
Submitted:
20 May 2024
Posted:
21 May 2024
You are already at the latest version
Abstract
Sesame seeds are abundant in sesamin, which exerts health-promoting effects such as extending the lifespan of adult Drosophila and suppression of oxidative stress accumulation by activating the Nrf2 transcription factor. Here, we investigated whether sesamin activated Nrf2 in larval tissues and induced the expression of Nrf2 target genes. In sesamin-fed larvae, Nrf2 was activated in the central nervous system (CNS), gut, and salivary glands. However, ectopic expression of Keap1 (Nrf2 inhibitor) in glial cells inhibited sesamin-induced Nrf2 activation in all tissues except in the neurons, indicating that sesamin activates Nrf2 in glia. We labeled the astrocytes as well as cortex and surface glia with red fluorescence to identify these glial cell types in which Nrf2 was activated; we observed their activation in both cell types. These data suggest that sesamin may stimulate the expression of antioxidative genes in glial cells. Among the 17 candidate Nrf2 targets, the mRNA levels of Cyp6a2 and Cyp6g1 in cytochrome P450 were elevated in the CNS, gut, and salivary glands of sesamin-fed larvae. However, this did not lead to resistance against imidacloprid, which is detoxified by these enzymes. Our results suggest that sesamin may exert similar health-promoting effects on the human CNS and digestive tissues.
Keywords:
Sesamin
; Nrf2
; glial cells
; cytochrome P450
; Drosophila
1. Introduction
Sesame has been consumed since ancient times as a highly nutritious food. Sesame seeds are rich in various lignans including sesamin, sesaminol, and sesamol [1]. However, sesamin has been particularly studied for its potential as a supplement owing to its health-promoting effects [2,3,4]. A previous study has demonstrated that sesamin prolonged the lifespan of adult Drosophila by inhibiting and delaying the aging phenotype of muscles, midgut stem cells, and neurons [5]. Additionally, it mitigated age-dependent loss of dopaminergic neurons due to the accumulation of oxidative stress in the adult brain, suggesting that the antioxidant effect of sesamin is closely linked to its anti-aging effects [5,6]. Living organisms are constantly exposed to oxidative stress that is generated both inside and outside the body. Although moderate oxidative stress has beneficial effects such as the activation of the immune system and other organs, excessive oxidative stress damages lipids, proteins, and the DNA [7,8]. Furthermore, oxidative stress is closely linked to the pathogenesis of diseases such as cancer and diabetes [9]. Organisms possess antioxidant systems that produce antioxidant enzymes and antioxidants [10,11]. One of the most critical antioxidant systems is the Keap1–Nrf2 system for transcriptional regulation. The transcription factor Nf-E2-related factor (Nrf2) plays an indispensable role in maintaining redox homeostasis in the body. Nrf2 target genes include those encoding enzymes involved in oxidative stress removal, detoxification, metabolism, and other crucial metabolic pathways [12]. Under non-oxidative stress, Nrf2 forms a complex with the repressor Kelch-like ECH-associated protein 1 (Keap1), which is immediately ubiquitinated and degraded by proteasomes. In contrast, under oxidative stress, the SH group of Keap1 is oxidized, eliminating the interaction between Nrf2 and Keap1. This dissociates the inhibitor and stabilizes Nrf2, which migrates to the nucleus. There it binds to a consensus sequence called the antioxidant response element (ARE), which promotes the transcription of target genes [13,14,15,16,17]. An experimental system was established to monitor Nrf2/Cnc activation in the Drosophila body by creating an ARE–GFP reporter with a GFP gene downstream of this sequence [18]. Generally, Keap1–Nrf2 binding is disrupted by reactive oxygen species (ROS) generated during metabolic processes or by external environmental stimuli such as UV light or some chemicals. The disruption activates Nrf2, which promotes the transcription of antioxidant genes and protects the cells from oxidative stress [19,20,21,22,23]. Thus, the Keap1–Nrf2 system protects organisms against endogenous and exogenous oxidative stress. In mice, Nrf2 expression in the liver decreases with age [24]. Additionally, Nrf2 deficiency increases oxidative stress and promotes aging [25]. Therefore, identifying effective anti-oxidation and aging substances in the diet is crucial for preventing oxidative damage-associated diseases and delaying the aging progression.
Although several in vitro and in vivo studies have demonstrated the antioxidant effect of sesamin, direct evidence indicating its anti-aging effect in mammals, including humans, is yet to be obtained [26,27,28,29,30]. In contrast, Drosophila studies reported that the antioxidant effect of sesamin is mediated by Nrf2 activation [5,6]. Oxidative damage to intracellular biomaterials by endogenous reactive oxygen species has been proposed as a factor in the aging process [31]. The results of the Drosophila studies provide important insights into the mechanism underlying the anti-oxidative and anti-aging effects exerted by sesamin. In this study, we investigated whether sesamin activates Nrf2 in the central nervous system (CNS) and digestion-related tissues during larval development, as observed in adults. ARE–GFP reporter is useful for this investigation because it enables the study of Nrf2 activation with high sensitivity [18].
Neurons and glial cells are more involved in age-related neurodegenerative diseases [32]. Although sesamin exerts inhibitory and retardative effects on the aging of the nervous system, few studies have examined its effects on glial cells in the CNS using model organisms including Drosophila. Mice and rats are the most commonly used animal models in brain research. Insect models such as Drosophila are also being increasingly used because of their usefulness owing to the high similarity to the nervous system of mammals. Although the number of neurons in the Drosophila CNS is 1,000–10,000 times lesser than that in mammals [33], the neurons, glutamate, dopamine, serotonin, acetylcholine, GABA, and other neurotransmitters are used in the CNS for neural activity as in humans. Additionally, several genes and regulators involved in CNS development and function are shared between humans and Drosophila [34]. Drug metabolism, mechanisms of action, and the effects on neural activity and behavior in Drosophila are similar to those observed in mammals [35].
In addition to its anti-oxidative effect in the CNS of Drosophila, the consumption of sesamin suppresses the oxidative damage-dependent phenotype that appears in the gut of Drosophila adults [5]. The Drosophila gut is responsible for the digestion and absorption of nutrients. It gradually becomes less functional with age owing to an activated inflammatory response and metabolic decline [36]. Moreover, metabolic enzymes such as CYP450 are abundantly expressed in intestinal epithelial cells. In Drosophila adults, sesamin is also metabolized to its intermediate metabolite SC1 [5], which is absorbed and metabolized in intestinal epithelial cells similar to that observed in mammals. Sesamin and its metabolic products may alter gene expression throughout the body. Furthermore, the structure and function of the gastrointestinal tract exhibit several other similarities between humans and Drosophila. Therefore, Drosophila is a valuable experimental animal for studying the effects of chemicals on the CNS and digestion-related tissues.
In this study, based on findings regarding the effects of sesamin in Drosophila adults, we investigated whether sesamin induces the activation of the transcription factor Nrf2 in larvae. We also investigated the subsequent antioxidant responses in the CNS and digestive tissues during larval development. Sesamin-fed larvae showed Nrf2 activation in the CNS, salivary glands, and gut. Next, we investigated whether the neurons or glial cells within the CNS of sesamin-fed larvae presented stronger Nrf2 activation. Consequently, we found that sesamin activated Nrf2 in glial cells rather than in neurons in the larval CNS. This study discusses how Nrf2 activation in glial cells influences CNS development. Next, we examined the endogenous genes whose transcription was altered by sesamin. We found that the mRNA levels of several drug-metabolizing enzymes were increased in the CNS, gut, and salivary glands of the sesamin-fed larvae. Since upregulation of these enzymes has been reported in Drosophila mutant strains that are resistant to the neurotransmitter-blocking insecticide, we further investigated whether sesamin influences sensitivity to the drug. As its antioxidant and anti-aging effects were observed in adults, sesamin mediates antioxidant and detoxifying effects via Nrf2 activation during larval development. These results suggest that sesamin may exert similar effects on the human CNS and digestive tissues. This is an important finding with regard to the health benefits of sesamin.
2. Materials and Methods
2.1. Fly Stocks and Culture
To monitor ARE-dependent transcription, the ARE-GFP reporter (ARE-GFP), in which the cDNA for GFP was placed after ARE sequences on which the Nrf2/Cnc transcription factor binds, was used. A stock harboring the ARE-GFP was a gift from D. Bohmann (University of Rochester Medical Center, Rochester, New York, USA). P{EPgy2}Keap1[EY02632] (UAS-Keap1) (#15427) was used for the overexpression of Keap1. The Gal4/UAS system was used for ectopic gene expression in Drosophila. The following fly stocks were obtained from the Bloomington Drosophila Stock Center (Bloomington, IN, USA). To visualize all neurons in larval CNSs, we used P{GawB}elavC155; P{tubP-GAL80ts}10 (elav-Gal4) (#67058). To visualize specific types of glial cells in larval brains, the following Gal4-drivers were used: P{GAL4}repo (repo-Gal4) (#7415) for expression in all glial cells, P{Eaat1-GAL4.R}2 (Eaat1-Gal4) (#8849) for expression in astrocytes, and P{nrv2-GAL4.S}3; P{nrv2-GAL4.S}8 (Nrv2-Gal4) (#6797) for expression in cortex and surface glial cells. The P{UAS-tdTom.S}3 (UAS-RFP) (#36328) stock was used in combination with the appropriate Gal4 driver for labeling the neurons and the glial cells by expression of Red Fluorescence Protein (RFP).
A standard cornmeal diet containing 7.2 g agar, 100 g glucose, 40 g dried yeast, and 40 g cornmeal per liter was used as fly food. All ingredients were mixed well and boiled for 10 min, as described previously [37]. After cooling to 75 ℃, 5 mL of 10% parahydroxybenzonate dissolved in ethanol and 5 mL of propionic acid were added as anti-septic agents to the diet all fly stocks were maintained on a regular cornmeal diet at 25 ℃, except for the overexpression experiments performed at 28 ℃.
2.2. Chemical Feeding
For sesamin feeding, early third instar larvae were collected and separated into males and females. The larvae were reared in a single plastic vial containing the Drosophila instant medium (Formula 4-24® Instant Drosophila Medium, Blue; Carolina Biological Supply Company, Burlington, NC, USA) at a 0.3g/ml concentration. Sesamin (Nacalai Tesque, Kyoto, Japan) dissolved in DMSO solution (Dimethyl sulfoxide; Wako Pure Chemical Industries, Ltd., Osaka, Japan) was added to final concentrations of sesamin 1 mg/mL, DMSO 1% in the instant medium. As a control diet, the DMSO 1% alone was added to the instant medium. For the imidacloprid-feeding experiment, adults were mated in a standard cornmeal diet supplemented with 1mg/ml sesamin and 1% DMSO or the diet with 1% DMSO alone as a control, were removed after the egg-laying for 8h. 72 h later, larvae were collected and reared on a standard cornmeal diet containing imidacloprid (Tokyo chemical industry, Tokyo, Japan) 2 µM and sesamin 1 mg/ml with DMSO 1%. The individuals were kept at 25 °C.
2.3. Observation of GFP and RFP Fluorescence
To visualize GFP fluorescence in the whole body, the ARE-GFP larvae at the early third instar were collected and fed the instant medium supplemented sesamin 1 mg/mL and DMSO 1% for 24 h, while the medium with 1% DMSO alone was used as a control diet. The GFP fluorescence was observed under a stereo fluorescence microscope (SZX7; Olympus, Tokyo, Japan). To visualize GFP and/or RFP fluorescence in the CNSs, guts, and salivary glands, third-instar larvae harboring the ARE-GFP were collected and fed sesamin 1 mg/mL and DMSO 1% for 24 h. To visualize neurons or glial cells in larval CNSs, the flies carrying pan-neuronal, neuron-specific, pan-glial, or glial-specific Gal4 drivers were crossed with those with UAS-RFP. Third instar larvae were collected and raised for 24 h on diets supplemented with sesamin 1 mg/mL and DMSO 1%. The CNS, gut, or salivary gland was collected from the larvae as described previously [5,38]. The whole CNSs, gut, or salivary glands were fixed in 4% paraformaldehyde for 30 min, washed with PBST (phosphate-buffered saline containing 0.1% Triton X-100), and subsequently blocked with 10% normal goat serum for 30 min. All samples were mounted in Vectashield (Vector Laboratories, Burlingame, CA, USA). The GFP and/or RFP fluorescence was observed using an Olympus laser scanning confocal microscope (Fv10i; Olympus, Tokyo, Japan) or a stereo fluorescence microscope (SZX7; OLYMPUS, Tokyo, Japan). The brightness and contrast of the entire images were adjusted using the Fv10i software.
2.4. Survival Rate of Larvae Fed the Diet Supplemented with Drugs
After the egg-laying of adults for 8h, the hatched larvae were reared on a standard cornmeal diet supplemented with sesamin 1mg/ml and DMSO 1%. The the late 2nd-instar larvae were collected and transferred to a standard cornmeal diet with imidacloprid and 1mg/ml sesamin with 1% DMSO. The number of pupae was counted, and the survival rate from larvae to pupa was calculated.
2.5. Quantitative Reverse Transcription Polymerase Chain Reaction
For quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis, total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) from the CNSs, gut, salivary glands of larvae fed diets with or without sesamin 1mg/mL for 24 h. cDNA synthesis was performed using a PrimeScript II High Fidelity RT-PCR kit (Takara, Shiga, Japan) with oligo dT primers. qRT-PCR was performed using TB Green Premix Ex Taq Ⅱ (Takara, Shiga, Japan) and Thermal Cycler Dice® Real Time System Ⅲ (Takara, Shiga, Japan). RP49 was used as the normalization reference. Relative mRNA levels were quantified using Thermal Cycler Dice® Real Time System Ⅲ version 6.0.1 (Takara, Shiga, Japan). The primers used were as follows:
RP49-Fw, 5’-TTCCTGGTGCACAACGTG-3’,
RP49-Rv, 5’-TCTCCTTGCGCTTCTTGG-3’,
GFP-Fw, 5’-AAGCTGACCCTGAAGTTCATCTGC-3’,
GFP-Rv, 5’-CTTGTAGTTGCCGTCGTCCTTGAA-3’,
Cyp6a2-Fw, 5’- TTCACCACCGATGTGATTGGC-3’,
Cyp6a2-Rv, 5’- TCGGGCATCATGCGCATT-3’,
Trx2-Fw, 5’-ATGGACAGCTGACCAAGGCATC-3’,
Trx2-Rv, 5’-CCCACTTAGATATTGGCCTTGATG-3’,
GPx-Fw, 5’-GGTCGATGTGAATGGAGACA,
GPx-Rv, 5’-CCCTCCTTGTTCACCAGAAA,
Sod3-Fw, 5’-AGCTGGAGGGATTGAAGGAG-3’,
Sod3-Rv, 5’-GGGGCCACCGTGATCAAC-3’,
Gclm-Fw, 5’-AGGATTCCAACGTCAGCAGG-3’,
Gclm-Rv, 5’-AATCTGCTGCTTGAGGGCAT-3’
Nqo1-Fw, 5’-TTTCTCCTGGTGCCATTACC-3’,
Nqo1-Rv, 5’-CTGCAGCAGGATAGGTCCTC-3’,
TrxR1-Fw, 5’- CGTTCTATTGTGCTGCGTGG-3’,
TrxR1-Rv, 5’- AGCTTGCCATCATCCTGCTT-3’,
Catalase-Fw, 5’-TTTCTCCTGGTGCCATTACC-3’,
Catalase-Rv, 5’-CTGCAGCAGGATAGGTCCTC-3’
dGB-Fw, 5’-CCATGAGGGGTATGATCAGTG-3’,
dGB-Rv, 5’-ATTTATGTGCTGGCCAATGTG-3’,
Aldh1-Fw, 5’- TCCGAGGGAGATAAGGCTGA- 3’,
Aldh1-Rv, 5’- GAATGCCTTGTCCCGATCCA -3’,
Jafrac1-Fw, 5’-ACCGAGATCATTGCGTTCTC-3’,
Jafrac1-Rv, 5’-AAGTGGGTGAACTGGCTGTC-3’,
Gclc-Fw, 5’-ATGACGAGGAGAATGAGCTG-3’,
Gclc-Rv, 5’-CCATGGACTGCAAATAGCTG-3’,
Pgd-Fw, 5’-GGAATGTGTGAACGGGAAAGTGGAG-3’,
Pgd-Rv, 5’-AGGACTCGTGGCGCGAGGTG-3’,
Adh-Fw, 5’-AAACTGGCCCCCATTACCG-3’,
Adh-Rv, 5’-CAAGTCCAGTTTCCAGATG-3’,
HO-Fw, 5’-ACCATTTGCCCGCCGGGATG-3’,
HO-Rv, 5’-AGTGCGACGGCCAGCTTCCT-3’,
Zw-Fw, 5’-AAGCGCCGCAACTCTTTG-3’,
Zw-Rv, 5’-AGGGCGGTGTGATCTTCC-3’,
p62-Fw, 5’-CGTAAGGACCTTCTGGATCG-3’,
p62-Rv, 5’- CGTCGTGGATGGTGAAATTG -3’,
Cyp6g1-Fw, 5’- GCCCGCTGCGATCCCCAT-3’,
Cyp6g1-Rv, 5’- CCT TTCCAATCTCCTGCATA-3’,
All qRT-PCR experiments were performed in triplicate, and an average of three replicates in each group were considered. The ∆∆Ct method was used to determine the differences in target gene expression relative to the reference Rp49 gene expression.
2.6. Quantitation of GFP or RFP-Positive Area in CNS
To measure the intensity of GFP fluorescence and the GFP-positive area in the larval whole body or CNSs in larvae, fluorescence images were acquired using a stereo fluorescence microscope, and the microscopic images were analyzed using the Image J software (National Institutes of Health, Bethesda, MD, USA). The GFP intensity or fluorescence area in the larval CNSs was measured using the Fv10i software and quantified using the Image J software.
2.7. Statistical Analysis
Statistical analyses were performed using GraphPad Prism (Version 9, GraphPad Software, San Diego, CA, USA). The Student’s t-test, Welch’s t-test, or Pearson’s chi-square test was used to compare the two groups. p-values less than 0.05 were judged to be statistically significant.
3. Results
3.1. Sesamin Consumption Activated Nrf2/Cnc in Specific Tissues of Drosophila Larvae
Based on a previous claim that sesamin-induced Nrf2/Cnc transcription factor activation produced antioxidant effect in Drosophila adults [6], we investigated whether Nrf2/Cnc was activated in sesamin-fed larvae using an ARE–GFP reporter. GFP fluorescence was not observed in 0.5%, 1% and 2% DMSO-fed larvae for 24h (Figure 1A,c’,g,’e’,i’,k’,m’). The most intense GFP fluorescence was observed in the ARE–GFP expressing larvae that were fed 2 mg/ml of sesamin in 2% DMSO for 24 h (Figure 1A,d’,f’,h,’j‘,l’,n’). GFP fluorescence was observed in the larval head region, indicating that sesamin strongly activated Nrf2/Cnc in the larval tissues. GFP fluorescence was quantified by measuring the GFP-positive regions in whole larval body images captured using a stereo-fluorescence microscope (Figure 1B). The positive area increased as DMSO concentration increased. The largest positive area was observed in the larvae that were fed 2 mg/ml of sesamin in 2% DMSO. The second largest area was observed in the larvae that were fed 1 mg/ml of sesamin in 1% DMSO. In subsequent experiments, we used either feeding condition for sesamin administration.
3.2. Sesamin-Consumption Activated Nrf2/Cnc in the CNS and Digestion-related Tissues of Drosophila Larvae
We identified the tissues showing intense GFP fluorescence in the head region of ARE-GFP-expressing larvae after sesamin consumption by examining the presence of sesamin-induced GFP reporter expression in the larval CNS. The late second instar larvae were fed a diet of 1 or 2 mg/ml of sesamin in DMSO, and DMSO alone as control for 24 h. The CNS from the third instar larvae were observed for GFP fluorescence using confocal microscopy (Figure 2A). GFP fluorescence in the CNS was barely detected in the 1% DMSO-fed larvae (Figure 2A,a–c,f,i,l), whereas it was relatively higher in the 2% DMSO-fed larvae (Figure 2B). The concentration of DMSO in subsequent experiments was set at 1% because the diet containing 2% DMSO elicited a relatively higher GFP fluorescence intensity in the CNS than that of the 1% DMSO-containing diet. In contrast, the fluorescence intensity in the CNS of sesamin-fed larvae increased significantly compared with that of the controls. The highest increase in GFP fluorescence intensity was observed in 2 mg/mL sesamin (2% DMSO)-fed larvae. However, the GFP fluorescence intensity decreased in the CNS of larvae fed 2 mg/mL of sesamin under 1% DMSO conditions compared to larvae fed 1 mg/mL of sesamin under the same conditions. Fluorescence intensity fluctuated less in females compared to males. Based on the results of these preliminary experiments, sesamin dissolved in 1% DMSO was mixed with feed to a concentration of 1 mg/mL and fed to the third-instar larvae in subsequent experiments. Hereafter, this diet is referred to as the sesamin-containing diet. Larvae in the control group were fed a diet containing only 1% DMSO. Hereafter, this diet is referred to as the control diet. Since the GFP fluorescence difference among the individuals between males and females, the female third instar larvae, which showed less difference in fluorescence between individuals, were used in subsequent experiments. Next, GFP mRNA levels of the ARE–GFP reporter were quantified in the CNS of sesamin-containing diet-fed larvae using qRT-PCR (Figure 2C). The third instar larval females that consumed a sesamin-containing diet for 24 h showed an average 39.2-fold increase in GFP mRNA level compared with that in control diet-fed larvae.
To identify other tissues that show intense GFP reporter expression in the head of sesamin-fed larvae, we examined whether sesamin induced GFP reporter expression in the salivary gland and larval gut, both of which are located in the larval head along with the CNS. After consuming the sesamin-containing diet for 24 h, intense GFP fluorescence was observed, particularly in the salivary glands. Stronger GFP fluorescence was observed in the anterior midgut and was relatively less intense in the hindgut. Quantifying the GFP mRNA levels using qRT-PCR (three independent trials for each condition) showed an average 98.9-fold and 4,896.6-fold increase in the mRNA levels in the gut and salivary glands, respectively, of sesamin-fed larvae compared with that of the control. The GFP fluorescence intensity indicates the intensity of Nrf2 activation. Thus, these results indicate that sesamin activates Nrf2 in larval CNSs, salivary glands, and gut.
3.3. Activation of Nrf2/Cnc in Glial Cells of the Larval CNS Following Sesamin Consumption
As sesamin induced Nrf2/Cnc activation in the Drosophila CNS, we examined whether sesamin activated Nrf2 in the neurons and/or glial cells in the CNS. We labeled all neurons in the CNS based on ectopic RFP expression (elav-Gal4/+; ARE-GFP/+; UAS-RFP/+) and examined ARE–GFP reporter expression in the RFP fluorescent areas. The fluorescence was observed throughout the larval brain and ventral ganglia in the CNS of sesamin-fed larvae, whereas only faint GFP fluorescence was observed in the CNS of non-sesamin fed larvae. This GFP-positive region overlapped considerably with the distribution of the neurons. However, intense Nrf2/Cnc activation was also observed in areas with few neurons (Figure 3A). In contrast, when all glial cells were labeled with specifically expressing RFP and their distribution in the CNS was observed (ARE-GFP/+; repo-Gal4/UAS-RFP), the ARE–GFP fluorescence was found to approximately coincide with the RFP-positive region showing ARE-GFP-related GFP fluorescence after sesamin consumption (Figure 3B). This suggests that strong Nrf2 activation by sesamin occured primarily in glial cells.
Thus, we confirmed that sesamin activated Nrf2 in glial cells rather than in neurons. We induced the ectopic expression of the Nrf2 inhibitor Keap1 in all neurons and glial cells in the CNS and examined whether ARE–GFP fluorescence was lost or reduced in the sesamin-fed larvae. Keap1 expression was induced in all neurons (elav-Gal4/+; ARE-GFP/+; UAS-Keap1EY/+) in the CNS of the larvae that were fed sesamin for 24 h. In non-feeding controls, GFP fluorescence was not observed, confirming the absence of Nrf2 activation (Figure 3C,a–d,D). Although distinctive GFP fluorescence was observed in the CNS of sesamin-fed larvae (Figure 3C,e,D), the induced expression of Keap1 in all neurons little reduced ARE–GFP fluorescence (Figure 3C,f,D). In contrast, the intense GFP fluorescence (Figure 3C,g,D) was no longer observed (Figure 3C,h,D) in the sesamin-fed larvae that overexpressed Keap1 in all glial cells of the CNS (ARE-GFP/+; repo-Gal4/UAS-Keap1EY). These results suggest that Nrf2 activation in the larval CNS by sesamin is primarily induced in glial cells rather than in neurons.
3.4. Sesamin Activated Nrf2 in the Astrocytes, Cortex, and Surface Glia
As previously mentioned, sesamin activates the Nrf2 transcription factor in glial cells within the larval CNS. Next, we investigated the types of glial cells in which Nrf2 is activated. Using the Gal4/UAS system, which allows ectopic gene expression, we performed astrocyte (Figure 4A,a,b, Eaat1-Gal4 /ARE-GFP; UAS-RFP/+)- or cortex- and surface glia (Figure 4D,a,b, Nrv2-Gal4/ARE-GFP; UAS-RFP/+)-specific RFP labeling of the glial cells within the CNS,. After being fed sesamin for 24 h, we examined whether the RFP fluorescence areas indicating each type of glial cell overlapped with those of Nrf2-activated cells in the CNS. Stronger GFP fluorescence was observed in the CNS of the sesamin-fed larvae (Figure 4A,b’) than in the controls (Figure 4a’). GFP fluorescence intensity in each glial cell region was quantified on a 0–255 scale. The average fluorescence intensity in the sesamin-fed larvae was 72.0 (57.4% increase) compared with 45.7 in the controls (Figure 4B). Similarly, in the cortex and surface glial regions, the average fluorescence intensity in the sesamin-fed group was 86.2 (73.2% increase, Figure 4D,b’,E) compared with 49.8 in the controls (Figure 4D,a’,E). As the fluorescence detected in the CNS of the controls was weak, we measured the size of the positive areas that expressed each glia marker at a stronger GFP fluorescence intensity than the average fluorescence intensity of the controls (without sesamin consumption). The lower limit of the fluorescence threshold was set as the average fluorescence intensity of the controls (threshold, 46–255 for astrocytes; 50–255 for cortex and surface glia). We found that 23.8% of astrocytes in the sesamin-fed larvae emitted stronger GFP fluorescence than that of the control, which is a 16.2% increase in fluorescence expressing area compared with the 7.6% of the controls (Figure 4C). Among the cortex and surface glial cells of the sesamin-fed larvae, 31.1% of fluorescence emitting regions was detected, which was a 28.2% increase compared with the 2.9% in the controls (Figure 4F). These results indicate that sesamin consumption activates Nrf2 in astrocytes as well as cortex and surface glial cells.
3.5. Sesamin-Consumption in Larvae Elevated the mRNA Levels of Several Genes Encoding Enzymes in Cytochrome P450
Using the ARE–GFP reporter, we showed that sesamin consumption induced a pronounced upregulation of Nrf2-dependent transcription in the CNS, salivary glands, and gut. As this GFP reporter has an artificial regulatory region harboring up to four consecutive ARE sequences aligned in tandem before the GFP cDNA, it causes GFP to be excessively expressed in response to Nrf2 activation. To quantify the upregulation of the Nrf2-targeted endogenous genes, we performed quantitative PCR using RNA prepared from the CNS of sesamin-fed larvae. The mRNA levels of the GFP gene transcribed from ARE–GFP increased 40-fold on average than that in the control larvae reared on a diet without sesamin (Figure 2C).
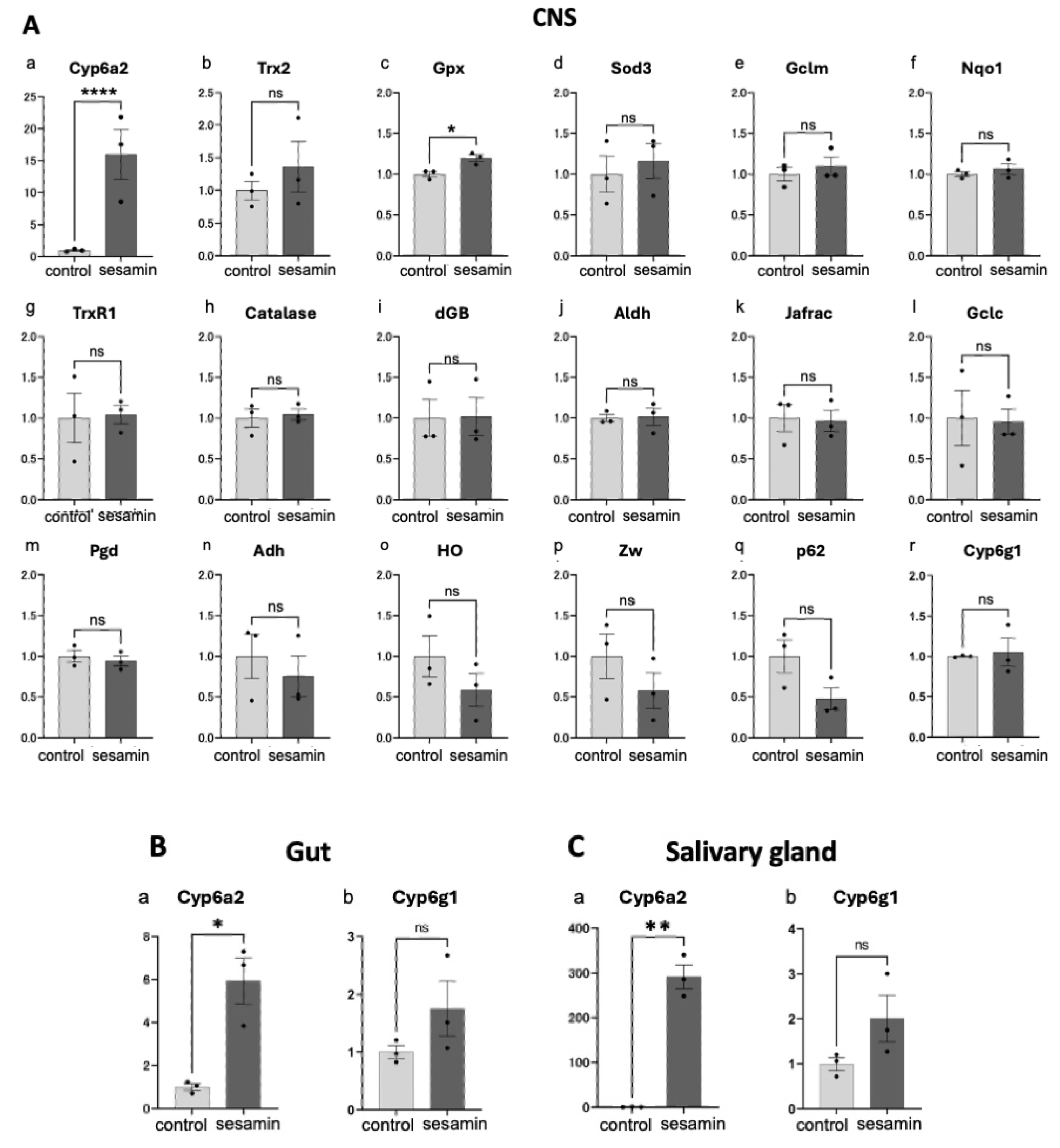
Next, we attempted to identify the endogenous genes whose transcription was induced by sesamin. Only a few Nrf2/Cnc target genes have been identified in Drosophila. Among the known Nrf2 target genes in mammals, we selected 17 genes whose orthologues were conserved in Drosophila as candidates for Nrf2 targets [39]. qRT-PCR was performed to quantify the target mRNA levels in the CNS of the third instar larvae (40 larvae/experiment) that were fed diets supplemented with sesamin or only 1% DMSO (control) for 24 h using synthesized cDNAs as templates (Figure 5A). Among these 17 genes, the mRNA level of the gene encoding Cyp6a2 (Figure 5A,a), which is a member of the cytochrome p450 family, increased 16.0-fold in the CNS of sesamin-fed larvae compared with than of the controls. Among the remaining 16 candidate Nrf2 target genes, mRNA levels of Trx2 (Figure 5A,b), Gpx (Figure 5A,c) and Sod3 (Figure 5A,d) were slightly increased than that of the controls (36.0%, 19.6%, and 16.0% increase, respectively). The mRNA levels of other nine genes, namely, Gclm (Figure 5A,e), Nqo1 (Figure 5A,f), TrxR1 (Figure 5A,g), Catalase (Figure 5Ah), dGB (Figure 5A,i), Aldh (Figure 5A,j), Jafrac1 (Figure 5A,k), Gclc (Figure 5A,l), Pgd (Figure 5A,m), and Cy6g1 (Figure 5A,r) remained essentially unchanged after feeding (<10% change in each case). In contrast, the mRNA levels of Adh (Figure 5A,n), HO (Figure 5A,o), Zw (Figure 5A,p), and p62 (Figure 5A,q) rather reduced (24.7%, 41.6%, 42.4%, and 52.5% reduction, respectively).
As sesamin significantly increased Cyp6a2 mRNA levels in the larval CNS, we investigated whether its mRNA levels were altered in other larval tissues involved in absorption and metabolism, such as the gut and salivary gland. Cyp6a2 mRNA levels in the gut and salivary glands of the sesamin-fed larvae showed significant 5.9-fold (Figure 5B,a) and 291.7-fold (Figure 5C,a) increase than that of the controls. Drosophila Cyp6a2 is an ortholog of human cytochrome P450 family 3 subfamily A member 43 (Cyp3a4), which encodes an enzyme involved in insecticide metabolism. Whether this gene is a Nrf2 target is unknown; nevertheless, we investigated whether sesamin consumption increased its expression. Consequently, the mRNA levels of this gene in gut and salivary gland of sesamin-fed larvae increased 1.8-fold (Figure 5B,b) and 2.0-fold (Figure 5C,b) than that in the controls. In summary, two genes, Cyp6a2 and Cyp6g1, which encode cytochrome P450 enzymes, were upregulated in tissues related to the Drosophila larval digestive system after sesamin consumption.
3.6. Resistance to the Neonicotinoid Insecticide Imidacloprid Was not Observed in Sesamin-Fed Larvae
The mRNA levels of the cytochrome P450 drug-metabolizing enzyme Cyp6a2 increased notably in the CNS, gut, and salivary glands of the sesamin-fed larvae. A similar drug-metabolizing enzyme, Cyp6g1, was also induced in the gut and salivary glands of larvae. Cyp6g1 is involved in the metabolism of neonicotinoids that have toxic effects on neuronal connections in Drosophila [40], and we predicted that accelerated metabolism of this chemical would eventually lead to its rapid degradation in organisms, potentially inducing drug resistance. Therefore, we investigated whether larvae reared on sesamin exhibit resistance to the neonicotinoid insecticide—imidacloprid. First, we determined the suitable concentration of imidacloprid for the study by examining the pupation rates. Drosophila larvae failed to pupate or were killed at the larval stage (survival rate 30%) at insecticide concentrations in the range of 3–100 µM. The pupation rate was 100% (16 out of 16 larvae) for the standard diet without the drug. In contrast, the pupation rates were 31.1% (14 of 45 larvae) for 3 µM, 31.0% (13 of 42 larvae) for 10 µM, 28.6% (10 of 35 larvae) for 30 µM, and 0% (0 out of 19 larvae) for 100 µM of the drug. An imidacloprid concentration of 3 µM was selected for the subsequent experiments as the corresponding survival rate was approximately 30%. To induce Cyp6a2 and Cyp6g1 expression in advance, the standard cornmeal food supplemented with 1 mg/mL of sesamin and 1% DMSO was fed to the first instar larvae before drug administration. Parent flies were placed in vials with the sesamin-containing or control diets for 8 h and allowed to lay eggs. After removing the parent flies, the hatched larvae were reared on the same diet for 72 h. The second instar larvae (50 larvae) were transferred to a diet containing 3 µM of imidacloprid and 1 mg/mL of sesamin with 1% DMSO (the control diet contained 3 µM of imidacloprid and 1% DMSO) from the second instar stage until they pupated. We found that 26.8±2.7% of the larvae (67 of 250 larvae) that were fed diets containing sesamin, DMSO, and 3 µM imidacloprid pupated. In contrast, the pupation rate was 27.6±4.9% (69 of 250) among the controls. However, no significant difference was observed between the two groups (p > 0.05). These results indicate that the induction of Cyp6a2 and Cyp6g1 expression by sesamin in the digestion-related tissues did not confer resistance to 3 µM imidacloprid.
4. Discussion
4.1. Activation of Nrf2/Cnc in Specific Larval Tissues by Sesamin
Sesamin is an abundantly occurring compound in sesame seeds. Its health benefits such as its antioxidant [27], antihypertensive [41], and alcohol-metabolizing [42] properties have been previously reported. Previous studies have revealed that sesamin elicits its antioxidant properties in Drosophila adults by activating the Nrf2/Cnc transcription factor, which is required for oxidative stress response [5]. Moreover, Nrf2 activation may be caused by the inhibition of Nrf2–Keap1 binding by sesamin [6]. Additionally, sesamin activates Nrf2/Cnc in larval tissues. Strong Nrf2 activation was observed in the CNS, salivary glands, and gut, including the upper midgut and hindgut. These results are consistent with those of previous studies showing that sesamin activates Nrf2 in the adult brain and gut [5]. The activation of Nrf2 in intestinal epithelial cells by sesamin may be transmitted to the CNS via neural networks. Drosophila enteric nerves connect to two regions: the region consisting of the foregut, proventriculus, and upper midgut and the hindgut with no nerve extensions or connections in the majority of the midgut, which is the largest region of the intestinal tract [43,44,45]. Therefore, Drosophila intestinal epithelial cells, in which Nrf2 is activated during the absorption of sesamin, may transmit signals to the CNS via neural networks. Alternatively, sesamin may act directly on the CNS via the hemolymph after absorption by intestinal epithelial cells. In mammalian liver, sesamin is metabolized to polyphenol-like metabolite SC1, which is present as an intermediate product. Sesamin is also metabolized to SC1 in the gut epithelial cells of Drosophila adults [46]. It is metabolized to SC1 in intestinal epithelial cells, absorbed, released into the larval hemolymph, and transported to the CNS through the larval hemolymph. A rigid barrier in mammals known as the blood–brain barrier prevents most drugs and other substances in the blood from entering the brain [47]. In contrast, the blood–brain barrier in Drosophila is relatively more permeable than that in mammals [48]. Therefore, SCI, which is a water-soluble metabolite of sesamin, may enter the CNS directly via the hemolymph without using a neural network connected to the gut cells. We showed that sesamin consumption induced higher Nrf2 activation in some tissues but not in others. This variation in the degree of Nrf2 activation by sesamin among larval tissues could be attributed to variation in sensitivity to sesamin or efficiency of its uptake in different tissues. Hence, the mechanisms underlying the cell-specific effects of sesamin need to be clarified in future studies.
4.2. Activation of Nrf2/Cnc in Larval Glial Cells in CNS by Sesamin
This study demonstrated intense Nrf2 activation in glial cells within the CNS of sesamin-fed larvae. The ectopic expression of Keap1, which is an Nrf2 inhibitor, in glial cells repressed most sesamin-induced Nrf2 activation in the CNS. However, Nrf2 induction in the neurons did not change to a significant level. This suggests that Nrf2 activation in the CNS occurs primarily in glial cells. Glial cells are intimately involved in the physiological actions of the entire CNS, including developmental and nutritional processes, the immune system, and electrical insulation of neurons [49,50,51]. Glial cells are as crucial as neurons in maintaining brain integrity and function [52]. Glial cell dysfunction is closely associated with various neurological disorders such as autism, schizophrenia, and neurodegeneration. Nrf2 activation in glial cells suppresses Parkinson’s phenotype in a Drosophila disease model [53]. Nrf2 activation in the glial cells rather than in neurons elicited a greater protective effect against oxidative stress in the CNS. In this study, sesamin strongly activated Nrf2 in glial cells of the larval CNS. Sesamin consumption activates Nrf2 in specific neurons such as glutamatergic, cholinergic, and dopaminergic neurons in the adult brain. Furthermore, it suppresses neuronal loss associated with oxidative stress accumulation [5,6]. Although Nrf2 activation in glial cells of the adult brain was not examined in this study, the neuroprotective effect of sesamin may be reasonably attributed to the activation of Nrf2 in glial cells associated with neurons in the Drosophila adult’s brain. Nrf2-dependent expression of antioxidative genes in glial cells may improve the oxidative condition of neurons, which could maintain neuronal homeostasis in the CNS.
The activation of NF-κB transcription factor-mediated signaling in innate immune pathways was accelerated with oxidative stress and vascular inflammation in aging in rats [54]. During neuroinflammation observed in neurodegenerative disease models, the microglia are activated and secrete excessive cytokines that stimulate astrocytes to secrete extracellular factors that induce neuronal cell death [55]. Recently, the production of neuronal death-inducing factors in astrocytes was reported to be suppressed by inhibiting the NF-κB pathway via Nrf2 activation in astrocytes [56]. We found that sesamin activates Nrf2 in astrocytes within the larval CNS. Therefore, sesamin-induced Nrf2 activation in astrocytes may suppress neurodegeneration by suppressing excess cytokine production in the astrocytes. We plan to study this further in future studies. Thus, sesamin is a promising health-promoting agent that suppresses the progression of human neurodegenerative diseases and alleviates their symptoms.
4.3. Induction of Cytochrome P450 Drug-Metabolizing Gene Expression in the Larval CNS and Digestion-Related Tissues by Sesamin and Its Effects on the Organism
This study demonstrates that sesamin consumption significantly increases the mRNA level of Cyp6a2 in the larval CNS and salivary glands and that there is a tendency for it to increase the level of Cyp6g1 in salivary glands and gut. The Cyp6a2 gene is an ortholog of the human CYP3A4 gene, which encodes an enzyme that is involved in the metabolism of hypnotic sedatives, antidepressants, and antiepileptic drugs. Cyp6g1 can also metabolize dichlorodiphenyltrichloroethane (DDT) and imidacloprid, which are used as pesticides [57]. Furthermore, Drosophila wild-type stocks that show resistance to DDT also exhibit elevated expression of Cyp6g1 and Cyp6a2 genes [58,59]. Thus, sesamin-induced elevated expression of these genes could imply accelerated drug metabolism; that is, the drug would be rapidly degraded in the body. The sesamin-administered larvae were fed imidacloprid to examine whether resistance was conferred against the pesticides. Contrary to expectations, no significant changes were observed in drug-resistance when compared with that of the controls. As amino acid substitution in Cyp6a2 was observed when reporting altered activity in resistant stocks [60], increased Cyp6a2 expression as that observed in this study may not be sufficient to impart resistance; alternatively, the elevated expression levels of the two cytochrome P450 genes could have been insufficient. Additionally, we cannot exclude the fact that the imidacloprid concentration used in this study was considerably high; hence, the sesamin-induce increase in Cyp6a2 and Cyp6g1 expression may be insufficient to eliminate a substantial concentration of the toxin. Neonicotinoids, including imidacloprid, disrupt insect neurotransmission by binding to nicotinic acetylcholine receptors (nAchR). Although the effects of these toxins on mammals have not been fully elucidated, they are known to adversely affect the human brain—particularly neural development [61]. The present study found that sesamin activates Nrf2 in the developing larval CNS. Therefore, if the detoxifying effects of sesamin on neonicotinoids can be verified, similar adverse effects on human neural development may be possibly prevented. DDT is absorbed through the insect epidermis and acts on Na+ channels on reaching the nerve terminals. DDT is metabolized in the body into highly bioaccumulating metabolites. Although these pesticides are no longer used, they remain bioaccumulated in the environment. They are associated with the pathogenesis of certain types of cancers and Alzheimer’s disease [61,62]. DDT is difficult to metabolize and excrete but the Cyp6g1 and Cyp6a2 produced in the DDT-resistant mutants may metabolize and detoxify DDT. Sesamin would provide a new utility if it could promote DDT detoxification by upregulation of drug-metabolizing enzymes. Therefore, the effect of sesamin on DDT detoxification should be investigated in future studies.
Author Contributions
Conceptualization, Y.H.I.; methodology, A.T. and Y.H.I.; validation, A.T. and Y.H.I.; formal analysis, A.T.; investigation, A.T.; data curation, Y.H.I.; writing—original draft preparation, A.T.; writing—review and editing, Y.H.I.; visualization, Y.H.I and E.K.; supervision, Y.H.I.; project administration, Y.H.I.; funding acquisition, Y.H.I. and E.K. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially supported by the Grant-in-Aid for Scientific Research (C) 17K07500 to Y.H.I.
Institutional Review Board Statement
Ethical review and approval were waived for this study because this experimental plan is not restricted under Japanese law.
Informed Consent Statement
Not applicable.
Data Availability Statement
The study did not report any data.
Acknowledgments
We thank Dirk Bohmann for providing the fly stock. We also thank Bloomington Drosophila Stock Center, Kyoto Stock Center, and Vienna Drosophila Resource Center for providing fly stocks. We acknowledge Akiko Watanabe and Mina Platt for their technical assistance.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Andargie, M.; Vinas, M.; Rathgeb, A.; Möller, E.; Karlovsky, P. Lignans of sesame (Sesamum indicum L.): A Comprehensive Review. Molecules 2021, 26, 883. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, K.; Kitagawa, Y.; Akamatsu, T.; Hirose, N.; Sugano, M.; Shimizu, S.; Yamada, H. Protective effects of sesamin against liver damage caused by alcohol or carbon tetrachloride in rodents. Ann. Nutr. Metab. 1993, 37, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Ashakumary, L.; Rouyer, I.; Takahashi, Y.; Ide, T.; Fukuda, N.; Aoyama, T.; Hashimoto, T.; Mizugaki, M.; Sugano, M. Sesamin, a sesame lignan, is a potent inducer of hepatic fatty acid oxidation in the rat. Metabolism: Clinical and Experimental 1999, 48, 1303. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gao, Y.; Li, S.; Yang, J. Effect of sesamin on pulmonary vascular remodeling in rats with monocrotaline-induced pul monary hypertension. China journal of Chinese materia medica 2015, 40, 1355–1361. [Google Scholar] [PubMed]
- Le, T.D.; Nakahara, Y.; Ueda, M.; Okumura, K.; Hirai, J.; Sato, Y.; Takemoto, D.; Tomimori, N.; Ono, Y.; Nakai, M.; Shibata, H.; Inoue, Y.H. Sesamin suppresses aging phenotypes in adult muscular and nervous systems and intestines in a Drosophila senescence-accelerated model. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1826–1839. [Google Scholar] [PubMed]
- Le, T.D.; Inoue, Y.H. Sesamin Activates Nrf2/Cnc-dependent transcription in the absence of oxidative stress in Drosophila adult brains. Antioxidants 2021, 10, 924. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, S.; Hiraku, Y.; Oikawa, S. Mechanism of guanine-specific DNA damage by oxidative stress and its role in carcinogenesis and aging. Mutat. Res. 2001, 488, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Glei, M.; Schaeferhenrich, A.; Claussen, U.; Kuechler, A.; Liehr, T.; Weise, A.; Marian, B.; Sendt, W.; Pool-Zobel, B.L. Comet fluorescence in situ hybridization analysis for oxidative stress–induced DNA damage in colon cancer relevant genes. Toxicol. Sci. 2007, 96, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, A.; Idelchik, M.d.P. S.; Melendez, J.A. Redox control of senescence and age-related disease. Redox Biol. 2016, 11, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Hedge, M.; Lortz, S.; Drinkgern, J.; Lenzen, S. Relation between antioxidant enzyme gene expression and antioxidative defense status of insulin-producing cells. Skelet. Muscle 1997, 46. [Google Scholar]
- Kirkman, H.N.; Rolfo, M.; Ferraris, A.M.; Gaetani, G.F. Mechanisms of protection of catalase by NADPH: KINETICS AND STOICHIOMETRY. J. Biol. Chem. 1999, 274, 13908–13914. [Google Scholar] [CrossRef] [PubMed]
- Hirotsu, Y.; Katsuoka, F.; Funayama, R.; Nagashima, T.; Nishida, Y.; Nakayama, K.; Engel, J.D.; Yamamoto, M. Nrf2-MafG heterodimers contribute globally to antioxidant and metabolic networks. Nucleic Acids Res. 2012, 40, 10228–10239. [Google Scholar] [CrossRef] [PubMed]
- Moi, P.; Chan, K.; Asunis, I.; Cao, A.; Kan, Y.W. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proc. Natl. Acad. Sci. U.S.A. 1994, 91, 9926–9930. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, R.; Jaiswal, A.K. Nrf1 and Nrf2 positively and c-Fos and Fra1 negatively regulate the human antioxidant response element-mediated expression of NAD(P)H:quinone oxidoreductase1 gene. Proc. Natl. Acad. Sci. U.S.A. 1996, 93, 14960–14965. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; Yamamoto, M.; Nabeshima, Y. An Nrf2/Small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef] [PubMed]
- McMahon, M.; Swift, S.R.; Hayes, J.D. Zinc-binding triggers a conformational-switch in the cullin-3 substrate adaptor protein KEAP1 that controls transcription factor NRF2. Toxicol. Appl. Pharmacol. 2018, 360, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Kang, M.-I.; Watai, Y.; Tong, K.I.; Shibata, T.; Uchida, K.; Yamamoto, M. Oxidative and electrophilic stresses activate Nrf2 through inhibition of Ubiquitination activity of Keap1. Mol.Cell. Biol. 2006, 26, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, N.; Bohmann, D. A versatile ΦC31 based reporter system for measuring AP-1 and Nrf2 signaling in Drosophila and in tissue culture. PLoS One 2012, 7, e34063. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Cole, R.N.; Itoh, K.; Wakabayashi, N.; Katoh, Y.; Yamamoto, M.; Talalay, P. Direct evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidants. Proc. Natl. Acad. Sci. U. S. A. 2002, 99, 11908–11913. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, M.; Xiong, Y. BTB protein Keap1 targets antioxidant transcription factor Nrf2 for ubiquitination by the Cullin 3-Roc1 ligase. Mol Cell Biol. 2005, 25, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Hartenstein, V.; Tepass, U.; Gruszynski-Defeo, E. Embryonic development of the stomatogastric nervous system in Drosophila. J. Comp. Neurol. 1994, 350, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Kanayama, M.; Shibata, T.; Itoh, K.; Kobayashi, A.; Yamamoto, M.; Uchida, K. Ebselen, a seleno-organic antioxidant, as an electrophile. Chem. Res. Toxicol. 2006, 19, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Rachakonda, G.; Xiong, Y.; Sekhar, K.R.; Stamer, S.L.; Liebler, D.C.; Freeman, M.L. Covalent modification at Cys151 dissociates the electrophile sensor Keap1 from the ubiquitin ligase CUL3. Chem. Res. Toxicol. 2008, 21, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.-H.; Yen, G.-C. Differential expressions of antioxidant status in aging rats: The role of transcriptional factor Nrf2 and MAPK signaling pathway. Biogerontol. 2007, 8, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Schmidlin, C.J.; Dodson, M.B.; Madhavan, L.; Zhang, D.D. Redox regulation by NRF2 in aging and disease. Free Radic. Biol. Med. 2019, 134, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, T.; Nishijima, Y.; Shibata, H.; Kiso, Y.; Ohnuki, K.; Fushiki, T.; Moritani, T. Protective effect of sesamin administration on exercise-induced lipid peroxidation. Int. J. Sports Med. 2003, 24, 530–534. [Google Scholar] [PubMed]
- Nakai, M.; Harada, M.; Nakahara, K.; Akimoto, K.; Shibata, H.; Miki, W.; Kiso, Y. Novel Antioxidative Metabolites in Rat Liver with Ingested Sesamin. J. Agric. Food Chem. 2003, 51, 1666–1670. [Google Scholar] [CrossRef] [PubMed]
- Kiso, Y. Antioxidative roles of sesamin, a functional lignan in sesame seed, and it’s effect on lipid- and alcohol-metabolism in the liver: A DNA microarray study. BioFactors 2004, 21, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Ou, H.C.; Wu, C.M.; Lee, I.T.; Lin, S.Y.; Lin, L.Y.; Tsai, K.L.; Lee, S.D.; Sheu, W.H.H. Sesamin mitigates inflammation and oxidative stress in endothelial cells exposed to oxidized low-density lipoprotein. J. Agric. Food Chem. 2009, 57, 11406–11417. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, D.; Yasutake, Y.; Tomimori, N.; Ono, Y.; Shibata, H.; Hayashi, J. Sesame lignans and vitamin E supplementation improve subjective statuses and anti-oxidative capacity in healthy humans with feelings of daily fatigue. Glob. J. Health Sci. 2015, 7, 1–10. [Google Scholar] [CrossRef]
- Hagen, T.M. Oxidative Stress, Redox Imbalance, and the Aging Process. Antioxid. Redox Sign. 2003, 5, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, R.; Samokhina, E.; Rossetti, I.; Morley, J.W.; Buskila, Y. Neuromodulation of glial function during neurodegeneration. Front. cell. neurosci. 2020, 14. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.A.; Krantz, D.E. Drosophila melanogaster as a genetic model system to study neurotransmitter transporters. Neurochem. Int. 2014, 73, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Lessing, D.; Bonini, N.M. Maintaining the brain: Insight into human neurodegeneration from Drosophila melanogaster mutants. Nature Rev. Genet. 2009, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Nichols, C.D. Drosophila melanogaster neurobiology, neuropharmacology, and how the fly can inform CNS drug discovery. Pharmacol. Therapeut. 2006, 112, 677–700. [Google Scholar] [CrossRef] [PubMed]
- Rera, M.; Clark, R.I.; Walker, D.W. Intestinal barrier dysfunction links metabolic and inflammatory markers of aging to death in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 2012, 109, 21528–21533. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Hirai, J.; Yasukawa, T.; Nakahara, Y.; Inoue, Y.H. A correlation of reactive oxygen species accumulation by depletion of superoxide dismutases with age-dependent impairment in the nervous system and muscles of Drosophila adults. Biogerontol. 2015, 16, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Araki, M.; Kurihara, M.; Kinoshita, S.; Awane, R.; Sato, T.; Ohkawa, Y.; Inoue, Y.H. Anti-tumour effects of antimicrobial peptides, components of the innate immune system, against haematopoietic tumours in Drosophila mxc mutants. Dis. Model Mech. 2019, 12, dmm037721. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of Nrf2 in oxidative stress and toxicity. Ann. Rev. Pharmacol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Fusetto, R.; Denecke, S.; Perry, T.; O’Hair, R.A.J.; Batterham, P. Partitioning the roles of CYP6G1 and gut microbes in the metabolism of the insecticide imidacloprid in Drosophila melanogaster. Sci. Rep. 2017, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Nakano, D.; Kwak, C.J.; Fujii, K.; Ikemura, K.; Satake, A.; Ohkita, M.; Takaoka, M.; Ono, Y.; Nakai, M.; Tomimori, N.; Kiso, Y.; Matsumura, Y. Sesamin metabolites induce an endothelial nitric oxide-dependent vasorelaxation through their antioxidative property-independent mechanisms: possible involvement of the metabolites in the antihypertensive effect of sesamin. J. Pharmacol. Exp. Ther. 2006, 318, 328–35. [Google Scholar] [CrossRef] [PubMed]
- Tsuruoka, N.; Kidokoro, A.; Matsumoto, I.; Abe, K.; Kiso, Y. Modulating effect of sesamin, a functional lignan in sesame seeds, on the transcription levels of lipid- and alcohol-metabolizing enzymes in rat liver: A DNA microarray study. Biosci. Biotechnol. Biochem. 2005, 69, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Hartenstein, V.; Tepass, U.; Gruszynski-Defeo, E. Embryonic development of the stomatogastric nervous system in Drosophila. J. Comp. Neurol. 1994, 350, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Spiess, R.; Schoofs, A.; Heinzel, H.-G. Anatomy of the stomatogastric nervous system associated with the foregut in Drosophila melanogaster and Calliphora vicina third instar larvae. J. Morphol. 2008, 269, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Cognigni, P.; Bailey, A.P.; Miguel-Aliaga, I. Enteric neurons and systemic signals couple nutritional and reproductive status with intestinal homeostasis. Cell Metab. 2011, 13, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Ikushiro, S.; Kamakura, M.; Ohta, M.; Sakaki, T. Metabolism of sesamin by cytochrome P450 in human liver microsomes. Drug Metab. Dispos. 2010, 38, 2117–2123. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Blood-brain barrier drug targeting: the future of brain drug development. Mol. Interv. 2003, 3, 90. [Google Scholar] [CrossRef] [PubMed]
- Hindle, S.J.; Bainton, R.J. Barrier mechanisms in the Drosophila blood-brain barrier. Front. Cell Neurosci. 2014, 8, 414. [Google Scholar] [CrossRef] [PubMed]
- Ullian, E.M.; Sapperstein, S.K.; Christopherson, K.S.; Barres, B.A. Control of synapse number by glia. Science 2001, 291, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu, C.S.; Tani, M.; Ransohoff, R.M.; Wysocka, M.; Hilliard, B.; Fujioka, T.; Murphy, S.; Tighe, P.J.; Das Sarma, J.; Trinchieri, G.; Rostami, A. Astrocytes as antigen-presenting cells: Expression of IL-12/IL-23. J. Neurochem. 2005, 95, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Edgar, J.M.; Garbern, J. The myelinated axon is dependent on the myelinating cell for support and maintenance: Molecules involved. J. Neurosci. Res. 2004, 76, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Edenfeld, G.; Stork, T.; Klämbt, C. Neuron-glia interaction in the insect nervous system. Curr. Opin. Neurobiol. 2005, 15, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Wang, B.; Wang, X.; Smith, W.W.; Zhu, Y.; Liu, Z. Activation of Nrf2 in astrocytes suppressed PD-Like phenotypes via antioxidant and autophagy pathways in rat and Drosophila models. Cells 2021, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Orosz, Z.; Labinskyy, N.; Rivera, A.; Xiangmin, Z.; Smith, K.; Csiszar, A. Increased mitochondrial H2O2 production promotes endothelial NF-κB activation in aged rat arteries. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H37–H47. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; Wilton, D.K.; Frouin, A.; Napier, B.A.; Panicker, N.; Kumar, M.; Buckwalter, M.S.; Rowitch, D.H.; Dawson, V.L.; Dawson, T.M.; … Barres, B.A. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Nakano-Kobayashi, A.; Canela, A.; Yoshihara, T.; Hagiwara, M. Astrocyte-targeting therapy rescues cognitive impairment caused by neuroinflammation via the Nrf2 pathway. Proc. Natl. Acad. Sci. U.S.A. 2023, 120, e2303809120. [Google Scholar] [CrossRef] [PubMed]
- Joußen, N.; Heckel, D.G.; Haas, M.; Schuphan, I.; Schmidt, B. Metabolism of imidacloprid and DDT by P450 CYP6G1 expressed in cell cultures of Nicotiana tabacum suggests detoxification of these insecticides in Cyp6g1-overexpressing strains of Drosophila melanogaster, leading to resistance. Pest Manag. Sci. 2008, 64, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Daborn, P.; Boundy, S.; Yen, J.; Pittendrigh, B.; French-Constant, R. DDT resistance in Drosophila correlates with Cyp6g1 over-expression and confers cross-resistance to the neonicotinoid imidacloprid. Mol. Genet. Genomics 2001, 266, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Pedra, J.H.F.; McIntyre, L.M.; Scharf, M.E.; Pittendrigh, B.R. Genome-wide transcription profile of field- and laboratory- selected dichlorodiphenyltrichloroethane (DDT)-resistant Drosophila. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 7034–7039. [Google Scholar] [CrossRef]
- Amichot, M.; Tarès, S.; Brun-Barale, A.; Arthaud, L.; Bride, J.M.; Bergé, J.B. Point mutations associated with insecticide resistance in the Drosophila cytochrome P450 Cyp6a2 enable DDT metabolism. Eur. J. Biochem. 2004, 271, 1250–1257. [Google Scholar] [CrossRef]
- Kimura-Kuroda, J.; Komuta, Y.; Kuroda, Y.; Hayashi, M.; Kawano, H. Nicotine-Like Effects of the neonicotinoid insecticides acetamiprid and imidacloprid on cerebellar neurons from neonatal rats. PLOS ONE 2012, 7, e32432. [Google Scholar] [CrossRef] [PubMed]
- Cocco, P.; Blair, A.; Congia, P.; Saba, G.; Flore, C.; Ecca, M.R.; Palmas, C. Proportional mortality of dichloro-diphenyl-trichloroethane (DDT) workers: A preliminary report. Arch. Environ. Health 1997, 52, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.R.; Roy, A.; Shalat, S.L.; von Stein, R.T.; Hossain, M.M.; Buckley, B.; Gearing, M.; Levey, A.I.; German, D.C. Elevated serum pesticide levels and risk for Alzheimer disease. JAMA Neurol. 2014, 71, 284–290. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Observation of ARE-GFP fluorescence in the whole body of larvae fed sesamin. (A) Stereo fluorescence microscopy images of whole body GFP fluorescence of ARE-GFP larvae. a-n are bright field stereomicroscopic images of larvae, and a’-n’ are fluorescence stereomicroscopic images of the same field of view. Green indicates ARE-GFP fluorescence. The fluorescence is stronger in larvae fed sesamin than in controls. Scale bar is 1.0 mm. (B) Quantification of GFP-positive regions in ARE-GFP larvae. The numbers of male and female larvae used in this assay are noted at the bottom. The vertical axis of the graph shows the number of pixels of GFP-positive regions in the whole larval body. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001: Welch’s t-test. Bars indicate SEM.
Figure 1.
Observation of ARE-GFP fluorescence in the whole body of larvae fed sesamin. (A) Stereo fluorescence microscopy images of whole body GFP fluorescence of ARE-GFP larvae. a-n are bright field stereomicroscopic images of larvae, and a’-n’ are fluorescence stereomicroscopic images of the same field of view. Green indicates ARE-GFP fluorescence. The fluorescence is stronger in larvae fed sesamin than in controls. Scale bar is 1.0 mm. (B) Quantification of GFP-positive regions in ARE-GFP larvae. The numbers of male and female larvae used in this assay are noted at the bottom. The vertical axis of the graph shows the number of pixels of GFP-positive regions in the whole larval body. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001: Welch’s t-test. Bars indicate SEM.
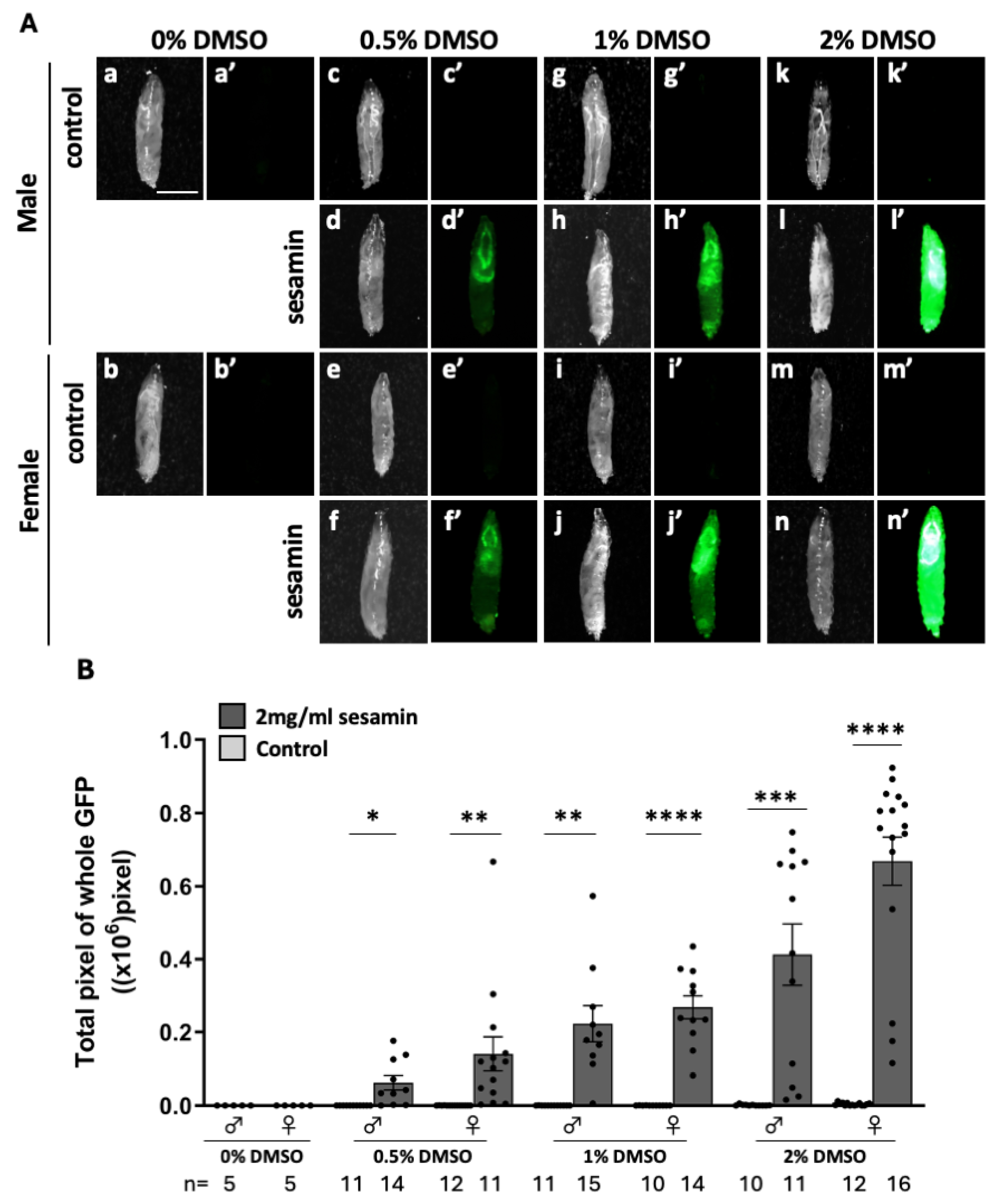
Figure 2.
GFP fluorescence of ARE-GFP reporter in the larval CNS and digestive-related tissues and quantitation of GFP mRNA in the larvae fed sesamin. (A, D, F) Confocal fluorescence microscopy images of the larval CNSs (A), gut (B), salivary glands (D) of ARE-GFP larvae. Note that intense GFP fluorescence was strongly observed, in the midgut and hindgut of sesamin-feeding larvae. Green indicates GFP fluorescence. The scale bars indicate 100 µm (A, F) or 1 mm (D). (B) Mean (relative) GFP fluorescence intensity in the CNS of ARE-GFP larvae. The vertical axis of the graph shows the relative values of the mean GFP fluorescence intensity in the CNSs of larval males reared on the diet without sesamin (supplemented with DMSO 1%), as a mean value of 1. (C, E, G) Relative expression levels of GFP in the CNS (C), gut (E) or salivary gland (G) in female larvae fed the diets supplemented with sesamin 1mg/mL (DMSO 1%) (control; light gray bars; sesamin-feeding; dark gray bars) were quantified by qRT-PCR. GFP mRNA levels increased from 39- to 4,897-fold on average. B; the numbers of male and female larvae used are noted at the bottom. C, E, G; n=3 (40 larvae/experiment, triplicates). n.s. = no significant, ****p<0.0001: Student’s t-test or Welch’s t-test). Bars indicate SEM.
Figure 2.
GFP fluorescence of ARE-GFP reporter in the larval CNS and digestive-related tissues and quantitation of GFP mRNA in the larvae fed sesamin. (A, D, F) Confocal fluorescence microscopy images of the larval CNSs (A), gut (B), salivary glands (D) of ARE-GFP larvae. Note that intense GFP fluorescence was strongly observed, in the midgut and hindgut of sesamin-feeding larvae. Green indicates GFP fluorescence. The scale bars indicate 100 µm (A, F) or 1 mm (D). (B) Mean (relative) GFP fluorescence intensity in the CNS of ARE-GFP larvae. The vertical axis of the graph shows the relative values of the mean GFP fluorescence intensity in the CNSs of larval males reared on the diet without sesamin (supplemented with DMSO 1%), as a mean value of 1. (C, E, G) Relative expression levels of GFP in the CNS (C), gut (E) or salivary gland (G) in female larvae fed the diets supplemented with sesamin 1mg/mL (DMSO 1%) (control; light gray bars; sesamin-feeding; dark gray bars) were quantified by qRT-PCR. GFP mRNA levels increased from 39- to 4,897-fold on average. B; the numbers of male and female larvae used are noted at the bottom. C, E, G; n=3 (40 larvae/experiment, triplicates). n.s. = no significant, ****p<0.0001: Student’s t-test or Welch’s t-test). Bars indicate SEM.
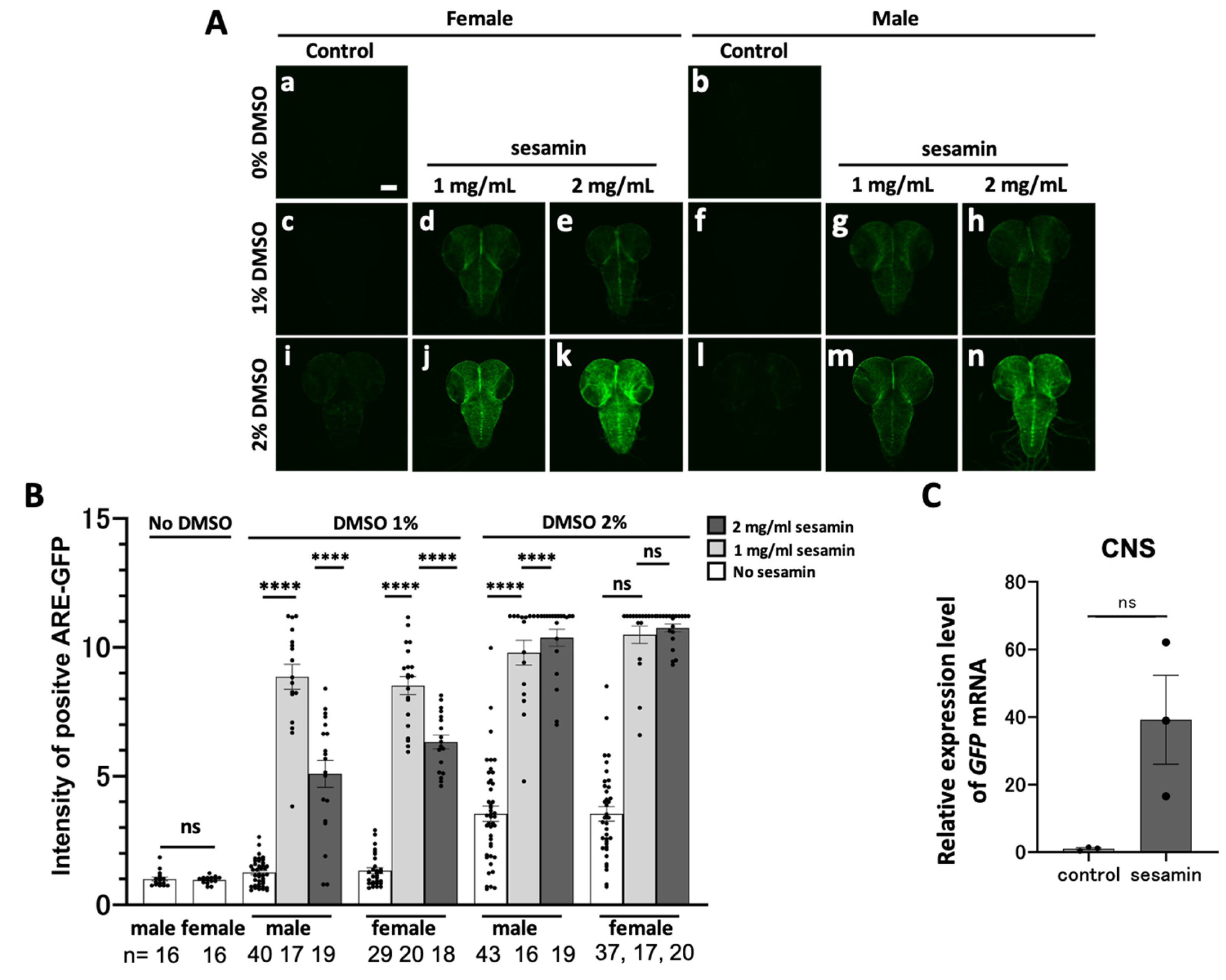
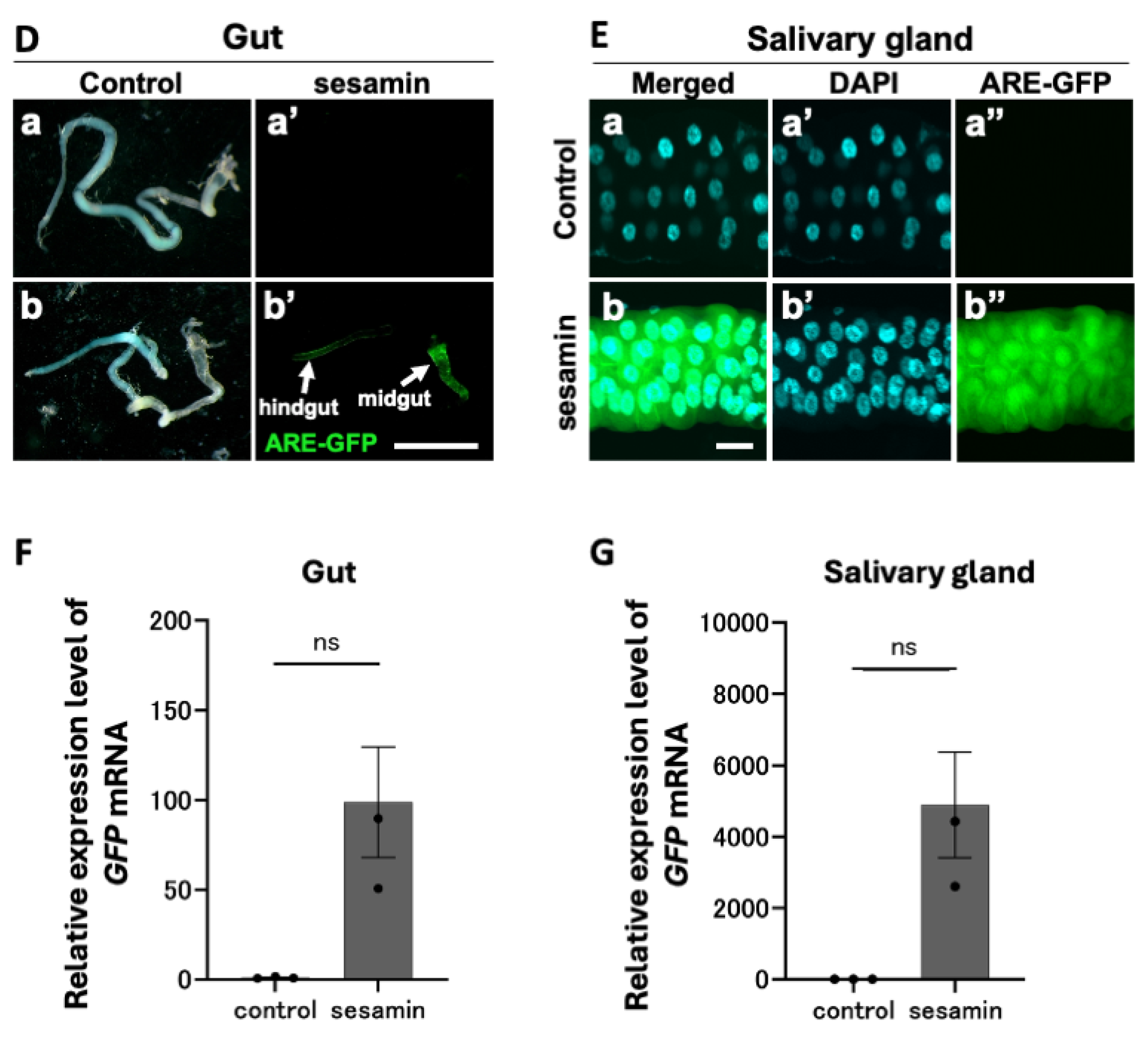
Figure 3.
Observation of pan-glial cells within the larval CNS and promotion of its expression by sesamin. (A, B) Confocal fluorescence microscopy images of GFP fluorescence in all neurons (elav-Gal4/+; ARE-GFP/+; UAS-RFP/+ (A)) or all glial cells (ARE-GFP/+; repo-Gal4/UAS-RFP (B)) within the CNS of larvae. GFP fluorescence is observed in glial cells. Red indicates all neurons or all glial cells, and green indicates ARE-GFP. Scale bar is 100 µm. (C) Confocal fluorescence microscopy images of GFP fluorescence in CNS in ARE-GFP larvae harboring ectopic overexpression of Keap1 (elav-Gal4/+; ARE-GFP/+; UAS-Keap1EY/+ (Cb, f), ARE-GFP/+; repo-Gal4/UAS-Keap1EY (Cd, h)). (D) Quantification of GFP-positive regions in CNS in ARE-GFP larvae harboring ectopic overexpression of Keap1. GFP regions were reduced in sesamin-fed larvae overexpressing Keap1 in all glial cells. (D; elav-Gal4/+ (control; n=21, sesamin; n=24), elav-Gal4/+; UAS-Keap1EY/+ (control; n=26, sesamin; n=20), repo-Gal4/+ (control; n=11, sesamin; n=20), n.s.=no significant, **p<0.01, ****p<0.0001; Student’s t-test or Welch’s t-test). Bars indicate SEM.
Figure 3.
Observation of pan-glial cells within the larval CNS and promotion of its expression by sesamin. (A, B) Confocal fluorescence microscopy images of GFP fluorescence in all neurons (elav-Gal4/+; ARE-GFP/+; UAS-RFP/+ (A)) or all glial cells (ARE-GFP/+; repo-Gal4/UAS-RFP (B)) within the CNS of larvae. GFP fluorescence is observed in glial cells. Red indicates all neurons or all glial cells, and green indicates ARE-GFP. Scale bar is 100 µm. (C) Confocal fluorescence microscopy images of GFP fluorescence in CNS in ARE-GFP larvae harboring ectopic overexpression of Keap1 (elav-Gal4/+; ARE-GFP/+; UAS-Keap1EY/+ (Cb, f), ARE-GFP/+; repo-Gal4/UAS-Keap1EY (Cd, h)). (D) Quantification of GFP-positive regions in CNS in ARE-GFP larvae harboring ectopic overexpression of Keap1. GFP regions were reduced in sesamin-fed larvae overexpressing Keap1 in all glial cells. (D; elav-Gal4/+ (control; n=21, sesamin; n=24), elav-Gal4/+; UAS-Keap1EY/+ (control; n=26, sesamin; n=20), repo-Gal4/+ (control; n=11, sesamin; n=20), n.s.=no significant, **p<0.01, ****p<0.0001; Student’s t-test or Welch’s t-test). Bars indicate SEM.
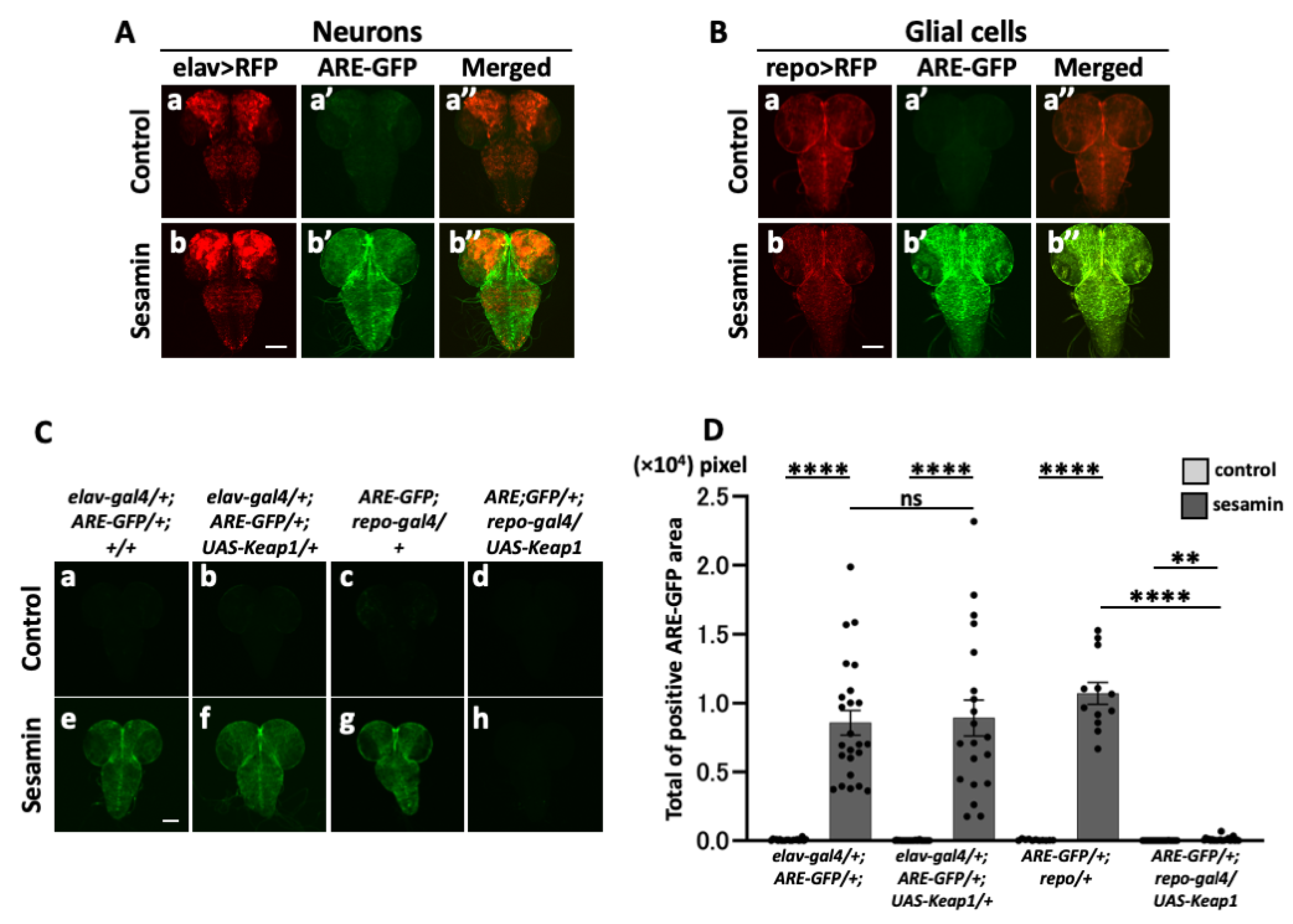
Figure 4.
Observation of ARE-GFP in specific glial cells within the CNS of larvae and its expression enhanced by sesamin. (A, D) Confocal fluorescence microscopy images of GFP fluorescence in the glial cells in the CNS of Eaat1-Gal4/ARE-GFP; UAS-RFP/+ larvae (A) or Nrv2-Gal4/ARE-GFP; Nrv2-Gal4/UAS-RFP larvae (D). Red indicates astrocytes (A) or cortex and surface glial cells, and green indicates ARE-GFP. The scale bar is 100 µm. (B, E) The intensity of the GFP fluorescence in and around astrocytes (B) or cortex and surface glial cells (E) (quantified from 0 to 255). The intensity in each glial cell was significantly increased in the larvae fed sesamin compared to controls. (C, F) Percentage of ARE-GFP region in astrocytes or cortex and surface glial cells. In each glial cell, the percentage was significantly increased compared to controls (n=12 (control), n=18 (sesamin) (B, C), n=16 (control), n=18 (sesamin) (E, F), ****p<0.0001, Welch’s t- test). Bars indicate SEM.
Figure 4.
Observation of ARE-GFP in specific glial cells within the CNS of larvae and its expression enhanced by sesamin. (A, D) Confocal fluorescence microscopy images of GFP fluorescence in the glial cells in the CNS of Eaat1-Gal4/ARE-GFP; UAS-RFP/+ larvae (A) or Nrv2-Gal4/ARE-GFP; Nrv2-Gal4/UAS-RFP larvae (D). Red indicates astrocytes (A) or cortex and surface glial cells, and green indicates ARE-GFP. The scale bar is 100 µm. (B, E) The intensity of the GFP fluorescence in and around astrocytes (B) or cortex and surface glial cells (E) (quantified from 0 to 255). The intensity in each glial cell was significantly increased in the larvae fed sesamin compared to controls. (C, F) Percentage of ARE-GFP region in astrocytes or cortex and surface glial cells. In each glial cell, the percentage was significantly increased compared to controls (n=12 (control), n=18 (sesamin) (B, C), n=16 (control), n=18 (sesamin) (E, F), ****p<0.0001, Welch’s t- test). Bars indicate SEM.
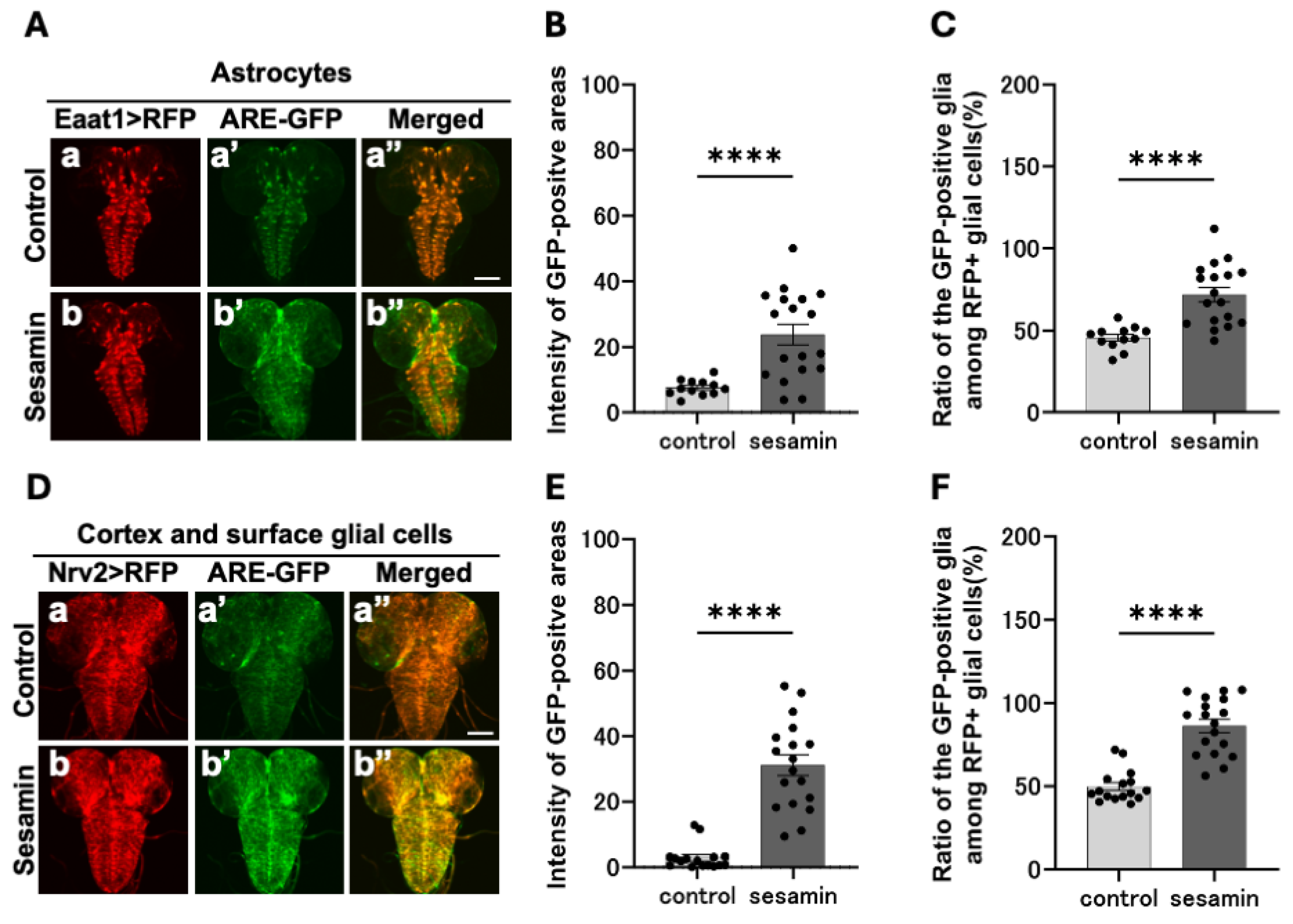
Figure 5.
Quantification of mRNA for Nrf2-regulated genes in the CNSs and digestive-related tissues in larvae reared on sesamin. (A) Relative expression levels of each gene in larvae fed sesamin (control; light gray bars; sesamin; dark gray bars) were quantified by qRT-PCR. Total RNA was isolated from CNSs (A), gut (B), or salivary glands (C) of larvae reared on diets containing sesamin 1 mg/mL, DMSO 1%, and those from larvae reared on diets containing 1% DMSO only as controls for 24 hours. Cyp6a2 mRNA levels were significantly increased in the CNS, gut, and salivary glands in larvae fed sesamin (n=3). (ns, not significant, *p < 0.05; **p < 0.01, Welch’s t-test). Bars indicate SEM.
Figure 5.
Quantification of mRNA for Nrf2-regulated genes in the CNSs and digestive-related tissues in larvae reared on sesamin. (A) Relative expression levels of each gene in larvae fed sesamin (control; light gray bars; sesamin; dark gray bars) were quantified by qRT-PCR. Total RNA was isolated from CNSs (A), gut (B), or salivary glands (C) of larvae reared on diets containing sesamin 1 mg/mL, DMSO 1%, and those from larvae reared on diets containing 1% DMSO only as controls for 24 hours. Cyp6a2 mRNA levels were significantly increased in the CNS, gut, and salivary glands in larvae fed sesamin (n=3). (ns, not significant, *p < 0.05; **p < 0.01, Welch’s t-test). Bars indicate SEM.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



