
Submitted:
21 May 2024
Posted:
21 May 2024
You are already at the latest version
Abstract
Transplanted organs experience several episodes of ischemia and Ischemia-reperfusion. The graft injury resulting from ischemia-reperfusion (IRI) remains a significant obstacle to the successful survival of transplanted grafts. Temperature plays an important role in cellular metabolic rates since biochemical reactions are highly temperature dependent. Therefore, ischemia-triggered degradative reactions could be mitigated by lowering temperature. Whether a local hypothermia on liver before blockage of blood flow protects liver grafts against IRI has not been investigated. In this study, we applied local hypothermia to mouse donor livers for a specific duration before stopping blood flow to liver lobes, a procedure called "liver precooling." Mouse donor liver temperature in control groups was controlled at 37ºC. Subsequently, the liver donors were preserved in cold University of Wisconsin solution for various durations followed by orthotopic liver transplantation. Liver graft injury, function and inflammation were assessed at 1- and 2-days post-transplantation. Liver precooling exhibited a significant improvement in graft function, revealing more than a 47% decrease in plasma aspartate transaminase (AST) and alanine aminotransferase (ALT) levels, coupled with a remarkable reduction of approximately 50% in liver graft histological damage compared to the control group. The protective effects of liver precooling were associated with the preservation of mitochondrial function, substantial reduction in hepatocyte cell death, and a significantly attenuated inflammatory response. A retrospective analysis of patient data revealed a close correlation between hypothermia and enhanced liver graft function. Taken together, reduction of the cellular metabolism and enzymatic activity to a minimum level before ischemia protects against IRI during transplantation.
Keywords:
hypothermia
; mitochondria
; liver transplantation
; graft function
Introduction
Liver transplantation is a life-saving procedure and is often the only treatment option available for patients with end-stage liver disease and acute liver failure[1,2,3,4]. A significant limitation of liver transplantation is the unavoidable hepatic ischemia/reperfusion injury (IRI), which poses a critical risk factor for the survival of transplanted grafts[5,6,7,8]. Furthermore, donation after circulatory death (DCD) is a growing and increasingly important source of liver grafts for transplantation, helping address the shortage of liver donors. However, outcomes from DCD transplants have historically shown a relative inferiority when compared to liver transplantation from a living donor or donors after brain death[9,10]. These adverse outcomes primarily stem from the significant graft injury caused by mandatory warm ischemia during DCD organ recovery.
The injury induced by IR primarily occurs due to the prolonged metabolic demands imposed on a warm ischemic organ and the insufficient supply of oxygen to fulfill these requirements. One approach to enhance the liver’s tolerance to ischemia is by lowering the organ’s temperature[11]. A 50% reduction in cellular metabolic rate occurs with every 10°C decrease in temperature according to the Arrhenius equation. Building on this principle, the invention of cold preservation solution in late 1960′s allowed successful storage of organs in a simple hypothermia solution for up to 15 hours[12,13,14,15]. Recently machine perfusion (MP) at different temperatures has been tried on DCD liver grafts, however, the application of MP to DCD liver transplantation has remained challenging[16,17]. The optimal parameters for the use of MP in DCD livers have not been well established[14,18]. Numerous experimental and clinical studies have been conducted and various drug products have been tested in clinical trials in the peri-transplant period. While some have demonstrated early beneficial effects on IRI, their benefit to long-term graft is yet to be fully demonstrated[19,20,21,22,23,24]. The pathophysiological changes associated with IRI in liver transplantation are not yet well defined, although they have been studied extensively. Future double-blinded, randomized, large-scale clinical trials will be necessary to assess the advantages of MP technology.
Temperature significantly influences cellular metabolic rates as biochemical reactions are greatly sensitive to temperature variations[,25,26]. Therefore, it seems quite likely that most ischemia-triggered degradation reactions can be lessened by lowering tissue temperature[,27,28,29]. Therapeutic hypothermia has been used in surgeries, including liver surgery, since 1960s[30,31,32], however, its significance is still controversial and not conclusive[33,3435,36,37]. Whether inducing local hypothermia in the liver before the stopping of blood flow protects against IRI has not been explored. In the present study, we demonstrated that reduction of the cellular metabolism and oxygen consumption in the liver to a minimum level by local hypothermia before ischemia ameliorates IRI during transplantation.
Materials and Methods
Animals
The use of animals in the experiments was conducted following the approved guidelines from the Institutional Animal Care and Use Committee at the University of South Florida and the National Institutes of Health’s Guide for the Care and Use of Laboratory Animals. Male C57BL/6J mice, aged 12 weeks with a body weight of about 28-32g were obtained from Jackson Lab (Indianapolis, IN). After arrival, the animals were housed in a temperature-controlled environment with 12:12h light-dark cycle and ad libitum access to water and food for 1 week before experiments. The animals were randomly divided into control or precooling groups and donors or recipients based on the requirement of experiments. All chemicals were purchased from Sigma-Aldrich (St. Louis, MO) unless otherwise indicated. Animals were euthanized as needed according to the guidelines set forth by the American Veterinary Medicine Association.
Liver Transplantation with or without Pre-Cooling
Orthotopic liver transplantation (OLT) was performed in mice of the same sex, employing the cuff technique as described in the literature[38]. Briefly, donor mice were anesthetized using isoflurane and the body temperature was kept at 37°C. A midline abdominal incision was made to expose the infra hepatic vena cava (IHVC) and supra hepatic vena cava (SHVC) from the left diaphragmatic vein, right suprarenal vein, and right renal vein. A 22G stent was then inserted into the bile duct, followed by exposing the portal vein (PV) from the pyloric vein. A polystyrene plate was used to isolate the liver lobes from the animal body. The isolated liver lobes were then surrounded with 4 cm length Penrose tubing (3/4” Diameter) filled with 4° (precooling) or 37°C (control) water and circulated with a pump for 30minutes as shown in Figure S. Liver tissue temperature changes were monitored with a flexible probe (Homeothermic Monitoring System, Harvard Apparatus from Holliston, MA, USA) placed between median and left lateral lobes during pre-cooling. The liver was then flushed with 5 mL of cold University of Wisconsin (UW) solution via PV and removed. At last, the obtained graft was preserved in 4°C UW solution for 6 hours. The cuff was prepared before implantation as previously described[38].
Recipient mice were anesthetized, and a midline abdominal incision was made. The anastomosis of SHVC was performed with a running 10-0 suture after removing the original liver. During the recipient operation, two-cuff anastomosis technology was utilized to restore the blood flow of the PV and the IHVC. A biliary stent was used to connect the bile duct. Finally, the abdomen was sutured shut, and the mice were given time to recover.
The sham group also underwent similar procedures and time courses except for the liver transplantation.
Liver Graft Injury and Oxidative Stress Biomarkers
The function of the liver graft was assessed through the measurement of plasma aspartate transaminase (AST), alanine aminotransferase (ALT), and total bilirubin levels by Antech Diagnostics at 1- and 2-days following transplantation. Oxidative stress was evaluated by measuring the liver malondialdehyde (MDA) and the carbonyl content levels as described previously [39, 40]. Briefly, liver tissue was homogenized and incubated with 2,4-dinitrophenylhydrazine. The carbonyl content was determined by utilizing a spectrophotometer set at 370 nm, relying on the reaction between carbonyl groups and 2,4-dinitrophenylhydrazine to produce a 2,4-dinitrophenylhydrazone. Liver MDA levels were assessed using high-performance liquid chromatography at 250 nm with an LC-18 DB column[39].
Histopathological Examinations
At the end of experiment, liver specimens were fixed in 4% paraformaldehyde and embedded in paraffin. Liver samples were sectioned into 4-μm slices and stained with hematoxylin and eosin (H&E). The severity of liver injury was graded from 0–4 for sinusoidal congestion, vacuolization of hepatocyte cyto-plasm, and parenchymal necrosis as described by Suzuki et al. [41]. For statistical analysis, at least 5 visual fields of each specimen were randomly selected and photographed under the microscope (200×, Olympus BX53), and analyzed by Fiji/ImageJ.
Hepatocyte cell death was assessed by Terminal dUTP Nick-End Labeling (TUNEL) staining with in situ Cell Death Detection Kit (Roche, catalog no. 11684795910) according to manufacturer’s instructions. TUNEL-positive cell/nuclei were quantified as the percentage (%) of TUNEL and DAPI double-positive cells relative to total cells (DAPI-positive cells). For statistical analysis, 5 visual fields of each specimen were randomly selected and photographed with fluorescence microscope (Keyence BZ-X710) and analyzed with Fiji/ImageJ.
Immunohistochemical staining (IHC) was performed as previously described[42]. Briefly, the liver slices were subjected to antigen retrieval with a sodium citrate buffer and permeabilized with 0.3% Triton-X 100 in PBS, blocked with 5% normal goat serum, and probed with the primary antibody Caspase 3 (Abcam, ab184787) at the dilution of 1:1000 overnight followed by incubation with Goat anti-Rabbit lgG H&L (HRP) (Abcam, ab97051). The percentage of Caspase 3 positive cells were analyzed with Fiji/ImageJ.
All morphometric analyses were performed blinded to the experimental procedures.
Mitochondria Isolation and Function Evaluation
One separate group of recipients were euthanized 1 day after transplantation for liver mitochondria activity and function evaluation. Mitochondria were isolated and the complex activities and the oxygen consumption rates were measured with a Seahorse XFe24 Analyzer (Agilent Technologies, Inc.) as previously described[43]. Fresh liver samples of 400mg were washed with 4°C isolation buffer (210 mM mannitol, 70 mM sucrose, 5 mM HEPES, 1 mM EGTA, 0.5% BSA pH 7.2) and homogenized using a glass Dounce Homogenizer. The homogenate was centrifuged at 1000×g for 5 min at 4°C. The supernatant, devoid of fat, was gathered and centrifuged at 12,000×g for 10 minutes at 4°C, repeated twice, to yield a mitochondrial pellet, subsequently resuspended in a small volume of isolation buffer. The mitochondrial protein concentration was quantified using a BCA protein assay kit (Pierce, 23227).
Mitochondrial Complex I Activity Mitochondrial OXPHOS complex I (NADH dehydrogenase) enzyme activity was assessed with a Complex I Enzyme Activity Microplate Assay Kit (Colorimetric) (ab109721; Abcam) following the manufacturer’s instruction. Briefly, isolated mitochondria samples with the concentration of 5.5 mg/ml were loaded to the wells of the microplate and incubated for 3 hours at room temperature. Then, 200μl of assay solution was added to each well, and the optical density (OD) at 450 nm was monitored using a microplate reader for 30 minutes at room temperature.
Mitochondrial oxygen consumption rates (OCR) were assessed with a Seahorse XFe24 Analyzer (Agilent Technologies, Inc.) as previously described[43, 44]. Two micrograms of freshly prepared mitochondria were used in this measurement. The respiratory stocks were loaded into the drug ports of a hydrated sensor cartridge in the following order: (A) ADP (1mM, final), (B) oligomycin (2 µM, final), (C) carbonyl-cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP, 4 µM final) and (D) rotenone + antimycin A (2 µM final). A minimum of three cycles of OCR measurements were conducted in the respiration assay. The basal respiration (state 2), phosphorylating respiration in the presence of ADP (state 3), resting respirations with oligomycin (state 4o), maximal uncoupling respiration in the presence of FCCP (state 3u), and the response to antimycin A was determined. Results were analyzed using the Wave software (Agilent Technologies, Inc.) export option and GraphPad Prism.
Cytokine Profiles of Liver Lysates
At the end of experiment, liver tissue sections were harvested, homogenized, and treated with protease/phos-phatase inhibitor cocktail (5872, Cell Signaling Technol-ogy). The levels of cytokines were evaluated by the Proteome Profiler Mouse XL Cytokine Array (ARY028, R&D Systems, Boston, USA) for 111 different analytes involved in the inflammatory response, according to manufacturer’s instructions. Array panels were visualized and captured with a ChemiDoc Imager (Bio-Rad, Inc.). Pixel densities of the cytokine blot spots were analyzed using ImageJ software (National Institutes of Health, Bethesda, MD) and normalized using reference spots.
Real Time PCR
Total RNA isolation and real-time PCR were performed as previously described[45]. Briefly, total RNA was extracted from fresh liver tissue using Trizol reagent (Invitrogen, Carlsbad, CA, USA) and converted to cDNA using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). The primer pairs used for amplification of the interested genes were described in Table 1. β-actin was the invariant control in the experiment. Quantitative PCR analysis was then performed using CFX96 Real-Time Detection System (Chromo4, Bio-Rad, CA, USA) and iQ SYBR Green Supermix (iTaq SYRB, Bio-Rad, CA, USA), according to the protocol of the manufacturer. The amount of each cDNA relative to the β-actin was determined using the 2-ΔΔCt method.
Patient Data
To assess the potential extrapolation of our mouse transplant results to humans, we retrospectively reviewed patient data from liver donors and their corresponding liver transplant recipients from the electronic health records at Tampa General Hospital (TGH). Donor and recipient age, sex, race, and medical histories were extracted from electronic health records at TGH. Patients’ protected health information, including name, birthdate, medical record numbers, etc., were stored on an encrypted, password protected SharePoint site accessible only by approved TGH members. De-identified data were utilized for the statistical analysis. The donors’ body temperature within 30 min before vascular clamping during the procurement and recipients’ serum AST and ALT levels at 1 day post operation have been analyzed. We divided the patients into two groups based on the donors’ body temperature: hypothermia (<36.5oC) and normal temperature (>36.5oC). We then analyzed the correlation between the donor’s body temperature and the function of the transplanted graft.
Statistics
Experimental values are presented as mean ± SEM unless otherwise indicated in the figure legends. Statistical analysis was performed using Prism 10 (GraphPad Software; CA) or SigmaPlot13.0 software. Statistical tests for each dataset are specified in the figure legends, where statistical significance is defined as P < 0.05. Comparisons of the mitochondria bioenergetic analysis and liver function were done by two-way analysis of variance (ANOVA) followed by Tukey multiple comparison test. Comparisons of the datasets of histology and RT-PCR were done using Student’s t test. Patient data were analyzed using SPSS version 24 statistical analysis software (IBM Corp. SPSS Statistics for Windows, Version 25.0. Armonk, NY). The possible linear association between donors’ body temperature and recipients’ serum enzymes was tested using the Pearson’s product moment correlation coefficient.
Results
Liver Temperature Changes during Pre-Cooling
The experimental procedure was depicted in Figure 1. During precooling, the core of the liver lobes and body temperatures were monitored with a Signals 2-Channel Alarm Thermometer assembled with two microprobes. One microprobe was positioned under the median lobe and between the left lateral and the right lateral lobes, the second one was used to measure the body temperature, which was maintained at 36.8°C-37°C utilizing an auxiliary heating lamp and warm water circulating blankets. The liver temperature dropped from 37°C to about 8°C within 8 minutes and maintained at 6-8°C for 30 minutes. Then the liver was flushed with cold UW solution. The temperature of the liver immediately decreased to about 4°C. For the livers without precooling, the temperature was maintained at 36.8°C-37°C before liver flushing and gradually decreased to about 13°C after flushing (Figure 2, n=5/group). Variations in temperature alterations could exert a notable influence on oxygen consumption and ATP preservation, stemming from the distinct declines in metabolic rates.
Male C57BL/6J mice were randomly divided into the groups as described in the methods. For the donor in control group (A), liver lobes were surrounded with 37°C water circulation for 30 min followed by cold storage and OLT and liver graft function evaluation. For the donor in the precooling group (B), liver lobes were surrounded with 4°C water circulation for 30 min followed by cold storage and OLT and liver graft function evaluation. (n=5)
For the precooling group, the liver temperature dropped from 37°C to about 8°C within 8 minutes and maintained at 6-8°C for 30 minutes and quickly decreased to about 4 °C upon flushing with cold saline. In the livers without precooling, the temperature was maintained at 36.8°C-37°C and gradually decreased to about 13°C after flushing.
Pre-Cooling Preserved Mitochondria Activities and Function
Mitochondria are the primary source of ATP generation through oxidative phosphorylation. Mitochondrial metabolism is sensitive to temperature. In cold ischemia, ATP depletion leads to an increase in the osmotic gradient, causing mitochondrial matrix swelling, which in turn inhibits mitochondrial respiration[46].
To investigate the impact of pre-cooling on mitochondria function preservation by slowing the mitochondria metabolism rate and delaying ATP depletion, we isolated mitochondria from liver tissue post-transplantation through sequential centrifugations. We first tested the influence of precooling on the activities of Complex I in the isolated liver mitochondria. Figure 3A illustrated that OLT significantly decreased the activity of Complex I in the control group’s isolated liver mitochondria by 47% compared to the sham group. Notably, precooling demonstrated a substantial preservation on Complex I activity in the recipient liver mitochondria, indicated by approximately 50% higher activity than the control group (n=5/group).
Next, we aimed to affirm their consequent occurrence in the mitochondria function by measuring the OCR in isolated mitochondrial of both precooling and control groups using Seahorse XFe24 Analyzer. As shown in Figure 3. B and C, basal OCR was significantly higher in the precooling group than the control group (n=5/group). State 3 phosphorylating respiration induced by addition of ADP was about 30% greater in the precooling group than that in the control group. There was a minor difference observed between the control and precooling groups in state 4o respiration, which was not statistically significant. FCCP (state 3μ) increased OCR for both groups, but the increase was greater in the precooling group than that in the control group. These studies demonstrated that liver precooling attenuated the overall mitochondrial damage, albeit an unremarkable preservation in mitochondrial function.
Liver Precooling Protected Transplanted Graft Function Against IRI
Liver pre-cooling was applied 30 min before the liver procurement to the C57BL/6 donor mice, followed by cold storage for 6 hours. Liver transplantation was then performed. Graft functions were evaluated by measurement of plasma AST, ALT, and total bilirubin levels by Antech Diagnostics at 1- and 2-days following transplantation. All recipients were tested in similar basal liver chemistry before transplantation. Recipients from donors with liver pre-cooling (n=5) exhibited significantly lower liver enzyme levels. The plasma ALT and AST were 42%-46% lower in the precooling group than the control group as shown in Figure 4A,B. Total bilirubin was about 53% lower in the precooling groups than the control group (Figure 4C). These data indicated that precooling pretreatment protects against liver IRI and preserved graft hepatocellular function.
Liver Precooling Attenuated Liver Cell Death
Histopathology analysis by H&E staining revealed more severe sinusoidal congestion, vacuolization, hepatocytes necrosis in the control group than precooling group at 2 days after OLT. TUNEL staining showed 43% less cell death in the pre-cooling group than control group, which was consistent with the serum ALT levels and H&E staining (Figure 4D). Caspase-cascade system plays a crucial role in liver IRI. Apoptotic active Caspases -3 directly caused hepatocellular apoptosis and reflected the progress of apoptosis in the ischemic liver graft. Next, we assessed the expression of cleaved Caspases-3 by immunohistochemistry analysis. Along with the TUNEL assay, the activity of Caspases-3 was significantly repressed in the liver graft tissue in the precooling group compared with the control group.
The enhanced functional parameters observed correlated with a reduction in histopathological evidence of injury. This indicates that the implementation of pre-cooling effectively shielded the donor liver from ischemic injury during cold storage and consequently improved the function of the transplanted graft.
Liver Precooling Suppressed Oxidative Stress and Inflammatory Response
During liver transplantation, IRI triggers a complex inflammatory program involving cytokines and chemokines. Following liver IRI, inflammatory cytokines and chemokines stimulate the release of reactive oxygen species (ROS), cytokines, myeloperoxidase (MPO), and various mediators, amplifying tissue damage. To evaluate the hepatoprotective effects of precooling in the context of oxidative stress and inflammatory stimulation, we compared oxidative damage parameters and cytokine profiles in liver lysates between precooling and control groups.
The cytokine array revealed that precooling treatment led to a decreased expression of more than 20 cytokines and inflammatory mediators. The heat map illustrated cytokines and inflammatory mediators with a reduction in expression by more than 2-fold compared to the control group (Figure 5A,B), including CCL6/C10, Chitinase 3-like 1, MMP9, Pentraxin 2/SAP. Subsequently, we conducted RT-PCR to assess the mRNA levels of these and inflammatory mediators. Precooling of the liver led to a substantial decrease ranging from 30% to 80% in the mRNA expression of CCL6/C10, Chitinase 3-like 1, MMP9, and Pentraxin 2/SAP in the precooling group compared to the non-precooling group (Figure 5C). Liver precooling significantly reduced oxidative damage parameters assessed in this study. Specifically, liver carbonyl levels, indicative of protein oxidative damage, were measured significantly lower in the precooling group than control group post-transplantation. While both precooling and control groups showed a significant increase in total MDA levels, the magnitude of this increase was notably lower in the precooling group compared to the control group (Figure 5D). This suppression of the oxidative stress and inflammatory response, alongside a reduction in metabolic rate and oxygen consumption, contributes to the protective effects of precooling on liver grafts, thereby mitigating ischemic injury during liver transplantation.

Table 1.
Primer pairs for the RT-PCR.

Donor Body Temperature during Liver Procurement Surgery Correlates with Transplanted Graft Outcomes
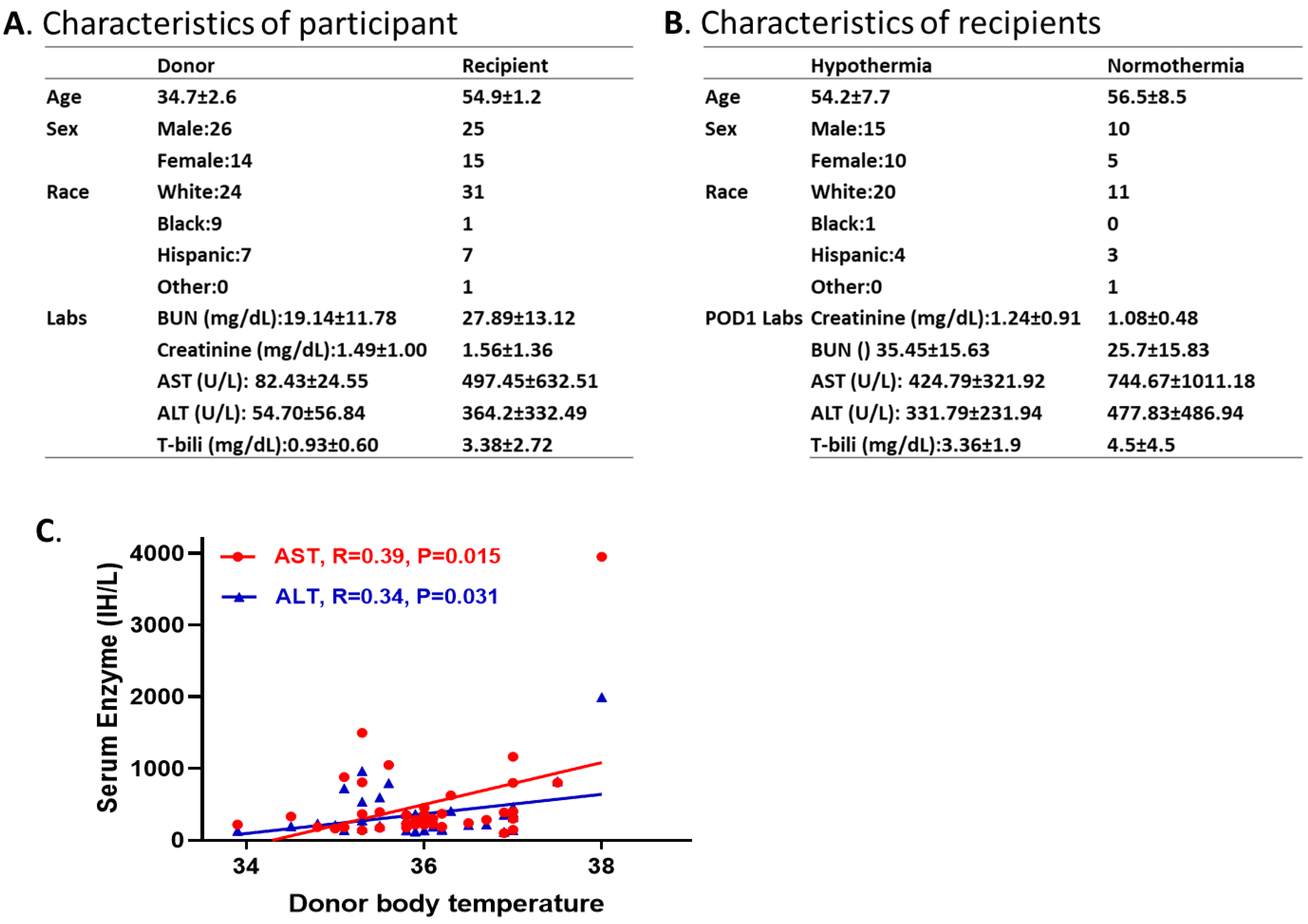
We collected and analyzed 40 donor and recipient pairs who underwent deceased liver transplants performed at TGH. The data of the donor (14 females and 26 males; age 12-66 years old) body temperature and recipient (15 females and 25 males; age 27-69 years old) liver chemistry were obtained from electronic health records at TGH. None of the donors in this cohort had any prior history of liver disease or significant alcohol consumption (exceeding 20 g/day). Additionally, all participants tested negative for hepatitis B, C, or D viruses, as well as other known viral infections. Demographic characteristics of the participants and the outcomes of the recipients are presented in Figure 6A,B, including age, sex, race, and liver chemistry. Possible associations between the donors’ temperature and recipients’ liver graft function were explored. A lower donor body temperature closely correlated with improved liver graft function in recipients, with the correlation coefficients of 0.39 and 0.34 between donor body temperature and recipient AST and ALT values, respectively, at 1-day post-surgery. These results are consistent with our animal studies and provide additional support and rational for our hypothesis that liver precooling protects transplanted graft function. Additionally, these preliminary patient data showed strong support in clinical relevance and significance.
Discussion
Overcoming IRI and preserving organ quality are crucial challenges in transplantation surgery. Ongoing studies aim to find new methodologies for reducing liver IRI, as there are currently no clinically approved strategies to completely protect the liver from IRI. In this study, we explored precooling as a drug-free approach with profound protective effects against IRI during liver transplantation.
The protective effect of hypothermia is linked to a decrease in liver metabolism, leading to a subsequent reduction in oxygen requirement as temperature decreases. Prior research has established the protective effects of mild hypothermia against neurological impairment, cardiac arrest, ischemic stroke, and kidney damage[32,47,48]. However, these studies typically involved systemic application, limiting temperature reduction due to potential adverse effects on the overall system such as hypotension, cardiac arrhythmias, and metabolic acidosis[49]. Selective liver hypothermia or topical liver cooling avoids the systemic side effects and has been proven to be effective in some clinical trials [50,51]. Nevertheless, in all these reports, the utilization of hypothermia during organ procurement or implantation was emphasized, while the warm ischemic period occurring after vascular clamping and before organ perfusion was overlooked. In our ongoing research, we have implemented localized and topical precooling to specifically tackle this overlooked aspect. By circulating 4°C water around the liver lobes, we achieved continuous liver temperature reduction without affecting core temperature. This method rapidly lowered the liver temperature to 6-8°C, reducing the metabolic rate of the liver cells before vasculature clamping and surpassing the reduction achieved with conventional systemic mild hypothermia. Consequently, it enabled a more significant decrease in the oxygen requirement.
Precooling is expected to not only lower the energy demand but also diminish the accumulation of superoxide radicals, delaying ATP decline and consequently mitigating oxidative stress during ischemia. Elevated ROS production during liver ischemia-reperfusion triggers an oxidative stress response, leading to cellular damage and apoptosis[52]. Mitochondria are identified as the primary source of ROS product[53]. Notably, moderate hypothermia has demonstrated protective effects on isolated perfused rat livers by mitigating oxidative stress[54,55]. The reactive oxygen metabolite levels were reported decreased during the hypothermic stage of treatment which returned to control-group levels after rewarming. In our current investigation, we observed that levels of liver MDA and carbonyls remained significantly lower in the precooling group compared to the control group post-transplantation. This observation suggests that while ROS generation may return to basal levels after rewarming, the activation of antioxidant defenses may impede protein damage and lipid peroxidation cascades.
The reduced oxidative damage may be correlated with the preserved mitochondrial respiratory enzyme activity. Studies have shown that mitochondrial ROS production exhibits an inverse relationship with oxidative phosphorylation rates[56,57]. Our results revealed a notably higher activity of Complex I within the precooling group than control group, suggesting that precooling likely mitigates the impairment of respiratory enzymes induced by IRI. The enhanced function of mitochondrial respiratory enzymes may play a pivotal role in sustaining the integrity of the mitochondrial respiratory chain, preserving membrane permeability, regulating Ca2+ levels, and fortifying mitochondrial defense mechanisms. Our high resolution respirometry data obtained with the Seahorse XF Analyzer confirmed that precooling preserved mitochondria function and alleviated IR-induced mitochondrial injury in livers.
Liver contains all the subtypes of lymphocytes as resident cells [58,59,60,61], acting as the source for the product of cytokines and chemokines[58, 62, 63]. These molecules, in turn, activate neutrophils during the late phase of reperfusion injury[64,65], and induce the release of ROS, cytokines, myeloperoxidase and various other mediators, all of which amplify the tissue damage[65,66,67,68]. It is widely recognized that an inflammatory response, dependent on neutrophils, contributes to IRI. Kato et al. demonstrated that hypothermia, with temperatures as low as 25°C, attenuates the hepatic inflammatory response induced by IRI[69]. In this study, we investigated if local precooling could blunt the inflammatory response following transplantation by using the Proteome Profiler Mouse XL Cytokine Array kit. We identified more than 20 inflammatory mediators involved in the inflammatory response that were significantly suppressed by the precooling. Further investigations will be necessary to discern the specific mediators within the inflammatory cascade that are modulated by pre-cooling.
Our current study provides pre-clinical evidence that application of locally pre-cooling can protect against IRI-induced acute liver injury. Its application in the mouse model of liver transplantation revealed protection of liver graft function, which is associated with attenuated mitochondrial oxidative stress, suppressed inflammatory response and subsequently decreased hepatocytes necrosis, fibrosis and apoptosis. Although direct application to humans is currently unfeasible, our retrospective analysis of 40 donor and recipient patient datasets, including organ donors and their respective liver transplant recipients, underscores a correlation between the donor’s body temperature and transplanted graft function. This retrospective analysis raises the promising prospect that the technique could be translated to human applications. Although we only analyzed a small size of data, a good correlation between donors’ temperature and recipients’ graft function has been achieved. We expect a significant correlation will be obtained if we increase the sample size as described in the protocol.
There are some limitations in our work. First, we only aimed to assess the effects of precooling on the short-term of graft function rather than long-term function. The detailed mechanisms of the antioxidant enzymes, which contribute to oxidative stress in mitochondria and cytokines will be covered in the continuous study. We only select the adult recipients for this study as pediatric liver transplantation recipients may have specific etiologies and outcomes with justified independent pediatric guidelines which are widely different from adult patients. Only a limited amount of patient data was analyzed in this retrospective study. Further research with a larger patient cohort is necessary to validate the findings presented here. In summary, our study describes a new strategy of local pre-cooling that attenuated oxidation stress and inflammatory response in the liver, ameliorated hepato-cellular damage and improved transplanted liver graft function. This technique has the potential to translate to humans not only in the improvement of graft survival, but also in the expansion of donor pool.
Author Contributions
MT, JG, RL and LW designed the research study and generated the draft of the manuscript. MT established the liver temperature changing pattern. LW conducted the OLT with assistance from MT. MT, JG and CP collected the specimen and performed the measurement. JG and YF performed histology and analyzed the data. CE and CP performed the mitochondria bioenergetic evaluation and analysis. JG and CP performed the cytokine array and RT-PCR experiment and analysis. MT, RL and LW analyzed human data. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Funding
This work was supported by American Heart Associate Career Development Grant (18CDA34110441) and National Institutes of Health Grant (R01DK122050).
Institutional Review Board Statement
All animal use and welfare adhered to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. This study followed the protocol approved by the Institutional Animal Care and Use Committee (IACUC, IS000011615R) at the University of South Florida.
Data Availability Statement
All the data and materials supporting the findings of this study are available within the article. Further enquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare that they have no competing financial interests or personal relationships that could have potentially influenced the findings presented in this paper.
References
- Onghena, L., et al., Quality of life after liver transplantation: State of the art. World J Hepatol, 2016. 8(18): p. 749-56. [CrossRef]
- Zaydfudim, V., et al., Reduction in corticosteroids is associated with better health-related quality of life after liver transplantation. J Am Coll Surg, 2012. 214(2): p. 164-73. [CrossRef]
- Wang, G.S., et al., Health-related quality of life after liver transplantation: the experience from a single Chinese center. Hepatobiliary Pancreat Dis Int, 2012. 11(3): p. 262-6. [CrossRef]
- Masala, D., et al., Quality of life and physical activity in liver transplantation patients: results of a case-control study in Italy. Transplant Proc, 2012. 44(5): p. 1346-50. [CrossRef]
- Zhai, Y., et al., Ischaemia-reperfusion injury in liver transplantation--from bench to bedside. Nat Rev Gastroenterol Hepatol, 2013. 10(2): p. 79-89.
- Franco-Gou, R., et al., New preservation strategies for preventing liver grafts against cold ischemia reperfusion injury. J Gastroenterol Hepatol, 2007. 22(7): p. 1120-6. [CrossRef]
- Land, W.G., The role of postischemic reperfusion injury and other nonantigen-dependent inflammatory pathways in transplantation. Transplantation, 2005. 79(5): p. 505-14. [CrossRef]
- Ikeda, T., et al., Ischemic injury in liver transplantation: difference in injury sites between warm and cold ischemia in rats. Hepatology, 1992. 16(2): p. 454-61. [CrossRef]
- Hashimoto, K., Liver graft from donation after circulatory death donor: Real practice to improve graft viability. Clin Mol Hepatol, 2020. 26(4): p. 401-410. [CrossRef]
- Reich, D.J., et al., ASTS recommended practice guidelines for controlled donation after cardiac death organ procurement and transplantation. Am J Transplant, 2009. 9(9): p. 2004-11. [CrossRef]
- van Golen, R.F., et al., Organ cooling in liver transplantation and resection: how low should we go? Hepatology, 2015. 61(1): p. 395-9.
- Lee, C.Y. and M.J. Mangino, Preservation methods for kidney and liver. Organogenesis, 2009. 5(3): p. 105-12. [CrossRef]
- Teoh, N.C. and J. Hartley, Modifications to the UW: simple solutions to the ‘sticky’ problem of preservation injury in liver transplantation. J Gastroenterol Hepatol, 2007. 22(10): p. 1564-5. [CrossRef]
- Matsuno, N. and E. Kobayashi, Challenges in machine perfusion preservation for liver grafts from donation after circulatory death. Transplant Res, 2013. 2(1): p. 19. [CrossRef]
- Guibert, E.E., et al., Organ Preservation: Current Concepts and New Strategies for the Next Decade. Transfus Med Hemother, 2011. 38(2): p. 125-142.
- Ravikumar, R., et al., Liver Transplantation After Ex Vivo Normothermic Machine Preservation: A Phase 1 (First-in-Man) Clinical Trial. Am J Transplant, 2016. 16(6): p. 1779-87. [CrossRef]
- Nasralla, D., et al., A randomized trial of normothermic preservation in liver transplantation. Nature, 2018. 557(7703): p. 50-56.
- Henry, S.D. and J.V. Guarrera, Protective effects of hypothermic ex vivo perfusion on ischemia/reperfusion injury and transplant outcomes. Transplant Rev (Orlando), 2012. 26(2): p. 163-75. , . [CrossRef]
- Yamanaka, K., et al., A systematic review of pharmacological treatment options used to reduce ischemia reperfusion injury in rat liver transplantation. PLoS One, 2014. 10(4): p. e0122214. [CrossRef]
- Whitsett, M. and J. Levitsky, Medication nonadherence in liver transplantation. Clin Liver Dis (Hoboken), 2017. 10(6): p. 157-160. [CrossRef]
- Lee, S.G., A complete treatment of adult living donor liver transplantation: a review of surgical technique and current challenges to expand indication of patients. Am J Transplant, 2015. 15(1): p. 17-38. [CrossRef]
- Fernandez, L., et al., Preconditioning protects liver and lung damage in rat liver transplantation: role of xanthine/xanthine oxidase. Hepatology, 2002. 36(3): p. 562-72. [CrossRef]
- Casillas-Ramirez, A., et al., Past and future approaches to ischemia-reperfusion lesion associated with liver transplantation. Life Sci, 2006. 79(20): p. 1881-94.
- Cannistra, M., et al., Hepatic ischemia reperfusion injury: A systematic review of literature and the role of current drugs and biomarkers. Int J Surg, 2016. 33 Suppl 1: p. S57-70. [CrossRef]
- Lee, M.P. and A.R. Gear, The effect of temperature on mitochondrial membrane-linked reactions. J Biol. Chem, 1974. 249(23): p. 7541-7549. [CrossRef]
- Geiser, F., Metabolic rate and body temperature reduction during hibernation and daily torpor. Annu. Rev. Physiol, 2004. 66: p. 239-274. [CrossRef]
- Tian, G., et al., A comparison of UW cold storage solution and St. Thomas’ solution II: a 31P NMR and functional study of isolated porcine hearts. J Heart Lung Transplant, 1991. 10(6): p. 975-85.
- Keon, W.J., et al., Cardiac transplantation: the ideal myocardial temperature for graft transport. Ann Thorac Surg, 1988. 46(3): p. 337-41. [CrossRef]
- Bernard, M., et al., The influence of temperature on metabolic and cellular protection of the heart during long-term ischemia: a study using P-31 magnetic resonance spectroscopy and biochemical analyses. Cryobiology, 1998. 37(4): p. 309-17. [CrossRef]
- Benson, D.W., et al., The use of hypothermia after cardiac arrest. Anesth Analg, 1959. 38: p. 423-8. [CrossRef]
- Williams, G.R., Jr. and F.C. Spencer, The clinical use of hypothermia following cardiac arrest. Ann Surg, 1958. 148(3): p. 462-8. [CrossRef]
- Yamanaka, N., C.L. Dai, and E. Okamoto, Historical evolution of hypothermic liver surgery. World J Surg, 1998. 22(10): p. 1104-7. [CrossRef]
- Zeiner, A., et al., Mild resuscitative hypothermia to improve neurological outcome after cardiac arrest. A clinical feasibility trial. Hypothermia After Cardiac Arrest (HACA) Study Group. Stroke, 2000. 31(1): p. 86-94. [CrossRef]
- Pacini, D., et al., Visceral organ protection in aortic arch surgery: safety of moderate hypothermia. Eur J Cardiothorac Surg, 2014. 46(3): p. 438-43. [CrossRef]
- Nolan, J.P., et al., Therapeutic hypothermia after cardiac arrest: an advisory statement by the advanced life support task force of the International Liaison Committee on Resuscitation. Circulation, 2003. 108(1): p. 118-21. [CrossRef]
- Niemann, C.U., et al., Therapeutic Hypothermia in Deceased Organ Donors and Kidney-Graft Function. N Engl J Med, 2015. 373(5): p. 405-14.
- Hessel, E.A., 2nd, Therapeutic hypothermia after in-hospital cardiac arrest: a critique. J Cardiothorac Vasc Anesth, 2014. 28(3): p. 789-99. [CrossRef]
- Yokota, S., et al., Orthotopic mouse liver transplantation to study liver biology and allograft tolerance. Nat Protoc, 2016. 11(7): p. 1163-74. [CrossRef]
- Mendonca, R., et al., Sensitive and selective quantification of free and total malondialdehyde in plasma using UHPLC-HRMS. J Lipid Res, 2017. 58(9): p. 1924-1931. [CrossRef]
- Levine, R.L., Carbonyl modified proteins in cellular regulation, aging, and disease. Free Radic Biol Med, 2002. 32(9): p. 790-6. [CrossRef]
- Suzuki, S., et al., The beneficial effect of a prostaglandin I2 analog on ischemic rat liver. Transplantation, 1991. 52(6): p. 979-83. [CrossRef]
- Wang, L., et al., Graft function assessment in mouse models of single- and dual-kidney transplantation. Am J Physiol Renal Physiol, 2018. 315(3): p. F628-F636. [CrossRef]
- Chen, W., et al., Reducing ischemic kidney injury through application of a synchronization modulation electric field to maintain Na(+)/K(+)-ATPase functions. Sci Transl Med, 2022. 14(635): p. 10.1126/scitranslmed.abj4906. [CrossRef]
- McCrimmon, A., et al., Comprehensive assessment of mitochondrial respiratory function in freshly isolated nephron segments. Am J Physiol Renal Physiol, 2020. 318(5): p. F1237-F1245. [CrossRef]
- Wang, L., et al., Cross-sex transplantation alters gene expression and enhances inflammatory response in the transplanted kidneys. American Journal of Physiology - Renal Physiology, 2017. 313(2): p. F326-F338.
- Kalogeris, T., et al., Cell biology of ischemia/reperfusion injury. Int Rev Cell Mol Biol, 2012. 298: p. 229-317.
- Meier, R.P.H., et al., Intra-Abdominal Cooling System Limits Ischemia-Reperfusion Injury During Robot-Assisted Renal Transplantation. Am J Transplant, 2018. 18(1): p. 53-62. [CrossRef]
- Heijnen, B.H., et al., Decrease in core liver temperature with 10 degrees C by in situ hypothermic perfusion under total hepatic vascular exclusion reduces liver ischemia and reperfusion injury during partial hepatectomy in pigs. Surgery, 2003. 134(5): p. 806-17.
- Hildebrand, F., et al., Pathophysiologic changes and effects of hypothermia on outcome in elective surgery and trauma patients. Am J Surg, 2004. 187(3): p. 363-71. [CrossRef]
- Grezzana Filho, T.J.M., et al., Induction of selective liver hypothermia prevents significant ischemia/reperfusion injury in Wistar rats after 24 hours. Acta Cir Bras, 2020. 35(2): p. e202000205. [CrossRef]
- Azoulay, D., et al., In situ hypothermic perfusion of the liver versus standard total vascular exclusion for complex liver resection. Ann Surg, 2005. 241(2): p. 277-85. [CrossRef]
- Spencer, N.Y., et al., Hepatocytes produce TNF-alpha following hypoxia-reoxygenation and liver ischemia-reperfusion in a NADPH oxidase- and c-Src-dependent manner. Am J Physiol Gastrointest Liver Physiol, 2013. 305(1): p. G84-94. [CrossRef]
- He, H., et al., Role of mitochondria on UV-induced skin damage and molecular mechanisms of active chemical compounds targeting mitochondria. J Photochem Photobiol B, 2022. 232: p. 112464. [CrossRef]
- Choi, S., et al., Mild hypothermia provides significant protection against ischemia/reperfusion injury in livers of obese and lean rats. Ann Surg, 2005. 241(3): p. 470-6. [CrossRef]
- Zar, H.A. and J.R. Lancaster, Jr., Mild hypothermia protects against postischemic hepatic endothelial injury and decreases the formation of reactive oxygen species. Redox Rep, 2000. 5(5): p. 303-10. [CrossRef]
- Cadenas, E. and K.J. Davies, Mitochondrial free radical generation, oxidative stress, and aging. Free Radic Biol Med, 2000. 29(3-4): p. 222-30. [CrossRef]
- Balaban, R.S., S. Nemoto, and T. Finkel, Mitochondria, oxidants, and aging. Cell, 2005. 120(4): p. 483-95.
- Wang, Y. and C. Zhang, The Roles of Liver-Resident Lymphocytes in Liver Diseases. Front Immunol, 2019. 10: p. 1582. [CrossRef]
- Caldwell, C.C., J. Tschoep, and A.B. Lentsch, Lymphocyte function during hepatic ischemia/reperfusion injury. J Leukoc Biol, 2007. 82(3): p. 457-64. [CrossRef]
- Calne, R.Y., Immunological tolerance--the liver effect. Immunol Rev, 2000. 174: p. 280-2. [CrossRef]
- Tiegs, G. and A.W. Lohse, Immune tolerance: what is unique about the liver. J Autoimmun, 2010. 34(1): p. 1-6. [CrossRef]
- Vardanian, A.J., R.W. Busuttil, and J.W. Kupiec-Weglinski, Molecular mediators of liver ischemia and reperfusion injury: a brief review. Mol Med, 2008. 14(5-6): p. 337-45. [CrossRef]
- Zwacka, R.M., et al., Redox gene therapy for ischemia/reperfusion injury of the liver reduces AP1 and NF-kappaB activation. Nat Med, 1998. 4(6): p. 698-704.
- Shibuya, H., et al., Tumor necrosis factor-induced, superoxide-mediated neutrophil accumulation in cold ischemic/reperfused rat liver. Hepatology, 1997. 26(1): p. 113-20.
- Ahmed, O., et al., Hepatic stellate cell activation markers are regulated by the vagus nerve in systemic inflammation. Bioelectron Med, 2023. 9(1): p. 6. [CrossRef]
- Nakamura, K., S. Kageyama, and J.W. Kupiec-Weglinski, The Evolving Role of Neutrophils in Liver Transplant Ischemia-Reperfusion Injury. Curr Transplant Rep, 2019. 6(1): p. 78-89. [CrossRef]
- Peralta, C., M.B. Jimenez-Castro, and J. Gracia-Sancho, Hepatic ischemia and reperfusion injury: effects on the liver sinusoidal milieu. J Hepatol, 2013. 59(5): p. 1094-106. [CrossRef]
- Wen, Y., et al., Hepatic macrophages in liver homeostasis and diseases-diversity, plasticity and therapeutic opportunities. Cell Mol Immunol, 2021. 18(1): p. 45-56. [CrossRef]
- Kato, A., et al., Mechanisms of hypothermic protection against ischemic liver injury in mice. Am J Physiol Gastrointest Liver Physiol, 2002. 282(4): p. G608-16. [CrossRef]
Figure 1.
Experimental procedure.
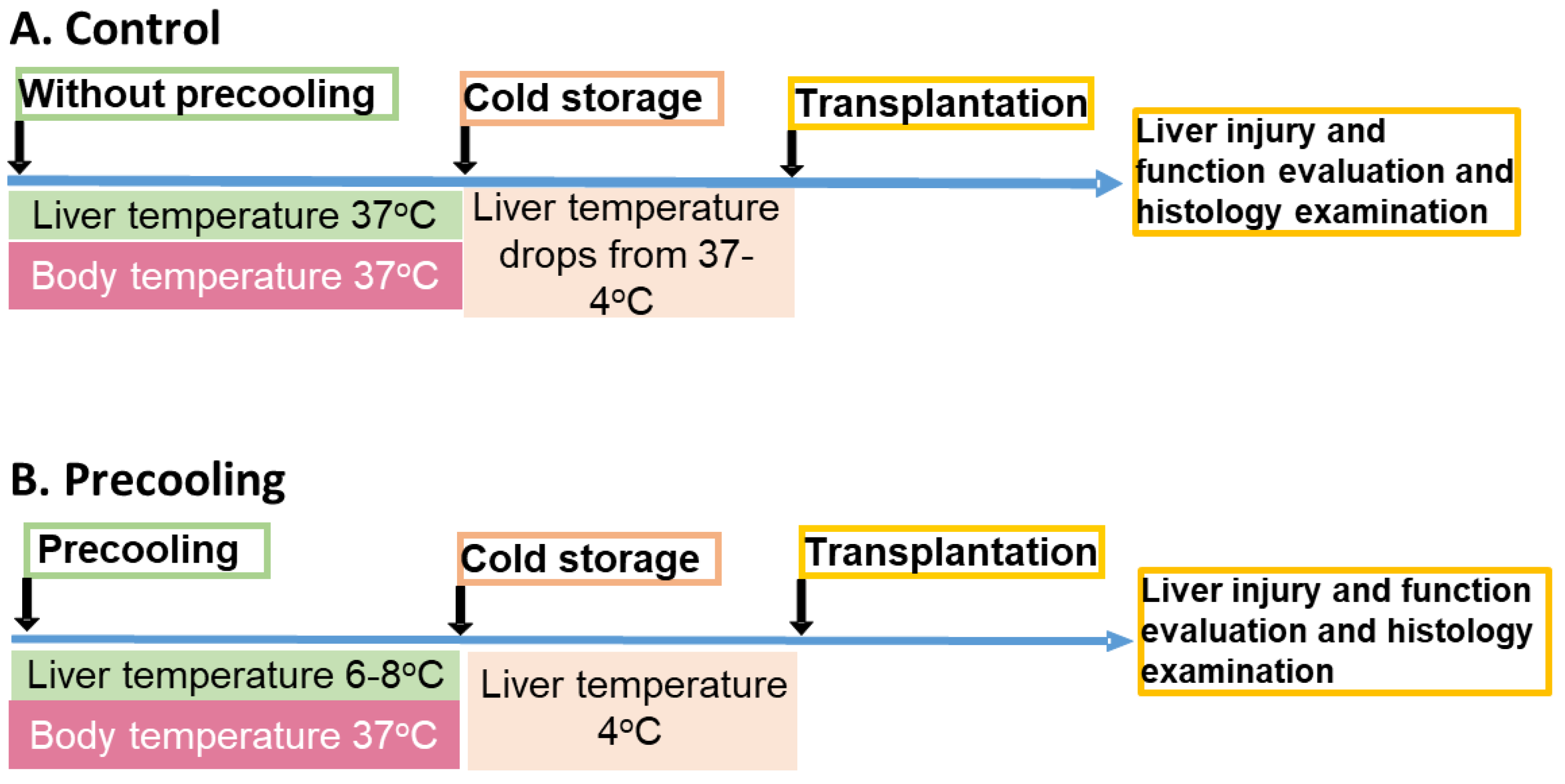
Figure 2.
Liver temperature changes during the precooling.
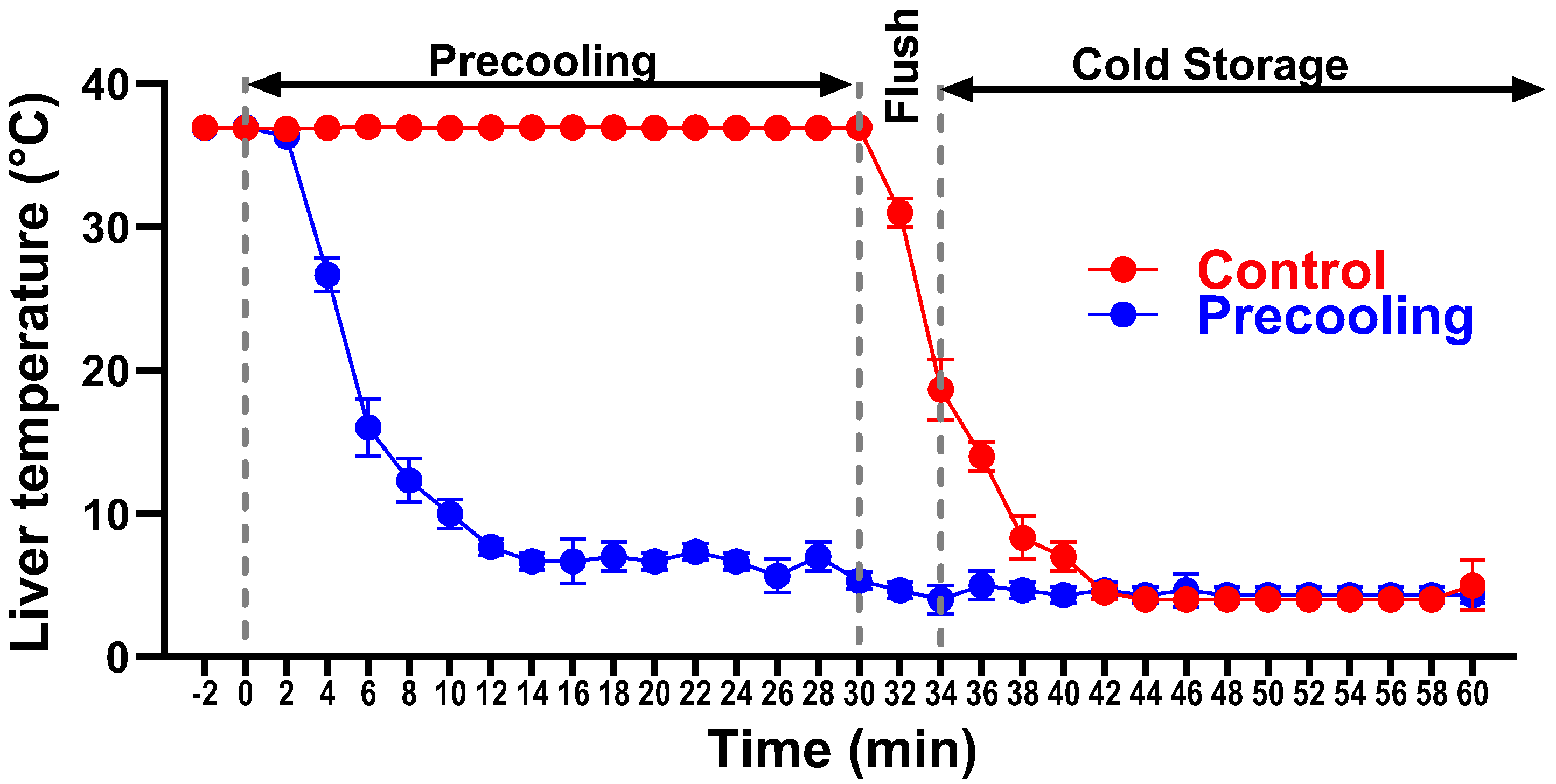
Figure 3.
Liver mitochondrial function is preserved with the application of precooling. A, Mitochondria Complex I activities were preserved in the precooling group. B, OCR traces of isolated mitochondria from liver grafts with or without precooling, expressed as picomoles of O2 per minute, under basal conditions and after the injection of adenosine diphosphate (ADP; 1 mM), oligomycin (Oligo; 2 µM), carbonyl-cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP; 4 µM), and antimycin A (AA) + rotenone (2 µM). C, Analysis of mitochondrial respiratory parameters obtained from normalized XFe24 graphs (B). Data are presented as means ± SEM. Statistical significance was assessed by one-way ANOVA followed by Tukey multiple comparisons test (*P < 0.05, ***P < 0.001, and ****P < 0.0001, ns, not significant; n = 5 mice per group).
Figure 3.
Liver mitochondrial function is preserved with the application of precooling. A, Mitochondria Complex I activities were preserved in the precooling group. B, OCR traces of isolated mitochondria from liver grafts with or without precooling, expressed as picomoles of O2 per minute, under basal conditions and after the injection of adenosine diphosphate (ADP; 1 mM), oligomycin (Oligo; 2 µM), carbonyl-cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP; 4 µM), and antimycin A (AA) + rotenone (2 µM). C, Analysis of mitochondrial respiratory parameters obtained from normalized XFe24 graphs (B). Data are presented as means ± SEM. Statistical significance was assessed by one-way ANOVA followed by Tukey multiple comparisons test (*P < 0.05, ***P < 0.001, and ****P < 0.0001, ns, not significant; n = 5 mice per group).
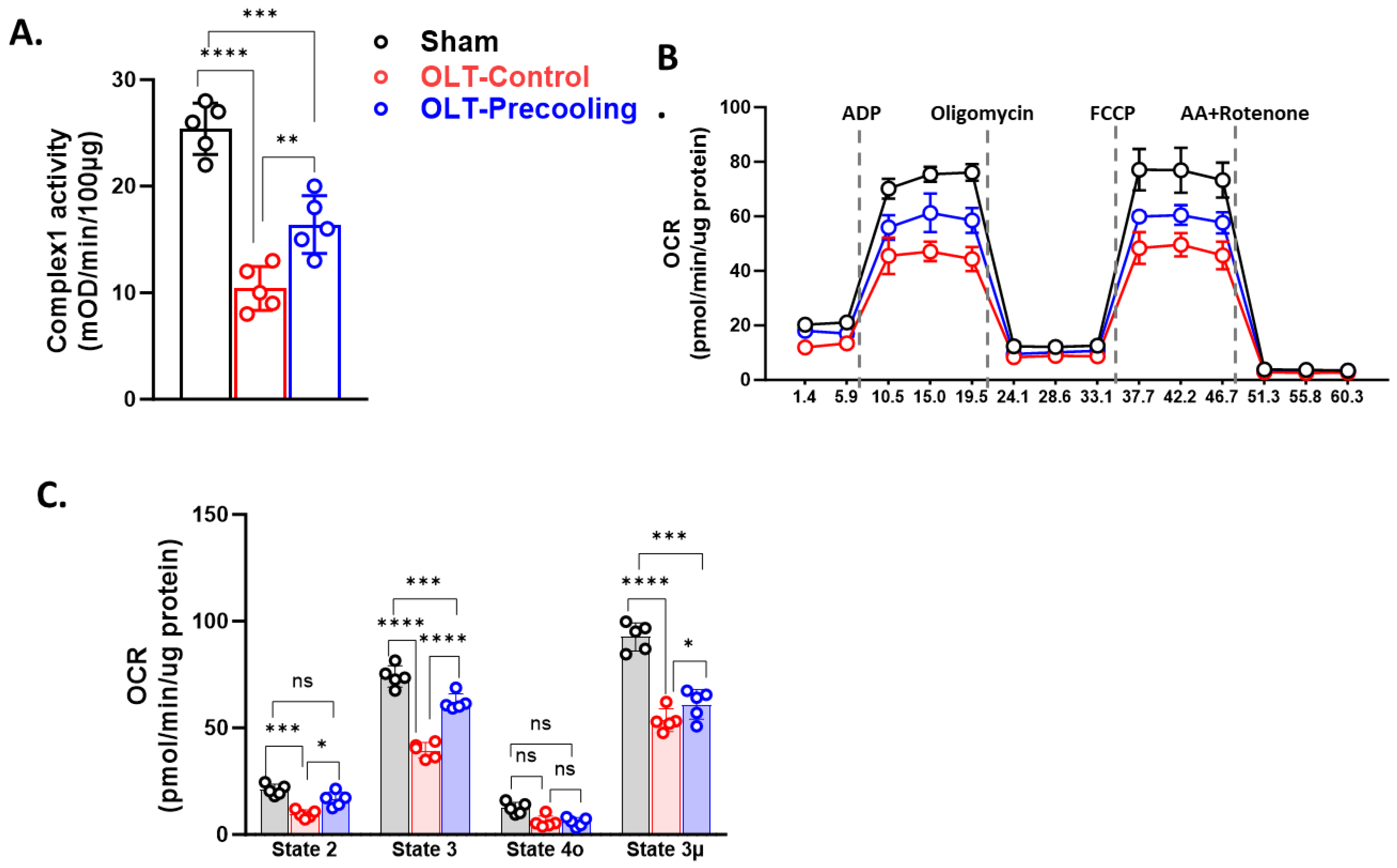
Figure 4.
IRI–induced liver graft injury is attenuated by the application of precooling. Plasma ALT (A), AST (B) and total bilirubin (C) levels measurements on day 1 and 2 after transplant demonstrated decreased liver injury by the application of precooling. (*P < 0.05, **P< 0.01, ***P < 0.001 and ****P < 0.0001, Statistical significance was assessed by two-way ANOVA followed by Tukey multiple comparisons test. n = 5 mouse recipients.) (D) Representative liver tissue sections stained with H&E, TUNEL, and immunofluorescent anti-Caspase-3. (E) Quantification of sinusoidal congestion, vacuolization and hepatocytes necrosis area over whole-liver sections, TUNEL-positive cells/nuclei as the percentage (%) of TUNEL, and the percentage of Caspase 3 positive cells/nuclei. Scale bar is 50µm. Data are presented as means ± SEM. One-way ANOVA followed by Tukey multiple comparisons test **P< 0.01, ***P < 0.001, n = 5 liver sections per group).
Figure 4.
IRI–induced liver graft injury is attenuated by the application of precooling. Plasma ALT (A), AST (B) and total bilirubin (C) levels measurements on day 1 and 2 after transplant demonstrated decreased liver injury by the application of precooling. (*P < 0.05, **P< 0.01, ***P < 0.001 and ****P < 0.0001, Statistical significance was assessed by two-way ANOVA followed by Tukey multiple comparisons test. n = 5 mouse recipients.) (D) Representative liver tissue sections stained with H&E, TUNEL, and immunofluorescent anti-Caspase-3. (E) Quantification of sinusoidal congestion, vacuolization and hepatocytes necrosis area over whole-liver sections, TUNEL-positive cells/nuclei as the percentage (%) of TUNEL, and the percentage of Caspase 3 positive cells/nuclei. Scale bar is 50µm. Data are presented as means ± SEM. One-way ANOVA followed by Tukey multiple comparisons test **P< 0.01, ***P < 0.001, n = 5 liver sections per group).
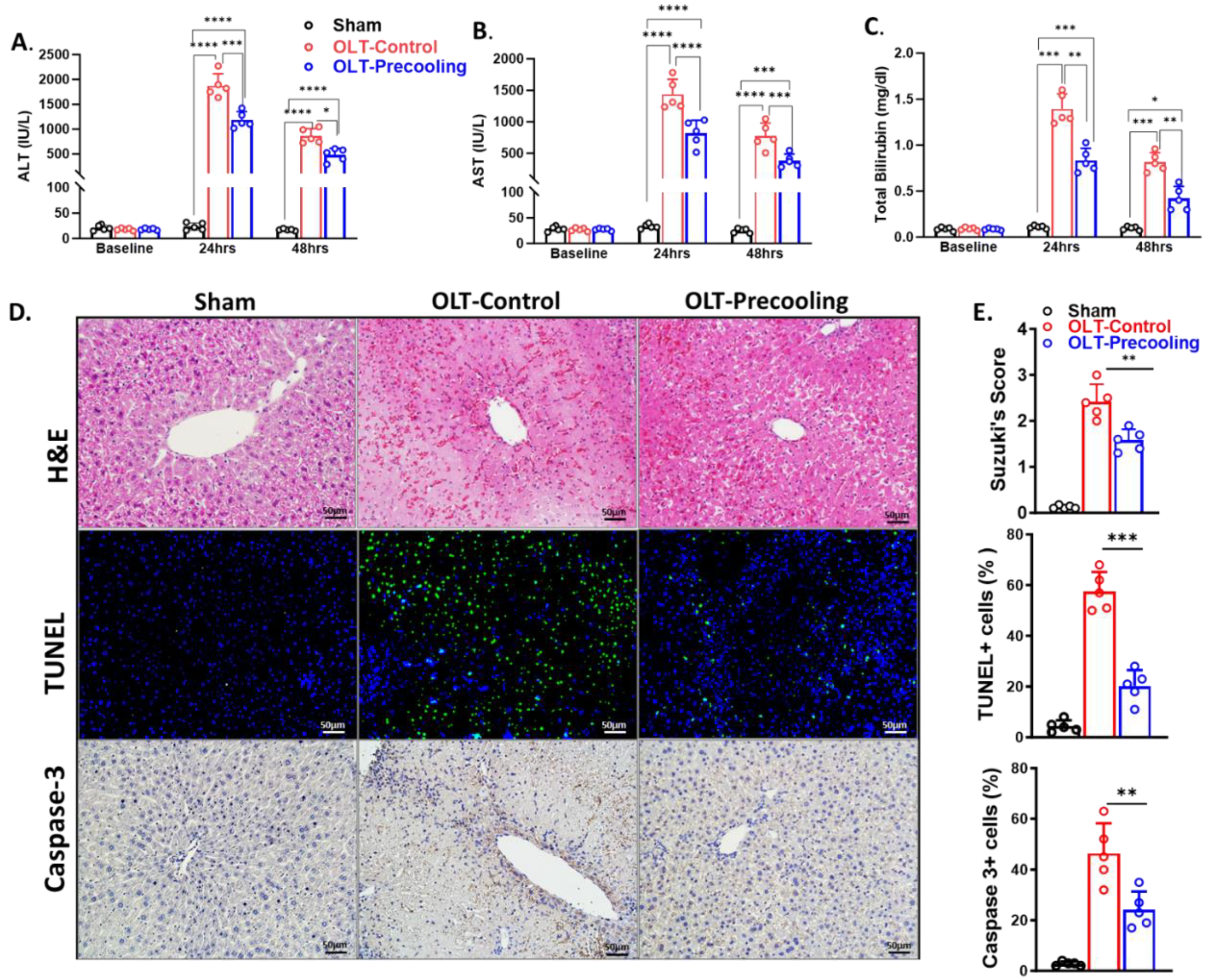
Figure 5.
Liver precooling suppressed inflammatory response and oxidative stress. A. Cytokine Arrays detection of multiple analytes in mouse liver graft tissues of control and precooling group. Each blot represents immunoreactive labeling against the respective antibodies. The blots labeled with colored squares represent cytokines that exhibited significant differences between the two groups. B. Heatmap representation of quantification of cytokine profile of control (C1, and C2) and precooling group (P1 and P2). Colored squares highlighted the most depressed cytokines by precooling. (n = 2 biologically independent samples/group). C. RT-PCR confirmed the mRNA levels of the genes exhibiting the most significant differences between the control and precooling groups. (**P< 0.01 and ****P < 0.0001, Statistical significance was assessed by One-way ANOVA followed by Tukey multiple comparisons test. n = 5 mouse recipients.) D. Liver carbonyl and MDA levels demonstrated reduced oxidative damage in the precooling group than control group. (*P < 0.05, **P< 0.01, ***P < 0.001 and ****P < 0.0001, Statistical significance was assessed by two-way ANOVA followed by Tukey multiple comparisons test. n = 5 mouse recipients.).
Figure 5.
Liver precooling suppressed inflammatory response and oxidative stress. A. Cytokine Arrays detection of multiple analytes in mouse liver graft tissues of control and precooling group. Each blot represents immunoreactive labeling against the respective antibodies. The blots labeled with colored squares represent cytokines that exhibited significant differences between the two groups. B. Heatmap representation of quantification of cytokine profile of control (C1, and C2) and precooling group (P1 and P2). Colored squares highlighted the most depressed cytokines by precooling. (n = 2 biologically independent samples/group). C. RT-PCR confirmed the mRNA levels of the genes exhibiting the most significant differences between the control and precooling groups. (**P< 0.01 and ****P < 0.0001, Statistical significance was assessed by One-way ANOVA followed by Tukey multiple comparisons test. n = 5 mouse recipients.) D. Liver carbonyl and MDA levels demonstrated reduced oxidative damage in the precooling group than control group. (*P < 0.05, **P< 0.01, ***P < 0.001 and ****P < 0.0001, Statistical significance was assessed by two-way ANOVA followed by Tukey multiple comparisons test. n = 5 mouse recipients.).
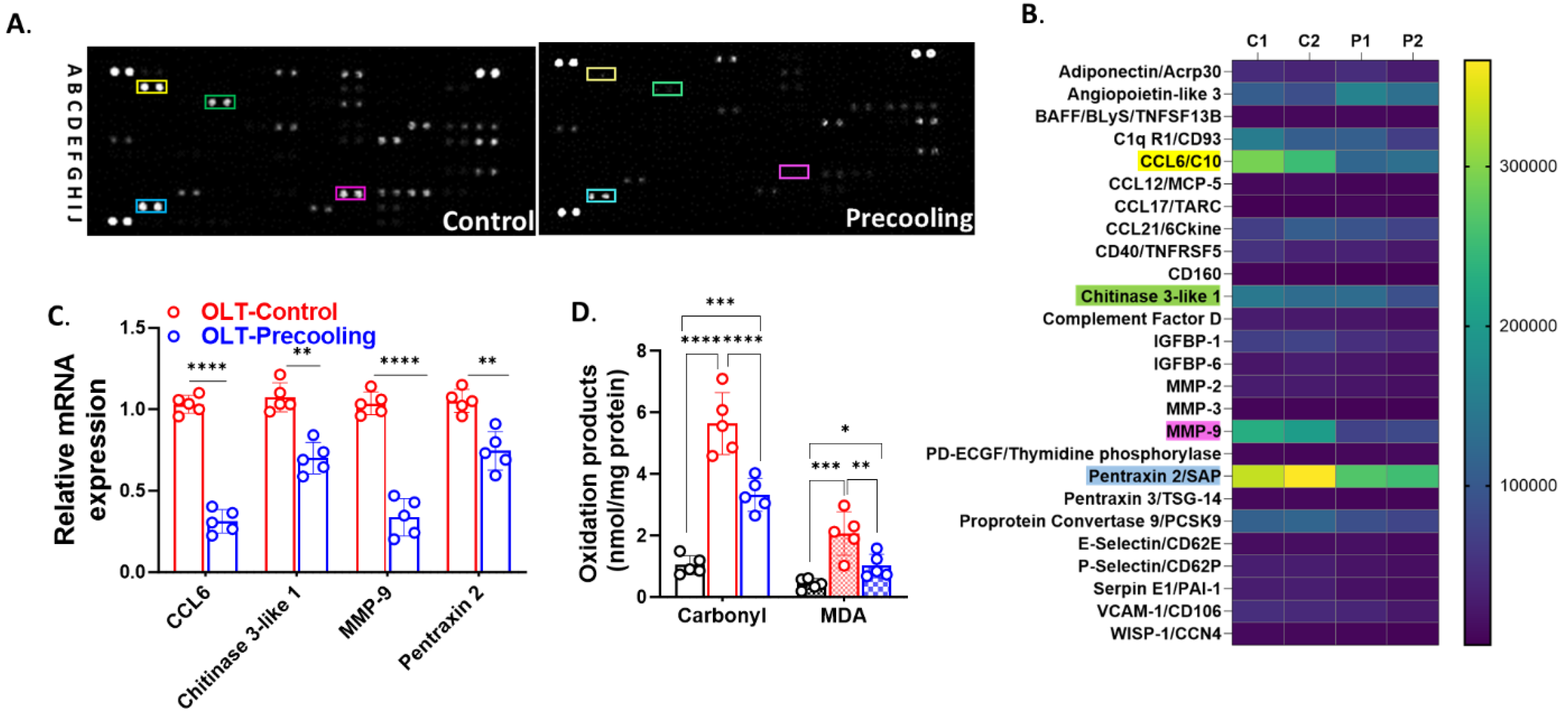
Figure 6.
The association between donors’ body temperature and the recipients’ liver enzyme levels. A. Demographic characteristics of the participants. B. Demographic characteristics of the recipients. C. A lower donor body temperature correlates with improved liver graft function in recipients, with correlation coefficients of 0.39 and 0.34 between donor body temperature and recipients’ AST and ALT values. Statistical difference was calculated by Pearson’s R.
Figure 6.
The association between donors’ body temperature and the recipients’ liver enzyme levels. A. Demographic characteristics of the participants. B. Demographic characteristics of the recipients. C. A lower donor body temperature correlates with improved liver graft function in recipients, with correlation coefficients of 0.39 and 0.34 between donor body temperature and recipients’ AST and ALT values. Statistical difference was calculated by Pearson’s R.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



