR proteins: An evolved resource for rational engineering of plant immunity. Proc. Natl. Acad. Sci. 2018, 109, 20119–20123. [Google Scholar] [CrossRef] [PubMed]
- Anbu, S.; Swart, V.; Van Den Berg, N. Unmasking the invaders: NLR-mal function in plant defense. Front. Plant. Sci. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Marla, S.R.; Chu, K.; Chintamanani, S.; Multani, D.S.; Klempien, A.; DeLeon, A.; Johal, G.S. Adult plant resistance in maize to northern leaf spot is a feature of partial loss-of-function alleles of Hm1. PloS. Pathog. 2018, 14, e1007356. [Google Scholar] [CrossRef] [PubMed]
- Tettey, C.K.; Yan, Z.Y.; ZHAO, M.S.; Chao, G.E.N.G.; TIAN, Y.P.; LI, X.D. Tomato mottle mosaic virus: Characterization, resistance gene effectiveness, and quintuplex RT-PCR detection system. J. Integr. Agricul. 2022, 21, 2641–2651. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, J.; Wu, Z.; VandenLangenberg, K.; Wehner, T.C.; Wen, C.; Weng, Y. STAYGREEN, STAY HEALTHY: A loss-of-susceptibility mutation in the STAYGREEN gene provides durable, broad-spectrum disease resistances for over 50 years of US cucumber production. New Phytol. 2019, 221, 415–430. [Google Scholar] [CrossRef]
- Johal, G.S.; Briggs, S.P. Reductase activity encoded by the HM1 disease resistance gene in maize. Science 1992, 258, 985–987. [Google Scholar] [CrossRef]
- Butterbach, P.; Verlaan, M.G.; Dullemans, A.; Lohuis, D.; Visser, R.G.; Bai, Y.; Kormelink, R. Tomato yellow leaf curl virus resistance by Ty-1 involves increased cytosine methylation of viral genomes and is compromised by cucumber mosaic virus infection. Proc. Natl. Acad. Sci. 2014, 111, 12942–12947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yin, Z.; White, F. TAL effectors and the executor R genes. Front. Plant Sci. 2015, 6, 641. [Google Scholar] [CrossRef] [PubMed]
- Nowack, M.K.; Holmes, D.R.; Lahaye, T. TALE-induced cell death executors: An origin outside immunity? Trends. Plant. Sci. 2022, 27, 536–548. [Google Scholar] [CrossRef]
- Ji, Z.; Guo, W.; Chen, X.; Wang, C.; Zhao, K. Plant Executor Genes. Int. J. Mol. Sci. 2022, 23, 1524. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Wang, J.; Zeng, X.; Gu, K.; Qiu, C.; Yang, X.; Yin, Z. The rice TAL effector–dependent resistance protein XA10 triggers cell death and calcium depletion in the endoplasmic reticulum. Plant Cell. 2014, 26, 497–515. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, X.; Fan, Y.; Gao, Y.; Zhu, Q.; Zheng, C.; Zhao, K. XA23 is an executor R protein and confers broad-spectrum disease resistance in rice. Mol. Plant. 2015, 8, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.; Yang, B.; Tian, D.; Wu, L.; Wang, D.; Sreekala, C.; Yin, Z. R gene expression induced by a type-III effector triggers disease resistance in rice. Nature 2005, 435, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Römer, P.; Hahn, S.; Jordan, T.; Strauss, T.; Bonas, U.; Lahaye, T. Plant pathogen recognition mediated by promoter activation of the pepper Bs3 resistance gene. Science 2007, 318, 645–648. [Google Scholar] [CrossRef]
- Strauss, T.; van Poecke, R.M.; Strauß, A.; Römer, P.; Minsavage, G.V.; Singh, S.; Lahaye, T. RNA-seq pinpoints a Xanthomonas TAL-effector activated resistance gene in a large-crop genome. Proc. Natl. Acad. Sci. 2012, 109, 19480–19485. [Google Scholar] [CrossRef]
- Zeng, X.; Tian, D.; Gu, K.; Zhou, Z.; Yang, X.; Luo, Y.; Yin, Z. Genetic engineering of the Xa10 promoter for broad-spectrum and durable resistance to Xanthomonas oryzae pv. oryzae. Plant Biotechnol. J. 2015, 13, 993–1001. [Google Scholar] [CrossRef]
- Jiang, N.; Yan, J.; Liang, Y.; Shi, Y.; He, Z.; Wu, Y.; Peng, J. Resistance genes and their interactions with bacterial blight/leaf streak pathogens (Xanthomonas oryzae) in rice (Oryza sativa L.)—An updated review. Rice 2020, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gallas, N.; Li, X.; von Roepenack-Lahaye, E.; Schandry, N.; Jiang, Y.; Wu, D.; Lahaye, T. An ancient cis-element targeted by Ralstonia solanacearum TALE-like effectors facilitates the development of a promoter trap that could confer broad-spectrum wilt resistance. Plant Biotech. J. 2024, 22, 602–616. [Google Scholar] [CrossRef] [PubMed]
- Vannier, N.; Agler, M.; Hacquard, S. Microbiota-mediated disease resistance in plants. PLoS Pathog. 2019, 15, e1007740. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.; Ahmed, T.; Ijaz, U.; Shahid, M.; Nazir, M.M.; Azizullah; White, J.C.; Li, D.; Song, F. Bio-functionalized manganese nanoparticles suppress Fusarium wilt in watermelon (Citrullus lanatus L.) by infection disruption, host defense response potentiation, and soil microbial community modulation. Small 2023, 19, 2205687. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, J.; Zhang, H.; Ji, G.; Zeng, L.; Li, Y.; Chen, W. Bacterial blight induced shifts in endophytic microbiome of rice leaves and the enrichment of specific bacterial strains with pathogen antagonism. Front. Plant. Sci. 2020, 11, 963. [Google Scholar] [CrossRef]
- Sahu, K.P.; Kumar, A.; Patel, A.; Kumar, M.; Gopalakrishnan, S.; Prakash, G.; Gogoi, R. Rice blast lesions: An unexplored phyllosphere microhabitat for novel antagonistic bacterial species against Magnaporthe oryzae. Microb. Ecol. 2021, 81, 731–745. [Google Scholar] [CrossRef]
- Tamura, T.; Shinzato, N.; Ito, M.; Ueno, M. Microbial secondary metabolite induction of abnormal appressoria formation mediates control of rice blast disease caused by Magnaporthe oryzae. Phytopathology 2019, 167, 156–162. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Metagenomics methods for the study of plant-associated microbial communities: A review. J. Microbiol. Methods. 2020, 170, 105860. [Google Scholar] [CrossRef]
- Mechan Llontop, M.E.; Sharma, P.; Aguilera Flores, M.; Yang, S.; Pollok, J.; Tian, L.; Vinatzer, B.A. Strain-level identification of bacterial tomato pathogens directly from metagenomic sequences. Phytopathology 2020, 110, 768–779. [Google Scholar] [CrossRef]
- Mendes, L.W.; Raaijmakers, J.M.; De Hollander, M.; Sepo, E.; Gómez Expósito, R.; Chiorato, A.F.; Carrión, V.J. Impact of the fungal pathogen Fusarium oxysporum on the taxonomic and functional diversity of the common bean root microbiome. Environ. Microbiol. 2023, 18, 68. [Google Scholar] [CrossRef]
- Kwak, M.J.; Kong, H.G.; Choi, K.; Kwon, S.K.; Song, J.Y.; Lee, J.; Kim, J.F. The Rhizosphere microbiome structure alters to enable wilt resistance in tomato. Nat Biotechnol. 2018, 36, 1100–1109. [Google Scholar] [CrossRef]
- Crews, T.E.; Carton, W.; Olsson, L. Is the future of agriculture perennial? Imperatives and opportunities to reinvent agriculture by shifting from annual monocultures to perennial polycultures. Glo. Sustain. 2018, 1, e11. [Google Scholar] [CrossRef]
- Clare, S.J.; Çelik Oğuz, A.; Effertz, K.; Karakaya, A.; Azamparsa, M.R.; Brueggeman, R.S. Wild barley (Hordeum spontaneum) and landraces (Hordeum vulgare) from Turkey contain an abundance of novel Rhynchosporium commune resistance loci. Theor. Appl. Genet. 2023, 136, 15. [Google Scholar] [CrossRef]
- Fu, D.; Uauy, C.; Distelfeld, A.; Blechl, A.; Epstein, L.; Chen, X.; Dubcovsky, J. A kinase-START gene confers temperature-dependent resistance to wheat stripe rust. Science 2009, 323, 1357–1360. [Google Scholar] [CrossRef]
- Wang, H.; Sun, S.; Ge, W.; Zhao, L.; Hou, B.; Wang, K.; Kong, L. The horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef]
- Kawashima, C.G.; Guimarães, G.A.; Nogueira, S.R.; MacLean, D.; Cook, D.R.; Steuernagel, B.; Brommonschenkel, S.H. A pigeonpea gene confers resistance to Asian soybean rust in soybean. Nat. Biotechnol. 2016, 34, 661–665. [Google Scholar] [CrossRef]
- Martin, M.J.; Chicaiza, O.; Caffarel, J.C.; Sallam, A.H.; Druka, A.; Waugh, R.; Steffenson, B.J. Development of barley introgression lines carrying the leaf rust resistance genes Rph1 to Rph15. Crop Sci. 2020, 60, 282–302. [Google Scholar] [CrossRef]
- Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; He, Z. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science 2017, 355, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhu, X.; Shen, Y.; He, Z. Genetic characterization and fine mapping of the blast resistance locus Pigm (t) tightly linked to Pi2 and Pi9 in a broad-spectrum resistant Chinese variety. Theor. Appl. Genet. 2006, 113, 705–713. [Google Scholar] [CrossRef]
- Jaganathan, D.; Bohra, A.; Thudi, M.; Varshney, R.K. Fine mapping and gene cloning in the post-NGS era: Advances and prospects. Theor. Appl. Genet. 2020, 133, 1791–1810. [Google Scholar] [CrossRef]
- Thoen, M.P.; Davila Olivas, N.H.; Kloth, K.J.; Coolen, S.; Huang, P.P.; Aarts, M.G.; Dicke, M. Genetic architecture of plant stress resistance: Multi-trait genome-wide association mapping. New Phytol. 2017, 213, 1346–1362. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Wang, Y.; Peng, S.; Zhang, Y.; Xiao, Y.; Wang, D.; Wang, G.L. Dissection of the genetic architecture of rice resistance to the blast fungus Magnaporthe oryzae. Mol Plant. Pathol. 2016, 17, 959–972. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.H.; Kang, H.; Xu, Y.; Peng, Y.; Wang, D.; Gao, L.; Wang, G.L. Genome-wide association study identifies an NLR gene that confers partial resistance to Magnaporthe oryzae in rice. Plant. Biotech. J. 2020, 18, 1376–1383. [Google Scholar] [CrossRef] [PubMed]
- Genger, R.K.; Nesbitt, K.; Brown, A.H.D.; Abbott, D.C.; Burdon, J.J. A novel barley scald resistance gene: Genetic mapping of the Rrs15 scald resistance gene derived from wild barley, Hordeum vulgare ssp. spontaneum. Plant Breed. 2005, 124, 137–141. [Google Scholar] [CrossRef]
- Hofmann, K.; Silvar, C.; Casas, A.M.; Herz, M.; Büttner, B.; Gracia, M.P.; Schweizer, G. Fine mapping of the Rrs1 resistance locus against scald in two large populations derived from Spanish barley landraces. Theor. Appl. Genet. 2013, 126, 3091–3102. [Google Scholar] [CrossRef]
- Ntoukakis, V.; Saur, I.M.; Conlan, B.; Rathjen, J.P. The changing of the guard: The Pto/Prf receptor complex of tomato and pathogen recognition. Curr. Opin. Plant. Biol. 2014, 20, 69–74. [Google Scholar] [CrossRef]
- Arora, S.; Steuernagel, B.; Gaurav, K.; Chandramohan, S.; Long, Y.; Matny, O.; Wulff, B.B. Resistance gene cloning from a wild crop relative by sequence capture and association genetics. Nat Biotech. 2019, 37, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Martín, J.; Steuernagel, B.; Ghosh, S.; Herren, G.; Hurni, S.; Adamski, N.; Wulff, B.B. Rapid gene isolation in barley and wheat by mutant chromosome sequencing. Genome Biol. 2016, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell. 2021, 184, 1621–1635. [Google Scholar] [CrossRef]
- Gao, W.; Long, L.; Tian, X.; Xu, F.; Liu, J.; Singh, P.K.; Song, C. Genome editing in cotton with the CRISPR/Cas9 system. Front. Plant. Sci. 2017, 8, 1364. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, X.; Chen, X.; Zhou, J.M. From plant immunity to crop disease resistance. J. Genet. Genom. 2022, 49, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Porteus, M.H.; Carroll, D. Gene targeting using zinc finger nucleases. Nat. Biotechnol. 2005, 23, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Joung, J.K.; Sander, J.D. TALENs: A widely applicable technology for targeted genome editing. Nat. Rev. 2013, 14, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Kumar, R.; Kumar, V.; Won, S.Y.; Shukla, P. Engineering disease resistant plants through CRISPR-Cas9 technology. GM Crops. Food. 2021, 12, 125–144. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, M.S.; Yang, Y. Versatile applications of the CRISPR/Cas toolkit in plant pathology and disease management. Phytopathology 2021, 111, 1080–1090. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Zhao, K. Enhanced rice blast resistance by CRISPR/Cas9-targeted mutagenesis of the ERF transcription factor gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef]
- Pyott, D.E.; Sheehan, E.; Molnar, A. Engineering of CRISPR/Cas9-mediated potyvirus resistance in transgene-free Arabidopsis plants. Mol Plant Pathol. 2016, 17, 1276–1288. [Google Scholar] [CrossRef]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Gal-On, A. Development of broad virus resistance in non-transgenic cucumber using CRISPR/Cas9 technology. Mol. Plant. Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef]
- Brauer, E.K.; Balcerzak, M.; Rocheleau, H.; Leung, W.; Schernthaner, J.; Subramaniam, R.; Ouellet, T. Genome editing of a deoxynivalenol-induced transcription factor confers resistance to Fusarium graminearum in wheat. Mol. Plant-Micro. Inter. 2020, 33, 553–560. [Google Scholar] [CrossRef]
- Koseoglou, E.; van der Wolf, J.M.; Visser, R.G.; Bai, Y. Susceptibility reversed: Modified plant susceptibility genes for resistance to bacteria. Trends. Plant. Sci. 2022, 27, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Gomez, M.A.; Lin, Z.D.; Moll, T.; Luebbert, C.; Chauhan, R.D.; Vijayaraghavan, A.; Bart, R. Simultaneous CRISPR/Cas9-mediated editing of cassava eIF4E isoforms nCBP-1 and nCBP-2 confers elevated resistance to cassava brown streak disease. BioRxiv 2017, 209874. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lin, D.; Zhang, Y.; Deng, M.; Chen, Y.; Lv, B.; Gao, C. Genome-edited powdery mildew resistance in wheat without growth penalties. Nature 2022, 602, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, L.; Zeilmaker, T.; Giovannini, O.; Salvagnin, U.; Masuero, D.; Franceschi, P.; Moser, C. Simultaneous editing of two DMR6 genes in grapevine results in reduced susceptibility to downy mildew. Front. Plant. Sci. 2023, 14. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, S.P.; Iqbal, H.M.; Parra-Saldivar, R.; Varjani, S.; Tong, Y.W. Genetic modifications associated with sustainability aspects for sustainable developments. J. Bioeng. 2022, 13, 9509–9521. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, U.; Shafique, A.; Hasnain, N.; Babar, N.I.; Zameer, R.; Azeem, F. Current and future perspectives of genetically modified organisms in North America. GMOs and Political Stance 2023, 151–163. [Google Scholar] [CrossRef]
- Avila-Quezada, G.D.; Golinska, P.; and Rai, M. Engineered nanomaterials in plant diseases: Can we combat phytopathogens? Appl Microb. Biotechol. 2022, 106, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Dkhar, D.S.; Kumari, R.; Chandra, P. Chemically engineered unzipped multiwalled carbon nanotube and rGO nanohybrid for ultrasensitive picloram detection in rice water and soil samples. Sci. Rep. 2023, 13, 9899. [Google Scholar] [CrossRef]
- Chen, W.X.; Chen, K.F.; Chang, K.L.; Chen, W.H.; Lin, C.H.; Chen, C.H.; and Peng, Y.P. Photoelectrochemical degradation of TMAH by green synthesized silver modified titanate nanotube arrays. J. Environ. Chem. Engin. 2023, 11, 110625. [Google Scholar] [CrossRef]
- Sidhu, A.; Bala, A.; Singh, H.; Ahuja, R.; Kumar, A. Development of MgO-sepoilite nanocomposites against phytopathogenic fungi of rice (Oryzae sativa): A green approach. ACS Omega 2020, 5, 13557–13565. [Google Scholar] [CrossRef] [PubMed]
- Ranpariya, B.; Salunke, G.; Karmakar, S.; Babiya, K.; Sutar, S.; Kadoo, N.; Ghosh, S. Antimicrobial synergy of silver-platinum nanohybrids with antibiotics. Front. Microbiol. 2021, 11, 610968. [Google Scholar] [CrossRef] [PubMed]
- Tyczewska, A.; Woźniak, E.; Gracz, J.; Kuczyński, J.; Twardowski, T. Towards food security: Current state and prospects of agrobiotechnology. Trends Biotechnol. 2018, 36, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Bucchini, L.; Goldman, L.R. Starlink corn: A risk analysis. Environ. Health. Perspect. 2002, 110, 5–13. [Google Scholar] [CrossRef] [PubMed]
- US EPA. Starlink™ corn regulatory information. Pesticides. 2017, US EPA.
- Fagan, J.; Traavik, T.; Bøhn, T. The Seralini affair: Degeneration of science to re-science? Environ Sci. Eur. 2015, 27, 1–9. [Google Scholar] [CrossRef]
- Séralini, G.E.; Clair, E.; Mesnage, R.; Gress, S.; Defarge, N.; Malatesta, M.; de Vendômois, J.S. Republished study: Long-term toxicity of a Roundup herbicide and a Roundup-tolerant genetically modified maize. Environ. Sci. Eur. 2014, 26, 1–17. [Google Scholar] [CrossRef] [PubMed]
- De Santis, B.; Stockhofe, N.; Wal, J.M.; Weesendorp, E.; Lalles, J.P.; van Dijk, J.; Kleter, G. Case studies on genetically modified organisms (GMOs): Potential risk scenarios and associated health indicators. Food. Chem. Toxicol. 2018, 117, 36–65. [Google Scholar] [CrossRef] [PubMed]
- Han, S.M.; Lee, B.; Won, O.J.; Hwang, K.S.; Suh, S.J.; Kim, C.G.; Park, K.W. Gene flows from herbicide resistant genetically modified rice to conventional rice (Oryza sativa L.) cultivars. J. Ecol. Environ. 2015, 38, 397–403. [Google Scholar] [CrossRef]
- Umurzokov, M.; Bo Bo, A.; Ruziev, F.; Jia, W.Q.; Le, T.; Cho, M.K.; Park, K.W. Pollen-mediated exclusive gene flow from transgenic crops. Inter. J. Pest. Manag. 2021, 67, 260–268. [Google Scholar] [CrossRef]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef]
- Singh, S.; Singh, V.; Lawton-Rauh, A.; Bagavathiannan, M.V.; Roma-Burgos, N. EPSPS gene amplification primarily confers glyphosate resistance among Arkansas Palmer amaranth (Amaranthus palmeri) populations. Weed Sci. 2018, 66, 293–300. [Google Scholar] [CrossRef]
- Anjanappa, R.B.; Gruissem, W. Current progress and challenges in crop genetic transformation. J. Plant. Physiol. 2021, 261, 153411. [Google Scholar] [CrossRef] [PubMed]
- El-Shetehy, M.; Moradi, A.; Maceroni, M.; Reinhardt, D.; Petri-Fink, A.; Rothen-Rutishauser, B.; Schwab, F. Silica nanoparticles enhance disease resistance in Arabidopsis plants. Nat Nanotechnol. 2021, 16, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Hajiahmadi, Z.; Shirzadian-Khorramabad, R.; Kazemzad, M.; Sohani, M.M. Enhancement of tomato resistance to Tuta absoluta using a new efficient mesoporous silica nanoparticle-mediated plant transient gene expression approach. Hortic Sci. 2019, 243, 367–375. [Google Scholar] [CrossRef]
- Kumar, S.; Nehra, M.; Dilbaghi, N.; Marrazza, G.; Tuteja, S.K.; Kim, K.H. Nanovehicle for plant modifications towards pest-and disease-resistance traits. Trends. Plant Sci. 2020, 25, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Saintenac, C.; Lee, W.S.; Cambon, F.; Rudd, J.J.; King, R.C.; Marande, W.; Kanyuka, K. Wheat receptor-kinase-like protein Stb6 controls gene-for-gene resistance to fungal pathogen Zymoseptoria tritici. Nat. Genet. 2018, 50, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Kurth, J.; Wei, F.; Elliott, C.; Valé, G.; Yahiaoui, N.; Keller, B.; Somerville, S.; Wise, R.; Schulze-Lefert, P. Cell-autonomous expression of barley Mla1 confers race-specific resistance to the powdery mildew fungus via a Rar1 -independent signaling pathway. Plant Cell. 2001, 13, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Halterman, D.; Zhou, F.; Wei, F.; Wise, R.P.; Schulze-Lefert, P. The MLA6 coiled-coil, NBS-LRR protein confers AvrMla6-dependent resistance specificity to Blumeria graminis f. sp. hordei in barley and wheat. Plant J. 2001, 25, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Brueggeman, R.; Druka, A.; Nirmala, J.; Cavileer, T.; Drader, T.; Rostoks, N.; Kleinhofs, A. The stem rust resistance gene Rpg5 encodes a protein with nucleotide-binding-site, leucine-rich, and protein kinase domains. Proc. Natl. Acad. Sci. 2008, 105, 14970–14975. [Google Scholar] [CrossRef]
- Srichumpa, P.; Brunner, S.; Keller, B.; Yahiaoui, N. Allelic series of four powdery mildew resistance genes at the Pm3 locus in hexaploid bread wheat. Plant Physiol. 2005, 139, 885–895. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, H.; Gong, Y.; Tao, Y.; Jiang, L.; Zuo, W.; Xu, M. An atypical thioredoxin imparts early resistance to sugarcane mosaic virus in maize. Mol Plant. 2017, 10, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Collins, N.; Drake, J.; Ayliffe, M.; Sun, Q.; Ellis, J.; Hulbert, S.; Pryor, T. Molecular characterization of the maize Rp1-D rust resistance haplotype and its mutants. Plant Cell. 1999, 11, 1365–1376. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Lin, X.; Poland, J.; Trick, H.; Leach, J.; Hulbert, S. A maize resistance gene functions against bacterial streak disease in rice. Proc. Natl. Acad. Sci. 2005, 102, 15383–15388. [Google Scholar] [CrossRef] [PubMed]
- Chintamanani, S.; Multani, D.S.; Ruess, H.; Johal, G.S. Distinct mechanisms govern the dosage-dependent and developmentally regulated resistance conferred by the maize Hm2 gene. Mol. Plant-Micro. Interact. 2008, 21, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, Q.; Wang, W.; Li, Y.; Guo, Y.; Zhang, D.; Xu, M. A transposon-directed epigenetic change in ZmCCT underlies quantitative resistance to Gibberella stalk rot in maize. New Phytol. 2017, 215, 1503–1515. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; He, Y.; Kabahuma, M.; Chaya, T.; Kelly, A.; Borrego, E.; Balint-Kurti, P. A gene encoding maize caffeoyl-CoA O-methyltransferase confers quantitative resistance to multiple pathogens. Nat Genet. 2017, 49, 1364–1372. [Google Scholar] [CrossRef]
- Zhou, B.; Qu, S.; Liu, G.; Dolan, M.; Sakai, H.; Lu, G.; Wang, G.L. The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea. Mol. Plant-Microb. Interact. 2006, 19, 1216–1228. [Google Scholar] [CrossRef]
- Bryan, G.T.; Wu, K.S.; Farrall, L.; Jia, Y.; Hershey, H.P.; McAdams, S.A.; Valent, B. A single amino acid difference distinguishes resistant and susceptible alleles of the rice blast resistance gene Pi-ta. Plant Cell. 2000, 12, 2033–2045. [Google Scholar] [CrossRef]
- Tsunoda, Y.; Jwa, N.S.; Akiyama, K.; Nakamura, S.; Motomura, T.; Kamihara, K.; Kawasaki, S. Cloning of the rice blast resistance gene PI-B. In Advances in rice blast research; Springer: Dordrecht, 2000; pp. 9–16. [Google Scholar] [CrossRef]
- Chen, D.X.; Chen, X.W.; Lei, C.L.; Wang, Y.P.; Li, S.G. Rice blast resistance of transgenic rice plants with Pi-d2 gene. Rice Sci. 2010, 17, 179–184. [Google Scholar] [CrossRef]
- Qu, S.; Liu, G.; Zhou, B.; Bellizzi, M.; Zeng, L.; Dai, L.; Wang, G.L. The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding site–leucine-rich repeat protein and is a member of a multigene family in rice. Genetics 2006, 172, 1901–1914. [Google Scholar] [CrossRef]
- Cesari, S.; Thilliez, G.; Ribot, C.; Chalvon, V.; Michel, C.; Jauneau, A.; Rivas, S.; Alaux, L.; Kanzaki, H.; Okuyama, Y.; Morel, J.B. The rice resistance protein pair RGA4/RGA5 recognizes the Magnaporthe oryzae effectors AVR-Pia and AVR1-CO39 by direct binding. Plant Cell. 2013, 25, 1463–1481. [Google Scholar] [CrossRef]
- Yoshimura, S.; Yamanouchi, U.; Katayose, Y.; Toki, S.; Wang, Z.X.; Kono, I.; Kurata, N.; Yano, M.; Iwata, N.; Sasaki, T. Expression of Xa1, a bacterial blight-resistance gene in rice, is induced by bacterial inoculation. Proc. Natl. Acad. Sci. 1998, 95, 1663–1668. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.S.; McCouch, S.R. The rice bacterial blight resistance gene xa5 encodes a novel form of disease resistance. Mol. Plant-Microb. Interact. 2004, 17, 1348–1354. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, J.; Pan, J.; Gu, Z.; Chen, X.; Jin, Y.; Ma, B. Identification and molecular mapping of the rice bacterial blight resistance gene allelic to Xa7 from an elite restorer line Zhenhui 084. Europ. J. Plant Pathol. 2009, 125, 235–244. [Google Scholar] [CrossRef]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; Fauquet, C. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cao, Y.; Yang, Z.; Xu, C.; Li, X.; Wang, S.; Zhang, Q. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J. 2004, 37, 517–527. [Google Scholar] [CrossRef]
- Gómez-Gómez, L.; Boller, T. FLS2: An LRR receptor–like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol Cell. 2000, 5, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Mackey, D.; Holt III, B.F.; Wiig, A.; Dangl, J.L. RIN4 interacts with Pseudomonas syringae type III effector molecules and is required for RPM1-mediated resistance in Arabidopsis. Cell 2002, 108, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Axtell, M.J.; Staskawicz, B.J. Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell 2003, 112, 369–377. [Google Scholar] [CrossRef]
- Dixon, M.S.; Jones, D.A.; Keddie, J.S.; Thomas, C.M.; Harrison, K.; Jones, J.D. The tomato Cf-2 disease resistance locus comprises two functional genes encoding leucine-rich repeat proteins. Cell 1996, 84, 451–459. [Google Scholar] [CrossRef]
Figure 1.
Components of plant disease resistance mechanisms involved in pathogen detection, signal transduction, and defense response.
Figure 1.
Components of plant disease resistance mechanisms involved in pathogen detection, signal transduction, and defense response.
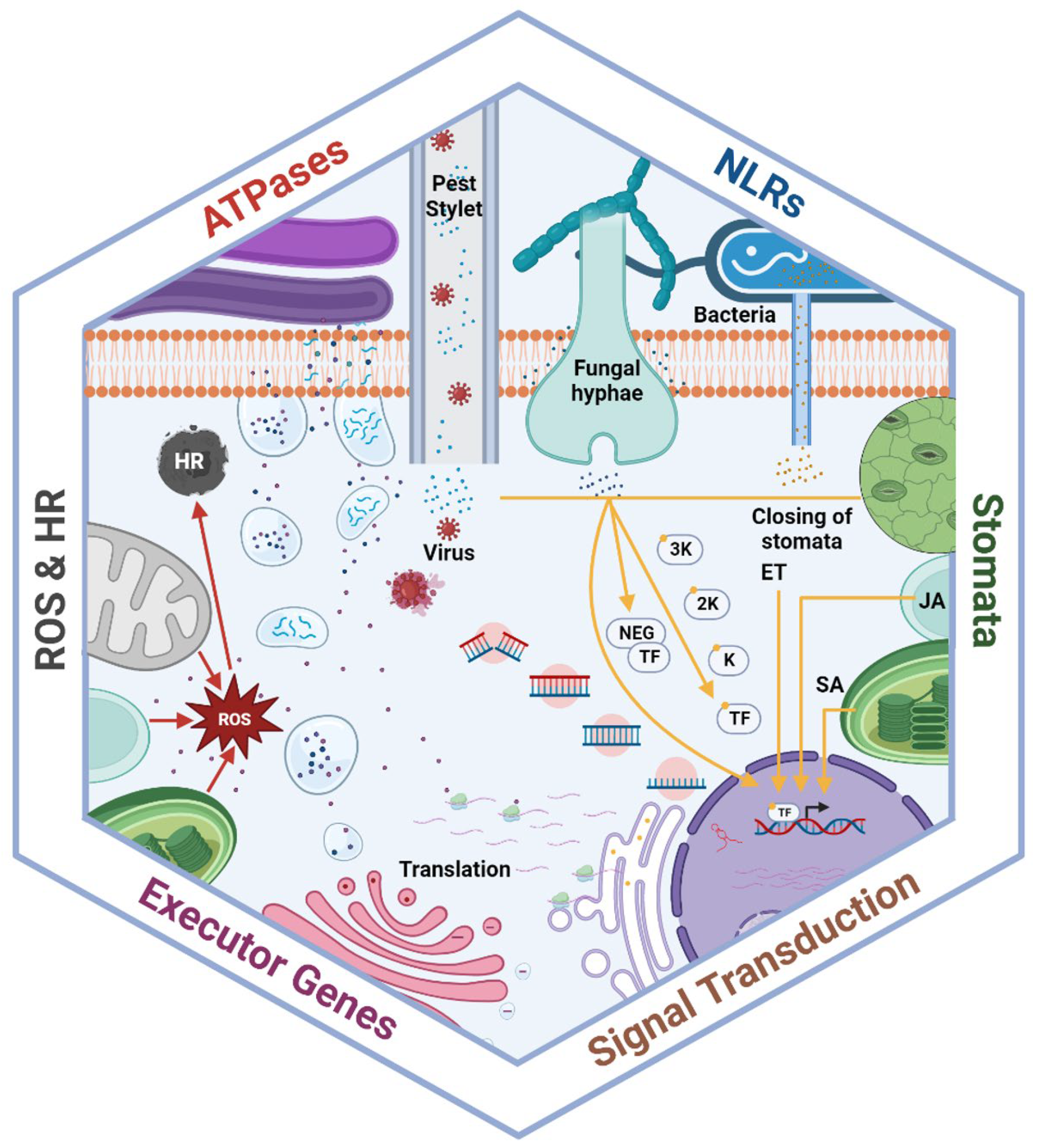
Figure 2.
illustration of development of disease-resistant variety in plants by targeting susceptibility (S) gene/s via CRISPR/Cas9 system.
Figure 2.
illustration of development of disease-resistant variety in plants by targeting susceptibility (S) gene/s via CRISPR/Cas9 system.
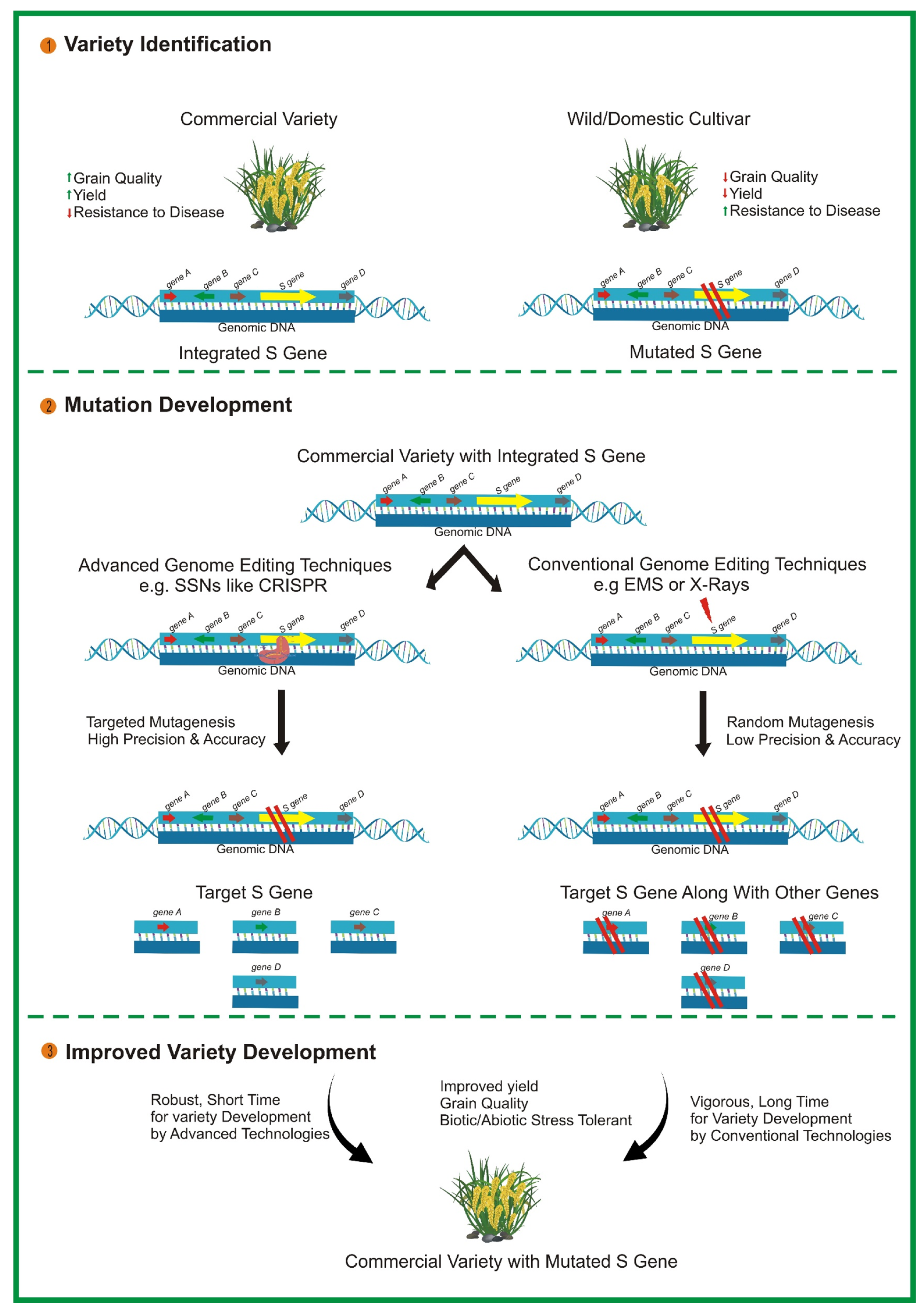
Figure 3.
Mechanistic action of nanohybrids induced pathogen resistance in plants.
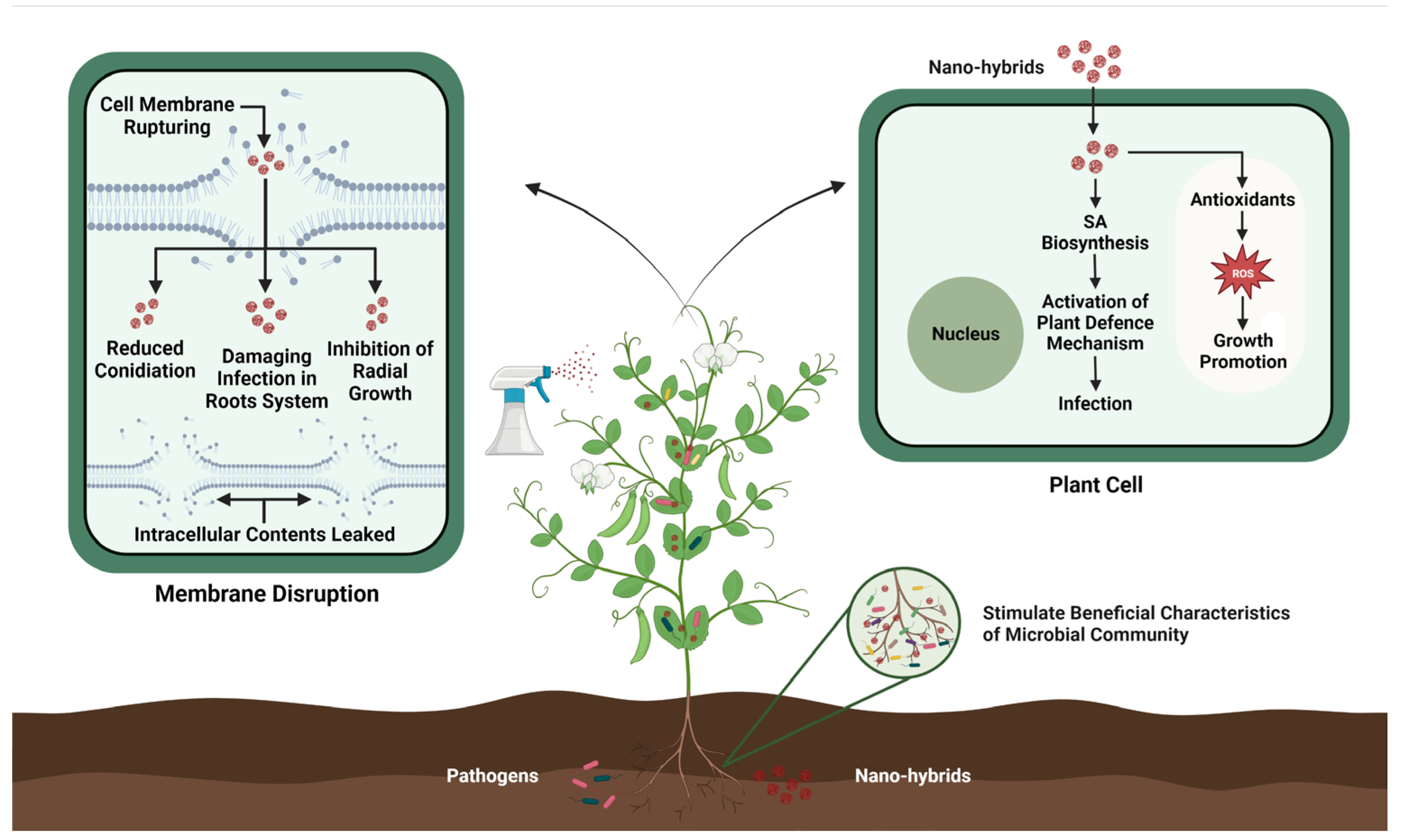
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



