Submitted:
13 May 2024
Posted:
14 May 2024
You are already at the latest version
Abstract
Dengue infection is caused by the Dengue virus (DENV) and is transmitted to humans by infected female Aedes aegypti and Aedes albopictus mosquitoes. There are nearly 100 million new dengue cases yearly in more than 120 countries with a five-fold increase in incidence over the past four decades. While many patients experience a mild illness, a subset suffer from severe disease which can be fatal. Dysregulated immune responses are central to the pathogenesis of dengue, and haematologic manifestations are a prominent feature of severe disease. While thrombocytopaenia and coagulopathy are major causes of bleeding in severe dengue, leucocyte abnormities are emerging as important markers of prognosis. In this review, we provide our perspective on the clinical aspects and pathophysiology of haematologic manifestations in dengue. We also discuss the key gaps in our current practice and areas to be addressed by future research.
Keywords:
Dengue
; Immunohematology
; Thrombocytopaenia
; Immune dysregulation
; Haemostatis
Introduction
Dengue infection is caused by the Dengue virus (DENV), a positive-sense, single-stranded RNA virus which belongs to the Flaviviridae family and is transmitted by Aedes aegypti and Aedes albopictus mosquitoes.[1] DENVs comprise four serotypes, which are distinguished based on differences in viral structural and non-structural proteins. [2] Infection with each serotype confers lifelong immunity for the causative serotype but not for others.[3] The incidence of dengue is on the increase, with 100-400 million new infections each year, and the highest prevalence being in countries with tropical climates.[4] It is noteworthy that only a quarter of these patients become symptomatic.[5] Clinical manifestations may be limited to mild fever, headache and myalgia, or may progress to severe dengue.
The 2009 World Health Organisation ( WHO) criteria classifies dengue according to three levels of severity including dengue with and without warning signs and severe dengue with the aim of guiding management. [6] Dengue infection is most commonly diagnosed using the enzyme linked immunosorbent assay (ELISA) which measures dengue immunoglobulin G (IgG) and immunoglobulin M (IgM) antibodies.[7] The alternative being the non-structural antigen 1 ( NS-1), or the dengue reverse transcriptase PCR (RT-PCR) assay.[8] The management of dengue comprises mainly supportive therapy, which is adequate for most patients.[9] However, a subset of patients develops severe disease for which the optimal management remains uncertain.
Severe dengue occurs in 500 000 people per year, with a mortality rate of 10% among hospitalised patients.[10] A dysregulated host immune response, including antibody dependent enhancement (ADE) [11] is thought to drive most of the complications associated with severe dengue, in particular the haematologic manifestations.[12] Haematologic abnormalities are a key feature of dengue, with coagulopathy (Figure1), thrombocytopaenia (Figure 2) and leucocytes abnormalities (Figure 3) being well described. The pathogenesis and management of these are the subject of active research. In this review, we provide our perspective on the immuno- haematological aspects of dengue infection with a focus on their biological and clinical implications.
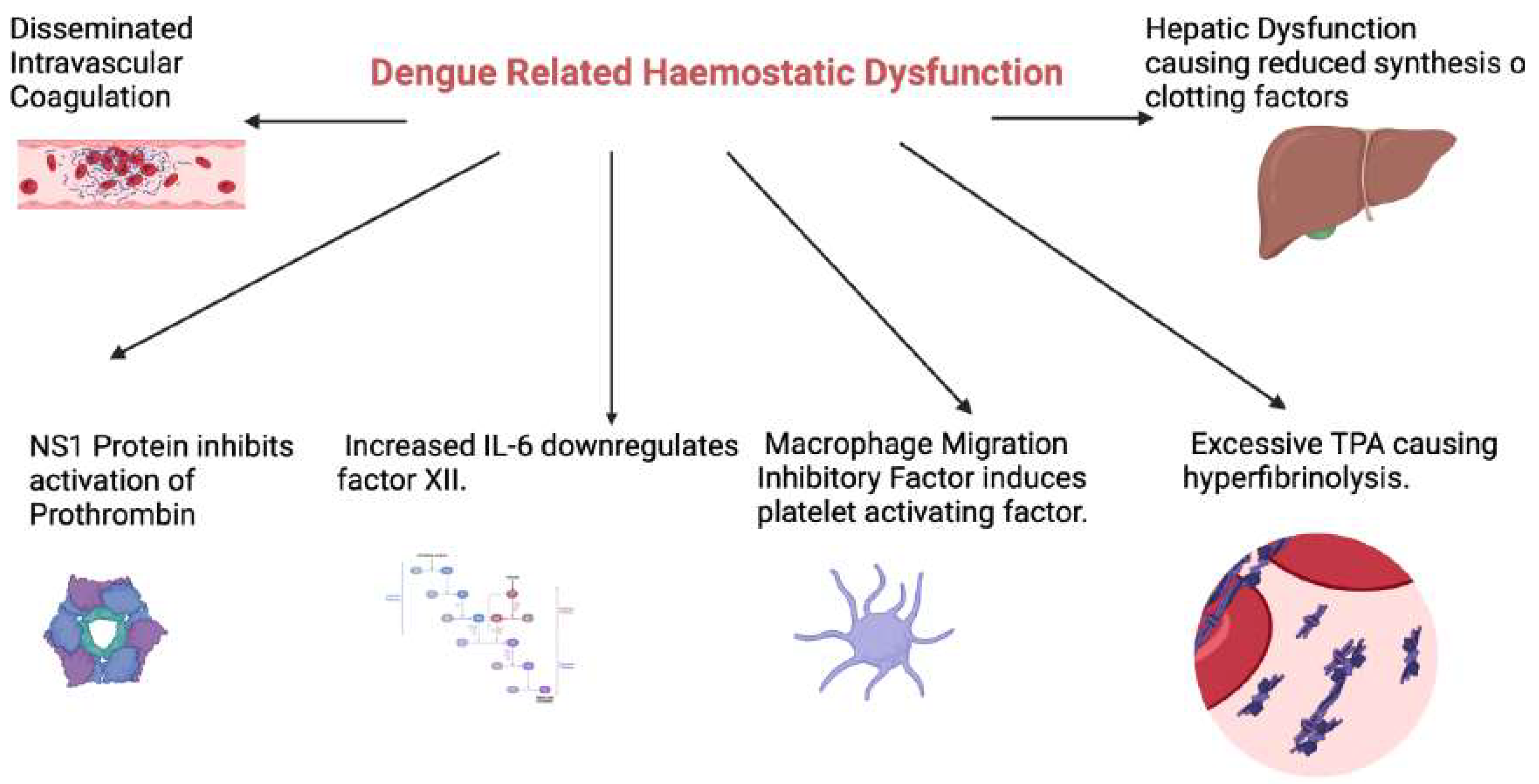
Figure 1.
An overview of the mechanisms underlying dengue related haemostatic derangements. NS1: Dengue nonstructural antigen 1, IL-6: Interleukin 6, TPA: Tissue plasminogen activator. (Figure created with Biorender.com).
Figure 1.
An overview of the mechanisms underlying dengue related haemostatic derangements. NS1: Dengue nonstructural antigen 1, IL-6: Interleukin 6, TPA: Tissue plasminogen activator. (Figure created with Biorender.com).
Haemostatic Abnormalities in Dengue Infection
Derangement of laboratory coagulation assays is relatively common in dengue infection. A meta-analysis of forty-two studies reported the frequency of prolonged activated partial thromboplastin time (aPTT) and prothrombin time (PT) among patients with dengue at 42.9% and 16.5% respectively.[13] Despite the high frequency of prolonged APTT and PT, clinically significant bleeding manifestations are less common.[14] In the event of bleeding, blood transfusion requirements are low. In a retrospective study done over 7 years in a tertiary hospital in South India, clinically significant bleeding manifestations occurred in 44.7% of patients with dengue infection but only 1.2% of them required packed cell transfusion.[15]
The specific mechanism underlying coagulation abnormalities associated with dengue remains unclear. Various pathogenic mechanisms have been proposed. The aPTT and PT serve as indicators for the intrinsic and extrinsic pathways of the coagulation system, respectively.[16] Prolongation of PT and aPTT may result from either decreased synthesis of specific factors or an increased consumption of these factors.16 The NS1 protein can bind to both thrombin and prothrombin.[17] While binding to thrombin has no effects, it inhibits the activation of prothrombin. [18] The prolonged aPTT in dengue patients is due to a deficiency in the intrinsic pathway, possibly stemming from impaired coagulation factor synthesis from liver failure.[19] An imbalance between coagulation and fibrinolysis is also postulated to cause haemorrhagic complications in DHS and DSS.[20] DENV induced macrophage migration inhibitory factor promotes bleeding by inducing platelet activating factor.[21] Furthermore, increased IL-6 down-regulates the production of factor XII, which initiates the intrinsic coagulation pathway, resulting in prolongation of the aPTT. [22]. Excessive tissue plasminogen activator (tPA) production in DHS/DSS leads to hyperfibrinolysis and further exacerbates the bleeding tendency.[23]
Indeed, dengue patients with bleeding complications were shown to have higher levels of tPA, D-dimer and reduced thrombin formation compared to those without bleeding.[24] Similar results were demonstrated in children with DHF, where von Willebrand factor antigen(vWF: Ag), tissue factor(TF) and plasminogen activator inhibitor(PAI-1) were increased, but ADAMTS-13 (a dis-integrin and metalloprotease with thrombospondin repeats) was significantly reduced.[25] Disseminated intravascular coagulation (DIC) is also a major contributor to dengue related coagulopathy and was recently reported to occur in 26% of patients with severe disease, in addition to being a predictive factor for early mortality.[26] Taken together, these data suggest that reduced thrombin formation and increased fibrinolytic activity are the key contributing factors to bleeding complications in dengue infection. It is noteworthy that dengue related coagulopathy is a result of both the direct effects of the virus on the haemostatic system as well as the dysregulated host immune response.[27]
There is currently a paucity of predictive factors to predict bleeding manifestations due to dengue. Further studies are required to identify patients who may need closer monitoring of their coagulation system and potentially earlier intervention. Furthermore, the optimal management of dengue related coagulopathy needs to be established, especially with regards to the indication and timing for the use of blood products.
Thrombocytopaenia
Thrombocytopenia is a major contributor to the haemorrhagic tendency in dengue infection, occurring in up to 79% of cases, with the elderly being at higher risk.[28] Dengue related thrombocytopaenia typically occurs on days 4-6 and recovers by day 10.[29] The platelet nadir is variable, ranging from 40–90 x 10^9/l with a median nadir of 65 10^9/l according to a prospective observational study done among 2300 children with dengue infection.[30]
In the acute phase of secondary dengue, the DENV binds to platelets, forming platelet-associated IgM (PA-IgM), resulting in platelet destruction.[31] In young patients with DHF, platelet aggregation is decreased, and the plasma levels of platelet factor-4 and beta thrombomodulin are increased, contributing to bleeding complications.[32] Interleukin-1B, released from platelet microparticles via inflammasome activation, also promotes vascular permeability, leading to plasma leakage.[33] It is noteworthy that levels of platelet microparticles and plasma P-selectin correlate with the onset and recovery from thrombocytopenia in dengue infection.[34] In addition, complement C5b-9 mediated apoptosis of activated platelets and clearance of IgG bound platelets have been observed to occur on days 4-6 of dengue infection, accounting for the significant drop in platelet count from day four onwards.[35] The consumption of platelets is also accelerated by DENV-induced disseminated intravascular coagulation (DIC) as discussed above.
DENV-induced bone marrow suppression is proposed to occur within 3 to 4 days of dengue infection.[36] The reduction in haematopoiesis is likely a protective mechanism to limit the injury to the bone marrow progenitor cells during elimination of dengue-infected cells.[37] Bone marrow suppression is likely to contribute significantly to DENV related thrombocytopaenia. The fact that the immature platelet fraction (a surrogate for thrombopoiesis) in the peripheral blood is significantly lower in patients with severe dengue supports this hypothesis.[38] This is however difficult to study definitively, as bone marrow aspiration/biopsy is rarely performed in patients with Dengue infection. A variety of clinical parameters have been proposed which may enable early identification of patients who are likely to develop severe thrombocytopaenia.[39,40,41,42] Further studies are required to validate these findings and derive globally applicable prognostic scores for dengue related thrombocytopaenia.
Leucocyte Abnormalities
Monocytes and Macrophages
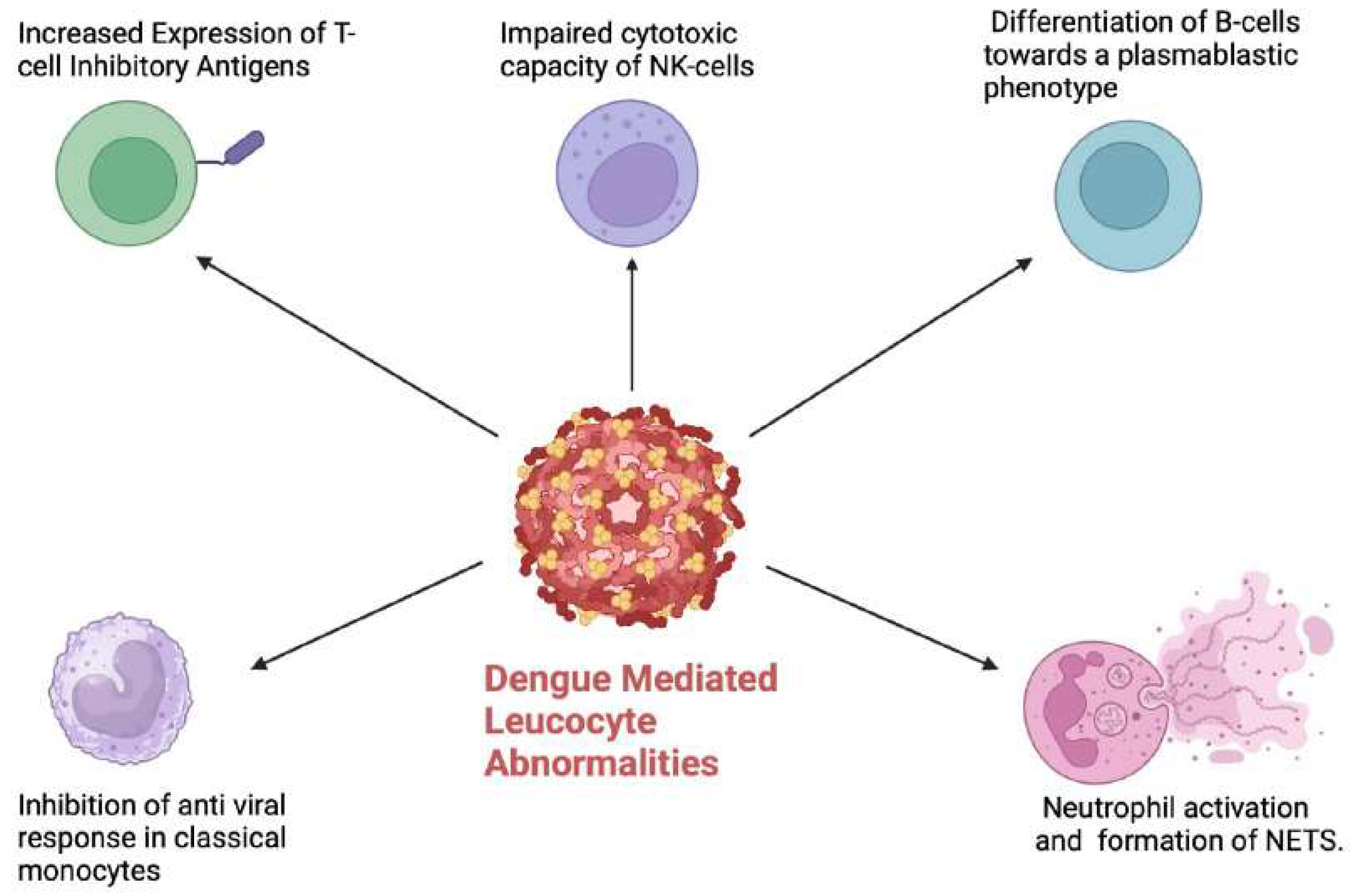
Monocytes are a key target of the DENV and while being crucial for the host anti-viral response, they also contribute to ADE. [43] Several monocyte subsets have been implicated in the pathogenesis of dengue,[44] and the DENV appears to have a predilection to infect some subsets over others. The biological role of specific monocyte subsets in dengue remains an active field of research.[45,46] CD14+/CD16+ intermediate monocytes were found to be increased in paediatric patients with dengue up until day 3 of infection followed by a decline from day 4 onwards.[47] Interestingly, intracellular staining demonstrated DENV infection in the classical (CD14bright/CD16-) and intermediate monocytes subsets but not the non-classical monocytes. The role of monocytes was reinforced in this study through the detection of raised IL-10 mRNA levels from CD14+ monocytes compared to other subsets of peripheral blood mononuclear cells. The increased IL-10 levels resulted in upregulation of pathways inhibiting the anti-viral response including the suppression of JAK-STAT signalling.[48] Furthermore, in -vitro infection of monocytes with the DENV resulted in suppression of Interferon-Beta and induced nitric oxide synthesis with sub-neutralising concentrations of DENV specific antibodies.[49] Collectively, these data suggest that the DENV “hijacks” monocytes to downregulate innate anti-viral immune responses.
DENV infected macrophages have been detected in the tissues of patients with dengue, [50]and macrophage recruitment to the endothelium has been implicated in the development of haemorrhage.[51] It has been postulated that macrophages at different anatomic sites may have distinctive roles in the response to dengue infection, with some subsets being permissive to DENV.[52] Indeed, it is possible that macrophage dysfunction underlies dengue related Hemophagocytic lymphohistiocytosis (HLH). HLH is characterized by inappropriate macrophage stimulation resulting in a cytokine storm and a severe systemic inflammatory response.[53] Variable levels of haemo-phagocytosis were seen on post-mortem studies of patients with severe dengue, in contrast to those with mild disease.[54] In a retrospective single centre study, mortality due to severe dengue was 22% and approximately 40% of these patients had dengue related HLH.[55] The phenotype of macrophages in dengue associated HLH has not been adequately characterised and is an important area to be addressed for future research.
Further studies are required to further unravel the biological significance of the monocyte/macrophage lineage specially in the pathogenesis of severe dengue. Integration of single cell RNA sequencing with flow cytometry at serial time points of the disease would provide crucial information on the evolution of monocyte subsets and their function. Differential gene expression analysis between monocyte subsets infected by DENV and those which are not would also be of significant interest. Ideally, changes in monocytes subsets should be studied in the context of other immune cell populations including lymphocytes, as it is likely that a complex interplay exists between these populations.[56]
Lymphocytes
Lymphocytes play a crucial role in the immune response against viral infections including dengue.[57] Interestingly, a lower mean lymphocyte count was reported as a biomarker to distinguish dengue from non-dengue viral fevers[58] , while a higher percentage of lymphocytes in the WBC differential was associated with a shorter hospital stay in a study from India.[59]A lower absolute lymphocyte count was also reported as a predictor of severe dengue in a separate study.[60]These conflicting data suggest that a specific subset of lymphocytes (rather than total lymphocytes) may be more relevant in the pathophysiology of dengue infection.
The finding of “atypical” or “reactive” lymphocytes in dengue infection has been well described for several years. [61] Atypical lymphocytes (AL) are readily detected by evaluation of the peripheral blood film[62] , and more recently by automated haematology analysers.[63] Current analysers identify AL based on their light scatter properties which generates the AL count (a research parameter) as part of the full blood count read out. These data have been used by several groups to describe the kinetics of AL and their potential prognostic significance.[64] AL can be detected at presentation, are known to peak during the defervescing phase of dengue, and are associated with more severe disease.64 Specifically, the percentage of AL at presentation was associated with more severe thrombocytopaenia64, as well as haemorrhage, shock, and respiratory compromise.[65] It is reasonable to postulate that a higher AL count reflects greater immune dysregulation resulting in more severe thrombocytopaenia and other clinical manifestations.
T-cells play a significant role in the host response to dengue infection. Patients with severe dengue were shown to have an increase in markers of T-cell activation, as well as cytolytic granule proteins.[66] In contrast, other studies have shown that the inhibitory CTLA4 antigen was expressed at higher levels in T-cells from patients with dengue suggesting that T-cell responses against DENV maybe attenuated in some patients.[67]It is noteworthy that T-regulatory (T-reg) cells are increased in dengue infection, however they are not associated with adverse clinical outcomes and are of the naïve T reg subtype with limited suppressive capacity.[68] These data suggest a heterogeneity of T-cell subsets in dengue which maybe evolving as the disease progresses. Single-cell RNA sequencing (scRNAseq) at serial time points during dengue infection demonstrated unique effector T cell clusters expressing skin homing signature genes on day 1 of illness which was associated with defervescence.[69] Further studies using high dimensional profiling of T-cells in dengue are required to elucidate the key T-cell subsets involved in the progression to severe disease. Prospective studies looking at the correlation between T-cell subsets and clinical outcomes DENV would be of significant interest and may generate new hypotheses to be addressed through translational research.
While B-cells are important in generating the humoral response against viral infections, they can also be a target for infection by the DENV. [70] Indeed, virus-inclusive single-cell RNA-Seq (viscRNA-Seq) analysis demonstrated that naïve B-cells contained the highest amount of DENV RNA among the immune cells analysed in patients with severe dengue.[71]. The DENV has been proposed to bind to B-cells via CD300a and is internalised via clathrin mediated endocytosis.[72] Infection of B-cells by the DENV was shown to result in their proliferation as well as differentiation to a plasmablastic phenotype.[73] Indeed, skin and gut homing genes were upregulated in plasma cells and plasmablasts of dengue patients during the febrile period [74], which maybe a signature of entry into the critical phase. Furthermore, the Fcy receptors FcyRIIB and Fc like receptor LILRB1 expressed by B cells , are upregulated in dengue infection, and are implicated in ADE. [75] It is possible that the AL detected by automated haematology analysers represent a subset of B-cells which have been infected by DENV. This may explain their association with more severe disease. Future studies should focus on delineating the phenotype of DENV infected B-cells in clinical samples at serial time points to confirm this hypothesis.
Natural killer (NK) cells are a key part of the innate immune response against viral infections and are charactered by their cytotoxic function.[76] NK cells secrete cytolytic granules to eliminate dengue infected cells.[77] Costa et al demonstrated that in a murine model, IFN-y production by human NK cells was important in controlling DENV replication.[78] The significance of NK cells in the immune response against dengue has since been corroborated in clinical studies. The CD56 bright immunoregulatory subset of NK cells was found to be increased in patients with mild disease but notably not in those with severe dengue.[79] Similarly, the cytotoxic CD56 dim NK cell subset showed increased cytolytic capability ( based on the expression of CD69, NKP30,Granzyme B and IFNγ) in those with mild compared to severe dengue. [79] Importantly, the impaired cytotoxic capacity of NK cells in those with severe disease persisted even up to a week after infection.[79] Mass cytometry by time of flight (CyTOF) analysis was able to delineate unique NK cell signatures associated with dengue infection in paediatric and adult patients, respectively.[80] Similar studies evaluating NK cell subsets via high dimensional immune profiling at presentation and at serial time points during the disease would be of significant value, especially if NK cell phenotypes at presentation can predict severity of disease. The impact of dengue vaccines on T/NK cell subsets and their function is another important area to be addressed by future research.
Neutrophils
Neutropenia is a well-recognised feature of many viral infections including dengue.[81] The neutrophil nadir in dengue infection typically occurs on day 4-6 and may coincide with defervescence.[82] It is noteworthy that severe neutropenia is not associated with disease severity or mortality.[82] Although neutropenia is common in dengue, there is clear evidence of neutrophil activation during dengue infection as evidenced by elevated levels of IL-8 and TNF alpha.[83,84]
Neutrophils in dengue also over-express the activation marker CD66B, which potentiates endothelial adhesion and the generation of reactive oxygen species. [85] Patients with severe dengue also have increased neutrophil elastase levels, which implies that enhanced neutrophil activation can also be associated with severe disease. The generation of neutrophil extracellular traps (NETs) (web like chromatin structures which trap and destroy pathogens) has recently been demonstrated in dengue. [86] In addition to their antimicrobial functions, NETs are also proposed to have pro-inflammatory properties, and may exacerbate vascular permeability. Indeed, increased levels of NETs components were found in the serum of patients with severe dengue.
The occurrence of NETs in dengue is thought to be mediated by platelet extracellular vesicles which in turn promote signalling via C-type lectin domain containing 5 A (CLEC5A) and Toll Like Receptor 2 (TLR2)[87]. Dual blockade of CLEC5A and TLR2 reduced virus induced inflammation in a murine model.87 Collectively, these data support a prominent role for neutrophils in generating the pro inflammatory milieu associated with severe dengue. Future studies should address the neutrophil subsets involved in this process and how they interact with other immune cells as well as virus infected cells in driving the pathogenesis of severe dengue.
Mast Cells
Mast cells are a key component of the host response against parasitic infection[88], however their role in viral infections remains less clear. Mast cell activation, as measured by mast cell-derived mediators vascular endothelial growth factor (VEGF), tryptase, and chymase, has been implicated in the pathogenesis of dengue infection. However, the evidence related to the role of mast cell activation is mixed, both in mild and severe dengue.[89] Although elevated levels of tryptase are associated with severe dengue [90], further research is required to determine the true clinical relevance of Mast cells in dengue infection.
Eosinophils and Basophils
The role of eosinophils and basophils in the pathophysiology of dengue infection remains relatively under-studied. Low eosinophil counts seem to occur in the acute phase of dengue while eosinophilia has been reported later in the disease with a peak on days 9-10.[91] There have been no studies looking at whether eosinophil or basophil counts correlate with the severity of dengue. An important confounding factor to be considered in future studies is the possibility of concurrent parasitic infection (which can also cause an eosinophilia or basophilia) which maybe prevalent in the same populations susceptible to dengue.
Implications for Prognosis and Risk Stratification
Early identification of patients at risk of severe dengue infection is crucial to enable patients at risk to be offered closer monitoring and appropriate investigations. Given that dengue occurs predominantly in resource limited settings, this would also allow the prioritisation of scarce health care resources to those patients who need it most. Risk stratification for dengue is currently based on clinical parameters which often lack specificity. There are currently no validated laboratory investigations which can predict the severity of dengue infection. [92] There is however emerging data on haematologic and immunologic parameters which have potential utility in this space. [93]
A raised haematocrit has been proposed as a marker of haemoconcentration and hence a surrogate for vascular leakage in dengue infection. The WHO guideline on dengue incorporates a rising haematocrit of more than 20% as a marker of severity.[94] Indeed, It is noteworthy that a haematocrit value of greater than 40% has been associated with clinical capillary leak syndrome.[95,96] Other haematologic parameters which may have utility include the AL count which is predictive of thrombocytopaenia as well as other adverse clinical outcomes.[97] Accurately predicting the severity of thrombocytopaenia, as well as the timing of its recovery is also of great clinical value. The immature platelet fraction (IPF) is an index of thrombopoiesis which quantifies reticulated platelets that have recently been released from the bone marrow.[98] An IPF of more than 10% after defervescence predicts subsequent platelet recovery to a haemostatic level of more than 60 x 10^9/L within 72hours. [99] In a prospective study conducted among 240 patients, an IPF greater than 7.25% on day 3 of illness had a sensitivity of 88% for predicting platelet recovery to > 60 x10^9/L on day 8 of illness. Within 48hours after the peak IPF % was reached, platelet recovery was observed regardless of severity.[100]
The increased risk of adverse outcomes with secondary dengue infection is well documented. Indeed , there is a correlation between the serologic subtype of dengue and various haematologic parameters.[101] Recent studies have also demonstrated that patients who are triple positive for IgG, IgM and NS1 were more likely to develop severe dengue related thrombocytopaenia compared to those who were only NS1 positive.[102] Given the promising data supporting immuno-haematologic indices as predictors of outcome in dengue, future studies should evaluate the role of combining clinical with laboratory parameters to create composite predictive scores.
Implications for Management
There is no licensed antiviral treatment for dengue infection. The mainstay of management remains supportive care, including management of intravascular volume and bleeding.[103] Transfusions of platelets, fresh frozen plasma, and cryoprecipitate, can be lifesaving in selected patients with severe bleeding. The indication for prophylactic platelet transfusions in dengue infection however, remains uncertain and is not recommended at present. [104] The normal response to platelet transfusion is an increase in the platelet count at 10min to 1h post transfusion.[105] However, as dengue mediated thrombocytopaenia is at least partly immune mediated 105, transfused platelets are likely to be destroyed at an accelerated rate which leads to a suboptimal platelet increment.
It has been hypothesised that patients with severe thrombocytopaenia (i.e: platelet count less than 10 x 109/L) are more likely to have an immune mediated platelet destruction and are less likely to benefit from platelet transfusion.105 A single centre randomised controlled trial (RCT) demonstrated that although platelet transfusions increased platelet counts, they had no impact on bleeding related outcomes, and were associated with significant adverse effects.[106] Given the potential side effects of transfusion and their questionable benefit, therapies targeting the immune dysregulation in dengue as a means to treat dengue related thrombocytopaenia are likely to be a more worthwhile avenue for future research.
Steroids have been used for many years to address the dysregulated host immune response which causes much morbidity in dengue. The evidence supporting this practice is however weak. Although non-randomised trials of corticosteroids given for severe dengue showed possible benefits, there is insufficient evidence to justify the routine use of steroids. [107] A RCT in Vietnam comparing high dose (2mg/kg) and low dose (0.5mg/kg) prednisolone for 3 days in early dengue infection showed no reduction in the incidence of shock or other complications. [108] Both low dose and high dose dexamethasone were trialled in patients with dengue fever with thrombocytopaenia in a separate study where the control group demonstrated a higher rise in mean platelet counts on all 4 days of assessment, although bleeding manifestations were not significantly improved. [109] An important concern raised with regards to steroid use in dengue is the potential for immunosuppression leading to increased viral replication. [110] These findings were backed up by a recent systemic review which showed that there was insufficient evidence to justify the use of corticosteroids in DHF and DSS.[111]Larger RCTs are required to evaluate the role of steroids in dengue, given that there may be certain subgroups of patients who may benefit. These trials should incorporate translational correlatives to identify biomarkers which may be used as predictors of response to steroids.
It is noteworthy that conventional anti-viral therapies have not shown significant clinical benefit in dengue infection, with balapiravir, and celgosivir[112] among others having been evaluated in clinical trials. Given these findings, repurposing of anti-parasitic agents against dengue has been actively explored by several groups. Ivermectin inhibits DENV replication in vitro by targeting host nuclear import proteins that are required for nuclear localisation of the dengue NS5 protein.[113] A phase 3 RCT demonstrated faster clearance of NS1 antigen-aemia in the ivermectin treated arm, however there were no clear improvements in viraemia or clinical outcomes.[114] The anti-helminth agent Doxycycline was also shown to reduce viral replication in vitro by inhibiting NS2B-NS2 protease activity. [115] Indeed, an RCT of 231 patients demonstrated reduced mortality in the doxycycline treated arm compared to dengue patients treated with standard supportive care, suggesting the potential of doxycycline as a dengue therapeutic. In addition to repurposing of older drugs, several novel compounds targeting the DENV are in development.[116] JNJ -A07 for example, was identified via a cell-based screen and inhibits the interaction between the NS4B and NS3 proteins.[117,118] This molecule showed a favourable toxicity profile and induced rapid clearance of viraemia in murine models, suggesting potential for future evaluation in clinical trials. [118] The impact of these novel agents on the immune profile of dengue patients as well as the role of host immunity on the clinical responses should be studied in future clinical trials.
Conclusions and Future Directions
Our understanding of the immuno-haematologic manifestations of dengue infection has seen significant growth in the recent past. While the host immune response plays a pivotal role in the pathogenesis and progression of dengue, our understanding of the underlying mechanisms remains incomplete. A clear delineation of immune cell subsets driving severe dengue at each stage of disease, as well as the corresponding cytokine profile would be of critical importance. The role of the host gut microbiome in the progression of dengue is an active area of study, and while the interaction between gut flora and the host immune response is becoming well established[119], its impact on haematologic manifestations including bleeding remains unknown. Given that the gut microbiome is amenable to therapeutic intervention, this would be a key area for future investigations.
Further improvements in patient outcomes will require early prediction of severe disease as well as novel therapeutics targeting the immune response. Future trials of dengue viral therapeutics and vaccines should include correlative translational research, including high-dimensional immune profiling analyses. This will provide an integrated picture of the host immune system in relation to haematologic parameters in dengue infection and inform how best it can be modulated to improve clinical outcomes.
References
- Bhatt, S., Gething, P.W., Brady, O.J., Messina, J.P., Farlow, A.W., Moyes, C.L., Drake, J.M., Brownstein, J.S., Hoen, A.G., Sankoh, O., Myers, M.F., George, D.B., Jaenisch, T., Wint, G.R.W., Simmons, C.P., Scott, T.W., Farrar, J.J. and Hay, S.I. (2013). The global distribution and burden of dengue. Nature, 496(7446), pp.504–507. [CrossRef]
- Murugesan, A. and Manoharan, M. (2020). Dengue Virus. Emerging and Reemerging Viral Pathogens, pp.281–359. [CrossRef]
- Roy, S.K. and Bhattacharjee, S. (2021). Dengue virus: epidemiology, biology, and disease aetiology. Canadian Journal of Microbiology, 67(10), pp.687–702. [CrossRef]
- Shrestha DB, Budhathoki P, Gurung B, Subedi S, Aryal S, Basukala A, Aryal B, Adhikari A, Poudel A, Yadav GK, Khoury M, Rayamajhee B, Shrestha LB. Epidemiology of dengue in SAARC territory: a systematic review and meta-analysis. Parasit Vectors. 2022 Oct 24;15(1):389. [CrossRef] [PubMed] [PubMed Central]
- Kularatne, S.A. and Dalugama, C. (2022). Dengue infection: Global importance, immunopathology, and management. Clinical Medicine, 22(1), pp.9–13. [CrossRef]
- Organization, W.H. (2009). Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control. [online] Google Books. World Health Organization. Available at:https://books.google.com.sg/books?hl=en&lr=&id=dlc0YSIyGYwC&oi=fnd&pg=PP2&dq=World+Health+Organization+Dengue:+Guidelines+for+Diagnosis [Accessed 30 Dec. 2023].
- Pal, S., Dauner, A.L., Mitra, I., Forshey, B.M., Garcia, P., Morrison, A.C., Halsey, E.S., Kochel, T.J., and Wu, S.-J.L. (2014). Evaluation of Dengue NS1 Antigen Rapid Tests and ELISA Kits Using Clinical Samples. PLoS ONE, 9(11), p.e113411. [CrossRef]
- Bandyopadhyay, S., Lum, L.C.S. and Kroeger, A. (2006). Classifying dengue: a review of the difficulties in using the WHO case classification for dengue haemorrhagic fever. Tropical Medicine and International Health, 11(8), pp.1238–1255. [CrossRef]
- Tayal, A., Kabra, S.K. and Lodha, R. (2022). Management of Dengue: An Updated Review. Indian Journal of Pediatrics, 90(2). [CrossRef]
- Salles T.S., da Encarnação Sá-Guimarães T., de Alvarenga E., Guimarães-Ribeiro V., de Meneses M., de Castro-Salles P.F., et al. History, epidemiology and diagnostics of dengue in the American and Brazilian contexts: a review. Parasit Vectors. 2018;11(1):264. [CrossRef]
- Khanam A, Gutiérrez-Barbosa H, Lyke KE, Chua JV. Immune-Mediated Pathogenesis in Dengue Virus Infection. Viruses. 2022 Nov 21;14(11):2575. [CrossRef] [PubMed]
- Malavige, G.N., Jeewandara, C. and Ogg, G.S. (2020). Dysfunctional Innate Immune Responses and Severe Dengue. Frontiers in Cellular and Infection Microbiology, [online] 10, p.590004. [CrossRef]
- Adane, T. and Getawa, S. (2021). Coagulation abnormalities in Dengue fever infection: A systematic review and meta-analysis. PLOS Neglected Tropical Diseases, 15(8), p.e0009666. [CrossRef]
- Patel, Govind R.1; Thanvi, Indu2; Nadeem, Mohammad2; Kanwaria, Rahul2. Coagulation abnormalities and their relationship with bleeding manifestations in patients with dengue-A single center observational study. Asian Pacific Journal of Tropical Medicine 16(2):p 65-71, February 2023.
- John, K.J., Gunasekaran, K., Prasad, J.D., Mathew, D., Das, S., Sultan, N., Abraham, A.M. and Iyyadurai, R. (2019). Predictors of Major Bleeding and Mortality in Dengue Infection: A Retrospective Observational Study in a Tertiary Care Centre in South India. Interdisciplinary Perspectives on Infectious Diseases, 2019, pp.1–7. [CrossRef]
- Ruttmann, T. (2006). Coagulation for the clinician. South African Journal of Surgery. Suid-Afrikaanse Tydskrif Vir Chirurgie, [online] 44(1), pp.22, 24–26, 28–30; passim. Available at: https://pubmed.ncbi.nlm.nih.gov/16619987/.
- Lin SW, Chuang YC, Lin YS, Lei HY, Liu HS, Yeh TM. Dengue virus nonstructural protein NS1 binds to prothrombin/thrombin and inhibits prothrombin activation. J Infect. 2012 Mar;64(3):325-34. [CrossRef] [PubMed]
- Isarangkura PB, Pongpanich B, Pintadit P, Phanichyakarn P, Valyasevi A. Hemostatic derangement in dengue haemorrhagic fever. Southeast Asian J Trop Med Public Health. 1987 Sep;18(3):331-9. [PubMed]
- Teerasarntipan, T., Chaiteerakij, R., Komolmit, P., Tangkijvanich, P. and Treeprasertsuk, S. (2020). Acute liver failure and death predictors in patients with dengue-induced severe hepatitis. World Journal of Gastroenterology, 26(33), pp.4983–4995. [CrossRef]
- Azeredo, E.L. de, Monteiro, R.Q. and de-Oliveira Pinto, L.M. (2015). Thrombocytopenia in Dengue: Interrelationship between Virus and the Imbalance between Coagulation and Fibrinolysis and Inflammatory Mediators. Mediators of Inflammation, [online] 2015, pp.1–16. [CrossRef]
- Factors contributing to the disturbance of coagulation and fibrinolysis in dengue virus infection. (2013). Journal of the Formosan Medical Association, [online] 112(1), pp.12–17. [CrossRef]
- Azeredo, E.L. de, Monteiro, R.Q. and de-Oliveira Pinto, L.M. (2015). Thrombocytopenia in Dengue: Interrelationship between Virus and the Imbalance between Coagulation and Fibrinolysis and Inflammatory Mediators. Mediators of Inflammation, [online] 2015, pp.1–16. [CrossRef]
- Huang, Y.-H., Lei, H.-Y., Liu, H.-S., Lin, Y.-S., Chen, S.-H., Liu, C.-C. and Yeh, T.-M. (2003). Tissue plasminogen activator induced by dengue virus infection of human endothelial cells. Journal of Medical Virology, 70(4), pp.610–616. [CrossRef]
- Azeredo, E.L. de, Monteiro, R.Q. and de-Oliveira Pinto, L.M. (2015). Thrombocytopenia in Dengue: Interrelationship between Virus and the Imbalance between Coagulation and Fibrinolysis and Inflammatory Mediators. Mediators of Inflammation, [online] 2015, pp.1–16. [CrossRef]
- Sosothikul, D., Seksarn, P., Pongsewalak, S., Thisyakorn, U. and Lusher, J. (2007). Activation of endothelial cells, coagulation, and fibrinolysis in children with Dengue virus infection. Thrombosis and Haemostasis, [online] 97(4), pp.627–634. Available at: https://pubmed.ncbi.nlm.nih.gov/17393026/ [Accessed 29 Dec. 2023].
- Nurnaningsih, null, Sunbanu, S.E., Rusmawatiningtyas, D., Arguni, E., Makrufardi, F. and Kumara, I.F. (2022). Disseminated intravascular coagulation initial score as a predictor of mortality in children with dengue shock syndrome: A retrospective cohort study. Annals of Medicine and Surgery (2012), [online] 79, p.103890. [CrossRef]
- Factors contributing to the disturbance of coagulation and fibrinolysis in dengue virus infection. (2013). Journal of the Formosan Medical Association, [online] 112(1), pp.12–17. [CrossRef]
- Castilho, B.M., Silva, M.T., Freitas, A.R.R., Fulone, I. and Lopes, L.C. (2020). Factors associated with thrombocytopenia in patients with dengue fever: a retrospective cohort study. BMJ Open, [online] 10(9), p.e035120. [CrossRef]
- Ojha, A., Nandi, D., Batra, H., Singhal, R., Annarapu, G.K., Bhattacharyya, S., Seth, T., Dar, L., Medigeshi, G.R., Vrati, S., Vikram, N.K. and Guchhait, P. (2017). Platelet activation determines the severity of thrombocytopenia in dengue infection. Scientific Reports, [online] 7(1). [CrossRef]
- Lam, P.K., Ngoc, T.V., Thu Thuy, T.T., Hong Van, N.T., Nhu Thuy, T.T., Hoai Tam, D.T., Dung, N.M., Hanh Tien, N.T., Thanh Kieu, N.T., Simmons, C., Wills, B. and Wolbers, M. (2017). The value of daily platelet counts for predicting dengue shock syndrome: Results from a prospective observational study of 2301 Vietnamese children with dengue. PLOS Neglected Tropical Diseases, 11(4), p.e0005498. [CrossRef]
- SAITO, M., OISHI, K., INOUE, S., DIMAANO, E.M., ALERA, M.T.P., ROBLES, A.M.P., ESTRELLA, B.D., KUMATORI, A., MOJI, K., ALONZO, M.T., BUERANO, C.C., MATIAS, R.R., MORITA, K., NATIVIDAD, F.F. and NAGATAKE, T. (2004). Association of increased platelet-associated immunoglobulins with thrombocytopenia and the severity of disease in secondary dengue virus infections. Clinical and Experimental Immunology, 138(2), pp.299–303. [CrossRef]
- Bridget A. Wills, Emmanuelle E. Oragui, Alick C. Stephens, Olufunmilayo A. Daramola, Nguyen Minh Dung, Ha Thi Loan, Nguyen Vinh Chau, Mary Chambers, Kasia Stepniewska, Jeremy J. Farrar, Michael Levin, Coagulation Abnormalities in Dengue Hemorrhagic Fever: Serial Investigations in 167 Vietnamese Children with Dengue Shock Syndrome, Clinical Infectious Diseases, Volume 35, Issue 3, 1 August 2002, Pages 277–285. [CrossRef]
- Pan, P., Zhang, Q., Liu, W., Wang, W., Yu, Z., Lao, Z., Zhang, W., Shen, M., Wan, P., Xiao, F., Shereen, M.A., Zhang, W., Tan, Q., Liu, Y., Liu, X., Wu, K., Liu, Y., Li, G. and Wu, J. (2019). Dengue Virus Infection Activates Interleukin-1β to Induce Tissue Injury and Vascular Leakage. Frontiers in Microbiology, [online] 10. [CrossRef]
- Gomes de Azevedo-Quintanilha I, Campos MM, Teixeira Monteiro AP, Dantas do Nascimento A, Calheiros AS, Oliveira DM, Dias SSG, Soares VC, Santos JDC, Tavares I, Lopes Souza TM, Hottz ED, Bozza FA, Bozza PT. Increased platelet activation and platelet-inflammasome engagement during chikungunya infection. Front Immunol. 2022 Sep 15;13:958820. [CrossRef]
- Ojha, A., Nandi, D., Batra, H., Singhal, R., Annarapu, G.K., Bhattacharyya, S., Seth, T., Dar, L., Medigeshi, G.R., Vrati, S., Vikram, N.K. and Guchhait, P. (2017). Platelet activation determines the severity of thrombocytopenia in dengue infection. Scientific Reports, [online] 7(1). [CrossRef]
- La Russa, V.F. and Innis, B.L. (1995). Mechanisms of dengue virus-induced bone marrow suppression. Baillière’s Clinical Haematology, 8(1), pp.249–270. [CrossRef]
- La Russa, V.F. and Innis, B.L. (1995). 11 Mechanisms of dengue virus-induced bone marrow suppression. Baillière’s Clinical Haematology, 8(1), pp.249–270. [CrossRef]
- Abeysuriya V, Seneviratne SL, de Mel P, Clarice CSH, de Mel C, Chandrasena L, Yip C, Yap ES, de Mel S. The immature platelet fraction, a predictive tool for early recovery from dengue-related thrombocytopenia: a prospective study. Trans R Soc Trop Med Hyg. 2022 May 2;116(5):424-432.
- de Mel, S., Thilakawardana, B.U., de Mel, P., Clarice, C.S.H., Shalindi, M., de Mel, C., Chandrasena, L., Yip, C., Yap, E.-S., Seneviratne, S.L. and Abeysuriya, V. (2020). Triple positivity for nonstructural antigen 1, immunoglobulin M and immunoglobulin G is predictive of severe thrombocytopaenia related to dengue infection. Journal of Clinical Virology, 129, p.104509. https://doi.org/10.1016/j.jcv.2020.104509. 1 Rai, Dr.A., Azad, Dr.S., Nautiyal, Dr.S. and Acharya, Dr.S. (2019). Correlation between hematological and serological parameters in dengue patients- an analysis of 2022 cases. Tropical Journal of Pathology and Microbiology, 5(8), pp.547–554. [CrossRef]
- Abeysuriya, V., Choong, C.S.H., Thilakawardana, B.U., de Mel, P., Shalindi, M., de Mel, C., Chandrasena, L., Seneviratne, S.L., Yip, C., Yap, E.-S. and de Mel, S. (2020). The atypical lymphocyte count: a novel predictive factor for severe thrombocytopenia related to dengue. Transactions of The Royal Society of Tropical Medicine and Hygiene, 114(6), pp.424–432. [CrossRef]
- Donaldson CD, De Mel S, Clarice CSH, Thilakawardana BU, De Mel P, Shalindi M, et al. Admission ultrasonography as a predictive tool for thrombocytopenia and disease severity in dengue infection. Trans R Soc Trop Med Hyg. 2021;115: 1396–1402. [PubMed]
- de Mel, S., Thilakawardana, B.U., de Mel, P., Clarice, C.S.H., Shalindi, M., de Mel, C., Chandrasena, L., Yip, C., Yap, E.-S., Seneviratne, S.L. and Abeysuriya, V. (2020). Triple positivity for nonstructural antigen 1, immunoglobulin M and immunoglobulin G is predictive of severe thrombocytopaenia related to dengue infection. Journal of Clinical Virology, 129, p.104509. [CrossRef]
- Sun P, Bauza K, Pal S, Liang Z, Wu SJ, Beckett C, Burgess T, Porter K. Infection and activation of human peripheral blood monocytes by dengue viruses through the mechanism of antibody-dependent enhancement. Virology. 2011 Dec 20;421(2):245-52. https://doi.org/10.1016/j.virol.2011.08.026. Epub 2011 Oct 26. [CrossRef] [PubMed]
- The monocyte-macrophage-mast cell axis in dengue pathogenesis - Journal of Biomedical Science. Wan et al.https://jbiomedsci.biomedcentral.com/articles/10.1186/s12929-018-0482-9.
- Durbin AP, Vargas MJ, Wanionek K, Hammond SN, Gordon A, Rocha C, Balmaseda A, Harris E. Phenotyping of peripheral blood mononuclear cells during acute dengue illness demonstrates infection and increased activation of monocytes in severe cases compared to classic dengue fever. Virology. 2008 Jul 5;376(2):429-35. [CrossRef] [PubMed]
- Kwissa M, Nakaya HI, Onlamoon N, Wrammert J, Villinger F, Perng GC, Yoksan S, Pattanapanyasat K, Chokephaibulkit K, Ahmed R, Pulendran B. Dengue virus infection induces expansion of a CD14(+)CD16(+) monocyte population that stimulates plasmablast differentiation. Cell Host Microbe. 2014 Jul 9;16(1):115-27. [CrossRef] [PubMed]
- Singla, M., Kar, M., Sethi, T., Kabra, S.K., Lodha, R., Chandele, A. and Medigeshi, G.R. (2016). Immune Response to Dengue Virus Infection in Pediatric Patients in New Delhi, India—Association of Viremia, Inflammatory Mediators and Monocytes with Disease Severity. PLOS Neglected Tropical Diseases, 10(3), p.e0004497. [CrossRef]
- Tsai, T.-T., Chuang, Y.-J., Lin, Y.-S., Chang, C.-P., Wan, S.-W., Lin, S.-H., Chen, C.-L. and Lin, C.-F. (2014). Antibody-Dependent Enhancement Infection Facilitates Dengue Virus-Regulated Signaling of IL-10 Production in Monocytes. PLoS Neglected Tropical Diseases, 8(11), p.e3320. [CrossRef]
- Malavige, G.N., Jeewandara, C. and Ogg, G.S. (2020). Dysfunctional Innate Immune Responses and Severe Dengue. Frontiers in Cellular and Infection Microbiology, [online] 10, p.590004. [CrossRef]
- Jessie K, Fong MY, Devi S, Lam SK, Wong KT. Localization of dengue virus in naturally infected human tissues, by immunohistochemistry and in situ hybridization. J Infect Dis. 2004 Apr 15;189(8):1411-8. [CrossRef] [PubMed]
- Chen HC, Hofman FM, Kung JT, Lin YD, Wu-Hsieh BA. Both virus and tumor necrosis factor alpha are critical for endothelium damage in a mouse model of dengue virus-induced hemorrhage. J Virol. 2007 Jun;81(11):5518-26. [CrossRef] [PubMed]
- Wan, SW., Wu-Hsieh, B.A., Lin, YS. et al. The monocyte-macrophage-mast cell axis in dengue pathogenesis. J Biomed Sci 25, 77 (2018). [CrossRef]
- Kan FK, Tan CC, von Bahr Greenwood T, et al. Dengue Infection Complicated by Hemophagocytic Lymphohistiocytosis: Experiences From 180 Patients With Severe Dengue. Clin Infect Dis. 2020;70(11):2247-2255.
- Hasliana Azrah Ab-Rahman, Wong, P.-F., Rahim, H., Juraina Abd-Jamil, Kian Lam Tan, Sulaiman, S., Lum, C.-S., Syarifah-Faridah Syed-Omar and AbuBakar, S. (2015). Dengue death with evidence of hemophagocytic syndrome and dengue virus infection in the bone marrow. SpringerPlus, 4(1). [CrossRef]
- Kan, F.K., Tan, C.C., Von Bahr Greenwood, T., Khalid, K.E., Supramaniam, P., Hed Myrberg, I., Tan, L.H. and Henter, J.-I. (2020). Dengue Infection Complicated by Hemophagocytic Lymphohistiocytosis: Experiences From 180 Patients With Severe Dengue. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America, [online] 70(11), pp.2247–2255. [CrossRef]
- Ahmad, R., Suzilah, I., Wan Najdah, W.M.A., Topek, O., Mustafakamal, I. and Lee, H.L. (2018). Factors determining dengue outbreak in Malaysia. PLOS ONE, 13(2), p.e0193326. [CrossRef]
- Kurane, I., Innis, B.L., Nimmannitya, S., Nisalak, A., Meager, A., Janus, J., and Ennis, F.A. (1991). Activation of T lymphocytes in dengue virus infections. High levels of soluble interleukin 2 receptor, soluble CD4, soluble CD8, interleukin 2, and interferon-gamma in sera of children with dengue. Journal of Clinical Investigation, 88(5), pp.1473–1480. [CrossRef]
- 3 Salvatory Kalabamu, F. and Maliki, S. (2021). Use of Haematological Changes as a Predictor of Dengue Infection among Suspected Cases at Kairuki Hospital in Dar Es Salaam, Tanzania: A Retrospective Cross Sectional Study. East African Health Research Journal, 5(1), pp.91–98. [CrossRef]
- Ananda Rao, A., U, R.R., Gosavi, S. and Menon, S. (2020). Dengue Fever: Prognostic Insights From a Complete Blood Count. Cureus. [CrossRef]
- Salvatory Kalabamu, F. and Maliki, S. (2021). Use of Haematological Changes as a Predictor of Dengue Infection among Suspected Cases at Kairuki Hospital in Dar Es Salaam, Tanzania: A Retrospective Cross Sectional Study. East African Health Research Journal, 5(1), pp.91–98. [CrossRef]
- Tsai, J.-J., Liu, L.-T., Chang, K., Wang, S.-H., Hsiao, H.-M., Clark, K.B. and Perng, G.C. (2011). The importance of hematopoietic progenitor cells in dengue. Therapeutic Advances in Hematology, 3(1), pp.59–71. [CrossRef]
- Thisyakorn, U., Nimmannitya, S., Ningsanond, V. and Soogarun, S. (1984). Atypical lymphocyte in dengue hemorrhagic fever: its value in diagnosis. The Southeast Asian Journal of Tropical Medicine and Public Health, [online] 15(1), pp.32–36. Available at: https://pubmed.ncbi.nlm.nih.gov/6740378/ [Accessed 29 Dec. 2023].
- Clarice, C.S.H., Abeysuriya, V., de Mel, S., Uvindu Thilakawardana, B., de Mel, P., de Mel, C., Chandrasena, L., Seneviratne, S.L., Yip, C., and Yap, E.S. (2019). Atypical lymphocyte count correlates with the severity of dengue infection. PLOS ONE, 14(5), p.e0215061. [CrossRef]
- Chaloemwong, J., Tantiworawit, A., Rattanathammethee, T., Hantrakool, S., Chai-Adisaksopha, C., Rattarittamrong, E. and Norasetthada, L. (2018). Useful clinical features and hematological parameters for the diagnosis of dengue infection in patients with acute febrile illness: a retrospective study. BMC Hematology, 18(1). [CrossRef]
- Abeysuriya, V., Choong, C.S.H., Thilakawardana, B.U., de Mel, P., Shalindi, M., de Mel, C., Chandrasena, L., Seneviratne, S.L., Yip, C., Yap, E.-S. and de Mel, S. (2020). The atypical lymphocyte count: a novel predictive factor for severe thrombocytopenia related to dengue. Transactions of The Royal Society of Tropical Medicine and Hygiene, 114(6), pp.424–432. [CrossRef]
- Azeredo, E.L., Zagne, S.M., Alvarenga, A.R., Nogueira, R.M., Kubelka, C.F., and Oliveira-Pinto, L.M. de (2006). Activated peripheral lymphocytes with increased expression of cell adhesion molecules and cytotoxic markers are associated with dengue fever disease. Memórias do Instituto Oswaldo Cruz, 101(4), pp.437–449. [CrossRef]
- Adikari, T.N., Kamaladasa, A., Fernando, R.H., Fernando, S.M., Perera, T.M.K., Gomes, L., Jayaratne, S., Ogg, G.S. and Malavige, G.N. (2014). High CTLA-4 expression in T cells in patients with acute dengue infection. International Journal of Infectious Diseases, 21, p.330. [CrossRef]
- Jayaratne, H.E., Wijeratne, D., Fernando, S., Kamaladasa, A., Gomes, L., Wijewickrama, A., Ogg, G.S. and Malavige, G.N. (2017). Regulatory T-cells in acute dengue viral infection. Immunology, 154(1), pp.89–97. [CrossRef]
- Jantarika Kumar Arora, Anunya Opasawatchai, Tiraput Poonpanichakul, Natnicha Jiravejchakul, Waradon Sungnak, Anavaj Sakuntabhai, Pratap Singhasivanon, Swangjit Suraamornkul, Tawatchai Yingtaweesak, Khajohnpong Manopwisedjaroen, Nada Pitabut, Oranart Matangkasombut, Sarah A. Teichmann, Ponpan Matangkasombut, Varodom Charoensawan, Single-cell temporal analysis of natural dengue infection reveals skin-homing lymphocyte expansion one day before defervescence, iScience,Volume 25, Issue 4,2022,104034,ISSN 2589-0042. [CrossRef]
- Correa, A.R.V., Berbel, A.C.E.R., Papa, M.P., Morais, A.T.S. de, Peçanha, L.M.T. and Arruda, L.B. de (2015). Dengue Virus Directly Stimulates Polyclonal B Cell Activation. PLOS ONE, 10(12), p.e0143391. [CrossRef]
- Zanini, F., Robinson, M.L., Croote, D., Sahoo, M.K., Sanz, A.M., Ortiz-Lasso, E., Albornoz, L.L., Rosso, F., Montoya, J.G., Goo, L., Pinsky, B.A., Quake, S.R. and Einav, S. (2018). Virus-inclusive single-cell RNA sequencing reveals the molecular signature of progression to severe dengue. Proceedings of the National Academy of Sciences, [online] 115(52), pp.E12363–E12369. [CrossRef]
- Carnec, X., Meertens, L., Dejarnac, O., Perera-Lecoin, M., Hafirassou, M.L., Kitaura, J., Ramdasi, R., Schwartz, O. and Amara, A. (2015). The Phosphatidylserine and Phosphatidylethanolamine Receptor CD300a Binds Dengue Virus and Enhances Infection. Journal of Virology, 90(1), pp.92–102. [CrossRef]
- Vinit Upasani, Thi, H., Auerswald, H., Laurent, D., Heng Sothy, Duong, V., Rodenhuis-Zybert, I.A., Philippe Dussart and Tineke Cantaert (2021). Direct Infection of B Cells by Dengue Virus Modulates B Cell Responses in a Cambodian Pediatric Cohort. Frontiers in Immunology, 11. [CrossRef]
- Arora, J.K., Opasawatchai, A., Poonpanichakul, T., Jiravejchakul, N., Sungnak, W., Sakuntabhai, A., Singhasivanon, P., Suraamornkul, S., Yingtaweesak, T., Manopwisedjaroen, K., Pitabut, N., Matangkasombut, O., Teichmann, S.A., Matangkasombut, P. and Charoensawan, V. (2022). Single-cell temporal analysis of natural dengue infection reveals skin-homing lymphocyte expansion one day before defervescence. iScience, [online] 25(4). [CrossRef]
- Upasani, V., Vo, H.T.M., Ung, S., Heng, S., Laurent, D., Choeung, R., Duong, V., Sorn, S., Ly, S., Rodenhuis-Zybert, I.A., Dussart, P. and Cantaert, T. (2019). Impaired Antibody-Independent Immune Response of B Cells in Patients With Acute Dengue Infection. Frontiers in Immunology, [online] 10, p.2500. [CrossRef]
- Vivier, E., Tomasello, E., Baratin, M., Walzer, T. and Ugolini, S. (2008). Functions of natural killer cells. Nature Immunology, 9(5), pp.503–510. [CrossRef]
- Mathew, A. (2018). Defining the role of NK cells during dengue virus infection. Immunology, 154(4), pp.557–562. [CrossRef]
- Costa, V.V., Ye, W., Chen, Q., Teixeira, M.M., Preiser, P., Ooi, E.E. and Chen, J. (2017). Dengue Virus-Infected Dendritic Cells, but Not Monocytes, Activate Natural Killer Cells through a Contact-Dependent Mechanism Involving Adhesion Molecules. mBio, 8(4). [CrossRef]
- Snehal Shabrish, Karnik, N.D., Gupta, V., Priya Bhate and Manisha Madkaikar (2020). Impaired NK cell activation during acute dengue virus infection: A contributing factor to disease severity. Heliyon. [CrossRef]
- McKechnie, J.L., Beltrán, D., Ferreira, A.-M.M., Vergara, R., Saenz, L., Vergara, O., Estripeaut, D., Araúz, A.B., Simpson, L.J., Holmes, S., López-Vergès, S. and Blish, C.A. (2020). Mass Cytometry Analysis of the NK Cell Receptor–Ligand Repertoire Reveals Unique Differences between Dengue-Infected Children and Adults. ImmunoHorizons, 4(10), pp.634–647. [CrossRef]
- Muralidharan, A. and Reid, S.P. (2021). Complex Roles of Neutrophils during Arboviral Infections. Cells, 10(6), p.1324. [CrossRef]
- Thein, T.-L., Wong, J.G.X., Lye, D.C., Hao, Y., Wilder-Smith, A. and Leo, Y.-S. (2014). Severe Neutropenia in Dengue Patients: Prevalence and Significance. The American Journal of Tropical Medicine and Hygiene, [online] 90(6), pp.984–987. [CrossRef]
- Wang SSY, Chng WJ, Liu H, de Mel S. Tumor-Associated Macrophages and Related Myelomonocytic Cells in the Tumor Microenvironment of Multiple Myeloma. Cancers (Basel). 2022 Nov 17;14(22):5654. https://doi.org/10.3390/cancers14225654. [CrossRef] [PubMed]
- Screaton, G., Mongkolsapaya, J., Yacoub, S. and Roberts, C. (2015). New insights into the immunopathology and control of dengue virus infection. Nature Reviews Immunology, [online] 15(12), pp.745–759. [CrossRef]
- Jenne, Craig N., Wong, Connie H.Y., Zemp, Franz J., McDonald, B., Rahman, Masmudur M., Forsyth, Peter A., McFadden, G. and Kubes, P. (2013). Neutrophils Recruited to Sites of Infection Protect from Virus Challenge by Releasing Neutrophil Extracellular Traps. Cell Host & Microbe, 13(2), pp.169–180. [CrossRef]
- Jenne, Craig N., Wong, Connie H.Y., Zemp, Franz J., McDonald, B., Rahman, Masmudur M., Forsyth, Peter A., McFadden, G. and Kubes, P. (2013). Neutrophils Recruited to Sites of Infection Protect from Virus Challenge by Releasing Neutrophil Extracellular Traps. Cell Host & Microbe, 13(2), pp.169–180. [CrossRef]
- Sung, P.-S., Huang, T.-F. and Hsieh, S.-L. (2019). Extracellular vesicles from CLEC2-activated platelets enhance dengue virus-induced lethality via CLEC5A/TLR2. Nature Communications, [online] 10(1), pp.1–13. [CrossRef]
- Rathore, A.P.S., Mantri, C.K., Aman, S.A.B., Syenina, A., Ooi, J., Jagaraj, C.J., Goh, C.C., Tissera, H., Wilder-Smith, A., Ng, L.G., Gubler, D.J. and St. John, A.L. (2019). Dengue virus–elicited tryptase induces endothelial permeability and shock. Journal of Clinical Investigation, 129(10), pp.4180–4193. [CrossRef]
- Sherif, N.A., Zayan, A.H., Elkady, A.H., Ghozy, S., Ahmed, A.R., Omran, E.S., Taha, E.A., Eldesoky, E.A., Ebied, A., Tieu, T., Maraie, N., Kamel, M.G., Ngo, H.T., Mattar, O.M., Hirayama, K. and Huy, N.T. (2019). Mast cell mediators in relation to dengue severity: A systematic review and meta-analysis. Reviews in Medical Virology, 30(1). [CrossRef]
- Rathore, A.P.S., Mantri, C.K., Aman, S.A.B., Syenina, A., Ooi, J., Jagaraj, C.J., Goh, C.C., Tissera, H., Wilder-Smith, A., Ng, L.G., Gubler, D.J. and St. John, A.L. (2019). Dengue virus–elicited tryptase induces endothelial permeability and shock. Journal of Clinical Investigation, 129(10), pp.4180–4193. [CrossRef]
- Fujimoto, D.E. and Koifman, S. (2014). Clinical and laboratory characteristics of patients with dengue hemorrhagic fever manifestations and their transfusion profile. Revista Brasileira de Hematologia e Hemoterapia, 36(2), pp.115–120. [CrossRef]
- Clarice, C.S.H., Abeysuriya, V., de Mel, S., Uvindu Thilakawardana, B., de Mel, P., de Mel, C., Chandrasena, L., Seneviratne, S.L., Yip, C., and Yap, E.S. (2019). Atypical lymphocyte count correlates with the severity of dengue infection. PLOS ONE, 14(5), p.e0215061. [CrossRef]
- Moallemi S, Lloyd AR, Rodrigo C. Early biomarkers for prediction of severe manifestations of dengue fever: a systematic review and a meta-analysis. Sci Rep. 2023 Oct 14;13(1):17485. [CrossRef] [PubMed]
- Comprehensive Guidelines for Prevention and Control of Dengue and Dengue Hemorrhagic Fever | GHDx. [online] Available at: https://ghdx.healthdata.org/record/comprehensive-guidelines-prevention-and-control-dengue-and-dengue-hemorrhagic-fever [Accessed 29 Dec. 2023].
- Balasubramanian, S., Anandnathan, K., Shivbalan, So., Datta, M. and Amalraj, E. (2004). Cut-off Hematocrit Value for Hemoconcentration in Dengue Hemorrhagic Fever. Journal of Tropical Pediatrics, 50(2), pp.123–124. [CrossRef]
- Balasubramanian, S., Anandnathan, K., Shivbalan, So., Datta, M. and Amalraj, E. (2004). Cut-off Hematocrit Value for Hemoconcentration in Dengue Hemorrhagic Fever. Journal of Tropical Pediatrics, 50(2), pp.123–124. [CrossRef]
- Clarice, C.S.H., Abeysuriya, V., de Mel, S., Uvindu Thilakawardana, B., de Mel, P., de Mel, C., Chandrasena, L., Seneviratne, S.L., Yip, C., and Yap, E.S. (2019). Atypical lymphocyte count correlates with the severity of dengue infection. PLOS ONE, 14(5), p.e0215061. [CrossRef]
- Looi, K.W., Matsui, Y., Kono, M., Samudi, C., Kojima, N., Ong, J.X., Tan, C.A., Ang, C.S., Tan, P.H.Y., Shamnugam, H., Sekaran, S.D., Syed Omar, S.F. and Lum, L.C.S. (2021). Evaluation of immature platelet fraction as a marker of dengue fever progression. International Journal of Infectious Diseases, 110, pp.187–194. [CrossRef]
- Chuansumrit, A., Apiwattanakul, N., Sirachainan, N., Paisooksantivatana, K., Athipongarporn, A., Tangbubpha, N., Kadegasem, P., Tangnararatchakit, K. and Yoksan, S. (2019). The use of immature platelet fraction to predict time to platelet recovery in patients with dengue infection. Paediatrics and International Child Health, 40(2), pp.124–128. [CrossRef]
- Abeysuriya, V., Seneviratne, S.L., de Mel, P., Clarice, C.S.H., de Mel, C., Chandrasena, L., Yip, C., Yap, E.-S. and de Mel, S. (2021). The immature platelet fraction, a predictive tool for early recovery from dengue-related thrombocytopenia: a prospective study. Transactions of The Royal Society of Tropical Medicine and Hygiene, 116(5), pp.424–432. [CrossRef]
- Rai, Dr.A., Azad, Dr.S., Nautiyal, Dr.S. and Acharya, Dr.S. (2019). Correlation between hematological and serological parameters in dengue patients- an analysis of 2022 cases. Tropical Journal of Pathology and Microbiology, 5(8), pp.547–554. [CrossRef]
- de Mel, S., Thilakawardana, B.U., de Mel, P., Clarice, C.S.H., Shalindi, M., de Mel, C., Chandrasena, L., Yip, C., Yap, E.-S., Seneviratne, S.L. and Abeysuriya, V. (2020). Triple positivity for nonstructural antigen 1, immunoglobulin M and immunoglobulin G is predictive of severe thrombocytopaenia related to dengue infection. Journal of Clinical Virology, 129, p.104509. [CrossRef]
- World Health Organisation (2009). Dengue guidelines, for diagnosis, treatment, prevention, and control. [online] www.who.int. Available at: https://www.who.int/publications/i/item/9789241547871.
- Lye, D.C., Archuleta, S., Syed-Omar, S.F., Low, J.G., Oh, H.M., Wei, Y., Fisher, D., Ponnampalavanar, S.S.L., Wijaya, L., Lee, L.K., Ooi, E.-E., Kamarulzaman, A., Lum, L.C., Tambyah, P.A. and Leo, Y.-S. (2017). Prophylactic platelet transfusion plus supportive care versus supportive care alone in adults with dengue and thrombocytopenia: a multicentre, open label, randomised, superiority trial. The Lancet, 389(10079), pp.1611–1618. [CrossRef]
- Assir, M.Z.K., Kamran, U., Ahmad, H.I., Bashir, S., Mansoor, H., Anees, S.B. and Akram, J. (2013). Effectiveness of Platelet Transfusion in Dengue Fever: A Randomized Controlled Trial. Transfusion Medicine and Hemotherapy, 40(5), pp.362–368. [CrossRef]
- Carr, J.M., Kruskall, M.S., Kaye, J.A., and Robinson, S.H. (1986). Efficacy of platelet transfusions in immune thrombocytopenia. The American Journal of Medicine, [online] 80(6), pp.1051–1054. [CrossRef]
- de Mel, S., Thilakawardana, B.U., de Mel, P., de Silva, A.P., de Mel, C., Chandrasena, L., Seneviratne, S.L. and Abeysuriya, V. (2020). The impact of empirical hydrocortisone therapy on clinical outcomes in dengue fever: a retrospective chart review. Transactions of The Royal Society of Tropical Medicine and Hygiene, 114(8), pp.632–634. [CrossRef]
- Tam, D.T.H., Ngoc, T.V., Tien, N.T.H., Kieu, N.T.T., Thuy, T.T.T., Thanh, L.T.C., Tam, C.T., Truong, N.T., Dung, N.T., Qui, P.T., Hien, T.T., Farrar, J.J., Simmons, C.P., Wolbers, M. and Wills, B.A. (2012). Effects of short-course oral corticosteroid therapy in early dengue infection in Vietnamese patients: a randomized, placebo-controlled trial. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America, [online] 55(9), pp.1216–1224. [CrossRef]
- Kc, S., Ka, M., Gowdappa Hb and A Bhograj (2013). Effect of High Dose of Steroid on Plateletcount in Acute Stage of Dengue Fever with Thrombocytopenia. Journal of Clinical and Diagnostic Research. [CrossRef]
- Kularatne, S. a. M., Walathara, C., Mahindawansa, S.I., Wijesinghe, S., Pathirage, M.M.K., Kumarasiri, P.V.R. and Dissanayake, A.M.S.D.M. (2009). Efficacy of low dose dexamethasone in severe thrombocytopenia caused by dengue fever: a placebo-controlled study. Postgraduate Medical Journal, [online] 85(1008), pp.525–529. [CrossRef]
- Panpanich, R., Sornchai, P. and Kanjanaratanakorn, K. (2006). Corticosteroids for treating dengue shock syndrome. The Cochrane Database of Systematic Reviews, [online] (3), p.CD003488. [CrossRef]
- Low JG, Sung C, Wijaya L, et al. Efficacy and safety of celgosivir in patients with dengue fever (CELADEN): a phase 1b, randomised, double-blind, placebo-controlled, proof-of-concept trial. Lancet Infect Dis. 2014;14(8):706–15. [CrossRef]
- Palanichamy Kala, M., St. John, A.L. & Rathore, A.P.S. Dengue: Update on Clinically Relevant Therapeutic Strategies and Vaccines. Curr Treat Options Infect Dis 15, 27–52 (2023). [CrossRef]
- Suputtamongkol Y, Avirutnan P, Mairiang D, et al. Ivermectin Accelerates circulating nonstructural protein 1 (NS1) clearance in adult dengue patients: a combined phase 2/3 randomized double-blinded placebo controlled trial. Clin Infect Dis. 2021;72(10):e586–93. [CrossRef]
- Rothan HA, Mohamed Z, Paydar M, Rahman NA, Yusof R. Inhibitory effect of doxycycline against dengue virus replication in vitro. Arch Virol. 2014;159(4):711–8. [CrossRef]
- Palanichamy Kala, M., St. John, A.L. & Rathore, A.P.S. Dengue: Update on Clinically Relevant Therapeutic Strategies and Vaccines. Curr Treat Options Infect Dis 15, 27–52 (2023). [CrossRef]
- Zou J, Lee LT, Wang QY, et al. Mapping the interactions between the NS4B and NS3 proteins of dengue virus. J Virol. 2015;89(7):3471–83. [CrossRef]
- Kaptein SJF, Goethals O, Kiemel D, Marchand A, Kesteleyn B, Bonfanti JF, Bardiot D, Stoops B, Jonckers THM, Dallmeier K, Geluykens P, Thys K, Crabbe M, Chatel-Chaix L, Münster M, Querat G, Touret F, de Lamballerie X, Raboisson P, Simmen K, Chaltin P, Bartenschlager R, Van Loock M, Neyts J. A pan-serotype dengue virus inhibitor targeting the NS3-NS4B interaction. Nature. 2021 Oct;598(7881):504-509. Epub 2021 Oct 6. Erratum in: Nature. 2021 Nov;599(7883):E2. [CrossRef] [PubMed]
- Wu-Chuang A, Rojas A, Bernal C, Cardozo F, Valenzuela A, Romero C, Mateos-Hernández L, Cabezas-Cruz A. Influence of microbiota-driven natural antibodies on dengue transmission. Front Immunol. 2024 Mar 15;15:1368599. https://doi.org/10.3389/fimmu.2024.1368599. [CrossRef] [PubMed]
Figure 2.
An overview of the mechanisms behind dengue related thrombocytopaenia. IgM: Immunoglobulin M, IgG: Immunoglobulin G. (Figure created with Biorender.com).
Figure 2.
An overview of the mechanisms behind dengue related thrombocytopaenia. IgM: Immunoglobulin M, IgG: Immunoglobulin G. (Figure created with Biorender.com).
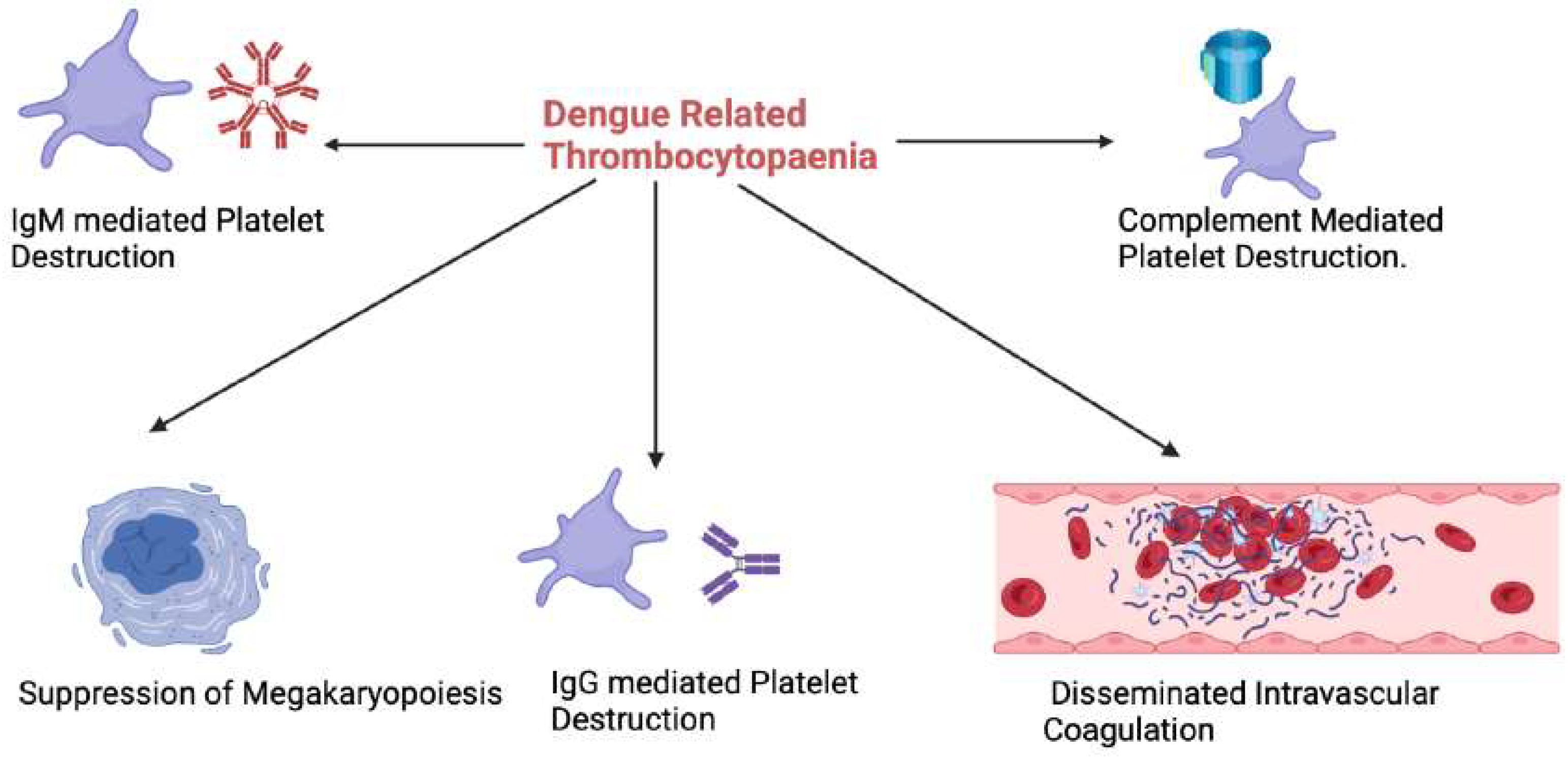
Figure 3.
Dengue mediated leucocyte abnormalities resulting in a dysregulated host immune response. NETS: Neutrophil extracellular traps. (Figure created with Biorender.com).
Figure 3.
Dengue mediated leucocyte abnormalities resulting in a dysregulated host immune response. NETS: Neutrophil extracellular traps. (Figure created with Biorender.com).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



