
Submitted:
13 May 2024
Posted:
13 May 2024
You are already at the latest version
Abstract
Tumor microenvironment (TME) is composed of various cellular components such as tumor cells, stromal cells including fibroblasts, adipocytes, mast cell, lymphatic vascular cells and infiltrating immune cells, macrophages, dendritic cells and lymphocytes. The intricate interplay between these cells influences tumor growth, metastasis and therapy failure. Significant advancements in breast cancer therapy have resulted in a substantial decrease in mortality. However, existing cancer treatments frequently result in toxicity and nonspecific side effects. Therefore, improving targeted drug delivery and increasing the efficacy of drugs are crucial for enhancing the treatment outcome and reducing the burden of toxicity. In this review, we provided an overview of how tumor and stroma-derived osteopontin (OPN) plays a key role in regulating the oncogenic potential of various cancers including breast. Next, we have dissected the signalling network by which OPN regulates tumor progression through interaction with selective integrins and CD44 receptors. This review addresses the latest advancement in role of various splice variants of OPN in cancer progression and OPN-mediated tumor-stromal interaction, EMT, CSCs enhancement, immunomodulation, metastasis, chemoresistance, metabolic reprogramming and further suggest that OPN might be a potential therapeutic target and prognostic biomarker for the evolving landscape of cancer management.
Keywords:
breast cancer
; osteopontin (OPN)
; tumor associated macrophages
; cancer associated fibroblast
; immune modulation
1. Introduction
An estimated new cases of cancer incidences are 19.3 million and cancer related deaths are 10.0 million according to GLOBOCAN 2020 [1]. Recent data showed that female breast cancer accounts for the second most commonly occurring cancer after lung globally. Tumor microenvironment (TME) consists of heterogenous cell subpopulations including infiltrating immune cells like tumor-associated macrophages (TAMs), dendritic cells (DCs), lymphocytes and other stromal cells like cancer-associated fibroblasts (CAFs), endothelial cells, pericytes, and extracellular matrix (ECM) [2]. All these components form a complex network through cell-cell and cell-matrix interactions to shape the microenvironment conducive to tumor progression and metastasis. These cells enhance several hallmarks of cancer via secreting a rich array of cytokines and chemokines [2].
Osteopontin (OPN), a sialic acid rich, non-collagenous, chemokine like, glycosylated, phospoprotein is expressed in various cell types such as osteoblasts, osteoclasts, epithelial cells, endothelial cells and immune and stromal cells [3]. OPN has emerged as a pivotal mediator facilitating intracellular crosstalk within the breast TME. Due to variations in post-translational modifications (PTMs) and proteolytic cleavage, the molecular weight of OPN ranges from 44 to 75 kDa [3]. OPN plays a crucial role in several normal physiological processes including vascularisation, bone remodelling and immue-regulation as well as pathological processes including tumor progression, metastasis, immunosuppression, angiogenesis and chemoresistance [4,5]. The overexpression of OPN has been linked to poor prognosis in various malignancies, including breast, lung cancer, glioblastoma, colorectal cancer, hepatocellular carcinoma, bladder cancer, melanoma, and acute myeloid leukemia [6]. It also serves as diagnostic as well as prognostic biomarker in different cancers [6].
In the present scenario, the diverse function attributes to OPN in tumor progression are directly linked to its structural features binding to specific integrins and CD44. Moreover, OPN- mediated signalling activates various oncogenic molecules and promotes tumor growth as well as metastasis [3]. Interestingly, OPN also governs immunosuppression by activating macrophages and suppressing T-cell activation in various cancers [7]. Single cell and spatial transcriptomics analyses, revealed a correlation between interactions of CAF and OPN+ TAM with the progression of cancer [8].
This review comprehensively covers recent progress in the field of OPN with a focus on elucidating its structural features, immune modulatory properties, its role in regulating CAFs, influencing epithelial-mesenchymal transition (EMT), contributing to the enrichment of cancer stem cells (CSCs), promoting metastasis, fostering therapy resistance, and influencing metabolic regulation. The goal of this review is to provide the latest progress in elucidating the role of OPN in shaping the TME. We also discuss recent treatment strategies to target OPN and its receptors for the management of cancers using small molecules inhibitors, monoclonal antibodies and aptamer-based approaches.
2. OPN Structure and Function
Structural Architecture and Splice Variants of OPN
OPN is a member of SIBLING (small integrin binding ligand N-linked glycoprotein) family of ECM-associated chemokine like aspartic acid rich, glycosylated phospho-sialoprotein [9]. It consists of various domains such as arginine-glycine-aspartic acid (RGD) containing integrin binding, two calcium binding, MMP and thrombin cleavage and CD44 binding site [10].
The αvβ3, αvβ5, αvβ1and other integrins bind OPN through RGD motif whereas α9β1 and α4β7 integrins interact with OPN through SVVYGLR motif. The C-terminal region of OPN is responsible for the interaction with CD44 [11] (Figure 1). Moreover, OPN also manifests in five structural isoforms, distinguished by exon deletions, resulting from alternative splicing, exon shuffling, and other PTMs [12]. PTMs including Ser/Thr phosphorylation, tyrosine sulfation and glycosylation, contribute to the molecular weight variation of OPN, ranging from 44-75 kDa. These modifications induce both structural alterations and functional modifications deepening our understanding of the diverse roles of OPN [3]. Moreover, these five isoforms such as OPN-a, OPN-b, OPN-c, OPN-4 and OPN-5 are mainly involved in various tumorigenic events [13] (Figure 2). OPN-a, the full-length isoform consists of seven exons whereas OPN-b and OPN-c lacks exon 5 and exon 4 respectively. In addition, OPN-4 lacks both exon 4 and 5 while OPN 5 contains seven exons along with a translation start sequence positioned between the third and fourth exons. More recently, additional splice variants of OPN-5 have been reported which are denoted as OPN-5b, OPN-5c, OPN-5d and OPN-5e [13] (Figure 2). However, OPN-5c and OPN-5d have additional 9 bp insertion into the extra exon in between exon 3 and 4 [13]. Apart from secreted OPN (OPN-s), intra cellular OPN (OPN-i) is also found to orchestrate various hallmarks of cancer. Translation of OPN-s originates at the 5’ AUG start codon while OPN-i translation begins at the downstream of non-AUG codon. In addition, PTMs in OPN-i deletes the 16-aa signal sequence from the N-terminus which allows it to localize within the cytoplasm [14].
3. OPN Expression in Various Cancers
OPN exhibits high levels of expression in a wide range of tumor types, such as breast, ovarian, cutaneous, head and neck, thyroid, lung, liver, esophageal, gastric, pancreatic, colorectal, kidney, prostate, bladder and glioblastoma [15]. Tumor, stromal, and tumor- infiltrating myeloid cells express high levels of OPN within the heterogeneous TME [6]. Clinical research has demonstrated a correlation between OPN expression in various tumor tissues, plasma, serum and has been shown to be correlated with an advanced tumor stage, grade, size, invasiveness, metastasis, poor survival rate of patients in a variety of human malignancies [6]. Furthermore, the differential expression level of OPN isoforms vary depending upon the cancer types as shown in Table 1. In general, OPN-a is expressed in various malignancies and is involved in promoting cancer progression, metastasis, angiogenesis, immunosuppression and drug resistance. In breast cancer, OPN-a and OPN-c are expressed at higher levels whereas high level of OPN-b is expressed in lung cancer and are associated with poor survival and relapse [16]. However, the correlation of OPN expression with its clinical implications in various cancers remain to be explored.
4. Role of OPN in Tumor Progression
OPN regulates cancer progression by influencing several hallmarks of cancers through the interaction with its receptors. This section mainly focuses on the mechanism by which tumor and stroma derived-OPN promotes the tumor growth.
4.1. OPN Receptors
4.1.1. Integrin Receptors
Integrins, a heterodimeric cell surface receptor consisting of α and β subunits, are a cell adhesion molecule that bind with matricellular and extracellular matrix (ECM) proteins [15]. Mammals have been shown to possess 18α and 8β subunits which constitute 24 different integrins. Both the integrin α and β subunits are type I transmembrane proteins with a short (~30-40 aa) cytoplasmic domain, substantial extracellular domain, and single transmembrane domain [15]. OPN interacts with integrins αvβ3, α8β1, α5β1, αvβ1, αvβ6 and αvβ5 through GRGDS motif whereas binds with integrins α9β1, α4β1 and α4β7 via the ELVTDFPTDLPAT and/or SVVYGLR sequences [11]. The expression of integrins are low in normal adult epithelia whereas significantly high in most of the cancer cell types and tumor tissues [17]. The αvβ3 integrin not only binds to vitronectin but also interacts with other ECM proteins such as OPN, fibronectin and laminin in various cell types [18]. Expression of αvβ5 is high in non-small cell lung cancer (NSCLC), prostate cancer, lung cancer, gastric cancer as well as in associated stromal cells [19]. The α5β1 integrin is an essential angiogenic marker correlating with tumor malignancy, invasiveness and development of metastasis [19]. α8β1 integrin is mostly expressed in smooth muscle cells [20]. The α4β1integrin is overexpressed in leukocytes including eosinophils, lymphocytes, monocytes, macrophages, NK cells, basophils, and mast cells whereas α9β1integrin is over expressed in macrophages and other immune cells [21]. OPN is involved in the activation of several signal transduction pathways via integrins that promote tumor metastasis, migration, adhesion and survival [22].
4.1.2. CD44 Receptors
Apart from integrins, OPN also binds to CD44, a type I transmembrane glycoprotein composed of intracellular, extracellular and transmembrane domains [23]. Alternative splicing of CD44 genes result in the generation of two isoforms: CD44s (standard) and CD44v (variant) [23]. CD44v isoforms can have a single variant exon as seen in CD44v6 and CD44v7 or multiple variants as observed in CD44v4-v5 and CD44v3-v10 [24]. Thrombin cleavage of OPN results in two fragments that interacts with CD44 independent of RGD sequence [25]. One of which is situated downstream of the RGD motif and shown to be overlap with SVVYGLR domain, because binding of OPN and CD44 competes with α9β1 integrin but not with αvβ3 [26]. Interaction of OPN with CD44 activates multiple signalling pathways which in turn promote tumor growth. OPN is reported to enhance the radiation resistance by maintaining stemness in the adjacent cells through activation of CD44 in glioma [27]. In addition, OPN secreted by macrophages bind to CD44 and promote invasiveness by activating the Rac-specific guanine nucleotide exchange factor, TIAM1 in bladder cancer [28]. It has been reported that OPN-CD44 interaction activates c-Jun-NH (2)-kinase (JNK) signalling cascade which drives the tumorigenicity in colorectal cancer [29]. Interestingly, binding of OPN with CD44 leads to cleavage of CD44 intra-cellular domain by γ-secretase, which in turn governs the stemness characteristics in glioma [27]. Thus, targeting the OPN-CD44 axis might open a new dimension for cancer therapy.
4.1.3. Receptor-Mediated Signalling
OPN interacts with various integrins as well as other co-receptors collectively and activates downstream complex signalling cascade such as PI3K/Akt, p38/MAPK/ERK, JNK, Ras/Raf/MEK/ERK, JAK/STAT and TIAM1/Rac1pathway thereby inducing the oncogenic gene expression in various cancer cells [15]. The activation of these pathways in cancer cells regulate several pathological processes such as cell adhesion, migration, invasion, metastasis, proliferation, tumor growth, survival, chemoresistance, stemness, angiogenesis, and immune suppression. In the following section, we highlighted the OPN-mediated activation of PI3K/Akt and p38/MAPK/ERK signalling pathways in various cancer cells [15]. Moreover, we depicted such OPN-mediated signalling pathways in Figure 3.
PI3K/Akt Signalling
The PI3K/Akt signalling cascade plays crucial role in regulating several cellular processes associated with tumorigenesis. Dysregulation of this pathway has been linked to tumor growth, metastasis, EMT, immunosuppression and drug resistance. In colorectal cancer cells, OPN has been identified as a key factor that induces stem cell-like properties and cell invasion via the PI3K-Akt-GSK/β-catenin pathway [30]. Silencing of OPN downregulates migration and invasion but induces apoptosis and autophagy via inactivating the PI3K/Akt/mTOR pathway [31]. OPN promotes tumor progression and angiogenesis in oral cancer through the activation of PI3K/Akt/mTOR signalling cascade [32]. Moreover, OPN- αvβ3 interaction increases HIF-1α expression, which in turn trans-activates TCF12 gene expression [33]. TCF12 engages in transcriptional repression of the VE-cadherin gene by interacting with histone deacetylases and EZH2, promoting the endothelial-mesenchymal (EndoMT) transition [33]. OPN expression is regulated by the EGF/PI3K signaling pathway in HepG2 cells [34]. Additionally, OPN upregulates angiogenesis via activation of PI3K/Akt and ERK1/2 pathways in breast cancer whereas promotes tumor progression via PI3K/Akt/Twist signalling axis in HCC [35]. OPN knockdown reduced Akt phosphorylation and down-regulated the expression of VEGF and MMP-2 in gastric cancer [36]. Silencing calpain subunit 4 (Capn4) downregulates OPN expression and suppresses the migration of ovarian cancer cells [37]. Interestingly, OPN has also been reported to modulate drug resistance through PI3K/Akt pathway in several cancers. For example, OPN overexpression increased PI3K, p-ERK1/2, and ERCC1 expressions in lung cancer and caused cisplatin resistance, but OPN silencing decreased this effect [38]. Similarly, blocking OPN-mediated PI3K/Akt signalling pathway reversed OPN-induced cisplatin resistance in HCC cells [39]. Similarly, OPN is upregulated in EGFR-TKI resistant NSCLC cells thereby activating PI3K/Akt pathway leading to downregulation of EGFR-TKI-induced apoptosis while augmenting EMT [40]. Moreover, epoxyazadiradione, a limonoid inhibits the growth of breast tumors by depolarizing the mitochondria and inducing caspase-dependent apoptosis through the inhibition of PI3K/Akt pathway [41]. A splice variant of OPN, OPN-c has been reported to be involved in promoting tumorigenesis and proliferation by activating PI3K/Akt pathway in ovarian cancer [42].
p38/MAPK Signalling
The binding of OPN to it’s receptors activates MAPK pathway which in turn regulates several vital processes like EMT, chemoresistance and senescence [15]. It is observed that OPN- induced the PI3K/Akt and MAPK/Erk1/2 cascade promotes EMT which in turn is associated with increase cell migration, invasion, and proliferation in lung cancer [43]. OPN-mediated stimulation of the MAPK pathway is essential for the cell growth and metastasis in HCC [44]. The binding of OPN to αvβ3 stimulates VEGF expression via MAPK pathway leading to increased prostate cancer cell invasion [45]. However, OPN stimulates NF-κB nuclear translocation through the PI3K/Akt and MAPK pathways, which in turn increases HIF-1α to support the growth and survival of gastric cancer cells [46]. It was revealed that OPN-knockout breast cancer cells exhibit increased levels of cyclophosphamide-induced apoptosis when compared with control [47]. The expression of Immediate Early Response 2 (IER2) triggers senescence in melanoma cells via p53/MAPK/Akt-dependent manner and results in poor prognosis in melanoma patients [48]. Doxorubicin-treated breast cancer cells inhibit caspase- 3-induced apoptosis via activating the MAPK pathways in response to OPN [49].
Other Signalling
Many other signalling pathways are also involved in OPN-mediated regulation of tumor progression, angiogenesis and metastasis. For instance, OPN interacts with CD44 and integrins and mediates several signalling networks such the JAK/STAT and NIK pathway to trigger gene expression that mediates invasion, metastasis, and angiogenesis. Intracellular signalling pathways can be triggered by the c-Src-dependent transactivation of EGFR through interaction with OPN and integrin via the Brk/NF-κB signalling pathway that ultimately regulates VEGF expression in breast cancer cells [50]. Under hypoxic conditions, OPN triggers integrin-linked kinase (ILK)/Akt-mediated NF-κB activation, which results in HIF-1α-dependent VEGF production in breast cancer cells and subsequent angiogenesis [15]. NF-κB and HIF-1α are involved downstream of OPN signalling and induce a CSC-like phenotype in HCC [6].
4.2. Multifaceted Functions of OPN in Tumor Progression
OPN has been shown to stimulate the progression of cancer by activating specific signalling pathways. These components collectively create an intracellular "signalling traffic network" that controls the expression of various oncogenic molecules which are essential for initiating tumorigenesis , regulating EMT, stemness, angiogenesis, metastasis and drug resistance. The OPN-regulated various oncogenic functions are depicted in Figure 4.
4.1.1. EMT
EMT is a multifaceted process that occurs when epithelial cells lose their adhesion capacity and acquire a mesenchymal phenotype. The loss of epithelial integrity is characterized by downregulation of E- cadherin and upregulation of N-cadherin, fibronectin, vimentin and [11]. OPN modulates many EMT-related transcription factors such as Twist, Snail, and Slug in various solid cancers including breast. OPN also plays a critical role in initiating EMT through activation of Twist in breast cancer [51]. Interestingly, Butti et al have demonstrated that OPN- educated fibroblasts induce EMT in breast cancer cells through the secretion of CXCL12 and promotes breast cancer progression [52]. Moreover, OPN triggers an autocrine MAPK intracellular signalling cascade which leads to activation of Twist and upregulation of Bmi1 in MDA-MB-231 cells [53]. Similarly, Snail is also involved in the upregulation of genes linked with mesenchymal and invasive characteristics [51]. OPN interacts directly with Runx2 resulting in induction of Snail-dependent EMT in mammary epithelial cells [54]. OPN-specific aptamer reduces Snail expression thereby abrogating EMT using in vitro breast cancer models [51]. In addition, OPN induces NF-κB activation and increases the expression of both ZEB1 and ZEB2 resulting in the acquisition of an EMT-like phenotype in breast cancer cells [55]. The intricate interplay between the tumor and its surrounding environment is a crucial regulatory component of EMT and OPN is proven to be a key player in the tumor-stroma interaction. Tumor derived OPN upregulates Cox-2 expression in TAMs leading to an enhancement in angiogenesis and melanoma growth [56]. Myofibroblasts produced directly by OPN or by other mechanisms secrete a variety of chemokines, TNFβ, TGFβ, IL-6, sphingosine-1-phosphate (S1P) and angiotensin II (Ang II) which promotes EMT [57]. In both in vitro and in vivo breast cancer models, tumor-derived OPN have been shown to regulate the transformation of tissue-resident normal mammary fibroblasts into tumor-supporting CAFs [58].
Both tumor-derived and exogenous OPN can trigger the transition of mesenchymal stem cells (MSCs) to CAFs in breast cancer models. This is accomplished by activating MSCs to generate TGFβ, thereby initiating a feedback loop that drives the CAF phenotype [59]. OPN is found to upregulate HIF-1α which leads to Twist 1 activation followed by EMT. Remarkably, OPN-s initiates cancer metastasis by inducing the EMT, whereas OPN-i activates the MET to promote metastatic formation [60]. Therefore, targeting OPN/αvβ3 integrin and OPN/CD44 signalling cascade may control epithelial-mesenchymal plasticity in various cancers.
4.1.2. Enrichment of CSC
CSCs, also known as tumor-initiating cells, are a subpopulation of cells within a neoplasm that exhibit distinctive characteristics, including self-renewal capacity, pluripotency, and the ability to generate distinct progeny, mirroring the heterogenous inherent in the primary tumor [61]. These cells possess a unique ability to sustain tumorigenesis by maintaining their undifferentiated state, resisting conventional anti-cancer therapies, and orchestrating the hierarchical organization of the tumor mass [61,62,63]. CSCs are majorly characterized by the expression of CD44, CD24, ALDH1 and CD133 in breast cancer [61].
Several factors secreted by tumor and stroma, cell-cell contacts or cell-matrix interactions are reported to impact the stemness of cancer cells through autocrine, paracrine and juxtracrine mechanisms [11]. OPN emerges as a critical player in the enrichment of CSCs for its attribute to interact with its receptor and a recognized stem cell marker, CD44 [64]. OPN along with the stem cell marker CD44, shapes a perivascular niche that promotes CSCs phenotype and radiation resistance in glioma. The γ-secretase-cleaved intracellular domain of CD44 interacts with OPN that enriches stem cell phenotype and glioma growth through CBP/p300-dependent activation of HIF-2α [27]. Furthermore, ALDHhiCD44+CD24- stem cells successfully metastasize to bone, where bone-derived OPN is implicated in promoting stem-like phenotype in breast cancer cells, thereby influencing metastasis. The mechanism underlying OPN-induced CSC-facilitated metastasis involves CD44 and RGD-dependent cell surface integrins, which augment the functional response to bone-derived OPN, potentially through activation of WNK-1 and PRAS40- related pathways [65]. Thus bone metastasis can be targeted by disrupting this dynamic interaction. However, another study reported the contrasting role of osteoclast-derived OPN in bone metastasis, suggesting a counterintuitive option for the treatment of breast cancer-associated bone metastasis [66]. Mechanistically, the Lrp5-overexpressing osteoclast-derived OPN regulates Hsp90ab1 (Hsp90 beta) and moesin (MSN). Importantly, Hsp90ab1 immuno-precipitated latent
TGFβ and inactivated TGFβ, whereas MSN interacted with CD44 thereby inhibiting CD44 pathway [66]. This interaction suggests a potential impact on CSCs highlighting the complexity of the regulatory network established by OPN. The dichotomous role of OPN signalling may arise from the existence of multiple isoforms of both OPN and its receptors, each exhibiting context- dependent functions. Hu et al demonstrated that CD44v exhibits significantly higher lung metastatic potential as compared to CD44s in CD24-/CD44+ CSC population [67]. Modulating the CD44v/CD44s ratio through epithelial splicing regulatory protein 1 (ESRP1) expression influenced lung metastasis without affecting the stemness. CD44v, responsive to OPN in the lung environment, enhances cancer cell invasiveness and promotes lung metastasis, distinguishing it from CD44s [67]. Their findings identify a subset of metastatic breast CSCs marked by CD44v expression and respond to OPN, suggesting CD44v and OPN as key regulators of CSCs and metastasis.
4.1.3. Chemoresistance
The primary contributor to the ineffectiveness of chemotherapy in treating cancer stemness from the emergence of drug resistance against conventional therapeutic interventions. Chemoresistance can be induced by the interplay between the heterogenous cell population within the TME [68]. This interplay between several intrinsic factors such as tumor mutation load, heterogeneity, ECM and epigenetic modifications, along with extrinsic factors such as pH, hypoxia, paracrine signalling, and other stromal cells which triggers chemoresistance [69]. Mechanistically, chemoresistance results due to upregulation of various multidrug resistance efflux pump such as ABCC 1-9, ATP binding cassette (ABC) transporters and P-glycoprotein (P-gp) [70]. Elevated level of OPN mRNA expression is associated with poor disease free survival (DFS) and overall survival (OS) in a large cohort of breast cancer patients treated with adjuvant chemotherapy in clinical trials [71]. Overexpression of ABC transporters reported to be a key factor in drug resistance as it can mediate the efflux of various drugs thereby decreasing the intracellular concentration of the drug. Stromal OPN increases ABCG2 expression and enriches the side population (SP) through ERK2-dependent pathway in melanoma [72]. In prostate cancer, it has been shown that the activation of FAK leads to increased expression of P-gp upon the binding of secreted OPN to αvβ3 integrin [73]. Furthermore, Yi et al has shown that binding of OPN to αvβ3 through activating the PI3K/Akt/GSK3β/β-catenin signalling cascade results in cell survival and sorafenib insensitivity in FLT3-ITD mutant AML cells [74]. OPN causes aberrant activation of the PI3K/Akt signalling pathway via CD44 and αvβ3 integrin while blocking OPN could reverse the chemoresistance of cisplatin in HCC [39]. Upon binding to αvβ3, OPN induces autophagy thereby maintaining Foxo3a stability, which increases tumor growth and resistance to epirubicin and cisplatin in HCC cells [75]. Hyperactivated EMT has also been shown to be linked with increased treatment resistance in cancer cells and this may be induced by abnormal activation of multiple signalling pathways that drive the EMT phenotype [76]. Moreover, OPN induces the expression of N-cadherin, vimentin, Twist, Slug, MMP9 by activating the GLI-dependent Hedgehog signalling pathway in breast cancer. In this study, they have also reported that OPN upregulated the expression of drug resistant related proteins like ABCB1 and ABCG2 which in turn promote the EMT and induces efflux of therapeutic drugs such as paclitaxel, cisplatin and doxorubicin [77].
4.1.4. Angiogenesis
Angiogenesis, a pivotal process in tumor growth, is intricately regulated by the VEGF. It has been shown that OPN secreted by the tumor cells has the ability to enhance the metastatic potential and angiogenesis by regulating VEGF [78]. For example, upregulation of OPN by TBX3iso1 in breast cancer cells leads to angiogenesis using in vivo mice models [79]. ADAM8 activates OPN expression through the JAK/STAT3 pathway thereby enhancing angiogenesis in U87 cells and primary macrophages [80]. In addition, high levels of ADAM8, a proteolytically active member of the ADAM family is found in several malignancies contribute to tumor cell migration and invasion and are associated with poor patient prognosis [80]. Chakraborty et al. have shown that OPN promotes VEGF-dependent angiogenesis via the activation of the Brk/NF-κB/ATF-4 signalling pathway in paracrine, autocrine and juxtracrine manner in breast cancer [81]. Moreover, under hypoxic condition, OPN modulates HIF1α-induced VEGF expression via ILK/NF-κB signalling cascade which ultimately culminates in breast cancer progression and angiogenesis [82]. OPN induced ICOSL promotes angiogenesis and cell migration in breast cancer [83]. The interaction between SPP1+ macrophages and endothelial cells promotes angiogenesis through VEGF-A-VEGFR1/R2 [84].
4.1.5. Metastasis
In several cancers, there is a correlation between high level of OPN expression and metastasis. OPN facilitates the molecular process that dictate the development of metastatic lesions, including inhibition of apoptosis, ECM degradation and remodeling, cellular migration, host immune cell evasion and neovascularization [85]. Breast cancer cells spontaneously metastasized to lung in mouse model and the level of OPN is high in lung lesion [86]. The level of serum OPN is closely correlated with lymph node metastasis in small cell lung cancer suggesting that OPN acts as predictor of OS [87]. OPN secreted by tumor- associated cells, increased the expression of CD44v6 in colon cancer stem cells through Wnt/β- catenin pathway thereby promoting migration and metastasis [88]. In gastric cancer, increased OPN expression is linked to lymph node and distant metastasis [89]. The SNP at locus -443 of OPN promoter and related haplotypes (Ht2 and Ht3) considerably boost the activity and expression level of OPN promoter which significantly increased tumor growth and lung metastasize in HCC [90]. Lysosomal-associated membrane protein 3 (LAMP3) may control the invasion and metastasis by regulating downstream signalling of OPN in osteosarcoma [91]. OPN acts as diagnostic predictor with specificity and sensitivity of 96.15% and 52.94% respectively in bone metastasized breast cancer patients [92]. The osteolytic bone metastasis in breast cancer is significantly inhibited in conditional knocked down of OPN in nude rat models [93]. The neutralization of OPN effectively mitigated the enhanced osteoclast development and bone metastasis induced by Fam20C deficiency [94]. Combination of erufosine with OPN knockdown enhances anti-metastatic effect for the control of skeletal metastases using breast cancer cells [93]. Breast cancer bone metastasis is thought to be significantly influenced by RUNX2 which is regulated by OPN-αvβ3/CD44 axis [95]. The premetastatic niche in bone marrow requires OPN-dependent migration of CAF to promote the stemness using in vivo breast cancer model [96]. Therefore, OPN is a vital regulator of bone and lung metastases in breast cancer.
4.1.6. Cancer Cell Metabolism
Through metabolic alterations linked to carcinogenesis, transformed cells can persist in abnormal growth and invade various tissues by avoiding tissue homeostasis and utilizing an array of internal signalling pathways along with a variety of local tissue and whole-body resources. Significantly, the stromal cells in the TME, and the transformed cells themselves, all undergo metabolic remodelling in various cancers [97]. This promotes the accumulation and dissemination of cancer cells, weakens the immune system to prevent tumor growth, and increases the lethality associated with cancer [97]. This can be accomplished by rewiring the glucose metabolic pathways, which confer drug resistance and facilitate metastasis [98]. Tumor cells require glucose as a metabolic energy source for survival and proliferation. Glucose transporters (GLUTs) facilitate aerobic glycolysis, commonly called the Warburg effect, by delivering glucose into the cytosol. The class I glucose transporters GLUT1 and GLUT3 are sensitive to hypoxia and have a strong affinity for glucose [99]. Numerous genes including OPN, VEGF, PDK1 (pyruvate dehydrogenase kinase 1), iNOS, LDHA (lactate dehydrogenase A), EPO, GLUT1 and GLUT3 are regulated by HIF-1 under hypoxic conditions [100,101]. It has been reported that GLUT1 and GLUT3 upregulation is linked to a poor prognosis in breast cancer [102]. Additionally, poor survival in NSCLC is associated with overexpression of GLUT1 [103]. OPN is upregulated in hypoxic environments, that leads to increased expression of GLUT1 and GLUT3 via αvβ3 integrins mediated PI3K/Akt and p38 MAPK pathways. This process ultimately results in the development of osteosarcoma [99].
5. OPN-Mediated TME Regulation
The ECM, stromal cells, immune cells, matricellular proteins, fibroblasts, cytokines and growth factors define the complex microenvironment surround the tumors. Recent evidence suggests that tumor cell-TME interaction modulates tumorigenesis, tumor cell invasion, metastasis, chemoresistance, and immune response (Figure 5).
5.1. CAF
The reciprocal interaction between tumor cells and fibroblasts plays a crucial role in the progression of breast cancer [104,105]. OPN emerges as a key regulator in facilitating the crosstalk between tumor and stromal fibroblasts, contributing to breast cancer advancement [62,104,106]. Sharon et al demonstrated that OPN has the ability to reprogram normal fibroblasts, inducing a proinflammatory state that supports the growth of breast cancer. In this study, the authors have revealed that the OPN-reprogrammed fibroblasts secrete various proinflammatory cytokines, including CXCL1, CXCL2, COX-2, and IL-6. The ability of OPN to reprogram mammary fibroblasts is found to be reliant on signalling through CD44 and αvβ3 integrin [107]. Butti et al have reported that tumor cell-derived OPN, triggers the differentiation of fibroblasts into CAFs through activation of Twist1 [52]. Mechanistically, OPN induces Twist1-dependent myofibroblastic gene expression such as α-SMA, FSP, FAP, SDF1and PDGFRβ by binding to CD44 and αvβ3 integrins, activating Akt and ERK signalling pathways. OPN-driven CAFs then release CXCL12, inducing EMT and angiogenesis. OPN and CXCL12 are identified as crucial components perpetuating this crosstalk [52]. Costa et al have identified FAP+ CAFs (CAF-S1) subset enriched mainly in TNBC and it induces the immunosuppressive microenvironment by secreting CXCL12 [108]. The induction of FAP and CXCL12 in CAFs by OPN suggested that the reprogramming of CAFs may play an immunosuppressive role through CXCL12 secretion. Moreover, studies also implicated the involvement of OPN autocrine signalling in the generation of CAFs using the loss of function studies in fibroblasts [52,107]. To support this observation, it has been indicated that CAFs actively secrete OPN while it’s silencing in α-SMA+ CAF attenuated the growth of colonies using breast cancer cells [109]. The 3D cell culture and animal model data revealed that blocking CAF-derived OPN effectively prevented lung metastasis in breast cancer. Intriguingly, this study also highlighted a correlation between OPN expression and tumor invasiveness in patient specimens [110]. Collectively, the studies demonstrate that OPN plays an important role in CAF and cancer cell interaction leading to breast cancer progression.
The senescent fibroblasts exhibit characteristics reminiscent of myofibroblasts (CAFs), serving as potential regulators of senescence-associated cancers. Senescent fibroblasts induce preneoplastic growth through OPN [111]. Interestingly, the reduction of OPN levels through RNAi did not affect the induction of senescence in fibroblasts, however, it had a profound impact on diminishing the growth-promoting activities of senescent fibroblasts [112]. Another study has shown that altering Tiam1 expression in senescent fibroblasts induces the invasive and migratory potential, EMT, and CSC characteristics by upregulating OPN in breast cancer cells [110].
Although resident fibroblasts contribute majorly to CAF population, generation of CAFs from different cellular sources including MSCs have been well documented [113]. Tumor-derived OPN has also exhibited a very significant role in the generation of CAFs from MSCs in breast cancer. OPN induces expression of various myofibroblast markers including α-SMA, FSP-1, CXCL12. OPN-educated MSCs also produce higher levels of CXCL5 through integrins and activate c-Jun and OPN-interacted MSCs induce breast cancer metastasis [114]. Moreover, OPN instigates integrin-dependent MSC expression via TGF-β1 to facilitate acquisition of CAF phenotype. OPN activates the TGF-β expression transcriptionally via MZF-1 transcription factor. This study concluded that tumor-derived OPN induces MSC-CAF trans-differentiation to enhance tumor growth and metastasis via the OPN–MZF1–TGF-β1 signalling pathway [59]. These studies show that OPN may be a potential fibrogenic factor in breast tumor microenvironment to facilitate growth and metastasis of cancer cells.
5.2. Adipocytes
Adipose tissue (AT) is the site of inflammatory responses linked to obesity, which trigger a range of cytokines and modify metabolic regulation [115]. It has been demonstrated that OPN causes inflammatory signalling in adipocytes and is primarily produced by macrophages in obese AT [116]. Obese humans and mice exhibit elevated levels of OPN in plasma circulating levels as well as in AT macrophages [117]. MMP-2 and MMP-9 are highly expressed in various cancer cells and targeting MMPs with its inhibitor may act as important therapeutics in cancer [118].
5.3. Osteoclast
Osteoclastic cells have originated from monocytic lineage involved in bone remodelling, bone resorption and ossification [119]. About 15 to 30% of breast cancer patients are prone to metastasized to bone [94]. Zuo et al have shown that neutralization of OPN downregulates Fam20C deficiency which reduces differentiation of osteoclast along with bone metastasis [94]. This study has established a correlation between OPN and osteoclastogenesis and suggested that OPN may act as a potential therapeutic target for breast cancer bone metastasis.
6. OPN in Immunomodulation
Within the heterogenous TME, tumor immune microenvironment (TIME) comprises spatially distributed immune cells such as lymphocytes, monocytes, dendritic cells and macrophages. These cells play a critical role in supporting tumor progression and metastasis via crosstalk with the cancer cells. The role of OPN in immunomodulation has been greatly appreciated in mediating tumor-immune cell interaction and creating an immunosuppressive TIME. OPN can reprogram immune cells such as macrophages to induce tumor growth, angiogenesis and metastasis by secreting pro-tumorigenic cytokines and growth factors. Further, these re-educated immune cells may also secrete OPN which further aids in tumor progression [11]. Besides this, OPN can also induce an immunosuppressive effect in TME by promoting anti- inflammatory macrophages and inhibiting T cell activation via various mechanisms [11].
6.1. OPN Modulates Macrophages into TAMs
Various experimental studies have identified a population of macrophages in TME known as TAMs linked with drug resistance and poor prognosis in various cancers. TAMs are categorized as proinflammatory M1 and anti-inflammatory M2 phenotypes. M1 macrophages are known to induce inflammatory cytokines for the anti-tumor Th1 cell response, whereas M2 macrophages are involved in promoting tumor growth, angiogenesis, metastasis, CSC regulation and immune suppression in cancer by secreting anti-inflammatory cytokines [120].
The unique structure and ability of OPN to bind integrins make it an efficient signalling molecule and predominantly promotes tumor progression by governing macrophage polarisation, activation, migration and immunosuppression [121]. Owing to the evidence that OPN+ (SPP1+) macrophages are critical determinant of tumor progression within TME but its comprehensive regulatory principles underpinning the tumor regulation is remain unclear. OPN induces polarization of macrophages into M2 type, characterized by CD163, CD206 and CD209 expression, to promote tumor progression. It also induces immunosuppressive ques in TME- mediated through the TAMs [6]. The monocytes treated with OPN rich conditioned media from cancer cells leads to an increase in M2 macrophages and co-implanted xenograft of OPN expressing tumor cells and monocytes leads to high tumor growth and poor survival in gastric cancer mouse model [122]. However, this effect of OPN in promoting M2 macrophages may not be universal as treatment of monocyte from healthy donor with recombinant OPN could not enhance the M2 population but it helped in the maintenance of M2 phenotype [123]. Besides promoting M2-type macrophages, OPN also plays a crucial role in recruitment of TAMs in TME by acting as a chemoattractant and inducing TAM migration. A study using OPN KO mice demonstrated that absence of OPN reduced the infiltration of macrophages in tumor while there was no effect in normal tissue [56]. Another report suggested that reduced macrophage infiltration and enhanced T cell activity are observed in OPN deficient in vivo glioma model. OPN deficiency also reduced immune-suppressive regulatory T cells in blood and sensitized glioma cells to direct CD8+ T cell cytotoxicity [123]. Tumor-derived OPN stimulates CSF-1 through activation of PI3K/Akt/p65 signalling leading to the infiltration of macrophages in HCC [124]. Hence, these reports suggested that OPN may induce or maintain M2-type macrophages depending upon pathological conditions and tissue type in cancer.
Single-cell RNA seq (scRNA seq) analyses reported that SPP1 is highly expressed in monocyte-derived TAMs compared to resident macrophages in breast cancer [125]. Moreover, TAM subpopulations were divided into two subtypes (Group 1: high expression of CD204, APOE, C1QA, TREM2, CADM1 and SPP1; Group 2: strong expression of CD206, S100A9 and FCN1) [126]. Similarly, pan cancer scRNA-seq data revealed that SPP1+ TAMs were predominantly enriched in association with EMT, hypoxia, angiogenesis and enhances tumor metastasis [127]. Additionally, single-cell and spatial analyses suggest that FAP+ fibroblasts and SPP1+ macrophages cooperate to create a desmoplastic milieu that hinders lymphocytes from penetrating the tumor core, hence decreasing the effectiveness of PD-L1 therapy [8]. At single cell level, SPP1+ TAM subset regulates gene signatures and serve as a novel characteristic markers for M2 TAM. It is associated with a worst prognosis, poor immune cell infiltration, and decrease immune checkpoint expression.
Lipid-associated macrophages, also known as foamy macrophages, are TAMs that are linked to breast cancer and display an M2-like gene profile, such as CD163 expression, and release different pro-tumor secretory factors [128]. Moreover, OPN has been reported to modulate the expression of IL6 and IL12, suppresses the expression of IL27 in dendritic cells, downregulate the expression of IL10 in monocytes, and function as a chemoattractant cytokine that recruits neutrophils and macrophages [6]. Furthermore, a different study found that the NLRP3 TAM, SPP1 TAM, and IL4I1 TAM niche are strongly linked with tumor nests that includes acute inflammation, hypoxia and diffuse tissue necrosis. Bill et al recently demonstrated the synergistic ratio of CXCL9 and SPP1 in TAMs. This ratio is not only defines TAM (beyond its M1 and M2) polarity but also collectively dictates patient outcomes, anti- tumor potential, immunosuppressive behaviour of TME [129] (Figure 6A).
6.2. Role of OPN in TAM-Mediated Tumor Progression
OPN-mediated interaction between tumor and macrophages in TME plays an important role in the promotion of various hallmarks of cancer including proliferation, angiogenesis, metastasis, evading immune destruction and CSC enrichment. OPN induces tumor fibrosis by acting as a promoter of both TAMs and CAFs [52]. CAFs recruit monocytes in tumor and help in polarization into TAMs whereas TAMs promote activation and proliferation of fibroblasts by secreting TGF-β and PDGFs [130]. OPN acts as a facilitator of these processes in TME. Tokuda et al reported that TAM-derived OPN activates hepatic stellate cells into CAFs and promotes malignancy in HCC [131]. Further, crosstalk between tumor cells and TAMs via OPN leads to tumor growth, angiogenesis and metastasis. The reciprocal crosstalk between TAMs and cancer cells via OPN/CD44 axis advances the tumorigenicity through activation of JNK pathway in colorectal carcinoma [29]. Similarly, Nakajima et al showed that TAM derived OPN acts as a key regulator of cancer progression through interacting with CD44v6 in colorectal cancer [132]. Kale et al demonstrated that tumor-derived OPN promotes macrophage-dependent tube formation ability of HUVEC by inducing Cox-2 expression in macrophages via ERK/p38-dependent signalling pathway in melanoma [56]. Furthermore, ADAM8 enhances angiogenic potential of macrophages by inducing OPN expression via JAK/STAT3 and NF-κB signalling in glioblastoma [133]. TAMs also function as regulators of stem cell enrichment and maintenance and thus contribute to chemotherapy resistance and tumor relapse. Radharani et al demonstrated that macrophages activated by treatment with cancer cell condition media positively regulate CSC- mediated tumor progression by IL-6-dependent activation of Jak/STAT pathway in breast cancer cells [134]. OPN acts as a mediator of crosstalk between TAMs and CSCs and play a significant role in maintenance of stemness in cancer. It has been also demonstrated that TAMs interact with CD44 in CSCs through OPN to regulate CSC-mediated tumor progression by activating the PI3K/Akt in colorectal carcinoma [88].
6.3. Role of OPN in Immune Evasion
Immune evasion is a key hallmark of cancer. Tumors employ various mechanisms to evade immune attack including restricting antigen recognition, inhibiting the immune system, inducing T cell exhaustion and reprogramming immune cells from tumor suppressor to promoter type. Cancer cells hijack inflammatory mechanisms to convert anti-tumor to tumor promoting immune cells that secretes pro-tumor factors that supports tumor growth and metastasis. Cancer cells also express immune checkpoint proteins to induce inhibitory signals leading to suppression of T-cell activity [135]. Cancer cells utilise different molecules and signalling pathways to modulate immune response in TIME. OPN has been identified as one such regulatory molecule which can modulate immune response in favour of tumor promotion. It is predominantly expressed in tumors and perform an important role in immune evasion in cancer. OPN plays a regulatory role in T cell activation, conversion of macrophages from M1 to M2 type and expression of immune checkpoint proteins [136].
6.4. OPN Inhibits T Cell Activation
OPN was initially recognized as a regulator of T cell activation and called an early T cell- activated gene (Eta-1) [3]. OPN modulates the adhesion, migration, and activation of inflammatory cells along with T-cell differentiation to govern the immune response against infection [6]. However, various reports have demonstrated the role of OPN in suppression of T- cell response in cancer. Myeloid-Derived Suppressor Cells (MDSCs), an immature granulocytic or monocytic myeloid cell population are known to suppress both adaptive and innate immune systems by altering T and NK cell functions. Granulocytic MDSCs are predominantly found in tumor and associated with downregulation of interferon regulatory factor-8 (IRF-8) in tumor [137]. OPN is highly expressed in MDSCs and suggested that IRF-8 negatively regulates the expression of OPN in MDSCs. MDSCs and tumor-derived OPN have been shown to suppress T- cell activation and T-cell mediated IFN-γ secretion by binding to CD44 on T-cells leading to poor patient survival [7]. Further, tumor and host-derived OPN rendered an immunosuppressive effect in the lung metastatic model of breast cancer. Interestingly, monocytic MDCSs were found to be the primary source of host-derived OPN [138]. Tumor-derived OPN is also associated with the recruitment of MDSCs at tumor site which might lead to MDSC-mediated suppression of T-cell activity. Allegrezza et al indicated that trametinib, a MEK inhibitor, induces anti-tumor T-cell activation by inhibiting tumor-derived OPN-mediated MDSC recruitment in breast cancer [139].
Further, researchers demonstrated that silencing OPN in colon cancer cells lead to a significant increase in the efficacy of the tumor-specific cytotoxic T cells in killing the target cells suggesting an immunosuppressive role of OPN [140]. Macrophage-mediated immunosuppressive effect of OPN is determined in the lung adenocarcinoma model where co-culture of macrophages with cancer cells led to diminished activity of CD4+ T-cells which was rescued upon OPN depletion [141]. TIME may be categorized into three different categories: (i) inflamed type, (ii) the immune- excluded type, and (iii) the immune-desert type [142]. The role of OPN and its function has been explored in inflamed and immune- excluded TIME where as its function in immune-desert need to be explored further (Figure 6 B).
6.5. OPN Regulates Immune Checkpoints
Immune checkpoint molecules are ligand-receptor complex that exhibit inhibitory or stimulatory effect upon immune responses [143]. Tumor cells regulate immune checkpoints and evade host immune surveillance leading to tumor progression. PD-L1 is one such immune checkpoint molecule on tumor cells which binds with its receptor, PD-1 on T cells and induces T cell dysfunction. It has been demonstrated that TAM derived OPN increased the expression of PD-L1 in NSCLC via NF-κB signalling resulting in suppression of anti-tumor immune response [144]. In another reports, PD-L1 expression was upregulated on HCC by OPN through induction of CSF1-CSF1R pathway in macrophages [145]. Zhang et al showed that OPN upregulates PD- L1 in macrophages to facilitate their polarization and immune escape in lung adenocarcinoma [141]. CRISPR based approaches may be utilised to identify the OPN-driven immune-modulatory genes in breast cancer using scRNA seq platform [129] (Figure 6C).
7. Osteopontin as a Therapeutic Target
Several therapeutic approaches have been reported in targeting OPN including blocking the upstream and downstream pathways, inhibition of OPN expression, immune checkpoint blockade and OPN inhibitors (Figure 7).
7.1. OPN Neutralizing Antibody-Mediated Cancer Therapy
OPN neutralizing antibody or synthetic peptide binds with OPN or its receptor CD44 or αvβ3 to inhibit the OPN-mediated cellular function [146]. A study in breast cancer cohorts showed a reduction in bone metastasis and osteoclast differentiation after the treatment with OPN neutralizing antibodies along with decreased precursors of osteoclast [94]. A humanized OPN antibody, hu1A12 recognizes the epitope N212APSD216 in full-length OPN adjacent to the calcium-binding domain and inhibits cell adhesion and migration in breast cancer [147]. The antibody also exhibited efficacy in reducing the primary tumor growth and spontaneous metastasis in vivo lung metastatic mouse models [147]. The OPN neutralizing antibody attenuates Slug-mediated tumor-enhancing ability in CRC patients [148]. The anti- OPN monoclonal antibody, AOM1 blocks the αvβ3 binding site as well as the thrombin cleavage site of OPN, thus effectively inhibiting OPN-αvβ3 integrin interaction and reducing cell migration in colon cancer [140]. Similarly, AOM1 is shown to prevent the tumor growth in metastatic lesions of NSCLC mouse model while no effect is observed at primary sites [149]. OPN neutralizing monoclonal antibodies (100D3 and 103D6) showed reduced tumor growth by attenuating the interaction of OPN with T-cells in colon cancer model [140]. Blocking the interaction of OPN with αvβ3 integrin resulted in decreased expression of MMP-2, ILK and uPA whereas blocking CD44 interaction resulted in decreased tumor growth in mouse mammary epithelial cancer cells [11]. However, it has been reported that OPN undergoes frequent turnover in healthy human cohorts. The high concentration of OPN in plasma, coupled with its rapid turnover, underscores the demand for enhanced therapeutic antibodies targeting OPN. These antibodies should facilitate high-dose administration within short time interval and exhibit extended pharmacokinetics compared to conventional antibodies. Thus, antibodies targeting OPN receptors, CD44 or integrin can be administered for OPN-targeted cancer therapy [146].
7.2. Small Molecule Inhibitors as a Potential Therapeutic Agent
Owing to the small volume and ease in accessibility to tumor sites, small molecule inhibitors have gained much importance in this current era of cancer therapeutics. One of the well- known small molecules, andrographolide exhibits its activity via suppressing c-Jun and down-regulating the PI3K/Akt signaling pathway thereby abrogating the OPN expression and reducing the breast tumor growth [150]. Bandopadhyay et al have reported that the expression of OPN is inhibited by usage of small molecules, 3-hydroxy-3-methylglutaryl CoA reductase (HMGR) inhibitor and parecoxib, a Cox-2 inhibitor in the treatment of ovarian cancer and colorectal cancer respectively [11]. It has been demonstrated that luteolin and curcumin suppresses the OPN expression in HCC and prostate cancer models respectively, where curcumin attenuates the VEGF expression through OPN/αvβ3 pathway [45,151]. Additionally, administration of bisphosphonate resulted in decreased expression of CD44/MMP-9 as well as reduced migration in prostate cancer cells. Bisphosphonate inhibits the Rho GTPase activity through disrupting the interaction of OPN with αvβ3 integrin [152].
7.3. Epigenetic Approaches
Epigenetic alterations can influence the growth and development of healthy cells, leading to neoplastic transformation. Interestingly, WDR5-H3K4me3 epigenetic axis modulates OPN expression leading to tumor immune evasion and anti-PD-1 immunotherapy escape in pancreatic cancer [153]. In addition, OPN induces DNA methylation via DNMT1 and renders the CD133+/CD44+ CSC subpopulation more sensitive to 5-azacytidine in HCC. Thus, the OPN-DNMT1 axis promotes aberrant DNA methylation while inhibiting CD133+/CD44+ sphere formation and migration. These findings suggest that OPN could be an appealing target for HCC resistance through methylome reprogramming. [154]. Bromodomain and extra-terminal domain (BET) inhibitors impeded cell proliferation, invasion and migration via decreased OPN expression through transcriptional inactivation of NF-кB2 [155]. It is also observed that miR-181c downregulates the expression of OPN thereby enhancing the chemosensitivity to adriamycin and decreased chemoresistance in breast cancer cells [156]. OPN siRNA encapsulated nanoparticle significantly downregulated OPN mRNA level along with enhanced inhibition of tumor growth in mouse mammary carcinoma model [157]. miR-196a knockout showed decreased expression of OPN and reduced lung metastasis in HCC [158]. Therefore, the identification and combination of epigenetic modulators of OPN with the conventional chemotherapeutic agents pose great potential in preventing tumor growth and recurrence.
7.4. OPN Aptamer
Aptamers are highly stable, 12-30 short ssRNA nucleotide sequences that are capable of adopting 3D structure of the target molecule so as to precisely bind with the protein ligands or small molecules [159]. The well-known OPN aptamer, OPN-R3 is tailored to bind with OPN and exhibited decrease in the cellular migration, invasion and adhesion in MDA-MB-231 cells [160]. In addition, the modified OPN-R3 aptamer has shown to reduce tumor growth using in vivo model [161].
7.5. Biomarker
Biomarkers constitute an evolving and dynamic strategy within the realm of cancer research. Numerous discoveries in biomarker identification have significantly contributed to the realms of cancer diagnosis, assessment of cancer progression and monitoring of recurrence risk post-treatment.
Interestingly, it is found that expression of OPN is a predictable biomarker not only for breast cancer but also for NSCLC, HCC, gastric, prostate and colorectal cancer. Levels of OPN expression can be effectively used to evaluate the histological grade of tumor, clinical stage, response to treatment, risk of recurrence after surgery, overall survival, and disease-free survival [10]. OPN as a prognostic and diagnostic biomarker in various cancer types are summarized in Table 2.
8. Conclusion and Future Perspective
OPN emerges as a pivotal regulator in the intricate interplay between cancer cells and the TME, exerting influence across various facets of cancer progression. The abundance of OPN within the TME is responsible for modulating the fate of tumor and stromal cells but yet its clinical relevance remains to be a major limiting factor. Our earlier reports have dissected several OPN- modulated fundamental signalling pathways in breast and other cancers [3,11,52,81]. Furthermore, its multifaceted role encompasses angiogenesis, CSCs, bone metastasis, cancer cell metabolism, and modulation of the TME, involving interactions with CAFs, adipocytes, osteoclasts and immune cells. In addition, OPN intricately regulates cancer cell metabolism, particularly in glucose metabolism, influencing glucose transporters and contributing to the Warburg effect [98]. Moreover, OPN plays a central role by affecting the bone microenvironment, promoting bone metastasis, and contributing to the formation of premetastatic niches [85]. Thus, OPN acts as a potential therapeutic target for mitigating osteoclast development and controlling skeletal metastases.
In TME, the influence of OPN spans interactions with CAFs, adipocytes and osteoclasts. OPN-reprogrammed CAFs contribute to proinflammatory states, angiogenesis, and EMT, emphasizing its role in cancer progression [52]. OPN significantly impacts immunomodulation within the TME, influencing macrophage polarization towards a tumor-promoting phenotype (M2 type) and contributing to an immunosuppressive microenvironment. Moreover, OPN is implicated in immune evasion mechanisms, including the inhibition of T-cell activation and the regulation of immune checkpoint molecules such as PD-L1. CAF and TAM interaction are majorly regulated by OPN and its associated signalling events causing upregulation of various tumor events. For example, scRNA-seq revealed that OPN mediates the interaction between OPN-PTGER4 and OPN-CD44 stimulating the crosstalk between HCC cells and macrophages [162]. Similarly, single-cell transcriptome sequencing identified three TAMs subgroups: C1Q+, FCN1+ and OPN (SPP1)+ TAMs whereas OPN (SPP1)+ TAMs modulate TIME via interaction with CAFs [163]. Zhang et al reported that OPN+ TAMs are tightly associated with CAFs and endothelial cells in modulating TME [164]. However, the complex molecular mechanism of OPN mediated TAM subset enrichment, regulation of metabolic switch and intricate crosstalk between CAF-TAM is poorly elucidated. Although, multiple studies have demonstrated the function of CAFs in regulation of TAM, comprehending the influence of TAM in controlling CAF phenotypes warrant further investigation.
Using single-cell and spatial transcriptomics, immunofluorescent labelling, and other methods, have advanced our understanding the role of OPN in inflamed and immune-excluded TME. However, its role in immune-desert needs further exploration [129]. Detailed mechanistic studies on OPN-regulated networks and core regulatory transcription factors governing OPN expression in tumor and immune cells are warranted.
Considering the central role of OPN in cancer progression, targeted therapies aimed at inhibiting OPN expression or its downstream signalling pathways hold promise. However, current interventions lack efficacy in both preclinical and clinical trials, necessitating further development of small molecules or antibodies to neutralize the effect of OPN. Stratifying breast cancer patient based on expression levels of OPN and associated signalling pathways could tailor treatment strategies and identify the patient subgroups more responsive to OPN-targeted therapies. Understanding the immunomodulatory role of OPN suggests exploring combination therapies involving OPN inhibition and immunotherapeutic agents, potentially enhancing the effectiveness of immune checkpoint inhibitors and promoting antitumor immune responses.
In summary, the diverse function of OPN in breast and other cancers underscore its significance as a potential therapeutic target and diagnostic marker. Thus, the continued research on role of OPN and its associated signalling networks will unravel new insights in developing innovative therapeutic strategies for precision medicine, ultimately improving patient outcomes in various cancers including breast.
Author Contributions
The authors (GCK, VKP, RB, DM, ANN, ASY, BM, SK, PM, SM, KK, SM, BS and NNVR) wrote the manuscript and prepared the figures and tables together. GCK, VKP, BM, ANN and RB edited the entire manuscript.
Funding
This work was supported by Science and Engineering Research Board (SERB), Govt. of India, (Project No. JCB/2023/000011) to GCK; Department of Biotechnology (DBT) (Project No. BT/PR-32388/TRM/120/242/2019), Govt of India to GCK and DST INSPIRE Fellowship Program (DST INSPIRE /2021/IF210059), Govt of India to VKP.
Acknowledgments
This work was supported by Science and Engineering Research Board (SERB), Govt. of India, (Project No. JCB/2023/000011) to GCK; Department of Biotechnology (DBT) (Project No. BT/PR-32388/TRM/120/242/2019), Govt of India to GCK and DST INSPIRE Fellowship Program (DST INSPIRE /2021/IF210059), Govt of India to VKP. Figures were created using BioRender.com and accessed with license.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021, 71, 209-249. [CrossRef]
- Mao X, Xu J, Wang W, Liang C, Hua J, Liu J et al. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: new findings and future perspectives. Mol Cancer. 2021, 20, 1-30. [CrossRef]
- Rangaswami H, Bulbule A, Kundu GC. Osteopontin: role in cell signaling and cancer progression. Trends Cell Biol. 2006, 16, 79-87. [CrossRef]
- Cho H-J, Cho H-J, Kim H-S. Osteopontin: a multifunctional protein at the crossroads of inflammation, atherosclerosis, and vascular calcification. Curr Atheroscler Rep. 2009, 11, 206-213. [CrossRef]
- Zhao H, Chen Q, Alam A, Cui J, Suen KC, Soo AP et al. The role of osteopontin in the progression of solid organ tumour. Cell Death Dis. 2018, 9, 1-15. [CrossRef]
- Moorman HR, Poschel D, Klement JD, Lu C, Redd PS, Liu K. Osteopontin: a key regulator of tumor progression and immunomodulation. Cancers. 2020, 12, 3379. [CrossRef]
- Klement JD, Paschall AV, Redd PS, Ibrahim ML, Lu C, Yang D et al. An osteopontin/CD44 immune checkpoint controls CD8+ T cell activation and tumor immune evasion. J Clin Invest. 2018, 128, 5549-5560.
- Qi J, Sun H, Zhang Y, Wang Z, Xun Z, Li Z et al. Single-cell and spatial analysis reveal interaction of FAP+ fibroblasts and SPP1+ macrophages in colorectal cancer. Nat Commun. 2022, 13, 1742. [CrossRef]
- Bellahcène A, Castronovo V, Ogbureke KU, Fisher LW, Fedarko NS. Small integrin-binding ligand N-linked glycoproteins (SIBLINGs): multifunctional proteins in cancer. Nat Rev Cancer. 2008, 8, 212-226. [CrossRef]
- Icer MA, Gezmen-Karadag M. The multiple functions and mechanisms of osteopontin. Clin Biochem. 2018, 59, 17-24. [CrossRef]
- Bandopadhyay M, Bulbule A, Butti R, Chakraborty G, Ghorpade P, Ghosh P et al. Osteopontin as a therapeutic target for cancer. Expert Opin Ther Targets. 2014, 18, 883-895. [CrossRef]
- Lok ZSY, Lyle AN. Osteopontin in vascular disease: Friend or foe? Arterioscler Thromb Vasc Biol. 2019, 39, 613-622.
- Briones-Orta MA, Avendaño-Vázquez SE, Aparicio-Bautista DI, Coombes JD, Weber GF, Syn W-K. Osteopontin splice variants and polymorphisms in cancer progression and prognosis. Biochim Biophys Acta Rev Cancer. 2017, 1868, 93-108. [CrossRef]
- Shinohara ML, Kim H-J, Kim J-H, Garcia VA, Cantor H. Alternative translation of osteopontin generates intracellular and secreted isoforms that mediate distinct biological activities in dendritic cells. Proc Natl Acad Sci U S A. 2008, 105, 7235-7239. [CrossRef]
- Kariya Y, Kariya Y. Osteopontin in cancer: mechanisms and therapeutic targets. Int J Transl Med. 2022, 2, 419-447.
- Kruger TE, Miller AH, Godwin AK, Wang J. Bone sialoprotein and osteopontin in bone metastasis of osteotropic cancers. Crit Rev Oncol Hematol. 2014, 89, 330-341. [CrossRef]
- Desgrosellier JS, Cheresh DA. Integrins in cancer: biological implications and therapeutic opportunities. Nat Rev Cancer. 2010, 10, 9-22. [CrossRef]
- Liu Z, Wang F, Chen X. Integrin αvβ3-targeted cancer therapy. Drug Dev Res. 2008, 69, 329-339.
- Ludwig BS, Kessler H, Kossatz S, Reuning U. RGD-binding integrins revisited: how recently discovered functions and novel synthetic ligands (re-) shape an ever-evolving field. Cancers. 2021, 13, 1711. [CrossRef]
- Zargham R, Wamhoff BR, Thibault G. RNA interference targeting α8 integrin attenuates smooth muscle cell growth. FEBS Lett. 2007, 581, 939-943. [CrossRef]
- Baiula M, Spampinato S, Gentilucci L, Tolomelli A. Novel ligands targeting α4β1 integrin: Therapeutic applications and perspectives. Front Chem. 2019, 7, 489. [CrossRef]
- Wei R, Wong JPC, Kwok HF. Osteopontin--a promising biomarker for cancer therapy. J Cancer. 2017, 8, 2173. [CrossRef]
- Senbanjo LT, Chellaiah MA. CD44: a multifunctional cell surface adhesion receptor is a regulator of progression and metastasis of cancer cells. Front Cell Dev Biol. 2017, 5, 18. [CrossRef]
- Hassn Mesrati M, Syafruddin SE, Mohtar MA, Syahir A. CD44: A multifunctional mediator of cancer progression. Biomolecules. 2021, 11, 1850. [CrossRef]
- Katagiri YU, Sleeman J, Fujii H, Herrlich P, Hotta H, Tanaka K et al. CD44 variants but not CD44s cooperate with β1-containing integrins to permit cells to bind to osteopontin independently of arginine-glycine-aspartic acid, thereby stimulating cell motility and chemotaxis. Cancer Res. 1999, 59, 219-226.
- Fnu G, Agrawal P, Kundu GC, Weber GF. Structural Constraint of Osteopontin Facilitates Efficient Binding to CD44. Biomolecules. 2021, 11, 813. [CrossRef]
- Pietras A, Katz AM, Ekström EJ, Wee B, Halliday JJ, Pitter KL et al. Osteopontin-CD44 signaling in the glioma perivascular niche enhances cancer stem cell phenotypes and promotes aggressive tumor growth. Cell Stem Cell. 2014, 14, 357-369. [CrossRef]
- Ahmed M, Sottnik JL, Dancik GM, Sahu D, Hansel DE, Theodorescu D et al. An osteopontin/CD44 axis in RhoGDI2-mediated metastasis suppression. Cancer Cell. 2016, 30, 432- 443. [CrossRef]
- Rao G, Wang H, Li B, Huang L, Xue D, Wang X et al. Reciprocal interactions between tumor- associated macrophages and CD44-positive cancer cells via osteopontin/CD44 promote tumorigenicity in colorectal cancer. Clin Cancer Res. 2013, 19, 785-797. [CrossRef]
- Cheng Y, Wen G, Sun Y, Shen Y, Zeng Y, Du M et al. Osteopontin promotes colorectal cancer cell invasion and the stem cell-like properties through the PI3K-AKT-GSK/3β-β/catenin pathway. Med Sci Monit. 2019, 25, 3014. [CrossRef]
- Zhang H, Guo M, Chen J-h, Wang Z, Du X-f, Liu P-x et al. Osteopontin knockdown inhibits αv, β3 integrin-induced cell migration and invasion and promotes apoptosis of breast cancer cells by inducing autophagy and inactivating the PI3K/Akt/mTOR pathway. Cell Physiol Biochem. 2014, 33, 991-1002.
- Dos Santos ES, Ramos JC, Roza ALOC, Mariz BALA, Paes Leme AF. The role of osteopontin in oral cancer: a brief review with emphasis on clinical applications. Oral Dis. 2022, 28, 326-335.
- Fan C-S, Chen W-S, Chen L-L, Chen C-C, Hsu Y-T, Chua KV et al. Osteopontin–integrin engagement induces HIF-1α–TCF12-mediated endothelial-mesenchymal transition to exacerbate colorectal cancer. Oncotarget. 2018, 9, 4998. [CrossRef]
- Zhang G, Zhao Z, Lin X. EGF/PI3K signaling pathway regulates the expression of osteopontin in liver cancer HepG2 cells. Zhonghua Yi Xue Za Zhi. 2003, 83, 1980-1983.
- Yu X, Zheng Y, Zhu X, Gao X, Wang C, Sheng Y et al. Osteopontin promotes hepatocellular carcinoma progression via the PI3K/AKT/Twist signaling pathway. Oncol Lett. 2018, 16, 5299- 5308. [CrossRef]
- Qin Y-C, Yan X, Yuan X-L, Yu W-W, Qu F-J. Osteopontin promotes gastric cancer progression via phosphatidylinositol-3-kinase/protein kinase B/mammalian target of rapamycin signaling pathway. World J Gastrointest Oncol. 2023, 15, 1544. [CrossRef]
- Chen Y, Wang G, Wang Y, Gao X, Wang K, Li J et al. Capn4 regulates migration and invasion of ovarian carcinoma cells via targeting osteopontin-mediated PI3K/AKT signaling pathway. Oncol Lett. 2019, 17, 564-570. [CrossRef]
- Liu D, Luo M, Hu J, Chen C, Mei H. Osteopontin enhances cisplatin resistance of human A549 lung cancer cells via stimulating the PI3K signaling pathway and upregulating ERCC1 expression. Transl Cancer Res. 2020, 9, 3258. [CrossRef]
- Ding K, Fan L, Chen S, Wang Y, Yu H, Sun Y et al. Overexpression of osteopontin promotes resistance to cisplatin treatment in HCC. Oncol Rep. 2015, 34, 3297-3303. [CrossRef]
- Chen J, Shi L, Qian Y, Jin Y, Dong N, Chen C et al. Epithelial-mesenchymal transition is associated with osteopontin-induced EGFR-TKI resistance in EGFR mutant non-small cell lung cancer. J Thorac Dis. 2023, 15, 3359. [CrossRef]
- Kumar D, Haldar S, Gorain M, Kumar S, Mulani FA, Yadav AS et al. Epoxyazadiradione suppresses breast tumor growth through mitochondrial depolarization and caspase-dependent apoptosis by targeting PI3K/Akt pathway. BMC Cancer. 2018, 18, 1-17. [CrossRef]
- Tilli TM, Franco VF, Robbs BK, Wanderley JLM, de Azevedo da Silva FR, de Mello KD et al. Osteopontin-c splicing isoform contributes to ovarian cancer progression. Mol Cancer Res. 2011, 9, 280-293. [CrossRef]
- Shi L, Hou J, Wang L, Fu H, Zhang Y, Song Y et al. Regulatory roles of osteopontin in human lung cancer cell epithelial-to-mesenchymal transitions and responses. Clin Transl Med. 2021, 11, e486. [CrossRef]
- Sun BS, Dong QZ, Ye QH, Sun HJ, Jia HL, Zhu XQ et al. Lentiviral-mediated miRNA against osteopontin suppresses tumor growth and metastasis of human hepatocellular carcinoma. Hepatology. 2008, 48, 1834-1842. [CrossRef]
- Gupta A, Zhou CQ, Chellaiah MA. Osteopontin and MMP9: associations with VEGF expression/secretion and angiogenesis in PC3 prostate cancer cells. Cancers. 2013, 5, 617-638. [CrossRef]
- Song G, Ouyang G, Mao Y, Ming Y, Bao S, Hu T. Osteopontin promotes gastric cancer metastasis by augmenting cell survival and invasion through Akt-mediated HIF-1α up-regulation and MMP9 activation. J Cell Mol Med. 2009, 13, 1706-1718.
- Pang H, Cai L, Yang Y, Chen X, Sui G, Zhao C. Knockdown of osteopontin chemosensitizes MDA- MB-231 cells to cyclophosphamide by enhancing apoptosis through activating p38 MAPK pathway. Cancer Biother Radiopharm. 2011, 26, 165-173. [CrossRef]
- Kyjacova L, Saup R, Rönsch K, Wallbaum S, Dukowic-Schulze S, Foss A et al. IER2-induced senescence drives melanoma invasion through osteopontin. Oncogene. 2021, 40, 6494-6512. [CrossRef]
- Graessmann M, Berg B, Fuchs B, Klein A, Graessmann A. Chemotherapy resistance of mouse WAP-SVT/t breast cancer cells is mediated by osteopontin, inhibiting apoptosis downstream of caspase-3. Oncogene. 2007, 26, 2840-2850. [CrossRef]
- Belli S, Esposito D, Servetto A, Pesapane A, Formisano L, Bianco R. c-Src and EGFR inhibition in molecular cancer therapy: what else can we improve? Cancers. 2020, 12, 1489. [CrossRef]
- Li NY, Weber CE, Mi Z, Wai PY, Cuevas BD, Kuo PC. Osteopontin up-regulates critical epithelial- mesenchymal transition transcription factors to induce an aggressive breast cancer phenotype. J Am Coll Surg. 2013, 217, 17-26. [CrossRef]
- Butti R, Nimma R, Kundu G, Bulbule A, Kumar TV, Gunasekaran VP et al. Tumor-derived osteopontin drives the resident fibroblast to myofibroblast differentiation through Twist1 to promote breast cancer progression. Oncogene. 2021, 40, 2002-2017. [CrossRef]
- Li NY, Weber CE, Wai PY, Cuevas BD, Zhang J, Kuo PC et al. An MAPK-dependent pathway induces epithelial-mesenchymal transition via Twist activation in human breast cancer cell lines. Surgery. 2013, 154, 404-410. [CrossRef]
- Shen Q, Christakos S. The vitamin D receptor, Runx2, and the Notch signaling pathway cooperate in the transcriptional regulation of osteopontin. J Biol Chem. 2005, 280, 40589-40598. [CrossRef]
- Chua H, Bhat-Nakshatri P, Clare S, Morimiya A, Badve S, Nakshatri H. NF-κB represses E- cadherin expression and enhances epithelial to mesenchymal transition of mammary epithelial cells: potential involvement of ZEB-1 and ZEB-2. Oncogene. 2007, 26, 711-724.
- Kale S, Raja R, Thorat D, Soundararajan G, Patil T, Kundu G. Osteopontin signaling upregulates cyclooxygenase-2 expression in tumor-associated macrophages leading to enhanced angiogenesis and melanoma growth via α9β1 integrin. Oncogene. 2014, 33, 2295-2306. [CrossRef]
- Takuwa Y, Ikeda H, Okamoto Y, Takuwa N, Yoshioka K. Sphingosine-1-phosphate as a mediator involved in development of fibrotic diseases. Biochim Biophys Acta Mol Cell Biol Lipids. 2013, 1831, 185-192. [CrossRef]
- Hu D, Li Z, Zheng B, Lin X, Pan Y, Gong P et al. Cancer-associated fibroblasts in breast cancer: Challenges and opportunities. Cancer Commun (Lond). 2022, 42, 401-434. [CrossRef]
- Weber CE, Kothari AN, Wai PY, Li NY, Driver J, Zapf MA et al. Osteopontin mediates an MZF1– TGF-β1-dependent transformation of mesenchymal stem cells into cancer-associated fibroblasts in breast cancer. Oncogene. 2015, 34, 4821-4833. [CrossRef]
- Jia R, Liang Y, Chen R, Liu G, Wang H, Tang M et al. Osteopontin facilitates tumor metastasis by regulating epithelial–mesenchymal plasticity. Cell Death Dis. 2016, 7, e2564-e2564. [CrossRef]
- Butti R, Gunasekaran VP, Kumar TV, Banerjee P, Kundu GC. Breast cancer stem cells: Biology and therapeutic implications. Int J Biochem Cell Biol. 2019, 107, 38-52. [CrossRef]
- Butti R, Kumar TV, Nimma R, Banerjee P, Kundu IG, Kundu GC. Osteopontin signaling in shaping tumor microenvironment conducive to malignant progression. Adv Exp Med Biol. 2021, 1329, 419-441.
- Yoshida GJ, Saya H. Molecular pathology underlying the robustness of cancer stem cells. Regen Ther. 2021, 17, 38-50. [CrossRef]
- Chen C, Zhao S, Karnad A, Freeman JW. The biology and role of CD44 in cancer progression: therapeutic implications. J Hematol Oncol. 2018, 11, 1-23. [CrossRef]
- Pio GM, Xia Y, Piaseczny MM, Chu JE, Allan AL. Soluble bone-derived osteopontin promotes migration and stem-like behavior of breast cancer cells. PLoS One. 2017, 12, e0177640. [CrossRef]
- Sun X, Li K, Hase M, Zha R, Feng Y, Li B-Y et al. Suppression of breast cancer-associated bone loss with osteoblast proteomes via Hsp90ab1/moesin-mediated inhibition of TGFβ/FN1/CD44 signaling. Theranostics. 2022, 12, 929.
- Hu J, Li G, Zhang P, Zhuang X, Hu G. A CD44v+ subpopulation of breast cancer stem-like cells with enhanced lung metastasis capacity. Cell Death Dis. 2017, 8, e2679-e2679. [CrossRef]
- Nikolaou M, Pavlopoulou A, Georgakilas AG, Kyrodimos E. The challenge of drug resistance in cancer treatment: a current overview. Clin Exp Metastasis. 2018, 35, 309-318. [CrossRef]
- Mansoori B, Mohammadi A, Davudian S, Shirjang S, Baradaran B. The different mechanisms of cancer drug resistance: a brief review. Adv Pharm Bull. 2017, 7, 339. [CrossRef]
- Ji X, Lu Y, Tian H, Meng X, Wei M, Cho WC. Chemoresistance mechanisms of breast cancer and their countermeasures. Biomed Pharmacother. 2019, 114, 108800. [CrossRef]
- Gu M, Zheng X. Osteopontin and vasculogenic mimicry formation are associated with response to neoadjuvant chemotherapy in advanced breast cancer. Onco Targets Ther. 2017, 10, 4121-4127. [CrossRef]
- Kumar S, Sharma P, Kumar D, Chakraborty G, Gorain M, Kundu GC. Functional characterization of stromal osteopontin in melanoma progression and metastasis. PloS One. 2013, 8, e69116. [CrossRef]
- Hsieh I-S, Huang W-H, Liou H-C, Chuang W-J, Yang R-S, Fu W-M. Upregulation of drug transporter expression by osteopontin in prostate cancer cells. Mol Pharmacol. 2013, 83, 968-977. [CrossRef]
- Yi H, Zeng D, Shen Z, Liao J, Wang X, Liu Y et al. Integrin alphavbeta3 enhances β-catenin signaling in acute myeloid leukemia harboring Fms-like tyrosine kinase-3 internal tandem duplication mutations: Implications for microenvironment influence on sorafenib sensitivity. Oncotarget. 2016, 7, 40387.
- Liu G, Fan X, Tang M, Chen R, Wang H, Jia R et al. Osteopontin induces autophagy to promote chemo-resistance in human hepatocellular carcinoma cells. Cancer Lett. 2016, 383, 171-182. [CrossRef]
- Sui H, Zhu L, Deng W, Li Q. Epithelial-mesenchymal transition and drug resistance: role, molecular mechanisms, and therapeutic strategies. Oncol Res Treat. 2014, 37, 584-589. [CrossRef]
- Das S, Samant RS, Shevde LA. Nonclassical activation of Hedgehog signaling enhances multidrug resistance and makes cancer cells refractory to Smoothened-targeting Hedgehog inhibition. J Biol Chem. 2013, 288, 11824-11833. [CrossRef]
- Chakraborty G, Jain S, Behera R, Ahmed M, Sharma P, Kumar V et al. The multifaceted roles of osteopontin in cell signaling, tumor progression and angiogenesis. Curr Mol Med. 2006, 6, 819- 830. [CrossRef]
- Krstic M, Hassan HM, Kolendowski B, Hague MN, Anborgh PH, Postenka CO et al. Isoform- specific promotion of breast cancer tumorigenicity by TBX3 involves induction of angiogenesis. Lab Invest. 2020, 100, 400-413. [CrossRef]
- Yang Y-F, Chang Y-C, Jan Y-H, Yang C-J, Huang M-S, Hsiao M. Squalene synthase promotes the invasion of lung cancer cells via the osteopontin/ERK pathway. Oncogenesis. 2020, 9, 78. [CrossRef]
- Chakraborty G, Jain S, Kundu GC. Osteopontin promotes vascular endothelial growth factor– dependent breast tumor growth and angiogenesis via autocrine and paracrine mechanisms. Cancer Res. 2008, 68, 152-161. [CrossRef]
- Raja R, Kale S, Thorat D, Soundararajan G, Lohite K, Mane A et al. Hypoxia-driven osteopontin contributes to breast tumor growth through modulation of HIF1α-mediated VEGF-dependent angiogenesis. Oncogene. 2014, 33, 2053-2064. [CrossRef]
- Raineri D, Dianzani C, Cappellano G, Maione F, Baldanzi G, Iacobucci I et al. Osteopontin binds ICOSL promoting tumor metastasis. Commun Biol. 2020, 3, 615. [CrossRef]
- Jiang X, Zhang X, Jiang N, Sun Y, Li T, Zhang J et al. The single-cell landscape of cystic echinococcosis in different stages provided insights into endothelial and immune cell heterogeneity. Front Immunol. 2022, 13, 1067338. [CrossRef]
- Wai PY, Kuo PC. Osteopontin: regulation in tumor metastasis. Cancer Metastasis Rev. 2008, 27, 103-118. [CrossRef]
- Rizwan A, Paidi SK, Zheng C, Cheng M, Barman I, Glunde K. Mapping the genetic basis of breast microcalcifications and their role in metastasis. Sci Rep. 2018, 8, 11067. [CrossRef]
- Xu C, Yuan Q, Wang W, Chi C, Zhang Q, Li L et al. Prognostic significance of serum osteopontin levels in small cell lung cancer. BMC Pulm Med. 2020, 20, 1-7. [CrossRef]
- Todaro M, Gaggianesi M, Catalano V, Benfante A, Iovino F, Biffoni M et al. CD44v6 is a marker of constitutive and reprogrammed cancer stem cells driving colon cancer metastasis. Cell Stem Cell. 2014, 14, 342-356. [CrossRef]
- Gu X, Gao X-S, Ma M, Qin S, Qi X, Li X et al. Prognostic significance of osteopontin expression in gastric cancer: a meta-analysis. Oncotarget. 2016, 7, 69666. [CrossRef]
- Dong QZ, Zhang XF, Zhao Y, Jia HL, Zhou HJ, Dai C et al. Osteopontin promoter polymorphisms at locus-443 significantly affect the metastasis and prognosis of human hepatocellular carcinoma. Hepatology. 2013, 57, 1024-1034. [CrossRef]
- Li Y, Du W, Han J, Ge J. LAMP3 promotes the invasion of osteosarcoma cells via SPP1 signaling. Mol Med Rep. 2017, 16, 5947-5953. [CrossRef]
- Guo J, Tong C-Y, Shi J-G, Li X-J, Chen X-Q. Deletion of osteopontin in non-small cell lung cancer cells affects bone metabolism by regulating miR-34c/Notch1 axis: a clue to bone metastasis. Eur J Histochem. 2023, 67, 3631. [CrossRef]
- Kovacheva M, Zepp M, Schraad M, Berger S, Berger MR. Conditional knockdown of osteopontin inhibits breast cancer skeletal metastasis. Int J Mol Sci. 2019, 20, 4918. [CrossRef]
- Zuo H, Yang D, Wan Y. Fam20C regulates bone resorption and breast cancer bone metastasis through osteopontin and BMP4. Cancer Res. 2021, 81, 5242-5254. [CrossRef]
- Li X-Q, Lu J-T, Tan C-C, Wang Q-S, Feng Y-M. RUNX2 promotes breast cancer bone metastasis by increasing integrin α5-mediated colonization. Cancer Lett. 2016, 380, 78-86. [CrossRef]
- Kuo MC, Kothari AN, Kuo PC, Mi Z. Cancer stemness in bone marrow micrometastases of human breast cancer. Surgery. 2018, 163, 330-335. [CrossRef]
- Pavlova NN, Zhu J, Thompson CB. The hallmarks of cancer metabolism: Still emerging. Cell Metab. 2022,34, 355-377. [CrossRef]
- Lei P, Wang W, Sheldon M, Sun Y, Yao F, Ma L. Role of glucose metabolic reprogramming in breast cancer progression and drug resistance. Cancers (Basel). 2023, 15, 3390. [CrossRef]
- Hsieh I-S, Yang R-S, Fu W-M. Osteopontin upregulates the expression of glucose transporters in osteosarcoma cells. PLoS One. 2014, 9, e109550. [CrossRef]
- Bui BP, Nguyen PL, Lee K, Cho J. Hypoxia-inducible factor-1: a novel therapeutic target for the management of cancer, drug resistance, and cancer-related pain. Cancers (Basel). 2022, 14, 6054. [CrossRef]
- Nishikawa M, Inoue A, Ohnishi T, Yano H, Ozaki S, Kanemura Y et al. Hypoxia-induced phenotypic transition from highly invasive to less invasive tumors in glioma stem-like cells: Significance of CD44 and osteopontin as therapeutic targets in glioblastoma. Transl Oncol. 2021, 14, 101137. [CrossRef]
- Zeng K, Ju G, Wang H, Huang J. GLUT1/3/4 as novel biomarkers for the prognosis of human breast cancer. Transl Cancer Res. 2020, 9, 2363.
- Szablewski L. Glucose transporters as markers of diagnosis and prognosis in cancer diseases. Oncol Rev. 2022, 16, 561. [CrossRef]
- Butti R, Kundu GC. The molecular dialogue between the tumor cells and fibroblasts. Oncotarget. 2023, 14, 462. [CrossRef]
- Butti R, Khaladkar A, Bhardwaj P, Prakasam G. Heterotypic signaling of cancer-associated fibroblasts in shaping the cancer cell drug resistance. Cancer Drug Resist. 2023, 6, 182-204. [CrossRef]
- Butti R, Ghosh P, Totakura KV, Venkata RNN, Nimma R, Kundu GC. Role of osteopontin in tumor microenvironment: a new paradigm in cancer therapy. Multi-Targeted Approach to Treatment of Cancer. Springer, 2015, pp 113-125.
- Sharon Y, Raz Y, Cohen N, Ben-Shmuel A, Schwartz H, Geiger T, Erez N. Tumor-derived osteopontin reprograms normal mammary fibroblasts to promote inflammation and tumor growth in breast cancer. Cancer Res. 2015, 75, 963-973. [CrossRef]
- Costa A, Kieffer Y, Scholer-Dahirel A, Pelon F, Bourachot B, Cardon M et al. Fibroblast heterogeneity and immunosuppressive environment in human breast cancer. Cancer Cell. 2018, 33, 463-479. e410. [CrossRef]
- Muchlińska A, Nagel A, Popęda M, Szade J, Niemira M, Zieliński J et al. Alpha-smooth muscle actin-positive cancer-associated fibroblasts secreting osteopontin promote growth of luminal breast cancer. Cell Mol Biol Lett. 2022, 27, 1-14. [CrossRef]
- Liu J, Xu K, Chase M, Ji Y, Logan JK, Buchsbaum RJ. Tiam1-regulated osteopontin in senescent fibroblasts contributes to the migration and invasion of associated epithelial cells. J Cell Sci. 2012, 125, 376-386. [CrossRef]
- Mori JO, Elhussin I, Brennen WN, Graham MK, Lotan TL, Yates CC et al. Prognostic and therapeutic potential of senescent stromal fibroblasts in prostate cancer. Nat Rev Urol. 2023, 1-16. [CrossRef]
- Krtolica A, Parrinello S, Lockett S, Desprez P-Y, Campisi J. Senescent fibroblasts promote epithelial cell growth and tumorigenesis: a link between cancer and aging. Proc Natl Acad Sci U S A. 2001, 98, 12072-12077. [CrossRef]
- Louault K, Li R-R, DeClerck YA. Cancer-associated fibroblasts: understanding their heterogeneity. Cancers (Basel). 2020, 12, 3108. [CrossRef]
- Mi Z, Bhattacharya SD, Kim VM, Guo H, Talbot LJ, Kuo PC. Osteopontin promotes CCL5- mesenchymal stromal cell-mediated breast cancer metastasis. Carcinogenesis. 2011, 32, 477-487. [CrossRef]
- Belanger C, Dupont P, Tchernof A. Adipose tissue intracrinology: potential importance of local androgen/estrogen metabolism in the regulation of adiposity. Horm Metab Res. 2002, 34, 737-745. [CrossRef]
- Kiefer FW, Zeyda M, Todoric J, Huber J, Geyeregger R, Weichhart T et al. Osteopontin expression in human and murine obesity: extensive local up-regulation in adipose tissue but minimal systemic alterations. Endocrinology. 2008, 149, 1350-1357. [CrossRef]
- Ahmad R, Al-Mass A, Al-Ghawas D, Shareif N, Zghoul N, Melhem M et al. Interaction of osteopontin with IL-18 in obese individuals: implications for insulin resistance. PLoS One. 2013, 8, e63944. [CrossRef]
- Gialeli C, Theocharis AD, Karamanos NK. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16-27. [CrossRef]
- Madel M-B, Ibáñez L, Wakkach A, De Vries TJ, Teti A, Apparailly F, Blin-Wakkach C. Immune function and diversity of osteoclasts in normal and pathological conditions. Front Immunol. 2019, 10, 1408. [CrossRef]
- Lin Y, Xu J, Lan H. Tumor-associated macrophages in tumor metastasis: biological roles and clinical therapeutic applications. J Hematol Oncol. 2019, 12, 1-16. [CrossRef]
- Kahles F, Findeisen HM, Bruemmer D. Osteopontin: A novel regulator at the cross roads of inflammation, obesity and diabetes. Mol Metab. 2014, 3, 384-393. [CrossRef]
- Lin C-N, Wang C-J, Chao Y-J, Lai M-D, Shan Y-S. The significance of the co-existence of osteopontin and tumor-associated macrophages in gastric cancer progression. BMC Cancer. 2015, 15, 1-10. [CrossRef]
- Wei J, Marisetty A, Schrand B, Gabrusiewicz K, Hashimoto Y, Ott M et al. Osteopontin mediates glioblastoma-associated macrophage infiltration and is a potential therapeutic target. J Clin Invest. 2019, 129, 137-149. [CrossRef]
- Liang K-H, Yeh C-T. OPN sesame. Hepatobiliary Surg Nutr. 2014, 3, 112.
- Komohara Y, Kurotaki D, Tsukamoto H, Miyasato Y, Yano H, Pan C et al. Involvement of protumor macrophages in breast cancer progression and characterization of macrophage phenotypes. Cancer Sci. 2023, 114, 2220. [CrossRef]
- Ramos RN, Missolo-Koussou Y, Gerber-Ferder Y, Bromley CP, Bugatti M, Núñez NG et al. Tissue- resident FOLR2+ macrophages associate with CD8+ T cell infiltration in human breast cancer. Cell. 2022, 185, 1189-1207. [CrossRef]
- Wei J, Chen Z, Hu M, He Z, Jiang D, Long J, Du H. Characterizing intercellular communication of pan-cancer reveals SPP1+ tumor-associated macrophage expanded in hypoxia and promoting cancer malignancy through single-cell RNA-seq data. Front Cell Dev Biol. 2021, 9, 749210. [CrossRef]
- Liu Z, Gao Z, Li B, Li J, Ou Y, Yu X et al. Lipid-associated macrophages in the tumor-adipose microenvironment facilitate breast cancer progression. Oncoimmunology. 2022, 11, 2085432. [CrossRef]
- Bill R, Wirapati P, Messemaker M, Roh W, Zitti B, Duval F et al. CXCL9: SPP1 macrophage polarity identifies a network of cellular programs that control human cancers. Science. 2023, 381, 515-524. [CrossRef]
- Buechler MB, Fu W, Turley SJ. Fibroblast-macrophage reciprocal interactions in health, fibrosis, and cancer. Immunity. 2021, 54, 903-915. [CrossRef]
- Tokuda K, Morine Y, Miyazaki K, Yamada S, Saito Y, Nishi M et al. The interaction between cancer associated fibroblasts and tumor associated macrophages via the osteopontin pathway in the tumor microenvironment of hepatocellular carcinoma. Oncotarget. 2021, 12, 333. [CrossRef]
- Nakajima T, Uehara T, Iwaya M, Matsuda K, Wada M, Nagaya T et al. Osteopontin expression in the invasive front stroma of colorectal adenocarcinoma is associated with tumor budding and prognosis. Pathol Res Pract. 2022, 240, 154190. [CrossRef]
- Li Y, Guo S, Zhao K, Conrad C, Driescher C, Rothbart V et al. ADAM8 affects glioblastoma progression by regulating osteopontin-mediated angiogenesis. Biol Chem. 2021, 402, 195-206. [CrossRef]
- Radharani N, Yadav AS, Nimma R, Kumar TS, Bulbule A, Chanukuppa V et al. Tumor-associated macrophage derived IL-6 enriches cancer stem cell population and promotes breast tumor progression via Stat-3 pathway. Cancer Cell Int. 2022, 22, 122. [CrossRef]
- Jhunjhunwala S, Hammer C, Delamarre L. Antigen presentation in cancer: insights into tumour immunogenicity and immune evasion. Nat Rev Cancer. 2021, 21, 298-312. [CrossRef]
- Tan Y, Zhao L, Yang Y-G, Liu W. The role of osteopontin in tumor progression through tumor- associated macrophages. Front Oncol. 2022, 12, 953283. [CrossRef]
- Waight JD, Netherby C, Hensen ML, Miller A, Hu Q, Liu S et al. Myeloid-derived suppressor cell development is regulated by a STAT/IRF-8 axis. J Clin Invest. 2013, 123, 4464-4478. [CrossRef]
- Sangaletti S, Tripodo C, Sandri S, Torselli I, Vitali C, Ratti C et al. Osteopontin shapes immunosuppression in the metastatic niche. Cancer Res. 2014, 74, 4706-4719. [CrossRef]
- Allegrezza MJ, Rutkowski MR, Stephen TL, Svoronos N, Perales-Puchalt A, Nguyen JM et al. Trametinib drives T-cell–dependent control of KRAS-mutated tumors by inhibiting pathological myelopoiesis. Cancer Res. 2016, 76, 6253-6265. [CrossRef]
- Klement JD, Poschel DB, Lu C, Merting AD, Yang D, Redd PS, Liu K. Osteopontin blockade immunotherapy increases cytotoxic T lymphocyte lytic activity and suppresses colon tumor progression. Cancers (Basel). 2021, 13, 1006. [CrossRef]
- Zhang Y, Du W, Chen Z, Xiang C. Upregulation of PD-L1 by SPP1 mediates macrophage polarization and facilitates immune escape in lung adenocarcinoma. Exp Cell Res. 2017, 359, 449- 457. [CrossRef]
- Hegde PS, Chen DS. Top 10 challenges in cancer immunotherapy. Immunity. 2020, 52, 17-35. [CrossRef]
- Wykes MN, Lewin SR. Immune checkpoint blockade in infectious diseases. Nat Rev Immunol. 2018, 18, 91-104. [CrossRef]
- Li Y, Liu H, Zhao Y, Yue D, Chen C, Li C et al. Tumor-associated macrophages (TAMs)-derived osteopontin (OPN) upregulates PD-L1 expression and predicts poor prognosis in non-small cell lung cancer (NSCLC). Thorac Cancer. 2021, 12, 2698-2709.
- Zhu Y, Yang J, Xu D, Gao X-M, Zhang Z, Hsu JL et al. Disruption of tumour-associated macrophage trafficking by the osteopontin-induced colony-stimulating factor-1 signalling sensitises hepatocellular carcinoma to anti-PD-L1 blockade. Gut. 2019, 68, 1653-1666. [CrossRef]
- Yan Z, Hu X, Tang B, Deng F. Role of osteopontin in cancer development and treatment. Heliyon.2023,9, e21055. [CrossRef]
- Dai J, Li B, Shi J, Peng L, Zhang D, Qian W et al. A humanized anti-osteopontin antibody inhibits breast cancer growth and metastasis in vivo. Cancer Immunol Immunother. 2010, 59, 355-366. [CrossRef]
- Amilca-Seba K, Tan TZ, Thiery J-P, Louadj L, Thouroude S, Bouygues A et al. Osteopontin (OPN/SPP1), a mediator of tumor progression, is regulated by the mesenchymal transcription factor Slug/SNAI2 in colorectal cancer (CRC). Cells. 2022, 11, 1808. [CrossRef]
- Shojaei F, Scott N, Kang X, Lappin PB, Fitzgerald AA, Karlicek S et al. Osteopontin induces growth of metastatic tumors in a preclinical model of non-small lung cancer. J Exp Clin Cancer Res. 2012, 31, 1-12. [CrossRef]
- Kumar S, S Patil H, Sharma P, Kumar D, Dasari S, G Puranik V et al. Andrographolide inhibits osteopontin expression and breast tumor growth through down regulation of PI3 kinase/Akt signaling pathway. Curr Mol Med. 2012, 12, 952-966.
- Im E, Yeo C, Lee E-O. Luteolin induces caspase-dependent apoptosis via inhibiting the AKT/osteopontin pathway in human hepatocellular carcinoma SK-Hep-1 cells. Life Sci. 2018, 209, 259-266. [CrossRef]
- Desai B, Rogers MJ, Chellaiah MA. Mechanisms of osteopontin and CD44 as metastatic principles in prostate cancer cells. Mol Cancer. 2007, 6, 1-16. [CrossRef]
- Lu C, Liu Z, Klement JD, Yang D, Merting AD, Poschel D et al. WDR5-H3K4me3 epigenetic axis regulates OPN expression to compensate PD-L1 function to promote pancreatic cancer immune escape. J Immunother Cancer. 2021, 9, e002624. [CrossRef]
- Gao X, Sheng Y, Yang J, Wang C, Zhang R, Zhu Y et al. Osteopontin alters DNA methylation through up-regulating DNMT1 and sensitizes CD133+/CD44+ cancer stem cells to 5 azacytidine in hepatocellular carcinoma. J Exp Clin Cancer Res. 2018, 37, 1-14.
- Deng G, Zeng F, Su J, Zhao S, Hu R, Zhu W et al. BET inhibitor suppresses melanoma progression via the noncanonical NF-κB/SPP1 pathway. Theranostics. 2020, 10, 11428.
- Han B, Huang J, Han Y, Hao J, Wu X, Song H et al. The microRNA miR-181c enhances chemosensitivity and reduces chemoresistance in breast cancer cells via down-regulating osteopontin. Int J Biol Macromol. 2019, 125, 544-556. [CrossRef]
- Ben-David-Naim M, Dagan A, Grad E, Aizik G, Nordling-David MM, Morss Clyne A et al. Targeted siRNA nanoparticles for mammary carcinoma therapy. Cancers (Basel). 2019, 11, 442. [CrossRef]
- Wang S-Y, Chen C-L, Hu Y-C, Chi Y, Huang Y-H, Su C-W et al. High expression of microRNA- 196a is associated with progression of hepatocellular carcinoma in younger patients. Cancers (Basel). 2019, 11, 1549. [CrossRef]
- Lakhin A, Tarantul V, Gening L. Aptamers: problems, solutions and prospects. Acta Naturae. 2013, 5, 34-43.
- Hunter C, Bond J, Kuo PC, Selim MA, Levinson H. The role of osteopontin and osteopontin aptamer (OPN-R3) in fibroblast activity. J Surg Res. 2012, 176, 348-358. [CrossRef]
- Talbot LJ, Mi Z, Bhattacharya SD, Kim V, Guo H, Kuo PC. Pharmacokinetic characterization of an RNA aptamer against osteopontin and demonstration of in vivo efficacy in reversing growth of human breast cancer cells. Surgery. 2011, 150, 224-230. [CrossRef]
- Liu L, Zhang R, Deng J, Dai X, Zhu X, Fu Q et al. Construction of TME and identification of crosstalk between malignant cells and macrophages by SPP1 in hepatocellular carcinoma. Cancer Immunol Immunother. 2022, 71, 121-136. [CrossRef]
- Song H, Lou C, Ma J, Gong Q, Tian Z, You Y et al. Single-cell transcriptome analysis reveals changes of tumor immune microenvironment in oral squamous cell carcinoma after chemotherapy. Front Cell Dev Biol. 2022, 10, 914120. [CrossRef]
- Zhang L, Li Z, Skrzypczynska KM, Fang Q, Zhang W, O’Brien SA et al. Single-cell analyses inform mechanisms of myeloid-targeted therapies in colon cancer. Cell. 2020, 181, 442-459. [CrossRef]
Figure 1.
Structural domains of full-length OPN and its receptors. N-terminus of OPN consists of poly-D region, calcium binding domain, and ELVTDFPTDLPAT sequence motif which interacts with α4β1 integrin. The central region consists of RGD domain, binds with other integrins such as αvβ3, αvβ1, αvβ5, αvβ6, αvβ8, α5β1 and α8β1; the SVVYGLR sequence binds to α9β1, α4β1 and α4β7 integrins. The C-terminal region includes another calcium- binding domain, MMP-cleavage site, and heparin-binding domain which facilitates the interaction of OPN with CD44. Adopted from Mirzaei et al, Asian Pacific J of Cancer Prevention, 19 (3), 2018 and Bandopadhyay et al, EOTT, 18 (8), 2014 with modifications.
Figure 1.
Structural domains of full-length OPN and its receptors. N-terminus of OPN consists of poly-D region, calcium binding domain, and ELVTDFPTDLPAT sequence motif which interacts with α4β1 integrin. The central region consists of RGD domain, binds with other integrins such as αvβ3, αvβ1, αvβ5, αvβ6, αvβ8, α5β1 and α8β1; the SVVYGLR sequence binds to α9β1, α4β1 and α4β7 integrins. The C-terminal region includes another calcium- binding domain, MMP-cleavage site, and heparin-binding domain which facilitates the interaction of OPN with CD44. Adopted from Mirzaei et al, Asian Pacific J of Cancer Prevention, 19 (3), 2018 and Bandopadhyay et al, EOTT, 18 (8), 2014 with modifications.
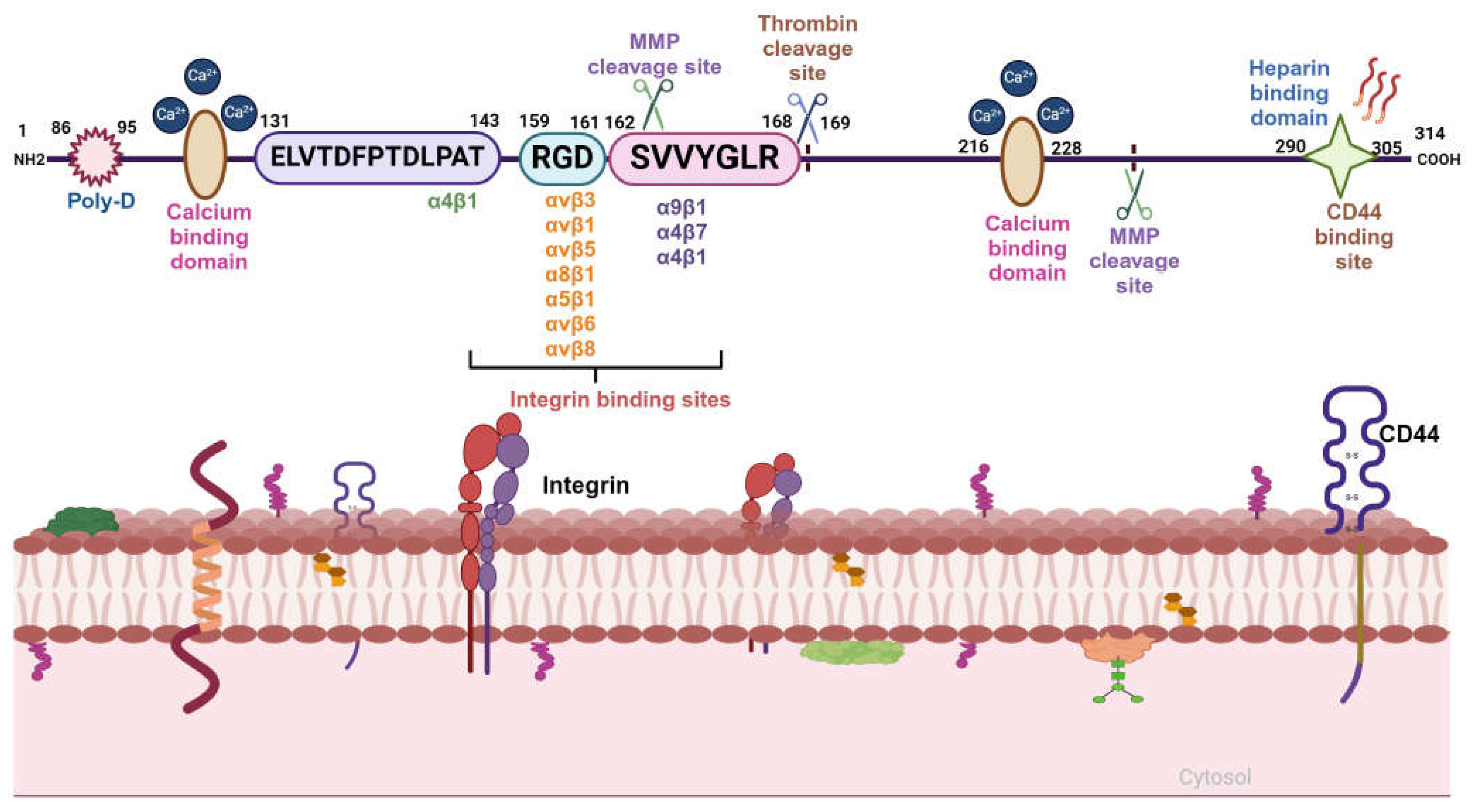
Figure 2.
Schematic illustration of the OPN gene and its splice variants. Alternative splicing of OPN transcript results in five splice variants, which are denoted as OPN- a, OPN-b, OPN-c, OPN-4, and OPN-5. OPN-a is a full-length variant that consists of 7 exons; OPN-b lacks exon 5; while in OPN-c exon 4 is absent. In OPN-4, both exon 4 and exon 5 are missing; whereas OPN-5 is the longest variant which consists of an additional exon, generated from a portion of intron 3. Additionally, four new sub-variants of OPN-5 (OPN-5b, OPN-5c, OPN- 5d, OPN-5e) have been identified. OPN-5a is the same as OPN-5; OPN-5b has the extra shortened exon while in OPN-5c has additional 9 base pairs in the 3’ region of extra exon. In OPN-5d, there is deletion of exon 5 with the addition of 9 base pairs in the 3’ region of the extra exon. OPN-5e lacks exon 5. Adopted from Bastos et al, Intl J of Mol Sci, 24, 2023 with modification.
Figure 2.
Schematic illustration of the OPN gene and its splice variants. Alternative splicing of OPN transcript results in five splice variants, which are denoted as OPN- a, OPN-b, OPN-c, OPN-4, and OPN-5. OPN-a is a full-length variant that consists of 7 exons; OPN-b lacks exon 5; while in OPN-c exon 4 is absent. In OPN-4, both exon 4 and exon 5 are missing; whereas OPN-5 is the longest variant which consists of an additional exon, generated from a portion of intron 3. Additionally, four new sub-variants of OPN-5 (OPN-5b, OPN-5c, OPN- 5d, OPN-5e) have been identified. OPN-5a is the same as OPN-5; OPN-5b has the extra shortened exon while in OPN-5c has additional 9 base pairs in the 3’ region of extra exon. In OPN-5d, there is deletion of exon 5 with the addition of 9 base pairs in the 3’ region of the extra exon. OPN-5e lacks exon 5. Adopted from Bastos et al, Intl J of Mol Sci, 24, 2023 with modification.
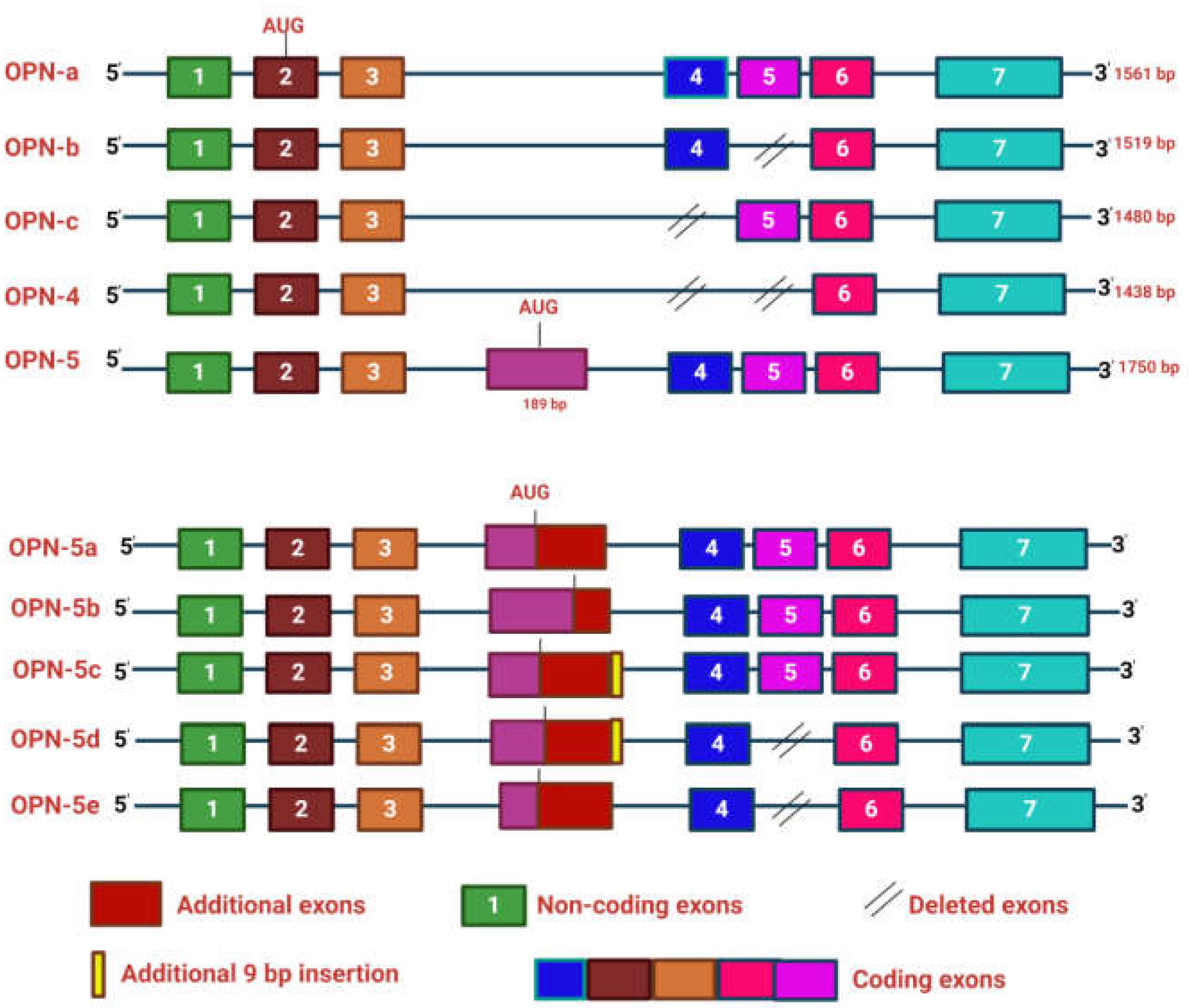
Figure 3.
Role of OPN in the regulation of various signalling pathways. OPN through its interaction with αvβ3, α4β1, α3β2, and α9β1 integrins, and CD44 receptor transduces multiple signalling pathways and their cross-talks such as FAK/MEK/ERK, PKCγ. /PKC/PI3K/Akt/mTOR, NIK/IBα/NFκB, JAK/STAT3, PI3K/Akt/β-catenin, NFκB/HIF1α/BMI1, c-Src/EGFR/MEK/ERK and MAPK pathways. These signalling cascades induce the activation of various tumor-promoting genes such as VEGF, MMPs, and COX-2, thereby inducing the tumor growth at the primary sites, angiogenesis, metastases at the distance sites, ECM remodeling, immune suppression, stemness, immune evasion, chemoresistance, migration, and survival. Adopted from Han et al, Oncology Letters, 17, 2019 and Ahmed et al EOTT, 15 (9), 2011 with modifications.
Figure 3.
Role of OPN in the regulation of various signalling pathways. OPN through its interaction with αvβ3, α4β1, α3β2, and α9β1 integrins, and CD44 receptor transduces multiple signalling pathways and their cross-talks such as FAK/MEK/ERK, PKCγ. /PKC/PI3K/Akt/mTOR, NIK/IBα/NFκB, JAK/STAT3, PI3K/Akt/β-catenin, NFκB/HIF1α/BMI1, c-Src/EGFR/MEK/ERK and MAPK pathways. These signalling cascades induce the activation of various tumor-promoting genes such as VEGF, MMPs, and COX-2, thereby inducing the tumor growth at the primary sites, angiogenesis, metastases at the distance sites, ECM remodeling, immune suppression, stemness, immune evasion, chemoresistance, migration, and survival. Adopted from Han et al, Oncology Letters, 17, 2019 and Ahmed et al EOTT, 15 (9), 2011 with modifications.
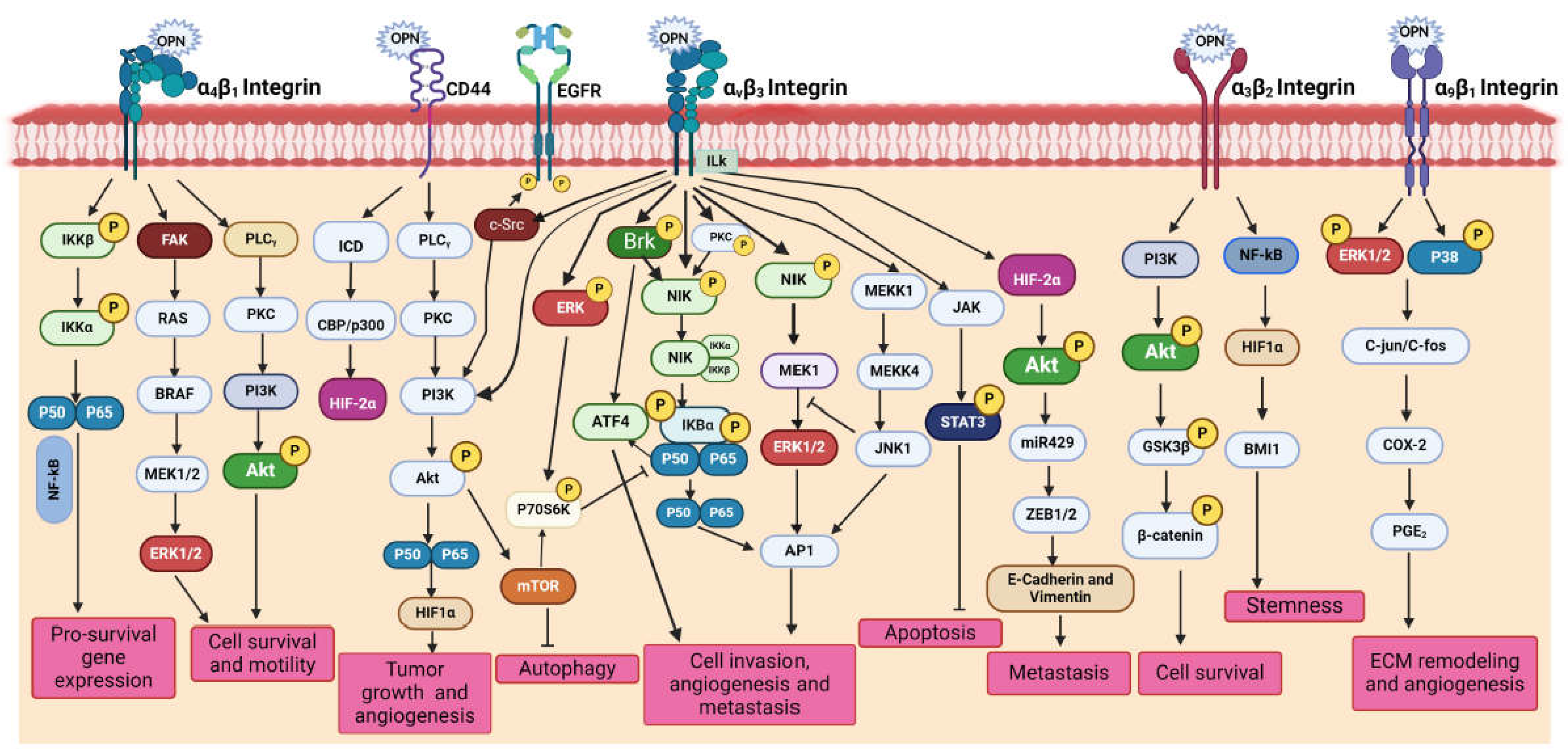
Figure 4.
Diagrammatic representation of multifaceted function of OPN in various tumors. OPN regulates the epithelial to mesenchymal transition (EMT) resulting in the loss of tight junctions, thereby enhancing metastasis, with the high expression of vimentin, slug, snail and fibronectin. Under hypoxic conditions, OPN induces activation of PI3K leading to phosphorylation of Akt, thereby upregulating VEGF dependent angiogenesis. OPN is responsible for metabolic function by activating HIF1α, under hypoxic conditions which further aids in the glycolytic process with high expression of VEGF, PDK1, LDHA, iNOS, GLUT1 and GLUT3. OPN further regulates CSC enrichment by activating a cascade of signalling pathways involving PI3K/Akt/mTOR, Hedgehog, MAPK, Wnt/β-catenin, JAK/STAT and Notch signalling. OPN- activated signalling pathways result in drug resistance. The interaction between OPN, αvβ3, and CD44 results in the activation of PI3K/Akt, FAK/MEK/ERK, EGFR, and Wnt/NFκB signalling cascades thereby aiding therapeutic resistance.
Figure 4.
Diagrammatic representation of multifaceted function of OPN in various tumors. OPN regulates the epithelial to mesenchymal transition (EMT) resulting in the loss of tight junctions, thereby enhancing metastasis, with the high expression of vimentin, slug, snail and fibronectin. Under hypoxic conditions, OPN induces activation of PI3K leading to phosphorylation of Akt, thereby upregulating VEGF dependent angiogenesis. OPN is responsible for metabolic function by activating HIF1α, under hypoxic conditions which further aids in the glycolytic process with high expression of VEGF, PDK1, LDHA, iNOS, GLUT1 and GLUT3. OPN further regulates CSC enrichment by activating a cascade of signalling pathways involving PI3K/Akt/mTOR, Hedgehog, MAPK, Wnt/β-catenin, JAK/STAT and Notch signalling. OPN- activated signalling pathways result in drug resistance. The interaction between OPN, αvβ3, and CD44 results in the activation of PI3K/Akt, FAK/MEK/ERK, EGFR, and Wnt/NFκB signalling cascades thereby aiding therapeutic resistance.
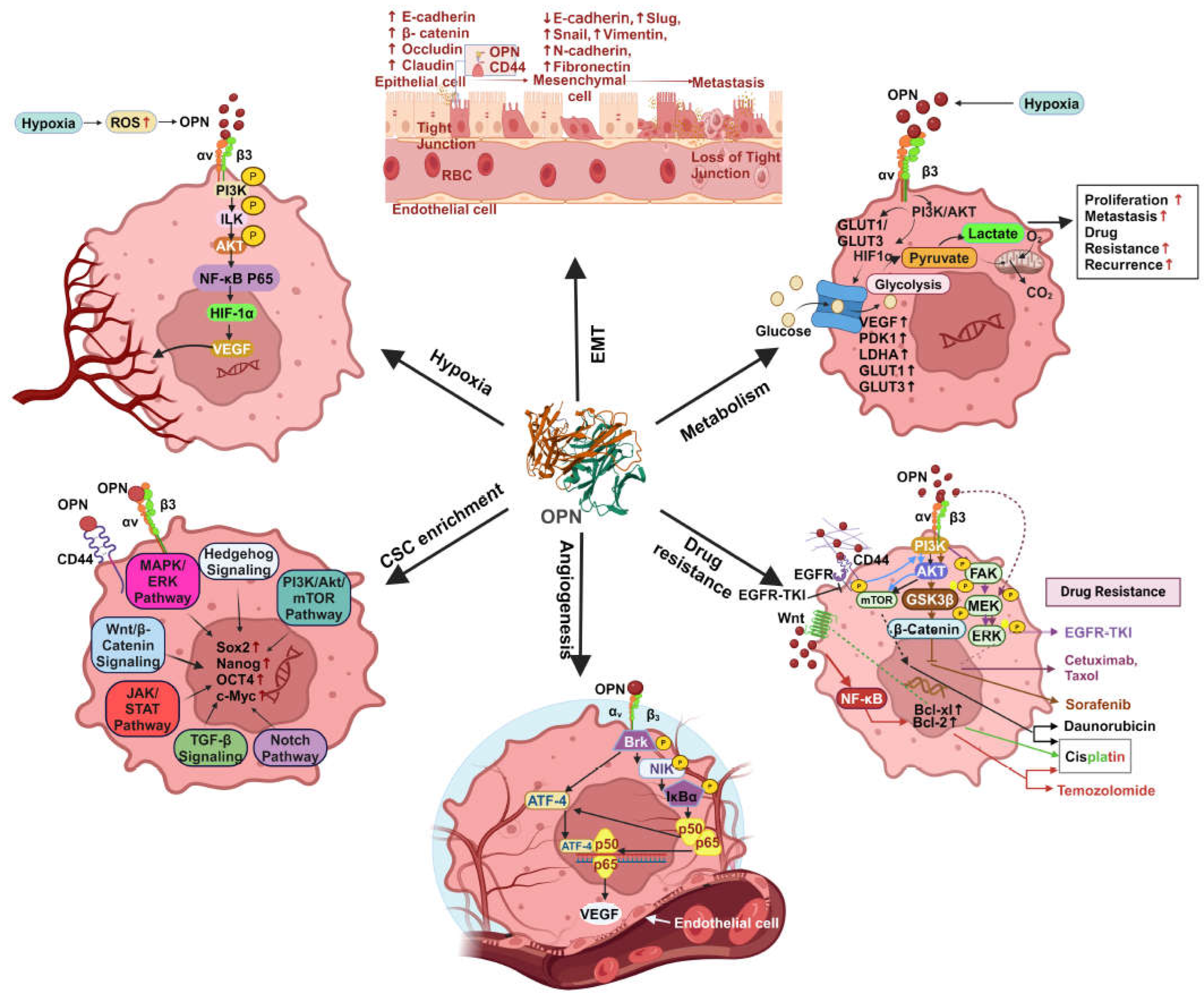
Figure 5.
Myofibroblast differentiation of resident fibroblasts and MSCs by tumor derived OPN. Tumor-derived OPN is involved in the transition of resident fibroblasts and MSCs into myofibroblast or CAFs. CAF-derived factors induce ECM deposition, EMT, angiogenesis, CSC enrichment, metabolic reprogramming, and tumor survival, resulting in the enhancement of tumor progression.
Figure 5.
Myofibroblast differentiation of resident fibroblasts and MSCs by tumor derived OPN. Tumor-derived OPN is involved in the transition of resident fibroblasts and MSCs into myofibroblast or CAFs. CAF-derived factors induce ECM deposition, EMT, angiogenesis, CSC enrichment, metabolic reprogramming, and tumor survival, resulting in the enhancement of tumor progression.
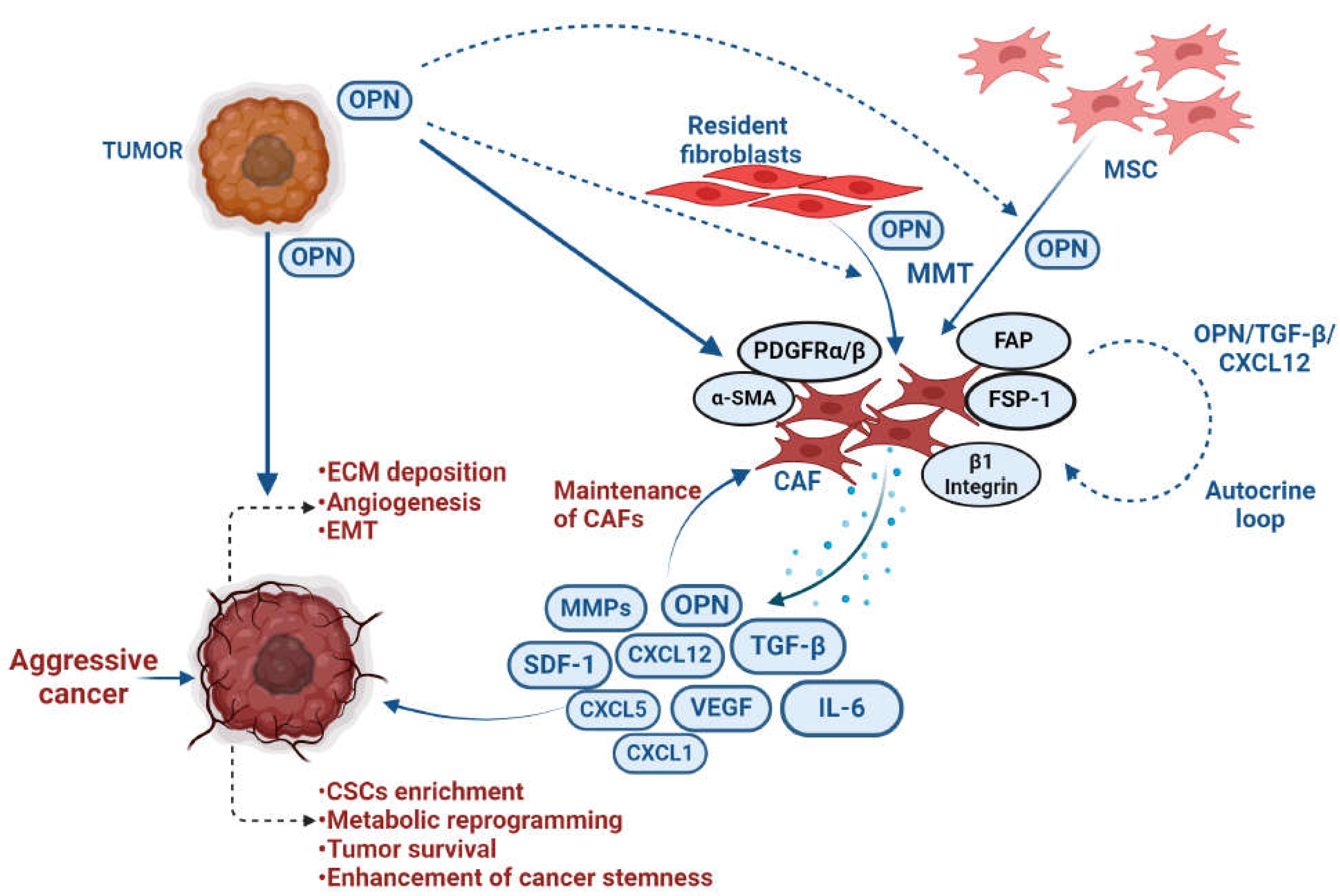
Figure 6.
Model depicting the role of OPN in tumor immune microenvironment (TIME): A. Role of OPN in shaping immunosuppressive TME: Tumor-derived OPN activates stromal cells by trans-differentiation of fibroblasts to myofibroblasts resulting in expansion of tumor. OPN- regulated PD-1/PDL1 interaction inhibits T-cell activation. Further, tumor cells induce lower expression of IRF8, resulting in overexpression of OPN thus leading to T-cell suppression. OPN via the NF-κB pathway, upregulates PD-L1 expression aiding in immune therapy escape. OPN induces the polarization of macrophages and the recruitment of monocytes. OPN activates TAM leading to angiogenesis, metastasis and enrichment of cancer stem cells via upregulation of various tumor-promoting factors like MMP-9, N-Cadherin, Vimentin, ICAM-1, COX-2, PGE-2, VEGF, Sox-2, Oct-3/4, Nanog and ALDH. The polarity ratio of CXCL9 and OPN (SPP1) determines the anti- and pro-tumorigenic properties of TAMs. B. Involvement of OPN in tumor immunity continuum: OPN is primarily associated in inflamed and immune excluded tumors whereas its role in immune dessert tumor is remains elusive. C. Schematic representations to identify OPN regulated immune cell heterogeneity in cancer: OPN regulated immune modulatory genes may be identified in TIME by CRISPR technology in breast cancer using scRNA seq. based platform.
Figure 6.
Model depicting the role of OPN in tumor immune microenvironment (TIME): A. Role of OPN in shaping immunosuppressive TME: Tumor-derived OPN activates stromal cells by trans-differentiation of fibroblasts to myofibroblasts resulting in expansion of tumor. OPN- regulated PD-1/PDL1 interaction inhibits T-cell activation. Further, tumor cells induce lower expression of IRF8, resulting in overexpression of OPN thus leading to T-cell suppression. OPN via the NF-κB pathway, upregulates PD-L1 expression aiding in immune therapy escape. OPN induces the polarization of macrophages and the recruitment of monocytes. OPN activates TAM leading to angiogenesis, metastasis and enrichment of cancer stem cells via upregulation of various tumor-promoting factors like MMP-9, N-Cadherin, Vimentin, ICAM-1, COX-2, PGE-2, VEGF, Sox-2, Oct-3/4, Nanog and ALDH. The polarity ratio of CXCL9 and OPN (SPP1) determines the anti- and pro-tumorigenic properties of TAMs. B. Involvement of OPN in tumor immunity continuum: OPN is primarily associated in inflamed and immune excluded tumors whereas its role in immune dessert tumor is remains elusive. C. Schematic representations to identify OPN regulated immune cell heterogeneity in cancer: OPN regulated immune modulatory genes may be identified in TIME by CRISPR technology in breast cancer using scRNA seq. based platform.
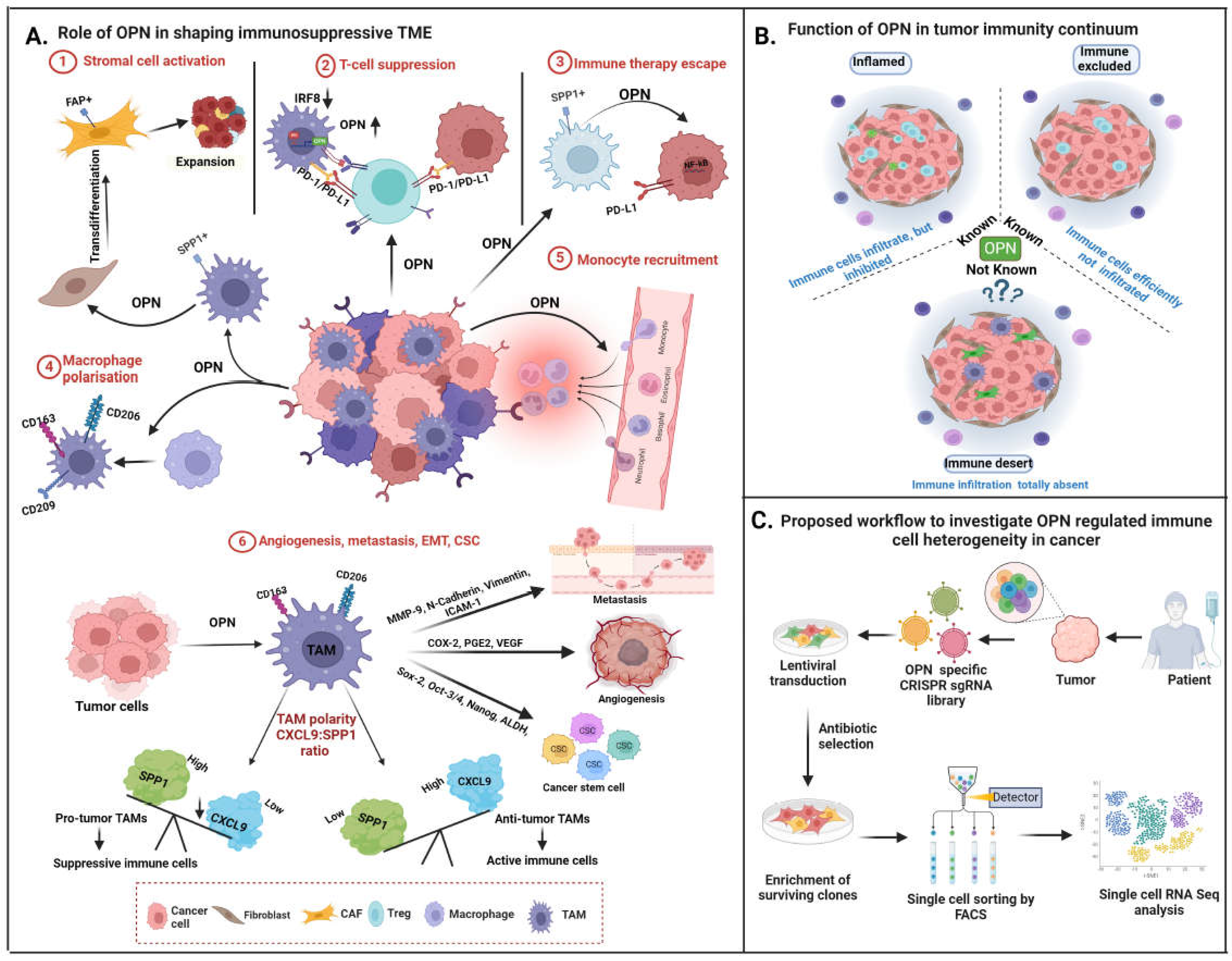
Figure 7.
OPN targeted novel therapeutic strategies. Small molecule inhibitors (andrographolide, curcumin, etc), OPN-specific siRNA, miRNA, shRNA, OPN aptamers, synthetic RGD peptides, OPN neutralizing antibodies, CD44 or integrin blocking antibodies, have been used recently as therapeutic approach to target OPN-integrin/CD44 axis which leads to downregulation of various oncogenic molecules and suppression of tumor progression by disrupting OPN regulated various signalling pathways in cancers.
Figure 7.
OPN targeted novel therapeutic strategies. Small molecule inhibitors (andrographolide, curcumin, etc), OPN-specific siRNA, miRNA, shRNA, OPN aptamers, synthetic RGD peptides, OPN neutralizing antibodies, CD44 or integrin blocking antibodies, have been used recently as therapeutic approach to target OPN-integrin/CD44 axis which leads to downregulation of various oncogenic molecules and suppression of tumor progression by disrupting OPN regulated various signalling pathways in cancers.
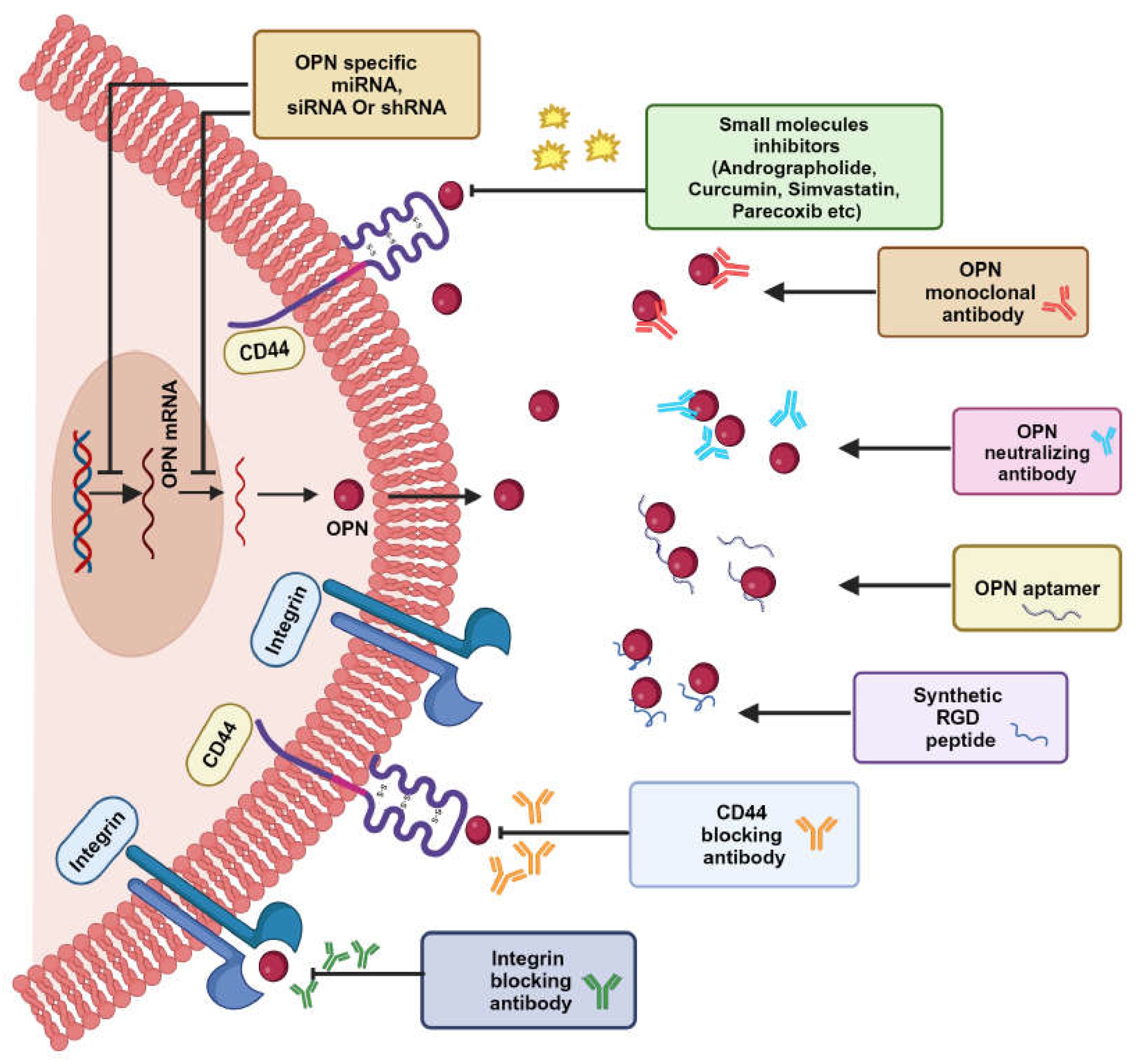

Table 1.
Expression of OPN in various cancers.
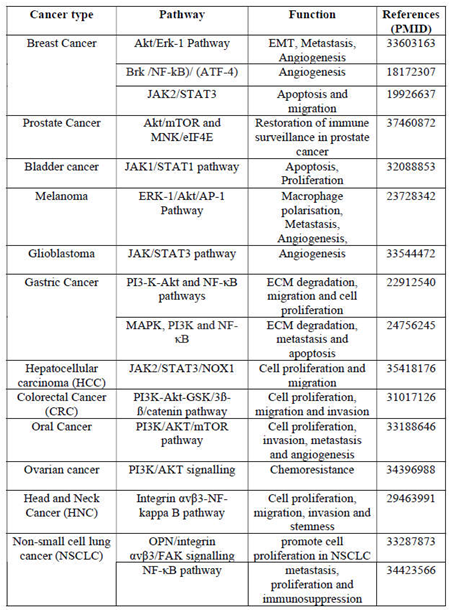
Table 2.
OPN as early prognostic & diagnostic biomarker in various cancers.
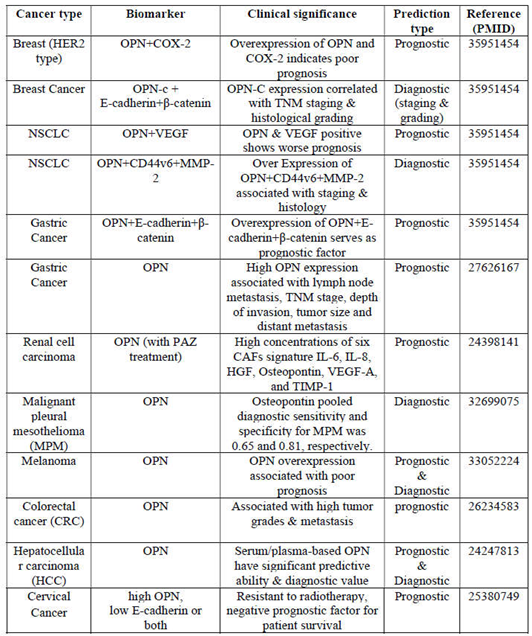
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



