
Submitted:
11 May 2024
Posted:
14 May 2024
You are already at the latest version
Abstract
Viral contamination in the aquaculture industry is a significant challenge that requires further research. Emerging evidence suggests that long noncoding RNAs (lncRNAs) play a role in regulating various biological processes, including immunity. However, most studies have focused on model organisms, and little is known about non-coding RNAs in the immune response to viral hemorrhagic septicemia (VHS) in rainbow trout. In this study, we investigated the listing of non-coding RNAs (ncRNAs) in rainbow trout challenged with VHS virus. We explored their potential role in defense-like responses based on transcriptomic analysis of the virus-treated and control groups. As a result of this research, 4207 lncRNAs were identified, of which 182 lncRNAs were differentially expressed between treated and untreated samples. Among 7753 lncRNA transcripts, 1920 were novel intergenic (class code “u”), and 3748 transcripts were potentially novel isoform of lncRNAs. Gene ontology analysis revealed biological pathways associated with the stress response and immune signaling. Pairs of genes such as TP63-MSTRG.62, IL10-MSTRG.9783, IL10-MSTRG.9784, TK1-MSTRG.22826, and MSTRG.17881-GLI1 showed cis interactions, while DOCK10-MSTRG.30015, MSTRG.36357-LOC101268921, and MSTRG.34190-ENPP2 showed trans interactions. In this study, the role of lncRNAs in regulating the expression of the target genes was clearly identified. This finding is consistent with the regulatory processes of immune system responses mediated by cytokines, as well as the regulation of oxygen levels in response to viral infection. The information presented here highlights the various regulatory mechanisms of lncRNAs in the expression of immune system genes in fish treated with VHS virus.
Keywords:
lncRNAs
; Oncorhynchus mykiss
; VHSV
; differential expression
; high-throughput sequencing
1. Introduction
Today, according to international organizations such as the FAO, the aquaculture industry is expanding worldwide. This rapid development has had numerous effects on the ecological situation, human health, and the economy of societies. Fish are one of the primary sources of human food globally, and aquaculture plays a crucial role in global food production[1]. Rainbow trout, scientifically known as Oncorhynchus mykiss, is a cold-water fish belonging to the Salmonidae family and the Salmoniformes order [2]. Viral hemorrhagic septicemia virus (VHSV), also known as Egtved disease, is a lethal infectious disease caused by a virus belonging to the Rhabdoviridae family and a new genus of rhabdoviruses called the VHSV virus, which is composed of RNA[3], The disease affects both farmed and wild trout of all ages and is one of the most significant diseases of these fish worldwide [4]. First time in 2013, there was a high incidence of disease recurrence and high mortality reported in rainbow trout farms in several provinces of Iran. The clinical symptoms were similar to sepsis [5].
Long noncoding RNAs are a vital group of biological sequences that play crucial roles in different biological processes. They modify gene expression networks by altering nuclear chromatin formation and modulating mRNA stability, translation, and post-translational modifications in the cytoplasm. Additionally, they can play significant roles in tissue development, signaling, and immunoregulatory functions [6]. Numerous studies have revealed the crucial role of lncRNAs in regulating cellular processes and maintaining homeostasis. In general, lncRNAs are more active in the cell nucleus compared to mRNAs. Their activity varies from one tissue to another, and they exhibit low expression and minimal protection[7]. Following transcription, some lncRNAs interact by forming secondary or tertiary structures with proteins, DNAs, or other RNAs to induce their regulatory functions[8].
Panero et al. (2016) examined the expression of non-coding RNA genes in three strains of rainbow trout after being infected with Flavobacterium psychrophilum. These three lines consist of a sensitive line, a resistant line, and a control line. A pairwise comparison between genetic lines and different infection statuses identified 556 differentially expressed of lncRNAs. Another study investigated the identification and analysis of differentially expressed lncRNAs in tilapia [9]. In total, 72,276 high-confidence lncRNAs were detected. Cho et al. (2017) conducted a comprehensive review of the integrated metabolic and transcriptomic responses to VHS virus in olive flounder fish. In this study, the amplification of viral hemorrhagic septicemia virus (VHSV) was measured continuously at 9, 24, 72, and 120 hours after infection. An increase in necrosis and its progression was observed 72 hours after infection. As a result, 1382 mRNA molecules and 72 metabolites were selected for identifying the mechanism of viral infection. Six KEGG metabolic pathways were identified, including carbohydrate metabolism, amino acid metabolism, lipid metabolism, transport and catabolism, cofactor and vitamin metabolism, and energy metabolism. Interestingly, these pathways are closely linked to immune function. Integrated analysis of the transcriptome and metabolome using the KEGG database revealed four renal responses to viral hemorrhagic septicemia virus (VHSV) infection in olive flounder. Among these responses, mechanisms related to the immune system and active protein synthesis were activated, while ATP synthesis mechanisms and antioxidant system activity were suppressed. The results indicated that inhibiting ATP synthesis and antioxidant systems, such as glutathione and peroxisome signaling, could lead to necrosis, while immune activation could cause inflammation in the kidney tissue of olive flounders infected with VHSV [10]. In another study, Wang et al. (2016) identified and evaluated the function of long noncoding intergenic RNAs (lincRNAs) in rainbow trout (Oncorhynchus mykiss)[11]. This report provides the first reference catalog of 9674 rainbow trout lincRNAs, which were identified through the analysis of RNA-seq data from 15 tissues.
Fish skin is an essential immune organ that serves as the first line of defense against the entry of foreign pathogens. In a study, researchers investigated the differences in innate immunity between the wild type of rainbow trout and its yellow mutant type by analyzing the complete transcriptome of the skin. A total of 2448 mRNAs, 1630 lncRNAs, 22 circRNAs and 50 miRNAs were differentially expressed. Among the expressed mRNAs, several key immune-related genes, including IFIH1, DHX58, TRIM25, ATP6V1E1, TAP1, TAP2, CD209, HSP90A.1, NLRP3, NLRC3, and several other metabolism-related genes (GSTP3, NAPRT), were identified. Gene ontology and database (KEGG) pathways related to innate immunity, including NAD+ ADP-ribosyltransferase activity, immune response, response to bacterial infection, and RIG-I receptor signaling, such as NOD-like receptor signaling and phagosome KEGG pathways, were identified. In addition, the role of long non-coding RNAs (lncRNAs) MSTRG.11484.2, MSTRG.32014.1, and MSTRG.29012.1 in the regulation of at least three immune-related genes (IFIH1, DHIRF) was determined. [12]. In another study, liver tissues from rainbow trout were sampled and analyzed using RNA-seq technology under heat stress (24 °C) and control conditions (18 °C). A total of 927 new lncRNAswere identified, and 428 differentially expressed lncRNAs were identified. GO and KEGG enrichment analyses revealed that numerous target genes associated with maintaining homeostasis and adapting to stress, including heat stress conditions, were expressed. Several regulatory pathways, including the thyroid hormone signaling pathway, the PI3K-Akt signaling pathway, and the estrogen signaling pathway, have also been implicated in heat stress. [13].
Due to the lack of information about the role of lncRNAs in regulating cellular processes under VHS infection, the aim of the present study was to investigate the regulatory effects of lncRNAs on differential gene expression in the spleen tissue of rainbow trout infected with VHSV.
2. Materials and Methods
2.1. Sample Collection and Library Construction
In the current study, the rainbow trout fish datasets from Ghaderzade et al. (2021) [14] were utilized to identify lncRNAs. According to this study, the procedures for sample collection and RNA library preparation are summarized as follows:
At the start of the experiment, 310 rainbow trout were kept for an 18-day acclimatization period under identical condition. Then, they were divided into four treatment groups and received doses of 0.06 and 0.03 cc of live viral hemorrhagic septicemia virus, as well as the same dosage of physiological serum, respectively. The virus concentration in the live virus injection group was measured at 4.5 x 104 PFU/ml. After 14 days post-injection, spleen tissues were collected from 30 fish in the 0.06 virus and 0.06 physiological serum treated groups. From each sample group, 3 replicates were prepared, resulting in 10 pooled samples from each group. Total RNA was extracted from spleen tissue using the standard TRIzol protocol. The extracted RNA was treated with DNase I enzyme at 37°C for 30 minutes. Subsequently, the quantity and quality of the extracted RNA were assessed using 1% agarose gel electrophoresis and Nanodrop spectrophotometry. The cDNA libraries were prepared using the TruSeq™ library preparation kit following the instructions provided by Illumina. RNA purification, cDNA library construction, and RNA sequencing were performed by BGI Company. Each library was paired-end sequenced (150 nt) on the Illumina HiSeq 2500 platform.
2.2. Data Analysis
2.2.1. Read Alignment and Transcript Assembly
The workflow of the bioinformatics analysis pipeline for this study is illustrated in Figure 1. The sequencing quality of the raw data was assessed using FastQC (version 0.11.5) software. Low-quality reads and adapter sequences were then eliminated using Trimmomatic software (version 0.32)[15]. Next, the reads with acceptable quality were aligned to the rainbow trout fish reference genome (GCF_013265735.2) using the HISAT2 aligner (version 2.2.0). The mapped reads of each sample were assembled separately using StringTie (v2.1.4)[16]. Then, all annotation (GTF) files created for six libraries were merged into a single unified transcriptome file using the 'StringTie-merge' option. Gffcompare (v0.9.8) was used to compare the unified transcripts with the reference genome (22).
2.2.2. Transcript Quantification and Differential Expression Analysis
The number of reads per gene in each sample was calculated using the StringTie -e option. Then, a Python script was utilized to generate a gene expression (read count) matrix for input into DESeq2. The analysis of differential gene expression was conducted using Deseq2 [17] based on normalized FPKM values. Genes with P-adjust < 0.05 and FC > 1 were considered to exhibit significant differential expression. Principal component analysis (PCA) was utilized to extract the primary features of the data, serving as indicators of the overall data state. The results were visualized using the ggplot2 package in R (version 4.2.2).
2.2.3. Identification and Classification of Putative lncRNAs
The assembled transcriptome was screened by StringTie to identify potential long non-coding RNAs (lncRNAs). Several filtration steps were carried out. First, transcripts shorter than 200 bp were discarded. Secondly, single-exon transcripts were removed. Third, transcripts with the class codes "u-j-o-i-u-x" were selected from the transcriptome file. Finally, to improve prediction accuracy, four different software programs with different algorithms, including CPC2 (http://cpc2.cbi.pku.edu.cn/)[18]; RNAsamba 0.2.5 [19], LncADeep1.0 [20], and Pleak.1.2 [21], were used to classify coding and noncoding transcripts. Transcripts that were identified by all four software programs were considered as lncRNAs and used in the subsequent steps of analysis.
2.2.4. Target Gene Prediction and PPI Network
To begin grasping the role of lncRNAs, they are commonly categorized into two groups: Cis-acting and trans-acting, depending on whether they regulate gene expression locally or distantly [22, 23]. It has been proposed that the capacity of lncRNAs to control nearby gene expression in cis could be due to the mature lncRNA transcript or the independence of lncRNAs themselves in carrying out regulatory functions [24]. DElncRNAs were identified by comparing the detected lncRNAs with the Deseq2 DE outputs and were distinguished from DEmRNAs. Prediction of cis-acting target genes was conducted using the sliding window strategy, and DEmRNAs within 5 Kbp upstream and downstream of neighboring DElncRNAs were identified using Bedtools software [25].
In the case of trans-acting genes, lncRNAs have been shown to interact with proteins and/or other RNA molecules in order to control the expression of specific target genes. [24]. For the prediction of trans-acting target genes, the RIBLAST[26] software was utilized with an interacting energy cutoff of -20 kcal/mol. Paired genes with the minimum interacting energy were filtered using a cutoff correlation rate of +95% and -95%. Additionally, Pearson correlation coefficients were calculated to assess the association between the expression profiles of DElncRNAs and DEmRNAs. A Pearson correlation coefficient cutoff of > ± 0.95 was considered a significant indicator. Finally, the trans-acting target genes were identified by combining the results of RIBLAST software and calculating the Pearson correlation coefficient.
Additionally, the coding genes targeted by trans and cis were analyzed for protein-protein interactions (PPI) using the geneMANIA plugin within cytoscape to explore how these genes interact within PPI networks. The CytoHubba plugin [27] in Cytoscape was utilized to identify the top 5 modules within the combined network by employing various topological algorithms. The integrated network was visualized using Cytoscape software version 3.10.2.
2.2.5. Gene Ontology and Networking Analysis
In this research, target mRNA genes involved in cis and trans interactions with lncRNAs were identified using the DAVID database to explore potential ontologies. Additionally, gene networking was visualized using zebrafish (Danio rerio) annotation information by Cytoscape software [28].
2.2.6. Novel lncRNAs and QTL Analysis
Given the limited number of all quantitative trait loci (QTLs) identified in rainbow trout, a study was conducted to locate expressed lncRNAs within all QTLs. The research involved analyzing the co-localization of relevant genes. Initially, QTL data was sourced from AnimalQTLdb [29], and then the differentially expressed lncRNAs were compared to those of the QTLs. Any newly discovered lncRNA genes with start and end positions falling within the QTL regions were deemed successfully annotated within the QTL.
2.2.7. Q-RT-PCR Validation
Real-time PCR was employed to validate the differential expression results of genes obtained from RNA-seq data for both mRNAs and lncRNAs. Due to reduced RNA quality, the results were compared with qPCR data from a previous experiment [14]. A DNAase- and RNAse-free kit from Cinaclon company was used to purify samples. The kit contained 50 mM EDTA reaction buffer, 2 mM MgCl, DNAseI, and RNAse-free components. RNA samples were extracted and used for cDNA synthesis using a kit from Thermo Fisher Scientific, following the manufacturer's instructions. The cDNA synthesis process involves several steps, including incubation, addition of reagents, and temperature adjustments. To compare Q-RT-PCR with RNA-Seq results, the mean values for each gene were converted into fold changes, and primers were designed using Oligo version 7 software.
3. Results
3.1. RNA-Seq Data Analysis
To identify the lncRNAs responsive to viral infection, six RNA-seq libraries from VHSV-infected rainbow trout were utilized. After filtering the raw data, the number of clean reads ranged from 40,035,219 to 53,829,279 for the six samples. These reads had a Q30 percentage value (where Q30 indicates that the probability of an incorrect base call is 1 in 1,000) ranging from 91.07 to 92.25. This indicates that the quality of reads from each library was reliable for the subsequent analysis steps. Then, the clean reads were mapped to the rainbow trout reference genome, and the mapping ratio was estimated to be higher than 88.89% for each library (Table 1). Finally, the StrigTie software assembled a total of 78,380 genes (166,522 transcripts).
3.2. Identification and Classification of Putative lncRNAs
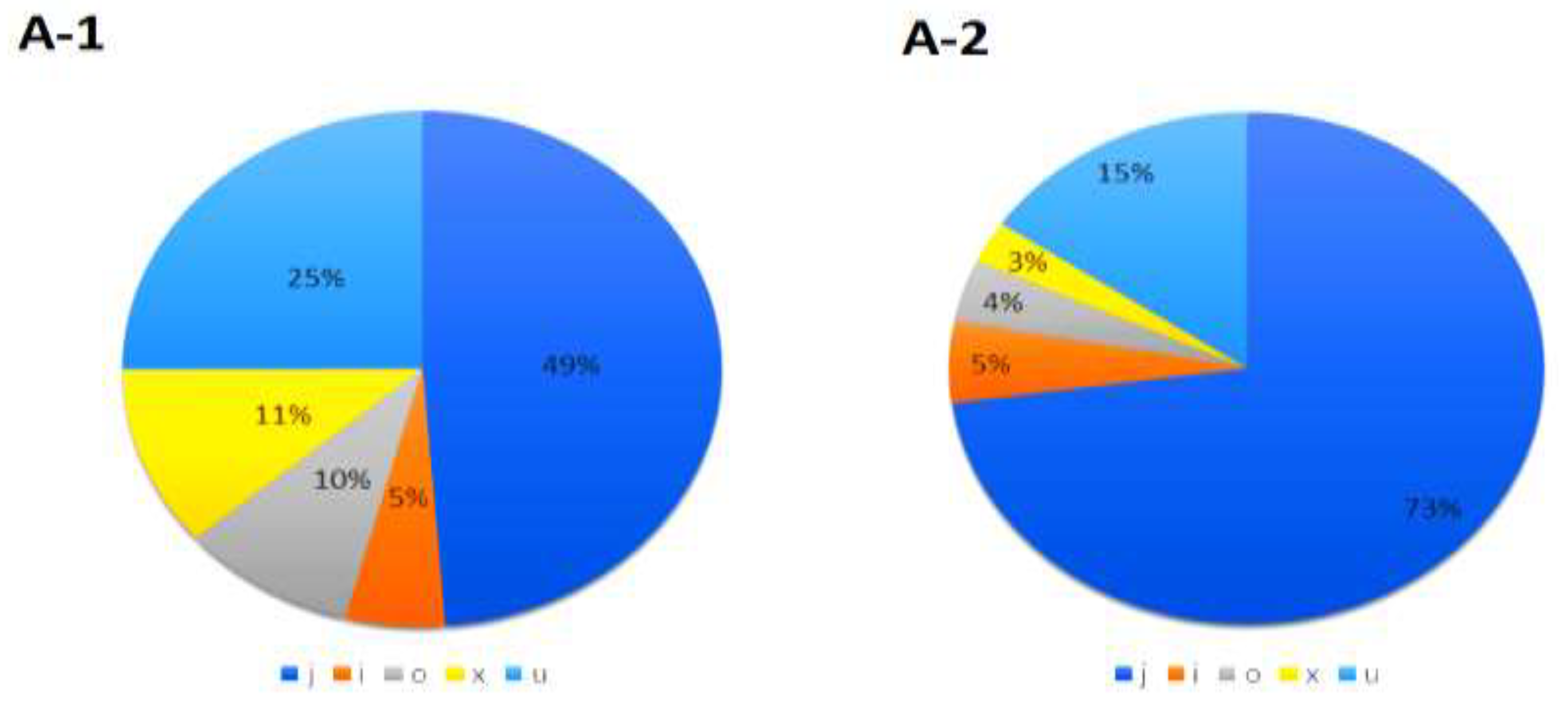
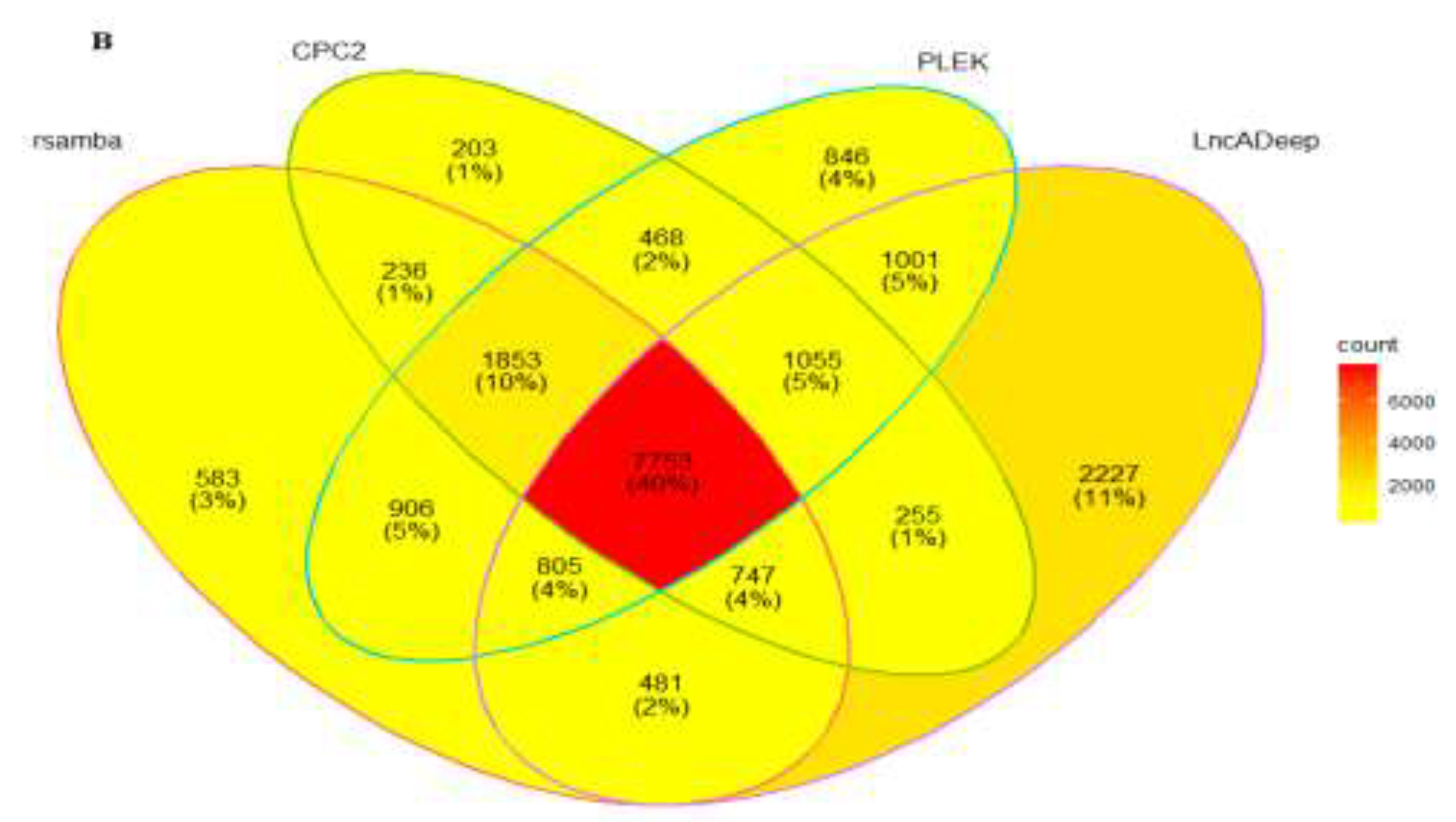
All coding and noncoding genes were analyzed using the same pipeline outlined in Figure 1. We utilized four types of prediction software to identify transcripts without coding potential or lncRNAs. The results revealed that CPC2, plek, lncADeep, and RNAsamba identified 14,687, 14,324, 13,363, and 12,570 transcripts as potential lncRNAs, respectively. Finally, 7753 transcripts (4207 genes) were identified as the final set of lncRNAs using the mathematical concept of intersection sets. The results of the analysis based on RNA-Seq data revealed that the identified lncRNAs were classified into five class codes based on their genomic location and direction of transcription. In this study, various types of long non-coding RNAs (lncRNAs) were identified, including 25% intergenic novel lncRNAs, 11% antisense lncRNAs, 5% intronic lncRNAs, 10% exonic overlap lncRNAs with reference transcripts, and 49% potentially novel isoform lncRNAs. The majority of the identified lncRNAs were classified as potentially novel isoform lncRNAs. Additionally, three out of four differentially expressed lncRNAs were also potentially novel isoforms.
Figure 2.
(A-1) Percentage of various class codes in all identified lncRNAs (7753 transcripts). (A-2) Distribution of various class codes in differentially expressed lncRNAs (308 transcripts). (B) The Venn diagram shows the overlapping noncoding genes and the percentage of their intersection.
Figure 2.
(A-1) Percentage of various class codes in all identified lncRNAs (7753 transcripts). (A-2) Distribution of various class codes in differentially expressed lncRNAs (308 transcripts). (B) The Venn diagram shows the overlapping noncoding genes and the percentage of their intersection.
3.3. Differentially Expressed lncRNAs
Differential expression analysis revealed that 182 lncRNA had significantly different expression in the virus-infected samples compared to the control group, meeting the criteria of |log2FoldChange|> 1 and FDR<0.05. of the 182 DElncRNAs, 73 were up-regulated and 109 were down-regulated (Figure 3, B). Three lncRNAs of MSTRG.33904, MSTRG.13838, and MSTRG.27745, exhibited maximum up-regulation of 25.8 fc and 1.48E-08 padj, 13.8 fc and 0.012 padj, and 12.7 fc and 0.026 padj, respectively.
The lncRNAs MSTRG.5754, MSTRG.33167, and MSTRG.35881 exhibited a maximum down-regulation of -27.1-fold change (fc) with a p-value of 1.78E-09, -20.8 fc with a p-value of 3.04E-06, and -15.01305686 fc with a p-value of 5.23E-32, respectively. Hierarchical clustering with a heatmap and PCA showed that the replicates of each condition clustered correctly (see figure 3, D). The average log2-FPKM of lncRNAs and mRNAs was estimated to be 3.42 and 3.75, respectively, in the control samples. Additionally, the average log2 FPKM values of long non-coding RNAs (lncRNAs) and messenger RNAs (mRNAs) in the virus-infected samples were 3.6 and 3.8, respectively, while those in the control samples were 3.2 and 3.6.
3.4. Target Prediction of the Candidate lncRNAs and Gene Networking
To understand how DElncRNAs regulate gene expression, we studied how changes in lncRNA levels affect nearby (cis) or distant (trans) mRNA expression. By analyzing the correlation between lncRNA:mRNA pairs within 5 kb regions, we found 53 DEmRNAs and 46 DElncRNAs that form cis-targeting pairs (see Table 5 in the supplementary material). For trans-acting genes, 38 DEmRNAs were influenced by 11 DElncRNAs.
3.5. LncRNAs Target Gene Networking
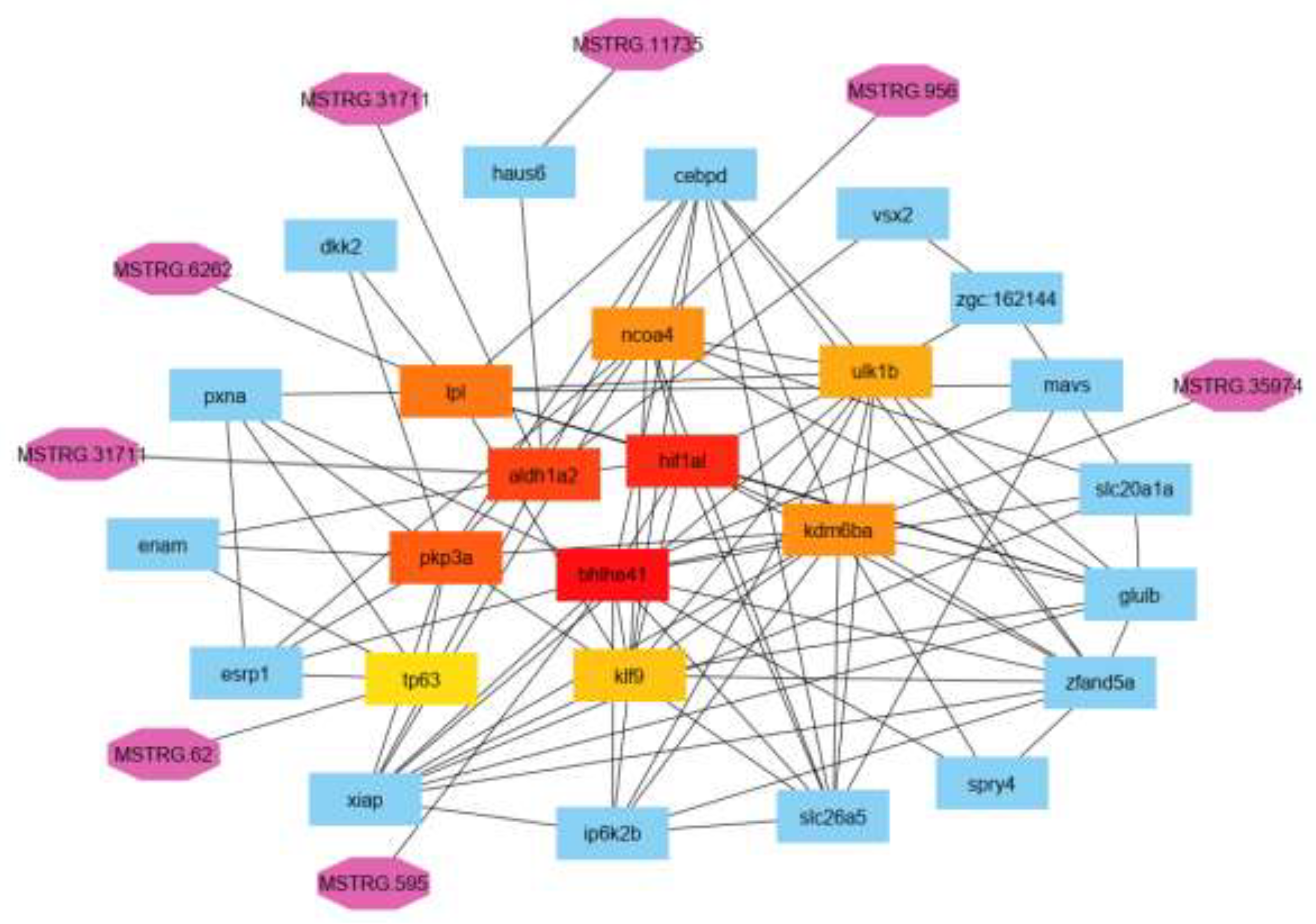
Depending on lncRNAs location and interactions with DNA, RNA, and proteins, they can influence chromatin function, regulate the formation and function of nuclear bodies without membranes, affect the stability and translation of mRNAs in the cytoplasm, and interfere with signaling pathways. These various roles can impact gene expression in a wide range of biological and disease-related scenarios, such as neuronal disorders, immune responses, or other crucial developmental processes. The fact that lncRNAs exhibit specific expression patterns in different tissues and under different conditions suggests that they could serve as potential biomarkers and may be targeted for clinical purposes [30]. To achieve that, all protein-coding genes regulated by lncRNAs were used to construct a protein-protein interaction network (see Figure 5).
Figure 4.
annotated paired mRNAs targeted with the lncRNA gene network.
3.6. Validation of RNA-Seq by qRT‒PCR
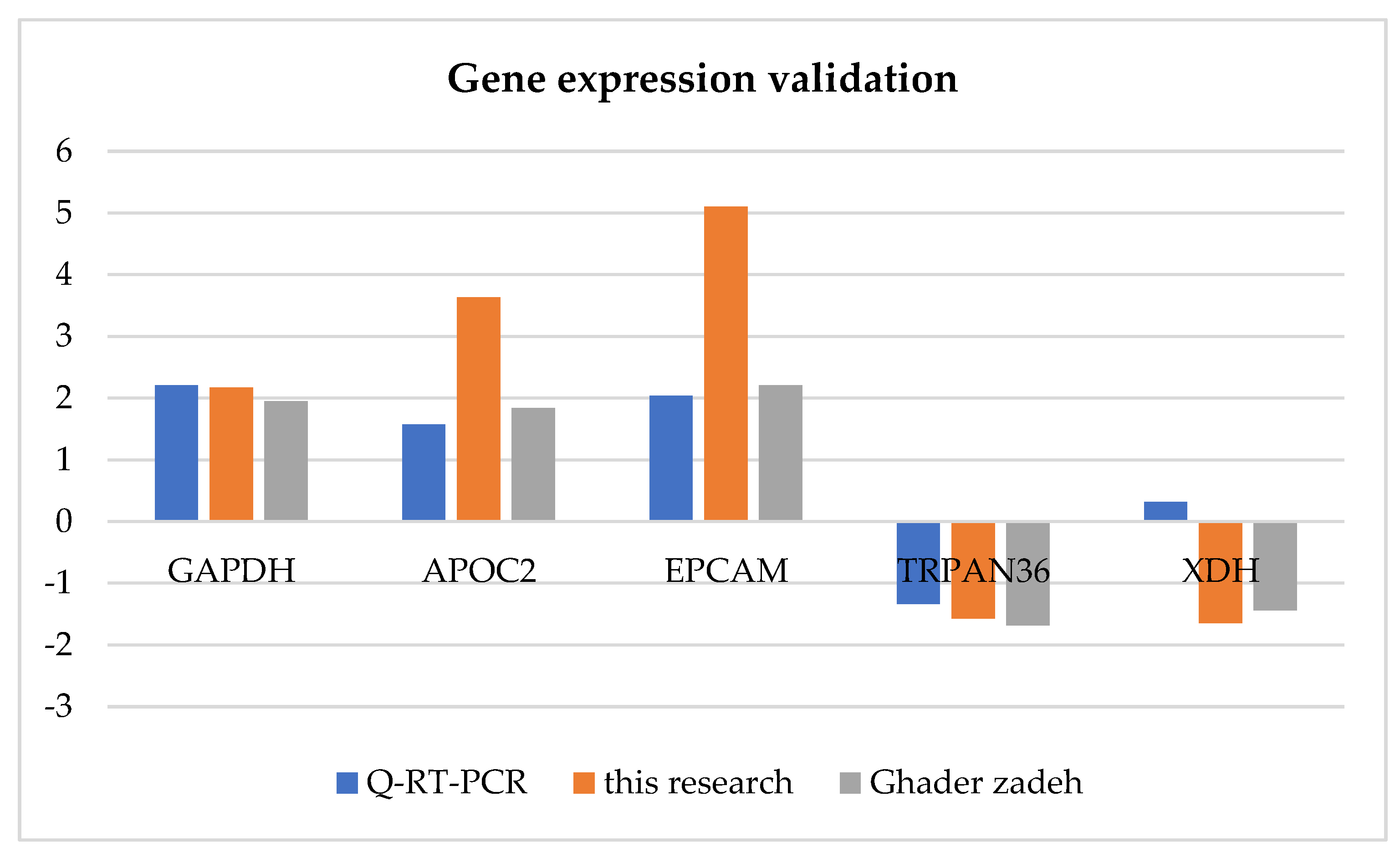
To validate the expression patterns of the mRNA determined by RNA sequencing, qRT-PCR was performed on four randomly selected mRNA transcripts in previously study [15]. As depicted in Figure 5, the results indicated that the expression patterns of all mRNA transcripts were in line with the RNA-seq data. These findings indicate that in most cases, the qRT-PCR results were consistent with those of RNA-Seq, suggesting that the identification pipeline is reproducible and reliable.
Figure 5.
Q-RT-PCR validation of some of the mRNA transcripts. Here, fold change indicates the ratio of average expression of mRNAs.
Figure 5.
Q-RT-PCR validation of some of the mRNA transcripts. Here, fold change indicates the ratio of average expression of mRNAs.
3.7. QTL Analysis
To identify the potential lncRNAs involved in viral response, coincidence of the expressed lncRNAs and all QTLs were investigated. For doing that, 2,329 QTLs including VHSVDD (157282) identified in 23 different publications were used from the rain bow trout QTL database Release #52 (Dec 27, 2023).
Due to the extensive spans of rainbow trout QTL regions, lncRNAs found within a 1 Mb range of the QTL were given additional consideration (500,000 kb upstream and downstream). Out of 182 expressed lncRNAs, MSTRG.26178 (downregulated) were successfully mapped to VHSVDD (157282) on chromosome 13. Seven lncRNAs, including MSTRG.18466, MSTRG.18477, MSTRG.18466, MSTRG.18477, MSTRG.23083, MSTRG.30905, and MSTRG.30997, were located in different QTL numbers (259577, 259578, 259578, 259723, 259605, 259606, 259608, 259609, 259611, 259612, 259613, 259614, 259615, 259616, 259617, 259837) on chromosome 13, associated with survival time and survival rate in bacterial cold-water disease.
4. Discussion
Due to the key roles of lncRNAs in many important biological processes they are currently of particular interest. The rapid development of high throughput sequencing methods has led to the discovery of thousands of lncRNAs in recent years. Rainbow trout, a commonly cultivated species in the aquaculture industry, are vulnerable to various diseases that can have a significant impact on their growth and survival [31]. Genetic and animal breeding of the immune system in rainbow trout are crucial aspects of aquaculture management. A robust immune system is essential for preventing disease outbreaks in aquaculture facilities and decreasing the dependence on antibiotics and other chemical treatments. This can be accomplished by measuring various immunological parameters, such as antibody production, phagocytic activity, cytokine expression, and complement activity [32]. While many lncRNAs have been identified in different organisms such as human and mouse, the systematic prediction and regulatory roles of lncRNAs has been poorly studied in aquatic species, particularly in rainbow trout. For example, the NONCODE database is an integrated knowledge database dedicated to ncRNAs in 38 species, including animals and plants. However, information on rainbow trout fish lncRNAs in this database is lacking. In pursuit of this objective, this research has contributed to identifying and utilizing lncRNA genes or markers to enhance genetic selection in this field. One example of successful disease challenge testing in rainbow trout is the selection for resistance to infectious hematopoietic necrosis virus (IHNV). IHNV is a highly contagious virus that causes severe mortality in salmonid species, including rainbow trout. By exposing fish to Infectious Hematopoietic Necrosis Virus (IHNV) under controlled conditions, researchers have identified individuals with enhanced resistance to the virus and incorporated them into breeding programs to develop lines of IHNV-resistant rainbow trout [33]. For example, researchers have identified certain genes that are associated with enhanced immune responses in rainbow trout. By selectively breeding individuals with these genes, breeders can produce offspring with improved immunity. This approach has been successful in other species such as chickens and pigs, where genetic selection for disease resistance has led to significant improvements in health and productivity.
When comparing FPKM-normalized expression values between DElncRNAs and DEmRNAs, as previously reported by [34], it was observed that the expression levels of lncRNAs were lower than those of mRNAs in both conditions.
It is now well-established that lncRNAs can regulate neighboring gene expression patterns by influencing RNA polymerase II activity [35] or by facilitating the phosphorylation of transcription factors [36]. They exhibit stage-selective regulation of lncRNAs and transcription factors (TFs) by modulating their functional activities in development of ovarian cancer [37]. There are some important genes in cis-acting which have previously been examined for their key role in biological processes. For example, LOC110527687 gene is located on chromosome 1 and is near the MSTRG.982 lncRNA on the same chromosome. LOC110527687 gene in rainbow trout is annotated as a Synuclein gene, with its most important functions including vesicle transport in neurons, transcriptional regulation, modulation of immune cell maturation and response, and a role as an antimicrobial peptide [38]. It has been confirmed that this gene plays a crucial role in viral infections in SARS-CoV-2[39]. Furthermore, upregulation of this gene has been observed in Parkinson's disease, similar to the present results [40]. Cellular alpha-synuclein levels have been shown to increase during certain viral and bacterial infections, and alpha-synuclein can influence immune cell function, suggesting that it may be important in pathogen-activated immune responses [41]. In addition, a recent study showed that α-synuclein is essential for the proper function of immune cells following pathogen invasion and lymphocyte maturation [42]. Previous studies have confirmed the role of the TP63, IL10, and LOC110502310 (suppressor of tumorigenicity 7 protein homolog) genes in responding to biological stress. The DOCK10 gene exhibited increased expression and had a cis-regulatory effect on the downregulation of the lncRNA MSTRG.30015. Docking cytokinesis (DOCK) proteins belong to a family of guanine nucleotide exchange factors (GEFs) for the Rho family of GTPases [43]. Our findings suggest that the expression of these genes is regulated through the cis interaction of MSTRG.62, MSTRG.9783, MSTRG.9784, and MSTRG.2952 lncRNAs. Therefore, these lncRNAs can play important roles in the biological responses of rainbow trout.
Gene networking from paired mRNAs with lncRNAs represents some modules. In rank one genes, BHLHE41 gene is co-expressed and interacts with 10 coding genes and MSTRG.595 lncRNA. This gene is upregulated just like the behavior that showed under cold stress in zebra fish [44]. Hypoxia-inducible factor 1 (HIF-1) is a transcription factor that is essential in responding to low oxygen levels in cells. When oxygen levels are normal, the activity of HIF-1 is controlled by breaking down its alpha subunit (HIF-1α), preventing the formation of an active complex in the nucleus that controls the expression of many genes. Significantly, HIF-1 is involved in cancer and infectious diseases, presenting potential new targets for treatment of these conditions [45]. ALDH1A2 is crucial for converting vitamin A into retinoic acid and plays a vital role in embryonic development, as shown in studies on model organisms. However, only a small number of genetic mutations in ALDH1A2 have been associated with birth defects in humans [46].
An analysis of binding energy and Pearson correlation was conducted to identify the trans-interaction between DElncRNAs and DEmRNAs, revealing that 93 mRNAs and 19 lncRNAs exhibit trans-acting interactions (see Table 6 in the Supplementary File). MSTG.982 gene in chromosome 1 downregulation had a trans action with the LOC110521060 gene, in chromosome 4, which is annotated as an inverted formin-2 (INF2) and down-regulated (log2fc= -1.33). The expression of INF2 increased rapidly when the cells were exposed to H2O2. This upregulation promoted HaCaT cell survival by reducing H2O2-mediated cell apoptosis. Additionally, the deletion of INF2 attenuated mitochondrial reactive oxygen species (ROS) overload, restored the cellular redox balance, sustained the mitochondrial membrane potential, improved mitochondrial respiratory function, and corrected mitochondrial dynamics disorder in an H2O2-mimicking oxidative stress microenvironment [47]. Also, this LncRNA had a trans action with the LOC110513729 gene, in chromosome 9, encoding the fibrinogen alpha chain and exhibited up-regulation (+4.82). Fibrinogen is a vital protein involved in platelet aggregation, essential for blood coagulation, and it also plays a role in the immune system of fish. The expression of Atlantic salmon (Salmo salar) infected with the bacterium Aeromonas salmonicida was up-regulated [48]. Fibrinogen is a crucial protein involved in platelet aggregation, essential for blood coagulation, and it also contributes to the immune system of fish. The expression of Atlantic salmon (Salmo salar) infected with the bacterium Aeromonas salmonicida was up-regulated and also had effects on the pathogenesis of endometriosis [49] just like the present study. LOC110534814, in chromosome 10, is annotated as the small subunit 4 (SLC43A2) of the large neutral amino acid transporter and is upregulated by 7.77-fold. Tumor cells outcompete T cells for methionine by overexpressing SLC43A2, which leads to T-cell exhaustion. SLC43A2 is positively correlated with markers of immune exhaustion. SLC43A2 may contribute to the development of a suppressive tumor microenvironment and regulate immune-related genes, thereby influencing the prognosis of liver hepatocellular carcinoma [50]. This gene trans-acted with the MSTRG.982 as well. The LOC118948728 gene was annotated as the RBP47 gene. Gene pairs, such as MSTRG.36357-LOC110534579, MSTRG.36357-PRLH2, MSTRG.36357-LOC101268921, MSTRG.36357-FAM78AB, MSTRG.36357-LOC110510746, MSTRG.36357-LOC110485438, and MSTRG.36357-SI:CH211-214J24.15 (GTPase IMAP family member 8 isoform X1 [Oncorhynchus keta]), were identified in trans position. The gene LOC118942903 is annotated as the Ig kappa-b4 chain C region. This gene is a significant quantitative trait locus (QTL) for resistance to Vibrio anguillarum in rainbow trout (66). This gene is a target gene for the lncRNAs MSTRG.27793, MSTRG.27770, and MSTRG.20675 in cis in action. The function of PRLH2, in chromosome 15, is to release prolactin hormone 2, which has been proven to be involved in inflammatory processes [51] which were in trans action with MSTRG.24034 in chromosome 16. The LOC101268921 gene, also known as mothers against decapentaplegic homolog 4 or SMAD4, plays a crucial role in the canonical TGF-β signaling pathway in humans. Its expression level is indicative of inactivation in various cancers [52], and is significantly associated with tumor development and prognosis in cancer patients [53, 54]. LOC110510746 is annotated as LIM domain-binding protein,1 in chromosome 17, upregulated had trans action with downregulated MSTRG.24034 gene in chromosome number 12. Viruses have developed various strategies to hijack the cellular gene expression system to facilitate their replication. Poly(A) binding proteins (PABPs), which are a vital family of gene expression factors, are frequently targeted by viruses. PABPs act not only as translation factors but also as key factors in mRNA metabolism [32].
Several studies have shown that cytokines play a crucial role in various inflammatory diseases. They can be used as biomarkers to identify the disease or track its progression. They may also serve as clinically useful indicators for treatments [55]. Researchers evaluated the significant role of long noncoding RNAs in the immunotoxicity induced by BPA in adrenal gland macrophages of common carp (Cyprinus carpio) using sequencing. KEGG analysis has revealed that exposure to BPA severely disrupts several immune-related signaling pathways, including the NF-kappa B, Toll-like receptor, B-cell receptor, Jak-STAT, and Hippo signaling pathways [56]. In our research, we also found that the PPAR signaling pathway, cell adhesion molecules, and Toll-like receptor signaling pathway play significant roles. In a study, a comprehensive transcriptome analysis was conducted on fish skin to investigate the inherent differences in immunity between the wild type of rainbow trout and its yellow mutant type. Several important immune-related genes were identified, including IFIH1, DHX58, TRIM25, ATP6V1E1, TAP1, TAP2, CD209, HSP90A.1, NLRP3, NLRC3, and various other metabolism-related genes. The gene ontology and databank (KEGG) pathways related to innate immunity, such as NAD+ADP-ribosyl transferase activity, immune response to bacterial infection, and RIG-I receptor signaling, including NOD-like receptor and phagosomal KEGG pathways, are presented. In addition, we identified the role of the lncRNAs MSTRG.11484.2, MSTRG.32014.1, and MSTRG.29012.1 in regulating at least two immune-related genes (IFIH1, DHIRF). Interferon, CD, and HSP genes were also expressed in our study [12]. The KEGG database identified the presence of the PPAR signaling pathway, cell adhesion molecules, and the Toll-like receptor signaling pathway. The PPAR signaling pathway regulates fat metabolism and releases energy by breaking down fat. PPARs can also regulate gene transcription by antagonizing several important signaling pathways through various repression mechanisms. Activated PPARs regulate the inflammatory response by controlling the expression of multiple genes involved in inflammation. PPARs are expressed in a wide variety of tissues and cells within the immune system, including macrophages, dendritic cells (DCs), T cells, and B cells. PPARs play a crucial role in regulating the immune response [57].The Toll-like receptor signaling pathway is the most recent pathway identified by KEGG. The innate immune system is the body's first line of defense, detecting, responding to, controlling, and destroying invading pathogens. Toll-like receptors (TLRs) detect the presence of pathogens and initiate immune responses (refer to Figure 26-4). To date, 17 different TLRs have been identified in more than 12 different fish species. Several studies have shown that fish-specific TLRs have functional properties similar to those of their mammalian counterparts [58]. Viruses' ability to infect cells is impacted by oxygen levels, with some thriving in low-oxygen conditions. The research aimed to identify lncRNAs and explore their regulatory impact on target mRNAs, including genes from the interleukin family. lncRNAs are associated with crucial genes involved in immunity and meeting the cell's energy requirements. Gene ontology pathways demonstrate the diverse roles of immune response, cytokines, and other biological processes. Functional signaling pathways such as PPAR, cell adhesion molecules, and the Toll-like signaling pathway indicate cell involvement with external agents.
Two lncRNAs, MSTRG.982 and MSTRG.36357, were identified as common to both the cis and trans acting groups and showed distinct target genes. At the cis-targeting level, MSTRG.982 showed interaction with only one DEmRNA, LOC110527687 (synuclein), while MSTRG.36357 demonstrated cis interaction with LOC118948728 (RNA-binding protein 47) DEmRNA. Some interacting genes, such as the TFAP2A gene, are crucial components of this network. They are involved in the production of a protein known as AP-2 alpha (AP-2α) transcription factor. This protein plays a role in regulating cell division and apoptosis. The expression of this gene was upregulated and it had a cis relationship with the downregulated lncRNA MSTRG.20058, as well as a trans relationship with the downregulated lncRNAs MSTRG.34190 and MSTRG.22890. The number of trans interactions of MSTRG.982 was greater than the number of cis interactions. As a result, MSTRG.982 interacted with 5 DEmRNAs, including LOC110506699, LOC110521060, FAM78AB, LOC110513729, and LOC110534814. The number of MSTRG.36357–DEmRNA pairs was 9, including si:ch211-214j24.15, LOC110485438, LOC110510746 (LIM domain-binding protein 1), FAM78AB, LOC101268921 (SMAD4), PRLH2, LOC110485438, LOC110534579 (E3 ubiquitin-protein ligase TRIM21), and LOC110510746 (LIM-domain-binding protein 1 (LDB1)).
In 2019, Gupta et al. conducted an analysis of the expression of mRNAs and lncRNAs in macrophages of cows with Johne's disease or paratuberculosis, revealing new insights into Mycobacterium avium subspecies paratuberculosis [59]. The study's findings revealed that 397 new candidate lncRNAs in macrophages of diseased animals exhibited differential expression as a result of the disease. In total, approximately 820 genes associated with the disease exhibited significant alterations. The combined analysis of lncRNAs and their associated coding genes has unveiled the regulatory role of lncRNAs in pathways associated with the immune response. Wentao Cai et al. (2018) conducted an experiment on two groups of Chinese Holstein cows, with each group consisting of three cows. The groups were categorized based on their milk protein production, with one group having high production (≥ 3.5%) and the other having low production (≤ 3.0%). The aim of the experiment was to identify long non-coding RNAs in the lncRNA genome and explore their potential relationship with milk protein production[60]. As a result, 31 lncRNAs and 18 novel genes were identified in cows with high milk protein and cows with low milk protein. Differentially expressed lncRNAs were identified for predicting target genes through bioinformatics analysis. This involved integrating mRNA data with information on differential expression, functional genes, Gene Ontology (GO) and gene pathways, Genome-Wide Association Studies (GWAS), QTL and network analysis. As a result, several lncRNAs have been identified and shown to be useful as selective markers for predicting milk protein content [60]. In another study by Panero et al. (2016), the differentiation of expression of non-coding RNA genes in three lines of rainbow trout in response to Flavobacterium psychrophilum infection was examined [61]. Comparisons of protein-encoded RNAs from different lines in the 556-infection state revealed expression variations in lncRNAs. There was a positive correlation between the number of different lncRNAs and protein-encoding genes. There was a strong positive and negative correlation between certain lncRNAs and protein-encoding genes, such as cytokines, chemokines, and molecular compounds related to immunity. The results of this study indicate the potential for regulation of protein genes by lncRNAs. This type of study helps to develop and identify genetic markers associated with rainbow trout resistance to VHS virus stress [62]. Cho et al. (2017) conducted a comprehensive review of the integrated metabolic and transcriptomic responses to VHS virus in olive flounder fish. In this study, viral hemorrhagic septicemia virus (VHSV) amplification was measured continuously at 9, 24, 72, and 120 hours after infection. A total of 1382 mRNA molecules and 72 metabolites were selected to investigate the mechanism of viral infection and integrated into the KEGG database. Six KEGG metabolic pathways were identified, including carbohydrate metabolism, amino acid metabolism, lipid metabolism, transport and catabolism, cofactor and vitamin metabolism, and energy metabolism. Interestingly, these pathways are closely related to immune function. Suppressed by VHSV infection. A decrease in amino acids, such as valine, leucine, and isoleucine, as well as their carnitines, was observed following VHSV infection. Furthermore, the levels of arachidonic acid were elevated. Integrated analysis of the transcriptome and metabolome using the KEGG database revealed four renal responses to VHSV infection in olive flounder. Among these responses, mechanisms related to the immune system and active protein synthesis were inhibited, while ATP synthesis mechanisms and antioxidant system activity were suppressed. The results showed that inhibition of ATP synthesis and antioxidant systems, such as glutathione and peroxisome signaling, could cause necrosis, while immune activation could lead to inflammation of kidney tissue in VHSV-infected olives [10]. Fish skin is an important immune organ that serves as the initial protective barrier against the entry of foreign pathogens. In this study, we investigated the differences in innate immunity between the wild type of rainbow trout and its yellow mutant type by analyzing the complete transcriptome of the skin. A total of 2448 mRNAs, 1630 lncRNAs, 22 circRNAs, and 50 miRNAs were differentially expressed. Among the expressed mRNAs, several key immune-related genes, including IFIH1, DHX58, TRIM25, ATP6V1E1, TAP1, TAP2, CD209, HSP90A.1, NLRP3, NLRC3, and several other metabolism-related genes (GSTP3, NAPRT), were identified. Gene Ontology and Databank (KEGG) pathways associated with innate immunity, such as NAD+ADP-ribosyltransferase activity, immune response to bacterial infection, and RIG-I receptor signaling, including NOD-like receptor signaling and phagosomal KEGG pathways, are depicted. In addition, the role of long non-coding RNAs (lncRNAs) MSTRG.11484.2, MSTRG.32014.1, and MSTRG.29012.1 in the regulation of at least three immune-related genes (IFIH1, DHIRF) was determined [12].
5. Conclusions
Considering the vital role of lncRNAs in various cellular processes, this work aimed to elucidate the significant role of lncRNAs in the context of VHS virus disease, a highly lethal and detrimental disease in aquaculture. Identification of the differentially expressed lncRNAs that were neighboring mRNAs related to VHS-infected fish group yielded novel insights into the regulatory mechanisms of lncRNAs. Furthermore, the co-expression analysis unveiled potential roles of the identified lncRNAs in the vicinity of pertinent QTLs, offering avenues for further investigation for genomic interventions targeting VHS disease. A network of regulatory interactions was constructed, and several significant modules related to various sections of metabolism, immune response, cytokines, and other processes were identified. By finding target genes with known functions in immune response, oxygen metabolism, mTOR signaling, it is speculated that novel lncRNAs may function through neighboring or co-expressed target genes, which might contribute to any response to this infection by regulating these protein coding genes. Our results not only improved the annotation of the rainbow trout genome by reporting the novel lncRNAs, but also provided new insights into the function of lncRNAs in rainbow trout fish infected by VHS virus.
Funding
We would like to thank the Sari Agricultural Sciences and Natural Resources University and Mobarak Andish Advanced Technologies for the financial support of this work.
References
- Food and A.O.o.t.U.N.F. Department, The state of world fisheries and aquaculture. 1996: Food and Agriculture Organization of the United Nations.
- Smith, G.R.; Stearley, R.F. The Classification and Scientific Names of Rainbow and Cutthroat Trouts. Fisheries 1989, 14, 4–10. [Google Scholar] [CrossRef]
- McAllister, P. Fish Disease Leaflet 83. Viral Hemorrhagic Septicemia of Fishes; US Fish and Wildlife Service, National Fisheries Research Center-Leetown, National Fish Health Research Laboratory: Kearneysville, VA, 2013. [Google Scholar]
- Olesen, N.J.; Korsholm, H. Control measures for viral diseases in aquaculture: eradication of VHS and IHN. Bulletin of the European Association of Fish Pathologists 1997, 17, 229–233. [Google Scholar]
- Bokaei, S.; et al. Investigation of Outbreaks of Viral Hemorrhagic Septicemia and Associated Risk Factors in Rainbow Trout Farms in Iran, 2014. Iranian Journal of Epidemiology 2017, 13, 253–261. [Google Scholar]
- Yao, R.-W.; Wang, Y.; Chen, L.-L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef]
- Cech, T.R.; Steitz, J.A. The Noncoding RNA Revolution—Trashing Old Rules to Forge New Ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef]
- Li, B.J.; Jiang, D.L.; Meng, Z.N.; Zhang, Y.; Zhu, Z.X.; Lin, H.R.; Xia, J.H. Genome-wide identification and differentially expression analysis of lncRNAs in tilapia. BMC Genom. 2018, 19, 1–12. [Google Scholar] [CrossRef]
- Cho, S.-Y.; Kwon, Y.-K.; Nam, M.; Vaidya, B.; Kim, S.R.; Lee, S.; Kwon, J.; Kim, D.; Hwang, G.-S. Integrated profiling of global metabolomic and transcriptomic responses to viral hemorrhagic septicemia virus infection in olive flounder. Fish Shellfish. Immunol. 2017, 71, 220–229. [Google Scholar] [CrossRef]
- Wang, J.; Fu, L.; Koganti, P.P.; Wang, L.; Hand, J.M.; Ma, H.; Yao, J. Identification and Functional Prediction of Large Intergenic Noncoding RNAs (lincRNAs) in Rainbow Trout (Oncorhynchus mykiss). Mar. Biotechnol. 2016, 18, 271–282. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Li, Y.; Liu, Z.; Zhao, L. Integrated Analysis of lncRNA and circRNA Mediated ceRNA Regulatory Networks in Skin Reveals Innate Immunity Differences Between Wild-Type and Yellow Mutant Rainbow Trout (Oncorhynchus mykiss). Front. Immunol. 2022, 13, 802731. [Google Scholar] [CrossRef]
- Quan, J.; Kang, Y.; Luo, Z.; Zhao, G.; Ma, F.; Li, L.; Liu, Z. Identification and characterization of long noncoding RNAs provide insight into the regulation of gene expression in response to heat stress in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part D: Genom. Proteom. 2020, 36, 100707. [Google Scholar] [CrossRef]
- Ghaderzadeh, M.; Rahimi Mianji, G.; Nejati Javaremi, A.; Shahbazian, N. Effect of Viral Hemorrhagic Septicemia (VHS) disease on the biometric parameters in rainbow trout (Oncorhynchus mykiss). Veterinary Research & Biological Products 2021, 34, 111–123. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Kovaka, S.; Zimin, A.V.; Pertea, G.M.; Razaghi, R.; Salzberg, S.L.; Pertea, M. Transcriptome assembly from long-read RNA-seq alignments with StringTie2. Genome Biol. 2019, 20, 1–13. [Google Scholar] [CrossRef]
- Love, M.; Anders, S.; Huber, W. Differential analysis of count data–the DESeq2 package. Genome Biology 2014, 15, 10.1186. [Google Scholar]
- Kang, Y.-J.; Yang, D.-C.; Kong, L.; Hou, M.; Meng, Y.-Q.; Wei, L.; Gao, G. CPC2: a fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef]
- Camargo, A.P.; Sourkov, V.; Pereira, G.A.G.; Carazzolle, M.F. RNAsamba: neural network-based assessment of the protein-coding potential of RNA sequences. NAR Genom. Bioinform. 2020, 2, lqz024. [Google Scholar] [CrossRef]
- Camargo, A.P.; Sourkov, V.; Pereira, G.A.G.; Carazzolle, M.F. RNAsamba: neural network-based assessment of the protein-coding potential of RNA sequences. NAR Genom. Bioinform. 2020, 2, lqz024. [Google Scholar] [CrossRef]
- Li, A.; Zhang, J.; Zhou, Z. PLEK: a tool for predicting long non-coding RNAs and messenger RNAs based on an improved k-mer scheme. BMC Bioinform. 2014, 15, 1–10. [Google Scholar] [CrossRef]
- Rinn, J.L.; Chang, H.Y. Genome Regulation by Long Noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Fukunaga, T.; Hamada, M. RIblast: an ultrafast RNA–RNA interaction prediction system based on a seed-and-extension approach. Bioinformatics 2017, 33, 2666–2674. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl. 4), S11. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Hu, Z.-L.; Park, C.A.; Reecy, J.M. Developmental progress and current status of the Animal QTLdb. Nucleic Acids Res. 2015, 44, D827–D833. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118, Correction in 2021, 22, 159, 10.1038/s41580-021-00330-4. [Google Scholar] [CrossRef]
- Senthamarai, M.D.; Rajan, M.R.; Bharathi, P.V. Current risks of microbial infections in fish and their prevention methods: A review. Microb. Pathog. 2023, 185, 106400. [Google Scholar] [CrossRef]
- Gao, J.; Tang, Y.-D.; Hu, W.; Zheng, C. When Poly (A) Binding Proteins Meet Viral Infections, Including SARS-CoV-2. Journal of Virology 2022, 96, e00136-22. [Google Scholar] [CrossRef]
- Vallejo, R.L.; Fragomeni, B.O.; Cheng, H.; Gao, G.; Long, R.L.; Shewbridge, K.L.; MacMillan, J.R.; Towner, R.; Palti, Y. Assessing Accuracy of Genomic Predictions for Resistance to Infectious Hematopoietic Necrosis Virus With Progeny Testing of Selection Candidates in a Commercial Rainbow Trout Breeding Population. Front. Veter- Sci. 2020, 7. [Google Scholar] [CrossRef]
- Mattick, J.S.; et al. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nature Reviews Molecular Cell Biology 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, H.; Lu, Y.; Feng, T.; Sun, W. LncRNA BC032020 suppresses the survival of human pancreatic ductal adenocarcinoma cells by targeting ZNF451. Int. J. Oncol. 2018, 52, 1224–1234. [Google Scholar] [CrossRef]
- Sadallah, S.; Eken, C.; Martin, P.J.; Schifferli, J.A. Microparticles (Ectosomes) Shed by Stored Human Platelets Downregulate Macrophages and Modify the Development of Dendritic Cells. J. Immunol. 2011, 186, 6543–6552. [Google Scholar] [CrossRef]
- Guo, Q.; et al. Dynamic TF-lncRNA regulatory networks revealed prognostic signatures in the development of ovarian cancer. Frontiers in Bioengineering and Biotechnology 2020, 8, 460. [Google Scholar] [CrossRef]
- Toni, M.; Cioni, C. Fish Synucleins: An Update. Mar. Drugs 2015, 13, 6665–6686. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, X.; Huang, Z.; Ma, K. SARS-CoV-2 proteins interact with alpha synuclein and induce Lewy body-like pathology in vitro. International Journal of Molecular Sciences 2022, 23, 3394. [Google Scholar] [CrossRef]
- Kasen, A.; et al. Upregulation of α-synuclein following immune activation: Possible trigger of Parkinson's disease. Neurobiology of Disease 2022, 166, 105654. [Google Scholar] [CrossRef]
- Reish, H.E.A.; Standaert, D.G. Role of α-Synuclein in Inducing Innate and Adaptive Immunity in Parkinson Disease. J. Park. Dis. 2015, 5, 1–19. [Google Scholar] [CrossRef]
- Grathwohl, S.; et al. Experimental colitis drives enteric alpha-synuclein accumulation and Parkinson-like brain pathology. bioRxiv 2018, 505164. [Google Scholar]
- Kunimura, K.; Uruno, T.; Fukui, Y. DOCK family proteins: key players in immune surveillance mechanisms. Int. Immunol. 2019, 32, 5–15. [Google Scholar] [CrossRef]
- Peng, L.-B.; Wang, D.; Han, T.; Wen, Z.; Cheng, X.; Zhu, Q.-L.; Zheng, J.-L.; Wang, P. Histological, antioxidant, apoptotic and transcriptomic responses under cold stress and the mitigation of blue wavelength light of zebrafish eyes. Aquac. Rep. 2022, 26. [Google Scholar] [CrossRef]
- Reyes, A.; Corrales, N.; Galvez, N.M.S.; Bueno, S.M.; Kalergis, A.M.; Gonzalez, P.A. Contribution of hypoxia inducible factor-1 during viral infections. Virulence 2020, 11, 1482–1500. [Google Scholar] [CrossRef]
- Leon, E.; Nde, C.; Ray, R.S.; Preciado, D.; Zohn, I.E. ALDH1A2-related disorder: A new genetic syndrome due to alteration of the retinoic acid pathway. Am. J. Med Genet. Part A 2022, 191, 90–99. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, C.; Yu, N.; Si, L.; Zhu, L.; Zeng, A.; Liu, Z.; Wang, X. INF2 regulates oxidative stress-induced apoptosis in epidermal HaCaT cells by modulating the HIF1 signaling pathway. Biomed. Pharmacother. 2019, 111, 151–161. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Cheng, J.; Xia, Y.; Fan, K.; Liu, Y.; Liu, P.-F. Identification and expression analysis of a fibrinogen alpha chain-like gene in Atlantic salmon (Salmo salar). Aquac. Rep. 2021, 22, 100919. [Google Scholar] [CrossRef]
- Chen, Y.; Li, H.; Cheng, H.-Y.; Rui-Qiong, M.; Ye, X.; Cui, H.; Hong-Lan, Z.; Chang, X.-H. Fibrinogen alpha chain is up-regulated and affects the pathogenesis of endometriosis. Reprod. Biomed. Online 2019, 39, 893–904. [Google Scholar] [CrossRef]
- Liao, Y.; Weng, J.; Chen, L.; Hu, N.; Yuan, X.; Wang, J.; He, F.; Cai, Y.; Huang, Q.; Wang, J.; et al. Comprehensive analysis of SLC43A2 on the tumor immune microenvironment and prognosis of liver hepatocellular carcinoma. Front. Genet. 2022, 13. [Google Scholar] [CrossRef]
- Gong, N.; Ferreira-Martins, D.; McCormick, S.D.; Sheridan, M.A. Divergent genes encoding the putative receptors for growth hormone and prolactin in sea lamprey display distinct patterns of expression. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Cicenas, J.; et al. KRAS, TP53, CDKN2A, SMAD4, BRCA1, and BRCA2 mutations in pancreatic cancer. Cancers 2017, 9, 42. [Google Scholar] [CrossRef]
- Rosic, J.; Dragicevic, S.; Miladinov, M.; Despotovic, J.; Bogdanovic, A.; Krivokapic, Z.; Nikolic, A. SMAD7 and SMAD4 expression in colorectal cancer progression and therapy response. Exp. Mol. Pathol. 2021, 123, 104714. [Google Scholar] [CrossRef]
- Ahmed, S.; Bradshaw, A.-D.; Gera, S.; Dewan, M.Z.; Xu, R. The TGF-β/Smad4 Signaling Pathway in Pancreatic Carcinogenesis and Its Clinical Significance. J. Clin. Med. 2017, 6, 5. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in inflammatory disease. International journal of molecular sciences 2019, 20, 6008. [Google Scholar] [CrossRef]
- Liu, S.; Pan, C.; Tang, Y.; Chen, F.; Yang, M.; Wang, K.-J. Identification of novel long non-coding RNAs involved in bisphenol A induced immunotoxicity in fish primary macrophages. Fish Shellfish. Immunol. 2020, 100, 152–160. [Google Scholar] [CrossRef]
- Daynes, R.A.; Jones, D.C. Emerging roles of PPARS in inflammation and immunity. Nat. Rev. Immunol. 2002, 2, 748–759. [Google Scholar] [CrossRef]
- Rebl, A.; Goldammer, T.; Seyfert, H.-M. Toll-like receptor signaling in bony fish. Veter- Immunol. Immunopathol. 2010, 134, 139–150. [Google Scholar] [CrossRef]
- Gupta, P.; Peter, S.; Jung, M.; Lewin, A.; Hemmrich-Stanisak, G.; Franke, A.; von Kleist, M.; Schütte, C.; Einspanier, R.; Sharbati, S.; et al. Analysis of long non-coding RNA and mRNA expression in bovine macrophages brings up novel aspects of Mycobacterium avium subspecies paratuberculosis infections. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Cai, W.; Li, C.; Liu, S.; Zhou, C.; Yin, H.; Song, J.; Zhang, Q.; Zhang, S. Genome Wide Identification of Novel Long Non-coding RNAs and Their Potential Associations With Milk Proteins in Chinese Holstein Cows. Front. Genet. 2018, 9, 281. [Google Scholar] [CrossRef]
- Marancik, D.; Gao, G.; Paneru, B.; Ma, H.; Hernandez, A.G.; Salem, M.; Yao, J.; Palti, Y.; Wiens, G.D. Whole-body transcriptome of selectively bred, resistant-, control-, and susceptible-line rainbow trout following experimental challenge with Flavobacterium psychrophilum. Front. Genet. 2015, 5, 453–453. [Google Scholar] [CrossRef]
- Paneru, B.; Al-Tobasei, R.; Palti, Y.; Wiens, G.D.; Salem, M. Differential expression of long non-coding RNAs in three genetic lines of rainbow trout in response to infection with Flavobacterium psychrophilum. Sci. Rep. 2016, 6, 36032–36032. [Google Scholar] [CrossRef]
Figure 1.
Analysis pipeline for the identification, annotation, cis- and trans-acting, and differential expression of lncRNAs and mRNAs in rainbow trout under viral treatments.
Figure 1.
Analysis pipeline for the identification, annotation, cis- and trans-acting, and differential expression of lncRNAs and mRNAs in rainbow trout under viral treatments.
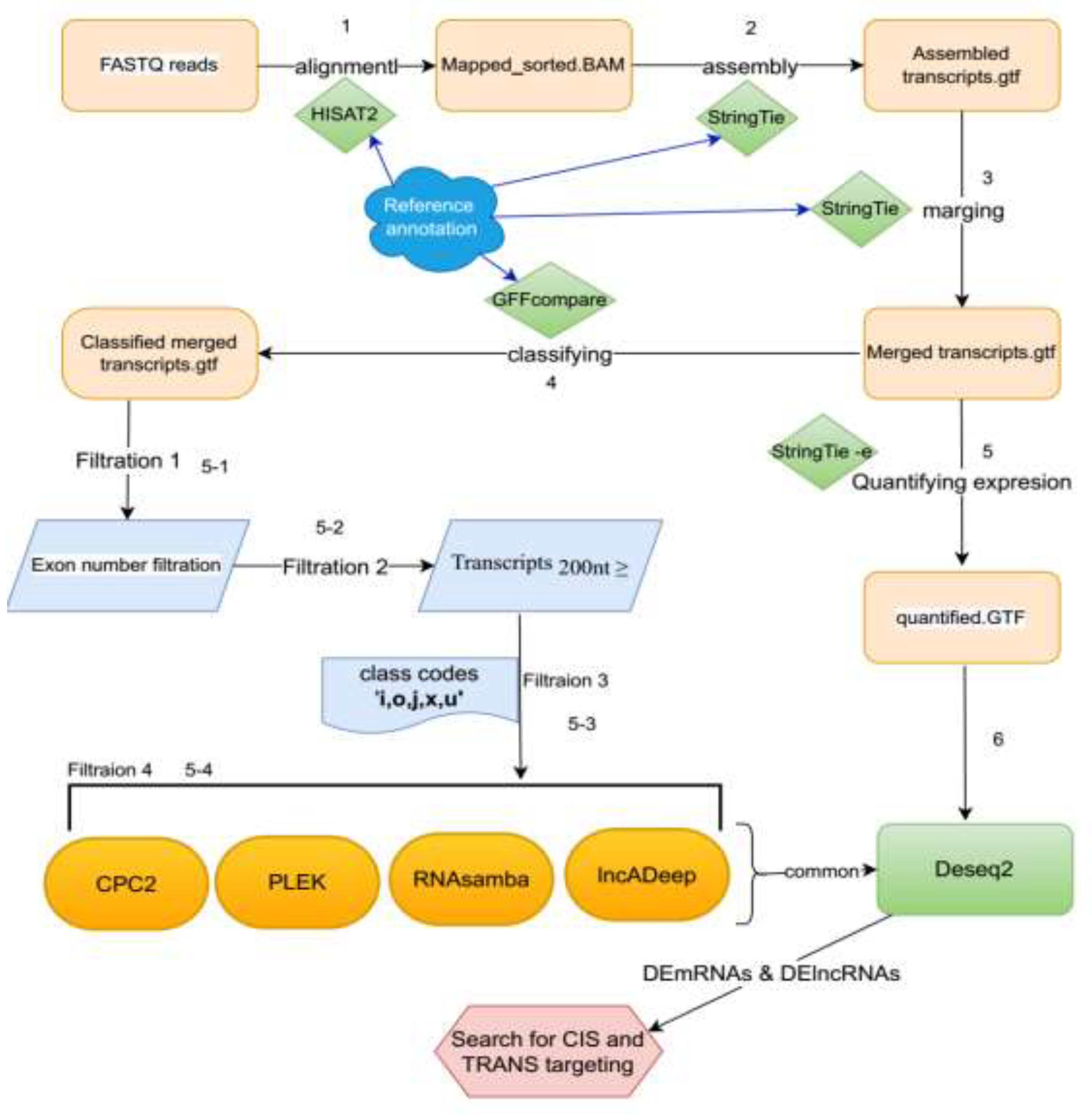
Figure 3.
(A) Volcano plot of lncRNAs. (B) Clustering analysis showing the expression profiles of DElncRNAs between control and virus groups. (C) PCA of expressed genes.
Figure 3.
(A) Volcano plot of lncRNAs. (B) Clustering analysis showing the expression profiles of DElncRNAs between control and virus groups. (C) PCA of expressed genes.
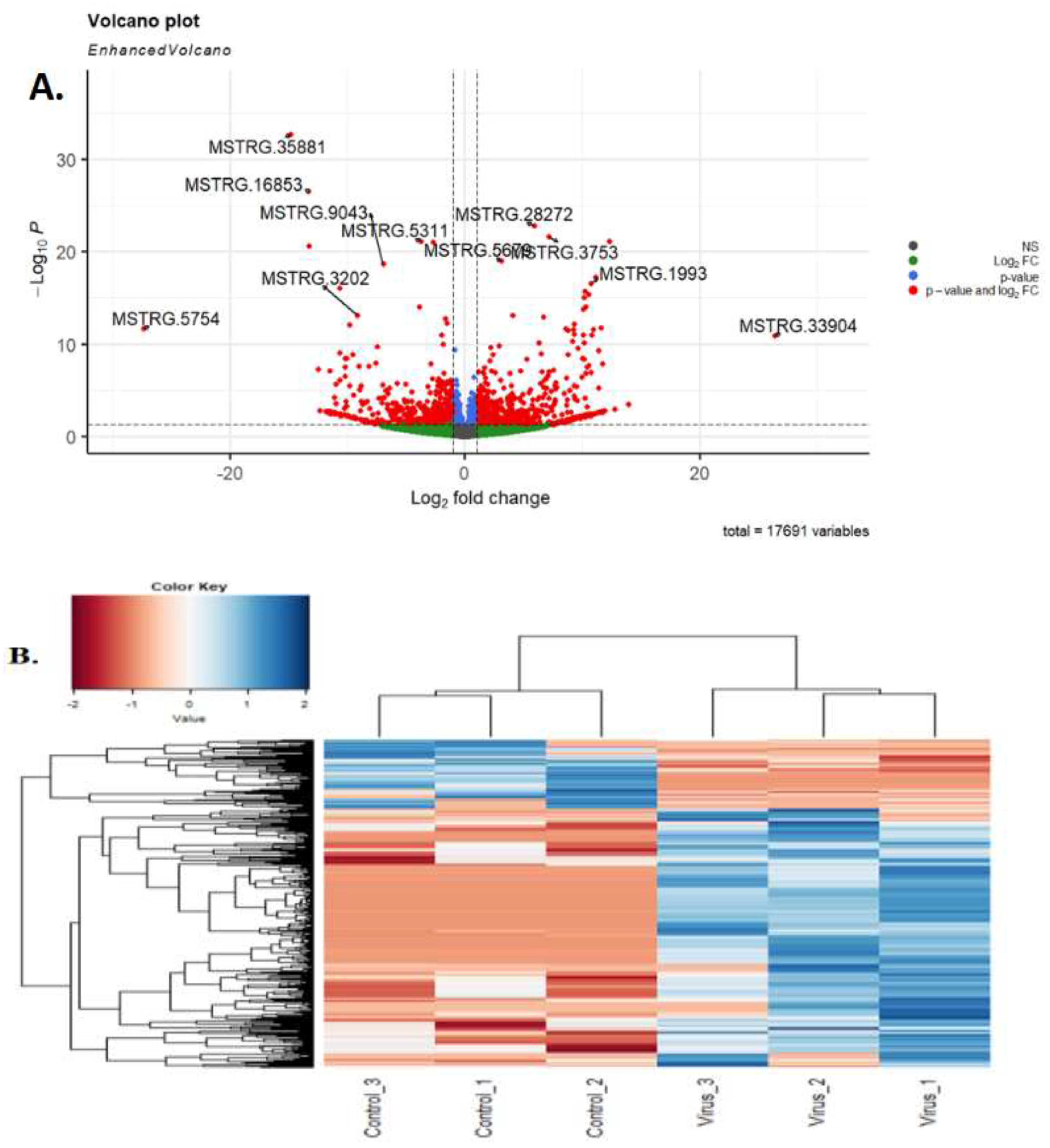
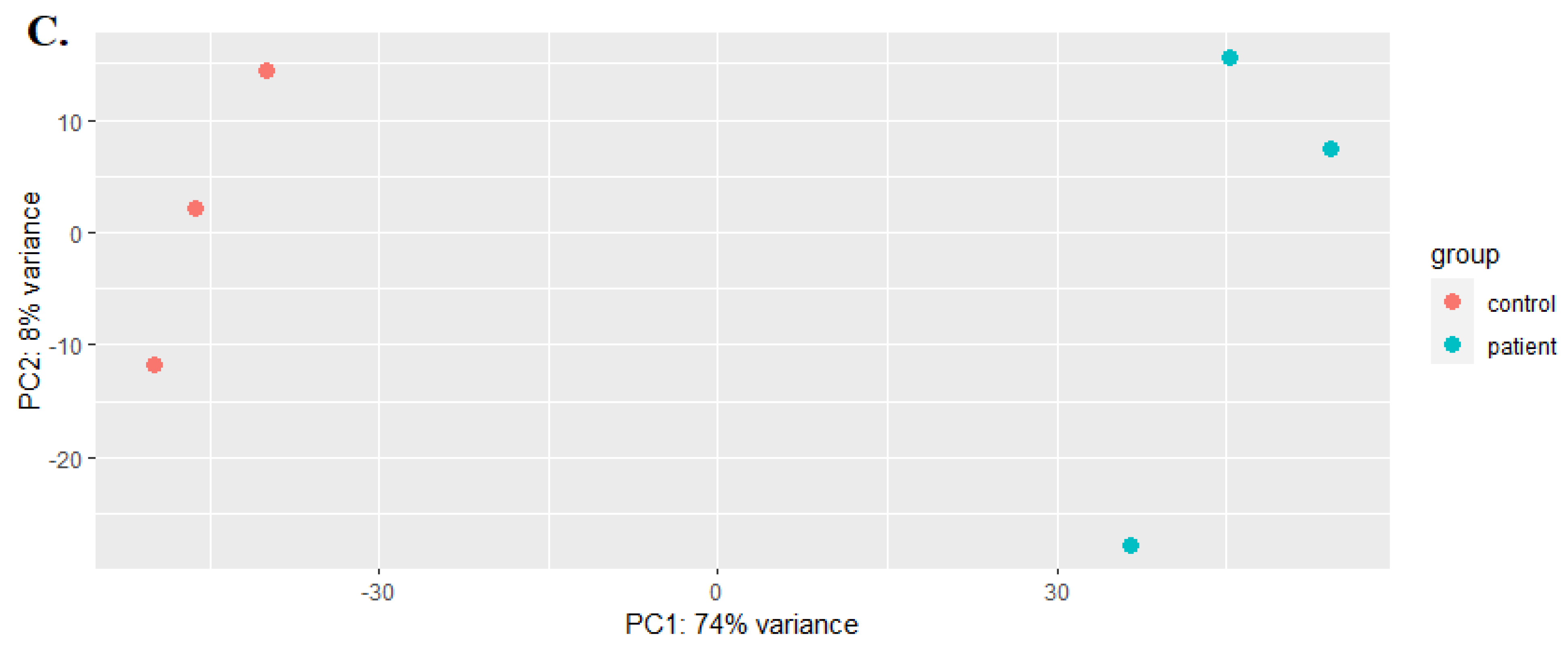

Table 1.
Quality statistics of different sample sequencing data.
| Sample | Raw Data | Valid Data | Valid Ratio (%) | Q20 (%) | Q30 (%) | Mapped Ratio (%) |
| Control | 42,374,402 | 42,087,999 | 99.32 | 99.88 | 92.25 | 91.17 |
| Control | 40,339,708 | 40,035,219 | 99.25 | 99.87 | 91.14 | 90.53 |
| Control | 54,456,286 | 53,829,279 | 98.85 | 99.85 | 91.07 | 90.25 |
| Treated | 45,502,028 | 45,364,200 | 99.70 | 99.91 | 91.87 | 89.92 |
| Treated | 48,998,728 | 48,930,236 | 99.86 | 99.93 | 91.77 | 90.23 |
| Treated | 44,135,524 | 43,796,603 | 99.23 | 99.88 | 91.40 | 88.89 |
Table 5.
displays the cis regulatory effects of DElncRNAs on DEmRNAs within a range of 5 kilobases upstream and downstream.
Table 5.
displays the cis regulatory effects of DElncRNAs on DEmRNAs within a range of 5 kilobases upstream and downstream.
| Tab. | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| mRNA | Regulation | lncRNA | class code | Regulation | mRNA | Regulation | lncRNA | class code | Regulation |
| TP63 | Down | MSTRG.62 | j | Up | LOC118944460 | Down | MSTRG.32002 | o | Down |
| LOC110524396 | Down | MSTRG.148 | x | Up | LOC118944461 | Down | MSTRG.32002 | o | Down |
| BHLHE41 | Down | MSTRG.595 | u | Up | LOC110528420 | Up | MSTRG.9557 | j | Up |
| LOC110525089 | Down | MSTRG.785 | j | Down | IL10 | Up | MSTRG.9783 | j | Up |
| LOC110526498 | Down | MSTRG.896 | o | Down | IL10 | Up | MSTRG.9784 | u | Down |
| NCOA4 | Down | MSTRG.956 | j | Down | LOC110529378 | Up | MSTRG.9966 | j | Down |
| LOC110527687 | Down | MSTRG.982 | j | Down | LOC110529379 | Down | MSTRG.9966 | j | Down |
| LOC110512766 | Down | MSTRG.1363 | j | Down | LOC110529615 | Up | MSTRG.10141 | j | Up |
| LOC110525147 | Down | MSTRG.1563 | j | Down | LOC118936275 | Down | MSTRG.10832 | j | Down |
| TUFT1A | Down | MSTRG.1680 | j | Up | LOC110530481 | Down | MSTRG.10832 | j | Down |
| LOC118937935 | Up | MSTRG.1706 | j | Up | LOC110530704 | Up | MSTRG.11013 | j | Up |
| LOC110485723 | Up | MSTRG.1993 | j | Up | HAUS6 | Down | MSTRG.11735 | j | Down |
| LOC110502310 | Down | MSTRG.2952 | j | Down | LOC110532684 | Down | MSTRG.12397 | j | Up |
| LOC110521085 | Up | MSTRG.5084 | j | Up | E1 | Down | MSTRG.12464 | j | Down |
| EFNA1B | Up | MSTRG.1700 | u | Down | LOC110533701 | Up | MSTRG.13116 | j | Up |
| LOC118937935 | Up | MSTRG.3327 | j | Up | LOC110497318 | Up | MSTRG.25735 | j | Up |
| LOC110516769 | Down | MSTRG.3347 | j | Down | LOC110497317 | Up | MSTRG.25735 | j | Up |
| LOC110520884 | Down | MSTRG.3347 | j | Down | LOC110497274 | Down | MSTRG.13985 | j | Down |
| LOC110520883 | Down | MSTRG.3347 | j | Down | LOC110497275 | Down | MSTRG.13985 | j | Down |
| GHR1 | Up | MSTRG.5679 | j | Down | LOC110514189 | Up | MSTRG.14211 | j | Up |
| LOC110523752 | Down | MSTRG.5854 | j | Down | LOC110535281 | Up | MSTRG.14419 | j | Up |
| SID1 | Up | MSTRG.6203 | j | Up | LOC110535279 | Down | MSTRG.14419 | j | Up |
| LPL | Down | MSTRG.6262 | j | Down | LOC110535540 | Down | MSTRG.14622 | j | Down |
| LOC110526114 | Down | MSTRG.7664 | j | Down | SLC20A1A | Up | MSTRG.14967 | j | Up |
| ALDH1A2 | Up | MSTRG.31711 | j | Up | SLC25A37 | Up | MSTRG.14992 | j | Up |
| LOC110506541 | Down | MSTRG.31713 | j | Down | NAALADL1 | Up | MSTRG.15351 | j | Down |
| LOC110506917 | Down | MSTRG.32002 | o | Down | LOC110537350 | Down | MSTRG.15881 | j | Down |
| Dock10 | Up | MSTRG.30015 | j | Down | |||||
Table 6.
TRANS acting pattern of DElncRNAs on DEmRNAs with interaction energy (<-25 kcal/mol).
| lncRNAs | Expr | mRNAs | Expr | InEnergy | lncRNAs | Expr | mRNAs | Expr | InEnergy |
|---|---|---|---|---|---|---|---|---|---|
| MSTRG.17204 | down | LOC110486166 | up | -20.1681 | MSTRG.24034 | down | LOC110503011 | down | -25.8338 |
| MSTRG.19333 | up | SI:DKEY-89B17.4 | down | -22.5963 | MSTRG.24034 | up | LOC110503701 | down | -22.4431 |
| MSTRG.19333 | up | LOC110486530 | up | -22.7976 | MSTRG.24034 | up | LOC110508925 | up | -23.4754 |
| MSTRG.19333 | up | LOC110486615 | up | -20.9064 | MSTRG.27872 | up | FAM78AB | down | -20.3565 |
| MSTRG.19333 | up | LOC110489230 | up | -20.1649 | MSTRG.27872 | up | LOC110504281 | up | -20.3359 |
| MSTRG.29125 | up | LOC110489230 | up | -21.5915 | MSTRG.34190 | up | ENPP2 | up | -21.8946 |
| MSTRG.29125 | up | LOC110490321 | down | -21.1108 | MSTRG.34190 | up | LOC110525147 | up | -22.0789 |
| MSTRG.29125 | up | LOC100136069 | up | -22.1927 | MSTRG.34190 | up | LOC110489230 | up | -27.3874 |
| MSTRG.34190 | down | MEOX2A | up | -20.8682 | MSTRG.34190 | down | TFAP2A | up | -20.5328 |
| MSTRG.34190 | down | CDH13 | down | -24.9856 | MSTRG.34190 | down | LOC118938874 | down | -20.3688 |
| MSTRG.34190 | down | LOC110500756 | down | -30.236 | MSTRG.34190 | down | LOC110491320 | up | -21.5654 |
| MSTRG.36357 | down | SI:CH211-214J24.15 | down | -21.568 | MSTRG.34190 | down | LOC110493456 | up | -23.9436 |
| MSTRG.36357 | down | LOC110485438 | down | -23.9051 | MSTRG.35974 | down | ANTXR1B | down | -22.1156 |
| MSTRG.36357 | down | LOC110510746 | up | -21.2892 | MSTRG.35974 | down | LOC110510746 | up | -20.399 |
| MSTRG.36357 | down | FAM78AB | down | -23.6439 | MSTRG.35974 | down | LOC110534579 | up | -22.3215 |
| MSTRG.36357 | down | LOC101268921 | up | -20.3122 | MSTRG.35974 | down | CHRNA2A | down | -22.1661 |
| MSTRG.36357 | down | PRLH2 | up | -20.1694 | MSTRG.35974 | down | KDM6BA | down | -21.7067 |
| MSTRG.36357 | down | LOC110534579 | up | -20.7044 | MSTRG.982 | down | LOC110506699 | down | -21.1293 |
| MSTRG.29125 | up | LOC110508925 | up | -20.2593 | MSTRG.982 | up | LOC110521060 | down | -20.433 |
| MSTRG.29125 | up | LOC110521826 | up | -28.0706 | MSTRG.982 | up | FAM78AB | down | -26.6405 |
| MSTRG.13869 | up | LOC100136069 | up | -25.5793 | MSTRG.982 | up | LOC110513729 | up | -20.1696 |
| MSTRG.13869 | up | LOC110534579 | up | -20.342 | MSTRG.982 | up | LOC110534814 | up | -22.5472 |
| MSTRG.13869 | up | SI:CH73-22O12.1 | up | -20.6675 | MSTRG.19333 | up | LOC110503011 | down | -21.2381 |
| MSTRG.22011 | up | LOC100136069 | up | -21.3084 | MSTRG.22011 | up | ZNF385A | up | -20.8434 |
| MSTRG.22011 | up | LOC118936283 | up | -21.8654 | MSTRG.22011 | up | LOC110510746 | up | -28.6177 |
| MSTRG.22011 | down | LOC110486615 | up | -20.4733 | MSTRG.24034 | down | LOC110489230 | up | -20.3093 |
| MSTRG.24034 | down | LOC110510746 | up | -31.7948 | MSTRG.24034 | down | PRLH2 | up | -21.9129 |
| MSTRG.24034 | down | ZNF385A | up | -20.3607 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



