
Submitted:
20 May 2024
Posted:
21 May 2024
You are already at the latest version
Abstract
Endometriosis is a prevalent chronic inflammatory disease characterized by a considerable delay between initial symptoms and diagnosis through surgery. The pressing need for a timely, non-invasive diagnostic solution underscores the focus of current research efforts. This study examines the diagnostic potential of the menstrual blood lipidome. The lipid profile of 39 samples (23 women with endometriosis and 16 patient of control group) was acquired using reverse-phase high-performance liquid chromatography-mass spectrometry with LipidMatch processing and identification. Profiles were normalized based on total ion counts. Significant differences in lipids were determined using the Mann-Whitney test. Lipids for the diagnostic model, based on logistic regression, were selected using a combination of variance importance projection filters and Akaike information criteria. Levels of ceramides, sphingomyelins, cardiolipins, triacylglycerols, acyl- and alkenyl-phosphatidylethanolamines, and alkenyl-phosphatidylcholines increased, while acyl- and alkyl-phosphatidylcholines decreased in cases of endometriosis. PE P-16:0/18:1 and CL 16:0_18:0_22:5_22:6 serve as marker lipids in the diagnostic model, exhibiting a sensitivity of 81% and specificity of 85%. The diagnostic approach based on dried spots of menstrual blood holds promise as an alternative to traditional non-invasive methods for endometriosis screening.
Keywords:
lipid
; endometriosis
; dried blood spot
; mass-spectrometry
; diagnostic
1. Introduction
Endometriosis is a chronic inflammatory disease that affects 10% of women of reproductive age. The progressive nature of endometriosis underscores the necessity of early diagnosis to conserve healthcare resources and improve patients’ quality of life through personalized approaches [1,2]. Timely diagnosis is hindered by the nonspecificity of clinical symptoms, which also characterize other diseases [3]. In some cases, endometriosis progresses without symptoms, leading to delays in diagnosis of more than 10 years [4,5].
While laparoscopy combined with histologic confirmation serves as the gold standard for diagnosing endometriosis, boasting a sensitivity of 94% and specificity of 79%, it is highly invasive and can result in significant delays [5,6]. Ultrasonography and magnetic resonance imaging of pelvic organs offer non-invasive diagnostic alternatives, but ultrasonography is operator-dependent and MRI is costly. The World Endometriosis Society has identified the development of reliable non-invasive diagnostic tools as one of the main directions in gynecology [7].
Endometriosis progression involves alterations in eutopic endometrial tissue, as demonstrated by changes in gene expression and the concentrations of proteins such as A, CYR61, annexin 1, osteopontin, and aromatase P450 [8]. These molecular alterations in tissue profiles lead to changes in uterine fluid, which can be utilized for biomarker discovery [9] [32]. Additionally, venous blood contains 122 potential biomarkers, including angiogenesis and growth factors, apoptosis markers, cell adhesion molecules, high-throughput markers, hormonal markers, immune system and inflammatory markers, oxidative stress markers, microRNAs, and tumor markers [10].
Menstrual blood serves as a non-invasive source of endometrial cells [11,12], which can be analyzed for potential endometriosis markers and easily collected from women using menstrual cups, specialized pads, or smart tampons [5,12,13]. Menstrual blood contains over 350 unique proteins not found in circulating blood or vaginal fluid [14]. Ji et al., in their study, observed increased menstrual blood levels of CXCL5 and IL1RN proteins in cases of endometriosis [15].
Additionally, alterations in the metabolomic profiles of tissue and fluid (blood, follicular fluid) occur in cases of endometriosis [16,17,18,19] with lipids playing a significant role in these profile changes [19]. Mass spectrometry analysis methods, coupled with chromatography or direct analysis (SELDI-, MALDI, shotgun), are effective for metabolic profiling of biological samples and have successfully been used in the search for endometriosis markers [19,20,21]. Dried blood spot samples require minimal biological material and have been successfully utilized in diagnostic research [22,23]. The aim of our study is to explore the possibilities of diagnosing endometriosis through the lipid profiles of dried menstrual blood spot.
2. Materials and Methods
2.1. Study Desing
This study involved samples from 23 patients with histologically verified endometriosis stages I-IV, as per the revised American Fertility Society (rAFS) classification, along with 16 patients comprising the control group. Menstrual blood samples from patients with endometriosis were collected prior to surgery.
Patients were invited on days 2-3 of their menstrual cycle (during the days of heaviest menstrual bleeding). Menstrual blood samples were collected on a gynecological chair, using a Cusco plastic speculum inserted and then removed, allowing the menstrual blood to flow onto a special form with five equally sized compartments. Application was made only on one side of the form, ensuring each drop of blood fully saturated the form. Drops could be larger than the compartments but never smaller. Blood drops were air-dried before being placed in an envelope for transportation. The collected sample was then placed in a plastic bag with a zip-lock and stored in a freezer at -20°C. Documentation was completed beforehand, including the patient's surname, first name, patronymic, age, endometriosis phenotype, outpatient card number, menstrual cycle day, and date of sample collection.
Inclusion criteria:
For all participants:
- -
- women aged 18-45 with regular menstrual cycles;
- -
- signed informed consent;
- -
- residing in Moscow.
For participants in the endometriosis group:
- -
- endometriosis confirmed by surgical intervention or instrumental visualization methods (ultrasound and/or magnetic resonance imaging of the pelvic organs);
- -
- presence of one or more endometriosis symptoms: dysmenorrhea and/or dyspareunia, and/or chronic pelvic pain, and/or infertility.
For participants in the control group:
- -
- absence of infertility (one or more childbirths);
- -
- regular menstrual cycle;
- -
- absence of severe dysmenorrhea;
- -
- absence of endometriosis confirmed by ultrasound investigation.
Exclusion criteria:
- -
- chronic diseases other than endometriosis (diabetes, hypertension, etc.);
- -
- infectious diseases: HIV and viral hepatitis (B and C);
- -
- hormonal medication intake within 3 months prior to study inclusion.
The main group consisted of 23 women (age 28 (25;33) years; BMI 19.4 (18.1;22.1) kg/m2) with external genital endometriosis (superficial endometriosis, ovarian endometriomas, deep endometriosis), identified by magnetic resonance imaging of the pelvic organs and confirmed histologically after laparoscopy. The control group included 16 women (age 27 (24;33) years; BMI 22.7 (19.7;24.9) kg /m2). Groups were comparable in age and BMI (p>0.05).
In the main group, in most cases there was a combination of various forms of endometriosis: superficial endometriosis with ovarian endometriomas in 26.1% (6/23), ovarian endometriomas and deep endometriosis in 43.5% (10/23), superficial endometriosis with deep endometriosis 17.4% (4/23) and isolated ovarian endometriomas were present in 13% of cases (3/23). The leading complaint of the majority of patients from the main group was dysmenorrhea (visual analog scale score was 7.0 (5.0; 9.0)), as well as abnormal uterine bleeding (heavy menstrual bleeding) and infertility.
2.2 Lipid Extraction
The lipid extract was obtained using a modified Folch method extraction procedure. The dried blood spot was removed and placed in a 2 mL microtube. To this, 480 µL of CHCl3/MeOH (2:1, v/v) was added, and the tube was vortexed using ultrasound for 15 minutes. Subsequently, 150 µL of HPLC-grade water was added, and the mixture was vortexed using ultrasound for another 15 minutes. The tube was then centrifuged at 15,000 rounds per minute for 10 minutes. After centrifugation, 150 µL of the organic layer was collected in a 1.5 mL Eppendorf tube, and 250 µL of CHCl3/MeOH (2:1, v/v) was added to the microtube again. The mixture was vortexed for 5 minutes and centrifuged at 15,000 rounds per minute for 10 minutes. Next, 250 µL of the organic layer was collected and added to the previous 150 µL. The lipid extract was dried under a nitrogen flow at 40°C and then redissolved in 150 µL of isopropanol/acetonitrile (1:1 v/v). The mixture was vortexed for 5 minutes and centrifuged at 15,000 rounds per minute for 5 minutes. Finally, 100 µL of the mixture was collected for chromatography-mass spectrometry analysis.
2.3 HPLC-MS Analysis
High-performance liquid chromatography coupled with mass spectrometry (HPLC-MS) was conducted using the 3000 Ultimate Nano LC system (Thermo Scientific, Waltham, MA, USA) and the Bruker MaXis Impact instrument (Bruker Daltonics, Bremen, Germany). Reverse-phase chromatography separation was achieved using a Zorbax XDB-C18 column (3.5 µm, 150 mm length, 0.5 mm inner diameter, Agilent, USA) with mobile phase A consisting of an acetonitrile/water mixture (60/40 v/v) and mobile phase B consisting of isopropanol/acetonitrile/water (90/8/2 v/v/v). Both mobile phases contained modifiers: 0.1% formic acid and 10 mM ammonium formate. The flow rate was set at µL/min for 22 minutes with a temperature of 50°C, and the composition of phase B changed during analysis as follows:
- 0-0.5 minutes: 15% B;
- linear gradient from 10% to 99% B over 15 minutes;
- 99% B for 4 minutes;
- linear gradient from 99% to 15% B over 15 minutes;
- 15% B for 2 minutes.
Electrospray ionization parameters were set as follows: a capillary voltage of 4.1 kV for positive ion mode and 3.0 kV for negative ion mode, spray gas pressure of 0.7 bar, drying gas flow rate of 6 L/min, and a drying gas temperature of 200°C. The mass spectra range was 100-1700 m/z with a resolution of 50,000. Tandem mass spectrometry was performed using data-dependent analysis with the following parameters: the three most abundant peaks were selected after a full mass scan, and the corresponding ions were fragmented by collision-induced dissociation using a collision energy of 35 eV and a mass exclusion time of 1 minute.
2.4 Data Processing
Data preprocessing and lipid identification were conducted using Koelmel's et al. pipeline with MzMine 2.3 and LipidMatch [24]. The peak areas of the lipids were normalized by the sum of peak areas (see Supplementary 1).
Statistically significant differences in lipids were identified using the Mann-Whitney test with a threshold of p<0.05. Feature selection for the diagnostic model was performed based on a combined dataset of negative ion mode and positive ion mode. This was achieved by a combination of variance important projection (VIP) filtering in orthogonal projection to latent structures (OPLS) and subsequent feature selection in logistic regression based on Akaike information criteria (AIC) [25,26]. The dependent value was assigned as 0 for the control group and 1 for the endometriosis group.
A threshold value for the dependent variable was calculated based on the results of receiver operating characteristic (ROC) curve analysis from cross-validation control. This analysis was conducted through 100 repetitions of a "train"/"test" split of 70/30. The threshold value was determined as the value that maximized the sum of sensitivity and specificity.
Search of metabolic pathways, associated with endometriosis, was performed by ConsenscusPathDB http://cpdb.molgen.mpg.de/[27] with over-representation analysis base on Wikipathways, SMPDB, KEGG, Reactome, PID, Biocarta, Ehmn, Humancyc, INOH, Netpath, Signallink.
3.1. Dried Menstrual Blood Lipids Profiling
In positive and negative ion modes, 98 and 107 lipids were identified, respectively (Supplementary 1, Figure S1 Supplementary2, and Figure S2 Supplementary2). These lipids belong to various classes, including (lyso)phosphatidylcholine, (lyso)phosphatidylethanolamine, cardiolipins, plasmanyl- and plasmenyl lipids, oxidized lipids, cholesterol esters, sphingomyelins, and triacylglycerols. Moreover, the levels of 40 lipids showed statistically significant differences (p<0.05) between endometriosis and control groups.
In the endometriosis group, menstrual blood ceramides, cardiolipins, oxidized lipids, PC 16:0_18:0, (plasmenyl-)phosphatidylethanolamines, plasmanyl-phosphatidylcholines, sphingomyelins, and triacylglycerols were increased. Conversely, the levels of (plasmenyl-)phosphatidylcholines, SM d18:0/18:0, and CerP d18:0/22:0 decreased in the endometriosis cases (Figure 1 Supplementary2 and Table 1). Figure 1 presents the volcano plot of lipids identified, emphasizing the potential markers with a fold change between endometriosis and control samples more than 1.5.

Table 2.
Relative level of statistically significant different lipids in dried menstrual blood spots, p-value (Mann-Whitney test), and fold change of median values. Cer-NS(NP, NDS) – (phytosphingosine, dihydrosphingosine) ceramide, CerP – ceramide-1-phosphate, CL – cardiolipine, oxCL – oxidized cardiolipins, PC – phosphatidylcholine, PC O- -plasmanylphosphatidylcholines, PC P- - plasmenylphosphatidylcholines, oxPC – oxidized phosphatidylcholine , PE – phoshpatidyl-ethanolamine, PE P- - plasmenylethanolamine, SM – sphingomyelin, TG – triacylglycerol, PG - phosphatidylglycerol.
Table 2.
Relative level of statistically significant different lipids in dried menstrual blood spots, p-value (Mann-Whitney test), and fold change of median values. Cer-NS(NP, NDS) – (phytosphingosine, dihydrosphingosine) ceramide, CerP – ceramide-1-phosphate, CL – cardiolipine, oxCL – oxidized cardiolipins, PC – phosphatidylcholine, PC O- -plasmanylphosphatidylcholines, PC P- - plasmenylphosphatidylcholines, oxPC – oxidized phosphatidylcholine , PE – phoshpatidyl-ethanolamine, PE P- - plasmenylethanolamine, SM – sphingomyelin, TG – triacylglycerol, PG - phosphatidylglycerol.
| Lipid | Control | Endometriosis | P | Fold change |
|---|---|---|---|---|
| Cer-NDS d18:0/24:0 | 0.58 (0.31;0.68) | 0.78 (0.53;1.26) | 0.03 | 1.34 |
| Cer-NP t18:0/26:0 | 0.68(0.28;0.91) | 1.19 (0.85;1.83) | 0.049 | 1.76 |
| Cer-NS d18:1/16:0 | 1.30(0.74;2.04) | 2.66 (1.29;3.69) | 0.02 | 2.05 |
| Cer-NS d18:1/24:1 | 0.11(0.10;0.36) | 0.30(0.14;0.52) | 0.007 | 2.87 |
| CerP d18:0/22:0 | 5.08 (3.97;5.70) | 3.52 (2.60;5.22) | 0.046 | 0.69 |
| CL 16:0_18:0_22:5_22:6 | 0.60(0.24;1.08) | 1.97 (0.79;3.09) | 0.001 | 3.31 |
| CL 18:0_18:0_18:1_20:1 | 0.23(0.14;0.28) | 0.32(0.19;0.47) | 0.03 | 1.43 |
| CL 18:1_18:1_18:1_20:3 | 0.81(0.43;1.11) | 1.64 (0.94;2.39) | 0.004 | 2.02 |
| CL 18:1_18:1_18:2_20:4 | 0.25(0.23;0.32) | 0.36(0.32;0.46) | <0.001 | 1.44 |
| CL 18:1_18:1_20:3_20:4 | 0.06(0.04;0.08) | 0.08(0.07;0.13) | 0.005 | 1.33 |
| OxCL 18:1_20:3(OOH)2_20:3(OOH)_20:3(OOH)2 | 0.04(0.03;0.07) | 0.10(0.05;0.18) | 0.03 | 2.38 |
| OxPC 16:0_18:1(OOO) | 0.13(0.09;0.16) | 0.24(0.15;0.37) | 0.009 | 1.95 |
| OxPC 16:0_20:4(OOH) | 0.13(0.07;0.17) | 0.24(0.15;0.30) | 0.001 | 1.90 |
| OxPC 18:1_18:3(OH) | 0.18(0.12;0.28) | 0.27(0.21;0.53) | 0.02 | 1.50 |
| OxPC 20:4_16:1(COOH) | 0.05(0.04;0.08) | 0.12(0.07;0.14) | 0.003 | 2.10 |
| PC 16:0_18:0 | 1.13 (0.78;2.35) | 2.86(1.25;0.34) | 0.02 | 2.52 |
| PC 16:0_18:1 | 10.10(4.54;14.82) | 3.41 (1.70;7.90) | 0.02 | 0.34 |
| PC 18:0_18:1 | 3.38 (2.50;3.90) | 1.98 (1.27;3.33) | 0.007 | 0.59 |
| PC 18:0_18:2 | 5.37 (3.68;6.82) | 2.29 (1.38;4.71) | 0.006 | 0.43 |
| PC 18:1_18:2 | 1.35 (1.06;2.48) | 0.78(0.67;1.30) | 0.03 | 0.58 |
| PE 16:0_18:1 | 0.32(0.28;0.35) | 0.38(0.34;0.41) | 0.004 | 1.18 |
| PE 16:0_18:2 | 0.15(0.10;0.29) | 0.22(0.16;0.29) | 0.04 | 1.48 |
| PE 16:0_20:4 | 0.37(0.30;0.45) | 0.51(0.44;0.57) | 0.005 | 1.40 |
| PE 18:1_18:2 | 0.12(0.08;0.16) | 0.15(0.14;0.17) | 0.01 | 1.34 |
| PG 18:1_18:1 | 0.33(0.22;0.40) | 0.42(0.31;0.58) | 0.04 | 1.29 |
| PC O-16:0/16:0 | 0.42(0.14;0.61) | 0.81(0.45;0.10) | 0.01 | 1.91 |
| PC O-16:0/20:4 | 0.16(0.11;0.18) | 0.22(0.19;0.25) | 0.003 | 1.37 |
| PC P-16:1/22:6 | 0.52(0.34;0.60) | 0.30(0.24;0.37) | 0.02 | 0.58 |
| PC P-18:0/18:1 | 0.46(0.39;0.55) | 0.39(0.34;0.43) | 0.03 | 0.84 |
| PE P-18:0/18:2 | 0.14(0.08;0.37) | 0.44(0.19;0.62) | 0.046 | 3.08 |
| PE P-18:0/20:4 | 2.48 (1.80;2.96) | 3.32 (2.56;3.62) | 0.03 | 1.34 |
| SM d18:0/18:0 | 0.24(0.17;0.30) | 0.20(0.14;0.23) | 0.01 | 0.83 |
| SM d18:1/24:0 | 4.05 (3.44;4.76) | 5.29 (3.98;6.08) | 0.02 | 1.31 |
| SM d20:0/18:2 | 0.27(0.18;0.36) | 0.36(0.31;0.61) | 0.003 | 1.32 |
| SM d22:6/20:2 | 0.09(0.04;0.12) | 0.12(0.08;0.15) | 0.047 | 1.37 |
| TG 16:1_18:1_18:2 | 1.72 (1.48;2.03) | 2.51 (1.66;2.91) | 0.03 | 1.46 |
| TG 16:1_18:2_18:2 | 0.28(0.19;0.33) | 0.38(0.29;0.52) | 0.01 | 1.37 |
| TG 18:1_18:2_18:2 | 1.48 (0.93;1.78) | 1.86 (1.36;2.73) | 0.03 | 1.26 |
| TG 18:1_18:2_18:3 | 0.58(0.39;1.21) | 1.10(0.07;1.61) | 0.01 | 1.92 |
| TG 18:1_22:3_8:0 | 2.18 (1.81;2.36) | 2.53 (2.20;2.95) | 0.02 | 1.16 |
Over-representation analysis revealed 168 statistically significant (FDR<0.05) enriched metabolic pathways (Table S1 Supplementary2). Glycerophospholipid biosynthesis/metabolism, sphingolipid de novo biosynthesis/metabolism, ceramide signaling, immune system, and acyl chain remodeling of CL/PE/PC present the most disturbed processes in endometriosis.
3.2. Diagnostic Model Creation
In the OPLS model utilized for feature preselection, the proportion of described dependent values was 63.2%, and the proportion of predicted dependent values was 42.2% (see Figure S3). The logistic regression model was finalized with the inclusion of cardiolipin CL 16:0_18:0_22:5_22:6 and plasmenylethanolamine PE P-16:0/18:1 (Figure 2) in the equation:
where Ix represents the relative part of the total ion chromatogram (TIC) as a percentage of compound x.
An optimal threshold value of 0.59 was determined, resulting in a sensitivity of 81% and a specificity of 85% (see Figure 3).
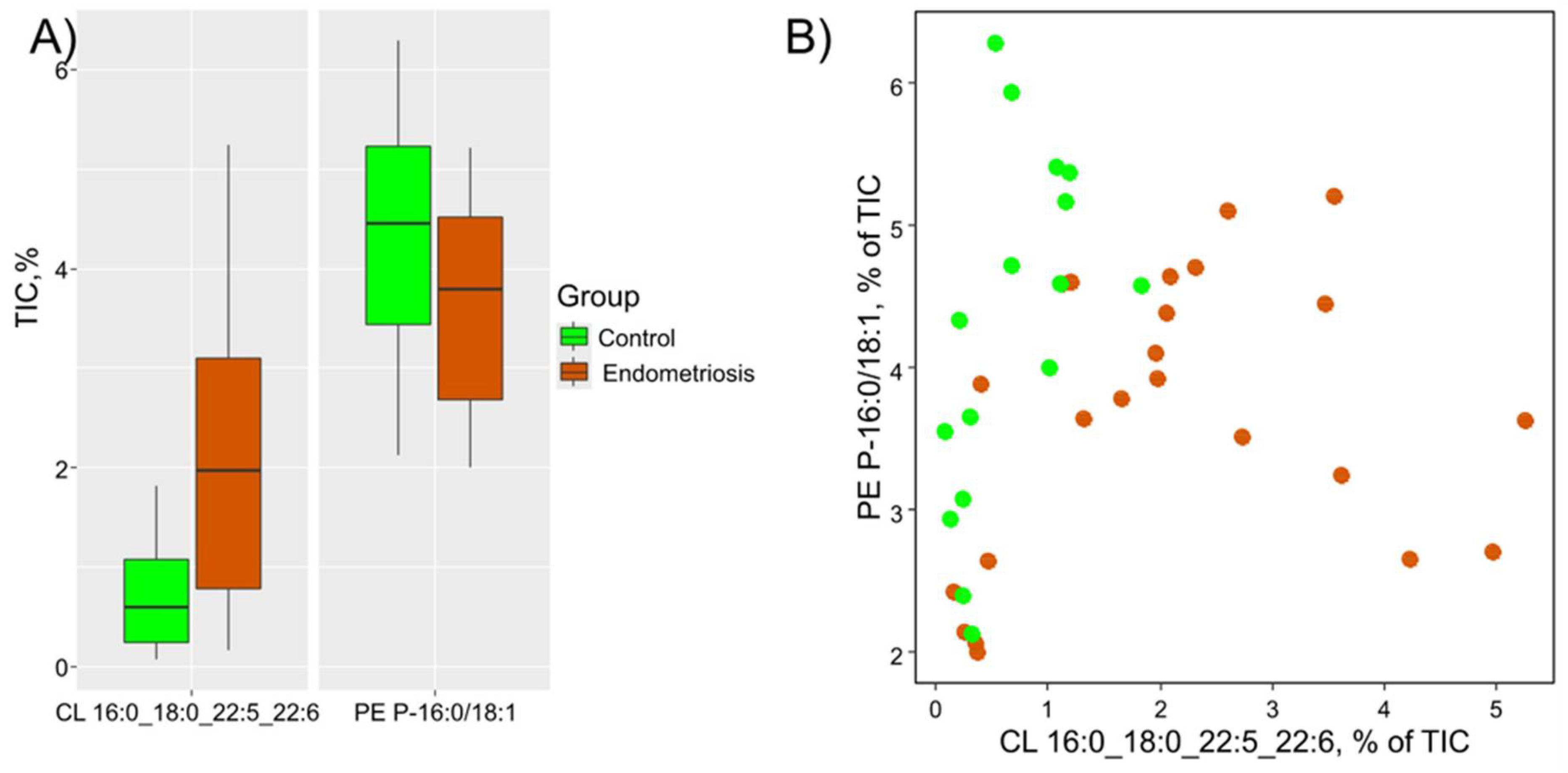
The level of cardiolipin CL 16:0_18:0_22:5_22:6 was markedly increased in menstrual blood of patients with endometriosis (FC = 3.3, p=0.001). Its significance in the equation corresponds to the calculated fold change of the median. Therefore, the menstrual blood from patients with endometriosis exhibits a higher level of CL 16:0_18:0_22:5_22:6 and a lower level of PE P-16:0/18:1 (Figure 4).
4. Discussion
Endometriosis is not just a medical condition but also a socially significant disease that impacts various aspects of women's lives. The profound impact of endometriosis goes beyond its physical symptoms and clinical manifestations, affecting women's emotional well-being, quality of life, relationships, work productivity, and overall societal participation [28]. The chronic nature of the disease, affecting only women of reproductive age, coupled with its unpredictable symptomatology (chronic pelvic pain, dysmenorrhea, dyspareunia, and gastrointestinal symptoms) and potential fertility implications, poses significant challenges in both diagnosis and management due to its chronic inflammatory nature [29]. As a progressive disease, early detection is essential to effectively preserve healthcare resources, enhance patient outcomes, and tailor treatment strategies through personalized approaches. To address the diagnostic challenges posed by the nonspecific symptoms of endometriosis, a multidimensional approach integrating medical history, physical examinations, imaging studies and minimally invasive procedures such as laparoscopy is crucial for accurate diagnosis [30]. Additionally, the development and validation of specific biomarkers and imaging modalities tailored to identify endometriosis early on could revolutionize diagnostic paradigms and enhance timely intervention.
Venous blood proteins such as VEGF, urocortin, CRP, TNF-alpha, IL-6, and follistatin or protein sets can possess a particular diagnostic strength [10], but their abnormal levels may also be caused by other diseases [31]. miRNA biomarkers also exhibit high sensitivity and specificity [32] but they entail high-cost analysis. Lipidomics, the comprehensive study of lipid molecules in biological systems, has emerged as a valuable tool in understanding the intricate interplay between lipids and various diseases [33]. Lipids play crucial roles in various cellular processes, including inflammation, immune response, and hormonal regulation, all of which are implicated in the pathogenesis of endometriosis [34]. By applying lipidomics approaches, researchers can dissect the lipid profiles in endometrial tissue, peritoneal fluid, blood, and other biological samples from individuals with endometriosis, offering deep insights into the lipid alterations associated with the disease. Studies employing lipidomics have highlighted dysregulated lipid metabolism in endometriosis, including changes in phospholipids, glycerolipids, sphingolipids, and sterols [35,36,37,38,39,40]. Furthermore, lipidomics investigations have unveiled the role of bioactive lipids, such as prostaglandins, and leukotrienes, in modulating inflammatory responses and pain perception in endometriosis [39,41]. Targeting specific lipid pathways through pharmacological interventions or dietary modifications may offer novel therapeutic strategies for managing endometriosis-related symptoms and improving patient outcomes [5]. Overall, the integration of lipidomics in endometriosis research provides a comprehensive understanding of lipid dysregulation in the disease pathogenesis, paving the way for personalized medicine approaches, biomarker discovery, and the development of targeted therapies tailored to individual lipid profiles.
Menstrual blood, known for its ease of collection and non-invasive nature, represents an abundant reservoir of endometrial cells [12]. The unique composition of menstrual blood, enriched with shed endometrial tissue during the menstrual cycle, presents a rich source of cells that reflect the dynamic changes occurring in the endometrium [42,43]. This biological fluid harbors a diverse array of cell types, including epithelial cells, mesenchymal stem cells, endothelial cells, and immune cells, each with distinct functional properties and regenerative potential [44]. Moreover, the non-invasive nature of collecting menstrual blood samples makes it an attractive option for longitudinal studies, biomarker discovery, and monitoring disease progression over time [11,12]. It is noteworthy that menstrual blood is the least researched sample in endometriosis research [32]. The convenience of collecting menstrual blood through self-collection using a menstrual cup, subsequently transferring it onto filter paper for air drying, presents dried menstrual blood spot samples as an excellent screening material [5,12,13].
Dried blood spots (DBS) are a sampling method used in screening programs to collect small, defined volumes of blood. This technique offers several advantages for screening purposes. The use of DBS enables easy collection, storage, and transportation of blood samples, as the dried spots on filter paper are stable at room temperature for extended periods [45]. This characteristic simplifies sample handling logistics and allows for decentralized sample collection, making it feasible to conduct screening programs in remote areas or community settings without sophisticated laboratory infrastructure [46]. Furthermore, DBS sampling can facilitate high-throughput screening efforts due to the ease of processing multiple samples in a cost-effective manner. Automation and standardized protocols for punching, eluting, and analyzing DBS samples can streamline the screening workflow, increase efficiency, and reduce manual labor requirements compared to traditional liquid blood sample processing methods [47]. Moreover, DBS samples are suitable for a wide range of screening tests, including genetic screening, infectious disease detection, drug monitoring, and biomarker analysis [48]. The versatility of DBS makes it a valuable tool in population-based screening programs aimed at early detection, disease prevention, and monitoring of at-risk individuals.
This study is a pioneering effort in conducting lipid profiling of menstrual blood. While previous research has primarily focused on the protein profile of menstrual blood in relation to endometriosis, our findings reveal substantial alterations in the lipidome of menstrual blood among patients with endometriosis, encompassing fifteen lipid classes. These changes likely signify a disruption in membrane lipids of shedding epithelium and immune cells, along with disturbances in intracellular lipid metabolism.
The observed trends in phosphatidylethanolamines and sphingomyelins align with alterations seen in the ectopic endometrium compared to the eutopic endometrium [49]. Notably, the levels of unsaturated phosphatidylcholines decrease in menstrual blood, in contrast to their elevation in the ectopic endometrium relative to the eutopic endometrium. Furthermore, the eutopic endometrium in patients with endometriosis also demonstrates pathological changes, underscoring the complex interplay between lipid profiles and the pathophysiology of this condition [49]. PC 16:0_18:1, PC 18:1_18:2, and PC 18:0_18:2 are decreased in menstrual blood samples in cases of endometriosis, consistent with phosphatidylcholines with the same length and saturation degree showing a similar direction of alteration in peritoneal fluid [40].
The level of plasmalogens was dramatically disturbed in endometriosis group. Moreover, PE P-16:0/18:1 was included in the diagnostic model. Research on the role of plasmalogens in this pathology is still in its early stages. However, Vouk et al. included plasma and peritoneal fluid plasmanylphosphatidylcholines in an endometriosis diagnostic model [50,51]. Studies in other disease contexts have suggested that plasmalogens can modulate inflammatory responses, regulate immune cell function, and protect cells from oxidative damage [52]. Given that chronic inflammation and immune dysregulation are key features of endometriosis, investigating the relationship between plasmalogens and these processes could provide valuable insights into the development and progression of the disease [2,28]. Moreover, plasmalogens possess both anti-inflammatory and antioxidant properties, making them potentially beneficial for alleviating the inflammatory responses and oxidative stress commonly linked to endometriosis. [53,54].
Phosphatidylglycerol PG 18:1_18:1 is one marker that increases in menstrual blood in cases of endometriosis, consistent with Feider et al.'s observation of increased levels of this lipid in endometriosis lesions compared to eutopic endometrium [55]. Moreover, we observed substantial increase of unsaturated triglycerides, mainly with oleic C18:1 and linoleic C18:2 fatty acids. Elevated levels of triglycerides have been associated with obesity, insulin resistance, and metabolic syndrome, all of which are risk factors for endometriosis. Studies have shown a potential link between high levels of triglycerides and an increased risk of developing endometriosis or experiencing more severe symptoms [56,57]. Adipose tissue is an active endocrine organ that secretes various adipokines and inflammatory mediators, which can promote inflammation and affect hormone levels, potentially contributing to the development and progression of endometriosis [58] Moreover, insulin resistance, which is associated with high triglyceride levels, may also play a role in the pathogenesis of endometriosis [59,60]. Insulin resistance can lead to increased levels of insulin and insulin-like growth factor 1 (IGF-1), which have been implicated in promoting the growth and proliferation of endometrial cells outside the uterus [61,62]. On the other hand, alterations in lipid metabolism, including changes in triglyceride levels, may be consequences rather than causes of endometriosis. Endometriosis itself can lead to systemic inflammation, oxidative stress, and metabolic dysregulation, which could influence lipid metabolism and result in changes in triglyceride levels.
According to our data, endometriosis leads to a significant increase in various fatty acids (such as C16:0, C18:0, C18:1, C18:2, C18:3, C20:3, C20:4) present in lipids like TG, PC, PE, and CL. Among these fatty acids, oleic acid C18:1 stands out as particularly prominent. This prevalent fatty acid, commonly found in adipocytes, likely plays a crucial role in fueling the energy needs of the pathologic endometrial cells as they proliferate and metastasize within the body. Interestingly, this lipid profile mirrors a similar pattern observed in other aggressive gynecological neoplasms, including cancer[63] In particular, accumulation of lipid drops, mainly composed of TGs, in tumour assosiated macrophages has been observed in a variety of cancers and is strongly associated with poor prognosis [64]. Other fatty acids with significant increase in endometriosis group are strongly linked with inflammation pathways [65]. Step-by-step convertion of linoleic acid (LA) C18:2 to gamma-linolenic (GLA) C18:3, then dihomo-γ-linolenic acid (DGLA) C20:3 and finally, arachidonic (AA) C20:4 acid, represents the vital fatty acid metabolism, resulting in a wide range of pro- and anti-inflammatory eicosanoids, such as prostaglandins and leukotrienes [66,67] . Triglycerides rich in arachidonate are actively taken up and stored in lipid droplets within a variety of cell types such as leukocytes, epithelial cells, and neoplastic cells. In pro-inflammatory enviroment, the lipid droplets may combine all the necessary enzymatic machinery responsible for producing eicosanoids derived from arachidonic acid [67,68]. Increased levels of prostaglandin E2 (PGE2) have been observed in ectopic endometrial tissue, peritoneal fluid, and serum of women with endometriosis, contributing to inflammation and pain associated with the disease [69,70]. Arachidonic acid-derived leukotrienes play a pivotal role in the recruitment and activation of immune cells [71], leading to chronic inflammation and tissue damage in endometriosis. Additionally, the dysregulation of enzymes involved in arachidonic acid metabolism, such as COX-2 and 5-lipoxygenase (5-LOX), has been reported in endometriotic lesions, further supporting the role of arachidonic acid-derived inflammatory mediators in the pathophysiology of endometriosis[72]. Oppositely, eicosanoids derived from DGLA, such as prostaglandin E1 (PGE1) and 15-hydroxyeicosatrienoic acid (15-HETrE), have been shown to exhibit anti-inflammatory properties [73]. These DGLA-derived eicosanoids can counteract the pro-inflammatory effects of arachidonic acid-derived eicosanoids in the body [74]. The balance between arachidonic acid-derived eicosanoids and DGLA-derived eicosanoids is crucial for maintaining optimal inflammatory responses in the body [75,76] Disruption of this balance, such as in conditions like endometriosis where arachidonic acid metabolism is dysregulated, can lead to chronic inflammation and disease progression.
In this study, cardiolipins were significantly elevated in menstrual blood of patient with endometriosis. Moreover, very-long-chain CL 16:0_18:0_22:5_22:6 (FC = 3.3, p=0.001) was included in the final logistic regression model for endometriosis diagnosis. Cardiolipins are a unique class of phospholipids found predominantly in the inner mitochondrial membrane, where they play essential roles in mitochondrial structure, function, and cellular metabolism [77]. While the focus on cardiolipins in endometriosis research is not as extensive as with other lipid classes like ceramides, there is growing interest in their potential implications in the pathogenesis of endometriosis [78]. Mitochondrial dysfunction has been implicated in endometriosis, and alterations in cardiolipin content or metabolism could contribute to this dysfunction [79,80,81]. Cardiolipins are crucial for maintaining mitochondrial membrane integrity, regulating energy production through oxidative phosphorylation, and influencing apoptotic pathways [82]. Dysregulation of cardiolipin levels or composition may lead to impaired mitochondrial function, increased oxidative stress, and altered apoptotic signaling, all of which are features associated with endometriosis. Moreover, cardiolipins can interact with immune cells and modulate inflammatory responses through the production of cytokines and reactive oxygen species.
Ceramides and sphingomyelins are identified as lipid species whose levels increase in menstrual blood in cases of endometriosis. The direction of alteration of Cer d18:0/24:0, Cer d18:1/16:0, Cer d18:1/24:1, SM d18:1/24:0 in our study aligns with the alteration of these lipids in peritoneal fluid as observed in Lee et al.'s study, although in endometrial tissue, these lipids exhibit the opposite direction of alteration [83]. Another study has also reported an association between higher concentrations of ceramides in peritoneal fluid and endometriosis progression [84]. Nevertheless, Dominguez et al. found decreased levels of ceramides in endometrial fluid in cases of ovarian endometriosis [38]. Ceramides, a class of sphingolipids, have been implicated in various physiological processes, including cell growth, differentiation, apoptosis, inflammation, and pain modulation [85,86]. In the context of endometriosis, ceramides have gained attention due to their potential role in the modulation of pain sensation [87,88]. Dysregulated ceramide metabolism has been linked to increased inflammation, oxidative stress, and aberrant cell signaling pathways, all of which are key features of endometriosis [84,89,90]. In particular, ceramides have been shown to promote the production of pro-inflammatory mediators and cytokines, exacerbating the inflammatory response in endometriotic lesions [91,92,93,94]. Moreover, ceramides have been implicated in the modulation of pain perception by activation of nociceptors, the sensitization of peripheral nerves, and the modulation of central pain processing [95,96]. By influencing neuronal excitability and neurotransmitter release, ceramides may contribute to the chronic pain experienced by individuals with endometriosis [97,98,99]. Furthermore, an inflammatory environment can trigger the growth and stimulation of nerve fibers, resulting in activation of pain signaling pathways [100].
The balance between sphingomyelins and ceramides, which are key components of cell membranes and signaling molecules, is critical for cellular function and homeostasis [101]. Sphingomyelins are synthesized from phosphatidylcholines and ceramides through the action of sphingomyelin synthase, primarily in the plasma and Golgi membranes of cells. On the other hand, sphingomyelinases catalyze the breakdown of sphingomyelins back into ceramides [102]. In endometriosis, the dysregulation of sphingolipid metabolism, including alterations in sphingomyelin synthesis and breakdown, may contribute to the observed increase in both ceramides and sphingomyelins.
Oxidized lipids (in particular, unsaturated PCs and CL) are potential markers of endometriosis, as well. Increase in lipid oxidation is a hallmark of endometriosis, related to oxidative stress and chronic inflammation [103,104,105,106,107]. In endometriosis, there is a distinct pattern of iron-dependent lipid peroxide buildup and increased resistance to ferroptosis, facilitated by the upregulation of redox enzymes like glutathione peroxidase 4 (GPx4) and superoxide dismutase (SOD) [108]. The presence of oxidized lipids in the peritoneal fluid and endometrial tissues has been reported in pathology [109,110,111,112,113,114,115,116,117]. These lipids can initiate inflammatory responses, activate immune cells, and promote the proliferation and survival of endometrial lesions [103]. Oxidized lipids can also interact with receptors on immune cells, such as toll-like receptors, leading to the production of pro-inflammatory cytokines and chemokines [118]. Additionally, oxidized lipids have been shown to disrupt cellular signaling pathways, promote oxidative stress, and impair the function of various cellular components, such as mitochondria and membranes [115,119]. These effects can impact cell survival, proliferation, and migration, which are critical processes in the pathogenesis of endometriosis.
5. Conclusions
The study unveiled notable variances in the lipid profile of dried menstrual blood spots, indicating its viability as a diagnostic instrument for endometriosis. The lipid biomarker-based model displayed encouraging levels of sensitivity and specificity for diagnosing endometriosis. Delving into the molecular composition of menstrual blood presents exciting possibilities for propelling research on pelvic diseases forward. Continued exploration into the biomarkers present in menstrual blood could potentially usher in enhanced diagnostic approaches and personalized treatment tactics for conditions such as endometriosis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary 1: Lipid profiles of samples; Figure S1 Supplementary2: Based ion chromatogram of dried blood spot lipids profile in A) positive ion mode, B) negative ion mode. CE – cholesterol ester, Cer – ceramide, CL – cardiolipine, LPC – lysophosphatidylcholine, LPE – lysophosphatidylethanolamine, PC – phosphatidylcholine, PC O- -plasmanylphosphatidylcholines, PC P- - plasmenylphosphatidylcholines, PE – phoshpatidylethanolamine, PE O- - plasmanylphosphatidylethanolamine, PE P- - plasmenylethanolamine, SM – sphingomyelin, TG – triacylglycerol, TG O- - plasmanyltriacylglycerol; Figure S2 Supplementary2: Average mass spectra of menstrual blood dried spot lipid profile. A) Positive ion mode. B) Negative ion mode; Figure S3 Supplementary2: Score plot of samples in space “predictive component of OPLS model – orthogonal component of OPLS model”; Table S1 Supplementary2: Statistically significant (FDR<0.05) enriched pathways in endometriosis.
Author Contributions
Conceptualization, V.F., S.P and N.S; Data curation, A.T., N.S., E.K. and V.C.; Formal analysis, S.P., E.K. and M.D.; Investigation, V.C., A.T., V.F., A.N. and N.S.; Methodology, V.C; S.P., V.F. and G.S.; Resources, G.S., S.P. and V.F.; Software, A.T., E.K. and V.C.; Supervision, N.S., S.P., V.F. and G.S.; Validation, N.S., V.F., A.N. and M.D..; Visualization, A.T., N.S., M.D. and E.K.; Project administration, V.F, S.P. and G.S.; Funding acquisition, V.F., S.P. and G.S.; Writing—original draft, N.S., A.T., V.C., M.D., A.N. and E.K.; Writing—review and editing, V.F., S.P. and G.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the state assignment of the Ministry of Healthcare of the Russian Federation (Registration No. 122020900125-8).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Biomedical Research Ethics Commission of the National Medical Research Center for Obstetrics Gynecology and Perinatology, named after academician V.I. Kulakov of the Ministry of Healthcare of Russian Federation.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are contained within the supplementary material (Supplementary 1.xlsx).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Simoens, S.; Dunselman, G.; Dirksen, C.; Hummelshoj, L.; Bokor, A.; Brandes, I.; Brodszky, V.; Canis, M.; Colombo, G.L.; Deleire, T.; et al. The burden of endometriosis: Costs and quality of life of women with endometriosis and treated in referral centres. Hum. Reprod. 2012, 27, 1292–1299. [Google Scholar] [CrossRef]
- Taylor, H.S.; Kotlyar, A.M.; Flores, V.A. Endometriosis is a chronic systemic disease: clinical challenges and novel innovations. Lancet 2021, 397, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Schliep, K.C.; Mumford, S.L.; Peterson, C.M.; Chen, Z.; Johnstone, E.B.; Sharp, H.T.; Stanford, J.B.; Hammoud, A.O.; Sun, L.; Buck Louis, G.M. Pain typology and incident endometriosis. Hum. Reprod. 2015, 30, 2427–2438. [Google Scholar] [CrossRef] [PubMed]
- Moss, K.M.; Doust, J.; Homer, H.; Rowlands, I.J.; Hockey, R.; Mishra, G.D. Delayed diagnosis of endometriosis disadvantages women in ART: A retrospective population linked data study. Hum. Reprod. 2021, 36, 3074–3082. [Google Scholar] [CrossRef] [PubMed]
- Giudice, L.C.; Oskotsky, T.T.; Falako, S.; Opoku-Anane, J.; Sirota, M. Endometriosis in the era of precision medicine and impact on sexual and reproductive health across the lifespan and in diverse populations. FASEB J. 2023, 37, e23130. [Google Scholar] [CrossRef] [PubMed]
- de Almeida Filho, D.P.; de Oliveira, L.J.; do Amaral, V.F. Accuracy of laparoscopy for assessing patients with endometriosis. Sao Paulo Med. J. 2008, 126, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.P.; Hummelshoj, L. Consensus on current management of endometriosis. Hum. Reprod. 2013, 28, 1552–1568. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Hull, M.; Fraser, I.; Miller, L.; Bossuyt, P.; Johnson, N.; Nisenblat, V. Endometrial biomarkers for the non-invasive diagnosis of endometriosis (Review). Cochrane Database Syst. Rev. 2016, 2016, CD012165. [Google Scholar] [CrossRef] [PubMed]
- Ametzazurra, A.; Matorras, R.; García-Velasco, J.A.; Prieto, B.; Simón, L.; Martínez, A.; Nagore, D. Endometrial fluid is a specific and non-invasive biological sample for protein biomarker identification in endometriosis. Hum. Reprod. 2009, 24, 954–965. [Google Scholar] [CrossRef]
- Nisenblat, V.; Bossuyt, P.M.M.; Shaikh, R.; Farquhar, C.; Jordan, V.; Scheffers, C.S.; Mol, B.W.J.; Johnson, N.; Hull, M.L. Blood biomarkers for the non-invasive diagnosis of endometriosis. Cochrane Database Syst. Rev. 2016, 2016, CD012179. [Google Scholar] [CrossRef]
- Bozorgmehr, M.; Gurung, S.; Darzi, S.; Nikoo, S.; Kazemnejad, S.; Zarnani, A.H.; Gargett, C.E. Endometrial and Menstrual Blood Mesenchymal Stem/Stromal Cells: Biological Properties and Clinical Application. Front. Cell Dev. Biol. 2020, 8, 497. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, K.; Bourdon, M.; Bartkowski, R.; Verbanck, M.; Chapron, C.; Marcellin, L.; Batteux, F.; Santulli, P.; Doridot, L. Menstrual Blood Donation for Endometriosis Research: A Cross-Sectional Survey on Women’s Willingness and Potential Barriers. Reprod. Sci. 2024. [Google Scholar] [CrossRef] [PubMed]
- Naseri, S.; Young, S.; Cruz, G.; Blumenthal, P.D. Screening for High-Risk Human Papillomavirus Using Passive, Self-Collected Menstrual Blood. Obstet. Gynecol. 2022, 140, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhou, B.; Prinz, M.; Siegel, D. Proteomic analysis of menstrual blood. Mol. Cell. Proteomics 2012, 11, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; Liu, Y.; Yan, L.; Zhang, Y.; Li, Y.; Zhu, Q.; Xia, W.; Ge, S.; Zhang, J. DIA-based analysis of the menstrual blood proteome identifies association between CXCL5 and IL1RN and endometriosis. J. Proteomics 2023, 289, 104995. [Google Scholar] [CrossRef] [PubMed]
- Samare-Najaf, M.; Razavinasab, S.A.; Samareh, A.; Jamali, N. Omics-based novel strategies in the diagnosis of endometriosis. Crit. Rev. Clin. Lab. Sci. 2023, 25, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Fiscus, J.; Fraison, É.; Renault, L.; Salle, B.; Panthu, B.; Labrune, E. Metabolic signature of follicular fluid to understand infertility-related diseases: a narrative review. Reprod. Biomed. Online 2023, 48, 103762. [Google Scholar] [CrossRef] [PubMed]
- Angioni, S.; Congiu, F.; Vitale, S.G.; D’Alterio, M.N.; Noto, A.; Monni, G.; Santoru, M.L.; Fanos, V.; Murgia, F.; Atzori, L. Gas Chromatography–Mass Spectrometry (GC–MS) Metabolites Analysis in Endometriosis Patients: A Prospective Observational Translational Study. J. Clin. Med. 2023, 12, 922. [Google Scholar] [CrossRef] [PubMed]
- Tomkins, N.E.; Girling, J.E.; Boughton, B.; Holdsworth-Carson, S.J. Is there a role for small molecule metabolite biomarkers in the development of a diagnostic test for endometriosis? Syst. Biol. Reprod. Med. 2022, 68, 89–112. [Google Scholar] [CrossRef]
- Yang, H.; Lau, W.B.; Lau, B.; Xuan, Y.; Zhou, S.; Zhao, L.; Luo, Z.; Lin, Q.; Ren, N.; Zhao, X.; et al. A mass spectrometric insight into the origins of bening gynecological sudorder. Mass Spectrom. Rev. 2017, 36, 450–470. [Google Scholar] [CrossRef]
- Tokarz, J.; Adamski, J.; Rižner, T.L. Metabolomics for diagnosis and prognosis of uterine diseases? A systematic review. J. Pers. Med. 2020, 10, 294. [Google Scholar] [CrossRef] [PubMed]
- Morales-Romero, B.; González de Aledo-Castillo, J.M.; Fernández Sierra, C.; Martínez Carreira, C.; Zaragoza Bonet, C.; Fernández Bonifacio, R.; Caro Miró, M.A.; Argudo-Ramírez, A.; López Galera, R.M.; García-Villoria, J. Plasma C24:0- and C26:0-lysophosphatidylcholines are reliable biomarkers for the diagnosis of peroxisomal β-oxidation disorders. J. Lipid Res. 2024, 65, 100516. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, J.; Jiang, Y.; Ju, Y.; He, J.; Yu, K.; Kan, G.; Zhang, H. Determination of amino acid metabolic diseases from dried blood spots with a rapid extraction method coupled with nanoelectrospray ionization mass spectrometry. Talanta 2024, 272, 125768. [Google Scholar] [CrossRef]
- Koelmel, J.P.; Kroeger, N.M.; Ulmer, C.Z.; Bowden, J.A.; Patterson, R.E.; Cochran, J.A.; Beecher, C.W.W.; Garrett, T.J.; Yost, R.A. LipidMatch: An automated workflow for rule-based lipid identification using untargeted high-resolution tandem mass spectrometry data. BMC Bioinformatics 2017, 18, 331. [Google Scholar] [CrossRef] [PubMed]
- Tokareva, A.O.; Chagovets, V. V.; Kononikhin, A.S.; Starodubtseva, N.L.; Nikolaev, E.N.; Frankevich, V.E. Comparison of the effectiveness of variable selection method for creating a diagnostic panel of biomarkers for mass spectrometric lipidome analysis. J. Mass Spectrom. 2021, 56, e4702. [Google Scholar] [CrossRef] [PubMed]
- Starodubtseva, N.L.; Tokareva, A.O.; Rodionov, V. V; Brzhozovskiy, A.G.; Bugrova, A.E.; Chagovets, V. V; Kometova, V. V; Kukaev, E.N.; Soares, N.C.; Kovalev, G.I.; et al. Integrating Proteomics and Lipidomics for Evaluating the Risk of Breast Cancer Progression : A Pilot Study. Biomedicines 2023, 11, 1786. [Google Scholar] [CrossRef]
- Kamburov, A.; Stelzl, U.; Lehrach, H.; Herwig, R. The ConsensusPathDB interaction database: 2013 Update. Nucleic Acids Res. 2013, 41, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E.; Yilmaz, B.D.; Sison, C.; Miyazaki, K.; Bernardi, L.; Liu, S.; Kohlmeier, A.; Yin, P.; Milad, M.; Wei, J.J. Endometriosis. Endocr. Rev. 2019, 40, 1048–1079. [Google Scholar] [CrossRef]
- Tanbo, T.; Fedorcsak, P. Endometriosis-associated infertility: aspects of pathophysiological mechanisms and treatment options. Acta Obstet. Gynecol. Scand. 2017, 96, 659–667. [Google Scholar] [CrossRef]
- Becker, C.M.; Bokor, A.; Heikinheimo, O.; Horne, A.; Jansen, F.; Kiesel, L.; King, K.; Kvaskoff, M.; Nap, A.; Petersen, K. ESHRE PAGES The members of the Endometriosis Guideline Core Group, Hum. Reprod. 2022, 2022, hoac009. [Google Scholar] [CrossRef]
- Han, Z.; Li, J.; Yi, X.; Zhang, T.; Liao, D.; You, J.; Ai, J. Diagnostic accuracy of interleukin-6 in multiple diseases: An umbrella review of meta-analyses. Heliyon 2024, 10, e27769. [Google Scholar] [CrossRef] [PubMed]
- Brulport, A.; Bourdon, M.; Vaiman, D.; Drouet, C.; Pocate-Cheriet, K.; Bouzid, K.; Marcellin, L.; Santulli, P.; Abo, C.; Jeljeli, M.; et al. An integrated multi-tissue approach for endometriosis candidate biomarkers: a systematic review. Reprod. Biol. Endocrinol. 2024, 22, 21. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.A.; Lamichhane, S.; Dickens, A.; McGlinchey, A.; Ribeiro, H.C.; Sen, P.; Wei, F.; Hyötyläinen, T.; Orešič, M. Systems biology approaches to study lipidomes in health and disease. Biochim. Biophys. Acta - Mol. Cell Biol. Lipids 2021, 1866, 158857. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Zhang, L.; Yan, F.; Wang, X. Clinical lipidomics: a new way to diagnose human diseases. Clin. Transl. Med. 2018, 7, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, C.N.; Torres-Reverón, A.; Appleyard, C.B. Metabolomics in endometriosis: challenges and perspectives for future studies. Reprod. Fertil. 2021, 2, R35–R50. [Google Scholar] [CrossRef] [PubMed]
- Braga, D.P.A.F.; Montani, D.A.; Setti, A.S.; Turco, E.G.L.; Oliveira-Silva, D.; Borges, E. Metabolomic profile as a noninvasive adjunct tool for the diagnosis of Grades III and IV endometriosis-related infertility. Mol. Reprod. Dev. 2019, 86, 1044–1052. [Google Scholar] [CrossRef]
- Li, J.; Gao, Y.; Guan, L.; Zhang, H.; Sun, J.; Gong, X.; Li, D.; Chen, P.; Ma, Z.; Liang, X.; et al. Discovery of phosphatidic acid, phosphatidylcholine, and phosphatidylserine as biomarkers for early diagnosis of endometriosis. Front. Physiol. 2018, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, F.; Ferrando, M.; Díaz-Gimeno, P.; Quintana, F.; Fernández, G.; Castells, I.; Simón, C. Lipidomic profiling of endometrial fluid in women with ovarian endometriosis. Biol. Reprod. 2017, 96, 772–779. [Google Scholar] [CrossRef]
- Sasamoto, N.; Zeleznik, O.A.; Vitonis, A.F.; Missmer, S.A.; Laufer, M.R.; Avila-Pacheco, J.; Clish, C.B.; Terry, K.L. Presurgical blood metabolites and risk of postsurgical pelvic pain in young patients with endometriosis. Fertil. Steril. 2022, 117, 1235–1245. [Google Scholar] [CrossRef]
- Starodubtseva, N.; Chagovets, V.; Borisova, A.; Salimova, D.; Aleksandrova, N.; Chingin, K.; Chen, H.; Frankevich, V. Identification of potential endometriosis biomarkers in peritoneal fluid and blood plasma via shotgun lipidomics. Clin. Mass Spectrom. 2019, 13, 21–26. [Google Scholar] [CrossRef]
- Andrieu, T.; Chicca, A.; Pellegata, D.; Bersinger, N.A.; Imboden, S.; Nirgianakis, K.; Gertsch, J.; Mueller, M.D. Association of endocannabinoids with pain in endometriosis. Pain 2022, 163, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Warren, L.A.; Shih, A.; Renteira, S.M.; Seckin, T.; Blau, B.; Simpfendorfer, K.; Lee, A.; Metz, C.N.; Gregersen, P.K. Analysis of menstrual effluent: Diagnostic potential for endometriosis. Mol. Med. 2018, 24, 1. [Google Scholar] [CrossRef] [PubMed]
- Shih, A.J.; Adelson, R.P.; Vashistha, H.; Khalili, H.; Nayyar, A.; Puran, R.; Herrera, R.; Chatterjee, P.K.; Lee, A.T.; Truskinovsky, A.M.; et al. Single-cell analysis of menstrual endometrial tissues defines phenotypes associated with endometriosis. BMC Med. 2022, 20, 315. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Qu, J.; Xiang, C. The multi-functional roles of menstrual blood-derived stem cells in regenerative medicine. Stem Cell Res. Ther. 2019, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Zailani, N.N.B.; Ho, P.C.L. Dried Blood Spots—A Platform for Therapeutic Drug Monitoring (TDM) and Drug/Disease Response Monitoring (DRM). Eur. J. Drug Metab. Pharmacokinet. 2023, 48, 467–494. [Google Scholar] [CrossRef] [PubMed]
- Sakhi, A.K.; Bastani, N.E.; Ellingjord-Dale, M.; Gundersen, T.E.; Blomhoff, R.; Ursin, G. Feasibility of self-sampled dried blood spot and saliva samples sent by mail in a population-based study. BMC Cancer 2015, 15, 265. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.J.; Mangwiro, Y.; Wake, M.; Saffery, R.; Greaves, R.F. Multi-omics analysis from archival neonatal dried blood spots: Limitations and opportunities. Clin. Chem. Lab. Med. 2022, 60, 1318–1341. [Google Scholar] [CrossRef] [PubMed]
- Malsagova, K.; Kopylov, A.; Stepanov, A.; Butkova, T.; Izotov, A.; Kaysheva, A. Dried blood spot in laboratory: Directions and prospects. Diagnostics 2020, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Chagovets, V. V.; Wang, Z.; Kononikhin, A.S.; Starodubtseva, N.L.; Borisova, A.; Salimova, D.; Popov, I.A.; Kozachenko, A. V.; Chingin, K.; Chen, H.; et al. Endometriosis foci differentiation by rapid lipid profiling using tissue spray ionization and high resolution mass spectrometry. Sci. Rep. 2017, 7, 2546. [Google Scholar] [CrossRef]
- Vouk, K.; Hevir, N.; Ribič-Pucelj, M.; Haarpaintner, G.; Scherb, H.; Osredkar, J.; Möller, G.; Prehn, C.; Rižner, T.L.; Adamski, J. Discovery of phosphatidylcholines and sphingomyelins as biomarkers for ovarian endometriosis. Hum. Reprod. 2012, 27, 2955–2965. [Google Scholar] [CrossRef]
- Vouk, K.; Ribič-Pucelj, M.; Adamski, J.; Rižner, T.L. Altered levels of acylcarnitines, phosphatidylcholines, and sphingomyelins in peritoneal fluid from ovarian endometriosis patients. J. Steroid Biochem. Mol. Biol. 2016, 159, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Bozelli, J.C.; Azher, S.; Epand, R.M. Plasmalogens and Chronic Inflammatory Diseases. Front. Physiol. 2021, 12, 730829. [Google Scholar] [CrossRef]
- Zoeller, R.A.; Lake, A.C.; Nagan, N.; Gaposchkin, D.P.; Legner, M.A.; Lieberthal, W. Plasmalogens as endogenous antioxidants: Somatic cell mutants reveal the importance of the vinyl ether. Biochem. J. 1999, 338, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Broniec, A.; Klosinski, R.; Pawlak, A.; Wrona-Krol, M.; Thompson, D.; Sarna, T. Interaction of plasmalogens and their diacyl analogs with singlet oxygen in selected model systems. Free Radic. Biol. Med. 2011, 50, 892–898. [Google Scholar] [CrossRef]
- Feider, C.L.; Woody, S.; Ledet, S.; Zhang, J.; Sebastian, K.; Breen, M.T.; Eberlin, L.S. Molecular Imaging of Endometriosis Tissues using Desorption Electrospray Ionization Mass Spectrometry. Sci. Rep. 2019, 9, 15690. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, Y.; Zhang, L.; Zhang, L. Association between endometriosis and metabolic syndrome: a cross-sectional study based on the National Health and Nutrition Examination Survey data. Gynecol. Endocrinol. 2023, 39, 2254844. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.; Rich-Edwards, J.; Rimm, E.B.; Spiegelman, D.; Forman, J.P.; Missmer, S.A. Association between Endometriosis and Hypercholesterolemia or Hypertension. Hypertension 2017, 70, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Kirichenko, T. V.; Markina, Y. V.; Bogatyreva, A.I.; Tolstik, T. V.; Varaeva, Y.R.; Starodubova, A. V. The Role of Adipokines in Inflammatory Mechanisms of Obesity. Int. J. Mol. Sci. 2022, 23, 14982. [Google Scholar] [CrossRef]
- Fahed, G.; Aoun, L.; Zerdan, M.B.; Allam, S.; Zerdan, M.B.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Pathophysiology and Management in 2021. Int. J. Mol. Sci. 2022, 23, 786. [Google Scholar] [CrossRef]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuñiga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef]
- Farland, L. V.; Degnan, W.J.; Harris, H.R.; Tobias, D.K.; Missmer, S.A. A prospective study of endometriosis and risk of type 2 diabetes. Diabetologia 2021, 64, 552–560. [Google Scholar] [CrossRef]
- Sekulovski, N.; Whorton, A.E.; Shi, M.; Hayashi, K.; MacLean, J.A. Insulin signaling is an essential regulator of endometrial proliferation and implantation in mice. FASEB J. 2021, 35, e21440. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Kong, W.; Suo, H.; Shen, X.; Newton, M.A.; Burkett, W.C.; Zhao, Z.; John, C.; Sun, W.; Zhang, X.; et al. Oleic Acid Exhibits Anti-Proliferative and Anti-Invasive Activities via the PTEN/AKT/mTOR Pathway in Endometrial Cancer. Cancers (Basel). 2023, 15, 5407. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Wang, Q.; Bi, E.; Ma, X.; Liu, L.; Yang, M.; Qian, J.; Yi, Q. Enhanced Lipid Accumulation and Metabolism Are Required for the Differentiation and Activation of Tumor-Associated Macrophages. Cancer Res. 2020, 80, 1438–1450. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiù, V.; Leuti, A.; Maccarrone, M. Bioactive lipids and chronic inflammation: Managing the fire within. Front. Immunol. 2018, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Stables, M.J.; Gilroy, D.W. Old and new generation lipid mediators in acute inflammation and resolution. Prog. Lipid Res. 2011, 50, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Bozza, P.T.; Viola, J.P.B. Lipid droplets in inflammation and cancer. Prostaglandins Leukot. Essent. Fat. Acids 2010, 82, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Rakhila, H.; Bourcier, N.; Akoum, A.; Pouliot, M. Abnormal Expression of Prostaglandins E2 and F2 α Receptors and Transporters in Patients with Endometriosis. Biomed Res. Int. 2015, 2015, 808146. [Google Scholar] [CrossRef]
- Mandelboum, S.; Manber, Z.; Elroy-Stein, O.; Elkon, R. Recurrent functional misinterpretation of RNA-seq data caused by sample-specific gene length bias. PLoS Biol. 2019, 17, e3000481. [Google Scholar] [CrossRef]
- Von Moltke, J.; Trinidad, N.J.; Moayeri, M.; Kintzer, A.F.; Wang, S.B.; Van Rooijen, N.; Brown, C.R.; Krantz, B.A.; Leppla, S.H.; Gronert, K.; et al. Rapid induction of inflammatory lipid mediators by the inflammasome in vivo. Nature 2012, 490, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.Z.; Yang, H.L.; Ha, S.Y.; Chang, K.K.; Mei, J.; Zhou, W.J.; Qiu, X.M.; Wang, X.Q.; Zhu, R.; Li, D.J.; et al. Cyclooxygenase-2 in endometriosis. Int. J. Biol. Sci. 2019, 15, 2783–2797. [Google Scholar] [CrossRef]
- Levin, G.; Duffin, K.L.; Obukowicz, M.G.; Hummert, S.L.; Fujiwara, H.; Needleman, P.; Raz, A. Differential metabolism of dihomo-γ-linolenic acid and arachidonic acid by cyclo-oxygenase-1 and cyclo-oxygenase-2: Implications for cellular synthesis of prostaglandin E1 and prostaglandin E2. Biochem. J. 2002, 365, 489–496. [Google Scholar] [CrossRef]
- Wang, X.; Lin, H.; Gu, Y. Multiple roles of dihomo-γ-linolenic acid against proliferation diseases. Lipids Health Dis. 2012, 11, 25. [Google Scholar] [CrossRef]
- Mustonen, A.M.; Nieminen, P. Dihomo-γ-Linolenic Acid (20:3n-6)—Metabolism, Derivatives, and Potential Significance in Chronic Inflammation. Int. J. Mol. Sci. 2023, 24, 2116. [Google Scholar] [CrossRef] [PubMed]
- Sergeant, S.; Rahbar, E.; Chilton, F.H. Gamma-linolenic acid, Dihommo-gamma linolenic, Eicosanoids and Inflammatory Processes. Eur. J. Pharmacol. 2016, 785, 77–86. [Google Scholar] [CrossRef]
- Ren, M.; Phoon, C.K.L.; Schlame, M. Metabolism and function of mitochondrial cardiolipin. Prog. Lipid Res. 2014, 55, 1–16. [Google Scholar] [CrossRef]
- May, K.E.; Conduit-Hulbert, S.A.; Villar, J.; Kirtley, S.; Kennedy, S.H.; Becker, C.M. Peripheral biomarkers of endometriosis: A systematic review. Hum. Reprod. Update 2010, 16, 651–674. [Google Scholar] [CrossRef]
- Ye, C.; Chen, P.; Xu, B.; Jin, Y.; Pan, Y.; Wu, T.; Du, Y.; Mao, J.; Wu, R. Abnormal expression of fission and fusion genes and the morphology of mitochondria in eutopic and ectopic endometrium. Eur. J. Med. Res. 2023, 28, 209. [Google Scholar] [CrossRef]
- Li, H.; Yang, H.; Lu, S.; Wang, X.; Shi, X.; Mao, P. Autophagy-dependent ferroptosis is involved in the development of endometriosis. Gynecol. Endocrinol. 2023, 39, 2242962. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, X.; Guo, J.; Wang, D.; Li, X.; Cheng, X.; Wang, X. CHCHD2 Regulates Mitochondrial Function and Apoptosis of Ectopic Endometrial Stromal Cells in the Pathogenesis of Endometriosis. Reprod. Sci. 2022, 29, 2152–2164. [Google Scholar] [CrossRef] [PubMed]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Role of cardiolipin in mitochondrial function and dynamics in health and disease: Molecular and pharmacological aspects. Cells 2019, 8, 728. [Google Scholar] [CrossRef]
- Lee, Y.H.; Tan, C.W.; Venkatratnam, A.; Tan, C.S.; Cui, L.; Loh, S.F.; Griffith, L.; Tannenbaum, S.R.; Chan, J.K.Y. Dysregulated sphingolipid metabolism in endometriosis. J. Clin. Endocrinol. Metab. 2014, 99, E1913–E1921. [Google Scholar] [CrossRef]
- Lee, Y.H.; Yang, J.X.; Allen, J.C.; Tan, C.S.; Chern, B.S.M.; Tan, T.Y.; Tan, H.H.; Mattar, C.N.Z.; Chan, J.K.Y. Elevated peritoneal fluid ceramides in human endometriosis-associated infertility and their effects on mouse oocyte maturation. Fertil. Steril. 2018, 110, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, G.; Wang, L.; Meng, L.; Ren, J.; Shang, L.; Li, D.; Bao, Y. Dysregulation of sphingolipid metabolism in pain. Front. Pharmacol. 2024, 15, 1337150. [Google Scholar] [CrossRef]
- Gonzalez, P.A.; Simcox, J.; Raff, H.; Wade, G.; Von Bank, H.; Weisman, S.; Hainsworth, K. Lipid signatures of chronic pain in female adolescents with and without obesity. Lipids Health Dis. 2022, 21, 80. [Google Scholar] [CrossRef]
- Chrobak, A.; Sieradzka, U.; Sozański, R.; Chełmońska-Soyta, A.; Gabryś, M.; Jerzak, M. Ectopic and eutopic stromal endometriotic cells have a damaged ceramide signaling pathway to apoptosis. Fertil. Steril. 2009, 92, 1834–1843. [Google Scholar] [CrossRef]
- Santulli, P.; Marcellin, L.; Noël, J.C.; Borghese, B.; Fayt, I.; Vaiman, D.; Chapron, C.; Méhats, C. Sphingosine pathway deregulation in endometriotic tissues. Fertil. Steril. 2012, 97, 904–911e5. [Google Scholar] [CrossRef]
- Lee, M.; Lee, S.Y.; Bae, Y.S. Functional roles of sphingolipids in immunity and their implication in disease. Exp. Mol. Med. 2023, 55, 1110–1130. [Google Scholar] [CrossRef] [PubMed]
- Palareti, G.; Legnani, C.; Cosmi, B.; Antonucci, E.; Erba, N.; Poli, D.; Testa, S.; Tosetto, A. Comparison between different D-Dimer cutoff values to assess the individual risk of recurrent venous thromboembolism: Analysis of results obtained in the DULCIS study. Int. J. Lab. Hematol. 2016, 38, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, Y.L.; Li, Y.K.; Chen, D.K.; He, J.F.; Yao, N. Functions of Sphingolipids in Pathogenesis During Host–Pathogen Interactions. Front. Microbiol. 2021, 12, 701041. [Google Scholar] [CrossRef]
- MacEyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef]
- Malan, T.P.; Porreca, F. Lipid mediators regulating pain sensitivity. Prostaglandins Other Lipid Mediat. 2005, 77, 123–130. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Yamashita, A.; Matsuda, M.; Kawai, K.; Sawa, T.; Amaya, F. NLRP2 inflammasome in dorsal root ganglion as a novel molecular platform that produces inflammatory pain hypersensitivity. Pain 2019, 160, 2149–2160. [Google Scholar] [CrossRef]
- Ji, R.R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and central sensitization in chronic and widespread pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef]
- Kajimoto, T.; Okada, T.; Yu, H.; Goparaju, S.K.; Jahangeer, S.; Nakamura, S. Involvement of Sphingosine-1-Phosphate in Glutamate Secretion in Hippocampal Neurons. Mol. Cell. Biol. 2007, 27, 3429–3440. [Google Scholar] [CrossRef]
- Weth-Malsch, D.; Langeslag, M.; Beroukas, D.; Zangrandi, L.; Kastenberger, I.; Quarta, S.; Malsch, P.; Kalpachidou, T.; Schwarzer, C.; Proia, R.L.; et al. Ablation of sphingosine 1-phosphate receptor subtype 3 impairs hippocampal neuron excitability in vitro and spatial working memory in vivo. Front. Cell. Neurosci. 2016, 10, 258. [Google Scholar] [CrossRef] [PubMed]
- Morotti, M.; Vincent, K.; Brawn, J.; Zondervan, K.T.; Becker, C.M. Peripheral changes in endometriosis-associated pain. Hum. Reprod. Update 2014, 20, 717–736. [Google Scholar] [CrossRef]
- Jamjoum, R.; Majumder, S.; Issleny, B.; Stiban, J. Mysterious sphingolipids: metabolic interrelationships at the center of pathophysiology. Front. Physiol. 2023, 14, 1229108. [Google Scholar] [CrossRef] [PubMed]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.F.P.; Samadder, A.N.; Agarwal, A.; Fernandes, L.F.C.; Abrão, M.S. Oxidative stress biomarkers in patients with endometriosis: Systematic review. Arch. Gynecol. Obstet. 2012, 286, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Mier-Cabrera, J.; Jiménez-Zamudio, L.; García-Latorre, E.; Cruz-Orozco, O.; Hernández-Guerrero, C. Quantitative and qualitative peritoneal immune profiles, T-cell apoptosis and oxidative stress-associated characteristics in women with minimal and mild endometriosis. BJOG An Int. J. Obstet. Gynaecol. 2011, 118, 6–16. [Google Scholar] [CrossRef]
- Thézénas, M.L.; De Leo, B.; Laux-Biehlmann, A.; Bafligil, C.; Elger, B.; Tapmeier, T.; Morten, K.; Rahmioglu, N.; Dakin, S.G.; Charles, P.; et al. Amine oxidase 3 is a novel pro-inflammatory marker of oxidative stress in peritoneal endometriosis lesions. Sci. Rep. 2020, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Scutiero, G.; Iannone, P.; Bernardi, G.; Bonaccorsi, G.; Spadaro, S.; Volta, C.A.; Greco, P.; Nappi, L. Oxidative Stress and Endometriosis: A Systematic Review of the Literature. Oxid. Med. Cell. Longev. 2017, 2017, 7265238. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Yoshimoto, C.; Matsubara, S.; Shigetomi, H.; Imanaka, S. Current Understanding of and Future Directions for Endometriosis-Related Infertility Research with a Focus on Ferroptosis. Diagnostics 2023, 13, 1926. [Google Scholar] [CrossRef] [PubMed]
- Ota, H.; Igarashi, S.; Kato, N.; Tanaka, T. Aberrant expression of glutathione peroxidase in eutopic and ectopic endometrium in endometriosis and adenomyosis. Fertil. Steril. 2000, 74, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Polak, G.; Barczyński, B.; Kwaśniewski, W.; Bednarek, W.; Wertel, I.; Derewianka-Polak, M.; Kotarski, J. Low-density lipoproteins oxidation and endometriosis. Mediators Inflamm. 2013, 2013, 12–15. [Google Scholar] [CrossRef]
- Murphy, A.A.; Santanam, N.; Parthasarathy, S. Endometriosis: A disease of oxidative stress? Semin. Reprod. Endocrinol. 1998, 16, 263–273. [Google Scholar] [CrossRef]
- Nasiri, N.; Moini, A.; Eftekhari-Yazdi, P.; Karimian, L.; Salman-Yazdi, R.; Arabipoor, A. Oxidative Stress Statues in Serum and Follicular Fluid of Women with Endometriosis Citation: Nasiri N, Moini A, Eftekhari-Yazdi P, Karimian L, Salman-Yazdi R, Arabipoor A. Oxidative stress statues in serum and follicular fluid of women with endometriosis. CELL JOURNAL(Yakhteh) Cell Journal(Yakhteh) Cell J 2017, 18, 582–587. [Google Scholar]
- Murphy, A.A.; Palinski, W.; Rankin, S.; Morales, A.J.; Parthasarathy, S. Macrophage scavenger receptor(s) and oxidatively modified proteins in endometriosis. Fertil. Steril. 1998, 69, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.A.; Palinski, W.; Rankin, S.; Morales, A.J.; Parthasarathy, S. Evidence for oxidatively modified lipid-protein complexes in endometrium and endometriosis. Fertil. Steril. 1998, 69, 1092–1094. [Google Scholar] [CrossRef] [PubMed]
- Szczepańska, M.; Koźlik, J.; Skrzypczak, J.; Mikołajczyk, M. Oxidative stress may be a piece in the endometriosis puzzle. Fertil. Steril. 2003, 79, 1288–1293. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.H.; Huang, H.C.; Hsieh, R.H.; Chen, S.C.; Tsai, M.C.; Tzeng, C.R. Oxidative damage and mitochondrial DNA mutations with endometriosis. Ann. N. Y. Acad. Sci. 2005, 1042, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Mier-Cabrera, J.; Genera-García, M.; De la Jara-Díaz, J.; Perichart-Perera, O.; Vadillo-Ortega, F.; Hernández-Guerrero, C. Effect of vitamins C and E supplementation on peripheral oxidative stress markers and pregnancy rate in women with endometriosis. Int. J. Gynecol. Obstet. 2008, 100, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.; Mandai, M.; Toyokuni, S.; Hamanishi, J.; Higuchi, T.; Takakura, K.; Fujii, S. Contents of endometriotic cysts, especially the high concentration of free iron, are a possible cause of carcinogenesis in the cysts through the iron-induced persistent oxidative stress. Clin. Cancer Res. 2008, 14, 32–40. [Google Scholar] [CrossRef]
- Kajihara, H.; Yamada, Y.; Kanayama, S.; Furukawa, N.; Noguchi, T.; Haruta, S.; Yoshida, S.; Sado, T.; Oi, H.; Kobayashi, H. New insights into the pathophysiology of endometriosis: From chronic inflammation to danger signal. Gynecol. Endocrinol. 2011, 27, 73–79. [Google Scholar] [CrossRef]
- Clower, L.; Fleshman, T.; Geldenhuys, W.J.; Santanam, N. Targeting Oxidative Stress Involved in Endometriosis and Its Pain. Biomolecules 2022, 12, 1055. [Google Scholar] [CrossRef]
Figure 1.
A volcano plot of lipids detected in the dried menstrual blood spots. Orange color marks lipids with a notable statistically significant differences between groups studied (p-value < 0.05, Mann-Whitney test) and a median fold change greater than 1.5 are labeled. Cer-NS – ceramide , Cer-NP – physphingosine ceramide, CL – cardiolipine, oxCL – oxidized cardiolipins, PC – phosphatidylcholine, PC O- -plasmanylphosphatidylcholines, PC P- - plasmenylphosphatidylcholines, oxPC – oxidized phosphatidylcholines, PE P- - plasmenylethanolamine, TG – triacylglycerol.
Figure 1.
A volcano plot of lipids detected in the dried menstrual blood spots. Orange color marks lipids with a notable statistically significant differences between groups studied (p-value < 0.05, Mann-Whitney test) and a median fold change greater than 1.5 are labeled. Cer-NS – ceramide , Cer-NP – physphingosine ceramide, CL – cardiolipine, oxCL – oxidized cardiolipins, PC – phosphatidylcholine, PC O- -plasmanylphosphatidylcholines, PC P- - plasmenylphosphatidylcholines, oxPC – oxidized phosphatidylcholines, PE P- - plasmenylethanolamine, TG – triacylglycerol.

Figure 2.
Base peak chromatogram (black), extracted ion chromatogram of PE P-16:0/18:1 (red) and extracted ion chromatogram of CL 16:0_18:0_22:5_22:6 (blue) with tandem mass spectra. Extracted ion chromatogram of CL 16:0_18:0_22:5_22:6 is scaled in five times. Negative ion mode.
Figure 2.
Base peak chromatogram (black), extracted ion chromatogram of PE P-16:0/18:1 (red) and extracted ion chromatogram of CL 16:0_18:0_22:5_22:6 (blue) with tandem mass spectra. Extracted ion chromatogram of CL 16:0_18:0_22:5_22:6 is scaled in five times. Negative ion mode.
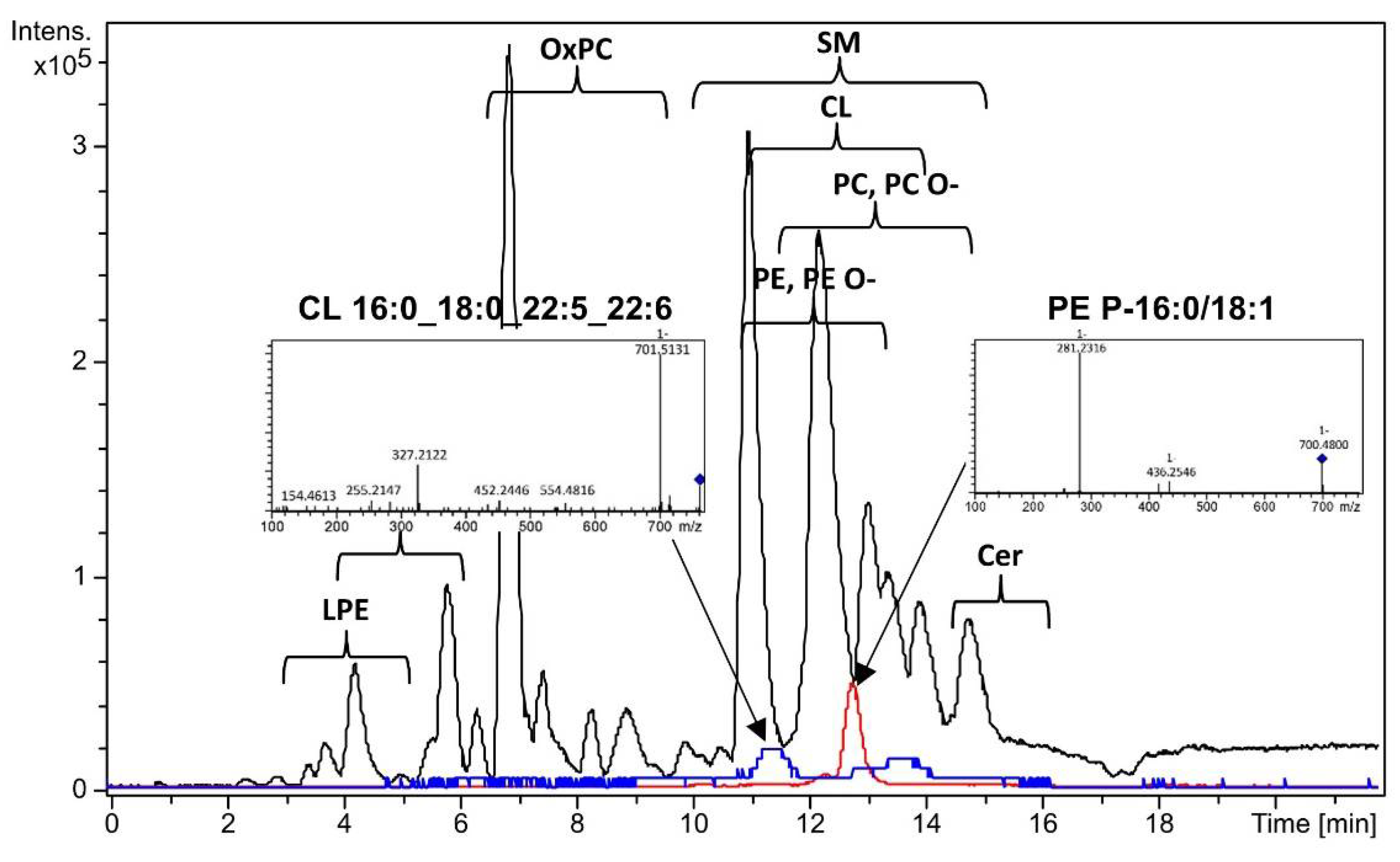
Figure 3.
Receiver operating characteristic curve obtained after cross-validation for logistic regression model developed (1). Coefficients in endometriosis diagnostic model, based on logic regression are presented underneath.
Figure 3.
Receiver operating characteristic curve obtained after cross-validation for logistic regression model developed (1). Coefficients in endometriosis diagnostic model, based on logic regression are presented underneath.
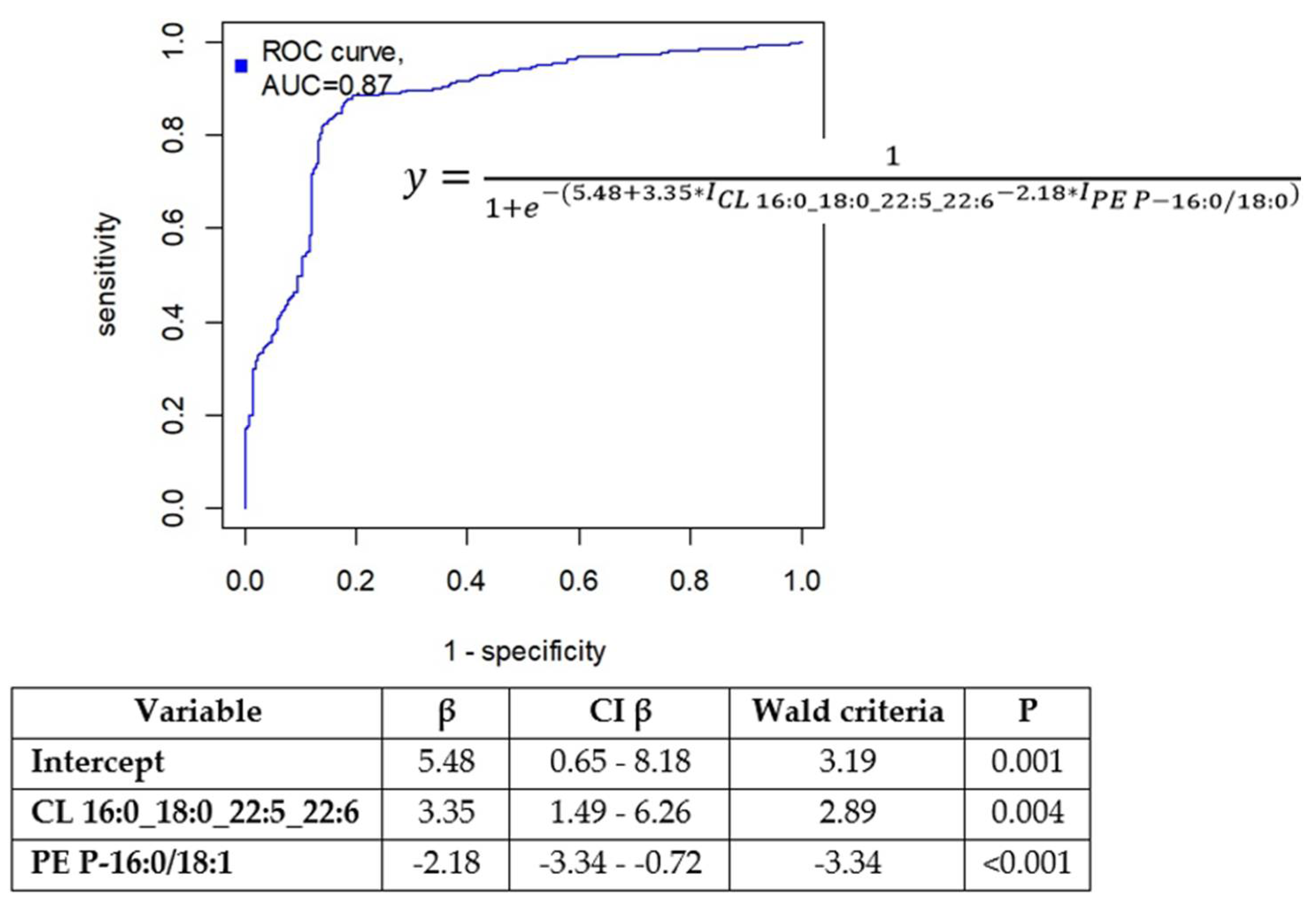
Figure 4.
Diagnostic potential of PE P-16:0/18:1 and CL 16:0_18:0_22:5_22:6: A) Boxplots illustrating the levels of these markers in menstrual blood; B) Dried menstrual blood spots plotted in the space of the relative proportion of PE P-16:0/18:1 and CL 16:0_18:0_22:5_22:6. The green points represent samples from the control group patients, while the ocher points represent samples from the endometriosis group.
Figure 4.
Diagnostic potential of PE P-16:0/18:1 and CL 16:0_18:0_22:5_22:6: A) Boxplots illustrating the levels of these markers in menstrual blood; B) Dried menstrual blood spots plotted in the space of the relative proportion of PE P-16:0/18:1 and CL 16:0_18:0_22:5_22:6. The green points represent samples from the control group patients, while the ocher points represent samples from the endometriosis group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



