
Submitted:
14 May 2024
Posted:
15 May 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Viral tropism is most commonly linked to receptor use, but host cell protease use can be a notable factor in susceptibility to infection. Here we review the use of host cell proteases by human viruses, focusing on those with primarily respiratory tropism, particularly SARS-CoV-2. We first describe the various classes of proteases present in the respiratory tract, as well as elsewhere in the body, and incorporate the targeting of these proteases as therapeutic drugs for use in humans. Host cell proteases are also linked to the systemic spread of viruses and play important roles outside of the respiratory tract; therefore, we address how proteases affect viruses across the spectrum of infections that can occur in humans, intending to understand the extrapulmonary spread of SARS-CoV-2.
Keywords:
proteases
; viruses
; respiratory proteases
; protease inhibitors
; SARS-CoV-2
1. Introduction
Among the hurdles that viruses must overcome is the inability to actively adapt to new environments. One way to overcome this hurdle is to utilize host components to modify and activate viral proteins. The human body contains hundreds of proteases, enzymes that process the peptide bonds of both proproteins and proteins through hydrolytic cleavage. Proteases are essential for countless basic human life processes such as hormone and neurotransmitter activation and release,. Likewise, proteolytic cleavage can be important for several steps in the life cycle of viruses; such as host cell entry, uncoating, and virion formation. While many viruses encode their own proteases – which are excellent drug targets – the ability to co-opt host cell proteases can be a major advantage – but one which has received comparably little attention for drug development. For respiratory viruses, the proteases present in the respiratory tract can be used for various viral functions, which can impact pathogenesis and cell tropism. As host proteases do not undergo the same selective evolutionary pressure as viral proteases, inhibitors that target host proteases may elicit a slower viral evolutionary response towards developing drug resistance. Identifying potent inhibitors of respiratory viruses has long been a goal for researchers and has become more of a priority with the advent of the COVID-19 pandemic. SARS-CoV-2, the causative agent of COVID-19, uses several proteases that were previously well known to be involved in the life cycle of other respiratory viruses, as well as illuminating new factors for viral infection. This has allowed researchers to explore foundational research on those viruses and use it as a guide for current drug development. This review will primarily focus on the proteases implicated in SARS-CoV-2 infection, the inhibitors used to disrupt that infection and the background of these proteases and protease inhibitors in other respiratory viruses.
2. Respiratory Proteases
Proteases recognize cleavage substrates through a variety of physical properties. The requirements that a substrate must meet in order to be cleaved vary by the protease of interest. Some proteases, for example, digestive enzymes such as trypsin, are quite general in their requirements and thus retain the ability to cleave a broad array of proteins. Others such as furin are much more specific. Among the properties that determine cleavability are the steric availability of the cleavage site to the protease, as well as the charge, polarity, and glycosylation status of the amino acids within and surrounding the cleavage site. Substrate cleavage sites are typically described using a numbering system introduced by Schechter and Berger [1]. The amino acids on the N terminal side of the cleavage site are numbered P1, P2, etc. with the numbering increasing as distance from the scissile bond increases. On the C terminal side, amino acids are similarly numbered, but as P1’, P2’, etc. as depicted in Figure 1.
Respiratory proteases conduct a variety of important roles in the respiratory tract. They are involved in ciliary function, ion transport, mucus expression, and the immune response, and their dysregulation is tied to multiple disease states [2]. Proteases are categorized by their catalytic residue, and there are four main types of proteases found in the respiratory tract: serine, cysteine, metallo-, and aspartyl proteases. An excellent review of host cell proteases and their role in the cell's immune response to respiratory virus infections has previously been published [3]. In this review, we shall focus on serine, cysteine, and metallo- proteases and their well-characterized roles in facilitating the cell entry and replication of respiratory viruses.
3. Serine Proteases
Serine proteases catalyze the cleavage of proteins via a nucleophile on a serine amino acid in the catalytic site of the enzyme. This serine can be a part of a catalytic triad or dyad. There are 50 different serine protease classes, further divided into 14 clans [4]. A wide variety of serine proteases have been found to proteolytically cleave the glycoproteins of respiratory viruses. Table 1 depicts select well-studied proteases, their cleavage preferences, and the viruses with which they are associated. Serine proteases are the most common class of protease present in the lungs and thus are often utilized by viruses.
3.1. Furin
Furin is a proprotein convertase that minimally cleaves at an R-X-X-R amino acid motif, a polybasic region that must be within an accessible portion of a protein. Furin is present in both the trans-Golgi network and endosomes, and it is essential for the processing of many host substrates crucial to homeostasis and the immune response. While the canonically recognized cleavage site is R-X-X-R, the “furin cleavage site” or FCS is actually much more complex, with furin interacting with its cleavage substrate at 30 amino acids spanning the cleavage site, with some amino acids interacting more strongly than others. A study looking at the interaction of furin with 130 known furin cleavage sites found that the properties of 20 of these amino acids were essential determinants of furin’s ability to cleave [41]. The canonical R-X-X-R motif lies within an 8 amino acids long core region of this larger motif. Within the core region, positively charged residues and flexibility are requirements, with specific amino acids needed at certain positions. Outside of the core region, the polarity of amino acids and accessibility to solvents are also determining factors of cleavage. Recent work on MHV has shown that what may appear to be minor changes in the FCS can have a pronounced effect [42]. In addition to specific amino acid side-chain requirements, it has been found that the glycosylation patterns of proteins (both N- and O-linked) can sometimes abrogate furin cleavage. Furin cleavage requires a pH of 6-8.5 and the presence of Ca2+ in order to be able to cleave [43].
Furin cleavage of influenza hemagglutinin (HA) has been extensively studied as a pathogenicity determinant in the context of highly pathogenic avian influenza [44]. The HA cleavage site is essential for influenza infectivity as, when cleaved, the N-terminus of the fusion peptide is exposed. However, the cleavage of HA exhibits a certain amount of variability. Individual strains of influenza (low pathogenicity or high pathogenicity) may carry either a monobasic or polybasic cleavage site, which determines whether or not their HA can undergo furin cleavage or needs to be cleaved by another protease; such as HAT, TMPRSS2, and other trypsin-like proteases. The presence of a monobasic or polybasic cleavage site is the defining characteristic that separates low-pathogenic viruses from highly pathogenic viruses [45]. This is due to furin being ubiquitous in host cell tissues, vs trypsin-like proteases which are only present in certain tissue types. It should be noted however that cells often express very low amounts of furin, and pathogenesis may be linked to expression levels in different tissues and cell types, eg endothelial cells [46].
Viral envelope proteins are typically cleaved at a single position, but in some cases, there are two distinct cleavage sites. This was first noted for SARS-CoV-1 and then with other coronaviruses. Depending on the individual virus, furin cleavage may be considered a priming event, before the activation protease subsequently cleaves adjacent to the fusion peptide, as with influenza HA, HIV-1 Env, and for paramyxo- and pneumoviruses—this activation is typically through non-furin proteases (see below). Respiratory syncytial virus (RSV, or human orthopneumovirus) is distinct in having two furin cleavage sites, a property shared with MERS-CoV, although the proteases used may be cell-type specific [13,47].
For SARS-CoV-2, furin cleavage of the spike (S) protein is important for the transmission and pathogenesis of the virus, the formation of syncytia, and the available entry pathway into the cell [48]. Furin processing allows for S to enter more easily into an “up” conformation which promotes receptor binding and acts as a “priming” event during virion formation [49]. This priming is essential for further proteolytic processing by either TMPRSS2 or other proteases. However, in the absence of this priming event, virion glycoproteins can instead be cleaved by cathepsins following uptake into the late endosome, leading to fusion with the endosomal membrane. This entry pathway takes more time but allows for the virus to enter a wider range of cells.
As SARS-CoV-2 continued to spread and evolve in the human population, the mutations that accumulated around the furin cleavage site (FCS) underscored the difference between evolutionary pressures in vivo and in vitro. While the virus readily loses the FCS via deletions and point mutations in vitro, the FCS has been strongly conserved in vivo [50]. Though several globally dominating variants have emerged with mutations near the FCS, these mutations have never interfered with furin processing. Rather these mutations seem to take on the role of regulating the level of furin cleavage that occurs (P681H), with some mutations increasing the level of cleavage (P681R) ([51,52,53,54]. While S entering the “up conformation” is advantageous in many ways, it does make the S more susceptible to immune recognition as well as making it less stable. This stability decrease has been lessened by other mutations in S such as D614G which may allow the S to undergo a higher level of cleavage with fewer disadvantages [55]. Modification of the FCS via point mutations also occurred extensively as different isolates of the prototype coronavirus MHV were isolated, and in certain other coronaviruses, (OC43) modulation of the FCS may occur through an indel, in addition to point mutations [56,57]. Thus modulation of furin cleavage through natural selection is a powerful means of regulating CoV tropism and pathogenesis; the virus continues to need to balance the efficiency of transmission benefit of furin cleavage with the need to evade the immune response, now that most people have some level of preexisting immunity.
For coronaviruses, furin-mediated cleavage (via priming at S1/S2) is linked to fusion activation at S2’, but it may also serve a more general role in virion maturation. In this case, furin may be playing an analogous role to that of other viral glycoproteins such as HIV-1 Env and filovirus GP. For the filoviruses EBOV and MARV, furin cleavage precedes the critical cathepsin-mediated cleavage events needed for virus entry, and it is interesting to note that the FCS is found in a quite different position in the GP protein sequence, implying a distinct maturation process between EBOV and MARV [58]. A similar role for furin is found for the DENV prM protein, where it controls the proportion of mature vs immature particles that form, which in turn may affect ADE and viral pathogenesis [59,60,61]. prM that is not cleaved by furin during virion formation can still be cleaved in the endosome during ADE by cathepsins. This alternative cathepsin cleavage route also can take over when SARS-CoV-2 S is not cleaved by furin, leading to furin cleavage being important for infection but not essential.
3.2. Coagulation Cascade Proteases (Plasmin, Thrombin, Factor Xa)
It is important to keep in mind that the host cell components that viruses take advantage of have important roles in maintaining homeostasis. The Blood Coagulation Cascade, which relies on several different proteases, is essential to stop blood loss following injury. Among the contributors to the cascade are Thrombin, Factor Xa, and plasmin. These proteases can be responsible for cleaving other proteins within the pathway, or they can activate various cell types to respond to injury via the cleavage of Protease Activated Receptors (PAR), which are ubiquitously expressed [62]. This cascade exists in a carefully regulated balance, and viral disruption of this balance in either direction can lead to pathology. Respiratory viral infections are known to increase the risk of deep vein thrombosis and pulmonary embolism, as well as other serious vascular complications [63]. This can be caused by either the upregulation of coagulant factors or the downregulation of anticoagulant proteins. Several respiratory viruses can utilize the coagulation proteases for glycoprotein cleavage. This has been posited to form a positive feedback loop, where the glycoprotein is cleaved leading to higher cell entry, which leads to the immune response of increased production of coagulation factors, which in turn leads to increased glycoprotein cleavage [9].
While influenza virus infection in the human respiratory tract is now known to be driven by trypsin-like proteases, the initial breakthrough for our understanding identified plasmin as the key activating protease [64]. The virus strain studied was the neuro-adapted isolate WSN, which had adapted to use plasmin during passage to make it trypsin-independent for growth in cell culture and in the murine brain [65]. Some clinical strains of influenza are also able to utilize plasmin for HA activation [30]. Early studies on SARS-CoV-1 showed a role for plasmin-activated cleavage, primarily at S2’ (R797), along with trypsin and TMPRSS11a; for SARS-CoV-2 plasmin has been indicated as one of the many proteases capable of activating spike [9,32]. Notably, RSV also is activated by plasmin, along with trypsin and thrombin [31].
Influenza HA activation, which is normally dependent on trypsin-like proteases or furin, is thought to lead to dependence on Factor Xa when the virus is adapted to grow in eggs, which is still a common method of growing influenza stocks (eg vaccine seeds)[66]. In contrast, SARS-CoV-2 does not appear to need adaptation to allow the use of a range of systemic secreted proteases such as plasmin, thrombin, and Factor Xa [9]. However, the impact of these proteases during an in vivo infection has yet to be fully elucidated.
3.4. Elastase
In vivo, elastase is produced mainly in the pancreas but also exists as a neutrophil-secreted form as part of the host’s innate immune defenses. The use of elastase as an activating protease has been reported for SARS-CoV-1, which may play a factor in promoting the initial inflammatory response to infection, but the situation for SARS-CoV-2 is less clear [24]. As with any virus-activating protease, the enzyme exists in tissue as part of a tightly regulated system, with neutrophil elastase proposed to be involved in ACE2 shedding and so have a protective effect for COVID [67]. Interestingly, engineering influenza HA to be elastase-cleaved has been used as a means to generate a live-attenuated vaccine virus [68]. Elastase-dependent influenza is attenuated, as elastase is not present in high enough levels during infection to make up for the lack of TMPRSS2 or Furin cleavage during virion formation. This highlights the fact that it is not enough to have a proteolytic cleavage site on the glycoprotein, the cleavage site needs to be able to be cleaved by specific proteases that are present in specific locations during infection.
3.3. Type II Transmembrane Serine Proteases (TTSP):
Type II Transmembrane Serine Proteases (TTSPs) all have a serine protease domain of chymotrypsin with a catalytic triad of histidine, aspartic acid, and serine [69]. Their catalytic triad leads to these proteases being considered trypsin-like. TTSPs are attractive proteases for viruses due to their location on the surface of epithelial cells, allowing for the possibility of glycoprotein processing during cell entry. There are at least 9 TTSPs that are found in the respiratory tract, and they are often involved in viral glycoprotein processing. Low pathogenic strains of influenza are known to rely on TTSPs for glycoprotein activation.

Table 2.
Characteristics of select Type II Transmembrane Serine Proteases.
| Type II Transmembrane Serine Proteases (TTSPs) | Cleavage Preferences | Exploiting Respiratory Viruses |
|---|---|---|
| DESC1 | R-R/A/L-L-A↓ [70] | Influenza virus [71], MERS-CoV [71], SARS-CoV-1 [71] |
| Human Airway Trypsin-like Protease | R/K↓ [72,73] | Influenza virus [74], HCoV-229E [75], SARS-CoV-1 [76], Mammalian Orthoreovirus [6] |
| Matriptase | Minimum: R/K [22]↓ Preferred: R-X(non-basic)-S-R↓ [70] |
Influenza A virus [77,78] |
| TMPRSS11a | Unconfirmed, Putative R/K↓ | SARS-CoV [32], Influenza virus [79], MERS-CoV [79] |
| TMPRSS13/MSPL | R/K↓, Preference for dibasic P2-P1 [80] | Influenza virus [71], SARS-CoV-1 [71], MERS-CoV [71], SARS-CoV-2 [81] |
| TMPRSS2 | R/K↓ [82] | Influenza A + B virus [74], SARS-CoV-2 [83], HMPV [84], HCoV-229E [75], MERS-CoV [85], SARS-CoV [85], HPIV [86], Mammalian Orthoreovirus [6] |
| TMPRSS4 | Unconfirmed, Putative R/K↓[87] | Influenza A virus [88], SARS-CoV-2 [89] |
3.3. TMPRSS2
TMPRSS2, or Transmembrane Serine Protease 2, is a member of the hepsin/TMPRSS subfamily of TTSPS, which consists of at least 7 similar proteases [69]. It cleaves at sterically available monobasic sites, where an arginine or lysine is present. The role of TMRPSS2 has yet to be fully elucidated, with it having proposed roles in fertility, inflammation moderation, prostate protection, and tumor suppression [90]. TMPRSS2 is present throughout the respiratory tract in the lung, bronchus, larynx, trachea, vocal folds, buccal mucosa, nasal mucosa, and tonsils [91]. The level of TMPRSS2 expression is tightly regulated in each tissue type, with over-expression of TMPRSS2 being associated with pathological states such as cancer. Inhibition of TMPRSS2 occurs naturally through host-made factors such as plasminogen inhibitor 1 and hepatocyte growth factor activator inhibitor (HAI-2). However, TMPRSS2 is also associated with the coagulation cascade proteases discussed above. TMPRRS2, trypsin, matriptase, and Factor Xa are all able to activate PAR2, which leads to the increased expression of matrix metalloproteases. These matrix metalloproteases are also known to be involved in viral life cycles, thus even when TMPRSS2 does not have a direct interaction with the virus, it may be still affecting the outcome of infection through other mechanisms.
Its involvement in many respiratory virus life cycles can be readily explained by its high level of expression throughout the respiratory tract. TMPRSS2 is involved in the direct proteolytic activation of many viral glycoproteins and was first identified as a viral cofactor for influenza [74]. TMPRSS2 processes the Influenza HA during virion formation, an essential step that renders HA fusion competent. TMPRSS2 also acts as a proteolytic activator during the cell entry of several coronaviruses, including SARS-CoV-1, SARS-CoV-2, HCoV-229E, and MERS-CoV. The activation of these viral glycoproteins by TMPRSS2 is an essential step for entry at the cell surface as it allows these glycoproteins to change into their fusion-competent conformations. TMPRSS2’s cleavage specificity is also broad enough that it is able to cleave cleavage sites that are traditionally considered to be “furin cleavage sites”. TMPRSS2 is interestingly also involved in the cleavage of ACE2, an additional host cell protease that will be discussed in depth later in this paper. This processing of ACE2 has been found to augment SARS-CoV-1 cell entry, in addition to TMPRSS2 processing of the SARS-CoV-1 S [92].
4. Cysteine Proteases - Cathepsins
Cathepsins are a type of cysteine protease that resemble Papain, leading to them also being referred to as Papain-Like Proteases. Their active site consists of a catalytic dyad with a histidine residue and a cysteine amino acid acting as the nucleophile [93]. Cathepsins are generally endosomal proteases but can also be found in other locations. For example, Cathepsin L is present in endosomes and lysosomes but is also secreted outside of the cell. The level of Cathepsin L in the blood is correlated to the level of pathogenesis following viral infection and is also elevated during other noninfectious pathological states [94]. Cathepsins need to be exposed to an acidic environment to be activated, and they prefer acidic and reducing conditions for cleavage. These proteases participate in a variety of virus life cycles and often are integral for alternative entry pathways. Cathepsins cleave viral glycoproteins in these so-called alternative pathways for several respiratory viruses, along with coronaviruses such as SARS-CoV-1, SARS-CoV-2, and MERS-CoV. For SARS-CoV-2, later variants show a preference for the cathepsin pathway in cell culture, despite this shift not occurring in vivo or in human airway organoids [95,96,97]. Beyond just glycoprotein cleavage, cathepsins also are used by viruses to modulate the immune response through several different pathways [98].
As a part of the Mammalian Orthoreovirus (MRV) life cycle, the virons’ capsids must be proteolytically digested away to become Infectious Subvirion Particles (ISVPs), which are then able to penetrate the endosomal membrane and gain entry into the cell [99]. This proteolytic digestion of the capsid has been attributed to several different cathepsins, making cysteine proteases essential for MRV infection. Specific point mutations in the MRV protein that undergoes cleavage have been found to increase the efficiency of cathepsin cleavage [100,101]. However, despite proteolytic cleavage being considered the rate-limiting step of infection, few strains of MRV have incorporated these mutations, and viruses with these mutations lose titer more rapidly at elevated temperatures when compared to wild-type virus [102]. This reaffirms that proteolytic cleavage requires a careful balance between cleavage efficiency and overall viral fitness. Cathepsin cleavage is intimately tied to the route of entry for reovirus, and has been reviewed in Mainou and Dermody 2012 [103].
Many other virus systems take advantage of cathepsin cleavage in endosomes. Perhaps the most notable of these is the Ebola virus, whereby, following non-specific binding at the cell surface and internalization, CatL and CatB degrade the filovirus GP down to an 18kDa ‘stub’ which is the fusion-active component upon engagement with the NPC-1 receptor [104]. Another not-classically respiratory virus (albeit transmitted via the oro-nasal route) is Nipah virus; in this case, the F protein enters recycling endosomes where it encounters cathepsins as part of the virus assembly pathway [105,106]. Cathepsins are also involved in RSV infection, via cleavage of the G (attachment) protein, and influenza infection via unspecified means [107].
Table 3.
Characteristics of select Cathepsins.
| Cysteine Proteases | Cleavage Determinants | Exploiting Respiratory Viruses |
|---|---|---|
| Cathepsin L | Prefers aromatic or aliphatic residues in P2 [108] | HCoV-229E [75], SARS-CoV-1 [109,110,111], SARS-CoV-2 [112], Mammalian Orthoreovirus [113], MERS-CoV [114], Nipah [115], RSV [107], Hendra [115] |
| Cathepsin B | Prefers an aromatic or aliphatic residue and tolerates a basic P2, an aromatic residue in P1’ and a P3' G [108] | Influenza A virus [116], Mammalian Orthoreovirus [113], Nipah [105], SARS-CoV-2 [117] |
| Cathepsin S | Prefers aliphatic residues in P2, G/E in P1 [108] | Mammalian Orthoreovirus [118], SARS-CoV-2, SARS-CoV-1, RSV [40] |
| Cathepsin W | W/F–L/V–G/A/R↓V–D/N/E/Q (Suggested [119]] | Influenza A virus [120], RSV [40] |
| Cathepsin K | Prefers non-aromatic hydrophobic residues in P2 [121] | SARS-CoV-2 [117] |
| Cathepsin V | Prefers hydrophobic residues in P2, P in P3 [122] | SARS-CoV-2 [117] |
4. Metalloproteases
Metalloproteases differ from serine and cysteine proteases in that instead of using an amino acid as a nucleophile in their hydrolytic reaction, metalloproteases use a water molecule that has been activated by a metallic cation. Some metalloproteases have been found to cleave viral glycoproteins, such as ADAM and MT-MMP. However, this function is less well studied from the perspective of viruses than other groups of proteases.
Many metalloproteases also impact the viral life cycle in less direct ways. Matrix metalloproteases (MMPS) in particular are important players in many pathways that regulate the immune response and wound repair. MMPS are regulated by Tissue Inhibitors of Metalloproteases (TIMPS), and the interruption of this careful balance between activity and inhibition is associated with many disease states [123]. Respiratory syncytial virus (RSV) is known to induce the expression of many MMPS. Drugs targeting these proteases were found to enhance viral clearance, inhibit syncytia formation, and prevent viral replication [40,124]. MMP-9 specifically is also implicated in other respiratory viruses, such as influenza, rhinovirus, and human parainfluenza virus [125,126,127]. The exact mechanism of action of these MMPs in enhancing viral infection is unknown and remains mainly as hypotheses, based on either MMPs role in restructuring the respiratory tract or their role in the immune response.
MMPs were directly shown to act on the coronavirus spike, first through a study of MHV where MMP- and ADAM-family proteases acted in a strain-dependent manner, and have more recently been added to the list of activating proteases for SARS-CoV-2 [128,129,130].
4.1. ACE2
Another metalloprotease of interest is Angiotensin Converting Enzyme II (ACE2) which, as the name suggests, regulates angiotensin II levels through hydrolytic cleavage. Its position at the cell surface allows it to function as a receptor for SARS-CoV-2. The S1 subunit of the spike protein binds to ACE2, which begins the process of the cell being infected [131]. This action does not directly cleave the spike protein, but it does allow for the spike protein to be cleaved by other proteases, such as the previously discussed TMPRSS2 [132]. This use of proteases as receptors, and not for their proteolytic capacity, is also seen in several other coronaviruses, and the pharmacological intervention of this receptor binding interaction is a major area of research. Drugs targeting the SARS-CoV-2 spike and ACE2 are not focused on blocking the proteolytic activity of ACE2. Rather, research focuses on ACE2 decoys that will bind the spike protein before the virus arrives at a cell, thereby preventing successful receptor binding and infection [133]. Recently ACE2 was also revealed to act as a regulator of TMPRSS2 activity. This regulation occurs through a non-catalytic mechanism but has been shown to increase TMPRSS2 activation of Influenza A and MERS-CoV [134]. When we think of the roles of host proteases in viral infection, it is tempting to think of them as having a straightforward interaction with viral glycoproteins, ie. ACE2 acts as a receptor for SARS-CoV-2 S. However, we must remember that these proteases exist in a complex and ever-changing environment, and they have important roles that they serve within the host. The relationship between proteases and viral infections cannot be simple, because the system that this relationship exists in is not simple.
Table 4.
Characteristics of select metalloproteases.
| Metalloprotease | Exploiting Respiratory Viruses | Role |
|---|---|---|
| MT-MMP | SARS-CoV-2 [130] | Proteolytic cleavage |
| ADAM | SARS-CoV-2 [130] | Proteolytic cleavage |
| ACE2 | SARS-CoV-2 [135], SARS-CoV [136], HcoV NL63 [137] | Influenza A virus [134], MERS-CoV [134] | Receptor| Regulation of TMPRSS2 |
| APN | HCoV-229E [138] | Receptor |
5. Protease Inhibitors –
Protease inhibitors, like proteases themselves, are a broad and diverse group. Inhibitors play an essential role in maintaining homeostasis, preventing proteases from being overly active. Due to this role, protease inhibitors have been found in many life forms, leading to a large group of existing inhibitors to explore as drug candidates. Inhibitors can be classified by the type of protease they inhibit or, like other enzyme inhibitors, by their mode of action. When an inhibitor binds to the enzyme through a covalent interaction, this is considered an irreversible reaction. These inhibitors fall into either the less specific group of affinity labeling inhibitors or the more specific suicide inhibitors, which require the protease to enzymatically act upon them to achieve inhibition [139]. For reversible inhibitors, inhibitors noncovalently interact with the protease, and the location of that interaction determines whether it is competitive, noncompetitive, or uncompetitive inhibition. Competitive inhibitors occupy the active site of the protease, noncompetitive inhibitors bind elsewhere on the protein, and uncompetitive inhibitors only bind to enzymes that are already bound to their substrates. Inhibitors can be proteins, peptides, small molecules, or antibodies. They can directly interact with the protease, or they can interact with molecules that proteases need to be active, ie. Ca2+ chelators that inhibit furin.
Because the kinetics of inhibition differ between the inhibitor types, whether an inhibitor is reversible or irreversible is a determinant of its appeal as an antiviral. Irreversible inhibitors are attractive because of how long they are active and the ease of which they can be designed, but they have more potential for negative side effects, especially if they are widely active on many proteases [140]. Both the length of activity and the potential for negative side effects stem from the fact that the inhibition is irreversible. Once a protease is inhibited, a cell will need to make new protease in order to regain proteolytic function. An irreversible inhibitor that is broadly acting could disrupt many processes involved in homeostasis. Therefore, irreversible inhibitors have a higher requirement for specificity and subcellular targeting design than reversible inhibitors. Competitive reversible inhibitors are also easy to design, due to their need to fit into the active site of the enzyme, but they require a high concentration to compete with other substrates. This can be a barrier when trying to inhibit the cleavage of viral proteins in subcellular compartments, as you need to get a large quantity of the inhibitor into those specific compartments. Uncompetitive and non-competitive reversible inhibitors do not have this same issue with concentration; however, these types of inhibitors are complicated to design. However, the problems of irreversible and competitive inhibitors both have the same solution, cellular and subcellular drug targeting. Directing inhibitors to the correct location would allow the concentration to reach a high level in the area of interest without the need for an extreme dose to the rest of the body. Ideally, the inhibitor would remain inert until it reached its target destination, lowering the chances of adverse side effects. Research is quickly advancing both in drug modifications that direct drugs to be taken up into specific cell compartments, as well as the creation of nanoparticle carriers which allow for precise drug targeting as well as having other benefits [141,142].
The human genome encodes for numerous protease inhibitors, as maintaining a careful balance of proteolytic activity is essential for homeostasis. However, these endogenous inhibitors also carry out important roles in the immune response to viral infections. The viruses that are inhibited by endogenous inhibitors are diverse and include coronaviruses, flaviviruses, and filoviruses among others [143]. This inhibition can be a direct interruption of the catalytic mechanism of cleavage, a rerouting of proteases to an alternate cellular compartment, or an interaction with the protease on one of its other functional domains. A comprehensive review of these endogenous inhibitors has been carried out by Lotke, Petersen, and Sauter [143]. Of particular interest among these inhibitors are Serpins.
Serine protease inhibitors (Serpins) are present in most types of life. These inhibitors mostly inhibit serine proteases but select serpins can also inhibit other types of proteases. Serpins contain cleavage motifs of the proteases that they inhibit, and upon their cleavage, they undergo a conformational change that covalently traps the protease in an irreversible interaction [144]. This conformational change is irreversible; thus, they are suicide inhibitors. Interestingly, poxviruses have been found to encode their own serpins. Poxvirus serpins have been found to exhibit anti-apoptotic and anti-inflammatory roles [145]. The consideration of the effects serpins have on the immune response is essential to keep in mind when considering serpins for the treatment of respiratory viruses.
6. Current Therapeutic Strategies Targeting Coronaviruses, Influenza viruses and Para-Myxoviruses
The COVID-19 pandemic prompted a resurgence of therapeutic development for respiratory viruses and while some drugs targeting SARS-CoV-2 are now approved for use in humans, many additional drugs are currently approved in development, including inhibitors of the viral polymerase and viral Mpro and 3CL-like proteases [146]. Recent oral inhibitors now available include Paxlovid® and while this remains in use, Molnupiravir® and the injectable Remdesivir® are no longer widely recommended, and most therapeutic monoclonal antibodies have been discontinued. Currently, the main therapeutics targeting influenza are the DAAs Tamiflu® and Relenza® (targeting the viral neuraminidase), with Baloxavir® also now available (targeting the viral polymerase). However, numerous influenza virus strains are emerging that are resistant to Tamiflu® and Relenza®, rendering the treatment ineffective. For paramyxoviruses and pnuemoviruses, ribavirin has been utilized for RSV treatment and shows similar in vitro neutralization of HMPV [147], but there are significant questions concerning its efficacy in clinical settings. Based on successes with RSV, humanized monoclonal antibodies, the primary prophylactic treatment for HRSV, are currently in development for HMPV, but the cost of such treatment is extremely high. Host-targeted approaches are also in development and have the notable potential benefit of limiting antiviral-resistant viruses emerging.
Inhibition of furin as an antiviral strategy.
Given its importance for viral infection, furin has received notable attention as a therapeutic option [148]. The first generation of furin inhibitors were peptide-based and lacked specificity, and were followed by serpins such as AT-PDX, which have high specificity and increased potential as therapeutic agents. Recent studies with bioavailable small molecule furin inhibitors such as BOS-981 that effectively inhibit SARS-CoV-2 show promise as candidate therapeutics, especially when combined with TTSP inhibitors (see below) [149]. Ca2+ chelators, Serine Protease inhibitors (Serpins), and proprotein convertase inhibitors have all been successful at inhibiting furin. Most known inhibitors of furin have inhibitory effects on other proteases as well, so the development of new, targeted furin inhibitors is still an area of intense interest. However, as some viruses are able to utilize multiple proteases for glycoprotein cleavage, it is important to design treatments that carefully balance the need to prevent off-target effects while still targeting the diverse set of proteases that viruses utilize.
Inhibition of TTSPs as an antiviral strategy
Inhibition of TTSPs is among the most promising alternative host-targeted approaches under consideration as therapeutics or prophylactics, for SARS-Cov-2 or other respiratory pathogens. The use of TTSP inhibitors was originally investigated for the treatment of influenza, as an alternative to inhibitors that target the influenza protease neuraminidase [150,151,152]. Camostat, a TTSP inhibitor repurposed as an anti-viral, is currently in clinical trials for COVID-19—although currently with only limited success [153]. Notably, camostat has relatively low potency and is taken orally, which may contribute to its limited success towards targeting respiratory disease. Impeding proteolytic activation of the viral HA via the serine protease inhibitor aprotinin [154], which has anti-fibrinolytic properties, has also been explored, including preclinical development and some limited clinical trials. The partial success of these trials reinforces the concept of targeting host cell-mediated proteolytic activation of viral glycoproteins as a viable therapeutic strategy. However, the development of aprotinin has limitations. First, being proteinaceous, indications are that aprotinin has a very short biological half-life, on the order of 0.7h [155]. Second, it is currently unclear exactly which proteases are active in the human respiratory tract, and so which proteases need to be targeted. While aprotinin is somewhat broadly acting, it is likely to only target a subset of proteases. Importantly, it is a bovine-derived product, likely contributing to its toxicity and removal from the market [156]. Therefore, the development of alternative protease-targeted therapeutics in addition to aprotinin is likely to significantly expand the therapeutic options available to treat respiratory viruses.
As proof-of-principle for this approach, a lead protease inhibitor comprising the kunitz-type, serine protease inhibitor SPINT2, also known as HAI-2, was tested [157]. SPINT2 is a 25 kDa protein found on the plasma membrane of a number of tissues, where it is mainly bound to the serine protease matriptase. Both matriptase and SPINT2 are initially located on the plasma membrane but are proteolytically released into the extracellular space. SPINT2 contains two Kunitz-type inhibitor domains that potently modulate the proteolytic activity of several trypsin-like serine proteases, including matriptase and hepatocyte growth factor activator; inhibition of these two proteases is thought to be the primary biological role of SPINT2. The properties of SPINT2 therefore made it an excellent proof of principle to efficiently inhibit viral replication, including both influenza and HMPV [156,158]. Small molecule approaches previously discovered included an initial lead compound IN-1 (N-0100) as an influenza therapeutic [77]. This was developed into N-0385, a small molecule peptidomimetic with a ketobenzothiazole warhead. In mice, N-0385 treatment reduced morbidity and mortality associated with influenza H1N1 and H3N2 infection, as well as proving highly effective in mice, to achieve progress toward the goal of prophylactic inhibition of a broad range of respiratory viruses [159].
7. Conclusions
The human genome encodes for hundreds of diverse proteolytic enzymes that are vital for the maintenance of homeostasis. These proteases are sometimes coopted for virion maturation, cell entry, and immune modulation, among other things. These proteases provide an additional area to focus on antiviral development. The COVID-19 pandemic has helped further the field of antiviral development but has also revealed complexities in the relationship between viruses and proteases that make designing antivirals more challenging. However, the field is readily rising to the challenge, and the field of antivirals targeting host cell proteases remains one of great interest.
Author Contributions
Writing—review and editing, B.L. and G.W.; supervision, G.W. All authors have read and agreed to the published version of the manuscript.
Funding
Work in the Author’s lab on this topic is funded by the National Institute of Health (NIH, grant number R01AI35270 to GW
Acknowledgments
We would like to thank all of the members of the Whittaker lab for their support, with a special acknowledgment to Annette Choi for many helpful discussions.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the writing of the manuscript.
References
- Schechter I, Berger A. On the size of the active site in proteases. I. Papain. Biochem Biophys Res Commun. 1967 Apr 20;27(2):157–62.
- McKelvey MC, Brown R, Ryan S, Mall MA, Weldon S, Taggart CC. Proteases, Mucus, and Mucosal Immunity in Chronic Lung Disease. Int J Mol Sci. 2021 May 9;22(9):5018. [CrossRef]
- Majchrzak M, Poręba M. The roles of cellular protease interactions in viral infections and programmed cell death: a lesson learned from the SARS-CoV-2 outbreak and COVID-19 pandemic. Pharmacol Rep. 2022 Dec 1;74(6):1149–65.
- Rawlings ND, Barrett AJ. Chapter 559 - Introduction: Serine Peptidases and Their Clans. In: Rawlings ND, Salvesen G, editors. Handbook of Proteolytic Enzymes (Third Edition) [Internet]. Academic Press; 2013 [cited 2023 Jul 13]. p. 2491–523. Available from: https://www.sciencedirect.com/science/article/pii/B9780123822192005597.
- Andersson MK, Enoksson M, Gallwitz M, Hellman L. The extended substrate specificity of the human mast cell chymase reveals a serine protease with well-defined substrate recognition profile. Int Immunol. 2009 Jan;21(1):95–104. [CrossRef]
- Nygaard RM, Golden JW, Schiff LA. Impact of Host Proteases on Reovirus Infection in the Respiratory Tract. Journal of Virology. 2012 Jan 15;86(2):1238–43. [CrossRef]
- Brown MA, Stenberg LM, Stenflo J. Coagulation Factor Xa. Handbook of Proteolytic Enzymes. 2013;2908–15.
- Du L, Kao RY, Zhou Y, He Y, Zhao G, Wong C, et al. Cleavage of spike protein of SARS coronavirus by protease factor Xa is associated with viral infectivity. Biochemical and Biophysical Research Communications. 2007 Jul 20;359(1):174–9. [CrossRef]
- Kastenhuber ER, Mercadante M, Nilsson-Payant B, Johnson JL, Jaimes JA, Muecksch F, et al. Coagulation factors directly cleave SARS-CoV-2 spike and enhance viral entry. van der Meer JW, editor. eLife. 2022 Mar 16;11:e77444.
- Hosaka M, Nagahama M, Kim WS, Watanabe T, Hatsuzawa K, Ikemizu J, et al. Arg-X-Lys/Arg-Arg motif as a signal for precursor cleavage catalyzed by furin within the constitutive secretory pathway. Journal of Biological Chemistry. 1991 Jul 5;266(19):12127–30. [CrossRef]
- Kawaoka Y, Webster RG. Sequence requirements for cleavage activation of influenza virus hemagglutinin expressed in mammalian cells. Proceedings of the National Academy of Sciences. 1988 Jan;85(2):324–8. [CrossRef]
- Peacock TP, Goldhill DH, Zhou J, Baillon L, Frise R, Swann OC, et al. The furin cleavage site in the SARS-CoV-2 spike protein is required for transmission in ferrets. Nat Microbiol. 2021 Jul;6(7):899–909.
- Millet JK, Whittaker GR. Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein. Proc Natl Acad Sci U S A. 2014 Oct 21;111(42):15214–9. [CrossRef]
- Collins PL, Huang YT, Wertz GW. Nucleotide sequence of the gene encoding the fusion (F) glycoprotein of human respiratory syncytial virus. Proc Natl Acad Sci U S A. 1984 Dec;81(24):7683–7. [CrossRef]
- Ortmann D, Ohuchi M, Angliker H, Shaw E, Garten W, Klenk HD. Proteolytic cleavage of wild type and mutants of the F protein of human parainfluenza virus type 3 by two subtilisin-like endoproteases, furin and Kex2. J Virol. 1994 Apr;68(4):2772–6. [CrossRef]
- Watanabe M, Hirano A, Stenglein S, Nelson J, Thomas G, Wong TC. Engineered serine protease inhibitor prevents furin-catalyzed activation of the fusion glycoprotein and production of infectious measles virus. J Virol. 1995 May;69(5):3206–10. [CrossRef]
- Ueo A, Kubota M, Shirogane Y, Ohno S, Hashiguchi T, Yanagi Y. Lysosome-Associated Membrane Proteins Support the Furin-Mediated Processing of the Mumps Virus Fusion Protein. J Virol. 2020 Jun 1;94(12):e00050-20. [CrossRef]
- Borgoño CA, Gavigan JA, Alves J, Bowles B, Harris JL, Sotiropoulou G, et al. Defining the extended substrate specificity of kallikrein 1-related peptidases. 2007 Nov 1;388(11):1215–25. [CrossRef]
- Milewska A, Falkowski K, Kulczycka M, Bielecka E, Naskalska A, Mak P, et al. Kallikrein 13 serves as a priming protease during infection by the human coronavirus HKU1. Science Signaling. 2020 Nov 24;13(659):eaba9902. [CrossRef]
- Li HX, Hwang BY, Laxmikanthan G, Blaber SI, Blaber M, Golubkov PA, et al. Substrate specificity of human kallikreins 1 and 6 determined by phage display. Protein Sci. 2008 Apr;17(4):664–72. [CrossRef]
- Hamilton BS, Whittaker GR. Cleavage Activation of Human-adapted Influenza Virus Subtypes by Kallikrein-related Peptidases 5 and 12 *. Journal of Biological Chemistry. 2013 Jun 14;288(24):17399–407. [CrossRef]
- Harris JL, Backes BJ, Leonetti F, Mahrus S, Ellman JA, Craik CS. Rapid and general profiling of protease specificity by using combinatorial fluorogenic substrate libraries. Proc Natl Acad Sci U S A. 2000 Jul 5;97(14):7754–9. [CrossRef]
- Golden JW, Schiff LA. Neutrophil elastase, an acid-independent serine protease, facilitates reovirus uncoating and infection in U937 promonocyte cells. Virology Journal. 2005 May 31;2(1):48. [CrossRef]
- Belouzard S, Madu I, Whittaker GR. Elastase-mediated Activation of the Severe Acute Respiratory Syndrome Coronavirus Spike Protein at Discrete Sites within the S2 Domain *. Journal of Biological Chemistry. 2010 Jul 23;285(30):22758–63. [CrossRef]
- Matsuyama S, Ujike M, Morikawa S, Tashiro M, Taguchi F. Protease-mediated enhancement of severe acute respiratory syndrome coronavirus infection. Proceedings of the National Academy of Sciences. 2005 Aug 30;102(35):12543–7. [CrossRef]
- Seidah NG. Chapter 730 - Proprotein Convertase 5. In: Rawlings ND, Salvesen G, editors. Handbook of Proteolytic Enzymes (Third Edition) [Internet]. Academic Press; 2013 [cited 2024 May 4]. p. 3305–10. Available from: https://www.sciencedirect.com/science/article/pii/B9780123822192007304.
- Basak A, Zhong M, Munzer JS, Chrétien M, Seidah NG. Implication of the proprotein convertases furin, PC5 and PC7 in the cleavage of surface glycoproteins of Hong Kong, Ebola and respiratory syncytial viruses: a comparative analysis with fluorogenic peptides. Biochem J. 2001 Feb 1;353(Pt 3):537–45.
- Castellino FJ. Chapter 648 - Plasmin. In: Rawlings ND, Salvesen G, editors. Handbook of Proteolytic Enzymes (Third Edition) [Internet]. Academic Press; 2013 [cited 2023 Jul 13]. p. 2958–68. Available from: https://www.sciencedirect.com/science/article/pii/B9780123822192006487.
- Xue F, Seto CT. Selective Inhibitors of the Serine Protease Plasmin: Probing the S3 and S3‘ Subsites Using a Combinatorial Library. J Med Chem. 2005 Nov 1;48(22):6908–17. [CrossRef]
- Tse LV, Marcano VC, Huang W, Pocwierz MS, Whittaker GR. Plasmin-Mediated Activation of Pandemic H1N1 Influenza Virus Hemagglutinin Is Independent of the Viral Neuraminidase. J Virol. 2013 May;87(9):5161–9. [CrossRef]
- Dubovi EJ, Geratz JD, Tidwell RR. Enhancement of respiratory syncytial virus-induced cytopathology by trypsin, thrombin, and plasmin. Infect Immun. 1983 Apr;40(1):351–8. [CrossRef]
- Kam YW, Okumura Y, Kido H, Ng LFP, Bruzzone R, Altmeyer R. Cleavage of the SARS Coronavirus Spike Glycoprotein by Airway Proteases Enhances Virus Entry into Human Bronchial Epithelial Cells In Vitro. PLOS ONE. 2009 Nov 17;4(11):e7870. [CrossRef]
- Hou Y, Yu T, Wang T, Ding Y, Cui Y, Nie H. Competitive cleavage of SARS-CoV-2 spike protein and epithelial sodium channel by plasmin as a potential mechanism for COVID-19 infection. Am J Physiol Lung Cell Mol Physiol. 2022 Nov 1;323(5):L569–77.
- Gallwitz M, Enoksson M, Thorpe M, Hellman L. The Extended Cleavage Specificity of Human Thrombin. PLoS One. 2012 Feb 27;7(2):e31756. [CrossRef]
- Rodriguez T, Dobrovolny HM. Quantifying the effect of trypsin and elastase on in vitro SARS-CoV infections. Virus Research. 2021 Jul 2;299:198423.
- Kirchdoerfer RN, Wang N, Pallesen J, Wrapp D, Turner HL, Cottrell CA, et al. Stabilized coronavirus spikes are resistant to conformational changes induced by receptor recognition or proteolysis. Sci Rep. 2018 Oct 24;8(1):15701.
- Schowalter RM, Smith SE, Dutch RE. Characterization of Human Metapneumovirus F Protein-Promoted Membrane Fusion: Critical Roles for Proteolytic Processing and Low pH. J Virol. 2006 Nov;80(22):10931–41. [CrossRef]
- Henrickson KJ. Parainfluenza Viruses. Clin Microbiol Rev. 2003 Apr;16(2):242–64.
- Thorpe M, Fu Z, Chahal G, Akula S, Kervinen J, de Garavilla L, et al. Extended cleavage specificity of human neutrophil cathepsin G: A low activity protease with dual chymase and tryptase-type specificities. PLoS One. 2018 Apr 13;13(4):e0195077. [CrossRef]
- Foronjy RF, Taggart CC, Dabo AJ, Weldon S, Cummins N, Geraghty P. Type-I interferons induce lung protease responses following respiratory syncytial virus infection via RIG-I-like receptors. Mucosal Immunology. 2015 Jan 1;8(1):161–75. [CrossRef]
- Tian S. A 20 Residues Motif Delineates the Furin Cleavage Site and its Physical Properties May Influence Viral Fusion. Biochem Insights. 2009 Jan 1;2:BCI.S2049. [CrossRef]
- Choi A, Kots ED, Singleton DT, Weinstein H, Whittaker GR. Analysis of the molecular determinants for furin cleavage of the spike protein S1/S2 site in defined strains of the prototype coronavirus murine hepatitis virus (MHV). Virus Res. 2024 Feb;340:199283.
- Molloy SS, Bresnahan PA, Leppla SH, Klimpel KR, Thomas G. Human furin is a calcium-dependent serine endoprotease that recognizes the sequence Arg-X-X-Arg and efficiently cleaves anthrax toxin protective antigen. Journal of Biological Chemistry. 1992 Aug 15;267(23):16396–402. [CrossRef]
- Böttcher-Friebertshäuser E, Klenk HD, Garten W. Activation of influenza viruses by proteases from host cells and bacteria in the human airway epithelium. Pathogens and Disease. 2013 Nov 1;69(2):87–100. [CrossRef]
- Lu X, Shi Y, Gao F, Xiao H, Wang M, Qi J, et al. Insights into Avian Influenza Virus Pathogenicity: the Hemagglutinin Precursor HA0 of Subtype H16 Has an Alpha-Helix Structure in Its Cleavage Site with Inefficient HA1/HA2 Cleavage. Journal of Virology. 2012 Dec;86(23):12861–70.
- Mayer G, Boileau G, Bendayan M. Sorting of Furin in Polarized Epithelial and Endothelial Cells: Expression Beyond the Golgi Apparatus. J Histochem Cytochem. 2004 May 1;52(5):567–79. [CrossRef]
- Krzyzaniak MA, Zumstein MT, Gerez JA, Picotti P, Helenius A. Host Cell Entry of Respiratory Syncytial Virus Involves Macropinocytosis Followed by Proteolytic Activation of the F Protein. PLOS Pathogens. 2013 Apr 11;9(4):e1003309. [CrossRef]
- Takeda M. Proteolytic activation of SARS-CoV-2 spike protein. Microbiol Immunol. 2022 Jan;66(1):15–23.
- Gobeil SMC, Janowska K, McDowell S, Mansouri K, Parks R, Manne K, et al. D614G Mutation Alters SARS-CoV-2 Spike Conformation and Enhances Protease Cleavage at the S1/S2 Junction. Cell Reports. 2021 Jan 12;34(2):108630.
- Sasaki M, Uemura K, Sato A, Toba S, Sanaki T, Maenaka K, et al. SARS-CoV-2 variants with mutations at the S1/S2 cleavage site are generated in vitro during propagation in TMPRSS2-deficient cells. PLOS Pathogens. 2021 Jan 21;17(1):e1009233.
- Akatsuki Saito, et al. Nature. 2021 [cited 2022 Sep 8]. Enhanced fusogenicity and pathogenicity of SARS-CoV-2 Delta P681R mutation | Nature. Available from: https://www.nature.com/articles/s41586-021-04266-9.
- Lubinski B, Frazier LE, Phan MVT, Bugembe DL, Cunningham JL, Tang T, et al. Spike protein cleavage-activation mediated by the SARS-CoV-2 P681R mutation: a case-study from its first appearance in variant of interest (VOI) A.23.1 identified in Uganda. bioRxiv. 2022 Mar 28;2021.06.30.450632.
- Lubinski B, Fernandes MHV, Frazier L, Tang T, Daniel S, Diel DG, et al. Functional evaluation of the P681H mutation on the proteolytic activation of the SARS-CoV-2 variant B.1.1.7 (Alpha) spike. iScience. 2022 Jan 21;25(1):103589.
- Liu Y, Liu J, Johnson BA, Xia H, Ku Z, Schindewolf C, et al. Delta spike P681R mutation enhances SARS-CoV-2 fitness over Alpha variant. Cell Reports. 2022 May 17;39(7):110829.
- Gellenoncourt S, Saunders N, Robinot R, Auguste L, Rajah MM, Kervevan J, et al. The Spike-Stabilizing D614G Mutation Interacts with S1/S2 Cleavage Site Mutations To Promote the Infectious Potential of SARS-CoV-2 Variants. J Virol. 96(19):e01301-22.
- Lau SKP, Li KSM, Li X, Tsang KY, Sridhar S, Woo PCY. Fatal Pneumonia Associated With a Novel Genotype of Human Coronavirus OC43. Front Microbiol [Internet]. 2022 Jan 14 [cited 2024 May 7];12. [CrossRef]
- Stodola JK, Dubois G, Le Coupanec A, Desforges M, Talbot PJ. The OC43 human coronavirus envelope protein is critical for infectious virus production and propagation in neuronal cells and is a determinant of neurovirulence and CNS pathology. Virology. 2018 Feb;515:134–49. [CrossRef]
- Structure of the Ebola virus glycoprotein bound to an antibody from a human survivor | Nature [Internet]. [cited 2024 Apr 9]. Available from: https://www.nature.com/articles/nature07082.
- Julie M. Murray, et al. Journal of General Virology. 1993 [cited 2022 Sep 8]. Processing of the dengue virus type 2 proteins prM and C-prM | Microbiology Society. [CrossRef]
- Dejnirattisai W, Jumnainsong A, Onsirisakul N, Fitton P, Vasanawathana S, Limpitikul W, et al. Cross-Reacting Antibodies Enhance Dengue Virus Infection in Humans. Science. 2010 May 7;328(5979):745–8. [CrossRef]
- Luo YY, Feng JJ, Zhou JM, Yu ZZ, Fang DY, Yan HJ, et al. Identification of a novel infection-enhancing epitope on dengue prM using a dengue cross-reacting monoclonal antibody. BMC Microbiology. 2013 Aug 29;13(1):194. [CrossRef]
- Schreuder H, Matter H. Serine Proteinases from the Blood Coagulation Cascade. In: Structural Biology in Drug Discovery [Internet]. John Wiley & Sons, Ltd; 2020 [cited 2024 Apr 9]. p. 395–422. [CrossRef]
- Franczuk P, Tkaczyszyn M, Kulak M, Domenico E, Ponikowski P, Jankowska EA. Cardiovascular Complications of Viral Respiratory Infections and COVID-19. Biomedicines. 2022 Dec 27;11(1):71. [CrossRef]
- Lazarowitz SG, Choppin PW. Enhancement of the infectivity of influenza A and B viruses by proteolytic cleavage of the hemagglutinin polypeptide. Virology. 1975 Dec 1;68(2):440–54. [CrossRef]
- Sun X, Tse LV, Ferguson AD, Whittaker GR. Modifications to the Hemagglutinin Cleavage Site Control the Virulence of a Neurotropic H1N1 Influenza Virus. J Virol. 2010 Sep;84(17):8683–90. [CrossRef]
- Gotoh B, Ogasawara T, Toyoda T, Inocencio NM, Hamaguchi M, Nagai Y. An endoprotease homologous to the blood clotting factor X as a determinant of viral tropism in chick embryo. EMBO J. 1990 Dec;9(12):4189–95. [CrossRef]
- Kummarapurugu AB, Hawkridge AM, Ma J, Osei S, Martin RK, Zheng S, et al. Neutrophil elastase decreases SARS-CoV-2 spike protein binding to human bronchial epithelia by clipping ACE-2 ectodomain from the epithelial surface. Journal of Biological Chemistry [Internet]. 2023 Jun 1 [cited 2024 May 1];299(6). Available from: https://www.jbc.org/article/S0021-9258(23)01848-3/abstract.
- Masic A, Booth JS, Mutwiri GK, Babiuk LA, Zhou Y. Elastase-Dependent Live Attenuated Swine Influenza A Viruses Are Immunogenic and Confer Protection against Swine Influenza A Virus Infection in Pigs. Journal of Virology. 2009 Oct;83(19):10198–210. [CrossRef]
- Bugge TH, Antalis TM, Wu Q. Type II Transmembrane Serine Proteases. J Biol Chem. 2009 Aug 28;284(35):23177–81.
- Béliveau F, Désilets A, Leduc R. Probing the substrate specificities of matriptase, matriptase-2, hepsin and DESC1 with internally quenched fluorescent peptides. The FEBS Journal. 2009;276(8):2213–26. [CrossRef]
- Zmora P, Blazejewska P, Moldenhauer AS, Welsch K, Nehlmeier I, Wu Q, et al. DESC1 and MSPL Activate Influenza A Viruses and Emerging Coronaviruses for Host Cell Entry. Journal of Virology. 2014 Oct 15;88(20):12087–97. [CrossRef]
- Yasuoka S, Ohnishi T, Kawano S, Tsuchihashi S, Ogawara M, Masuda K, et al. Purification, characterization, and localization of a novel trypsin-like protease found in the human airway. Am J Respir Cell Mol Biol. 1997 Mar;16(3):300–8. [CrossRef]
- Wysocka M, Spichalska B, Lesner A, Jaros M, Brzozowski K, Łęgowska A, et al. Substrate specificity and inhibitory study of human airway trypsin-like protease. Bioorganic & Medicinal Chemistry. 2010 Aug 1;18(15):5504–9. [CrossRef]
- Böttcher E, Matrosovich T, Beyerle M, Klenk HD, Garten W, Matrosovich M. Proteolytic Activation of Influenza Viruses by Serine Proteases TMPRSS2 and HAT from Human Airway Epithelium. Journal of Virology. 2006 Oct;80(19):9896–8. [CrossRef]
- Bertram S, Dijkman R, Habjan M, Heurich A, Gierer S, Glowacka I, et al. TMPRSS2 Activates the Human Coronavirus 229E for Cathepsin-Independent Host Cell Entry and Is Expressed in Viral Target Cells in the Respiratory Epithelium. Journal of Virology. 2013 Jun;87(11):6150–60. [CrossRef]
- Bertram S, Glowacka I, Müller MA, Lavender H, Gnirss K, Nehlmeier I, et al. Cleavage and Activation of the Severe Acute Respiratory Syndrome Coronavirus Spike Protein by Human Airway Trypsin-Like Protease. Journal of Virology. 2011 Dec 15;85(24):13363–72. [CrossRef]
- Beaulieu A, Gravel É, Cloutier A, Marois I, Colombo É, Désilets A, et al. Matriptase Proteolytically Activates Influenza Virus and Promotes Multicycle Replication in the Human Airway Epithelium. Journal of Virology. 2013 Apr 15;87(8):4237–51. [CrossRef]
- Whittaker GR, Straus MR. Human matriptase/ST 14 proteolytically cleaves H7N9 hemagglutinin and facilitates the activation of influenza A/Shanghai/2/2013 virus in cell culture. Influenza Other Respir Viruses. 2020 Mar;14(2):189–95.
- Zmora P, Hoffmann M, Kollmus H, Moldenhauer AS, Danov O, Braun A, et al. TMPRSS11A activates the influenza A virus hemagglutinin and the MERS coronavirus spike protein and is insensitive against blockade by HAI-1. Journal of Biological Chemistry. 2018 Sep 1;293(36):13863–73. [CrossRef]
- Kido H, Okumura Y. MSPL/TMPRSS13. FBL. 2008 Jan 1;13(2):754–8.
- Kishimoto M, Uemura K, Sanaki T, Sato A, Hall WW, Kariwa H, et al. TMPRSS11D and TMPRSS13 Activate the SARS-CoV-2 Spike Protein. Viruses. 2021 Mar;13(3):384.
- Yun Kim S, Park D, Oh M, Sellamuthu S, Park WJ. Detection of site-specific proteolysis in secretory pathways. Biochemical and Biophysical Research Communications. 2002 Aug 16;296(2):419–24.
- Mahmoud IS, Jarrar YB, Alshaer W, Ismail S. SARS-CoV-2 entry in host cells-multiple targets for treatment and prevention. Biochimie. 2020 Aug 1;175:93–8.
- Shirogane Y, Takeda M, Iwasaki M, Ishiguro N, Takeuchi H, Nakatsu Y, et al. Efficient Multiplication of Human Metapneumovirus in Vero Cells Expressing the Transmembrane Serine Protease TMPRSS2. Journal of Virology. 2008 Sep;82(17):8942–6. [CrossRef]
- Iwata-Yoshikawa N, Okamura T, Shimizu Y, Hasegawa H, Takeda M, Nagata N. TMPRSS2 Contributes to Virus Spread and Immunopathology in the Airways of Murine Models after Coronavirus Infection. J Virol. 2019 Mar 5;93(6):e01815-18. [CrossRef]
- Abe M, Tahara M, Sakai K, Yamaguchi H, Kanou K, Shirato K, et al. TMPRSS2 Is an Activating Protease for Respiratory Parainfluenza Viruses. J Virol. 2013 Nov;87(21):11930–5. [CrossRef]
- Esram P, Arumugam P. Development and Validation of an Enzymatic Assay for TMPRSS4: Evaluation of Molecular Inhibitors. Journal of Advanced Zoology. 2023 Oct 9;44(3):309–21. [CrossRef]
- Kühn N, Bergmann S, Kösterke N, Lambertz RLO, Keppner A, van den Brand JMA, et al. The Proteolytic Activation of (H3N2) Influenza A Virus Hemagglutinin Is Facilitated by Different Type II Transmembrane Serine Proteases. Journal of Virology. 2016 Apr 14;90(9):4298–307. [CrossRef]
- Zang R, Gomez Castro MF, McCune BT, Zeng Q, Rothlauf PW, Sonnek NM, et al. TMPRSS2 and TMPRSS4 promote SARS-CoV-2 infection of human small intestinal enterocytes. Sci Immunol. 2020 May 13;5(47):eabc3582.
- Epstein RJ. The secret identities of TMPRSS2: Fertility factor, virus trafficker, inflammation moderator, prostate protector and tumor suppressor. Tumor Biology. 2021 Jan 1;43(1):159–76. [CrossRef]
- Murza A, Dion SP, Boudreault PL, Désilets A, Leduc R, Marsault É. Inhibitors of type II transmembrane serine proteases in the treatment of diseases of the respiratory tract – A review of patent literature. Expert Opinion on Therapeutic Patents. 2020 Nov 1;30(11):807–24. [CrossRef]
- Heurich A, Hofmann-Winkler H, Gierer S, Liepold T, Jahn O, Pöhlmann S. TMPRSS2 and ADAM17 Cleave ACE2 Differentially and Only Proteolysis by TMPRSS2 Augments Entry Driven by the Severe Acute Respiratory Syndrome Coronavirus Spike Protein. J Virol. 2014 Jan;88(2):1293–307. [CrossRef]
- Kirschke H. Chapter 410 - Cathepsin L. In: Rawlings ND, Salvesen G, editors. Handbook of Proteolytic Enzymes (Third Edition) [Internet]. Academic Press; 2013 [cited 2023 Jul 13]. p. 1808–17. Available from: https://www.sciencedirect.com/science/article/pii/B9780123822192004105.
- Yadati T, Houben T, Bitorina A, Shiri-Sverdlov R. The Ins and Outs of Cathepsins: Physiological Function and Role in Disease Management. Cells. 2020 Jul 13;9(7):1679. [CrossRef]
- Du X, Tang H, Gao L, Wu Z, Meng F, Yan R, et al. Omicron adopts a different strategy from Delta and other variants to adapt to host. Sig Transduct Target Ther. 2022 Feb 10;7(1):1–3. [CrossRef]
- Willett BJ, Grove J, MacLean OA, Wilkie C, De Lorenzo G, Furnon W, et al. SARS-CoV-2 Omicron is an immune escape variant with an altered cell entry pathway. Nat Microbiol. 2022 Aug;7(8):1161–79.
- Mykytyn AZ, Breugem TI, Geurts MH, Beumer J, Schipper D, van Acker R, et al. SARS-CoV-2 Omicron entry is type II transmembrane serine protease-mediated in human airway and intestinal organoid models. J Virol. 97(8):e00851-23.
- Scarcella M, d’Angelo D, Ciampa M, Tafuri S, Avallone L, Pavone LM, et al. The Key Role of Lysosomal Protease Cathepsins in Viral Infections. Int J Mol Sci. 2022 Aug 13;23(16):9089. [CrossRef]
- Mainou BA. The Orchestra of Reovirus Cell Entry. Curr Clin Micro Rpt. 2017 Sep 1;4(3):142–9. [CrossRef]
- Baer GS, Dermody TS. Mutations in reovirus outer-capsid protein sigma3 selected during persistent infections of L cells confer resistance to protease inhibitor E64. Journal of Virology. 1997 Jul;71(7):4921–8. [CrossRef]
- Wilson GJ, Nason EL, Hardy CS, Ebert DH, Wetzel JD, Venkataram Prasad BV, et al. A Single Mutation in the Carboxy Terminus of Reovirus Outer-Capsid Protein σ3 Confers Enhanced Kinetics of σ3 Proteolysis, Resistance to Inhibitors of Viral Disassembly, and Alterations in σ3 Structure. Journal of Virology. 2002 Oct;76(19):9832–43.
- Doyle JD, Danthi P, Kendall EA, Ooms LS, Wetzel JD, Dermody TS. Molecular Determinants of Proteolytic Disassembly of the Reovirus Outer Capsid *. Journal of Biological Chemistry. 2012 Mar 9;287(11):8029–38. [CrossRef]
- Mainou BA, Dermody TS. In Search of Cathepsins: How Reovirus Enters Host Cells. DNA Cell Biol. 2012 Dec;31(12):1646–9. [CrossRef]
- Schornberg K, Matsuyama S, Kabsch K, Delos S, Bouton A, White J. Role of Endosomal Cathepsins in Entry Mediated by the Ebola Virus Glycoprotein. Journal of Virology. 2006 Apr 15;80(8):4174–8. [CrossRef]
- Diederich S, Sauerhering L, Weis M, Altmeppen H, Schaschke N, Reinheckel T, et al. Activation of the Nipah virus fusion protein in MDCK cells is mediated by cathepsin B within the endosome-recycling compartment. J Virol. 2012 Apr;86(7):3736–45. [CrossRef]
- Vogt C, Eickmann M, Diederich S, Moll M, Maisner A. Endocytosis of the Nipah Virus Glycoproteins. J Virol. 2005 Mar;79(6):3865–72. [CrossRef]
- Corry J, Johnson SM, Cornwell J, Peeples ME. Preventing Cleavage of the Respiratory Syncytial Virus Attachment Protein in Vero Cells Rescues the Infectivity of Progeny Virus for Primary Human Airway Cultures. J Virol. 2016 Jan 15;90(3):1311–20. [CrossRef]
- Biniossek ML, Nägler DK, Becker-Pauly C, Schilling O. Proteomic Identification of Protease Cleavage Sites Characterizes Prime and Non-prime Specificity of Cysteine Cathepsins B, L, and S. J Proteome Res. 2011 Dec 2;10(12):5363–73.
- Simmons G, Gosalia DN, Rennekamp AJ, Reeves JD, Diamond SL, Bates P. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc Natl Acad Sci USA. 2005 Aug 16;102(33):11876–81. [CrossRef]
- Kawase M, Shirato K, van der Hoek L, Taguchi F, Matsuyama S. Simultaneous Treatment of Human Bronchial Epithelial Cells with Serine and Cysteine Protease Inhibitors Prevents Severe Acute Respiratory Syndrome Coronavirus Entry. Journal of Virology. 2012 Jun 15;86(12):6537–45. [CrossRef]
- Belouzard S, Chu VC, Whittaker GR. Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites. Proceedings of the National Academy of Sciences. 2009 Apr 7;106(14):5871–6. [CrossRef]
- Zhao MM, Yang WL, Yang FY, Zhang L, Huang WJ, Hou W, et al. Cathepsin L plays a key role in SARS-CoV-2 infection in humans and humanized mice and is a promising target for new drug development. Sig Transduct Target Ther. 2021 Mar 27;6(1):1–12.
- Ebert DH, Deussing J, Peters C, Dermody TS. Cathepsin L and Cathepsin B Mediate Reovirus Disassembly in Murine Fibroblast Cells *. Journal of Biological Chemistry. 2002 Jul 5;277(27):24609–17. [CrossRef]
- Kleine-Weber H, Elzayat MT, Hoffmann M, Pöhlmann S. Functional analysis of potential cleavage sites in the MERS-coronavirus spike protein. Sci Rep. 2018 Nov 9;8(1):16597. [CrossRef]
- Pager CT, Craft WW, Patch J, Dutch RE. A mature and fusogenic form of the Nipah virus fusion protein requires proteolytic processing by cathepsin L. Virology. 2006 Mar 15;346(2):251–7. [CrossRef]
- Coleman MD, Ha SD, Haeryfar SMM, Barr SD, Kim SO. Cathepsin B plays a key role in optimal production of the influenza A virus. J Virol Antivir Res. 2018 Apr;7(1):1–20. [CrossRef]
- Bollavaram K, Leeman TH, Lee MW, Kulkarni A, Upshaw SG, Yang J, et al. Multiple sites on SARS-CoV-2 spike protein are susceptible to proteolysis by cathepsins B, K, L, S, and V. Protein Science. 2021;30(6):1131–43.
- Golden JW, Bahe JA, Lucas WT, Nibert ML, Schiff LA. Cathepsin S Supports Acid-independent Infection by Some Reoviruses *. Journal of Biological Chemistry. 2004 Mar 2;279(10):8547–57. [CrossRef]
- Brinkworth RI, Tort JF, Brindley PJ, Dalton JP. Phylogenetic relationships and theoretical model of human cathepsin W (lymphopain), a cysteine proteinase from cytotoxic T lymphocytes. The International Journal of Biochemistry & Cell Biology. 2000 Mar 1;32(3):373–84. [CrossRef]
- Edinger TO, Pohl MO, Yángüez E, Stertz S. Cathepsin W Is Required for Escape of Influenza A Virus from Late Endosomes. mBio. 2015 Jun 9;6(3):e00297.
- Brömme D. Chapter 409 - Cathepsin K. In: Rawlings ND, Salvesen G, editors. Handbook of Proteolytic Enzymes (Third Edition) [Internet]. Academic Press; 2013 [cited 2024 May 4]. p. 1801–7. Available from: https://www.sciencedirect.com/science/article/pii/B9780123822192004099.
- Brömme D. Chapter 414 - Cathepsin V. In: Rawlings ND, Salvesen G, editors. Handbook of Proteolytic Enzymes (Third Edition) [Internet]. Academic Press; 2013 [cited 2024 May 4]. p. 1831–4. Available from: https://www.sciencedirect.com/science/article/pii/B9780123822192004130.
- Cabral-Pacheco GA, Garza-Veloz I, Castruita-De la Rosa C, Ramirez-Acuña JM, Perez-Romero BA, Guerrero-Rodriguez JF, et al. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int J Mol Sci. 2020 Dec 20;21(24):9739. [CrossRef]
- Yeo SJ, Yun YJ, Lyu MA, Woo SY, Woo ER, Kim SJ, et al. Respiratory syncytial virus infection induces matrix metalloproteinase-9 expression in epithelial cells. Arch Virol. 2002 Feb 1;147(2):229–42. [CrossRef]
- Rojas-Quintero J, Wang X, Tipper J, Burkett PR, Zuñiga J, Ashtekar AR, et al. Matrix metalloproteinase-9 deficiency protects mice from severe influenza A viral infection. JCI Insight. 2018 Dec 20;3(24):e99022, 99022. [CrossRef]
- Tacon CE, Wiehler S, Holden NS, Newton R, Proud D, Leigh R. Human Rhinovirus Infection Up-Regulates MMP-9 Production in Airway Epithelial Cells via NF-κB. Am J Respir Cell Mol Biol. 2010 Aug;43(2):201–9.
- Elliott MB, Welliver RC, Laughlin TS, Pryharski KS, LaPierre NA, Chen T, et al. Matrix metalloproteinase-9 and tissue inhibitor of matrix metalloproteinase-1 in the respiratory tracts of human infants following paramyxovirus infection. J Med Virol. 2007 Apr;79(4):447–56. [CrossRef]
- Phillips JM, Gallagher T, Weiss SR. Neurovirulent Murine Coronavirus JHM.SD Uses Cellular Zinc Metalloproteases for Virus Entry and Cell-Cell Fusion. J Virol. 2017 Mar 29;91(8):e01564-16. [CrossRef]
- Yamamoto M, Gohda J, Kobayashi A, Tomita K, Hirayama Y, Koshikawa N, et al. Metalloproteinase-Dependent and TMPRSS2-Independent Cell Surface Entry Pathway of SARS-CoV-2 Requires the Furin Cleavage Site and the S2 Domain of Spike Protein. mBio. 2022 Aug 30;13(4):e0051922.
- Chan JFW, Huang X, Hu B, Chai Y, Shi H, Zhu T, et al. Altered host protease determinants for SARS-CoV-2 Omicron. Science Advances. 2023 Jan 20;9(3):eadd3867.
- Yan R, Zhang Y, Li Y, Xia L, Guo Y, Zhou Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science. 2020 Mar 27;367(6485):1444–8.
- Yu S, Zheng X, Zhou B, Li J, Chen M, Deng R, et al. SARS-CoV-2 spike engagement of ACE2 primes S2′ site cleavage and fusion initiation. Proceedings of the National Academy of Sciences. 2022 Jan 4;119(1):e2111199119.
- Zhang H, Lv P, Jiang J, Liu Y, Yan R, Shu S, et al. Advances in developing ACE2 derivatives against SARS-CoV-2. Lancet Microbe. 2023 May;4(5):e369–78.
- Heindl MR, Rupp AL, Schwerdtner M, Bestle D, Harbig A, De Rocher A, et al. ACE2 acts as a novel regulator of TMPRSS2-catalyzed proteolytic activation of influenza A virus in airway cells. Subbarao K, editor. J Virol. 2024 Mar 12;e00102-24. [CrossRef]
- Tai W, He L, Zhang X, Pu J, Voronin D, Jiang S, et al. Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cell Mol Immunol. 2020 Jun;17(6):613–20. [CrossRef]
- Li W, Moore MJ, Vasilieva N, Sui J, Wong SK, Berne MA, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003 Nov 27;426(6965):450–4. [CrossRef]
- Hofmann H, Pyrc K, van der Hoek L, Geier M, Berkhout B, Pöhlmann S. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci U S A. 2005 May 31;102(22):7988–93. [CrossRef]
- Funk CJ, Wang J, Ito Y, Travanty EA, Voelker DR, Holmes KV, et al. Infection of human alveolar macrophages by human coronavirus strain 229E. J Gen Virol. 2012 Mar;93(Pt 3):494–503. [CrossRef]
- Hajizadeh M, Moosavi-Movahedi Z, Sheibani N, Moosavi-Movahedi AA. An outlook on suicide enzyme inhibition and drug design. Journal of the Iranian Chemical Society. 2022;19(5):1575–92. [CrossRef]
- Ben-Tal AK Nir. Introduction to Proteins: Structure, Function, and Motion, Second Edition. 2nd ed. New York: Chapman and Hall/CRC; 2018. 988 p.
- Rajendran L, Knölker HJ, Simons K. Subcellular targeting strategies for drug design and delivery. Nat Rev Drug Discov. 2010 Jan;9(1):29–42. [CrossRef]
- Donahue ND, Acar H, Wilhelm S. Concepts of nanoparticle cellular uptake, intracellular trafficking, and kinetics in nanomedicine. Advanced Drug Delivery Reviews. 2019 Mar 15;143:68–96. [CrossRef]
- Lotke R, Petersen M, Sauter D. Restriction of Viral Glycoprotein Maturation by Cellular Protease Inhibitors. Viruses. 2024 Feb 22;16(3):332. [CrossRef]
- Stein P, Chothia C. Serpin tertiary structure transformation. Journal of Molecular Biology. 1991 Sep 20;221(2):615–21. [CrossRef]
- Nichols DB, De Martini W, Cottrell J. Poxviruses Utilize Multiple Strategies to Inhibit Apoptosis. Viruses. 2017 Aug;9(8):215. [CrossRef]
- Tarighi P, Eftekhari S, Chizari M, Sabernavaei M, Jafari D, Mirzabeigi P. A review of potential suggested drugs for coronavirus disease (COVID-19) treatment. Eur J Pharmacol. 2021 Mar 15;895:173890. [CrossRef]
- Wyde PR, Chetty SN, Jewell AM, Boivin G, Piedra PA. Comparison of the inhibition of human metapneumovirus and respiratory syncytial virus by ribavirin and immune serum globulin in vitro. Antiviral Res. 2003 Sep;60(1):51–9. [CrossRef]
- Thomas G, Couture F, Kwiatkowska A. The Path to Therapeutic Furin Inhibitors: From Yeast Pheromones to SARS-CoV-2. International Journal of Molecular Sciences. 2022 Jan;23(7):3435.
- Essalmani R, Jain J, Susan-Resiga D, Andréo U, Evagelidis A, Derbali RM, et al. Distinctive Roles of Furin and TMPRSS2 in SARS-CoV-2 Infectivity. J Virol. 96(8):e00128-22.
- 125. Someya A, Tanaka N, Okuyama A. Inhibition of influenza virus AWSN replication by a trypsin inhibitor, 6-amidino-2-naphthyl p-guanidinobenzoate. Biochemical and Biophysical Research Communications. 1990 May 31;169(1):148–52. [CrossRef]
- Hosoya M, Matsuyama S, Baba M, Suzuki H, Shigeta S. Effects of protease inhibitors on replication of various myxoviruses. Antimicrob Agents Chemother. 1992 Jul;36(7):1432–6. [CrossRef]
- Hosoya M, Shigeta S, Ishii T, Suzuki H, Clercq ED. Comparative Inhibitory Effects of Various Nucleoside and Nonnucleoside Analogues on Replication of Influenza Virus Types A and B In Vitro and In Ovo. The Journal of Infectious Diseases. 1993 Sep 1;168(3):641–6. [CrossRef]
- Gunst JD, Staerke NB, Pahus MH, Kristensen LH, Bodilsen J, Lohse N, et al. Efficacy of the TMPRSS2 inhibitor camostat mesilate in patients hospitalized with Covid-19-a double-blind randomized controlled trial. EClinicalMedicine. 2021 May;35:100849. [CrossRef]
- Zhirnov OP, Klenk HD, Wright PF. Aprotinin and similar protease inhibitors as drugs against influenza. Antiviral Res. 2011 Oct;92(1):27–36. [CrossRef]
- Grocott HP, Sheng H, Miura Y, Sarraf-Yazdi S, Mackensen GB, Pearlstein RD, et al. The effects of aprotinin on outcome from cerebral ischemia in the rat. Anesth Analg. 1999 Jan;88(1):1–7.
- Mangano DT, Tudor IC, Dietzel C, Multicenter Study of Perioperative Ischemia Research Group, Ischemia Research and Education Foundation. The risk associated with aprotinin in cardiac surgery. N Engl J Med. 2006 Jan 26;354(4):353–65.
- Hamilton BS, Chung C, Cyphers SY, Rinaldi VD, Marcano VC, Whittaker GR. Inhibition of influenza virus infection and hemagglutinin cleavage by the protease inhibitor HAI-2. Biochem Biophys Res Commun. 2014 Jul 25;450(2):1070–5. [CrossRef]
- Straus MR, Kinder JT, Segall M, Dutch RE, Whittaker GR. SPINT2 inhibits proteases involved in activation of both influenza viruses and metapneumoviruses. Virology. 2020 Apr;543:43–53. [CrossRef]
- Shapira T, Monreal IA, Dion SP, Buchholz DW, Imbiakha B, Olmstead AD, et al. A TMPRSS2 inhibitor acts as a pan-SARS-CoV-2 prophylactic and therapeutic. Nature. 2022;605(7909):340–8.
Figure 1.
Substrate (left) cleaved by protease (right) with the scissile bond marked by an arrow.
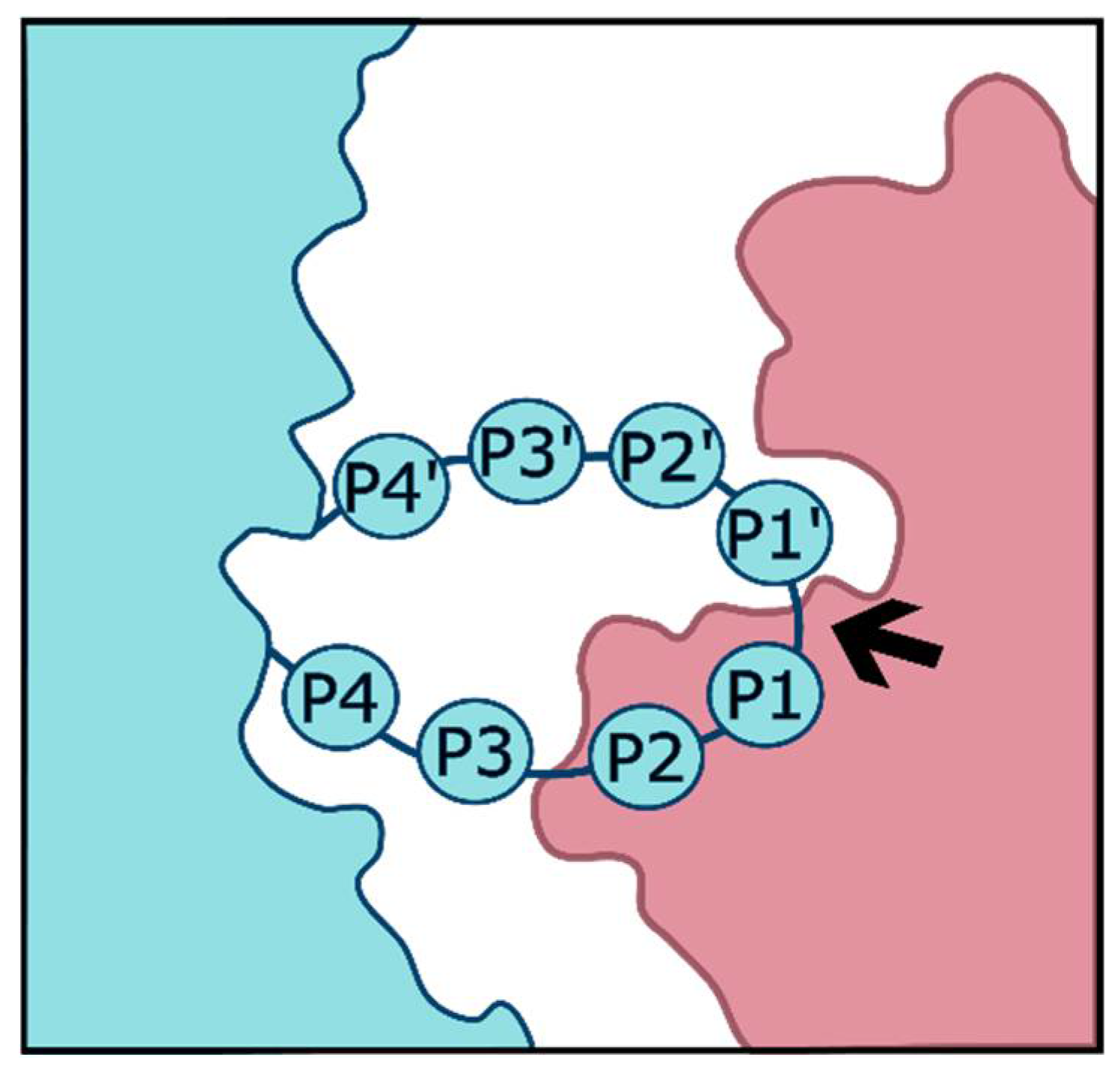
Table 1.
Characteristics of well-known serine proteases outside of the TTSP family.
| Serine Proteases | Cleavage Preferences | Exploiting Respiratory Viruses |
|---|---|---|
| Chymase | Aromatic amino acids at P1, aliphatic amino acids from P2-P4, S at P1’, E/D at P2’, A/V/G at P3’ [5] | Mammalian Orthoreovirus [6] |
| Factor Xa | Preference for PXG/AR↓XXD [7] | SARS-CoV-1 [8], SARS-CoV-2 [9] |
| Furin | R-X-X-R↓S [10] | Influenza virus [11], SARS-CoV-2 [12], MERS-CoV[13] , RSV [14], HPIV [15], Measles virus (MeV) [16], Mumps virus [17] |
| kallikrein-related peptidase 13 (KLK13) | V/Y-R/L/F/M-R↓ [18] | HKU-1 [19] |
| KLK1 | R/Y↓S/R [20] | Influenza virus [21] |
| KLK5 | X(aliphatic/aromatic)-R/K -X(polar/aliphatic)- R↓[18] | Influenza virus [21] |
| Neutrophil Elastase | A/V/I/T↓ [22] | Mammalian Orthoreovirus [23], SARS-CoV-1 [24,25] |
| PC5/6 | R-X-R/K-R↓ [26] | Influenza virus [27], RSV [27] |
| Plasmin | R/L↓ [28], preference for aromatic hydrophobic residue at P2 [29] | Influenza virus [30], RSV[31], SARS-CoV-1 [32], SARS-CoV-2 [33] |
| Thrombin | L-X-P-R↓S/A/G/T-X(aromatic)-R [34] | RSV [31], SARS-CoV-2 [9] |
| Trypsin | K/R↓[22] | SARS-CoV-1 [35,36], HMPV [37], RSV [31], HPIV [38] |
| Cathepsin G | Preference for F, Y, W, or L at P1 [39], S at P6, negatively charged amino acid in P2’ position | Mammalian Orthoreovirus [6], RSV [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



