Submitted:
19 May 2024
Posted:
20 May 2024
You are already at the latest version
Abstract
Drought and arsenic contamination pose a serious threat to global rice crop yield. Existing solutions focus on either heavy metal or drought remediation, but not both. Examination of the Oryza sativa metagenome via the National Center for Biotechnology Information (NCBI) database revealed promising candidates within the aquaporin gene family capable of efficiently mitigating both arsenic toxicity and drought stress. Rice cotyledons were genetically augmented with arsenic-resistant OsNIP2;1 and OsNIP3;2 genes, and drought-resistant OsPIP2;2 genes. In moderate drought conditions and 25 ppm arsenic-contaminated soil, transgenic plants showed drought tolerance and reduced translocation of arsenic from root to shoot. Soil arsenic levels plummeted from 25 ppm to 5 ppm, demonstrating the strategy's efficacy. Statistical significance was confirmed using T tests. Thus, aquaporin-augmented Oryza sativa offers a promising solution to mitigate both arsenic and drought stress in rice plants improving crop yield and facilitating soil decontamination.
Keywords:
Arsenic contamination
; Rice arsenic
; genetic augmentation
; bioremediation
Introduction
Rice cultivation faces significant environmental challenges, especially with drought and arsenic contamination [1]. Rice, known for its high water requirements, consumes nearly one-third of the world's freshwater resources for irrigation, as it is typically grown in flooded paddy fields to boost yields [2]. This method leads to substantial outflows which are then captured and reused downstream for higher water-use efficiency. Unfortunately, this groundwater contains arsenic. When rain is scarce, arsenic-laden groundwater is repeatedly used, causing arsenic buildup in the soil over time, a notable cause of arsenic contamination in rice [3] (Figure 1). In addition, arsenic contamination of soil is a known global public health concern. According to the EPA, soil arsenic levels ranging from 5 ppm to 20 ppm are generally considered safe [4]. Arsenic easily enters the environment through mining, smelting, and volcanic eruptions. Inorganic arsenate [As(V)] and arsenite [As(III)] are predominant in aquatic and soil environments. Current methods to enhance drought tolerance, such as improved breeding techniques, efficient water management practices, bio-stimulants, have limitations due to environmental variability, cost, and resource constraints. Current heavy metal remediation techniques, including landfilling, vitrification, and electrokinetics, are expensive and potentially toxic, making them suboptimal [4]. Moreover, these approaches often focus separately on either drought or heavy metal tolerance rather than addressing both simultaneously. Therefore, developing rice varieties capable of withstanding the coupled stress of arsenic and drought is paramount, addressing the impending food crisis while remediating arsenic-contaminated soil. To enhance rice plants' dual tolerance to arsenic and drought, the transporter mechanism for water and mineral movement was researched. Among the many genes associated with these processes, the aquaporin gene family emerged as a significant player in drought and salinity responses, stomatal regulation, and biotic stress responses [5,6]. Genes from the aquaporin family, namely Oryza sativa Plasma Membrane Intrinsic Proteins (OsPIPs) and Oryza sativa NOD26-like Intrinsic Proteins (OsNIPs), were selected for experimentation. Of the NIP genes, OsNIP2;1 and OsNIP3;2 were hyper expressed. OsNIP2;1 and OsNIP3;2 are influx only proteins facilitating As(III) entry into root tissues and are mainly expressed in primary and secondary roots, respectively [7]. The OsPIP2;2 gene is mainly expressed in primary plant roots, contributing to stomatal closure and water channel activation during drought stress. PIP2;2 protein also enhances the diffusion of proline, polyamines, and plant hormones such as ABA and Ethylene, produced under drought stress, which facilitates rapid stomatal closure, act as free radical scavengers, and aid in accumulating osmotically active compounds that protect cells from damage [8,9,10].
The aquaporin genes were obtained from a research laboratory at the University of Florida, and transgenic rice plants were developed using Agrobacterium tumefaciens, a gram-negative soil bacterium, infects plant cells and transfers genetic material. Once transgenic rice plants are developed, they will undergo various experimental conditions to test the hypothesis: "If Oryza sativa is transformed with OsPIP2;2, OsNIP2;1, and OsNIP3;2 genes and grown in drought-simulated arsenic-contaminated soil, it will accumulate significantly less arsenic in the leaves than non-transformed plants, exhibit improved overall plant health and substantially decrease arsenic in the soil." The project's objective was to devise a strategy for producing rice plants that can tolerate both drought and arsenic, prevent arsenic accumulation in rice grains for safe consumption, and decontaminate the soil of arsenic allowing crop cultivation in lands which were once non-arable.
Methods
The National Center for Biotechnology Information (NCBI) served as the platform for analyzing and comparing multiple Oryza sativa genes and their respective functionalities [11]. Once the desired family was identified, the protein sequences of the individual genes were compared using the BLAST (Basic Local Alignment Search Tool) bioinformatics tool to identify functional relationships between the selected genes to finally select the desired genes for experimentation.
Paddy seeds were germinated by soaking them in moisture for four days until cotyledons became visible. A. tumefaciens were transformed with OsPIP2;2, OsNIP2;1, and OsNIP3;2 genes with the help of a local biotech company. Transformation was confirmed by growing the Agrobacterium strains tagged with hygromycin-resistant genes in LB agar plates pre-treated with hygromycin. Small cuts were made on the cotyledons, and then transferred onto co-culture MS media along with transformed Agrobacterium and incubated for five days until a callus became visible in the plant tissue. The cotyledons with callus were incubated for sixty minutes in the MS resting media containing Kanamycin antibiotic to eliminate A. tumefaciens. Sterile cotyledons with callus were transferred into the regeneration media and incubated for fourteen days to facilitate root and shoot formation to finally develop transgenic rice plants (Figure 2). These were then transferred into the experimental pots. Pots were divided into one positive control, two negative control, and one experimental group.

Table 1.
Experimental and Control groups.
|
Positive control (P) Soil with arsenic & no plants |
Negative control (N1): No arsenic in soil & non-transgenic plant |
|
Negative control (N2) Soil with arsenic & non-transgenic plant |
Experimental group: Soil with arsenic & transgenic plants with NIP & PIP genes |
All experimental and N2 control plants were moderately drought-stressed by receiving only 40% water supply (400 ml) daily. N1 control pot received the regular water supply of 1000 ml each day. Soil arsenic levels were measured using Arsenic QuickTM Kit every two weeks until eight weeks in the control (P) pot and experimental pots. After eight weeks, arsenic levels were measured in the roots and leaves of the plants in the N2 control pot and experimental pots. At the end of 8 weeks, the plants from the N1 and N2 control and experimental groups were assessed for their length and chlorophyll content of the leaves using Vernier Go Direct Spectrophotometer. The Isatin test was conducted on the leaves of the N2 control and experimental group to detect markers of drought stress in the plants, such as proline or polyamines in the plant protoplast. Finally, statistical analysis was conducted using T tests.
Results
NCBI was used to analyze 32,000+ genes in O. sativa indica and O. sativa japonica subspecies. Among these genes, aquaporin family, particularly Plasma Membrane Intrinsic Proteins (PIPs) and NOD26-like Intrinsic Proteins (NIPs), were identified as the genes of interest. The protein sequences of selected PIPs and NIPs (OsPIP2;2, OsNIP2;1, and OsNIP3;2) were analyzed using BLAST (Basic Local Alignment Search Tool) bioinformatics tool for their functional relationship (Figure 3).
Rice plants transformed with OsPIP2;2, OsNIP2;1, and OsNIP3;2 genes using A. tumefaciens were successfully cultivated over eight weeks in drought simulated arsenic contaminated soil and results were analyzed. At the end of 8 weeks, arsenic level in the leaves of the dual stressed transgenic plants was undetectable compared to 15 ppm in the control (N2), length of the plants was 18” whereas those of control plants (N2) was 8" with p value 0.048 (Figure 4). In addition, the chlorophyll content of the leaves of the dual stressed plants was comparable to the leaves from N1 control pots under no arsenic or drought stress, while the chlorophyll content in the leaves of the N2 control plants was much lower compared to the N1 control and experimental plants (Figure 5).
After eight weeks, soil arsenic content decreased by 80% from 25 to 5 ppm in transgenic plants compared to only 40% from 25 ppm to 15 ppm in non-transgenic plants (Table 2) with p value 0.038.
Discussion and Conclusions
NCBI was utilized to untangle the complexity of the Oryza sativa metagenome. After identifying the gene family, using BLAST, it was concluded that the OsPIP2;2, OsNIP2;1, and OsNIP3;2 genes would provide the desirable results as they coded for the transport channels that facilitated movement of water and small ion particles across the cell membranes.
Both OsNIP2;1, and OsNIP3;2 genes were selected because OsNIP2;1 gene predominantly expressed in the primary root, while the OsNIP3;2 gene expressed in the lateral roots, and therefore expressing both increased the absorption surface area several thousand times. Both these genes encode for influx proteins which would only allow arsenic entry into the cell while preventing its efflux, which decreased root to shoot translocation eliminating arsenic accumulation in rice grain while simultaneously reducing arsenic level in the soil.
In addition, the spectrophotometric analysis of the chlorophyll content of experimental leaves were comparable to that of non-transgenic plant leaves in a non-drought environment, which showed that the genetic transformation of rice plants and arsenic decontamination in the soil did not negatively affect the plant health. Furthermore, the genes that were hyper expressed belonged to the Oryza sativa family which further increases the crop safety for consumption.
The results can be translated into developing a sustainable, ecofriendly and cost-effective solution for developing rice plants that are robust in withstanding the dual stress of drought and arsenic toxicity. Combined drought-arsenic tolerant rice will not only increase crop production by nearly 50%, it will help in remediating the soil of the arsenic and convert several acres of contaminated non-arable lands to arable lands in drought prone areas which would otherwise be unused.
Acknowledgments
I would like to thank my science fair sponsor Mrs. Julie Baker for her support and guidance throughout the project. I also would like to express my sincere gratitude to Dr. Lee Tarpley, distinguished professor at Texas A&M Agriculture Research Extension Center, Beaumont, Texas, who selected me for an internship and guided me on drought resistance mechanisms and plant transformation.
References
- Muehe, E. M., Wang, T., Kerl, C. F., Planer-Friedrich, B., & Fendorf, S. (2019). Rice production threatened by coupled stresses of climate and soil arsenic. Nature Communications, 10(1). [CrossRef]
- Satyapal, G. K., & Kumar, N. (2021). Arsenic: Source, distribution, toxicity and bioremediation. Arsenic Toxicity: Challenges and Solutions, 153-163. [CrossRef]
- Panda, D., Mishra, S. S., & Behera, P. K. (2021). Drought tolerance in rice: Focus on recent mechanisms and approaches. Rice Science, 28(2), 119-132. [CrossRef]
- Selvi, Adikesavan. (2019). Integrated Remediation Processes Toward Heavy Metal Removal/Recovery From Various Environments. Front. Environ. Sci., Volume 7, 22 May 2019. [CrossRef]
- Cai, L., Liu, G., Rensing, C., & Wang, G. (2009). Genes involved in arsenic transformation and resistance associated with different levels of arsenic-contaminated soils. BMC Microbiology,9(1). [CrossRef]
- Chen, Jian, and Barry P. Rosen. “Biosensors for Inorganic and Organic Arsenicals.” Biosensors, vol. 4, no. 4, 2014, pp. 494–512. [CrossRef]
- Sakurai, J., Ishikawa, F., Yamaguchi, T., Uemura, M., & Maeshima, M. (2005). Identification of 33 rice Aquaporin genes and analysis of their expression and function. Plant and Cell Physiology, 46(9), 1568-1577. [CrossRef] [PubMed]
- Garbinski, L. D., Rosen, B. P., & Chen, J. (2019). Pathways of arsenic uptake and efflux. Environment International, 126, 585-597. [CrossRef] [PubMed]
- Abedi, Tayebeh, and Amin Mojiri. “Arsenic Uptake and Accumulation Mechanisms in Rice Species.” Plants, vol. 9, no. 2, 2020, p. 129. [CrossRef]
- Chen, Y., Han, Y., Cao, Y., Zhu, Y., Rathinasabapathi, B., & Ma, L. Q. (2017). Arsenic transport in rice and biological solutions to reduce arsenic risk from rice. Frontiers in Plant Science, 8. [CrossRef]
- Sayers EW, Bolton EE, Brister JR, Canese K, Chan J, Comeau DC, Connor R, Funk K, Kelly C, Kim S, Madej T, Marchler-Bauer A, Lanczycki C, Lathrop S, Lu Z, Thibaud-Nissen F, Murphy T, Phan L, Skripchenko Y, Tse T, Wang J, Williams R, Trawick BW, Pruitt KD, Sherry ST. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022 Jan 7;50(D1):D20-D26. [CrossRef] [PubMed] [PubMed Central]
- 1Shackel, K. (2011). A plant-based approach to deficit irrigation in trees and vines. HortScience, 46(2), 173-177. [CrossRef]
Figure 1.
Diagram showing relationship between arsenic and drought. Decreased water and higher temperature reduces adsorption of arsenic resulting in increased arsenic availability for uptake by plants.
Figure 1.
Diagram showing relationship between arsenic and drought. Decreased water and higher temperature reduces adsorption of arsenic resulting in increased arsenic availability for uptake by plants.
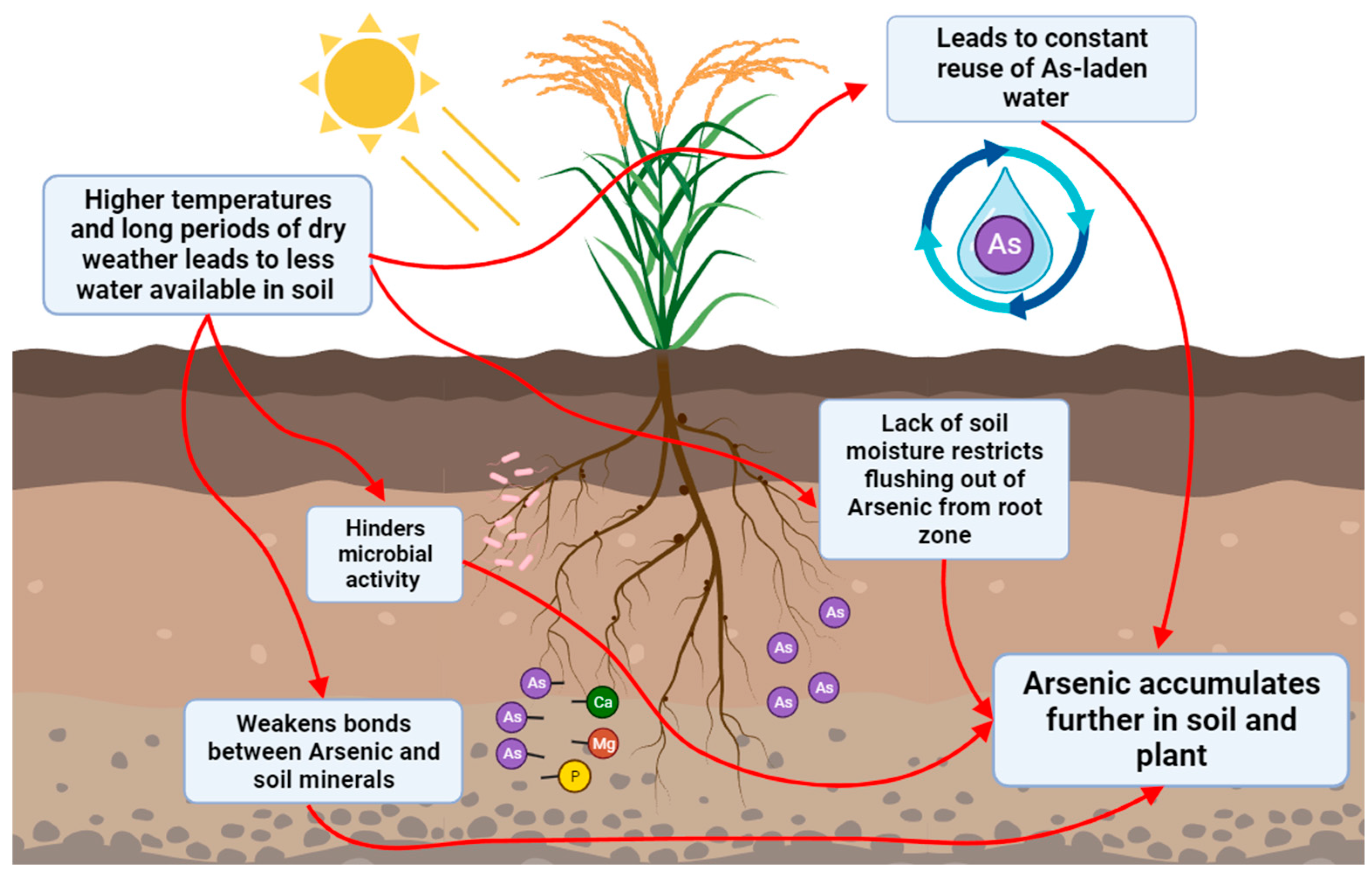
Figure 2.
Developing the Transgenic Rice plant using transformed A. tumefaciens. A) Rice cotyledons with the husk B) pCAMBIA1300 plasmid carrying OsNIP and OsNIP genes C) Transformed A. tumefaciens in LG agar plate D) Wounded rice cotyledons transformed with A. tumefaciens in MS tissue culture media E) Visible callus after 5 days F) Transgenic rice plant developed in regeneration media.
Figure 2.
Developing the Transgenic Rice plant using transformed A. tumefaciens. A) Rice cotyledons with the husk B) pCAMBIA1300 plasmid carrying OsNIP and OsNIP genes C) Transformed A. tumefaciens in LG agar plate D) Wounded rice cotyledons transformed with A. tumefaciens in MS tissue culture media E) Visible callus after 5 days F) Transgenic rice plant developed in regeneration media.

Figure 3.
Comparing the protein sequences of the individual genes using BLAST bioinformatics tool.
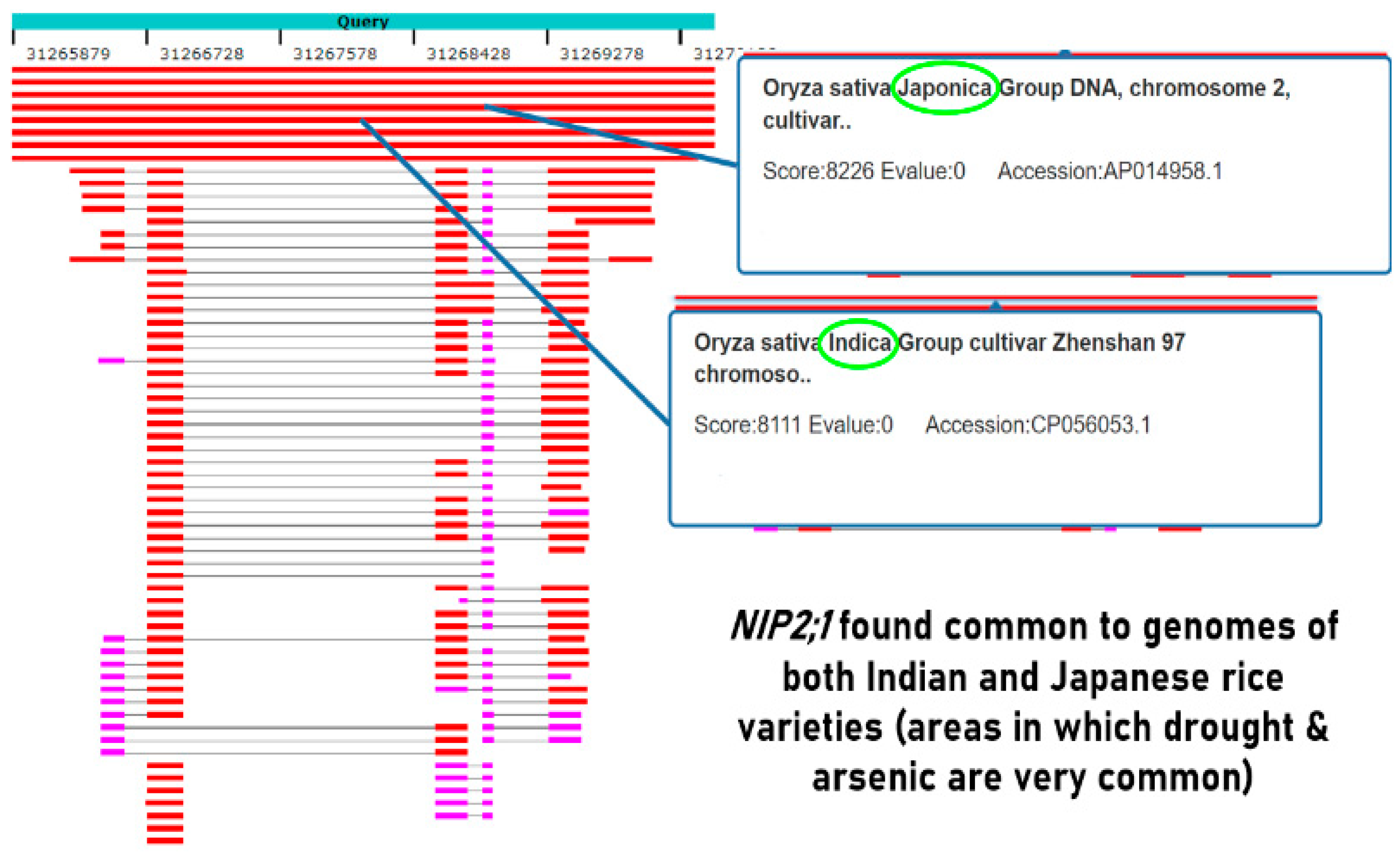
Figure 4.
Graph showing increase in plant height over time. At eight weeks, average length of the transgenic plants was much higher than non-transgenic control plants exposed to drought and arsenic, and similar to control plants not exposed to drought and arsenic.
Figure 4.
Graph showing increase in plant height over time. At eight weeks, average length of the transgenic plants was much higher than non-transgenic control plants exposed to drought and arsenic, and similar to control plants not exposed to drought and arsenic.
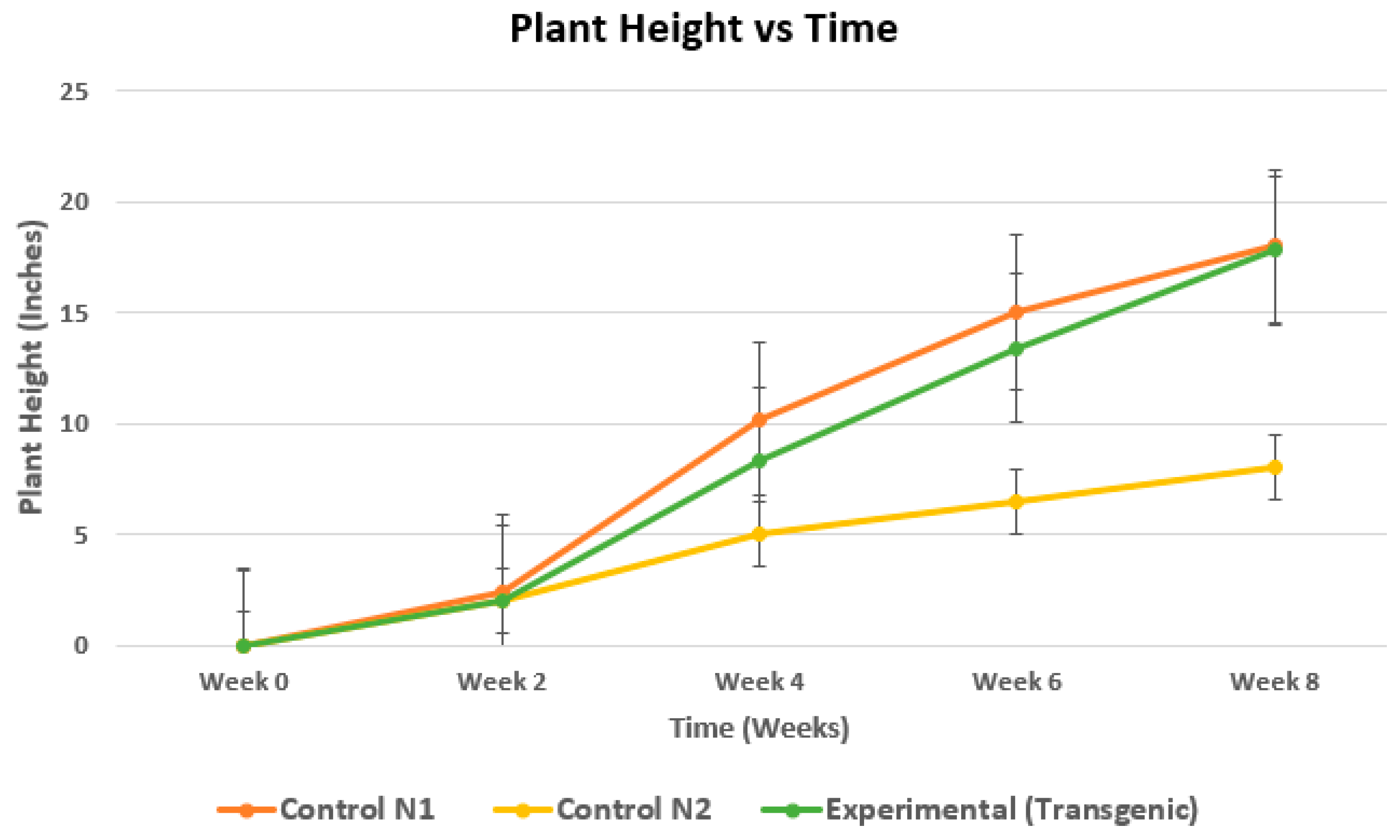
Figure 5.
Absorbance values measured by optical density. The absorbance values of all the transgenic plants (5 trials) subjected to the dual stress of arsenic and drought was similar and comparable to the non-transgenic plants in control pots (N1) not subjected to arsenic or drought stress, and higher than control (N2) subjected to dual stress.
Figure 5.
Absorbance values measured by optical density. The absorbance values of all the transgenic plants (5 trials) subjected to the dual stress of arsenic and drought was similar and comparable to the non-transgenic plants in control pots (N1) not subjected to arsenic or drought stress, and higher than control (N2) subjected to dual stress.
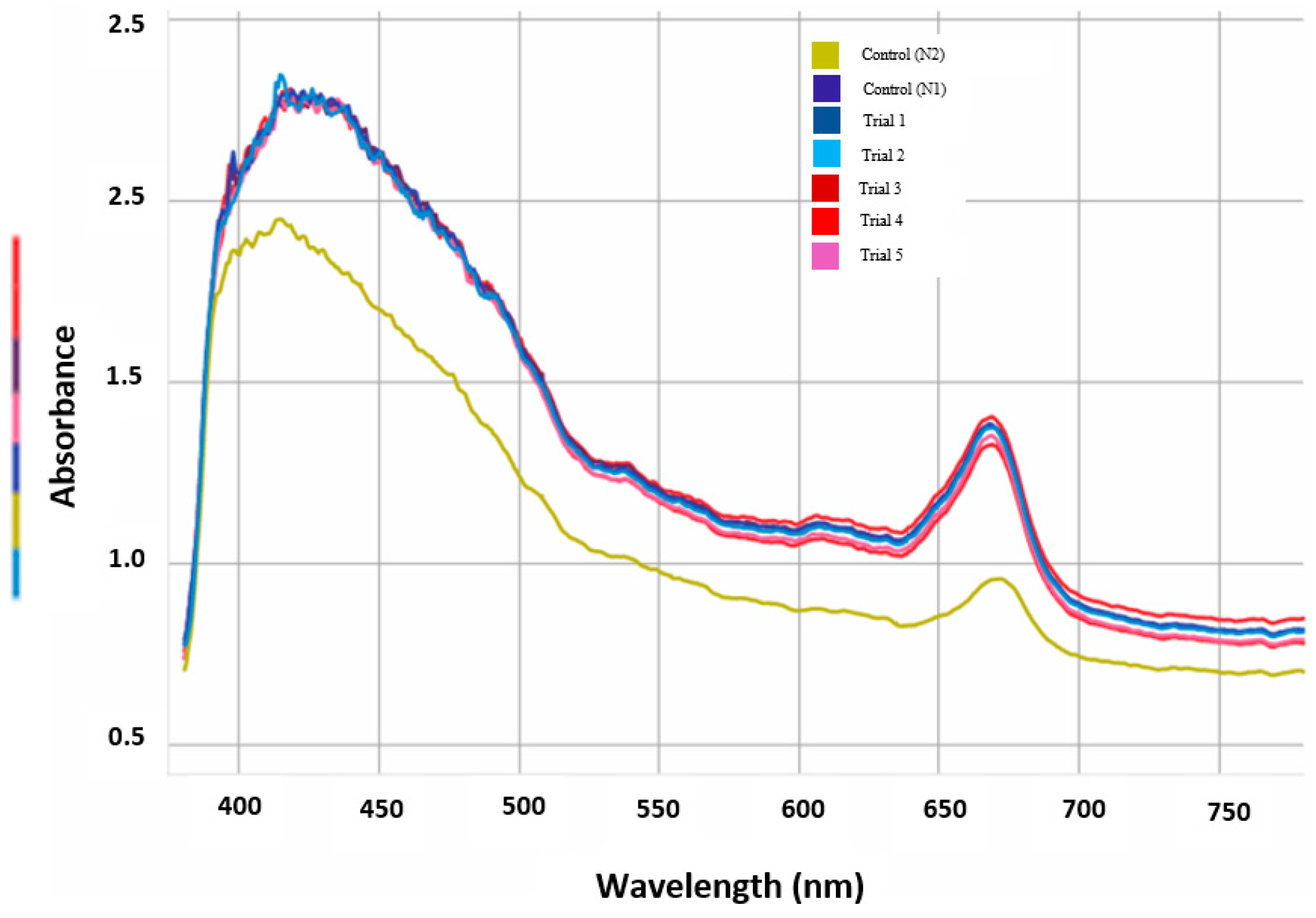
Table 2.
Soil Arsenic level at various stages of plant growth.
| Week 0 | Week 2 | Week 4 | Week 6 | Week 8 | |
|---|---|---|---|---|---|
| Control (P) | 25 ppm | 25 ppm | 25 ppm | 25 ppm | 25 ppm |
| Control N2 | 25 ppm | 25 ppm | 25 ppm | 20 ppm | 15 ppm |
| Transgenic | 25 ppm | 25 ppm | 15 ppm | 15 ppm | 5 ppm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



