
Submitted:
17 May 2024
Posted:
20 May 2024
You are already at the latest version
Abstract
Fusarium species are agriculturally important fungi with a broad host range and can be found as endophytic, pathogenic, or opportunistic parasites in many crop plants. This study aimed to identify Fusarium species in bare-rooted, dormant plants in Turkish grapevine nurseries using molecular identification methods and assess their pathogenicity. Asymptomatic dormant plants were sampled from grapevine nurseries (43) in different regions of the country, and fungi were isolated from plant roots and internal basal tissues. The Fusarium isolates were identified by performing gene sequencing (TEF1-α, RPB2) and phylogenetic analyses. Pathogenicity tests were carried out by inoculating mycelial agar pieces of isolates onto the stem or conidial suspensions into the rhizosphere of vines (1103 Paulsen rootstock). Laboratory tests revealed that Fusarium species were highly prevalent in Turkish grapevine nurseries (41 out of 43). Gene sequencing and phylogenetic analyses unraveled that 12 Fusarium species (F. annulatum, F. brachygibbosum, F. clavum, F. curvatum, F. falciforme, F. fredkrugeri, F. glycines, F. nanum, F. nematophilum, F. nirenbergiae, F. solani, and Fusarium spp.) existed in the ready-to-sale plants. Some of these species (F. annulatum, F. curvatum and F. nirenbergiae) consistently caused wood necrosis of seedling stems, rotting of the basal zone and roots, and reduced root biomass. Although the other nine species also caused some root rot and root reduction, their virulence was not as severe as the pathogenic ones, and they were considered opportunistic parasites or endophytic species. This study is the first detailed investigation of Fusarium species in grapevine nurseries in Türkiye and emphasizes the need to consider pathogenic species in producing healthy grapevine seedlings.
Keywords:
1103 Paulsen
; Fusarium
; grape
; identification
; pathogenicity
; rootstock
; Türkiye
1. Introduction
Grapevine sapling production is one of the most important agricultural sectors in Türkiye, and many young vines (2.5 to 3 million plants) are produced yearly in different geographical regions in the country [1]. The need for grapevine seedlings in the domestic market is relatively high, and this production needs to be increased to meet the demand for grapevine seedlings in Türkiye.
In grapevine nurseries, abiotic factors (unfavorable weather conditions, nutritional disorders, use of poor quality production materials, rootstock-scion incompatibility, etc.), nematodes, insects, and fungal pathogens cause the death of plants, and these factors bring about low productivity and economic losses every year. Fungal grapevine trunk disease (GTD) pathogens, which often settle on young seedlings with infected propagation materials, belonging to Botryosphaeriaceae, Diaporthaceae, and Diatrypaceae families, Cadophora, Cytospora, Phaeomoniella, Phaeoacremonium Seimatosporium genus, and soilborne fungi (Armillaria, Cylindrocarpon-like anamorphs, Fusarium, Macrophomina, Phytophthora, Rhizoctonia, and Verticillium sp.) are considered to be the main actors of plant mortality in the nurseries [2,3].
The genus Fusarium has an exceptional place in plant pathology, medical mycology, and the food industry as they are both plant and human pathogens and threaten human-animal health by producing mycotoxins in foods. To date, more than 400 Fusarium species have been identified, which nested in 23 different species complexes [4]. Most Fusarium species are soil-borne and are also called one of the ubiquitous fungal genera in mycology due to their endophytic, saprophytic, hemibiotrophic, or parasitic forms and strong competitive ability. Plant pathogenic species may result in significant crop damage and economic losses in some years by causing root and basal rots, damping-off, seed-tuber-fruit rots, wilt, and head blight diseases. According to the American Phytopathological Society, 83 out of 108 plant species in the field and horticultural crops are affected by one or more Fusarium diseases [5]. Many species in the Fusarium genus are true plant pathogens, while others are opportunists waiting for soil and environmental conditions to turn unfavorable for plants.
It has been pointed out that root rot-associated fungi considerably reduce young vine health and marketable sapling yield; Fusarium and Cylindrocarpon-like fungi were the main actors affecting plant vigor, and quality in grapevine nurseries. These fungi cause necrosis in the roots and basal tissues, leading to a reduction in hairy roots, retarded growth, and, the death of seedlings or young vines in later stages [6]. Research on the pathogenic roles and diversity of Fusarium species on plant death in grapevine nurseries and young vineyards has intensified in recent years. Highet and Nair [7] proved the infection of grapevine hairy roots by Fusarium oxysporum through transmission electron microscopy and pathogenicity tests and suggested it would be considered one of the fungi associated with root rot and decline in the nurseries. Reveglia et al. [8] revealed that the phytotoxins of Fusarium oxysporum (such as fusaric acid) and other potential metabolites have a critical role in the occurrence of these symptoms in seedlings and young vines in Australia. Vilvert et al. [9] claimed that Fusarium oxysporum f.sp. herbemontis is an important species responsible for decline and plant death in Brazilian grapevine nurseries, and it would be possible to control this pathogen using mycorrhizal fungi. Úrbez-Torres et al. [10] stated that Fusarium species were common in British Columbia (Canada) vineyards, but the most frequently isolated species might be as secondary pathogens on grapevine rootstock 3309C. Similarly, Bustamente et al. [11] suggested that Fusarium species isolated from grapevine nurseries and young vineyards in California (USA) are opportunistic pathogens attacking plants under stress. In contrast, Li et al. [12] reported that when the Fusarium isolates were inoculated into grapevine seedlings, they caused necrosis in the xylem vessels and basal regions of the plants resembling the infections of D. macrodidyma (a black-foot disease pathogen). Zhang et al. [13] reported for the first time Fusarium commune was a pathogen in grapevines causing leaf yellowing, stunting and root rot in Beijing Region, China. These studies indicate that Fusarium species on grapevines are a potential threat to nurseries and newly established vineyards and should not be underestimated. Furthermore, since these species can be found in the latent phase in plants, it is possible to spread them over large areas with marketable grapevine seedlings. Akgül and Ahioğlu [14] detected some Fusarium species in young vineyards, along with fungal pathogens associated with grapevine trunk diseases, in the southern Türkiye and confirmed the pathogenicity of these species. However, a nationwide study on the diversity and pathogenicity of Fusarium species in marketable grapevine saplings is needed. Therefore, this study aimed to reveal Fusarium diversity in dormant marketable plants in Turkish grapevine nurseries and to assess their pathogenicity.
2. Materials and Methods
2.1. Survey and Isolation of Fusarium Species
The survey was conducted in January 2021 in 43 grapevine nurseries in different geographical regions of Türkiye (in Adıyaman, Bursa, Denizli, Manisa, Mersin, Mersin, Tekirdağ, Tokat, and Urfa provinces). Ten dormant, commercially ready-for-sale seedlings from each nursery were randomly sampled and transported to the laboratory. The root and basal parts of the seedlings were washed under tap water and disinfected superficially with sodium hypochlorite solution (including >5% active clorine) for 3 minutes. Root and internal basal tissues (3-4 mm) were placed onto PDA (Potato dextrose agar, CondaLab; Spain) containing streptomycin-sulfate (250 mg×L-1), and the Petri plates were kept at 25°C in dark, for ten days to promote fungal colony growth. According to the morphological and microscopic characteristics detailed by Leslie and Summerell [15], a single spore was taken from the Fusarium colonies and purified on PDA for further stages. Ten Petri plates (containing seven tissue fragments in each) were used, and the isolation frequency of Fusarium colonies was calculated by proportioning the tissue number (Fusarium detected) to the total number (n=70).
2.2. Molecular Identification and Phylogenetic Analyses
A total of 60 Fusarium isolates were selected for molecular identification. They were grown on PDA at 25°C in the dark for seven days, and mycelium (56-60 mg) was harvested for DNA extraction. The genomic DNA was obtained following the CTAB protocol recommended by O'Donnell et al. [16] and diluted with 100 µl PCR grade water (Lonza) and stored at -18°C for further use. Translation elongation factor (TEF1-α) and second largest protein subunit of RNA polymerase II (RPB2) genes were amplified using the primers, EF1/EF2 and RPB2-5f2/fRPB2-7cr respectively [17]. The PCR reaction mixture contained 5 μl of buffer (10X Green Buffer, DreamTaq Green DNA Polymerase, Thermo-Scientific, USA), 2 μl of dNTPs mixture (10 mM each, Thermo Scientific, USA), 1 μl of forward and reverse primers (10 pmol·μl −1), 0.25 μl of Taq polymerase (DreamTaq Green DNA Polymerase, Thermo-Scientific), 39.75 μl PCR grade water and 1 μl genomic DNA (100 ng·μl −1). PCR amplifications were conducted in SimpliAmp A24811™ Thermal Cycler, Applied Biosystems, (USA) with the conditions detailed in the publications of O’Donnell et al. [16,17,18]. The PCR products were separated using gel electrophoresis in 1.5% agarose (Invitrogen) gel under 55V DC voltage, 250 mA current for 90 min. and were checked for DNA quality visually. After that, PCR products were sequenced bidirectionally via Sanger sequencing, derived chronogram files were trimmed from 3 and 5 prime with CLC main Workbench 5.5, and manual editing was done where necessary. Cleaned sequences were compared with those deposited in the NCBI GenBank database using the NCBIblastn suite (National Center for Biotechnology Information). Nucleotide sequences of TEF1-α and RPB2 genes were submitted to the NCBI GenBank, and the accession numbers were obtained. According to the nucleotide BLAST search results of TEF1-α and RPB2, a representative sequence dataset was used from the NCBI nucleotide database (Table 1) to perform the phylogenetic study. Constructed datasets for TEF1-α and RPB2 sequences were aligned individually via the ClustalW alignment tool in Geneious Prime 2019.1.3 software. After the alignment step, TEF1-α and RPB2 sequences were concatenated from end to end via Geneious Prime 2019.1.3 software for the multi-gene phylogenetic tree. Phylogenetic analyses were based on maximum likelihood (ML). The ML analysis was performed with IQ-TREE on the Galaxy Europe platform [19]. Model Finder was used to determine the best-fit model for the ML tree [20]. ML tree construction was performed under the TIM2e model with equal base frequencies and Invariable+Gamma with four categories (TIM2e+I+G4) nucleotide substitution model according to the Bayesian information criterion scores and weights (BIC and w-BIC). For the pseudoreplications of the ML tree, a 1000 ultrafast bootstrap parameter was used [21]. The alignment and the phylogenetic tree were deposited in TreeBASE under the study number S31385 (http://purl.org/phylo/treebase/phylows/study/TB2:S31385).
2.3. Pathogenicity Tests
Based on the identification results, 38 Fusarium isolates were selected for pathogenicity tests, and two types of inoculation methods were followed to evaluate from different aspects. In the first, the bark of the dormant cuttings was removed with a sterile cork-borer (3 mm), fresh mycelial agar discs of the isolates (10-day old) were placed on these wounds, and these points were wrapped with parafilm™. The cuttings were planted in the pots and grown in greenhouse conditions for four months. The inoculation points were scraped with a scalpel, and necrosis lengths in the wood tissues were measured and recorded [11]. Plants inoculated with an Ilyonectria liriodendri (black-foot disease pathogen) isolate (AFP115) were set as positive, and sterile agar-inoculated plants were set as healthy controls. In the second trial, 1103 Paulsen rootstock cuttings were planted in the plastic pots (0.85 L) containing sterile rooting mix (equal volumes of peat moss and perlite), and the pots were kept in lath house conditions (natural temperature, relative humidity and illumination). The Fusarium isolates were grown on PDA at 25°C for 15 days, and conidial suspensions (in sterile distilled water at 106 conidia·ml-1 concentration) were prepared using Thoma™ slides. Following root formation, conidial suspensions were poured into the root zone of the plants (20 ml per pot), and plants were grown lath house conditions for four months. The pathogenicity of the isolates was assessed based on root dry weight and necrosis length in the plants' basal zone (in wood tissues). The seedlings were uprooted from the pots, the roots were gently washed under tap water, and were harvested using a pruning shear. After briefly blotting with paper towels, the roots were held in a drying chamber at 65°C for 48 hours, then weighed using a precision balance, and weights were recorded [22]. Nevertheless, the bark of the cuttings was carefully peeled off with a knife, and the length of the necroses in the wood tissues was measured with a caliper. Pathogenicity tests were arranged according to the design of the randomized plot with six replications (two plants in a replicate), and twelve plant were used for each isolate. The trials were repeated once (2022 and 2023 years), and the data were subjected to statistical analysis. To clarify the virulence of each Fusarium species, an analysis of variance was performed again on the mean values of all isolates belonging to the same species.
2.4. Statistical Analyses
Analysis of variance (ANOVA) was performed on lengths of wood necrosis in basal parts and internodes of the stems and root dry weights. The data were checked for normality, and root square transformation was applied. Means were compared using Fisher’s least significant difference (LSD) test at the 5% significance level [23].
3. Results
3.1. Fungal Isolation and Prevalence of Fusarium Species
The first Fusarium colonies were aroused on the internal basal tissues and hairy roots of marketable, dormant grapevine plants after 5-6 days of incubation (at 24°C in the dark) in PDA media. Through morphological and microscopic examinations, 779 Fusarium colonies were detected in 3010 plant tissues (in 430 Petri dishes) plated for 43 grapevine nurseries. The isolation frequency of Fusarium species in these nurseries is shown in Table 2.
As shown in Table 1, Fusarium species were detected in 41 out of 43 grapevine nurseries, and the prevalence of these species in Turkish grapevine nurseries was calculated at 95.3%. The isolation frequency in nurseries ranged between 2.9 and 65.7%, while the overall average was 24.9%. Nevertheless, black foot, Petri disease pathogens, Botryosphaeriaceae fungi, Cytospora, Diaporthe, Truncatella species, and soil-borne plant pathogenic fungi (Macrophomina and Rhizoctonia) were also found (data not shown). Considering different geographical regions and morphological-microscopic characteristics, 121 Fusarium colonies were pre-selected for molecular-phylogenetic analyses.
3.2. Molecular Identification and Phylogenetic Analyses
Using the primers EF1/EF2 and RPB2-5f2/fRPB2-7cr, TEF1-α and RPB2 gene regions of the Fusarium isolates were amplified by end point PCR, and agarose gel electrophoresis revealed DNA bands with sizes ranging from 680 to 1600 bp (respectively). The initial approach for the identification of isolates relied on a blastn search of partial sequences of the TEF1-α and RPB2 gene. NCBIblastn search was performed with the nucleotide sequences of these regions, and the isolates were 99.2-100% similar to other Fusarium species in the GenBank. Afterwards, these sequences were aligned with the closest matching and nearly closest references obtained from GenBank (Table 1) for resolve the ambiguities. However, the NCBIblastn search results from the two gene regions of all isolates were not parallel, and the second gene region in some isolates matched with different Fusarium species. Yet, phylogenetic analyses conducted with concatenated nucleotide sequences clarified the ambiguity observed in the blastn results. The final phylogenetic dataset was contained a total of 150 taxa with 3040 nucleotide sites (89 references and 60 isolates) and Dactylonectria torresensis CBS 129086 as an outgroup at taxon level. Number of constant sites were 1787 (= 58.7829% of all sites) whereas there was no ambiguous constant sites. Number of parsimony informative sites and distinct site patterns were 1001 and 1598, respectivelly. Determining to the best fit model for ML tree, BIC score was found 43876.5755 whereas w-BIC score was 0.785. Total length of ML tree (sum of branch lengths) were 2.5465 whereas sum of internal branch lengths were 1.2670 (49.7559% of tree length) (data not shown in Figure 1).
In this study, 60 Fusarium isolates (Table 3) included in the phylogenetic analyses were clustered in six different species complexes, of which 38.3% were F. oxysporum (FOSC), 20% F. fujikuroi (FFSC), 18.3% F. solani (FSSC), 13.3% F. incarnatum-equiseti (FIESC), 6.7% F. sambucinum (FSAMSC) and 3.3% F. albidum. The Fusarium species clustered into these species complexes distributed in 12 species (Figure 1): F. annulatum (11 isolates-18.3%), F. brachygibbosum (4 isolates-6.7%), F. clavum (7 isolates- 11.6%) F. curvatum (12 isolates-20%), F. falciforme (one isolate-1.6%), F. fredkrugeri (one isolate-1.6%), F. glycines (3 isolates-5.0%), F. nanum (one isolate-1.6%), F. nematophilum (2 isolates-3.3%), F. nirenbergiae (2 isolates-3.3%), F. solani (10 isolates-16.7%), and Fusarium sp. (6 isolates-10.0%).
3.2. Pathogenicity of Fusarium Isolates and Species
In four-month pathogenicity tests, some Fusarium isolates inoculated on the plants' stems (considering the possibility of contamination to the vines through pruning wounds) produced lesions ranging from 5.9 to 12.0 mm (Figure 2). The lesion lengths of 10 of the 38 isolates inoculated on the plants in the first year were longer than those of the control and other isolates and statistically different.
The majority of these isolates belonged to the following species: F. annulatum (seven isolates), F. brachygibbosum (one isolate), F. nirenbergiae (one isolate), and I. liriodendri. In the second year, only 9 isolates had lesions longer than other Fusarium isolates and control statistically. They were F. annulatum (five isolates), F. nirenbergiae (two isolates), F. curvatum (one isolate) and I. liriodendri (Table 4). In both years, lesions caused by other Fusarium isolates were not significantly longer than those of the control. However, these isolates could be re-isolated from the point of inoculation (except from the non-inoculated control) at rates ranging from 15.2% to 53.8%.
In the second type of pathogenicity test, 38 isolates increased basal rot and significantly reduced root dry weight compared to the non- inoculated control. Due to the large number of isolates and replicates, there was a large variance among the means of isolates, resulting in many statistical groups. The Fusarium isolates, and I. liriodenri caused basal rot of the wood tissues in the basal region of the seedlings (Figure 3), and their lengths ranged from 3.6 to 37.0 mm in the first year and from 3.7 to 7.8 mm in the second year (Table 5). As in the case of stem necrosis, F. annulatum, F. curvatum, and I. liriodendri species were found to cause the most extended necrose length in basal rot formation. However, the wood necrosis induced by the other species was not as severe and consistent as that of these three species. The Fusarium isolates and I. liriodendri could be re-isolated from the basal necroses (except from the non-inoculated control) at rates ranging from 10.3% to 36.8%.
In parallel to basal rot, the Fusarium isolates and I. liriodendri decreased hair root formation and root dry weight in the inoculated plants compared with the non-inoculated control plants. In 30-35% of the plants inoculated with F. annulatum, F. curvatum, and I. liriodendri, shoots dried up, and plants died after one month. The average root dry weight recorded per plant in the first year varied between 0.022 and 0.344 g, while in the second year, these values were recorded between 0.599 and 1.463 g (Table 6).
The average values (necrose lengths in stem and basal part, and dry root weights) of the isolates (belonging to the same species) were considered, and analysis of variance was performed to the means to clarify the pathogenicity of each Fusarium species. When the pathogenicity of Fusarium species was evaluated according to the length of necrosis at the inoculation point, it was found that F. annulatum, F. brachygibbosum, F. nirenbergiae and Ilyonectria liriodendri caused wood necrosis. In contrast, the others did not show the same influence (Figure 4). Regarding the effects of Fusarium species on basal rot formation, it was determined that the three species causing the most extended necrosis in the first year were F. annulatum, I. liriodendri, and F. glycines; in the second year, I. liriodendri, F. nirenbergiae, and F. annulatum, respectively (Figure 5).
The effect of Fusarium species on root dry weight reduction was almost parallel to basal rot; when the results of both years were generally evaluated, F. annulatum, I. liriodendri, and F. nirenbergiae were found to be the most effective species (Figure 6).
4. Discussion
In this study, Fusarium fungi were found to be relatively common (95.3% of the nurseries) in bare-rooted plants ready for sale in Turkish grapevine nurseries. This rate is considerably higher than that found in North America and Canada, but it is close to that in the nurseries in France and Spain. Garnett et al. [24] investigated the fungal species associated with root rot of grapevines in two different vineyards in California and isolated a high proportion of Fusarium species (together with Rhizoctonia, Pythium, Macrophomina, Phytophthora fungi) in the sampled vines. Torres et al. [10] found Fusarium species in 43.9% of the seedlings ready for sale in four grapevine nurseries in Canada and determined that these species were isolated from the plants between 20.0 and 86.7%. Bustamente et al. [11] determined that the incidence of Fusarium species was 36.7% in young vineyards and 31.7% in nursery plants in California. However, Pintos et al., [25] detected 92% to 98% of Fusarium fungi, among other GTD pathogens, from plants sampled from two commercial grapevine nurseries in Spain and one in France.
In the current study, 12 distinct Fusarium species were found in six Fusarium species complexes in the grapevine nurseries, with the most common species complexes being F. oxysporum (38.3%), F. fujikuroi (20.0%), and F. solani (18.3%). The results revealed more diversity of Fusarium species than previous studies conducted in Canada and United States. Urbez-Torres et al. [10] reported that Fusarium species diversity was very low in four nurseries in British Columbia (Canada) and found only two species from two different species complexes (F. oxysporum and F. fujikuroi). However, Bustamente et al. [11] reported a high diversity of Fusarium in young vines in California (total, nine Fusarium species in six species complexes) and found five species (F. annulatum, Fusarium sp., F. solani, F. keratoplasticum, F. nirenbergiae) belonging to these complexes in the nurseries. The fungal isolation results in our study, the high Fusarium species diversity in ready-to-sale grapevine seedlings, and the presence of joint species (F. annulatum, F. brachygibbosum, F. clavum, F. nirenbergiae, F. solani) in the plants were consistent with the findings of Bustamente et al. [11]. However, in the abovementioned studies, F. avenaceum, F. ramigenum, F. culmorum, F. keratoplasticum, F. oxysporum, and F. proliferatum were not found in grapevine nurseries in Türkiye. Interestingly, although F. oxysporum is a large species complex, including 21 species [26], and F. oxysporum has an essential place in this complex, we could not detect F. oxysporum among the Fusarium species we isolated from vines. When phylogenetic analyses were performed, it was revealed that many isolates similar to this species were F. curvatum, F. glycines, and F. nirenbergiae. Similarly, although F. proliferatum has been reported as an important root rot pathogen in maize, soybean, tomato, and grapevine [10,27,28,29], we could not detect F. proliferatum among the available grapevine Fusarium isolates. These differences may have been made possible by detailed phylogenetic analyses using concatenated genes such as TEF1-α and RPB2, which are highly recommended to identify Fusarium. O’Donnell et al. [4] suggested that when identifying Fusarium species, the TEF1α and RPB2 gene regions should be amplified and concatenated to perform phylogenetic analyses if possible, and in case of financial limitations, the sequence of the TEF1α region might be sufficient.
Based on the pathogenicity results of Fusarium isolates inoculated on grapevine stems, F. annulatum, F. brachygibbosum, F. curvatum, and F. nirenbergiae were found to produce more considerable wood necrosis in comparison to the control and other Fusarium species. Reveglia et al. [7] widely isolated F. oxysporum isolates from grapevines showing young vine decline symptoms (in Italy) and investigated their phytotoxic metabolites. The fusaric acid purified from these isolates caused severe necrosis when injected into tobacco leaves, and they suggested that this metabolite may also cause root and basal rot in grapevines. Akgül and Ahioğlu [14] inoculated F. brachygibbosum isolates (obtained from three-year-old young grapevines) on the stems of grapevine seedlings and determined it to be a highly virulent species in woody tissues. Rajput et al. [30] investigated the pathogenicity of F. equiseti isolates isolated from the trunks of grapevines in the Kunar province of Afghanistan and reported that tissue necrosis occurred when this species was inoculated on woody shoots of three-year-old plants. Bustamente et al. [11] inoculated F. annulatum, F. nirenbergiae, and F. solani isolates from young grapevines on the stems of one-year-old vines and revealed that after seven months; these species produced longer necroses in the wood tissues of plants when compared to non-inoculated controls. The results of these studies support the view that the wounds occurring via disbudding of cuttings or basal cuts in the seedlings or wounds by removing vine suckers on trunks (in the vineyards) may be susceptible to Fusarium infections and that Fusarium species may be involved in young vine decline or trunk diseases.
Another outcome from the pathogenicity tests was that some Fusarium species (F. annulatum, F. curvatum, F. nirenbergiae, F. solani) significantly increased basal rot and reduced root biomass in the inoculated plants. Highet and Nair [7] investigated the effect of Fusarium oxysporum infections on root rot development in grapevines (cv. Semillon 5-25 years old) in New Zealand. When plants were inoculated with F. oxysporum, they observed the disintegration of bark cells (by transmission electron microscopy) and determined that Fusarium-infected root cells lacked cytoplasm compared to uninfected cells. Vilvert et al. [9] stated that F. oxysporum f.sp. herbemontis was an important fungal pathogen in Brazilian grapevine nurseries, causing basal rot, reduction in root biomass, and root rot symptoms in infected vines. Zhang et al. [13] detected several Fusarium species from young grapevines (cv. Red Globe) showing decline and leaf yellowing in vineyards in Beijing, China and found that F. commune was pathogenic among these species and associated with these symptoms. Li et al. [12] revealed that Fusarium isolates inoculated on grapevine seedlings caused not only a reduction in root biomass, root rot but also interveinal discolorations and coalescent necrosis on the leaves of the plant. When these isolates were inoculated together with Dactylonectria macrodidyma, the severity of the disease was further increased.
Regarding the pathogenicity of other Fusarium species, F. annulatum has also been reported to cause fruit-corm and root rot in crop plants such as melon and onion, as well as medicinal-aromatic plants such as Blettila striata L. and saffron, in addition to grapevine. [31,32,33,34]. These studies indicate that F. annulatum is pathogenic in many hosts. F. nirenbergiae and F. curvatum were other virulent species in the pathogenicity tests on the grapevine seedlings. When we reviewed the studies on this subject, we found only one study [11] in which F. nirenbergiae was detected as a pathogen in grapevine. However, in other studies, it has been reported to be pathogenic in crop plants such as maize, passion fruit, almond (in Portugal and Spain), and maple. Sanna et al. [29] investigated Fusarium species associated with post-emergence damping-off and root rot in maize and found that F. nirenbergiae caused a disease index of over 50% in some maize areas of Italy, as did F. verticilloides, F. annulatum and F. commune. Aiello et al. [35] identified F. nirenbergiae as the cause of root rot and wilt in passion fruit plants. Zhao et al. [36] identified it as the cause of wilt in maple trees (in China), and Moral-Lopez et al., [37] in almonds (in Portugal and Spain). Another virulent species, F. curvatum, which we identified in pathogenicity tests, has previously been found to be associated with dieback disease in Dendrobium officinale [38] and leaf spot of cherry [39] in China.
In this study, although other Fusarium species (F. clavum, F. falciforme, F. fredkrugeri, F. glycines, F. nanum, F. nematophilum, F. solani) from Turkish grapevine nurseries reduced root biomass in grapevine seedlings compared to the control, their virulence was not as consistent as F. annulatum, F. brachygibbosum, F. curvatum, and F. nirenbergiae. These results support the view that other species may be present in grapevine seedlings as endophytic or opportunistic parasites. Some studies also suggest that Fusarium species provide various benefits to plants by enhancing plant growth, and triggering production of secondary metabolites [40]. The rhizosphere of plants contains diverse microbial communities, such as actinomycetes, bacteria, fungi, and protozoa, which interact with plant roots and each other. Climatic conditions, soil texture, chemistry, and the plant species or cultivars may closely influence the formation of these communities in the rhizosphere [41]. These factors may affect the resistance of plants to pathogens and may also play a role in the transition of Fusarium species, which were found to be opportunistic in this study, to the pathogenic form. Pathogenic Fusarium species should be considered in grapevine nurseries, and various biological and chemical control possibilities should be investigated in the future.
Author Contributions
Davut Soner AKGÜL designed this study, archived the fungal isolates, performed statistical analyses, molecular studies and pathogenicity tests, wrote the paper. Serkan ÖNDER identified the species performing phylogenetic analyses and obtained GenBank accession numbers from NCBI. Nurdan GÜNGÖR SAVAŞ, Murat YILDIZ, İzzet BÜLBÜL and Mümine ÖZRASLANDAN surveyed the grapevine nurseries, obtained-archived the fungal isolates. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Çukurova University, Scientific Research Projects Department in project FBA-2021-13533.
Acknowledgments
The authors thank The Rectorate of Çukurova University for its financial contribution. Additionally, we thank Assoc. Prof. Adem Yağcı (Tokat Gazi Osman Paşa University), Dr. Yüksel Savaş, Metin Kesgin (Manisa Viticulture Research Institute), Kürşat Alp Arslan (Pistachio Research Institute), Mehmet Ali Kiracı (Tekirdağ Viticulture Research Institute), and Assoc. Prof. Arif Atak (Bursa Uludağ University) for their valuable contributions to this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Anonymous. Personal communication with the official staff. General Directorate of Plant Production at Turkish Ministry of Agriculture and Forestry 2023. [Google Scholar]
- Gubler, W.D.; Baumgartner, K.; Browne, G.T.; Eskalen, A.; Rooney-Latham, S. , Petit, E.; Bayramian, L.A. Root disease of grapevines. Australas. Plant Path. 2004, 33, 157–165. [Google Scholar] [CrossRef]
- Gramaje, D.; Armengol, J. Fungal trunk pathogens in the grapevine propagation process: Potential inoculum sources, detection, identification, and management strategies. Plant Dis. 2012, 95, 1040–1055. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K; Whitaker, B. K.; Laraba, I.; Proctor, R.H.; Brown, D.W.; Broders, K.; Kim, H.S.; McCormick, S.P.; Busman, M., Aoki, T.; Torres-Cruz, T.J.; Geiser, D.M. DNA sequence-based identification of Fusarium: A work in progress. Plant Dis. 2022, 106, 1597–1609. [Google Scholar] [CrossRef] [PubMed]
- Summerell, B.A. Resolving Fusarium: current status of the genus. Annu. Rev. Phytopathol. 2019, 57, 323–339. [Google Scholar] [CrossRef]
- Halleen, F.; Crous, P.W.; Petrini, O. Fungi associated with healthy grapevine cuttings in nurseries, with special reference to pathogens involved in the decline of young vines. Australas. Plant Pathol. 2003, 32, 47–52. [Google Scholar] [CrossRef]
- Highet, A.S.; Nair, N.G. Fusarium oxysporum associated with grapevine decline in the Hunter Valley, NSW, Australia. Australian Journal of Grape and Wine Research 1995, 1, 48–50. [Google Scholar] [CrossRef]
- Reveglia, P.; Cinelli, T.; Cimmino, A.; Masi, M.; Evidente, A. The main phytotoxic metabolite produced by a strain of Fusarium oxysporum inducing grapevine plant declining in Italy. Nat. Prod. Res. 2018, 32, 2398–2407. [Google Scholar] [CrossRef]
- Vilvert, E.; Costa, M.D.; Cangahuala-Inocente, G.C.; Lovato, P.E. Root proteomic analysis of grapevine rootstocks inoculated with Rhizophagus irregularis and Fusarium oxysporum f. sp. herbemontis. Rev. Bras. Cienc. Solo. 2017, 41, e0160134. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Boulé, J.; Hrycan, J.; O’Gorman, D.T. Potential role of Fusarium spp. in grapevine decline. Phytopathol. Mediterr. 2023, 62, 269–281. [Google Scholar] [CrossRef]
- Bustamente, M.I.; Todd, C.; Elfar, K.; Hamid, M.I.; Garcia, J.F.; Cantu, D.; Rolshausen, P.E.; Eskalen, A. Identification and pathogenicity of Fusarium species associated with young vine decline in California. Plant Dis. 2024, 108, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Zhang, W.; Zhang, J.; Wang, H.; Peng, J.; Wang, X.; Yan, J. Belowground microbiota analysis indicates that Fusarium spp. exacerbate grapevine trunk disease. Environmental Microbiome 2023, 18, 29. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, Y.Y.; Li, X.H.; Zhang, W.; Li, Y.H.; Wang, X.D.; Yan, J.Y. First report of Fusarium commune associated with root rot of grapevine in China. Plant Dis. 2023, 107, 1238. [Google Scholar] [CrossRef]
- Akgül, D.S.; Ahioğlu, M. Fungal pathogens associated with young grapevine decline in the Southern Turkey vineyards. BIO 42nd World Congress of Vine and Wine, Web of Conferences, 2019, 15, 01027.
- Leslie, F.J.; Summerell, A.B. ; The Fusarium Laboratory Manual. Blackwell Publishing, London, UK. 2006; pp 220.
- O’Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.; Schroers, H.J.; Chaverri, P.; … Thines, M. Fusarium: more than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Sarver, B.A.J.; Balajee, S.A.; Schroers, H.J.; … Geiser, D.M. Internet-accessible DNA sequence database for identifying Fusaria from human and animal infections. J. Clin. Microbiol. 2010, 48, 3708–3718. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Haeseler, A.V.; Lanfear, R. IQ-TREE 2: New Models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, A.V.; Jermiin, L.S. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Haeseler, A.V.; Minh, B.Q.; Vinh, L.S. ; UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Haag, P.; Bowen, P.; O’Gorman, D.T. ; Grapevine trunk diseases in British Columbia: Incidence and characterization of the fungal pathogens associated with esca and Petri diseases of grapevine. Plant Dis. 2014, 98, 456–468. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical procedures for agricultural research (2nd ed.). Wiley, UK. 1984, pp 680.
- Granett, J.; Omer, A.D.; Pessereau, P.; Walker, M.A. Fungal infections of grapevine roots in phylloxera-infested vineyards. Vitis 1998, 37, 39–42. [Google Scholar] [CrossRef]
- Pintos, C.; Redondo, V.; Costas, D.; Aguin, O.; Mansilla, P. Fungi associated with grapevine trunk diseases in nursery-produced Vitis vinifera plants. Phytopathol. Mediterr. 2018, 57, 407–424. [Google Scholar] [CrossRef]
- Lombard, L.; Sandoval-Denis, M.; Lamprecht, S.C.; Crous, P.W. Epitypification of Fusarium oxysporum clearing the taxonomic chaos. Persoonia 2019, 43, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.F.; Hwang, S.F.; Conner, R.L.; Gossen, B.D. First report of Fusarium proliferatum causing root rot in soybean (Glycine max L.) in Canada. Crop Prot. 2015, 67, 52–58. [Google Scholar] [CrossRef]
- Akbar, A.; Hussain, S.; Ullah, K.; Fahim, M.; Ali, G.S. Detection, virulence and genetic diversity of Fusarium species infecting tomato in Northern Pakistan. PloS One. [CrossRef]
- Sanna, M.; Martino, I.; Guarnaccia, V.; Mezzalama, M. Diversity and pathogenicity of Fusarium species associated with stalk and crown rot in maize in Northern Italy. Plants, 2023, 12, 3857. [Google Scholar] [CrossRef] [PubMed]
- Rajput, N.; Zaman, B.; Huo, C.; Cao, J.; Atiq, M.; Lodhi, A.M.; Syed, R.N.; Khan, B.; Iqbal, O.; Zhao, Z. First report of Fusarium equiseti causing stem rot disease of grape (Vitis vinifera L.) in Afghanistan. J. Plant Pathol. 2020, 102, 1277. [Google Scholar] [CrossRef]
- Parra, M.Á.; Gómez, J.; Aguilar, F.W.; Martínez, J.A. Fusarium annulatum causes Fusarium rot of cantaloupe melons in Spain. Phytopathol. Mediterr. 2022, 2022. 61, 269–277. [Google Scholar] [CrossRef]
- Mirghasempour, S.A.; Studholme, D.J.; Chen, W.; Zhu, W.; Mao, B. Molecular and pathogenic characterization of Fusarium species associated with corm rot disease in saffron from China. J. Fungi 2022, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Vetrova, S.; Alyokhina, K.; Engalycheva, I.; Kozar, E.; Mukhina, K.; Sletova, M.; Krivenkov, L.; Tikhonova, T.; Kameneva, A.; Frolova, S.; et al. Identification and pathogenicity of Fusarium species associated with onion basal rot in the Moscow Region of Russian Federation. J. Fungi 2024, 10, 331. [Google Scholar] [CrossRef]
- Zhang, H.; Sha, H.D.; Chen, W.L.; Mao, B.Z. First Report of Fusarium annulatum Causing Blight on Bletilla striata (Baiji) in China. Plant Dis. 2024, 108, 800. [Google Scholar] [CrossRef]
- Aiello, D.; Fiorenza, A.; Leonardi, G.R.; Vitale, A.; Polizzi, G. Fusarium nirenbergiae (Fusarium oxysporum Species Complex) causing the wilting of passion fruit in Italy. Plants 2021, 10, 2011. [Google Scholar] [CrossRef]
- Zhao, X.; Li, H.; Zhou, L.; Chen, F.; Chen, F. Wilt of Acer negundo L. caused by Fusarium nirenbergiae in China. J. For. Res. 2020, 31, 2013–2022. [Google Scholar] [CrossRef]
- López-Moral, A.; Antón-Domínguez, B.I.; Lovera, M.; Arquero, O.; Trapero, A.; Agusti-Brisach, C. Identification and pathogenicity of Fusarium species associated with wilting and crown rot in almond (Prunus dulcis). Sci. Rep. 2024, 14, 5720. [Google Scholar] [CrossRef] [PubMed]
- Mirghasempour, S.A.; Michailides, T.; Chen, W.; Mao, B. Fusarium spp. associated with Dendrobium officinale dieback disease in China. J. Fungi 2022, 8, 919. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, W.; Li, X.; Ji, S.; Chethana, K.W.T.; Hyde, K.D.; Yan, J. Fusarium species associated with cherry leaf spot in China. Plants 2022, 11, 2760. [Google Scholar] [CrossRef] [PubMed]
- Murali, M.; Naziya, B.; Ansari, M.A.; Alomary, M.N.; AlYahya, S.; Almatroudi, A.; Thriveni, M.C.; Gowtham, H.G.; Singh, S.B.; Aiyaz, M. Bioprospecting of rhizosphere-resident fungi: Their role and importance in sustainable agriculture. J. Fungi 2021, 7, 314. [Google Scholar] [CrossRef]
- Chauhan, P.; Sharma, N.; Tapwal, A.; Kumar, A.; Verma, G.S.; Meena, M.; Seth, C.S.; Swapnil, P. Soil microbiome: Diversity, benefits and interactions with plants. Sustainability 2023, 15, 14643. [Google Scholar] [CrossRef]
Figure 1.
Multi gene (TEF1-α and RPB2) Maximum Likelihood tree of Fusarium isolates. Circles of different sizes show bootstrap support values from 1000 replicates are indicated at the nodes. Bootstrap values less than 50% are not shown. The bold characters represent the Turkish isolates. Dactylonectria torresensis CBS 129086 was used for rooting the ML tree.
Figure 1.
Multi gene (TEF1-α and RPB2) Maximum Likelihood tree of Fusarium isolates. Circles of different sizes show bootstrap support values from 1000 replicates are indicated at the nodes. Bootstrap values less than 50% are not shown. The bold characters represent the Turkish isolates. Dactylonectria torresensis CBS 129086 was used for rooting the ML tree.
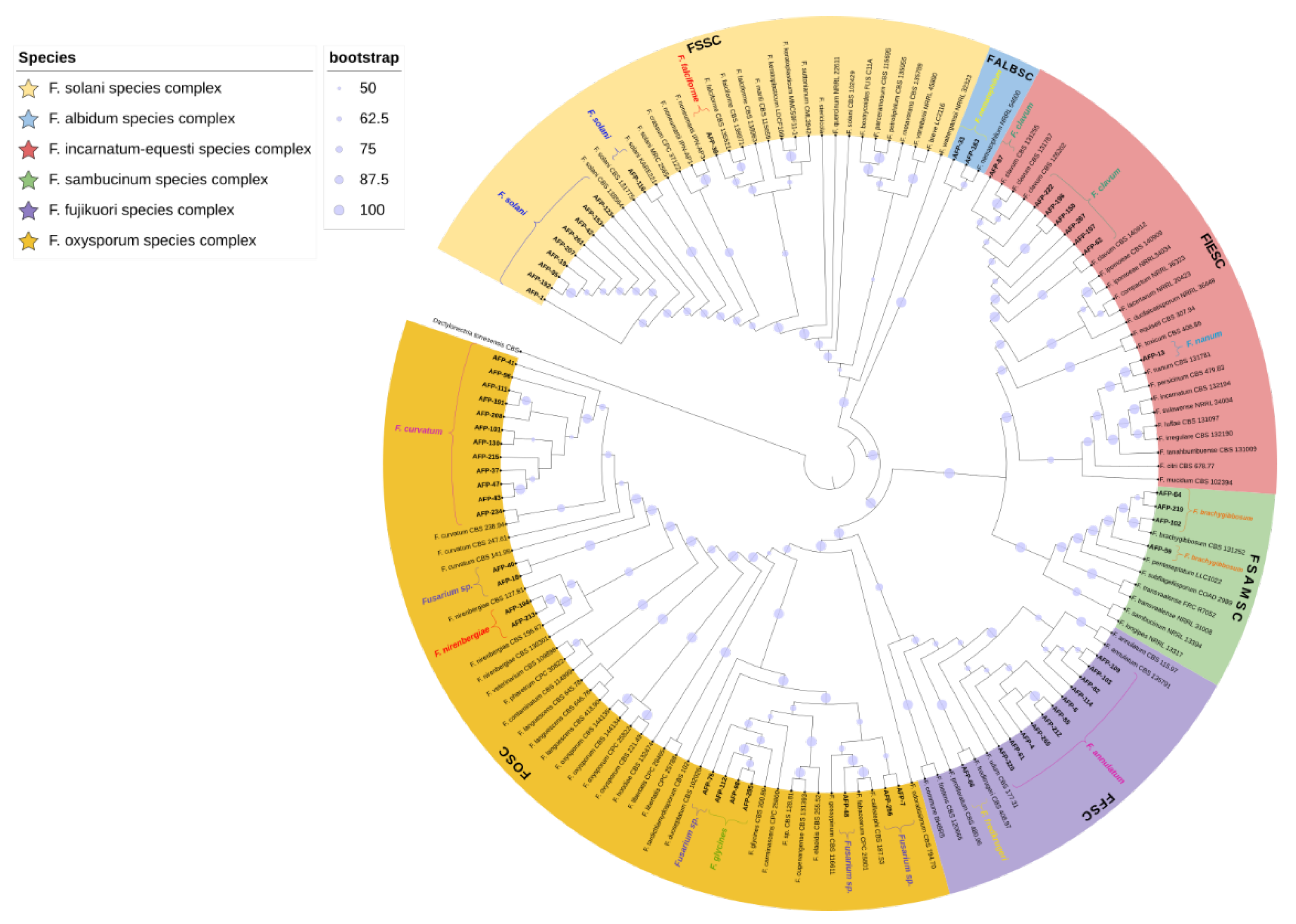
Figure 2.
Wood necrosis in the stems of grapevine seedlings induced by Fusarium and Ilynonectria liriodendri after four months of inoculation. a) I. liriodendri, b) F. annulatum, c) F. brachygibbosum, d) F. nirenbergiae, e) F. curvatum, f) F. solani, g) F. glycines, h) F. falciforme, i) Non-inoculated control.
Figure 2.
Wood necrosis in the stems of grapevine seedlings induced by Fusarium and Ilynonectria liriodendri after four months of inoculation. a) I. liriodendri, b) F. annulatum, c) F. brachygibbosum, d) F. nirenbergiae, e) F. curvatum, f) F. solani, g) F. glycines, h) F. falciforme, i) Non-inoculated control.

Figure 3.
Basal wood necrosis in grapevine seedlings induced by Fusarium and Ilynonectria liriodendri after four months of inoculation. a) I. liriodendri, b) F. annulatum, c) F. nirenbergiae, d) F. curvatum, e) F. glycines, f) F. solani, g) F. fredkrugeri, h) F. falciforme, i) Non-inoculated control.
Figure 3.
Basal wood necrosis in grapevine seedlings induced by Fusarium and Ilynonectria liriodendri after four months of inoculation. a) I. liriodendri, b) F. annulatum, c) F. nirenbergiae, d) F. curvatum, e) F. glycines, f) F. solani, g) F. fredkrugeri, h) F. falciforme, i) Non-inoculated control.

Figure 4.
The average lesion lengths in the wood tissues of grapevine seedlings (1103 Paulsen rootstock) induced by Fusarium species and I. liriodendri.
Figure 4.
The average lesion lengths in the wood tissues of grapevine seedlings (1103 Paulsen rootstock) induced by Fusarium species and I. liriodendri.

Figure 5.
The average lesion lengths in the basal wood tissues of grapevine seedlings (1103 Paulsen rootstock) induced by Fusarium species and I. liriodendri.
Figure 5.
The average lesion lengths in the basal wood tissues of grapevine seedlings (1103 Paulsen rootstock) induced by Fusarium species and I. liriodendri.
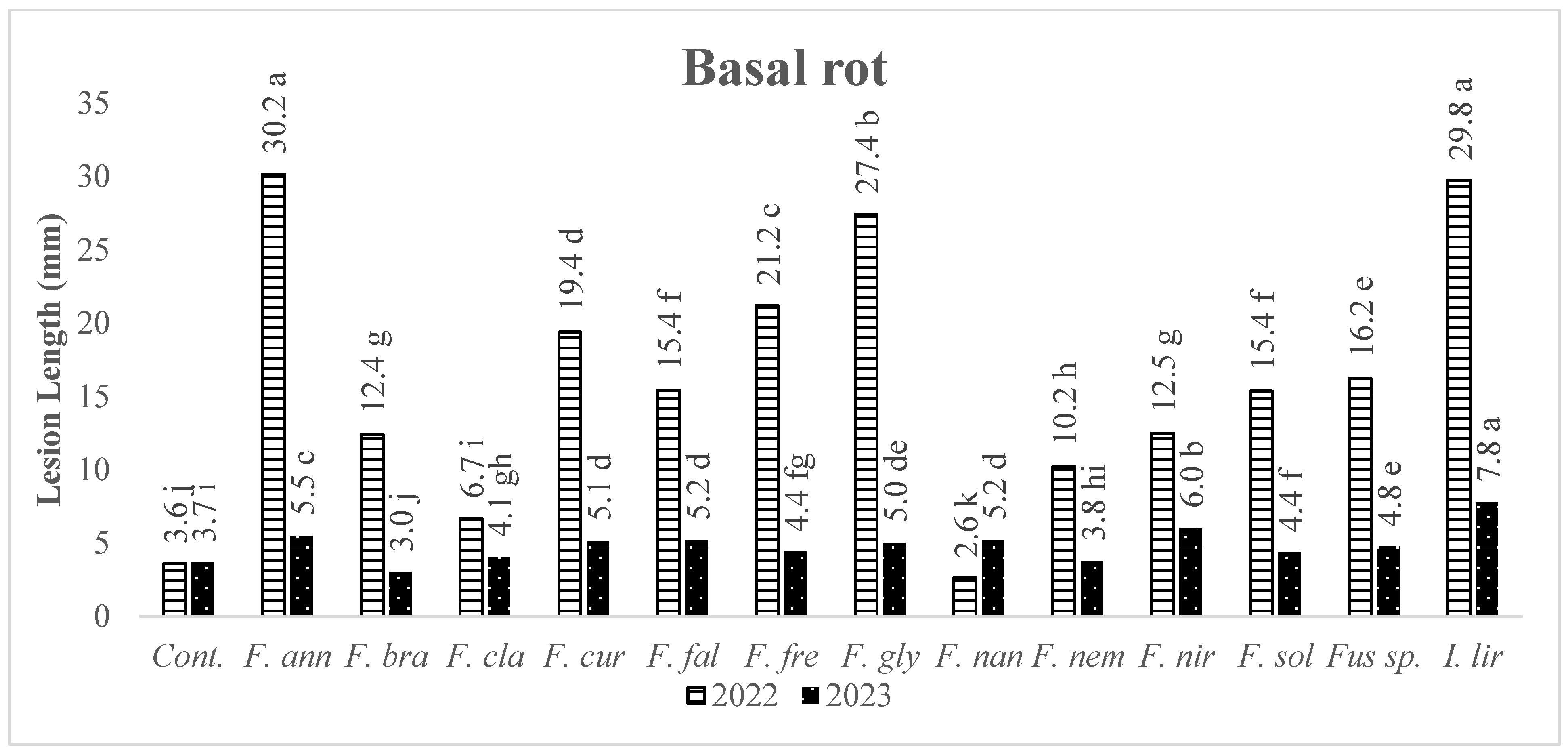
Figure 6.
The average dry root weight of grapevine seedlings (1103 Paulsen rootstock) induced by Fusarium species and I. liriodendri.
Figure 6.
The average dry root weight of grapevine seedlings (1103 Paulsen rootstock) induced by Fusarium species and I. liriodendri.
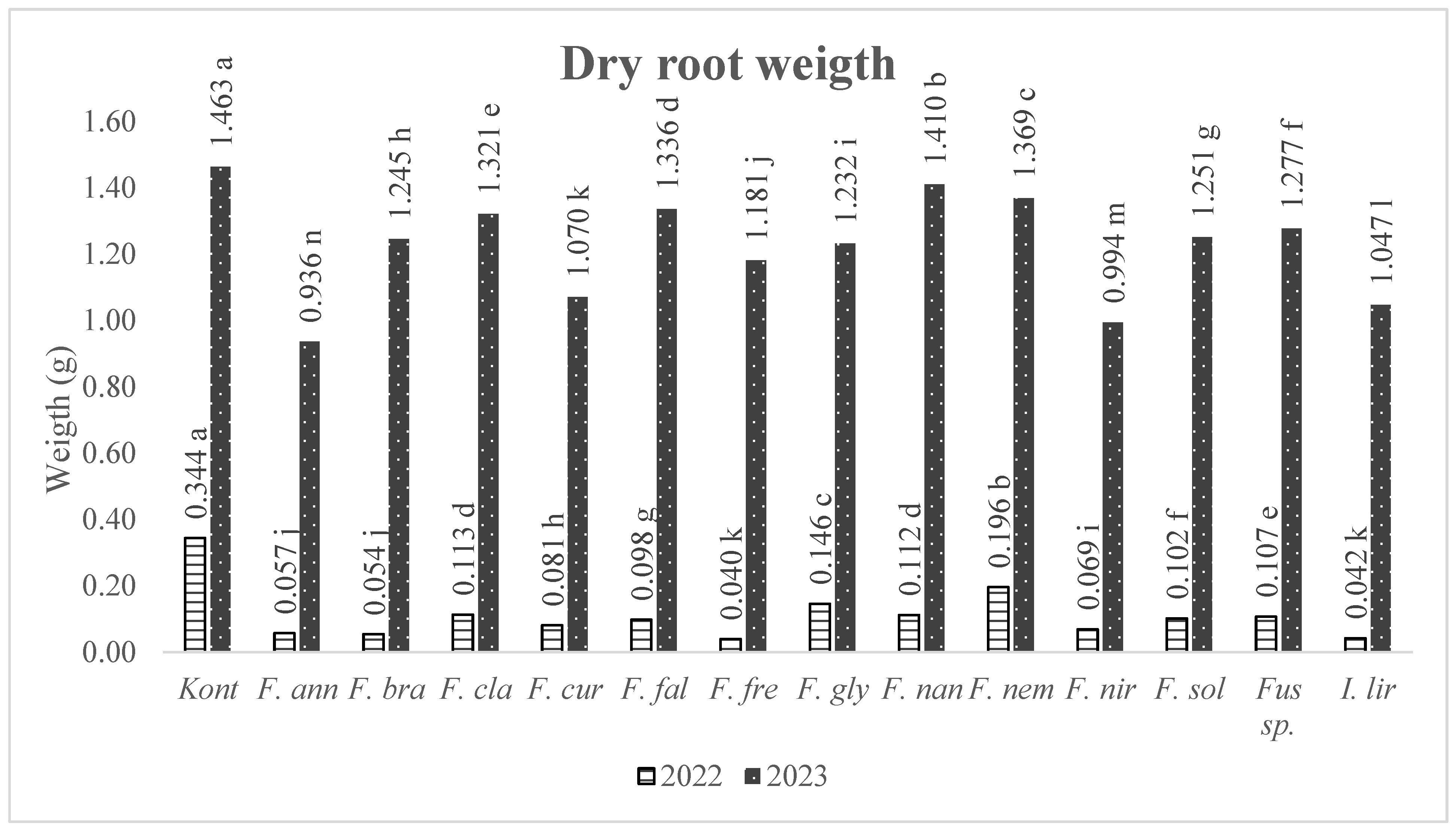

Table 1.
GenBank accession numbers of partial sequence of TEF1-α and RPB2 of references species used in the phylogenetic analyses.
Table 1.
GenBank accession numbers of partial sequence of TEF1-α and RPB2 of references species used in the phylogenetic analyses.
| Reference Species | Isolate | GenBank Accession Numbers | |
| TEF1-α | RPB2 | ||
| F. solani | CBS 138564 | KT272100 | KT272102 |
| " | CBS 131775 | JX118990 | JX237778 |
| " | KARE_221 | MK077042 | MK077080 |
| " | MRC_2565 | MH582420 | MH582410 |
| " | CBS 102429 | KM231936 | KM232376 |
| F. crassum | CPC_37122 | MW248760 | MW446594 |
| F. noneumartii | IPN-AP1 | OP902594 | OP902591 |
| " | IPN-AP3 | OP902596 | OP902593 |
| F. falciforme | CBS 135521 | KU711733 | KU604357 |
| " | CBS 138971 | KT716212 | KT716187 |
| " | CBS 138963 | KT716213 | KT716188 |
| F. martii | CBS 115659 | JX435156 | JX435256 |
| F. keratoplasticum | LDCF109 | OP184958 | OP186372 |
| " | MMC59F11-1 | MF069182 | MF069181 |
| F. suttonianum | CML3942 | MK158921 | MH709236 |
| F. stericicola | N/A | LR583659 | LR583888 |
| F. quercinum | NRRL:22611 | DQ246841 | EU329518 |
| F. bostrycoides | FUS C11A | PP105767 | PP125181 |
| F. parceramosum | CBS 115695 | JX435149 | JX435249 |
| F. petroliphilum | CBS 135955 | KU711768 | KU604337 |
| F. metavorans | CBS 135789 | KU711773 | KU604374 |
| F. vanettenii | NRRL 45880 | FJ240352 | JX171655 |
| F. breve | LC2116 | MW620163 | MW474688 |
| F. waltergamsii | NRRL 32323 | DQ246951 | EU329576 |
| F. nematophilum | NRRL_54600 | N/A | JX171664 |
| F. clavum | CBS 131255 | MN170460 | MN170393 |
| " | CBS 131787 | MN170461 | MN170394 |
| " | CBS 126202 | MN170456 | MN170389 |
| " | CBS 140912 | MN170462 | MN170395 |
| F. ipomoeae | CBS 140909 | MN170479 | MN170412 |
| " | NRRL 34034 | GQ505636 | GQ505814 |
| F. compactum | NRRL 36323 | GQ505648 | GQ505826 |
| F. lacertarum | NRRL 20423 | GQ505593 | GQ505771 |
| F. duofalcatisporum | NRRL 36448 | GQ505652 | GQ505830 |
| F. equiseti | CBS 307.94 | KR071777 | KU604327 |
| F. toxicum | CBS 406.86 | MN170508 | MN170441 |
| F. nanum | CBS 131781 | MN170487 | MN170420 |
| F. persicinum | CBS 479.83 | MN170495 | MN170428 |
| F. incarnatum | CBS 132194 | KF255470 | KF255542 |
| F. sulawense | NRRL 34004 | GQ505628 | GQ505806 |
| F. luffae | CBS 131097 | MN170482 | MN170415 |
| F. irregulare | CBS 132190 | MN170480 | MN170413 |
| F. tanahbumbuense | CBS 131009 | MN170506 | MN170439 |
| F. citri | CBS 678.77 | MN170453 | MN170386 |
| F. mucidum | CBS 102394 | MN170484 | MN170417 |
| F. brachygibbosum | CBS 131252 | JQ429334 | JX162526 |
| F. pentaseptatum | LLC1022 | OP487255 | OP486819 |
| F. subflagellisporum | COAD 2989 | MT774486 | MZ970426 |
| F. transvaalense | FRC R7052 | MW233161 | MW233505 |
| " | NRRL 31008 | MW233102 | MW233446 |
| F. sambucinum | NRRL 13394 | MW233064 | MW233407 |
| F. longipes | NRRL 13317 | MW233058 | MG282411 |
| F. annulatum | CBS 115.97 | MW401973 | MW402785 |
| " | CBS 135791 | MW402054 | MW402746 |
| F. udum | CBS 177.31 | MH484957 | MH484866 |
| F. fredkrugeri | CBS 408.97 | MW402126 | MW402814 |
| F. proliferatum | CBS 480.96 | MN534059 | MN534272 |
| F. foetens | CBS 120665 | MH485009 | MH484918 |
| F. commune | BHBR5 | OR900978 | OR888540 |
| F. odoratissimum | CBS 794.70 | MH484969 | MH484878 |
| F. callistephi | CBS 187.53 | MH484966 | MH484875 |
| F. fabacearum | CPC 25801 | MH485029 | MH484938 |
| F. gossypinum | CBS 116611 | MH484998 | MH484907 |
| F. elaeidis | CBS 255.52 | MH484965 | MH484874 |
| F. cugenangense | CBS 131393 | MH485019 | MH484928 |
| Fusarium sp. | CBS 128.81 | MH484975 | MH484884 |
| F. carminascens | CPC 25800 | MH485028 | MH484937 |
| F. glycines | CBS 200.89 | MH484979 | MH484888 |
| F. duoseptatum | CBS 102026 | MH484987 | MH484896 |
| F. tardichlamydosporum | CBS 102028 | MH484988 | MH484897 |
| F. libertatis | CPC 25788 | MH485024 | MH484933 |
| " | CPC 28465 | MH485035 | MH484944 |
| F. hoodiae | CBS 132474 | MH485020 | MH484929 |
| F. oxysporum | CBS 221.49 | MH484963 | MH484872 |
| " | CPC 25822 | MH485034 | MH484943 |
| " | CBS 144134 | MH485044 | MH484953 |
| " | CBS 144135 | MH485045 | MH484954 |
| F. languescens | CBS 413.90 | MH484981 | MH484890 |
| " | CBS 646.78 | MH484972 | MH484881 |
| " | CBS 645.78 | MH484971 | MH484880 |
| F. contaminatum | CBS 114899 | MH484992 | MH484901 |
| F. pharetrum | CPC 30822 | MH485042 | MH484951 |
| F. veterinarium | CBS 109898 | MH484990 | MH484899 |
| F. nirenbergiae | CBS 130301 | MH485017 | MH484926 |
| " | CBS 196.87 | MH484977 | MH484886 |
| " | CBS 127.81 | MH484974 | MH484883 |
| F. curvatum | CBS 141.95 | MH484985 | MH484894 |
| " | CBS 247.61 | MH484967 | MH484876 |
| " | CBS 238.94 | MH484984 | MH484893 |
| Dactylonectria torresensis | CBS 129086 | JF735870 | KM232347 |
Table 2.
Location of surveyed grapevine nurseries, rootstock/cultivars, isolation frequency of Fusarium species in Türkiye.
Table 2.
Location of surveyed grapevine nurseries, rootstock/cultivars, isolation frequency of Fusarium species in Türkiye.
| Nursery | Location | Rootstock or Cultivar | Isolation | ||
| Frequency (%) | |||||
| 1 | Bursa | 1103P-Trakya İlkeren | 32.9 | ||
| 2 | Mersin | 1103P- Victoria | 32.9 | ||
| 3 | Salihli, Manisa | Thompson Seedless | - | ||
| 4 | Salihli, Manisa | Sultana Seedless | 28.5 | ||
| 5 | Salihli, Manisa | Sultana Seedless | - | ||
| 6 | Salihli, Manisa | Sultana Seedless | 38.5 | ||
| 7 | Salihli, Manisa | 1103P / Sultana Seedless | 40.0 | ||
| 8 | Alaşehir, Manisa | Sultana Seedless | 34.3 | ||
| 9 | Alaşehir, Manisa | Sultana Seedless | 4.3 | ||
| 10 | Alaşehir, Manisa | Sultana Seedless | 24.3 | ||
| 11 | Sarıgöl, Manisa | Sultana Seedless | 58.6 | ||
| 12 | Salihli, Manisa | Sultana Seedless | 17.1 | ||
| 13 | Tekirdağ | Kober 5BB / Sultan 1 | 8.6 | ||
| 14 | Tekirdağ | Kober 5BB / Bozbey | 17.2 | ||
| 15 | Tekirdağ | 1103P-Tekirdağ Çekirdeksizi | 27.1 | ||
| 16 | Tekirdağ | 110R-Yapıncak | 2.9 | ||
| 17 | Denizli | 41B / Sultana Seedless | 20.0 | ||
| 18 | Denizli | 41B / Sultana Seedless | 40.0 | ||
| 19 | Denizli | 41B / Sultana Seedless | 65.7 | ||
| 20 | Denizli | 41B / Sultana Seedless | 50.0 | ||
| 21 | Denizli | 41B / Michele Palieri | 45.7 | ||
| 22 | Şanlıurfa | 1103P - Ergin Çekirdeksizi | 12.9 | ||
| 23 | Şanlıurfa | 110R - Horozkarası | 11.5 | ||
| 24 | Şanlıurfa | 99R - Çiloreş | 12.9 | ||
| 25 | Şanlıurfa | 1103P - Victoria | 5.7 | ||
| 26 | Manisa | 41B / Red Globe | 47.1 | ||
| 27 | Manisa | Kober 5BB / Royal | 20.0 | ||
| 28 | Manisa | 1103P - Sultana Seedless | 20.0 | ||
| 29 | Manisa | Kober 5BB - Sultana Seedless | 21.4 | ||
| 30 | Manisa | 1103P - Crimson Seedless | 17.1 | ||
| 31 | Manisa | 110R / Alicante Bouschet | 26.3 | ||
| 32 | Alaşehir, Manisa | 1103P - Thompson Seedless | 60.0 | ||
| 33 | Manisa | Kober 5BB / Ata Sarısı | 22.9 | ||
| 34 | Turgutlu, Manisa | Kober 5BB /Sultana Seedless | 28.6 | ||
| 35 | Manisa | Kober 5BB / Trakya İlkeren | 8.6 | ||
| 36 | Tokat | 1103P - Narince | 40.0 | ||
| 37 | Tokat | 1103P/Narince | 21.4 | ||
| 38 | Tokat | 1103P/Narince | 11.4 | ||
| 39 | Tokat | 1103P/Sultan7 | 12.9 | ||
| 40 | Tokat | 1103P/Narince | 20.0 | ||
| 41 | Tokat | Du Lot / Narince | 12.9 | ||
| 42 | Adıyaman | Kober 5BB / Hatun Parmağı | 17.9 | ||
| 43 | Mersin | 1103P / Victoria | 31.3 | ||
| Mean | 24.9 | ||||
Table 3.
Location of surveyed grapevine nurseries, cultivars, and the species found with their TEF1-α and RPB2 gene sequence numbers.
Table 3.
Location of surveyed grapevine nurseries, cultivars, and the species found with their TEF1-α and RPB2 gene sequence numbers.
| Isolate | Fusarium Species | Location | Rootstock / Cultivar | GenBank Accession Numbers | |
| TEF1-α | RPB2 | ||||
| AFP004 | Fusarium annulatum | Bursa | 1103 Paulsen | PP449277 | PP449217 |
| AFP006 | " | Bursa | 1103 Paulsen | PP449278 | PP449218 |
| AFP061 | " | Manisa | Kober 5BB | PP449279 | PP449219 |
| AFP082 | " | Tokat | 1103 Paulsen | PP449280 | PP449220 |
| AFP085 | " | Tokat | 1103 Paulsen | PP449281 | PP449221 |
| AFP103 | " | Manisa | Kober 5BB | PP449282 | PP449222 |
| AFP109 | " | Manisa | 110 Richter | PP449283 | PP449223 |
| AFP114 | " | Manisa | Kober 5BB | PP449284 | PP449224 |
| AFP212 | " | Tokat | 1103 Paulsen | PP449285 | PP449225 |
| AFP265 | " | Manisa | Sultana Seedless | PP449286 | PP449226 |
| AFP320 | " | Tekirdağ | 1103 Paulsen | PP449287 | PP449227 |
| AFP059 | Fusarium brachygibbosum | Manisa | Kober 5BB | PP449288 | PP449228 |
| AFP064 | " | Manisa | 41B | PP449289 | PP449229 |
| AFP102 | " | Manisa | Ramsey | PP449290 | PP449230 |
| AFP219 | " | Şanlıurfa | 1103 Paulsen | PP449291 | PP449231 |
| AFP062 | Fusarium clavum | Manisa | Kober 5BB | PP449292 | PP449232 |
| AFP087 | " | Tokat | 1103 Paulsen | PP449293 | PP449233 |
| AFP107 | " | Manisa | Ramsey | PP449294 | PP449234 |
| AFP150 | " | Tokat | 1103 Paulsen | PP449295 | PP449235 |
| AFP196 | " | Manisa | Ramsey | PP449296 | PP449236 |
| AFP222 | " | Tokat | 1103 Paulsen | PP449297 | PP449237 |
| AFP267 | " | Manisa | Sultana Seedless | PP449298 | PP449238 |
| AFP037 | Fusarium curvatum | Denizli | 140 Ruggeri | PP449299 | PP449239 |
| AFP041 | " | Denizli | 1103 Paulsen | PP449300 | PP449240 |
| AFP043 | " | Denizli | 140 Ruggeri | PP449301 | PP449241 |
| AFP047 | " | Denizli | 140 Ruggeri | PP449302 | PP449242 |
| AFP096 | " | Manisa | Kober 5BB | PP449303 | PP449243 |
| AFP101 | " | Manisa | Ramsey | PP449304 | PP449244 |
| AFP111 | " | Manisa | 110 Richter | PP449305 | PP449245 |
| AFP130 | " | Mersin | 1103 Paulsen | PP449306 | PP449246 |
| AFP191 | " | Tokat | 1103 Paulsen | PP449307 | PP449247 |
| AFP208 | " | Tokat | 1103 Paulsen | PP449308 | PP449248 |
| AFP215 | " | Şanlıurfa | 1103 Paulsen | PP449309 | PP449249 |
| AFP234 | " | Şanlıurfa | 1104 Paulsen | PP449310 | PP449250 |
| AFP038 | Fusarium falciforme | Denizli | 140 Ruggeri | PP449311 | PP449251 |
| AFP066 | Fusarium fredkrugeri | Manisa | Kober 5BB | PP449312 | PP449252 |
| AFP098 | Fusarium glycines | Manisa | Ramsey | PP449313 | PP449253 |
| AFP112 | " | Manisa | 110 Richter | PP449314 | PP449254 |
| AFP295 | " | Manisa | Sultana Seedless | PP449315 | PP449255 |
| AFP013 | Fusarium nanum | Mersin | 140 Ruggeri | PP449316 | PP449256 |
| AFP033 | Fusarium nematophilum | Manisa | Kober 5BB | PP449317 | PP449257 |
| AFP163 | " | Tokat | 1103 Paulsen | PP449318 | PP449258 |
| AFP194 | Fusarium nirenbergiae | Manisa | Ramsey | PP449319 | PP449259 |
| AFP213 | " | Tokat | 1103 Paulsen | PP449320 | PP449260 |
| AFP001 | Fusarium solani | Bursa | 1103 Paulsen | PP449321 | PP449261 |
| AFP019 | " | Manisa | 1103 Paulsen | PP449322 | PP449262 |
| AFP042 | " | Denizli | 140 Ruggeri | PP449323 | PP449263 |
| AFP095 | " | Manisa | Kober 5BB | PP449324 | PP449264 |
| AFP116 | " | Manisa | Kober 5BB | PP449325 | PP449265 |
| AFP123 | " | Mersin | 1103 Paulsen | PP449326 | PP449266 |
| AFP153 | " | Tokat | 1103 Paulsen | PP449327 | PP449267 |
| AFP192 | " | Manisa | Ramsey | PP449328 | PP449268 |
| AFP207 | " | Mersin | 1103 Paulsen | PP449329 | PP449269 |
| AFP261 | " | Manisa | Sultana Seedless | PP449330 | PP449270 |
| AFP007 | Fusarium sp. | Bursa | 1103 Paulsen | PP449331 | PP449271 |
| AFP018 | " | Manisa | 1103 Paulsen | PP449332 | PP449272 |
| AFP040 | " | Denizli | 140 Ruggeri | PP449333 | PP449273 |
| AFP048 | " | Denizli | 1103 Paulsen | PP449334 | PP449274 |
| AFP075 | " | Denizli | 140 Ruggeri | PP449335 | PP449275 |
| AFP256 | " | Manisa | Sultana Seedless | PP449336 | PP449276 |
Table 4.
Mean wood lesion lengths caused by Fusarium species in the inoculation points of 1103 Paulsen rootstock plants after four months.
Table 4.
Mean wood lesion lengths caused by Fusarium species in the inoculation points of 1103 Paulsen rootstock plants after four months.
| Isolates | 2022 | Lesion | Isolates | 2023 | Lesion | ||||
| Species | (mm) | Species | (mm) | ||||||
| AFP006 | F. annulatum | 12.0 | a* | AFP061 | F. annulatum | 8.9 | a* | ||
| AFP114 | F. annulatum | 11.9 | a | AFP115 | Ilyonectria liriodendri | 8.6 | a | ||
| AFP109 | F. annulatum | 11.0 | b | AFP213 | F. nirenbergiae | 8.4 | a | ||
| AFP103 | F. annulatum | 10.9 | bc | AFP103 | F. annulatum | 8.0 | b | ||
| AFP059 | F. brachygibbosum | 10.4 | bc | AFP265 | F. annulatum | 7.7 | bc | ||
| AFP115 | Ilyonectria liriodendri | 10.3 | c | AFP114 | F. annulatum | 7.5 | c | ||
| AFP265 | F. annulatum | 7.9 | d | AFP111 | F. curvatum | 7.5 | c | ||
| AFP194 | F. nirenbergiae | 7.1 | e | AFP194 | F. nirenbergiae | 7.5 | c | ||
| AFP061 | F. annulatum | 6.9 | e | AFP006 | F. annulatum | 6.3 | d | ||
| AFP004 | F. annulatum | 6.9 | e | AFP004 | F. annulatum | 5.9 | de | ||
| AFP096 | F. curvatum | 5.3 | f | AFP096 | F. curvatum | 5.2 | e | ||
| AFP213 | F. nirenbergiae | 5.3 | f | AFP109 | F. annulatum | 5.1 | e | ||
| AFP256 | Fusarium sp. | 5.3 | f | AFP059 | F. brachygibbosum | 5.1 | e | ||
| AFP043 | F. curvatum | 5.2 | f | AFP043 | F. curvatum | 5.1 | e | ||
| AFP101 | F. curvatum | 5.2 | f | AFP101 | F. curvatum | 5.0 | e | ||
| AFP038 | F. falciforme | 5.2 | f | AFP098 | F. glycines | 5.0 | e | ||
| AFP040 | Fusarium sp. | 5.2 | f | AFP019 | F. solani | 5.0 | e | ||
| AFP013 | F. nanum | 5.2 | f | AFP041 | F. curvatum | 5.0 | e | ||
| AFP130 | F. curvatum | 5.1 | f | AFP130 | F. curvatum | 5.0 | e | ||
| AFP019 | F. solani | 5.1 | f | AFP037 | F. curvatum | 5.0 | e | ||
| AFP037 | F. curvatum | 5.1 | f | AFP040 | Fusarium sp. | 5.0 | e | ||
| AFP075 | Fusarium sp. | 5.1 | f | AFP066 | F. fredkrugeri | 5.0 | e | ||
| AFP095 | F. solani | 5.1 | f | AFP123 | F. solani | 5.0 | e | ||
| AFP111 | F. curvatum | 5.1 | f | AFP256 | Fusarium sp. | 5.0 | e | ||
| AFP191 | F. curvatum | 5.1 | f | AFP191 | F. curvatum | 5.0 | e | ||
| AFP041 | F. curvatum | 5.1 | f | AFP038 | F. falciforme | 5.0 | e | ||
| AFP001 | F. solani | 5.1 | f | AFP001 | F. solani | 5.0 | e | ||
| AFP007 | Fusarium sp. | 5.1 | f | AFP018 | Fusarium sp. | 5.0 | e | ||
| AFP222 | F. clavum | 5.1 | f | AFP222 | F. clavum | 5.0 | e | ||
| AFP261 | F. solani | 5.1 | f | AFP261 | F. solani | 5.0 | e | ||
| AFP066 | F. fredkrugeri | 5.1 | f | AFP075 | Fusarium sp. | 5.0 | e | ||
| AFP033 | F. nematophilum | 5.0 | f | AFP033 | F. nematophilum | 5.0 | e | ||
| AFP048 | Fusarium sp. | 5.0 | f | AFP048 | Fusarium sp. | 5.0 | e | ||
| AFP062 | F. clavum | 5.0 | f | AFP062 | F. clavum | 5.0 | e | ||
| AFP123 | F. solani | 5.0 | f | AFP095 | F. solani | 5.0 | e | ||
| AFP196 | F. clavum | 5.0 | f | AFP196 | F. clavum | 5.0 | e | ||
| AFP098 | F. glycines | 5.0 | f | AFP013 | F. nanum | 5.0 | e | ||
| AFP018 | Fusarium sp. | 5.0 | f | AFP007 | Fusarium sp. | 5.0 | e | ||
| Non-inoculated Control | 5.0 | f | Non-inoculated Control | 5.0 | e | ||||
| LSD = 0.65 | LSD = 0.92 | ||||||||
*Means accompanied by same letter are not significantly different (P = 0.05) according to LSD tests.
Table 5.
Mean basal necrose lengths caused by Fusarium species in 1103 Paulsen rootstock plants after four months.
Table 5.
Mean basal necrose lengths caused by Fusarium species in 1103 Paulsen rootstock plants after four months.
| Isolates | 2022 | Basal | Isolates | 2023 | Basal | |||
| Fungal Species | Necrosis (mm) | Fungal Species | Necrosis (mm) | |||||
| AFP061 | F. annulatum | 37.0 | a | AFP115 | Ilyonectria liriodendri | 7.8 | a | |
| AFP103 | F. annulatum | 34.6 | ab | AFP004 | F. annulatum | 6.2 | ab | |
| AFP114 | F. annulatum | 30.8 | a-c | AFP101 | F. curvatum | 6.2 | ab | |
| AFP041 | F. curvatum | 29.8 | a-d | AFP194 | F. nirenbergiae | 6.2 | ab | |
| AFP115 | Ilyonectria liriodendri | 29.8 | a-d | AFP103 | F. annulatum | 6.0 | a-c | |
| AFP004 | F. annulatum | 28.4 | a-e | AFP111 | F. curvatum | 6.0 | a-c | |
| AFP109 | F. annulatum | 27.6 | b-f | AFP114 | F. annulatum | 5.8 | b-d | |
| AFP098 | F. glycines | 27.4 | b-f | AFP213 | F. nirenbergiae | 5.8 | b-d | |
| AFP006 | F. annulatum | 26.8 | b-g | AFP037 | F. curvatum | 5.6 | b-e | |
| AFP265 | F. annulatum | 26.4 | b-g | AFP109 | F. annulatum | 5.6 | b-e | |
| AFP019 | F. solani | 25.2 | c-h | AFP256 | Fusarium sp. | 5.4 | b-f | |
| AFP111 | F. curvatum | 23.8 | c-i | AFP006 | F. annulatum | 5.2 | b-f | |
| AFP018 | Fusarium sp. | 23.2 | c-i | AFP013 | F. nanum | 5.2 | b-f | |
| AFP256 | Fusarium sp. | 22.8 | c-i | AFP038 | F. falciforme | 5.2 | b-f | |
| AFP037 | F. curvatum | 21.2 | d-j | AFP048 | Fusarium sp. | 5.2 | b-f | |
| AFP066 | F. fredkrugeri | 21.2 | d-j | AFP061 | F. annulatum | 5.2 | b-f | |
| AFP075 | Fusarium sp. | 19.8 | d-j | AFP191 | F. curvatum | 5.2 | b-f | |
| AFP096 | F. curvatum | 19.0 | f-k | AFP040 | Fusarium sp. | 5.0 | b-f | |
| AFP123 | F. solani | 17.8 | g-l | AFP098 | F. glycines | 5.0 | b-f | |
| AFP101 | F. curvatum | 17.0 | h-m | AFP265 | F. annulatum | 5.0 | b-f | |
| AFP043 | F. curvatum | 16.0 | i-m | AFP075 | Fusarium sp. | 4.8 | b-g | |
| AFP007 | Fusarium sp. | 15.4 | i-n | AFP123 | F. solani | 4.8 | b-g | |
| AFP038 | F. falciforme | 15.4 | i-n | AFP001 | F. solani | 4.6 | b-g | |
| AFP130 | F. curvatum | 15.0 | i-n | AFP019 | F. solani | 4.6 | b-g | |
| AFP261 | F. solani | 15.0 | i-n | AFP041 | F. curvatum | 4.6 | b-g | |
| AFP191 | F. curvatum | 13.6 | j-o | AFP043 | F. curvatum | 4.6 | b-g | |
| AFP194 | F. nirenbergiae | 12.8 | j-o | AFP222 | F. clavum | 4.6 | b-g | |
| AFP059 | F. brachygibbosum | 12.4 | j-p | AFP007 | Fusarium sp. | 4.4 | b-g | |
| AFP213 | F. nirenbergiae | 12.2 | j-p | AFP066 | F. fredkrugeri | 4.4 | b-g | |
| AFP095 | F. solani | 10.4 | k-q | AFP096 | F. curvatum | 4.4 | b-g | |
| AFP033 | F. nematophilum | 10.2 | k-q | AFP130 | F. curvatum | 4.2 | c-g | |
| AFP040 | Fusarium sp. | 9.8 | l-q | AFP018 | Fusarium sp. | 4.0 | d-g | |
| AFP001 | F. solani | 8.6 | m-q | AFP095 | F. solani | 4.0 | d-g | |
| AFP196 | F. clavum | 8.4 | m-q | AFP261 | F. solani | 4.0 | d-g | |
| AFP062 | F. clavum | 6.6 | n-q | AFP033 | F. nematophilum | 3.8 | e-g | |
| AFP048 | Fusarium sp. | 6.4 | n-q | AFP062 | F. clavum | 3.8 | e-g | |
| AFP222 | F. clavum | 5.2 | o-q | AFP196 | F. clavum | 3.8 | e-g | |
| AFP013 | F. nanum | 3.6 | p-q | AFP059 | F. brachygibbosum | 3.7 | f-g | |
| Non-inoculated Control | 2.6 | q | Non-inoculated Control | 3.0 | g | |||
| LSD = 9.06 | LSD = 1.87 | |||||||
*Means accompanied by same letter are not significantly different (P = 0.05) according to LSD tests.
Table 6.
Mean root dry weights of 1103 Paulsen rootstock plants inoculated with Fusarium species after four months.
Table 6.
Mean root dry weights of 1103 Paulsen rootstock plants inoculated with Fusarium species after four months.
| Isolates | 2022 | Root Dry | Isolates | 2023 | Root Dry | |||
| Species | Weight (g) | Species | Weight (g) | |||||
| Non-inoculated Control | 0.344 | a* | Non-inoculated Control | 1.463 | a | |||
| AFP041 | F. curvatum | 0.224 | b | AFP222 | F. clavum | 1.436 | b | |
| AFP033 | F. nematophilum | 0.196 | c | AFP048 | Fusarium sp. | 1.425 | c | |
| AFP256 | Fusarium sp. | 0.182 | d | AFP062 | F. clavum | 1.417 | d | |
| AFP123 | F. solani | 0.150 | e | AFP013 | F. nanum | 1.410 | e | |
| AFP098 | F. glycines | 0.146 | f | AFP101 | F. curvatum | 1.399 | f | |
| AFP018 | Fusarium sp. | 0.134 | g | AFP033 | F. nematophilum | 1.369 | g | |
| AFP062 | F. clavum | 0.125 | h | AFP075 | Fusarium sp. | 1.353 | h | |
| AFP261 | F. solani | 0.125 | h | AFP261 | F. solani | 1.349 | i | |
| AFP048 | Fusarium sp. | 0.119 | i | AFP038 | F. falciforme | 1.336 | j | |
| AFP013 | F. nanum | 0.112 | j | AFP001 | F. solani | 1.327 | k | |
| AFP130 | F. curvatum | 0.111 | j | AFP095 | F. solani | 1.286 | l | |
| AFP196 | F. clavum | 0.109 | jk | AFP007 | Fusarium sp. | 1.282 | m | |
| AFP019 | F. solani | 0.108 | k | AFP019 | F. solani | 1.266 | n | |
| AFP109 | F. annulatum | 0.104 | l | AFP040 | Fusarium sp. | 1.251 | o | |
| AFP222 | F. clavum | 0.103 | l | AFP059 | F. brachygibbosum | 1.245 | p | |
| AFP038 | F. falciforme | 0.098 | m | AFP098 | F. glycines | 1.232 | q | |
| AFP004 | F. annulatum | 0.087 | n | AFP256 | Fusarium sp. | 1.231 | q | |
| AFP001 | F. solani | 0.083 | o | AFP066 | F. fredkrugeri | 1.181 | r | |
| AFP040 | Fusarium sp. | 0.082 | o | AFP191 | F. curvatum | 1.172 | s | |
| AFP007 | Fusarium sp. | 0.078 | p | AFP103 | F. annulatum | 1.160 | t | |
| AFP194 | F. nirenbergiae | 0.072 | q | AFP018 | Fusarium sp. | 1.118 | u | |
| AFP096 | F. curvatum | 0.070 | q | AFP196 | F. clavum | 1.108 | v | |
| AFP213 | F. nirenbergiae | 0.065 | r | AFP096 | F. curvatum | 1.101 | w | |
| AFP043 | F. curvatum | 0.063 | r | AFP006 | F. annulatum | 1.098 | x | |
| AFP114 | F. annulatum | 0.058 | s | AFP213 | F. nirenbergiae | 1.069 | y | |
| AFP059 | F. brachygibbosum | 0.054 | t | AFP043 | F. curvatum | 1.056 | z | |
| AFP101 | F. curvatum | 0.054 | t | AFP115 | Ilyonectria liriodendri | 1.047 | a1 | |
| AFP103 | F. annulatum | 0.051 | tu | AFP123 | F. solani | 1.026 | b1 | |
| AFP006 | F. annulatum | 0.050 | u | AFP111 | F. curvatum | 1.016 | c1 | |
| AFP075 | Fusarium sp. | 0.049 | uv | AFP109 | F. annulatum | 1.012 | d1 | |
| AFP095 | F. solani | 0.047 | vw | AFP037 | F. curvatum | 0.974 | e1 | |
| AFP037 | F. curvatum | 0.045 | wx | AFP041 | F. curvatum | 0.921 | f1 | |
| AFP111 | F. curvatum | 0.044 | x | AFP130 | F. curvatum | 0.919 | f1 | |
| AFP115 | Ilyonectria liriodendri | 0.042 | xy | AFP194 | F. nirenbergiae | 0.919 | f1 | |
| AFP066 | F. fredkrugeri | 0.040 | yz | AFP114 | F. annulatum | 0.901 | g1 | |
| AFP191 | F. curvatum | 0.038 | z | AFP004 | F. annulatum | 0.893 | h1 | |
| AFP265 | F. annulatum | 0.026 | a1 | AFP265 | F. annulatum | 0.880 | i1 | |
| AFP061 | F. annulatum | 0.022 | b1 | AFP061 | F. annulatum | 0.599 | j1 | |
| LSD = | 0.002 | LSD = | 0.003 | |||||
*Means accompanied by same letter are not significantly different (P = 0.05) according to LSD tests.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



