
Submitted:
10 May 2024
Posted:
13 May 2024
You are already at the latest version
Abstract
The in vitro cultivation of M. leprae has not been possible since it was described as causing leprosy, and the limitation of animal models for clinical aspects makes studies on leprosy and bacteria-human host interaction a challenge. Our aim was to standardize the ex vivo skin model (hOSEC) to maintenance and study of M. leprae as an alternative animal model. Bacillary suspensions were inoculated into human skin explants and sustained in DMEM medium until 60 days. Explants were evaluated by RT-PCR-16SrRNA, and cytokines gene expression. Viability and infectivity of bacilli recovered from explant (D28 and D60) were evaluated using the Shepard’s model. All explants were RT-PCR-16srRNA positive. Viability and infectivity of recovered bacilli from explants, analyzed after 5 months of inoculation in mice showed an average positivity of 31%, being the highest positivity in the D28 groups (80%). Furthermore, our work showed different pattern in cytokines gene expression (TGF-β, IL-10, IL-8 and TNF-α) in the presence of alive or dead bacilli. Although changes can be made for improve future experiments, our results have demonstrated that it is possible to use the hOSEC to maintain M. leprae for 60 days, interacting with the host system, an important step in the development of experimental models for studies on the biology of the bacillus, its interactions, and drug susceptibility.
Keywords:
Mycobacterium leprae
; leprosy
; hOSEC
; ex vivo skin culture
; animal use alternatives
; tissue culture techniques
1. Introduction
Mycobacterium leprae (M. leprae), or Hansen bacillus, described by Gerhard Henrik Armauer Hansen in 1873 [1], was the first bacterial pathogen identified and considered to be the cause of a human infectious disease; however, the fact that M. leprae cannot be grown in vitro still poses a challenge to researchers. M. leprae subverts the immune defense of the host, infects macrophages and Schwann cells of the peripheral nerve system in the skin [2,3] and causes leprosy, a chronic dermatoneurological disease, which continues to carry a strong social stigma, and is widespread throughout the world, mainly in Brazil and India [4].
Depending on the immunological response (humoral and cellular) of the host, leprosy presents a complex clinical spectrum [5]. After having contact with M. leprae, certain individuals do not become ill. Other individuals present with the disease, but do not have severe symptoms; the treatment of these individuals is efficient and uncomplicated. Other individuals, however, become ill, present important symptoms and complications during and/or after treatment (mainly in late diagnosis), and a few may become resistant to drugs [6].
One of the causes of delayed diagnosis of leprosy and the difficulty in studying M. leprae and its interactions with the host, is related to the inability of these mycobacteria to be cultured in vitro. Numerous attempts to cultivate in vitro using several different culture media have been unsuccessful in the maintenance the mycobacterium alive, in growth and induction of the pathology [7,8,9].
Currently, the only method for maintaining viable M. leprae in the laboratory is by in vivo inoculation, mainly passages into athymic nude mice, a laborious and time-consuming technique [10].
Although this in vivo experimental model has allowed important advances in M. leprae studies, it has limitations because it does not thoroughly reproduce the disease. The impossibility of in vitro M. leprae culture has hampered the study of basic aspects involved in disease transmission, genetic and immunological factors involved in resistance/susceptibility to the disease, and test of new therapeutic targets.
Although we have a model for cultivating of bacilli in mouse footpads, it is important to highlight the current advances in policies and laws related to restricting the use of animals in scientific experiments, through alternative methods [11,12].
Thus, human organotypic skin explant culture (hOSEC) is an ex vivo model of human skin, an alternative method, applied to study the skin and some interventions, so here, we used hOSEC to standardize its use in the maintenance of M. leprae, since the skin is the natural habitat of the bacillus, and we showed that the use of this model is feasible to maintain the viability and pathogenicity of M. leprae to study the interaction between the bacillus and the human skin.
2. Materials and Methods
2.1. hOSEC (human Organotypic Skin Explant Culture)
Fragments of healthy human skin were obtained from skin remaining from tummy tuck surgery after informed consent of patients in accordance with the ethical guidelines of the Research Ethics Committee of the Clinics Hospital at Ribeirão Preto Medical School, University of São Paulo (protocol number 1.744.888/2016).
Soon after the surgery, the skin tissues were manipulated inside a laminar flow hood and placed in phosphate-buffered saline (PBS, pH 7.2) plus 1.5% antibiotic solution (100 U/mL penicillin and 100 mg/mL streptomycin; Gibco®, Grand Island, NY, USA) overnight at 4°C for decontamination. Later, the subcutaneous tissue was removed with scissors, and the full-thickness skin (epidermis and dermis) was cut with an 8-mm diameter biopsy punch, the explants were placed with the dermal side facing down on pieces of filter paper (80 g/m2, 26 l/s m2 air permeability, 25 μm porosity) supported by metal grids in 6-well culture plates, and each grid supported three explants, similar as described by Frade and collaborators [13] (Figure 1). Approximately five milliliters of Dulbecco’s modified Eagle medium (DMEM; Gibco®) supplemented with 10% fetal bovine serum (FBS; Gibco®) and 1% antibiotic solution (100 U/mL penicillin and 100 mg/mL streptomycin) was added to each well until it reached the dermis. Every third day, two mL of exhausted medium was replaced with fresh medium. Experiments were performed on skin from four different individuals.
2.2. Inoculation of M. leprae
Before the explants were placed on the plates, each one was inoculated with 25 μL of a suspension containing 1,5x104 M. leprae bacilli. The bacilli in the suspension were obtained from continuous passage of the M. leprae Thai-53 strain into the footpads of athymic mice (NU-Foxn1nu) from the Lauro de Souza Lima Institute in Bauru, São Paulo-Brazil. The suspension was obtained following the method described by Trombone and collaborators [10]. Briefly, a mouse was euthanized, the footpads were remove and, cleaned with 70° ethanol, the bone tissue was discarded, and the remaining tissue was cut into small pieces and transferred to a tube containing Hanks' solution. The tissue was homogenized, trypsin was added, and the tube was incubated at 37°C for 1 hour. After trypsin inactivation, the suspension was centrifuged, and the supernatant was discarded. The pellet was re-suspended in 1 mL of saline solution (0.9% NaCl) and homogenized with a syringe and 12x0.7 needle. The bacilli were counted on a glass slide after Ziehl-Neelsen staining (ZN) and the percentage of viability determined by the Live/Dead BacLight Bacterial Viability kit (ThermoFisher Scientific, cat: L7007). The suspensions used had at least 86% viability, the dilution for inoculation was based only on the number of live bacilli determined by the Live/Dead BacLight kit (integral membrane). Equal volumes with equal amounts of bacilli were separated, one directed towards the inoculation of live bacilli and the other towards dead bacilli, which were obtained after autoclaving (autoclave at 121 °C for 40 min). An aliquot of the suspension was tested for contaminant microorganism growth on brain heart infusion (BHI) medium and Sabouraud medium, both solid and liquid, under aerobic and anaerobic conditions.
For inoculation into explants, the viable bacillary suspension (VML) was diluted in saline solution, and intradermal injection was performed using a microsyringe (Hamilton® Company- Reno, Nevada, USA). Others explants were inoculated with the same number of bacilli previously inactivated by autoclaving (DML), and saline solution (25 µL) was inoculated into control explants. After inoculation, the explants were placed on plates as previously described and incubated at 37°C in 5% CO2 for 4, 7, 14, 28, 60 days. Three explants from each group were collected for analysis using three different protocols (histomorphology, viability, enumeration/inoculation) starting at the initial time point (D0) and then on the other days (D4, D7, D14, D28 and D60).
2.3. Histomorphology analysis of hOSEC
The explants were embedded in paraffin for histomorphological analysis, the slides were prepared with 4-μm sections and hematoxylin and eosin staining (HE). The analysis was performed with a Leica® DM-4000B optical microscope with a Leica® DFC 280 camera connected to a computer using Leica Application Suite (LAS®) for capturing images (Leica Microsystems, Mannheim, Germany).
2.4. Molecular analysis of hOSEC
Explants were placed in TRIzol® (Life Technologies) and maintained at -80°C for molecular biology analysis. To isolate the RNA, the explants were thawed and thoroughly macerated with 2 mL of TRIzol® using a tissue homogenizer (Omni TH tissue homogenizer - Kennesaw, GA, USA), the tubes were maintained on ice during the process.
The RNA isolation technique followed the recommendations of the TRIzol® reagent manufacturer; briefly, to every 1 mL of macerated sample was added 200 μL of cold chloroform (J. T Baker. cat.: 9180-02), shaken and centrifuged (12,000 g at 4°C for 15 min). The upper aqueous phase was transferred to a new microtube containing 500 μL of isopropanol, incubated at -80°C for approximately 24 hours, and centrifuged at 12,000 g at 4°C for 10 min. The supernatant was discarded, and the pellet was washed with 1000 μL of 75% ethanol followed by centrifugation at 7,500 g at 4°C for 5 min. After drying, the pellet was eluted with 30 μL of diethyl pyrocarbonate water (DEPC) (Sigma-Aldrich, cat.: D5758), the amount of RNA was determined using a NanoVue® Plus spectrophotometer (GE Healthcare Life Sciences, Buckinghamshire, UK).
DNAse (RQ1 RNase-Free DNase, Promega, USA, cat.: M6101) was added to the RNA samples for DNA digestion according to the manufacturer’s protocol (Promega, cat.: M6101), and 500 ng was reverse-transcribed using random primers and the GoScript® Reverse Transcriptase System (Promega, cat.: A5001) according to the manufacturer’s protocol and incubated in a Bio-Rad T100 thermal cycler (Bio-Rad Laboratories, USA. cat.: 1861096). The cDNA was diluted 1:2, and 5 μL was used for RT-qPCR.
PCR was performed on a CFX96® Real-Time PCR Detection System (Bio-Rad, CFX96® Touch System, EUA. cat.: 184-5096) using 12,5 μL SYBR Green Master Mix (2x) (GoTaq qPCR Master Mix, Promega, cat.: A6002), 0.5 μL of each primer (10 μM) and 6.5 μL of nuclease-free water. The primer sequences and cycling protocol are shown in Table 1.
The cycle threshold (CT) value, positivity to sample, was considered only if the temperature melting (TM) was equal the reaction control [18,19,20], the expression rate of cytokines (TGF-β, TNF-α, IFN-γ, IL-1β, IL-10 and IL-8) was estimated for each explant with viable M. leprae and dead M. leprae comparing with saline group, using the 2-ΔΔCT formula (ΔΔCT = ΔCT test - ΔCT saline; ΔCT= target gene CT value –reference gene CT value) [19], and 18S rRNA was used as a reference gene, and the group saline as a normalize.
2.5. Viability using in vivo model
Following 28 and 60 days in hOSEC, the explants were processed to harvest bacilli for inoculation into the footpads of athymic mice. The fragments were cut into smaller pieces using scissors, transferred to tubes containing 1000 μL of saline solution, and homogenized by 3 pulses with a tissue homogenizer (Tissue Homogenizer Omni TH®) at a speed of 4 (14,450 rpm) for 15 sec. The tubes were always maintained on ice. Then, the homogenates were filtered through a cell strainer to eliminate the remaining debris, centrifuged at 12,000 g at 4°C for 10 min and suspended in 200 μL of saline.
These bacilli suspensions were taken, cooled, by road transport to the ILSL (Bauru-SP, Brazil) where following Shepard model [21], two hind footpads of mice were inoculated with 30 μL of the obtained bacillary suspension using a 30G needle and insulin syringe. For each analysis time (D28 and D60), there were 3 fragments of each skin, each fragment generated 200μL suspension that was divided into 3 animals (6 footpads/fragment). There were minimal of three animals per fragment suspension (D28 and D60) and three fragment of each time, of each skin. After five months, the animals were euthanized, and their foot pads were removed for molecular and histological analysis, Fite Faraco (FF) and Ziehl-Neelsen, being that the ZN was performed in one macerated foot pad, FF was performed in half of the second foot pad and the RT-PCR 16S rRNA was performed in the another half of the second foot pad.
The procedures were in accordance with the Ethical Principles in Animal Research and was approved by Local Animal Ethical Committee of the Ribeirão Preto Medical School, University of São Paulo (protocol number 026/2015-1).
2.6. Statistical analysis
Statistical analysis was performed using the GraphPad Prism 5 program, using One-Way ANOVA followed by Tukey's Test, for comparisons between all groups and T-Test for comparisons between two groups, with a 95% confidence interval, and values of significant p were: * p <0.05, ** p <0.01, *** p <0.001.
3. Results
3.1. Characterization of donor skin and histomorphology
The skin used for hOSEC was obtained from skin remaining from tummy tuck surgery in four white subjects (3 females and 1 male) between the ages of 25 and 56 years (median 46,5 years).
Upon histological examination, we did not observe any difference between skin with or without bacilli, in general, the hOSEC skin showed scattered cells and structures of glands, vessels, muscles, nerve fillets, and hair follicles, which were observed throughout the culture time. The skin maintained its natural architecture until the 7th day and was very similar to that observed at the initial time point, maintaining all layers of the epidermis, spinous, granulosa and cornea without visible changes to the dermo-epidermal junction. At subsequent time points, there was a progressive decrease in the number of keratinocyte layers, and the epidermis was more rectified, the corneal layer thickness was increased; however, the dermo-epidermal junction and the basal layer maintained regular. The papillary dermis was easily noticed until the 14th day, from the 28th day on, it became denser, with less delimitation. In the reticular dermis, in general, there was an increase in the thickness of the collagen fibers with a decrease in the spaces between them on the 28th day and mainly on the 60th day (Figure 2).
3.2. Viability of bacilli in hOSEC
To evaluate the viability of bacilli in the explants, 16S rRNA was used as a target RNA for RT-PCR, and all culture times until 60th day demonstrated amplification of this target, only four fragments showed undetected expression (Table 2).
3.3. Maintenance of viable and infective bacilli in hOSEC
After defining the bacillary viability in the explants until D60, we evaluated whether their infectiveness was also maintained by inoculating bacilli recovered from explants (D28 and D60) into the footpads of athymic mice. The in vivo experiment included 83 animals, 21,7% of the mice died before the time of analysis (<5 months) of causes unrelated to the experiment. Euthanasia was performed in the remaining mice after five months of inoculation, and the molecular analysis was performed in 43 animals and histological analysis in 65.
Among the mice that received suspensions from D28 explants (N=37), 10 (27,0%) showed positive microscopy for M. leprae by ZN, with counts from 1/100 to 470/100 bacilli/field, and 9 (24,3%) positivity by FF in the histology. Among the mice from D60 explants (N=28), 5 (17,9%) mice showed positive by ZN and 1 (5,6%) by FF.
By RT-PCR 16S rRNA analysis, only 15 animals inoculated with suspension from D28 explants and 28 from D60 were analyzed, among this, 6 (40,0%) from D28 and 3 (10,7%) from D60 showed positivity.
Interestingly, the higher positivity of the mice was found between the animals inoculated with suspension from fragments originated the male skin (Skin 4). All animals showed positivity after 5 months from inoculation in at least one of the analyses. In the viability analyses by 16S rRNA, 60% (6) of the animals from the D28 group and 42,9% (3) of the animals from the D60 group showed amplification of 16S rRNA (Table 3, Figure 3).
3.4. hOSEC for studying the interaction between M. leprae and human skin
Considering that we used an ex vivo model of human skin that remains metabolically active over time, in order to show whether M. leprae into skin could interfere with tissue immunity, the relative expression of TGF-β, TNF-α, IFN-γ, IL-1β, IL-10 and IL-8 was measured by RT-qPCR in three female skin. IFN-γ and IL-1β showed no detectable amplification in any of the explants, with or without bacilli, with respect to the other cytokines, the presence of M. leprae notably modulated the expression by skin as compared to the control (saline).
Notoriously, viable and dead M. leprae inoculated in the skin modulated differently the expression of the cytokines, which further reinforces that the two inoculums are different and that the bacillus remained viable during the culture.
In general, viable bacillus inhibits the expression of TGF-β, IL-10 and TNF-α, and on the contrary, skin with dead bacillus has increased expression of these same targets. At the analysis times of D0 and D60, the expression pattern by live bacillus was different, resembling the dead bacillus.
For IL-8, the opposite pattern of expression between viable and dead remains, however inversely, viable M. leprae stimulated expression and dead inhibited it (Figure 4).
4. Discussion
Described 150 years ago, the intriguing M. leprae has challenged and hampered leprosy research. Currently, there is no axenic medium capable of cultivating this mycobacterium and the animal models are limited for assessment of clinical aspects of the disease [22,23,24]. In the present study, we showed a new ex vivo model for maintaining viable M. leprae and to study the interaction between bacilli and human skin.
Human organotypic skin explant culture (hOSEC) is an ex vivo model of human skin that, in addition to containing keratinocytes and fibroblasts, maintains the complexity of skin composed of other cell types (melanocytes, Langerhans cells), extracellular matrix (glycosaminoglycans, collagen) and skin structures, such as nerve filaments, vessels and glands. Ex vivo skin is already used for studies on healing, for testing cosmetics and drug absorption [25]. In culture, some authors have shown that the skin maintains its natural architecture for 14 days [26,27]. Because of the long period of multiplication of M. leprae (approximately 14 days), we cultured skin for 60 days in the present study. Frade and collaborators [13], performed similar period studies and showed the maintenance of dermal junctions for 75 days in culture and the presence of cells in the basal layer stained for Ki-67, a marker of nuclear proliferation expressed in the cell cycle phases G1, S, G2 and M but not in G0.
We performed qualitative analysis of histological sections, in which it was possible see the histomorphology of the four skins used, and these skins maintained their natural architecture until the 14th day and their viability until the 60th day and exhibiting an intact dermoepidermal junctions, in both explants, regardless of the presence of M. leprae.
In the maintenance natural process of the skin, keratinocytes from the basal epidermal layer undergo differentiation after division and migrate to the horny layer during the maturation process, during which their interior is filled with keratins and their nucleus is hydrolytically degraded [28,29]. In the present study, by histology (Figure 2), we observed the keratinocytes maturation process and stratum corneum thickening. The finding of stratum corneum thickening and the reduction in keratinocyte layers indicate the maturation of the cells although without rapid keratinocyte replacement. Xu and collaborators [30] previously showed that cultured skin exhibits an approximate 20% decrease in the rate of proliferation of basal layer keratinocyte, leading to a reduced ability to maintain the thickness of the epidermis.
The most common natural habitat of M. leprae is the human skin, where it survives and multiplies mainly inside macrophages and Schwann cells, in a process that involves the axons of the peripheral nerve system, and has the ability to parasitize other types of cells that make up the skin [31,32]. Thus, it seemed to us promising to challenge and to evaluate the survival and proliferation of M. leprae in an ex vivo model of human skin considering that this model maintains the histomorphological characteristics of host skin, including epithelial cells, fibroblasts, Langerhans cells, glands, and nerves, in addition to maintaining a limited immune response against the pathogen compared to the complete immunity in the host.
Assays for determining bacillary viability besides the subjective morphological index include the use of fluorescent dyes as markers of membrane integrity [10,33]; the evaluation of cellular biochemical metabolism using radiorespirometry [34]; and the measurement of protein synthesis by molecular biology techniques. Due to its short half-life, RNA has been successfully used as an indicator of viability for several pathogens [35,36,37].
Molecular analysis is more sensitive and specific and is more reliable for evaluating cellular viability. Martinez and collaborators [20] demonstrated a good correlation between 16S rRNA RT-PCR and clinical disease and the better sensitivity of 16S rRNA than sodA targets for monitoring leprosy therapy. More recently, Collins and collaborator [38] mention that 16S rRNA is a stable marker, that could still be detected in dead bacilli. However, 16S rRNA has been used by other researchers to evaluate the viability of M. leprae in clinical, environmental and laboratory samples [14,39,40,41,42,43,44]. Here, we used 16S rRNA RT-PCR to showing molecular viability based on the presence of rRNA until the 60th day of the hOSEC incubation period; furthermore, to corroborate this find, we demonstrated that M. leprae, cultivated in the hOSEC, beyond maintaining viability, also had its infectivity preserved because it was able to infect the footpad of several athymic mice.
Several attempts to cultivate M. leprae in axenic medium have been unsuccessful or unreproducible and have resulted in the loss of the ability of the bacilli to infect animals, as described in the 1930s, [9] the tentative to reproduce the protocol used by Vaudremer (1935), who reported the long-term maintenance of M. leprae obtained from a leprosy patient in a culture medium formulation failed, the bacilli lost their pathogenic characteristics and became non-viable. Other groups, more recently, tried to use medium under microaerophilic conditions [8,45], but within weeks, the bacilli gradually lost their capacity to grow in artificial media and survived for no more than 36 weeks of incubation.
Here, we showed the maintenance of viable bacilli, well-stained bacilli and detectable levels of 16S rRNA in explants, in addition, it has been demonstrated that after 60 days in explants these bacilli were able to infect mice and remain viable for five months in the mouse footpads, as demonstrated by ZN/FF staining and RT-PCR of macerated footpads. Unfortunately, the numbers of animals analyzed was affected by the deaths before the analysis time and by an error in handling the paw after euthanasia, mainly in the group D60, but the results, mostly from Skin 4, are encouraging, showing the maintenance of infectious bacilli after culture in the hOSEC.
Amako and collaborators [7] cultivated M. leprae bacilli in modified Kirchner medium containing several nutrients (egg yolk extract, pyruvate and transferrin) and human plasma, and maintained the bacilli for over 120 days, but without observing signs of exponential growth, not all cells that constituted the colony as globi divided or survived, M. leprae seems to have unusual replication cycles. Ferreira and collaborators [39] showed improved conditions when M. leprae was grown in arthropod cells. In the hOSEC, our results showed viability and infectivity of bacilli for 60 days, however, others experiments will be necessary to address the rate of multiplication of the bacilli in this model.
Some recent studies that demonstrated successfully maintenance of viable and replicating bacilli in protozoa, enriched medium and arthropod cells [7,39,46], using large amounts of bacilli, on the order of 107, and incubation at low temperatures, approximately 30º to 32º, considered ideal temperatures for M. leprae growth [47,48]. In our experiment we have considered 37º C for skin maintenance in culture, but low temperature should be considered in future experiments with M. leprae.
In addition to maintain viable bacilli in the hOSEC, this model proved that it can be used as one useful tool to assess the interaction between M. leprae and human skin, because it has been demonstrated that the bacilli modulated gene expression in the skin. The cytokines TGF-β, TNF-α, IL-8 and IL-10 have important roles in the inflammatory process, responses to pathogens and in other processes, such as the healing process, cell differentiation and cell migration [49,50,51,52,53,54], processes that are certainly triggered in explants after the excision of the source tissue, and after interactions with the bacilli.
Regarding the processes of cell differentiation and migration, some studies have reported that bacilli interferes with them; for example, macrophages promote mycobacterial spread during early infection [31,55], in the Schwann cells, in which M. leprae shuts off the differentiation program, bacilli change the characteristics of differentiated cells and alters the expression of the genes involved in mesenchymal endothelial transition, promoting their survival and dissemination [3,31]. In this study, clearly, viable bacilli, unlike dead ones, inhibited TGF-β expression, an important molecule involved, among other processes, in cell differentiation and proliferation.
TNF-α and IL-10, molecules involved in the amplification of the inflammatory response, in the control and Th1/Th2 balance, and which in leprosy patients have an important role in the different forms and reactional states [56,57], at hOSEC had their expression rates reduced by viable bacilli and increased by dead bacilli, as compared to skin without bacilli. IL-8, with mainly chemotactic action on cell migration for immune response [58], also had its gene expression differentially modulated between viable and dead bacilli, showing very low rates when the tissue was challenged with dead bacilli, and showing an increase in its expression in the presence of viable bacilli.
This primary insight about differential modulation for viable and dead bacilli in the skin, reinforces the important consideration of the performance of dead M. leprae during and after multidrug therapy. Besides, it corroborates the discussion on the different performances of the expression profile induced by viable and dead bacilli, for example in neuropathy [59], and it is important for understanding the prognosis of patients, and the mechanisms that M. leprae uses to challenge and subvert the immune system. In addition, this experiment further reinforces that the two inocula (viable and dead M. leprae) are different and that the bacillus remained viable during the culture periods.
Our results successfully demonstrated the maintenance of viable M. leprae in an ex vivo human skin model for up to 60 days, besides to maintaining its infective potential, demonstrated by results from athymic mice, and showed the influence of M. leprae in modifying the immunological skin response. These important and unpublished findings support the development of further experimental models for studies of M. leprae biology and its interactions, as well as clinical, immunological and drug susceptibility mechanisms.
This is the first time that human skin remaining are used as a model to cultivate M. leprae, and it has been proved to be possible. This first work opens up prospects for further studies and improvements to the hOSEC model itself, as well as helping to reduce the number of experimental animals used, especially for leprosy research.
Author Contributions
“Conceptualization, N.A. Paula and M.A.C. Frade; methodology, N.A. Paula; M.N. Leite; D.F. Bertoluci; C.T. Soares and, P.S. Rosa; validation, N.A. Paula; P.S. Rosa and M.A.C. Frade; formal analysis, N.A Paula; investigation, N.A. Paula; D.F. Bertoluci, P.S. Rosa and C.T. Soares; resources, M.A.C. Frade; data curation, N.A. Paula; writing—original draft preparation, N.A. Paula; writing—review and editing, M.N. Leite; D.F. Bertoluci, P.S. Rosa; and M.A.C. Frade; visualization, N.A. Paula; M.A.C. Frade; supervision, M.A.C. Frade; project administration, N.A. Paula; M.A.C. Frade; funding acquisition, M.A.C. Frade. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by CAPES/CNPq (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior /Conselho Nacional de Desenvolvimento Científico e Tecnológico, FAEPA (Fundação de Apoio ao Ensino, Pesquisa e Assistência do Hospital das Clínicas da Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo), Centro de Referência em Dermatologia Sanitária com Ênfase em Hanseníase, Ministério da Saúde and Fundação Paulista contra a Hanseníase.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by Research Ethics Committee of the Clinics Hospital at Ribeirão Preto Medical School, University of São Paulo (protocol number 1.744.888/2016). The animal study protocol was in accordance with the Ethical Principles in Animal Research and was approved by Local Animal Ethical Committee of the Ribeirão Preto Medical School, University of São Paulo (protocol number 026/2015-1).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
We thank Endrigo Piva Pontelli for help in enrollment of the skin donors, and Milton Osório de Moraes and Leonardo Ribeiro from FioCruz - Rio de Janeiro by the discussion about technical protocols.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hansen, G.H.A.; Looft, C. Leprosy: in its Clinical & Pathological Aspects. Reviews of books - Translated by Norman Walker, M. D. Pp. xi., 162. Bristol: John Wright & Co. 1895, 73- 74.
- Singh, A.K.; Bishai, W.R. Partners in Crime: Phenolic Glycolipids and Macrophages. Trends Mol Med. 2017, 23, 981–983. [Google Scholar] [CrossRef] [PubMed]
- Hess, S.; Rambukkana, A. Cell Biology of Intracellular Adaptation of Mycobacterium leprae in the Peripheral Nervous System. Microbiol Spectr. 2019, 7(4). [CrossRef]
- Global leprosy (Hansen disease) update, 2021: moving towards interruption of transmission. (2020). Weekly epidemiological record. Available online: https://www.who.int/publications/i/item/who-wer9837-409-430 (accessed on 08/03/2024).
- Ridley, D.S.; Jopling, W.H. Classification of leprosy according to immunity: a five-group system. Int. J. Lepr. Other Mycobact. Dis. 1966, 34, 255–273. [Google Scholar] [PubMed]
- Rodrigues, L.C.; Lockwood, D.N. Leprosy now: epidemiology, progress, challenges, and research gaps. Lancet Infect Dis. 2011, 11, 464–70. [Google Scholar] [CrossRef]
- Amako, K.; Lida, K.; Saito, M.; Ogura, Y.; Hayashi, T.; Yoshida, S. Non-exponential growth of Mycobacterium leprae Thai-53 strain cultured in vitro. Microbiol Immunol. 2016, 60, 817–823. [Google Scholar] [CrossRef]
- Ishaque, M. Attempts to Grow Mycobacterium leprae in a Medium with Palmitic Acid as the Substrate. Int. J. Lepr. Other Mycobact. Dis. 1993, 61, 294–296. [Google Scholar] [PubMed]
- Lima, M.S. Cultura do Mycobacterium leprae (Verificação dos Trabalhos de Vaudremer). Trabalho do Laboratório de Bacteriologia e Sorologia do Departamento de Prophylaxia da Lepra. 1937, Paulo. 133-140. Available online: http://hansen.bvs.ilsl.br/textoc/revistas/1937/PDF/v5n2/v5n2a01.pdf.
- Hudson, M.; Balls, M. European experimental animal use declines ever so slightly. ATLA, 2010, 38, 529–532. [CrossRef]
- Ranganatha, N.; Kuppast, A. review on alternatives to animal testing methods in drug development. Academic Sciences. 2012, 5, p. 28-32.
- Frade, M.A.C.; Andrade, T.A.M.; Aguiar, A.F.C.L.; Guedes, F.A.; Leite, M.N.; Passos, W.R.; et al. Prolonged viability of human organotypic skin explant in culture method (hOSEC). An Bras Dermatol. 2015, 90, 347–350. [Google Scholar] [CrossRef]
- Trombone, A.P.F.; Pedrini, S.C.B.; Diório, S.M.; Belone, A.F.F.; Fachin, L.R.V.; Nascimento, D.C.; et al. Optimized Protocols for Mycobacterium leprae Strain Management: Frozen Stock Preservation and Maintenance in Athymic Nude Mice. J Vis Exp. 2014, 23. [Google Scholar] [CrossRef]
- Mohanty, P.S.; Naaz, F.; Katara, D.; Misba, L.; Kumar, D.; Dwivedi, D.K.; et al. Viability of Mycobacterium leprae in the environment and its role in leprosy dissemination. Indian J Dermatol Venereol Leprol. 2016, 82, 23–27. [Google Scholar] [CrossRef]
- Rho, H.W.; Lee, B.C.; Choi, E.S.; Choi, I.J.; Lee, Y.S.; Goh, S.H. Identification of valid reference genes for gene expression studies of human stomach cancer by reverse transcription-qPCR. BMC Cancer. 2010. [Google Scholar] [CrossRef]
- Ehlers, S.; Smith, K.A. Differentiation of T cell lymphokine gene expression: the in vitro acquisition of T cell memory. J. Exp. Med. 1991, 173, 25–36. [Google Scholar] [CrossRef]
- Negera, E.; Walker, S.L.; Bobosha, K.; Bekele, Y.; Endale, B.; Tarekegn, A.; et al. The effects of Prednisolone Treatment on cytokine expression in Patients with erythema nodosum leprosum reactions. Front Immunol. 2018, 9, 1–15, DOI: 10.3389/fimmu.2018.00189. eCollection 2018. [Google Scholar]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Martinez, A.N.; Lahiri, R.; Pittman, T.L.; Scollard, D.; Truman, R.; Moraes, M.O.; et al. Molecular determination of Mycobacterium leprae viability by use of real-time PCR. J. Clin. Microbiol. 2009, 47, 2124–2130. [Google Scholar] [CrossRef]
- Shepard, C.C. The experimental disease that follows the injection of human leprosy bacilli into foot-pads of mice. J Exp Med. 1960, 112: 445-454. [CrossRef]
- Oliveira, I.V.P.M.; Deps, P.D.; Antunes, J.M.A.P. Armadillos and leprosy: from infection to biological model. Rev Inst Med Trop São Paulo. 2019, 61:e44. [CrossRef]
- Adams, L.B. Susceptibility and resistance in leprosy: Studies in the mouse model. Immunol Rev. 2021 301(1):157-174. [CrossRef]
- Wang, H.; Maeda, Y.; Fukutomi, Y.; Makino, M.; An in vitro model of Mycobacterium leprae induced granuloma formation. BMC Infect Dis. 2013 20; 13:279. [CrossRef]
- Leite, M.N.; Viegas, J.S.R.; Praça, F.S.G.; Paula, N.A.; Ramalho L.N.Z.; Bentley, M.V.L.B.; Frade, M.A.C. Ex vivo model of human skin (hOSEC) for assessing the dermatokinetics of the anti-melanoma drug Dacarbazine. Eur J Pharm Sci. 2021 1:160:105769. [CrossRef]
- Danso, M.O.; Berkers, T.; Mieremet, A.; Hausil, F.; Bouwstra, J.A. An ex vivo human skin model for studying skin barrier repair. Exp. Dermatol. 2014, 24, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Garcia, J.; Sebastian, A.; Alonso-Rasgado, T.; Bayat, A. Optimization of an ex vivo wound healing model in the adult human skin: Functional evaluation using photodynamic therapy. Wound Repair Regen. 2015, 23, 685–702. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, J.; Green, H. Differentiation-related death of an established keratinocyte line in suspension culture. J. Cell. Physiol. 1978, 93,3, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.T.; Green, H. Differentiation of the epidermal keratinocyte in cell culture: formation of the cornified envelope. Cell. 1976, 9, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Hong, S.J.; Jia, S.; Zhao, Y.; Galiano, RD.; Mustoe, T.A. Application of a partial-thickness human ex vivo skin culture model in cutaneous wound healing study. Lab. Invest. 2012, 92, 584–599. [Google Scholar] [CrossRef]
- Masaki, T.; Qu, J.; Cholewa-Waclaw, J.; Burr, K.; Raaum, R.; Rambukkana, A. Reprogramming Adult Schwann Cells to Stem Cell-like Cells by Leprosy Bacilli Promotes Dissemination of Infection. Cell. 2012, 152, 51–67. [Google Scholar] [CrossRef]
- Teixeira, C.S.; Montalvão, P.P.; Oliveira, I.T.; Wachholz, P.A. Neoplastic cells parasitized by Mycobacterium leprae: report of two cases of melanocytic nevus and one of basal cell carcinoma. Surgical and Experimental Pathology. 2019, 2, 1–7. [Google Scholar] [CrossRef]
- Lahiri, R.; Randhawa, B.; Krahenbuhl, J. Application of a viability-staining method for Mycobacterium leprae derived from the athymic (nu/nu) mouse foot pad. J. Med. Microbiol. 2005, 54, 235–242. [Google Scholar] [CrossRef]
- Truman, R.W.; Krahenbuhl, J.L. Viable M. leprae as a research reagent. Int. J. Lepr. Other Mycobact. 2001, Dis. 69, 1–12. [Google Scholar]
- Baque, R.H.; Gilliam, A.O.; Robles, L.D.; Jakubowski, W.; Slifko, T.R. A real-time RT-PCR method to detect viable Giardia lamblia cysts in environmental waters. Water Res. 2011, 45, 3175–3184. [Google Scholar] [CrossRef]
- Montenegro, R.A.; Guarines, K.M.; Montenegro, L.M.L.; Lira, L.A.S.; Falcão, J.; Melo, F.L.; et al. Assessment of messenger RNA (mRNA) of Mycobacterium tuberculosis as a marker of cure in patients with pulmonary tuberculosis. J. Appl. Microbiol. 2014, 117, 266–272. [Google Scholar] [CrossRef]
- Turankar, R.P.; Turankara, R.P.; Lavania, M.; Darlongd, J.; Saib, K.S.R.S.; Senguptaa, U.; et al. Survival of Mycobacterium leprae and association with Acanthamoeba from environmental samples in the inhabitant areas of active leprosy cases: A cross sectional study from endemic pockets of Purulia, West Bengal. Infect. Genet. Evol. 72, 199-204. [CrossRef]
- Collins, J.H.; Lenz, S.M.; Raya, N.A.; Lahiria R.; Adams L.B. Assessment of esxA, hsp18, and 16S transcript expression as a measure of Mycobacterium leprae viability: A comparison with the mouse footpad assay. Lepr Rev (2023) 94, 7–18. [CrossRef]
- Ferreira, J.S.; Oliveira, D.A.S.; Santos, J.P.; Ribeiro, C.C.D.U.; Baêta, B.A.; Teixeira, R.C.; et al. Ticks as potential vectors of Mycobacterium leprae: Use of tick cell lines to culture the bacilli and generate transgenic strains. PLoS Negl Trop Dis. 12. 2018. [CrossRef]
- Lavania, M.; Katoch, K.; Katoch, V.M.; Gupta, A.K.; Chauhan, D.S.; Sharma, R.; et al. Detection of viable Mycobacterium leprae in soil samples: insights into possible sources of transmission of leprosy. Infect Genet Evol. 2008, 8, 627–631. [Google Scholar] [CrossRef]
- Martinez, A.N.; Talhari, C.; Moraes, M.O.; Talhari, S. PCR-Based Techniques for Leprosy Diagnosis: From the Laboratory to the Clinic. PLoS Negl Trop Dis. 2014, 8. [Google Scholar] [CrossRef]
- Phetsuksiri, B.; Rudeeaneksin, J.; Supapkul, P.; Wachapong, S.; Mahotarn, K.; Brennan, P.J. A simpliced reverse transcriptase PCR for rapid detection of Mycobacterium leprae in skin specimens. FEMS Immunol Med Microbiol. 2006, 48, 319–328. [Google Scholar] [CrossRef]
- Singh, I., Ahuja, M., Lavania, M., Pathak, V.K., Turankar, R.P., Singh, V., et al. Efficacy of fixed duration multidrug therapy for the treatment of multibacillary leprosy: A prospective observational study from Northern India. Indian J Dermatol Venereol Leprol. 2023. [CrossRef]
- Neumann, A.S.; Fontes, A.N.B.; Lopes, M.Q.P.; Suffys, P.N.; Moraes, M.O.; Lara, F.A. Heterogeneous persistence of Mycobacterium leprae in oral and nasal mucosa of multibacillary patients during multidrug therapy. Mem Inst Oswaldo Cruz, 2022, 117. [CrossRef]
- Ishaque, M. Growth of Mycobacterium leprae under low oxygen tension. Microbios. 1990, 64, 7–17. [Google Scholar] [PubMed]
- Wheat, W.H.; Casali, A.L.; Thomas, V.; Spencer, J.S.; Lahiri, R.; Williams, D.L.; et al. Long-term Survival and Virulence of Mycobacterium leprae in Amoebal Cysts. PLoS Negl Trop Dis. 2014, 8. [Google Scholar] [CrossRef]
- Balamayooran, G.; Pena, M.; Sharma, R.; Truman, R.W. The armadillo as an animal model and reservoir host for Mycobacterium leprae. Clin. Dermatol. 2015, 33, 108–115. [Google Scholar] [CrossRef]
- Kirchheimer, W.F.; Storrs, E.E. Attempts to establish the armadillo (Dasypus novemcinctus Linn.) as a model for the study of leprosy. I. Report of lepromatoid leprosy in an experimentally infected armadillo. Int. J. Lepr. Other Mycobact. Dis. 1971, 39, 693–702.
- Balbino, C.A.; Pereira, L.M.; Curi, R. Mecanismos envolvidos na cicatrização: uma revisão. Revista Brasileira de Ciências Farmacêuticas. 2005, 41, 27-51. DOI.org/10.1590/S1516-93322005000100004.
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [CrossRef]
- Clark, D.A.; Coker, R. Transforming growth factor-beta (TGF-β). Int. J. Biochem. Cell Biol. 1998, 30, 293–298. [Google Scholar] [CrossRef]
- Gailit, J.; Welch, M.P.; Clark, R.A.F. TGF-beta 1 stimulates expression of keratinocyte integrins during re-epithelialization of cutaneous wounds. J. Invest. Dermatol. 1994, 103, 221–227. [Google Scholar] [CrossRef]
- Rennekampff, H.O.; Hansbrough, J.F.; Kiessig, V.; Doré, C.; Sticherling, M.; Schröder, J.M. Bioactive interleukin-8 is expressed in wounds and enchances wound healing. J. Surg. Res. 2000, 93, 41–54. [Google Scholar] [CrossRef]
- Sato, Y.; Ohshima, T.; Konto, T. Regulatory role of endogenous interleukin-10 in cutaneous inflammatory response of murine wound healing. Biochem. Biophys. Res. Commum. 1999, 256, 194–199. [Google Scholar] [CrossRef]
- Davis, J.M.; Ramakrishnan, L. The Role of the Granuloma in Expansion and Dissemination of Early Tuberculous Infection. Cell. 2009, 136, 37–49. [Google Scholar] [CrossRef]
- Oktariana, D.; Saleh, I.; Hafy Z.; Liberty, I.A.; Salim E.M.; Legiran L. The Role of Interleukin-10 in Leprosy: A Review. Indian Journal Leprosy. 2022, 94: 321-334.
- Barnes, P.F.; Chatterjee, D.; Brennan, P.J.; Rea, T.H.; Modlin, R.L. Tumor Necrosis Factor Production in Patients with Leprosy. Infection and immunity. 1992, 1441–1446. [Google Scholar] [CrossRef]
- Matsushima, K.; Yang, D.; Oppenheim, J.J. Interleukin-8: An evolving chemokine. Cytokine. 2022, 153. [Google Scholar] [CrossRef]
- Souza, B.J.D.; Mendes, M.A.; Silva, G.M.S.; Sammarco-Rosa, P.; Moraes M.O.; Jardim, M,R.; et. al Gene Expression Profile of Mycobacterium leprae Contribution in the Pathology of Leprosy Neuropathy. Front. Med. 2022, 9:861586. [CrossRef]
Figure 1.
Photo of hOSEC explants on grids in culture plates. Explants of skin obtained from 0.8-cm diameter punch. The explants were placed on filter paper and metal grids in a 6-well culture plate with DMEM.
Figure 1.
Photo of hOSEC explants on grids in culture plates. Explants of skin obtained from 0.8-cm diameter punch. The explants were placed on filter paper and metal grids in a 6-well culture plate with DMEM.
Figure 2.
Histomorphology of skin from hOSEC. Sections of skin explants stained with hematoxylin and eosin, from left to right, the culture times are D0, D7, D14, D28 and D60. The images were obtained with a 40x objective and Leica Application Suite (LAS) software version 3.2.0 using the Stretch Image tool. C: corneal layer; K: epidermis, keratinocyte layer, DP: dermal papilla; arrow: basal layer, dermo-epidermal junction. Furthermore, no inflammatory processes were observed with the recruitment of defense cells.
Figure 2.
Histomorphology of skin from hOSEC. Sections of skin explants stained with hematoxylin and eosin, from left to right, the culture times are D0, D7, D14, D28 and D60. The images were obtained with a 40x objective and Leica Application Suite (LAS) software version 3.2.0 using the Stretch Image tool. C: corneal layer; K: epidermis, keratinocyte layer, DP: dermal papilla; arrow: basal layer, dermo-epidermal junction. Furthermore, no inflammatory processes were observed with the recruitment of defense cells.
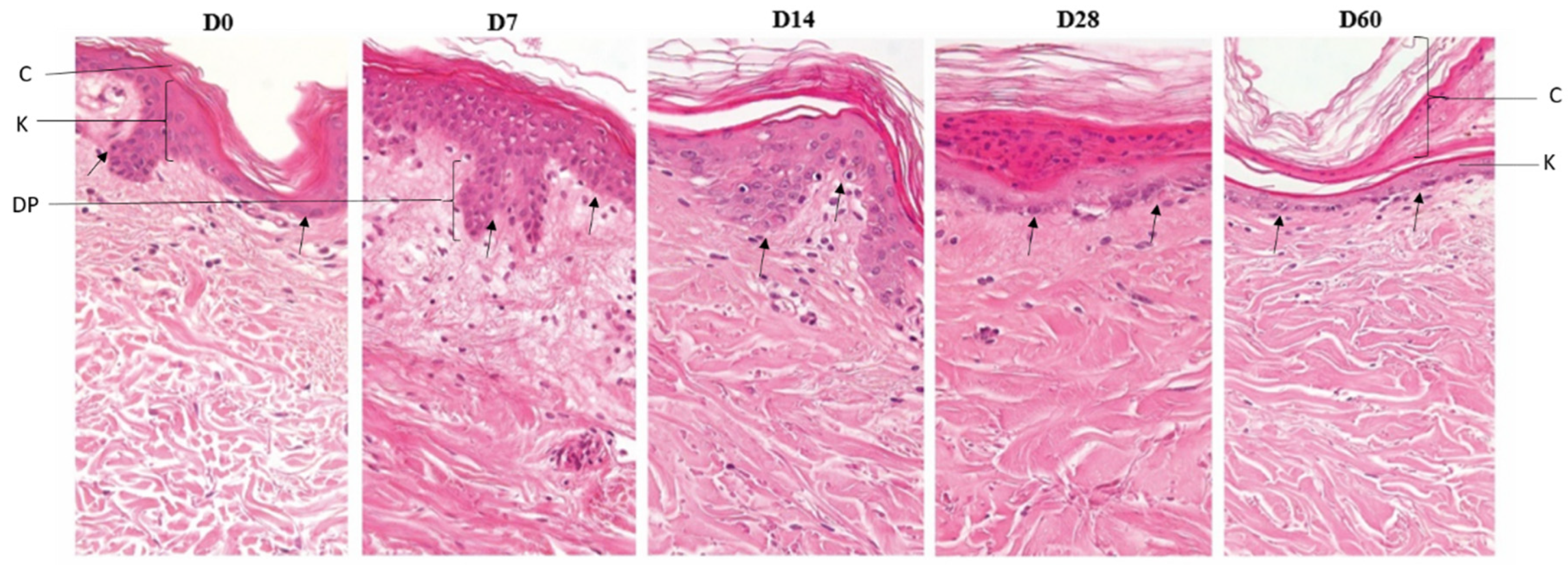
Figure 3.
M. leprae in skin from hOSEC and in footpads from athymic mice inoculated. Fite-Faraco staining (bacillus-specific stain) showing intact and well-stained bacilli. 1000x magnification. (A) Histological section from explant with M. leprae showing bacilli in the keratinocyte layer (arrowhead) and superficial and deep dermis regions (arrow). CL = corneal layer, SD = superficial dermis, DD = deep dermis. (B) Histological section from athymic mouse footpad after 5 months of inoculation with a suspension from D28 culture, the arrow showing bacilli.
Figure 3.
M. leprae in skin from hOSEC and in footpads from athymic mice inoculated. Fite-Faraco staining (bacillus-specific stain) showing intact and well-stained bacilli. 1000x magnification. (A) Histological section from explant with M. leprae showing bacilli in the keratinocyte layer (arrowhead) and superficial and deep dermis regions (arrow). CL = corneal layer, SD = superficial dermis, DD = deep dermis. (B) Histological section from athymic mouse footpad after 5 months of inoculation with a suspension from D28 culture, the arrow showing bacilli.
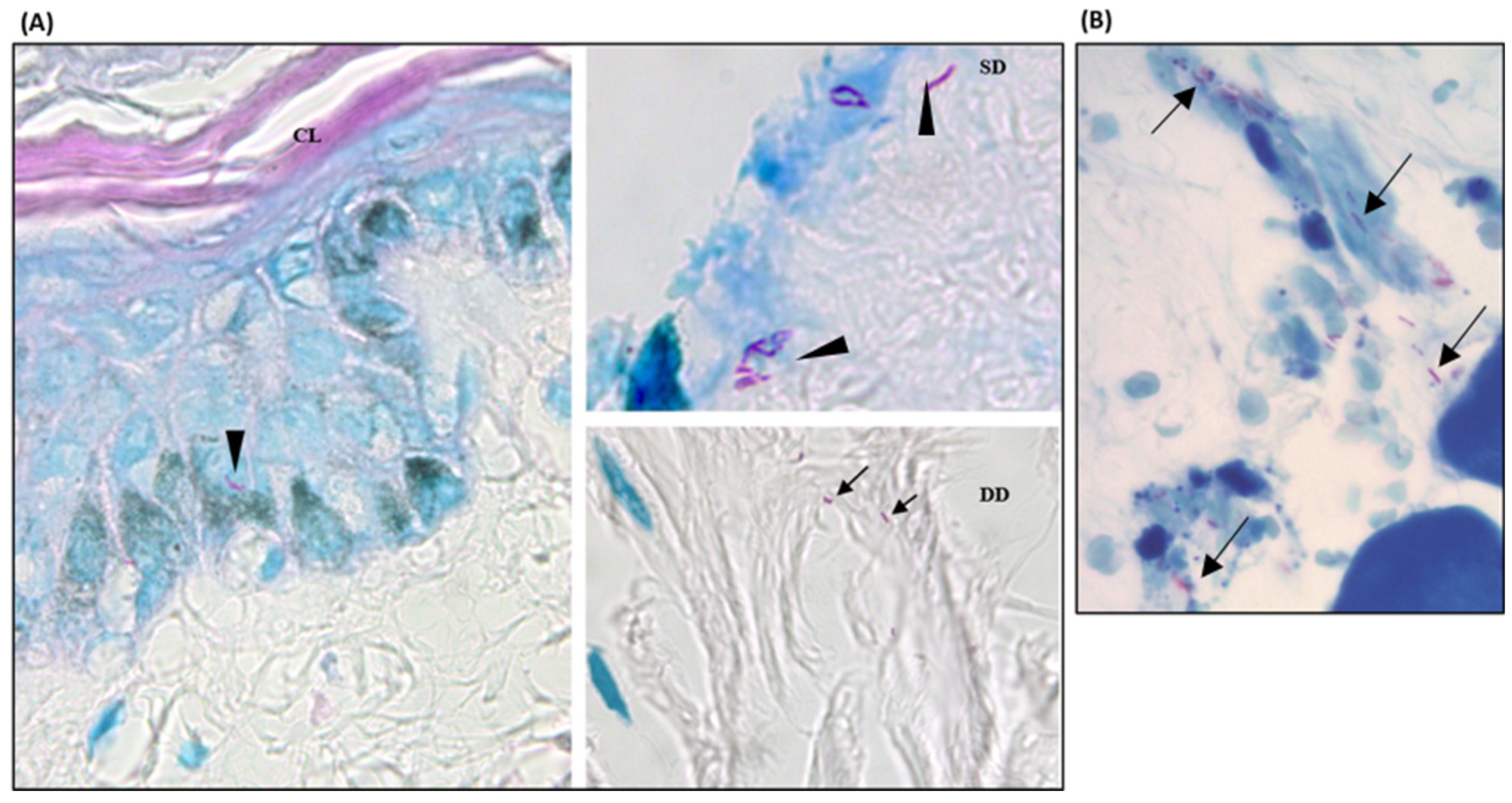
Figure 4.
Interaction between M. leprae and skin. Relative gene expression of cytokines (A) TGF-β, (B) IL-10, (C) TNF-α and (D) IL-8 in fragment with viable M. leprae (VML) and dead M. leprae (DML) comparing with saline group during the follow-up period, calculated by the 2ΔΔCT method based on CT value obtained from RT-PCR using 18S rRNA as reference gene and the Saline group as Normalizer. The relative expression of each cytokine in 3 female skin explants with viable M. leprae and dead M. leprae.
Figure 4.
Interaction between M. leprae and skin. Relative gene expression of cytokines (A) TGF-β, (B) IL-10, (C) TNF-α and (D) IL-8 in fragment with viable M. leprae (VML) and dead M. leprae (DML) comparing with saline group during the follow-up period, calculated by the 2ΔΔCT method based on CT value obtained from RT-PCR using 18S rRNA as reference gene and the Saline group as Normalizer. The relative expression of each cytokine in 3 female skin explants with viable M. leprae and dead M. leprae.
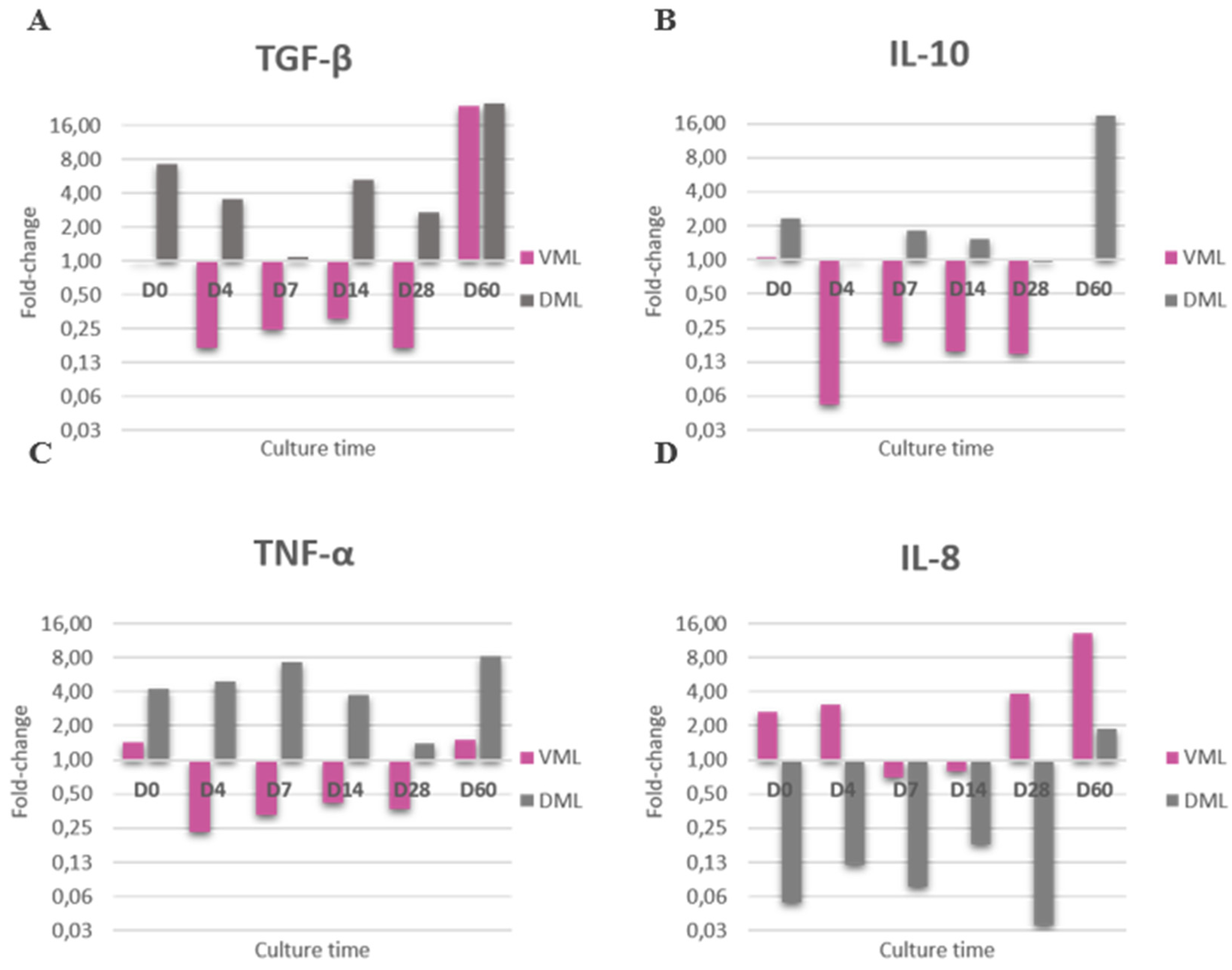

Table 1.
Primer sequences and cycling protocol.
| Target | Sequence | Cycling | Reference |
|---|---|---|---|
| 16S rRNA | 5' TCGAACGGAAAGGTCTCTAAAAAATC 3’ 5' CCTGCACCGCAAAAAGCTTTCC 3' |
2 min at 95°C; 45 cycles of 2 min at 94°C, 2 min at 60°C and 3 min at 72°C; and 10 min at 72°C | [14] |
| 18S rRNA | 5' GTAACCCGTTGAACCCCATT 3' 5' CCATCCAATCGGTAGTAGCG 3' |
2 min at 95°C; 38 cycles of 94°C for 2 min, 60°C for 2 min and 72°C for 3 min; and 72°C for 10 min. | [15] |
| IL-1β | 5’ CTTCATCTTTGAAGAAGAACCTATCTTCTT 3’ 5’ AATTTTTGGGATCTACACTCTCCAGCTGTA 3’ |
95°C for 2 min and 45 cycles of 95°C for 5 sec, 62°C for 10 sec and 72°C for 20 sec. | [16] |
| TGF-β | 5' ACATCAACGCAGGGTTCACT 3' 5' GAAGTTGGCATGGTAGCCC 3' |
95°C for 2 min and 45 cycles of 95°C for 5 sec, 60°C for 10 sec and 72°C for 20 sec. | [17] |
| TNF-α | 5’ TGGCTTTCACATACTGCTGGTA 3’ 5′ GCTGGTTATCTCTCAG CTCCA 3' |
95°C for 2 min and 45 cycles of 95°C for 5 sec, 60°C for 10 sec and 72°C for 20 sec. | [17] |
| IFN-γ | 5’ GGCTTTTCAGCTCTGCATCG 3’ 5’ TCTGTCACTCTCCTCTTTCCA 3’ |
95°C for 2 min and 45 cycles of 95°C for 5 sec, 60°C for 10 sec and 72°C for 20 sec. | [17] |
| IL-8 | 5’ ACCGGAAGGAACCATCTCAC 3’ 5’ AAACTGCACCTTCACACAGAG 3’ |
95°C for 2 min and 45 cycles of 95°C for 5 sec, 60°C for 10 sec and 72°C for 20 sec. | [17] |
| IL-10 | 5’ TGAGAACCAAGACCCAGACA 3’ 5’ TCATGGCTTTGTAGATGCCT 3’ |
95°C for 2 min and 45 cycles of 95°C for 5 sec, 60°C for 10 sec and 72°C for 20 sec. | [17] |
Table 2.
Cycle threshold of 16S rRNA by RT-PCR.
| Day | Skin 1 | Skin 2 | Skin 3 | Skin 4 |
|---|---|---|---|---|
| D0 | 28,05 | 34,73 | 30,29 | UR |
| 37,32 | 38,30 | 33,51 | UR | |
| ND | 35,34 | 34,95 | ND | |
| D4 | 28,66 | 32,66 | 28,74 | 29,21 |
| 28,45 | 31,70 | 29,84 | 28,17 | |
| 27,70 | 31,08 | 28,01 | 28,03 | |
| D7 | 29,21 | 30,75 | 33,35 | 28,62 |
| 29,77 | 30,27 | 33,19 | 29,16 | |
| 29,70 | 33,21 | 29,43 | 31,28 | |
| D14 | 29,71 | 29,80 | 29,80 | 29,12 |
| 29,01 | 29,19 | 28,92 | 30,10 | |
| 30,00 | 30,54 | 28,79 | 31,37 | |
| D28 | 31,03 | 30,65 | 28,37 | 33,13 |
| 31,27 | 32,37 | 31,61 | 29,31 | |
| 31,43 | 33,82 | 33,70 | ND | |
| D60 | 31,30 | UR | 29,81 | 30,61 |
| 37,17 | UR | 30,22 | ND | |
| 39,71 | UR | 32,90 | UR |
CT value of 16S rRNA RT-PCR in each explant inoculated with bacilli during the follow-up period. The lowest and highest CT values for each skin sample are in bold. ND= undetected; UR= not performed.
Table 3.
Positivity in footpads inoculated with bacilli from hOSEC after 28 and 60 days of culturing.
Table 3.
Positivity in footpads inoculated with bacilli from hOSEC after 28 and 60 days of culturing.
| Inoculum (nº of animals) | ZN (+) (%) | FF (+) (%) | RT-PCR (+) (%) | At least one microscopy analysis (+ ) (%) | ZN and RT-PCR (+) (%) | FF and RT-PCR (+) (%) | At least one microscopy analysis (+) and RT-PCR (+) (%) |
|---|---|---|---|---|---|---|---|
| D28 (10) | 6 (60,0) | 5 (50,0) | 6 (60,0) | 8 (80) | 4 (40,0) | 4 (40,0) | 6 (60,0) |
| D60 (7) | 3 (42,9) | 0 (0,0) | 3 (42,9) | 3 (42,9) | 1 (14,3) | 0 (0,0) | 1 (14,3) |
| Total (17) | 9 (52,9) | 5 (50,0) | 9 (52,9) | 11 (64,7) | 5 (29,4) | 4 (40,0) | 7 (41,2) |
Positivity (+) for M. leprae in the footpads of mice after 5 months of inoculation with bacilli suspension from male explants (Skin 4). D28 = inoculum from explants in culture for 28 days; D60 = inoculum from explants in culture for 60 days. ZN = Ziehl-Neelsen, performed in one macerated foot pad; FF = Fite-Faraco, performed in half of the second foot pad; RT-PCR= RT-PCR 16Sr RNA performed in the another half of the second foot pad.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



