
Submitted:
13 May 2024
Posted:
13 May 2024
You are already at the latest version
Abstract
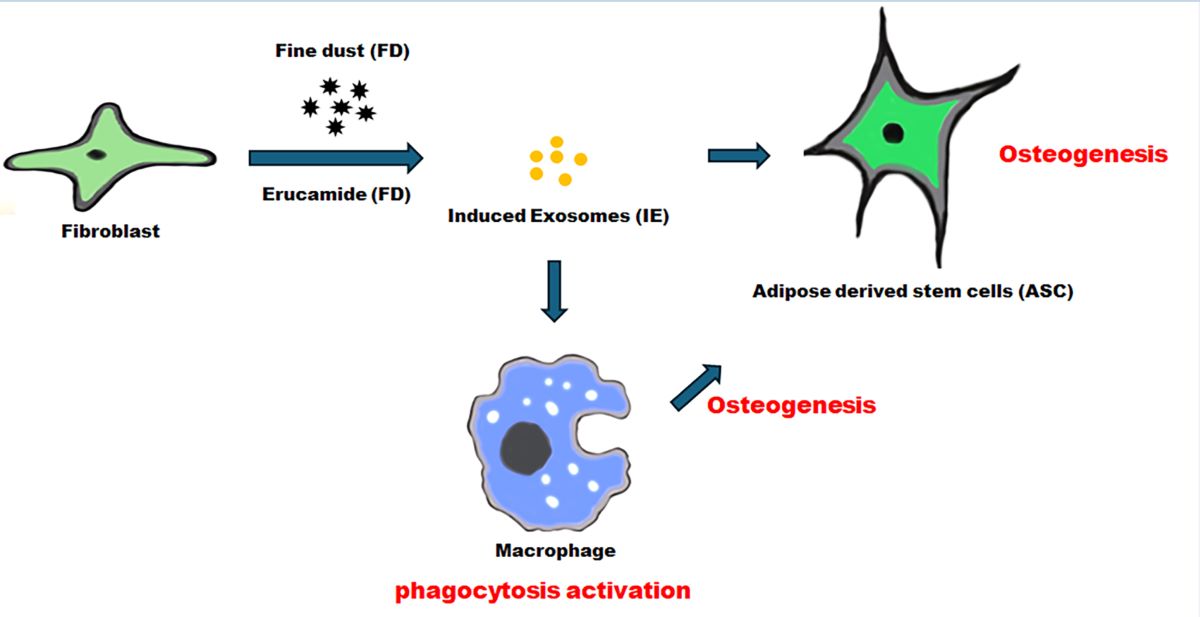
Fine dust causes various disorders, including cardiovascular, neurological, renal, reproductive, motor, systemic, respiratory, and cancerous diseases. Therefore, it is necessary to study functional materials to prevent these problems. In this study we used various methods, including quantitative PCR, flow cytometry, ELISA, and Alizarin O staining, to study the beneficial effects of erucic acid against fine dust. Erucic acid and erucic acid-induced exosomes (EIE) strongly suppress the attenuation of osteocytic differentiation and inflammation induced by fine dust. Erucic acid effectively suppresses inflammation in fibroblasts exposed to fine dust. Notably, despite exposure to fine dust, EIE promoted osteocytic differentiation in adipose-derived stem cells (ASC) and osteogenesis in macrophages, in addition to phagocytosis. Dramatic upregulation of RunX2 and BMP7 by EIE indicates that it is a strong activator of osteocytic differentiation in ASC and protects them from the effects of fine dust. Moreover, EIE induces immune activity and acts as an osteogenic trigger for macrophages. Therefore, this study suggests the potential use of erucic acid and erucic acid-induced exosomes (EIE) as biomaterials to prevent environment-borne diseases.
Keywords:
osteogenesis
; macrophage
; erucic acid
; fine dust
; stem cell
1. Introduction
Erucic acid, a monounsaturated omega-9 fatty acid, is found in seed oils of various plants, especially from the Brassicaceae family[1,2]. According to a previous study[3], rapeseed, mustard and wallflower seed oils contain high concentrations of erucic acid. This compound is used in the treatment of toxic oil syndrome and reduces cardiotoxicity in rats [4,5]. The beneficial effects of erucic acid include therapeutic effects in neurodegenerative diseases [2], obesity-induced metabolic disorders, melanomas, and diabetes. It also has anti-inflammatory and antioxidant properties [6]. Although erucic acid is classified as a toxicant, the maximum edible quantity for this compound is 7.5 mg/kg body weight/day [7].
Exosomes are approximately 40–100 nm in size, and almost all cell types secrete them into either the serum, urine, cerebrospinal fluid, ascites fluid, milk, or saliva [8,9]. Exosomes contain various molecules, including functional protein, carbohydrate, mRNA, microRNA (miRNA), and DNA molecules [10]. In general, cells exposed to a stimulant, upon induction, secrete exosomes that dramatically alter their components compared to unstimulated conditions [11]. Altered exosomes can have adverse or beneficial effects on the surrounding cells [11]. In addition, exosomes have various functions, including the modulation of the immune system, prognostic biomarkers for diseases, and cancerous activity in the human body [9,11]. Due to these characteristics, induced exosomes have the potential to be used as biofunctional materials in various fields, including pharmaceuticals, cosmetics, and foods [11].
Fine dust, or particulate matter, can be found in air, and fine dust particles sized 2.5 μm (PM2.5) and 10 μm (PM10) are very harmful to the human body [12]. In particular, elderly people and children are more susceptible to fine dust [13,14]. Fine dust has been shown to cause various disorders, including cardiovascular, neurological, renal, reproductive, motor, systemic, respiratory, and cancerous diseases. [15,16]. According to a recent study [17], fine dust suppresses the osteogenic differentiation of adipose-derived stem cells. Moreover, fine dust causes inflammation in the dermal tissues in humans and pets [17]. Skin inflammation causes adverse dermal immunity leading to differentiation of adipose-derived stem cells (ASC) in the subcutaneous fatty tissues of the skin [17,18]. When exposed to fine dust, dermal cells upregulate apoptotic proteins, including BAX and CytC, and downregulate antiapoptotic proteins, including AKT, P50, P52, and BCL-2 [17].
Osteogenesis is important for the bone regeneration associated with age-related bone diseases [19]. Changes in the expression of various genes occurs during the osteogenic stages of mesenchymal differentiation. The levels of RUNX2 and DLX5 are increased in preosteoblasts, while osteoponin and SOST-sclerostin are expressed in osteoblasts and osteocytes, respectively [17,20,21]. In contrast, p53 downregulates RUNX2 and osterix in ASCs [21]. Ordinarily, expression of VEGF and TGF-β, known as osteogenic activating markers, are down regulated in fibroblasts during inflammation. Additionally, aromatase, collagenase type 1, RUNX2, SAMD4, BMP6, and BMP7 are known activators of the osteogenic differentiation of ASCs [22-24]. In immune system of bone homeostasis, macrophages secrete cytokines, including TNF-α, IL-1, and IL-6, to inhibit bone formation and the cytokines, such as IL-4, IL-10, and IL-13, inhibit the differentiation of osteoclasts [25].
Various factors [26,27] and the deterioration of the environment have worsened the problem of fine dust [17]. Therefore, it is necessary to study functional materials to prevent the health problems that may arise from fine dust. This study investigates the potency of erucic acid and erucic acid-induced exosomes as biomaterials to prevent environment-borne diseases.
2. Results
We studied the biological functions of erucic acid and erucic acid-induced exosomes in the prevention of osteogenic suppression and inflammation caused by fine dust exposure to dermal cells, immune cells, and ASC.
2.1. Antiapoptotic function of erucic acid in fibroblasts exposed to fine dust
Based on the obtained results (Figure 1), treatment dosages for erucic acid and fine dust (PM10) were established as 250 μmole/mL and 20μg/mL, respectively, in fibroblasts (Figure 1a, b). Under the four conditions ( Con, E, FD, EFD), the concentrations exosomes isolated from fibroblasts were approximaely 20, 25, 15 and 20 μg/mL, respectively (Figure 1c, d). The levels of osteogenic activators were increased upon exposure to erucic acid in fibroblasts (Figure 2). Upon erucic acid exposure, the VEGF levels were approximately 6 times higher, while the levels of TGF-β were 4 times higher, than upon FD exposure (Figure 2a, b).
Although exposure to fine dust downregulated VEGF and TGF-β in fibroblasts, erucic acid prevented the suppression of the two markers in the cells (Figure 2a). Additionally, erucic acid enhanced the expression of the markers by approximately 3.2 times in comparison with the control (Figure 2a). Further, erucic acid induced the upregulation of antiapoptotic markers in fibroblasts in addition to protecting against fine dust (Figure 2b).
2.2. Protecting and Enhancing of osteocyic differentiation by the induced exosomes
The erucic acid-induced exosomes (EIE), isolated from fibroblasts, enhanced expression of the osteocyic differentiation markers, including aromatase, CT1 (collagenase type 1), RunX2, TGF-β, VEGF, SAMD4, BMP6, and BMP7 (Figure 3). In particular, RunX2, BMP 6, and BMP7 were dramatically upregulated in ASCs exposed to EIE. Despite exposure to fine dust, increased levels of the markers were observed in erucic-acid induced exosomes (EFDIE)(Figure 3a, b, c). Alizarin staining results (Figure 4) corresponded with the results shown in figure 3; ASCs were strongly differentiated into osteocytes at day 5, and then formed osteocytic colonies under EIE and EFDIE conditions on day 15 (Figure 4).
2.3. Activation of immunity in macrophages with induced exosomes
Macrophages exposed to fine dust showed attenuated phagocytic activity; in contrast, this activity was strongly enhanced under EIE and EFDIE conditions (Figure a to d). Compared with the FD condition, the phagocytic activity was approximately 10 and 7.5 times higher under EIE and EFIE conditions, respectively (Figure 5c, d). Upon evaluation of osteogenesis modulating cytokines (Figure 6), we found that osteogenesis inhibiting cytokines (TNF-α, IL-6) were downregulated in macrophages under EIE and EFDIE conditions, in contrast to FDIE condition (Figure 6a, b). However, osteogenesis activating cytokines were upregulated in macrophages under EIE and EFDIE conditions (Figure 6a, b). Notably, EIE significantly modulated IL-6 and IL-4 levels in macrophages (Figure 6b).
3. Discussion
In this study, we show that erucic acid and erucic acid-induced exosomes (EIE) directly affect the activation of osteocytic differentiation in ASCs, activation of osteogenesis in macrophages, activation of the immune response, and protection against fine dust in fibroblasts.
First, erucic acid prevented fibroblasts from being inflamed upon exposure to 50 ng/mL fine dust (S1) (Figure 2). Recently, fine dust has been found to contribute significantly to respiratory, circulatory, and skin diseases [28,29]. As erucic acid is considered a toxicant, this compound could be a better cosmetic biomaterial with greater safety for the skin than a functional food additive. According to previous studies [30,31], fine dust causes an increase in ROS in dermal cells, and here we show that erucic acid induced an increase in anti-ROS proteins, including AKT, P52, P50, and BCL2, in fibroblasts (Figure 2).
Second, two materials, erucic acid and erucic acid-induced exosomes (EIE) from fibroblasts, protected and activated osteocytic differentiation against fine dust. Erucic acid upregulated activators (VEGF, TGF-β) for osteocytic differentiation in fibroblasts upon exposure to fine dust (Figure 2a). Additionally, EIE increased the expression of the activating factors of osteocytic differentiation, including aromatase, CT1, RunX2, VEGF, TGF-β, BMP6, and BMP7, in adipose-derived stem cells upon exposure to fine dust (Figure 3 a, b, c). VEGF, TGF-β, BMP6, and BMP7 were reported as a triggers for osetocytic differentiation in stem cells [32,33]. Additionally, in the pre-osteoblast stage, CT1 and RunX2 are upregulated during stem cell osteocytic differentiation [34,35]. In particular, RunX2 acts as an activator of bone matrix protein synthesis in immature osteoblasts [35]. As observed in Figure 3, EIE intensely enhanced the expression of RunX2 and BMP 6, and BMP7. Based on the formation of calcium granules observed using Alizarin staining (Figure 4), we can conclude that EIE activates mineralization in differentiating ASCs. RunX2 plays a role of an activator of mineralization in immature osteoblasts [36]. A dramatic increase in the number of calcium granules (Figure 4) was induced upon increase in the RunX2 levels during osteogenesis. These results suggest that EIE triggers osteocytic differentiation and is a developmental activator of osteogenesis in stem cells. Moreover, EIE plays an effective protective role against fine dust in fibroblasts.
Third, EIE activates osteogenesis in macrophages involved in the immune activity of macrophages exposed to fine dust. Macrophages act as osteogenesis modulators and immune conductors in humans [36,37]. They affect bone formation via several factors [25]. Macrophages secrete cytokines, such as TNF-α, IL-1, and IL-6, which are associated with inhibiting bone formation and promoting bone resorption [25]. IL-4, IL-10, and IL-13 secreted by macrophages inhibit osteoclast differentiation and OSM, while VEGF, IGF, TGF-β, and BMP-2 activate osteoblast differentiation [25]. EIE activated upregulation of IL-4 and IL-13, in contrast with TNF-α and IL-6, in differentiating cells (Figure 6 a, b). Among the cytokines, TNF-α and IL-6 were affected the most strongly by EIE in the differentiating cells (Figure 6 a, b). These results suggest that EIE prevents upregulation of TNF-α and IL-6 by fine dust and activates osteogenesis through upregulation of IL-4 and IL-13, despite exposure to fine dust, in differentiating cells. The ordinary phagocytic activity of immune cells is attenuated by fine dust [38]. However, EIE protected the macrophages from fine dust (Figure 5). The activities in EFDIE-exposed cells were stronger than those in FDD-exposed cells, and their strength was approximately 7.5 times higher than that of FDD-exposed cells (Figure 5). Briefly, EIE effectively activates phagocytic activity and osteogenesis in macrophages.
4. Materials and Methods
4.1. Cell culture
Fibroblasts (Korea Cell Bank, Seoul, Korea) were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 100 µg/mL penicillin, and 100 µg/mL streptomycin at 37 °C and 5% CO2. Adipose derived stem cells (ASC) (Thermo Fisher Scientific, Waltham, MA, USA) were cultured in a MesenPRO RS™ Basal Medium, (Gibco, Thermo Fisher Scientific) with growth supplement (MesenPRO RS™ Growth Supplement, Thermo Fisher Scientific). Macrophages (KG-1, ATCC) were cultured in Iscove’s modified Dulbecco's medium (IMDM) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 100 µg/mL penicillin, and 100 µg/mL streptomycin at 37 °C and 5% CO2.
4.2. Cell viability test
To establish treatment dosages, fibroblasts were exposed to 5, 50, 100, 500, and 1000 µM of erucic acid (45629, Sigma-Aldrich, St. Louis, MO, USA) and 1, 5, 10, 20, and 50 µg/mL of PM10 (ERMCZ120; Sigma-Aldrich). For the induced exosomes (S1), ASCs and macrophages were exposed to 1, 5, 10, 50, or 100 ng/mL of induced exosomes form the four conditions (control, erucic acid, fine dust, and erucic acid + fine dust) for 1 d. To evaluate viability, all exposed cells were stained with Annexin V-conjugated propidium iodide (PI) (Invitrogen, Carlsbad, CA, USA) and analyzed using a flow cytometer (FACS calibur, BD Biosciences, San Jose, CA, USA) with the FlowJo 10.10 software (BD Biosciences).
4.3. Evaluation of the concentration of induced exosomes
After exposure to either of the four conditions (control, erucic acid, fine dust, and erucic acid + fine dust) for one day, the induced exosomes (CE, EIE, FDIE, and EFDIE) were isolated from the culture media of fibroblasts using the exoEasy Maxi Kit (QIAGEN, Hilden, Germany) and CD68 Exo-Flow Capture Kit (System Biosciences, Palo Alto, CA, USA), and the concentrations of the isolated exosomes were evaluated using the exosome standards kit (Sigma-Aldrich). Purified exosomes were evaluated using a flow cytometer (FACSCalibur) with the and FlowJo 10.10 software (BD Biosciences).
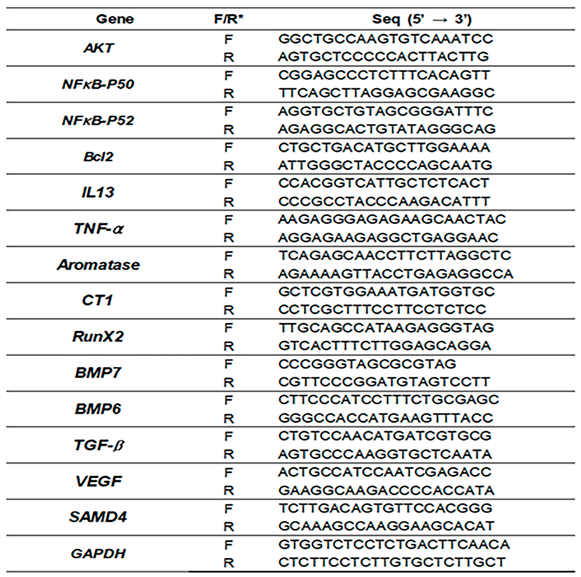
4.4. Quantitative PCR
Total RNA was extracted from cells (fibroblasts and ASCs) using the RiboEx reagent (GeneAll, Seoul, Korea). The RNA was then reverse transcribed into cDNA using a Maxime RT PreMix (iNtRON, Seongnam, Korea), and quantitative PCR was performed using specific primers (Table 1) with the following cycling parameters: 1 min at 95 °C, followed by 35 cycles of 35 s at 59 °C and 35 cycles of 1 min at 72 °C. The expression levels of the target genes in the samples were normalized to those of the housekeeping gene GAPDH, and the relative quantities of the target genes were determined with respect to those of the control.
4.5. Phagocytic activity Test
The cultured macrophages were exposed to the four types of induced exosomes (CE, EIE, FDIE, and EFDIE) for one day, and the exposed cells were treated with FITC-labeled E. coli particles using a Phagocytosis Assay Kit (ab235900, Abcam, Cambridge, UK). The treated cells were analyzed using a flow cytometer (BD FACSCalibur) with the FlowJo 10.7.0 software (BD Biosciences).
4.6. Evaluating of cytokine concentration
After the cultured macrophages were exposed to the four types of induced exosomes (CE, EIE, FDIE, and EFDIE) for one day, their culture media was isolated. Cytokines in the isolated media were evaluated using IL-4 and IL-6 ELISA kits ( ab46058 and ab178013, Abcam) and a microplate reader (AMR-100; Allsheng, Hangzhou, China).
4.7. Alizarin staining
After the cultured macrophages were exposed to the four types of induced exosomes (CE, EIE, FDIE, and EFDIE) for 5 and 15 days, the cultured cells were fixed with 2% paraformaldehyde for 12 h and stained using Alizarin O reagent (Sigma-Aldrich) for 40 min. Stained cells were evaluated using a fluorescence microscope (Eclipse Ts-2; Nikon, Shinagawa, Japan) and imaging software (NIS-elements V5.11 (Nikon).
4.8. Statistical analysis
All experiments were analyzed by one-way analysis of variance (ANOVA) with the post hoc test (Scheffe’s method) using Prism 7 software(GraphPad, San Diego, CA, USA).

Table 1.
The list of primers for qRT-PCR.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: viability result and full gels.
Author Contributions
Conceptualization and methodology: Hyunjung Kim; Writing—original draft preparation, writing—review and editing, supervision: Boyong Kim. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable. This study did not involve humans or vertebral animals.
Informed Consent Statement
Not applicable.
Acknowledgments
This study was supported by EVERBIO.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sahasrabudhe, M. Crismer values and erucic acid contents of rapeseed oils. Journal of the American Oil Chemists' Society 1977, 54, 323–324. [Google Scholar] [CrossRef]
- Goyal, A.; Dubey, N.; Verma, A.; Agrawal, A. Erucic acid: A possible therapeutic agent for neurodegenerative diseases. Current Molecular Medicine 2024. [Google Scholar] [CrossRef] [PubMed]
- Wani, I.A.; ul Ashraf, Z.; Muzzaffar, S. Erucic acid. In Handbook of plant and animal toxins in food; CRC Press: 2022; pp. 169–176.
- Galanty, A.; Grudzińska, M.; Paździora, W.; Paśko, P. Erucic acid—both sides of the story: a concise review on its beneficial and toxic properties. Molecules 2023, 28, 1924. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shang, L.; Deng, S.; Li, P.; Chen, K.; Gao, T.; Zhang, X.; Chen, Z.; Zeng, J. Peroxisomal oxidation of erucic acid suppresses mitochondrial fatty acid oxidation by stimulating malonyl-CoA formation in the rat liver. Journal of Biological Chemistry 2020, 295, 10168–10179. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.J.; Khan, S.A. Chemistry and Biological Activity of Mustard Oil: Therapeutic Benefits and Risk to Healthcare. Revista Brasileira de Farmacognosia 2024, 34, 65–79. [Google Scholar] [CrossRef]
- Wendlinger, C.; Hammann, S.; Vetter, W. Various concentrations of erucic acid in mustard oil and mustard. Food chemistry 2014, 153, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Tkach, M.; Théry, C. Communication by extracellular vesicles: where we are and where we need to go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Song, S.; Chen, N.; Liao, J.; Zeng, L. Stem cell-derived exosomes: A supernova in cosmetic dermatology. Journal of Cosmetic Dermatology 2021, 20, 3812–3817. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, S.; Du, C.; Zhang, Y.; Li, Y.; Chu, L.; Han, X.; Galons, H.; Zhang, Y.; Sun, H. Exosomes from different cells: Characteristics, modifications, and therapeutic applications. European Journal of Medicinal Chemistry 2020, 207, 112784. [Google Scholar] [CrossRef]
- Park, Y.; Lee, K.; Kim, S.W.; Lee, M.W.; Kim, B.; Lee, S.G. Effects of induced exosomes from endometrial cancer cells on tumor activity in the presence of Aurea helianthus extract. Molecules 2021, 26, 2207. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Z.; Shan, D.; Wu, Y.; Zhao, Y.; Li, C.; Shu, Y.; Linghu, X.; Wang, B. Adverse effects of exposure to fine particles and ultrafine particles in the environment on different organs of organisms. Journal of Environmental Sciences 2024, 135, 449–473. [Google Scholar] [CrossRef] [PubMed]
- Combes, A.; Franchineau, G. Fine particle environmental pollution and cardiovascular diseases. Metabolism 2019, 100, 153944. [Google Scholar] [CrossRef]
- Zhang, Z.; Dong, B.; Li, S.; Chen, G.; Yang, Z.; Dong, Y.; Wang, Z.; Ma, J.; Guo, Y. Exposure to ambient particulate matter air pollution, blood pressure and hypertension in children and adolescents: a national cross-sectional study in China. Environment international 2019, 128, 103–108. [Google Scholar] [CrossRef]
- Javadinejad, S.; Dara, R.; Jafary, F. Health impacts of extreme events. Safety in Extreme Environments 2020, 2, 171–181. [Google Scholar] [CrossRef]
- Xu, H.; Jia, Y.; Sun, Z.; Su, J.; Liu, Q.S.; Zhou, Q.; Jiang, G. Environmental pollution, a hidden culprit for health issues. Eco-Environment & Health 2022, 1, 31–45. [Google Scholar]
- Park, Y.; Shin, G.H.; Jin, G.S.; Jin, S.; Kim, B.; Lee, S.G. Effects of black jade on osteogenic differentiation of adipose derived stem cells under benzopyrene. Applied Sciences 2021, 11, 1346. [Google Scholar] [CrossRef]
- Wada, Y.; Ikemoto, T.; Morine, Y.; Imura, S.; Saito, Y.; Yamada, S.; Shimada, M. The differences in the characteristics of insulin-producing cells using human adipose-tissue derived mesenchymal stem cells from subcutaneous and visceral tissues. Scientific Reports 2019, 9, 13204. [Google Scholar] [CrossRef]
- Korzh, M.; Vorontsov, P.; Ashukina, N.; Maltseva, V. Age-related features of bone regeneration (literature review). Orthopaedics, Traumatology and Prosthetics 2021, 3, 92–100. [Google Scholar] [CrossRef]
- Velletri, T.; Xie, N.; Wang, Y.; Huang, Y.; Yang, Q.; Chen, X.; Chen, Q.; Shou, P.; Gan, Y.; Cao, G. P53 functional abnormality in mesenchymal stem cells promotes osteosarcoma development. Cell death & disease 2016, 7, e2015–e2015. [Google Scholar]
- Holleville, N.; Matéos, S.; Bontoux, M.; Bollerot, K.; Monsoro-Burq, A.H. Dlx5 drives Runx2 expression and osteogenic differentiation in developing cranial suture mesenchyme. Developmental biology 2007, 304, 860–874. [Google Scholar] [CrossRef]
- IGF, N.S.; but not Length, F.W. Aromatase Inhibition Causes Lower Bone Density Than Ovariectomy in Mice, an Effect Prevented by Bisphosphonates. W. Kozlow1, K.
- Bonewald, L.F. Cell–cell and cell–matrix interactions in bone. Intercellular Signaling in Development and Disease: Cell Signaling Collection.
- Mandal, C.C. Osteolytic metastasis in breast cancer: effective prevention strategies. Expert Review of Anticancer Therapy 2020, 20, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Huang, J.; Chen, C.; Wang, Y.; Hao, Z.; Chen, T.; Wang, J.; Li, J. Strategies of macrophages to maintain bone homeostasis and promote bone repair: a narrative review. Journal of Functional Biomaterials 2022, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R. Diabetic bone disease. Calcified tissue international 1991, 49, 155–160. [Google Scholar] [CrossRef]
- Harris, P.E.; Bouloux, P.-M.G. Metabolic bone disease. Endocrinology in Clinical Practice, ed. by PE Harris, PM Bouloux. CRC Press 2014, 243. [Google Scholar]
- Ritz, B.; Hoffmann, B.; Peters, A. The effects of fine dust, ozone, and nitrogen dioxide on health. Deutsches Ärzteblatt International 2019, 116, 881. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.E.; Cho, D.; Park, H.J. Air pollution and skin diseases: Adverse effects of airborne particulate matter on various skin diseases. Life sciences 2016, 152, 126–134. [Google Scholar] [CrossRef]
- Lee, C.-W.; Chi, M.-C.; Peng, K.-T.; Chiang, Y.-C.; Hsu, L.-F.; Yan, Y.-L.; Li, H.-Y.; Chen, M.-C.; Lee, I.-T.; Lai, C.-H. Water-soluble fullerenol C60 (OH) 36 toward effective anti-air pollution induced by urban particulate matter in HaCaT cell. International Journal of Molecular Sciences 2019, 20, 4259. [Google Scholar] [CrossRef]
- Kirindage, K.G.I.S.; Jayasinghe, A.M.K.; Cho, N.; Cho, S.H.; Yoo, H.M.; Fernando, I.P.S.; Ahn, G. Fine-dust-induced skin inflammation: Low-molecular-weight fucoidan protects keratinocytes and underlying fibroblasts in an integrated culture model. Marine Drugs 2022, 21, 12. [Google Scholar] [CrossRef]
- Amarasekara, D.S.; Kim, S.; Rho, J. Regulation of osteoblast differentiation by cytokine networks. International journal of molecular sciences 2021, 22, 2851. [Google Scholar] [CrossRef]
- Stein, G.S.; Lian, J.B. Molecular mechanisms mediating proliferation/differentiation interrelationships during progressive development of the osteoblast phenotype. Endocrine reviews 1993, 14, 424–442. [Google Scholar] [CrossRef]
- Pitkänen, S. In vitro and in vivo osteogenesis and vasculogenesis in synthetic bone grafts. 2020.
- Komori, T. Regulation of proliferation, differentiation and functions of osteoblasts by Runx2. International journal of molecular sciences 2019, 20, 1694. [Google Scholar] [CrossRef] [PubMed]
- Loi, F.; Córdova, L.A.; Zhang, R.; Pajarinen, J.; Lin, T.-h.; Goodman, S.B.; Yao, Z. The effects of immunomodulation by macrophage subsets on osteogenesis in vitro. Stem cell research & therapy 2016, 7, 1–11. [Google Scholar]
- Niu, Y.; Wang, Z.; Shi, Y.; Dong, L.; Wang, C. Modulating macrophage activities to promote endogenous bone regeneration: Biological mechanisms and engineering approaches. Bioactive materials 2021, 6, 244–261. [Google Scholar] [CrossRef] [PubMed]
- Meo, S.A.; Rasheed, S.; Khan, M.M.; Shujauddin, S.; Al-Tuwaijri, A.S. Effect of cement dust exposure on phagocytic function of polymorphonuclear neutrophils in cement mill workers. Int J Occup Med Environ Health 2008, 21, 133–139. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Establishment of treatment dosages for fine dust, erucic acid, and induction of exosomes in fibroblasts. Establishing treatment doses of fine dust and erucic acid for fibroblasts (a, b). Purification of exosomes isolated from fibroblasts (c). The results indicate the concentration of exosomes isolated from fibroblasts under condition (d). Con, control; FD, fine dust; E, erucic acid; ns, not significant; (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 1.
Establishment of treatment dosages for fine dust, erucic acid, and induction of exosomes in fibroblasts. Establishing treatment doses of fine dust and erucic acid for fibroblasts (a, b). Purification of exosomes isolated from fibroblasts (c). The results indicate the concentration of exosomes isolated from fibroblasts under condition (d). Con, control; FD, fine dust; E, erucic acid; ns, not significant; (* p < 0.05; ** p < 0.01; *** p < 0.001).
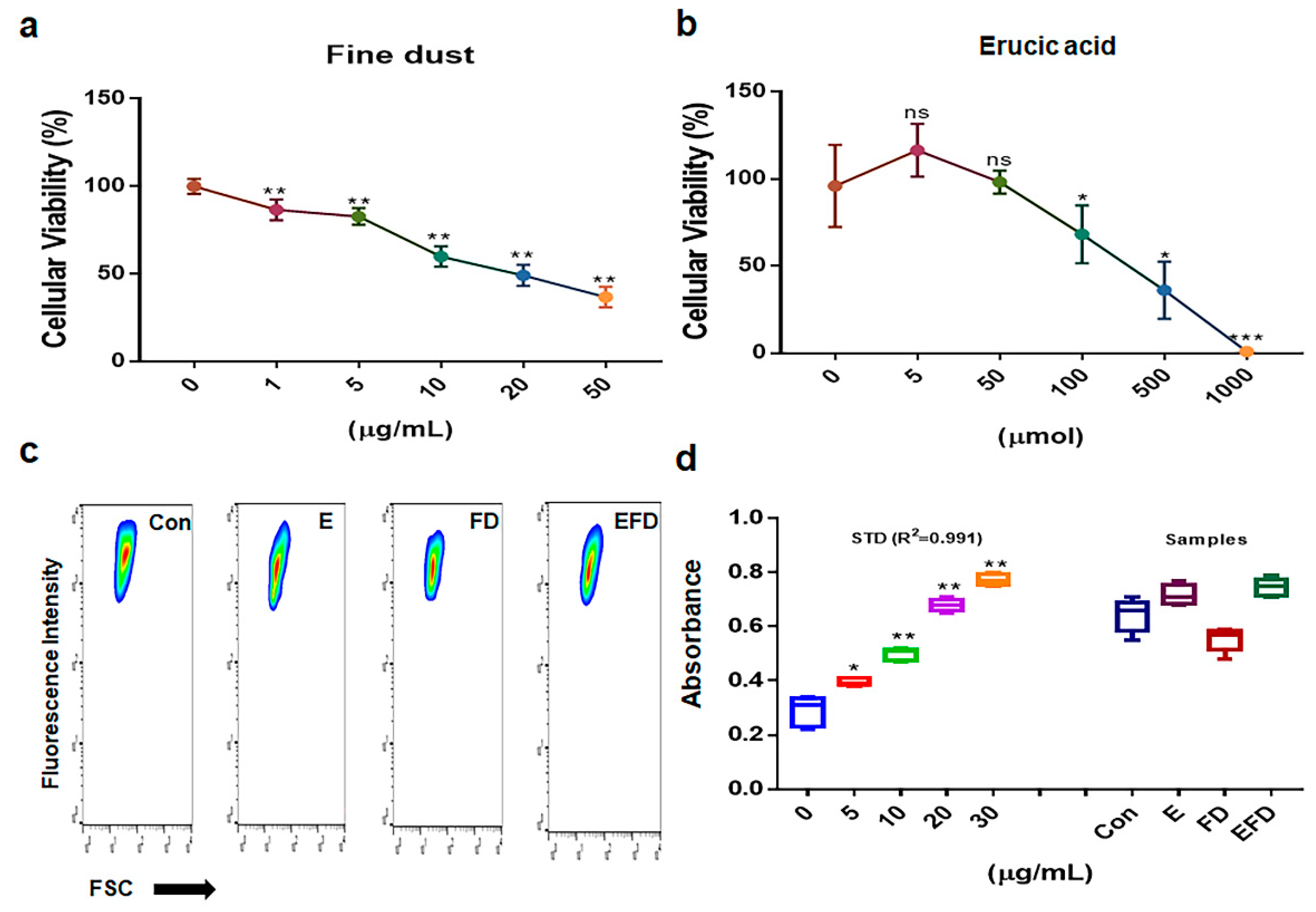
Figure 2.
The levels of osteogenic activators and anti-apoptotic markers in fibroblasts under various exposure conditions. Levels of osteogenic activators VEGF and TGF-β in fibroblasts under different conditions ( control, E, FD, and EFD)(a). Levels of anti-apoptotic markers in fibroblasts under different conditions (b). Con, control; FD, fine dust; E, erucic acid; EFD, exposure to FD after E exposure) ns, not significant; (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 2.
The levels of osteogenic activators and anti-apoptotic markers in fibroblasts under various exposure conditions. Levels of osteogenic activators VEGF and TGF-β in fibroblasts under different conditions ( control, E, FD, and EFD)(a). Levels of anti-apoptotic markers in fibroblasts under different conditions (b). Con, control; FD, fine dust; E, erucic acid; EFD, exposure to FD after E exposure) ns, not significant; (* p < 0.05; ** p < 0.01; *** p < 0.001).
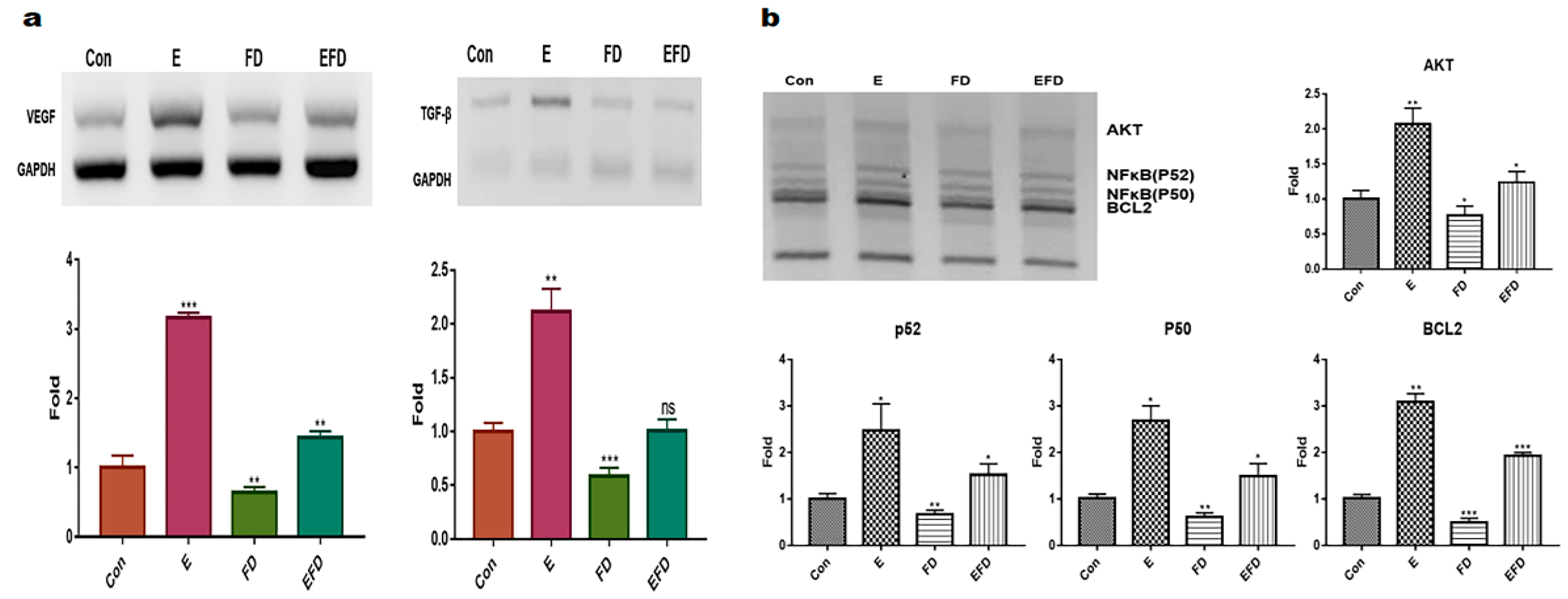
Figure 3.
The levels of osteocyic differentiation markers in adipose-derived stem cells in the induced exosomes isolated from various conditions. The levels of markers for osteocytic differentiation in adipose derived stem cells under various conditions (CE, EIE, FDIE, EFDIE) (a,b,c). The induced exosomes were isolated from fibroblasts under four conditions. CE, control induced exosomes; FDIE, fine dust-induced exosomes; EIE, erucic acid-induced exosomes; EFDIE, FD-induced exosomes after exposure, ns, not significant; (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 3.
The levels of osteocyic differentiation markers in adipose-derived stem cells in the induced exosomes isolated from various conditions. The levels of markers for osteocytic differentiation in adipose derived stem cells under various conditions (CE, EIE, FDIE, EFDIE) (a,b,c). The induced exosomes were isolated from fibroblasts under four conditions. CE, control induced exosomes; FDIE, fine dust-induced exosomes; EIE, erucic acid-induced exosomes; EFDIE, FD-induced exosomes after exposure, ns, not significant; (* p < 0.05; ** p < 0.01; *** p < 0.001).
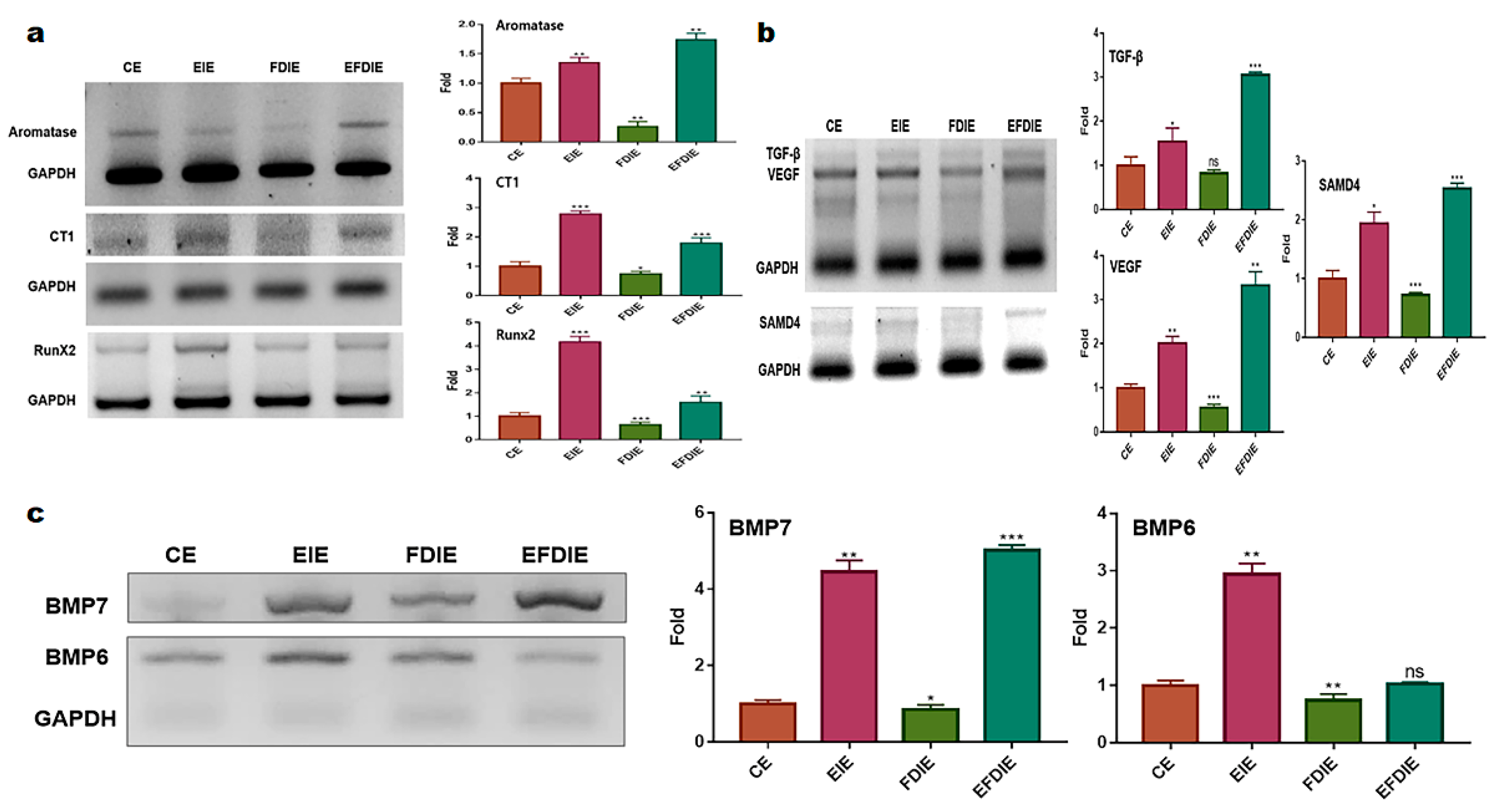
Figure 4.
Osteocytic differentiation in adipose-derived stem cells with the induced exosomes isolated from various conditions. Alizarin staining for osteocytic differentiation in adipose-derived stem cells under various conditions (CE, EIE, FDIE, EFDIE) (a,b,c). The red arrows indicate osteocytic colonies, and the black spots in cells of the stained images are calcium granules. The induced exosomes were isolated from fibroblasts under four conditions: CE, control induced exosomes; FDIE, fine dust-induced exosomes; EIE, erucic acid-induced exosomes; EFDIE, FD-induced exosomes after erucic acid exposure ; (* p < 0.05; *** p < 0.001), (the scale bars = 30μm).
Figure 4.
Osteocytic differentiation in adipose-derived stem cells with the induced exosomes isolated from various conditions. Alizarin staining for osteocytic differentiation in adipose-derived stem cells under various conditions (CE, EIE, FDIE, EFDIE) (a,b,c). The red arrows indicate osteocytic colonies, and the black spots in cells of the stained images are calcium granules. The induced exosomes were isolated from fibroblasts under four conditions: CE, control induced exosomes; FDIE, fine dust-induced exosomes; EIE, erucic acid-induced exosomes; EFDIE, FD-induced exosomes after erucic acid exposure ; (* p < 0.05; *** p < 0.001), (the scale bars = 30μm).
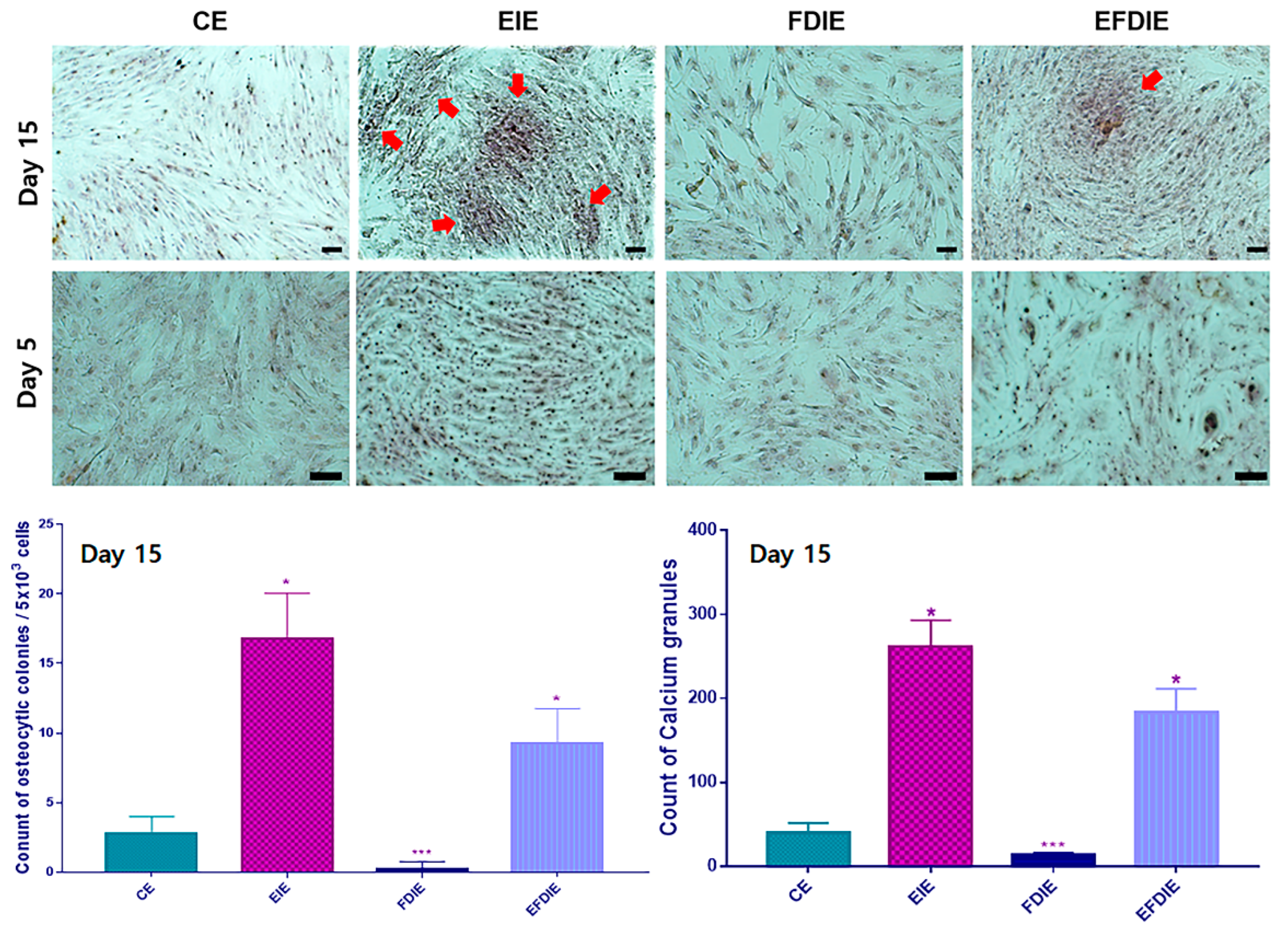
Figure 5.
Phagocytic activity of macrophages with induced exosomes isolated from various conditions. Stagger-offset (a) and overlapping (b) histograms of phagocytosis-positive cells. Dot plot for the phagocytosis-positive cells with the induced exosomes (c) and the results of counting the positive cells compared with the control (d). CE, control-induced exosomes; FDIE, fine dust-induced exosomes; EIE, erucic acid-induced exosomes; EFDIE, FD-induced exosomes after E exposure (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 5.
Phagocytic activity of macrophages with induced exosomes isolated from various conditions. Stagger-offset (a) and overlapping (b) histograms of phagocytosis-positive cells. Dot plot for the phagocytosis-positive cells with the induced exosomes (c) and the results of counting the positive cells compared with the control (d). CE, control-induced exosomes; FDIE, fine dust-induced exosomes; EIE, erucic acid-induced exosomes; EFDIE, FD-induced exosomes after E exposure (* p < 0.05; ** p < 0.01; *** p < 0.001).
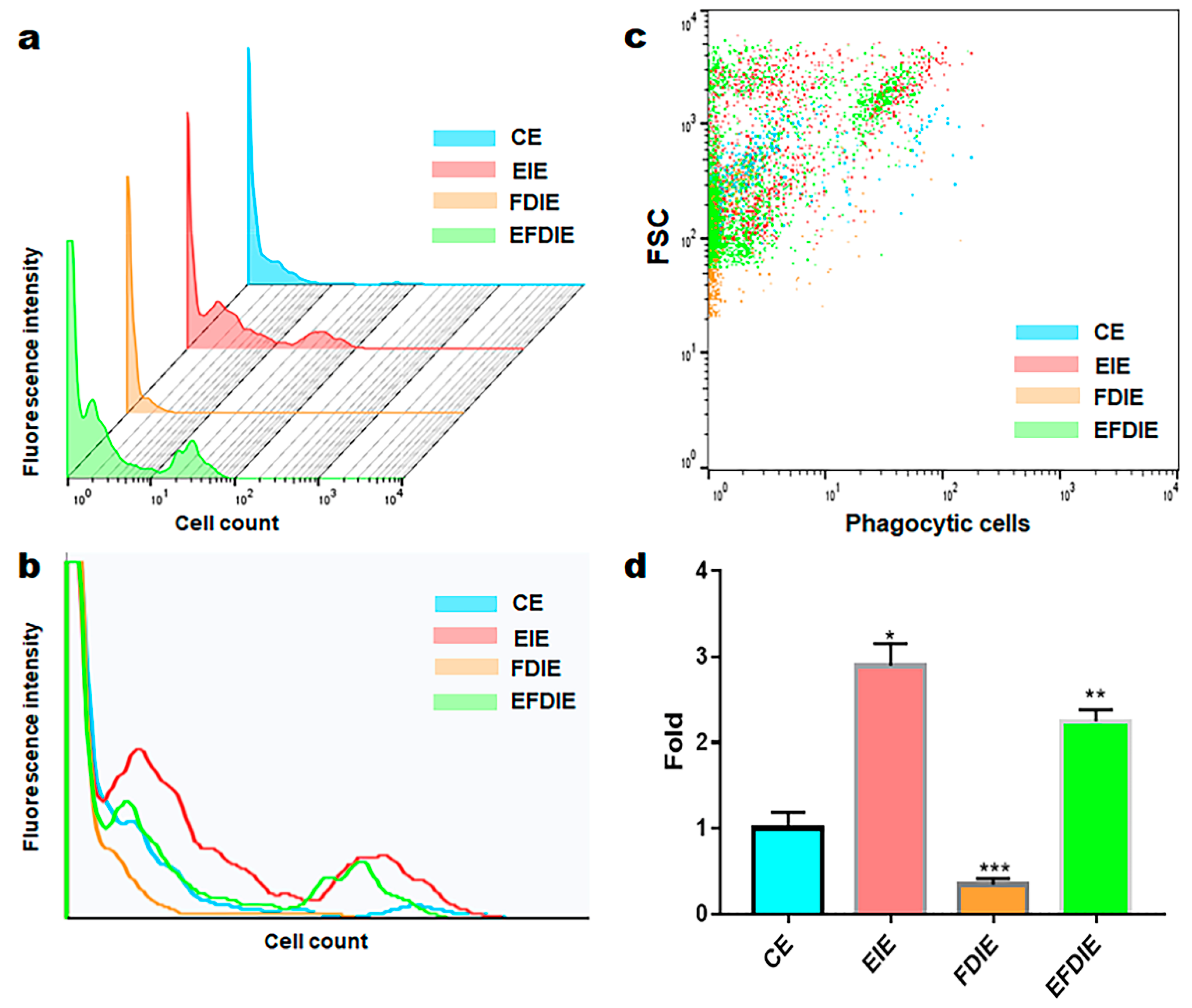
Figure 6.
The levels of the osteogenesis modulating cytokines in macrophages with induced exosomes. Panels (a) and (b) show the PCR and ELISA data, respectively. Cytokine levels in macrophages (a) and secreted cytokines in the supernatants of macrophages under various conditions. CE, control-induced exosomes; FDIE, fine dust-induced exosomes; EIE, erucic acid-induced exosomes; EFDIE, FD-induced exosomes after exposure; (* p < 0.05; ** p < 0.01; *** p < 0.001).
Figure 6.
The levels of the osteogenesis modulating cytokines in macrophages with induced exosomes. Panels (a) and (b) show the PCR and ELISA data, respectively. Cytokine levels in macrophages (a) and secreted cytokines in the supernatants of macrophages under various conditions. CE, control-induced exosomes; FDIE, fine dust-induced exosomes; EIE, erucic acid-induced exosomes; EFDIE, FD-induced exosomes after exposure; (* p < 0.05; ** p < 0.01; *** p < 0.001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



